
Method Article
Rekonstitution der Aktin-basierten Motilität mit kommerziell erhältlichen Proteinen
In diesem Artikel
Zusammenfassung
Dieses Protokoll beschreibt, wie Aktinkometen auf den Oberflächen von Perlen mit kommerziell erhältlichen Proteinbestandteilen hergestellt werden können. Solche Systeme ahmen die hervorstehenden Strukturen in Zellen nach und können verwendet werden, um physiologische Mechanismen der Kraftproduktion auf vereinfachte Weise zu untersuchen.
Zusammenfassung
Viele Zellbewegungen und Formänderungen sowie bestimmte Arten von intrazellulärer Bakterien- und Organellenmotilität werden durch das Biopolymer Aktin angetrieben, das ein dynamisches Netzwerk an der Oberfläche der Zelle, Organelle oder des Bakteriums bildet. Die biochemischen und mechanischen Grundlagen der Krafterzeugung während dieses Prozesses können untersucht werden, indem Aktin-basierte Bewegungen azellulär auf inerten Oberflächen wie Perlen reproduziert werden, die funktionalisiert und mit einem kontrollierten Satz von Komponenten inkubiert werden. Unter geeigneten Bedingungen baut sich an der Perlenoberfläche ein elastisches Aktinnetzwerk auf, das durch die durch das Netzwerkwachstum erzeugte Belastung aufbricht und einen "Aktinkometen" bildet, der die Perle vorwärts treibt. Solche Experimente erfordern jedoch die Reinigung einer Vielzahl verschiedener Aktin-bindender Proteine, die oft außerhalb der Reichweite von Nicht-Spezialisten liegen. Dieser Artikel beschreibt ein Protokoll zur reproduzierbaren Gewinnung von Aktinkometen und zur Motilität von Perlen unter Verwendung kommerziell erhältlicher Reagenzien. Kugelbeschichtung, Raupengröße und Motilitätsmischung können geändert werden, um den Effekt auf Raupengeschwindigkeit, Trajektorien und andere Parameter zu beobachten. Dieser Assay kann verwendet werden, um die biochemischen Aktivitäten verschiedener Aktin-bindender Proteine zu testen und quantitative physikalische Messungen durchzuführen, die Aufschluss über die Eigenschaften der aktiven Substanz von Aktinnetzwerken geben. Dies wird ein nützliches Werkzeug für die Gemeinschaft sein, das die Untersuchung der In-vitro-Aktin-basierten Motilität ohne Expertenwissen in der Aktin-bindenden Proteinreinigung ermöglicht.
Einleitung
Die Aktinpolymerisation in Zellen wird räumlich und zeitlich durch eine enge Regulation der Aktinfilamentkeimbildung stromabwärts der Zellsignalisierunggesteuert 1. Die Keimbildung erfolgt durch die Bildung eines Aktintrimers, und dann polymerisieren beide Enden des entstehenden Filaments spontan, obwohl ein Ende dynamischer ist (das Stachelende) als das andere (das spitze Ende)2. Wenn Keimbildung und Stachelendpolymerisation auf eine Oberfläche gerichtet sind, erzeugen sie genügend Kraft (im Pico- bis Nano-Newton-Bereich), um die Zellmembran für die Bewegung herauszudrücken und mikrometergroße Objekte innerhalb der Zelle mit ATP-Hydrolyse als Energiequellezu bewegen 3. Einige Beispiele sind Listeria monocytogenes-Bakterien, die Aktinkometen verwenden, um sich von Zelle zu Zelle auszubreiten, und Mitochondrien, bei denen die Bewegung auf Aktinkometen für die randomisierte Vererbung während der Mitose wichtig ist 4,5. Aktinkometen auf Endosomen und anderen intrazellulären Vesikeln sind an der Ablösung von Spendermembranen beteiligt 6,7,8.
Mit dem hier vorgestellten Verfahren werden die Signalaspekte der zellulären Aktinpolymerisation umgangen und die Aktinpolymerisation auf mikrometrischen Polystyrolkügelchen hergestellt, indem sie mit Aktivatoren der verzweigten Aktinkeimbildung, insbesondere der aktiven Domäne des menschlichen WASP-Proteins, VCA (auch WA oder WCA genannt)1, beschichtet werden. Die beschichteten Beads werden dann in einer Mischung inkubiert, die die für die Aktinpolymerisation notwendigen Bestandteile enthält, einschließlich des Hauptaktinpolymerisationsnukleators in Zellen, des Arp2/3-Komplexes, der durch VCA an der Perlenoberfläche aktiviert wird, um neue Filamente als Verzweigungen von den Seiten der Tochterfilamente1 zu bilden. Aktin polymerisiert zunächst gleichmäßig um die Perle herum, bricht dann aber spontan die Symmetrie, um einen Aktinkometen zu erzeugen, der die Perle nach vorne schiebt und dadurch zellartige hervorstehende Netzwerke und Kometen kontrolliert nachbildet. Ähnliche Ansätze mit Perlen und anderen beschichteten Oberflächen wurden in der Vergangenheit von uns und anderen verwendet, um die Biochemie und Biophysik der Aktinpolymerisation 9,10,11,12 zu untersuchen, aber für diese Experimente war umfangreiche Expertise in Aktin-bindenden Proteinen erforderlich. Das hier vorgestellte Protokoll beschreibt, wie Aktinkometen und Motilität vollständig mit kommerziell erhältlichen (oder bald verfügbaren) Reagenzien robust erzeugt werden können, wodurch dieser Ansatz für jedermann zugänglich gemacht wird, auch in einem Bildungsumfeld für die Vermittlung biophysikalischer Konzepte. Zu den wichtigsten Merkmalen gehören die Bedeutung eines schonenden und zuverlässigen Pipettierens, die Verwendung von Profilin-komplexem Monomer als Aktinquelle und die Wesentlichkeit der Verwendung eines hochaktiven Apr2/3-Komplexaktivators als Perlenbeschichtungsreagenz.
Protokoll
1. Vorbereitung der Puffer
HINWEIS: Verwenden Sie ultrareines H2O für alle Puffer. Es muss nicht steril sein. Alle in den Schritten 1.1-1.4 beschriebenen Lösungen werden mit einem 0,2-μm-Spritzenfilter filtriert, je nach Verwendung in Portionen von 500 μL-2 ml pro Röhrchen aliquot und bei -20 °C gelagert.
- Bereiten Sie 10% BSA vor, indem Sie 2 g BSA in ein 50-ml-konisches Röhrchen wiegen und bis zur 20-ml-Marke mitH2Ofüllen. Mischen Sie, bis das BSA aufgelöst ist (ca. 30 min), und füllen Sie dann das Volumen auf 20 ml auf.
HINWEIS: Verwenden Sie hochwertiges BSA (siehe Materialtabelle). Die 10% ige BSA-Lösung wird sowohl in der Perlenzubereitung als auch in der Motilitätsmischung verwendet. - Zur Perlenherstellung Xb-Puffer (10 mM HEPES, 0,1 M KCl, 1 mM MgCl2 und 0,1 mM CaCl2, pH 7,5) als 10x-Lösung herstellen und vor Gebrauch verdünnen (100 μL 10x Xb-Stammlösung + 900 μLH2O). Herstellen von Xb/1% BSA durch Mischen von 100 μL 10x Xb Stammlösung + 100 μL 10% BSA + 800 μLH2O.
- Bereiten Sie einen G-Puffer (2 mM Tris, 0,2 mM CaCl 2, 0,2 mM DTT,2 mM ATP) vor, der zur Verdünnung von monomerem Aktin (G-Aktin) verwendet wird. Stellen Sie sich auf pH 7 ein, nicht auf pH 8, wie traditionell verwendet (siehe Diskussion).
- Bereiten Sie den Motilitätspuffer MB13 vor (10 mM HEPES, 1,5 mM ATP, 3 mM DTT, 1,5 mM MgCl2, 1 mM EGTA, 50 mM KCl, 1% BSA, pH 7,5). Für einige Anwendungen ist 10x MB13 sinnvoll. Bereiten Sie jedoch 10x MB13 ohne BSA vor, da dies zu Problemen bei der pH-Einstellung führt. Fügen Sie BSA aus 10% Stammlösung (hergestellt in Schritt 1.1) hinzu, wenn 1x MB13 aus 10x MB13 rekonstituiert wird.
2. Herstellung von Proteinlösungen
HINWEIS: Verwenden Sie ultrareines H2O für alle Resuspensionen. Es muss nicht steril sein. Alle Proteine auf Eis behandeln und aliquot in vorgekühlte Röhrchen geben. Sanft manipulieren, um keine Blasen zu erzeugen, und niemals Wirbelproteinlösungen. Für die Lagerung von Vorräten bei -80°C ist ein Schockfrosten in flüssigem Stickstoff nicht notwendig. Passen Sie die aliquote Größe an, um mehr als etwa fünf Gefrier-Tau-Zyklen zu vermeiden, da dies die Aktivität eines der Proteine nicht zu beeinflussen scheint. Arbeitsaliquots können bei -20 °C für einige Wochen gelagert werden.
- Bereiten Sie G-Actin (Kaninchen-Skelettmuskel) Lösung wie unten beschrieben vor.
- Aktinpulver (siehe Materialtabelle) bei 4 °C pulsieren, um den Feststoff am Boden des Röhrchens zu sammeln.
- Fügen Sie H2O gemäß Herstellerangaben hinzu (1 mg Protein in 100 μL kaltem H2O für unmarkiertes Aktin, 100 μg Protein in 100 μL kaltemH2Ofür ATTO-markiertes Aktin).
- Lassen Sie es mindestens 15 Minuten auf Eis sitzen. Mischen Sie vorsichtig, indem Sie auf und ab pipettieren, lassen Sie es mindestens weitere 15 Minuten auf Eis sitzen und mischen Sie erneut. Pulszentrifuge bei 4 °C, um die Lösung am Boden des Röhrchens zu sammeln, und remixen.
- Bereiten Sie je nach Verwendung 10-50 μL Aliquots unmarkiertes Aktin und 20 μL Aliquots ATTO-markiertes Aktin vor. Lagern Sie die Aliquots bei -80 °C.
- Um Aktin-Oligomere, die sich während der Lyophilisation und des Gefrierens bilden, zu depolymerisieren, wird ein Aliquot resuspendiertes Aktin ~8-fach in G-Puffer verdünnt, der mit zusätzlichem ATP und DTT versetzt ist (z. B. zu 20 μL resuspendierter Aktinlösung aus Schritt 2.1.4, 134 μL G-Puffer, 0,32 μL 0,2 mM ATP und 0,16 μL 1 M DTT hinzufügen). Für die fluoreszierende Markierung fügen Sie etwa 10% markiertes Aktin hinzu; Fügen Sie beispielsweise 5 μL ATTO-markiertes Aktin zu 40 μL verdünntem unmarkiertem Aktin hinzu. Lassen Sie es auf Eis unter gelegentlichem Mischen (Pipettieren) mindestens einige Tage bis eine Woche depolymerisieren, bevor die Proteinkonzentration durch Bradford-Assay wie in Schritt 3 beschrieben gemessen wird.
HINWEIS: Verdünntes unmarkiertes und fluoreszierendes Aktin auf Eis in einem kalten Raum oder Kühlschrank aufbewahren; Niemals einfrieren oder erwärmen lassen. Die Zubereitung wird im Laufe der Zeit weiter depolymerisieren und kann bei richtiger Handhabung für mindestens 6 Monate verwendet werden.
- Arp2/3-Komplex (Schweinehirn) (siehe Materialtabelle) gemäß den Anweisungen des Herstellers (20 μg Protein in 20 μL kaltemH2O) mit der Sequenz des Pulszentrifugierens, Mischens usw. auf Eis resuspendieren, wie für Aktin in Schritt 2.1 beschrieben. Kombinieren Sie die Proteinlösungen aus dem Resuspendieren von zwei Pulverröhrchen, um einen größeren Vorrat für reproduzierbare Experimente zu erhalten. 2 μL Aliquots zubereiten und bei -80 °C lagern.
- Resuspendieren Sie Profilin (human-rekombinant) (siehe Materialtabelle) in einer 4x höheren Konzentration als in den Anweisungen des Herstellers angegeben (100 μg Protein in 25 μL kaltemH2O), mit der Sequenz von Pulszentrifugieren, Mischen usw. auf Eis wie für Aktin in Schritt 2.1. Kombinieren Sie die Proteinlösungen aus dem Resuspendieren von zwei Pulverröhrchen, bevor Sie die Proteinkonzentration bestimmen, um einen größeren Vorrat für reproduzierbare Experimente zu erhalten.
HINWEIS: Auf Eis in einem kühlen Raum oder Kühlschrank aufbewahren; Niemals einfrieren oder erwärmen lassen. Bei richtiger Handhabung ist resuspendiertes Profilin mindestens 6 Monate bis 1 Jahr gut. - Resuspendierendes Capping-Protein (α1β2, humanes rekombinant) (siehe Materialtabelle) gemäß den Anweisungen des Herstellers (50 μg Protein in 50 μL kaltemH2O), mit der Sequenz von Pulszentrifugieren, Mischen usw. auf Eis, wie für Aktin in Schritt 2.1 beschrieben. Kühlen Sie 50 μL Glycerin auf Eis und fügen Sie die 50 μL resuspendiertes Capping-Protein hinzu; vorsichtig mischen. Bei -20 °C lagern.
HINWEIS: Die Lösung friert nicht ein und die Aktivität ist robust, so dass die Lösung bei sorgfältiger Handhabung monate- oder sogar jahrelang als Aliquot aufbewahrt werden kann. Das rekombinante Kappierprotein der Maus, das in der Vergangenheit am häufigsten in In-vitro-Experimenten verwendetwurde 13, wird bald kommerziell verfügbar sein. - Resuspendieren Sie Gelsolin (human-rekombinant, His-markiert) (siehe Tabelle der Materialien) gemäß den Anweisungen des Herstellers (20 μg Protein in 20 μL kaltemH2O), mit der Sequenz von Pulszentrifugieren, Mischen usw. auf Eis, wie für Aktin in Schritt 2.1 beschrieben. Etwa 2 μL Gelsolin werden pro Versuchstag verwendet; Bereiten Sie daher große Aliquots (5-10 μL) vor und lagern Sie sie bei -80 °C.
HINWEIS: Das Protokoll mit Gelsolin wird als Alternative bereitgestellt. Die Verwendung von Capping-Protein anstelle von Gelsolin wird empfohlen, entweder gekauft oder gereinigt wie in13. - Resuspendieren Sie VCA (human WASP-VCA, GST-tagged) (siehe Materialtabelle) in einer 2x höheren Konzentration als in der Herstelleranleitung angegeben (500 μg Protein in 250 μL kaltemH2O), mit der Sequenz von Pulszentrifugieren, Mischen usw. auf Eis wie für Aktin in Schritt 2.1. 10 μL Aliquots herstellen und bei -80 °C lagern.
HINWEIS: Sobald SpVCA (humanes pVCA, Streptavidin und His-markiert) kommerzialisiert ist oder wenn eine Proteinreinigung14 möglich ist, wird die Verwendung von SpVCA anstelle von VCA empfohlen. VCA gibt Kometen unter den hier beschriebenen Bedingungen nicht reproduzierbar ab.
3. Messung der Proteinkonzentrationen
- Konstruieren Sie eine Bradford-Standardkurve aus zwei überlappenden seriellen Verdünnungen von BSA.
HINWEIS: Die Standardkurve muss nur alle paar Monate (oder noch seltener) erstellt werden, solange sich das Spektralphotometer nicht ändert.- Platzieren Sie in Reihe 1 eines Mikroröhrchen-Racks Röhrchen für die BSA-Verdünnungsserie #1: vier 2-ml-Röhrchen, gefolgt von vier 1,5-ml-Röhrchen. Platzieren Sie in Reihe 3 des Racks Röhrchen für die BSA-Verdünnungsserie #2 wie für die Serie #1. Legen Sie in Reihe 5 des Racks ein 2-ml-Röhrchen für jede zu messende Probe zusammen mit zwei 1,5-ml-Röhrchen. Fügen Sie ein einzelnes 1,5-ml-Röhrchen in Reihe 5 für den Rohling hinzu.
- Messen Sie das Bradford-Reagenz (siehe Materialtabelle) in die 1,5-ml-Röhrchen aus.
- Füllen Sie ein 15-ml-konisches Röhrchen oben mit Bradford-Reagenz, um das Pipettieren (überschüssiges Produkt wird in die Lagerflasche im Kühlschrank zurückgeführt) auf Eis zu erleichtern. Nehmen Sie 200 μL Bradford-Reagenz auf und werfen Sie es zurück in das konische Röhrchen, um die Pipettenspitze zu benetzen. Da die Lösung viskos ist, pipetten Sie langsam, damit die Lösung vollständig in die Spitze eindringen und sie verlassen kann, ohne Blasen zu bilden.
- Mit der "vornassen" Spitze pipetten Sie langsam 200 μL Bradford-Reagenz in jedes der 1,5-ml-Röhrchen im Rack (vier für jede BSA-Verdünnung, eines für den Rohling und zwei für jede zu messende Probe). Tun Sie dies zuerst, damit sich das Bradford-Reagenz gründlich auf Raumtemperatur erwärmen kann, bevor es mit Proteinlösungen gemischt wird. Geben Sie den restlichen Inhalt des 15-ml-konischen Röhrchens in die Flasche zurück.
- Messen Sie H 2 O in die2ml Röhrchen aus. Fügen Sie in Reihe 1 des Racks 1.990 μLH2O zum ersten Röhrchen und 900 μL zu den drei anderen Röhrchen hinzu. Für Zeile 3 fügen Sie 1.992,5 μL zum ersten Röhrchen und 900 μL zu den drei anderen Röhrchen hinzu. In jedes Probenröhrchen in Reihe 5 werden 2.000 μLH2Ogegeben.
HINWEIS: Verwenden Sie für alle Volumina größer als 1.000 μL eine 1.000 μL Pipette, aber verabreichen Sie die volle Menge durch zweimaliges Pipettieren. Es ist wichtig, alles vorzubereiten, bevor Sie mit den nachfolgenden Schritten beginnen, um zu vermeiden, dass Proteine über längere Zeit in H2O stark verdünnt sind. - Zur Herstellung der BSA-Verdünnungsserie #1 mischen Sie 10 μL kalibriertes 2 mg/ml BSA (siehe Materialtabelle) in das Röhrchen mit 1.990 μLH2O, um eine 10 μg/ml Lösung herzustellen. Daraus werden drei serielle Verdünnungen (5 μg/ml, 2,5 μg/ml und 1,25 μg/ml BSA) hergestellt, indem 900 μL jeder Lösung in das nächste Röhrchen (mit 900 μLH2O) überführt werden.
- Zur Herstellung der BSA-Verdünnungsserie #2 werden 7,5 μL kalibriertes 2 mg/ml BSA in das Röhrchen mit 1.992,5 μLH2Ogemischt, um eine 7,5 μg/ml Lösung herzustellen. Daraus werden drei serielle Verdünnungen (3,75 μg/ml, 1,875 μg/ml und 0,9375 μg/ml BSA) hergestellt, indem 900 μL jeder Lösung in das nächste Röhrchen (mit 900 μLH2O) überführt werden.
- Mischen und Ablesen der Absorption, um die Standardkurve zu erzeugen. Geben Sie 800 μL H2O in das Bradford-Reagenzrohr für den Rohling und starten Sie den Timer. Mischen Sie so effizient wie möglich ohne Blasen 800 μL jedes BSA-Standards mit 200 μL Bradford-Reagenz in den vorbereiteten Röhrchen. Sobald alle Standards mit Bradford-Reagenz gemischt wurden (< 5 min), gießen Sie jeden Standard in eine Einwegküvette und lesen Sie die Absorption bei 600 nm im Spektralphotometer ab, nachdem Sie die Maschine ausgeblendet haben.
HINWEIS: Die gleiche Küvette kann verwendet werden, um die gesamte Serie zu lesen, wenn der am wenigsten konzentrierte Standard zuerst gelesen wird und die Küvette zwischen den Lesevorgängen gut entleert ist. Wiederholen Sie die Standardkurve, bis die lineare Anpassung einen R-Wert von mindestens 0,99 aufweist. Erst nachdem das Pipettieren und schonende Mischen gemeistert ist, fahren Sie mit dem Lesen der Proben fort.
- Messung der Konzentrationen von Aktin- und Aktin-bindenden Proteinen
- In den 2 ml Röhrchen mit 2.000 μL H2O (hergestellt in Schritt 3.1.3) wird vorsichtig Folgendes untermischt: je 2 μLArp2/3-Komplex und Profilin, 4 μL markiertes G-Aktin, je 5 μL Gelsolin und VCA und 8 μL Capping-Protein (humanrekombinant). Man nimmt sofort 800 μL der Lösung und mischt es mit dem bereits vorbereiteten Bradford-Reagenz und wiederholt dann, um für jede Probe zwei Messwerte innerhalb von 5%-10% voneinander zu erhalten. Ein größerer Unterschied weist auf ein Problem mit der Neuaufhängung oder Handhabung hin. Lesen Sie innerhalb weniger Minuten nach dem Mischen mit Bradford-Reagenz.
HINWEIS: Wenn Sie eine Standardkurve von einem anderen Tag verwenden, muss zusätzlich zu den Proben nur der Rohling aus Schritt 3.1.6 vorbereitet werden.
- In den 2 ml Röhrchen mit 2.000 μL H2O (hergestellt in Schritt 3.1.3) wird vorsichtig Folgendes untermischt: je 2 μLArp2/3-Komplex und Profilin, 4 μL markiertes G-Aktin, je 5 μL Gelsolin und VCA und 8 μL Capping-Protein (humanrekombinant). Man nimmt sofort 800 μL der Lösung und mischt es mit dem bereits vorbereiteten Bradford-Reagenz und wiederholt dann, um für jede Probe zwei Messwerte innerhalb von 5%-10% voneinander zu erhalten. Ein größerer Unterschied weist auf ein Problem mit der Neuaufhängung oder Handhabung hin. Lesen Sie innerhalb weniger Minuten nach dem Mischen mit Bradford-Reagenz.
- Berechnen Sie die Konzentrationen von Aktin- und Aktin-bindenden Proteinen unter Verwendung der Standardkurve und des Verdünnungsfaktors. Wiederholen Sie die Messung für jede neue Resuspension. Molekulargewichte zur Umwandlung von mg/ml-Messwerten, die mit dem Bradford-Assay in μM erhalten wurden, sind: Aktin 43 kD, Arp2/3-Komplex 224 kD, Profilin 15 kD, Gelsolin 95 kD, Capping-Protein 68 kD (human-rekombinant) oder 63,5 kD (Maus-rekombinant), VCA 43 kD und SpVCA 54 kD (Monomermolekulargewicht).
4. Beschichtung von Perlen
- Die Zentrifuge wird auf 4 °C vorgekühlt und der rührtrockene Block (siehe Materialtabelle) auf 18 °C eingestellt.
- Die Perlen waschen: 50 μL Xb-Puffer in ein 1,5-ml-Mikrozentrifugenröhrchen pipettieren, 9 μl Perlensuspension mit 4,5 μm Durchmesser oder 2 μl Perlensuspension mit 1 μm Durchmesser (2,5 % w/v-Suspension) hinzufügen (siehe Materialtabelle). Gründlich mischen und die Proben bei 20.000 x g 10 min bei 4 °C zentrifugieren.
HINWEIS: Die Gesamtperlenoberfläche beider Perlengrößen beträgt 3 cm2:
abgeleitet durch Berechnung der Anzahl der Perlen und dann ihrer Gesamtoberfläche unter Verwendung klassischer Gleichungen für das Volumen und die Oberfläche von Kugeln. Andere Perlengrößen können verwendet werden, wenn die Mengen angepasst werden, um die Gesamtoberfläche bei3 cm 2 konstant zu halten. - Beschichten Sie die Perlen: Entfernen Sie vorsichtig den Überstand, ohne die Perlen zu stören, und resuspendieren Sie das Perlenpellet in 40 μL 2 μM SpVCA (oder 7 μM VCA) in Xb-Puffer durch sanftes Pipettieren. Rühren bei 18 °C, 1.000 U/min für 20 min.
- Überzogene Perlen waschen: Die Mischung (20.000 x g für 10 min bei 4 °C) zentrifugieren und den Überstand vorsichtig entfernen. Die Beads werden in 50 μL kaltem Xb/1% BSA resuspendiert und bei 20.000 x g für 10 min bei 4 °C zentrifugiert. Entfernen Sie den Überstand und wiederholen Sie den Waschschritt 1x.
- Resuspendieren Sie das beschichtete Perlenpellet aus Schritt 4.4 in 120 μL kaltem Xb/1% BSA für beide Perlengrößen, so dass die Menge an Perlenoberfläche/μL Perlenlösung gleich ist. Auf Eis im Kühlschrank oder Kühlraum aufbewahren. Beschichtete Perlen funktionieren noch mindestens einige Wochen normal.
5. Vorbereitung der Motilitätsmischung und der Objektträger für die Beobachtung
HINWEIS: Das Gesamtvolumen der Motilitätsmischung beträgt 8,4 μL, um einen Abstand von etwa 25 μm zwischen dem Objektträger und dem 18 mm x 18 mm großen Deckglas zu ermöglichen, so dass Perlen aller Größen (bis zu 10 μm Durchmesser) nicht zusammengedrückt werden. Die grundlegende Motilitätsmischung beträgt etwa 5 μM G-Aktin (10% markiertes fluoreszierendes Aktin) mit 5 μM Profilin, 50 nM Arp2/3-Komplex und 25 nM Capping-Protein (oder 240 nM Gelsolin).
- Bereiten Sie die Motilitätsreaktionsmischung vor. Die genauen Mengen an Aktin (und damit Profilin) hängen von der in Schritt 3.3 berechneten Konzentration ab, aber eine repräsentative Reaktion ist wie folgt. Auf Eis mischen, in dieser Reihenfolge: 3,2 μL MB13, 1,5 μL Profilin bei 30 μM verdünnt in MB13, 1 μL Capping-Protein bei 0,21 μM verdünnt in MB13 oder Gelsolin verdünnt auf 2 μM in MB13, 1 μL Arp2/3-Komplex bei 0,47 μM verdünnt in MB13, 0,2 μL Perlensuspension (Wirbel kurz vor Gebrauch), und 1,5 μL Aktin bei 30 μM im G-Puffer. Gut aber schnell mischen und den Timer starten.
- Erkennen Sie die gesamte Motilitätsreaktionsmischung auf einem Objektträger. Mit einem 18 mm x 18 mm großen Deckglas abdecken und das Deckglas mit geschmolzenem VALAP mit einem kleinen Pinsel versiegeln. VALAP ist eine Mischung aus Lanolin, Paraffin und Vaseline (siehe Materialtabelle) 1:1:1 nach Gewicht, geschmolzen und gerührt.
6. Mikroskopische Beobachtung
- Beobachten Sie Motilitätsreaktionen sofort mit einem 100x-Objektiv entweder auf einem aufrechten oder einem inversen Mikroskop (siehe Materialtabelle), das mit Phasenkontrast- und/oder Epifluoreszenzmikroskopie (GFP-Würfel, siehe Materialtabelle) ausgestattet ist. Die Beobachtungen werden bei Raumtemperatur (23-25 °C) durchgeführt.
- Um durchschnittliche Verschiebungsgeschwindigkeiten für eine ganze Population von Perlen zu erhalten, zeichnen Sie Phasenkontrast oder fluoreszierende Standbilder im Laufe der Zeit auf, indem Sie das gesamte Dia scannen. Messen Sie die Kometenlänge von Hand und plotten Sie über die Zeit. Die Steigung der linearen Passung ist die durchschnittliche Wachstumsgeschwindigkeit.
- Um die Geschwindigkeit einzelner Perlen zu bewerten, sammeln Sie Zeitrafferfilme in der Phasenkontrastmikroskopie. Abhängig von der Perlengeschwindigkeit und der erforderlichen Auflösung nehmen Sie alle 1-10 s Bilder auf. Verwenden Sie das Tracking-Tool eines beliebigen Bildverarbeitungsprogramms, um Perlengeschwindigkeiten und Trajektorien zu erhalten.
Ergebnisse
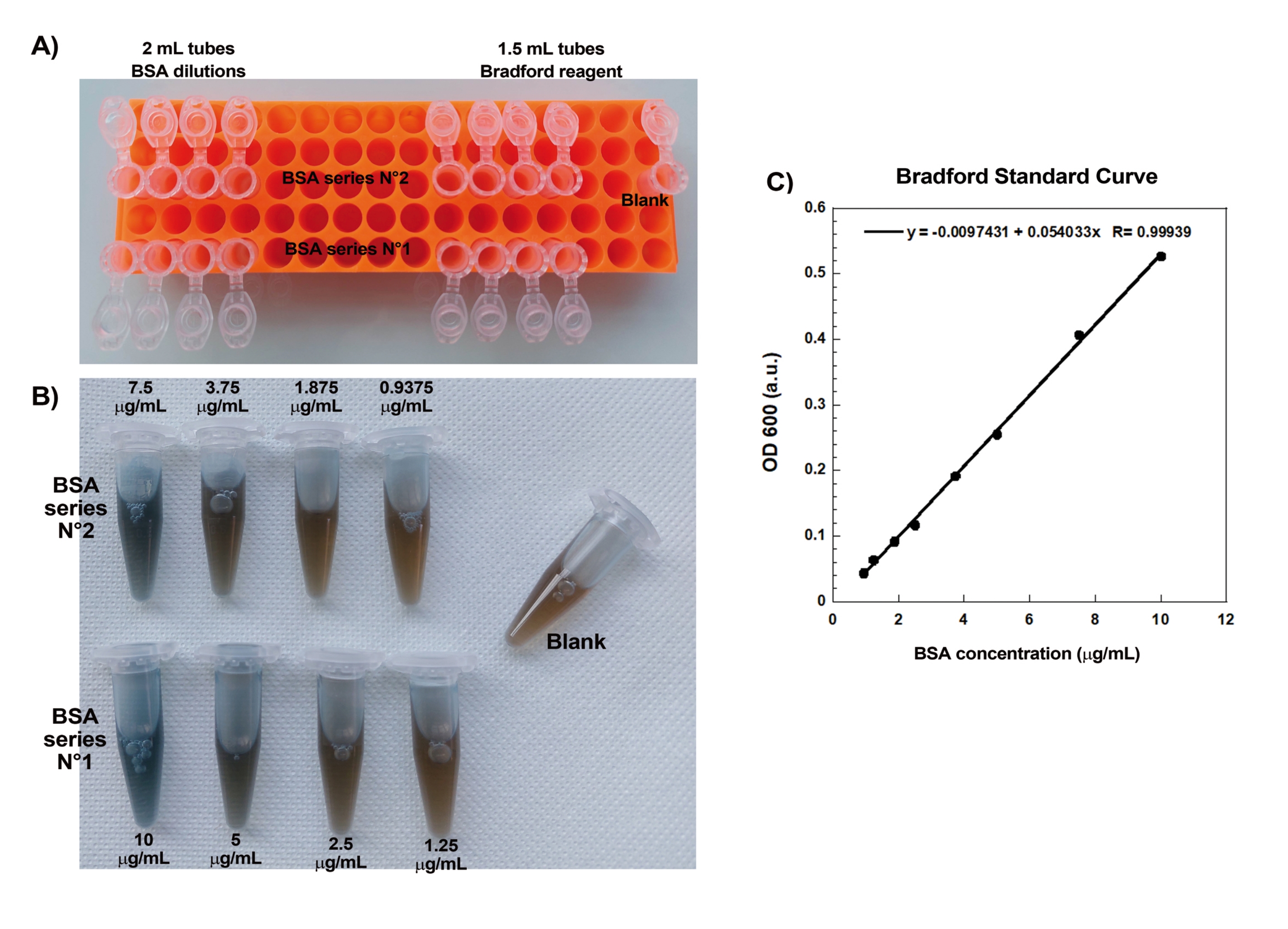
Einer der Schlüsselaspekte bei der reproduzierbaren Erzeugung von Aktinkometen auf Perlen ist das schonende und präzise Pipettieren empfindlicher Aktin-bindender Proteine. Das Generieren einer Bradford-Standardkurve ist eine gute Möglichkeit, die Pipettierfähigkeiten zu bewerten. Abbildung 1A,B zeigt die Rohre für die Standardkurve und ein Beispiel dafür, wie die beiden seriellen Verdünnungen von BSA aussehen, wenn sie mit Bradford-Reagenz gemischt werden. Beachten Sie den abgestuften blauen Farbton (höhere Proteinkonzentration ergibt eine blauere Lösung). Beim Ablesen im Spektralphotometer und Plotten ergeben diese Lösungen eine Standardkurve, wie in Abbildung 1C dargestellt. Um vorsichtiges Pipettieren zu üben, sollte der Assay wiederholt werden, bis der lineare Korrelationsfaktor 0,999 beträgt, wie gezeigt.
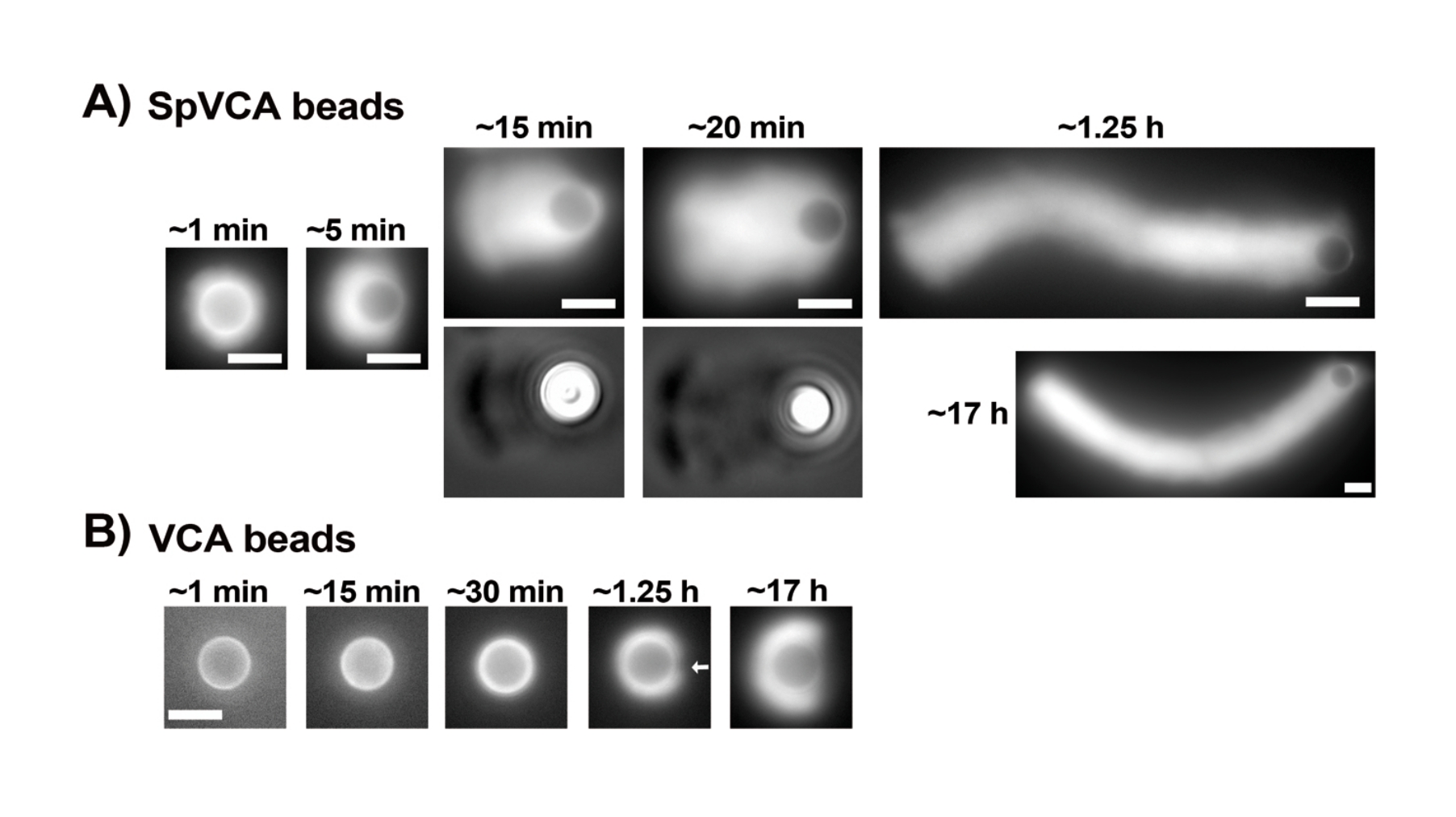
Sobald die Konzentrationen kommerziell resuspendierter Proteine sorgfältig mit dem Bradford-Assay bewertet wurden, werden beschichtete Beads und Motilitätsmischung hergestellt und miteinander gemischt. Abbildung 2A zeigt repräsentative Bilder der verschiedenen Stadien der Kometenentstehung: Aktinwolken bilden sich innerhalb von Minuten nach dem Mischen von SpVCA-beschichteten Kügelchen und Motilitätsmedium; Die Wolkenpolarisation tritt bei ~5 min und die Kometenproduktion bei 15-20 min auf. Die Aktinkometen, die sowohl mit Epifluoreszenz- als auch mit Phasenkontrastmikroskopie sichtbar sind (Abbildung 2A), verlängern sich noch viele Stunden, aber eine konsistente Geschwindigkeit wird nicht aufrechterhalten, so dass die Perlenmotilität normalerweise innerhalb von 1 h bewertet wird. Auf der anderen Seite dauert es 30 Minuten mit VCA-beschichteten Perlen, um helle Aktinwolken zu erhalten (Abbildung 2B), und es bilden sich keine Kometen, obwohl die Symmetrie bei 1-2 h zu brechen beginnt (Pfeil in Abbildung 2B) und Wolken nach nächtlicher Inkubation Polarisation zeigen.
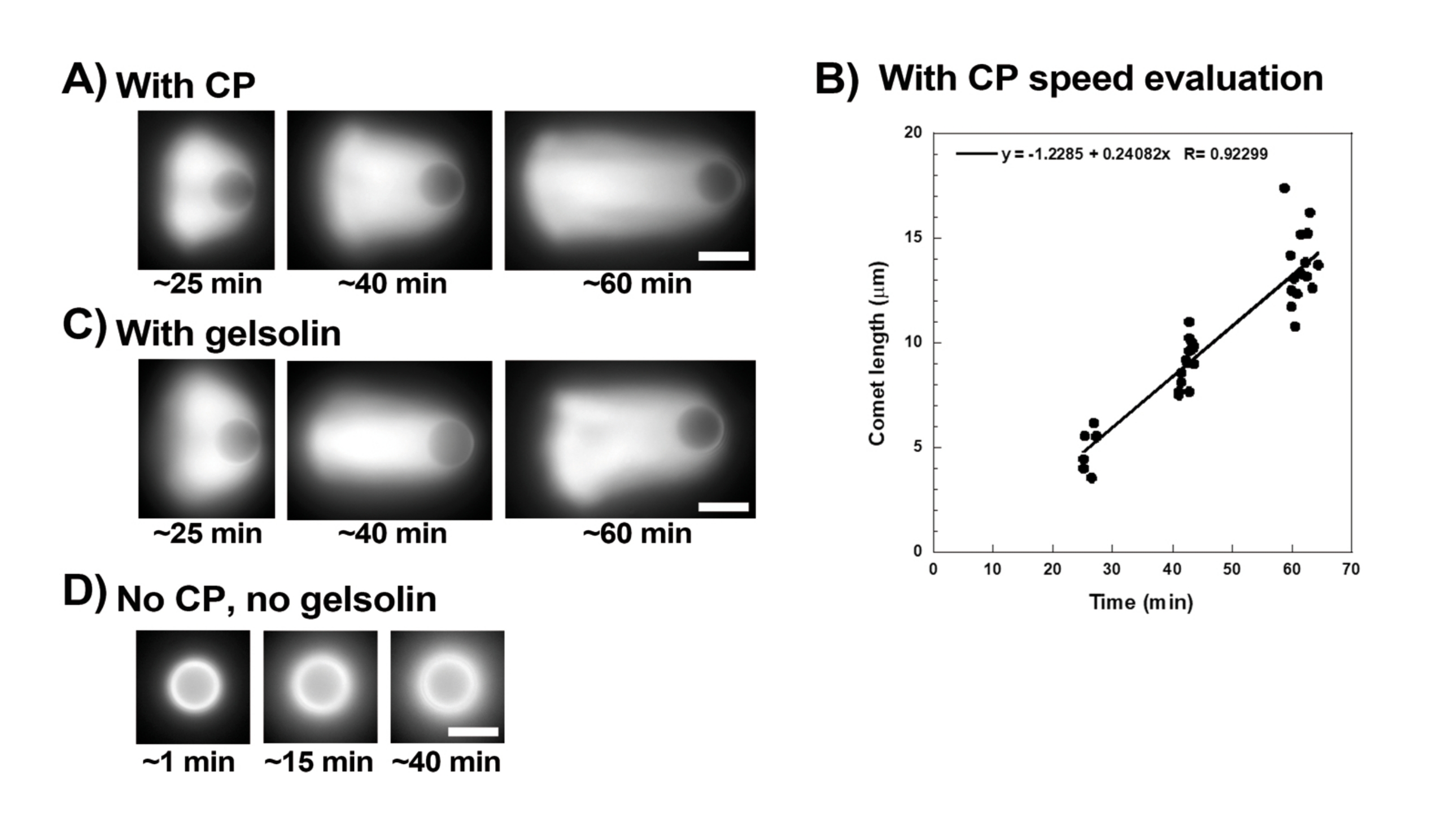
Abbildung 3 zeigt ein Beispiel für die Bewertung der Perlengeschwindigkeit in Gegenwart von Capping-Protein. Da alle Perlen ungefähr zur gleichen Zeit die Symmetrie brechen, werden "Pseudo-Zeitraffer"-Aufnahmen durchgeführt, bei denen das Dia gescannt und Bilder der gesamten Kometenpopulation im Laufe der Zeit aufgenommen werden (Abbildung 3A). Kometen depolymerisieren nicht; Daher kann die Zunahme der im Laufe der Zeit gemessenen Kometenlängen zur Berechnung der Verdrängungsgeschwindigkeit verwendet werden (Abbildung 3B). Gelsolin kann anstelle von Capping-Protein für die Kometenbildung verwendet werden, wenn 10x mehr Gelsolin hinzugefügt wird, um seine reduzierte Capping-Aktivität zu kompensieren. Kometen, die in Gegenwart von Gelsolin gebildet werden, sind qualitativ gleich und bewegen sich mit ungefähr der gleichen Geschwindigkeit wie Perlen mit Capping-Protein (Abbildung 3C). Die Verschließaktivität ist der Schlüssel zur Konzentration der Polymerisation an der Oberfläche der Perle, und wenn weder Capping-Protein noch Gelsolin in der Motilitätsmischung enthalten sind, polarisieren Aktinwolken niemals zu Kometen, obwohl sich helle Aktinwolken um die Perlen bilden (Abbildung 3D). Kometen auf Perlen können verwendet werden, um die Aktin-basierte Kraftproduktion in verschiedenen biochemischen Kontexten zu messen, indem die Motilitätsmischung verändert und das Motilitätsergebnis mit verschiedenen Mikromanipulationstechniken, zum Beispiel15, beobachtet wird.

Abbildung 1: Bradford-Standardkurve. (A) Abbildung der Einrichtung der Rohre für die Erstellung der Bradford-Standardkurve. Probenröhrchen werden nicht gezeigt. (B) Bild der beiden überlappenden BSA-Serienverdünnungen, sobald sie mit Bradford-Reagenz gemischt wurden. (C) Die Absorption bei 600 nm der unter (B) gezeigten Lösungen wird im Spektralphotometer gemessen und als Funktion der Proteinkonzentrationen der BSA-Lösungen aufgetragen. Die lineare Passung wird verwendet, um die Probenkonzentration zu berechnen. Der Korrelationsfaktor R der linearen Anpassung beträgt 0,999. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
{kind=link}

Abbildung 2: Kometenbildung auf SpVCA-beschichteten Perlen im Gegensatz zu VCA-beschichteten Perlen. (A) Repräsentative SpVCA-beschichtete Perlen (unterschiedliche Perlen in jedem Bild), die im Laufe der Zeit gezeigt werden. Die Zeit ab dem Zeitpunkt des Mischens wird angezeigt. Aktinwolken bilden sich sofort, und die Wolkenpolarisation ergibt Kometen, die sich stundenlang verlängern. (B) Repräsentative VCA-beschichtete Perlen (unterschiedliche Perlen in jedem Bild), die im Laufe der Zeit gezeigt werden. Mehr als 1 h ist notwendig, um die Anfänge der Aktinwolkenpolarisation (Pfeil) zu sehen, und selbst lange Inkubationen erzeugen keine Kometen. Alle Bilder sind von Perlen mit einem Durchmesser von 4,5 μm, Epifluoreszenzbildgebung von fluoreszierendem Aktin gepaart mit Phasenkontrastvisualisierung für die 15- und 20-Minuten-Zeitpunkte in (A), Maßstabsbalken = 5 μm. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
{kind=link}

Abbildung 3: Kometen- und Perlgeschwindigkeitsanalyse. (A) und (C) In Gegenwart von entweder Capping-Protein (CP) oder Gelsolin polarisieren Aktinwolken in den ersten 20 Minuten der Reaktion zu Kometen, und Kometen verlängern sich im Laufe der Zeit. Zeit vom Mischen für jedes Bild angegeben; Jedes Bild ist eine andere Perle. Es gibt eine gewisse Variabilität zwischen den Perlen und der Zubereitung, aber im Durchschnitt bewegen sich die Perlen unter den hier beschriebenen Standardbedingungen mit Mikron/Submikron pro Minute (0,2-1 μm/min). (B) Repräsentative Grafik, die die Bewertung der Kometenlänge (gesamte Perlenpopulation) im Zeitverlauf zeigt. Die Steigung der linearen Korrelation entspricht der mittleren Verdrängungsgeschwindigkeit, in diesem Fall 0,24 μm/min. (D) In Abwesenheit von Capping-Aktivität (kein Capping-Protein oder Gelsolin) bilden sich Aktinwolken um Perlen, aber Kometen bilden sich nicht. Alle Bilder sind von 4,5 μm Durchmesser Perlen, Epifluoreszenz-Bildgebung von fluoreszierendem Aktin, Maßstabsbalken = 5 μm. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
{kind=link}
Diskussion
Das hier beschriebene Protokoll beschreibt, wie Aktinnetzwerkwachstum auf Perlenoberflächen, Kometenbildung und Perlenmotilität mit kommerziell erhältlichen Proteinen erreicht werden können. Manchmal werden Kometen jedoch nicht reproduzierbar beobachtet oder sind inhomogen zwischen Objektträger und Deckglas. Die folgende Diskussion hebt einige wichtige Punkte im Protokoll hervor und schlägt einige Parameter vor, die angepasst werden können. Ein zu berücksichtigender Faktor ist, dass Kometenbildung und Perlengeschwindigkeit von der Temperatur beeinflusst werden, wobei Temperaturen weit über 25 ° C oder weit unter 23 ° C die Kometenbildung negativ beeinflussen und nicht reproduzierbare Daten liefern. Die Verwendung eines temperaturgesteuerten Mikroskops oder eines Mikroskops in einem klimatisierten Raum wird dringend empfohlen. Obwohl fluoreszenzmarkiertes Aktin oft in der Motilitätsmischung enthalten ist, um Kometen durch Fluoreszenzmikroskopie zu beobachten, sind Kometen, sobald sie mehr als einen Perlendurchmesser lang sind, auch durch Phasenkontrastmikroskopie als dunkler Abstrich neben der Perle sichtbar. Die Phasenkontrastvisualisierung ist besser für die Zeitraffer-Bildgebung geeignet, da eine gewisse Phototoxizität mit der Fluoreszenzbildgebung auch über die rotierende Scheibe verbunden ist. Da sich Perlen im Laufe der Zeit absetzen, erzeugt ein inverses Mikroskop weniger horizontale Perlendrift als ein aufrechtes und eignet sich besser für Filme. Die Verwendung von geschmolzenem VALAP zum Versiegeln von Objektträgern ist wichtig, da Substanzen wie Nagellack die Kometenbildung stören. Große Mengen VALAP können in einem Becherglas hergestellt und dann herausgeschöpft werden, um kleinere Becher nachzufüllen, die für ein schnelles Schmelzen besser geeignet sind. VALAP ist jahrelang bei Raumtemperatur gut.
Ein weiterer wichtiger technischer Aspekt ist die sorgfältige Vorbereitung von Puffer- und Motilitätsmischungen. Bei der Zubereitung von MB13 ist Vorsicht geboten, insbesondere bei der pH-Einstellung. Der pH-Wert von MB13 sollte schnell auf neutral mit NaOH eingestellt werden, um eine ATP-Hydrolyse zu vermeiden, aber nicht zu schnell, da die EGTA löslich wird, wenn sich der pH-Wert neutral nähert. EGTA ist ein Schlüsselbestandteil, da es das an Aktin gebundene Kalzium komplexiert und in der Motilitätsmischung die aktivere Magnesiumform16 ergibt. MB13, das zu schnell oder zu langsam vorbereitet wird, führt zu einer suboptimalen Kometenbildung oder sogar gar keiner Kometenbildung. Ein weiterer wichtiger Punkt ist, die KCl-Konzentration in der Motilitätsmischung sorgfältig zu verfolgen, wenn man mit Bedingungen spielt. Wenn beispielsweise 1x MB13 in der Reaktionsmischung verwendet und Profilin, Capping-Protein und der Arp2/3-Komplex in MB13 verdünnt werden, beträgt die endgültige KCl-Konzentration in der Motilitätsreaktion etwa 40-50 mM aufgrund der Verdünnung durch G-Puffer. Diese Konzentration liefert die besten Ergebnisse im Kometenassay, und mehr als 60 mM KCl verringert die Arp2/3-Komplexkeimbildungsaktivität.
Auf der Proteinseite ist ein kritischer technischer Aspekt bei der Gewinnung von Aktinkometen der richtige Umgang mit kommerziellen Aktin-bindenden Proteinen, insbesondere das präzise Pipettieren von Mikrolitermengen. Die Linearität der Bradford-Standardkurve ist ein guter Test für das Pipettieren und die Kurve kann dann für Routinemessungen der Proteinkonzentrationen verwendet werden. In der Tat ist es bei der Verwendung resuspendierter kommerzieller Proteine für das Kometenverfahren wichtig, die Proteinkonzentrationen immer zu überprüfen, da Chargenvariabilität und Benutzerfehler während der Resuspension zu Unterschieden zwischen tatsächlichen und erwarteten Konzentrationen führen können. Manchmal können kleine Unterschiede in der Proteinkonzentration dazu führen, dass Kometen komplett fehlen.
Ein weiterer wichtiger Aspekt des hier vorgestellten Verfahrens ist die Verwendung von profilinkomplexiertem G-Aktin als Brennstoff für die Polymerisation. Historisch gesehen verwendeten In-vitro-Systeme vorpolymerisiertes filamentöses Aktin (F-Aktin) als Aktinquelle: Depolymerisation in der Bulk-Feed-Polymerisation auf der Oberfläche10,17. Dies hatte den Vorteil, den G-Aktinspiegel zu kontrollieren, fügte jedoch eine Komplexitätsschicht hinzu, die zusätzliche Komponenten benötigte, um die Depolymerisation zu katalysieren. Da der Umsatz des Aktinnetzwerks für die Krafterzeugung und Motilität, die durch Keimbildung und Polymerisation an der Oberfläche der Perle angetrieben werden, nicht notwendig ist, während Aktin-Depolymerisationsfaktoren wie ADF / Cofilin auf die gealterten Netzwerke weit von der Oberfläche wirken18, erfolgt die meiste In-vitro-Rekonstitution der Aktin-basierten Motilität jetzt der Einfachheit halber ohne Umsatz. Es gibt jedoch einige Nachteile bei der Verwendung von G-Actin. Erstens sind bei der Verwendung von kommerziellem Aktin, das lyophilisiert wurde, Oligomere vorhanden. Die hier beschriebenen Depolymerisationsschritte sind sehr wichtig, um reproduzierbare Ergebnisse zu erhalten. Obwohl der G-Puffer traditionell auf pH 8 eingestellt ist, scheint ein niedrigerer pH-Wert (z. B. pH 7) in den in diesem Artikel beschriebenen Assays besser zu funktionieren, möglicherweise weil ein niedriger pH-Wert die Depolymerisation verstärkt19. Ein weiterer Nachteil der Verwendung von G-Aktin besteht darin, dass nach der Platzierung unter Salzbedingungen, die für die Polymerisation freizügig sind, spontane Keimbildung auftritt und sich F-Aktin sowohl in der Masse als auch auf der Perlenoberfläche bildet. Die Komplexierung von G-Aktin mit Profilin unterdrückt die spontane Keimbildung in der Bulk- und Point-End-Polymerisation, wodurch sowohl die Keimbildung als auch die Stachelendpolymerisation an der Oberflächefokussiert werden 20. Profilin-G-Aktin ist physiologisch relevant, da ein Großteil des Aktins in der Zelle in dieser Form vorliegt21. Hier wird ein 1:1-Verhältnis von Profilin:Aktin verwendet; höhere Verhältnisse (z.B. 3:1) hemmen jedoch die Polymerisation in der Masse gründlicher, obwohl höhere Verhältnisse auch den Arp2/3-Komplex und die Stachelenddehnung bis zu einem gewissen Grad hemmen22,23.
Die Capping-Aktivität ist auch für die Kometenbildung von entscheidender Bedeutung, da sie die Insertion von neuem Aktin an der Oberfläche über Keimbildungszyklen durch den oberflächenaktivierten Arp2/3-Komplex24,25 sicherstellt. Ohne Kappung brechen Aktinwolken die Symmetrie nicht, um Kometen zu bilden, da die Polymerisation an der Oberfläche nicht genug Spannung aufbaut, um die Wolke aufzubrechen26. In der Vergangenheit haben wir das zu Hause gereinigte rekombinante Mauskappenprotein13 verwendet, aber Tests, die für diesen Artikel durchgeführt wurden, zeigen, dass kommerziell erhältliches rekombinantes humanes Capping-Protein ebenso wirksam ist wie kommerziell erhältliches Gelsolin, obwohl 10x mehr Gelsolin verwendet werden muss, und für bestimmte Anwendungen ist es möglicherweise nicht geeignet, da es sowohl Aktin-Trennaktivität als auch Capping27 hat.
Schließlich liegt die Robustheit dieser Methode in der Verwendung eines sehr aktiven Arp2/3-Komplexaktivators, Streptavidin-pVCA (SpVCA)28. SpVCA umfasst die Profilin-G-Aktin-Bindungsdomäne von WASP (die p-Domäne) zusätzlich zur komplexen Arp2/3-Bindungsdomäne, da sich diese unter Profilin-G-Aktinbedingungen als am effizientesten erweist29. Noch wichtiger ist, dass die Verwendung des Streptavidin-Tags, das ursprünglich eingeführt wurde, um die Oberflächenfunktionalisierung über die Biotin-Streptavidin-Verbindung zu ermöglichen, den zusätzlichen Effekt hat, die Aktivierung des Arp2/3-Komplexes zu erhöhen, vermutlich aufgrund der Tatsache, dass Streptavidin ein Tetramer ist und somit den Aktivator clustert, von dem bekannt ist, dass er die Aktivität des Arp2/3-Komplexes erhöht30 . Das kommerziell produzierte SpVCA befindet sich derzeit in der Entwicklung und wird in Kürze zum Kauf angeboten. Es sollte ferner beachtet werden, dass, obwohl 40 μL 2 μM SpVCA routinemäßig verwendet werden, um 3 cm2 der Perlenoberfläche zu beschichten, andere Beschichtungskonzentrationen (höher und niedriger) ebenfalls funktionieren, und das Spielen mit diesen Bedingungen führt zu unterschiedlichen Kometenwachstumsgeschwindigkeiten und Morphologien. Wenn sich Kometen nicht bilden oder die Kometengröße auf dem Objektträger nicht homogen ist, sollten unterschiedliche Beschichtungsbedingungen sowie unterschiedliche KCl- und Profilinkonzentrationen in der Motilitätsmischung getestet werden. Die Konzentrationen von Aktin, Arp2/3-Komplex und Capping-Protein in der Motilitätsmischung können ebenfalls verändert werden, um die Kometenbildung zu optimieren, aber in unseren Händen führt die Änderung dieser Anteile oft zu verwirrenden Ergebnissen.
Zusammenfassend lässt sich sagen, dass die hier beschriebenen Verfahren Aktinanordnung auf Perlenoberflächen und Motilität erzeugen, aber jede Oberfläche, die mit SpVCA funktionalisiert werden kann, kann verwendet werden. In Fällen, in denen die Adsorption wie hier beschrieben nicht funktioniert, kann der Streptavidin-Anteil verwendet werden, um SpVCA nach der Biotinylierung an die interessierende Oberfläche zu binden. Die so gebildeten Aktinstrukturen, Kometen oder andere, können zum Testen verschiedener biochemischer und biophysikalischer Aspekte von Aktinnetzwerken verwendet werden und eignen sich besonders für physikalische Manipulationen mit Mikropipetten, optischen Pinzetten und Laserablationen 15,26,31,32. Zusätzlich zu seiner Verwendung für die Forschungsgemeinschaft eignet sich der hier beschriebene Ansatz als Lehrmittel für Studenten der Biophysik, um Konzepte der aktiven Materie wie Symmetriebrechung und Selbstorganisation zu studieren.
Offenlegungen
Die Autoren erklären, dass sie keine Interessenkonflikte mit dem Inhalt dieses Artikels haben.
Danksagungen
Wir danken den Mitgliedern unseres neuen Zuhauses bei LPENS herzlich für ihren herzlichen Empfang und insbesondere dem ABCDJ-Team für all ihre Hilfe und Unterstützung. J.P. dankt der finanziellen Unterstützung durch die Foundation ARC (Grant PJA 20191209604) und C.S. für die finanzielle Unterstützung durch die Human Frontiers Science Program Organization (Grant RGP0026/2020).
Materialien
| Name | Company | Catalog Number | Comments |
| Actin, rabbit muscle, Alexa Fluor 488 conjugate | Invitrogen (ThermoFisher Scientific) | A12373 (recently discontinued) | This product can be replaced with ATTO-488 actin from Hypermol. |
| Actin, rabbit muscle, ATTO-488 | Hypermol | 8153 | |
| Actin, rabbit skeletal muscle | Cytoskeleton | AKL99 | |
| Arp2/3 complex | Cytoskeleton | RP01P | |
| ATP | Sigma | A7699 | |
| BioSpectrometer, basic | Eppendorf | 035739 | |
| Bradford Reagent | Bio-Rad | 500-0006 | |
| BSA, high quality | Sigma | A3059 | |
| BSA standard 2 mg/mL (Pierce) | Thermo Scientific | 23209 | |
| Capping protein (a1b2, mouse recombinant) | Home-purified (Reference 13) | This product will soon be commercially available from Cytoskeleton. | |
| Capping protein (a1b2, human recombinant) | Hypermol | 8322 | |
| Cube, GFP: U-MNIBA3 or U-MWB2 | Olympus | discontinued | Any GFP cube, adapted to the microscope being used, can be used. |
| Dry block, agitating: ThermoMixer C (refrigerated) | Eppendorf | 035963 | |
| ** with SmartBlock, 24 microtubes 2 mL | Eppendorf | 035969 | |
| Gelsolin (human recombinant, His-tagged) | Cytoskeleton | HPG6 | |
| Lanolin | Sigma | 49909 | |
| Microcentrifuge 5427R + rotor | Eppendorf | 934126 | |
| Microscope, upright: BX51 | Olympus | discontinued | Any epifluorescence upright microscope equipped with phase contrast optics can be used. |
| Microscope, inverted: IX70 | Olympus | discontinued | Any epifluorescence inverted microscope equipped with phase contrast optics can be used. |
| Paraffin | Sigma | 76244 | |
| Petroleum jelly: Vaseline | Sigma | 16415 | |
| Pipettes Research Plus | Eppendorf | Gilson pipettes don't work as well for delivery of very small volumes (0.5 µL for example). | |
| **10 µL | 933954 | ||
| **2.5 µL | 933953 | These two sizes are essential, but the use of high-quality pipettes (a full Research Plus set for example) is recommended. | |
| Polystyrene carboxylate beads | Polysciences | ||
| **approx. 1 µm diameter | 08226 | ||
| **approx. 4.5 µm diameter | 17140-5 | ||
| Profilin 1 (human recombinant, untagged) | Cytoskeleton | PR02 | |
| SpVCA (human WASP pVCA domain, N-ter His-tag, C-ter Streptavidin tag) | Home-purified (Reference 14) | This product will soon be commercially available from Cytoskeleton. | |
| VCA (human WASP VCA domain, GST-tagged) | Cytoskeleton | VCG03 |
Referenzen
- Campellone, K. G., Welch, M. D. A nucleator arms race: cellular control of actin assembly. Nature Reviews Molecular Cell Biology. 11 (4), 237-251 (2010).
- Pollard, T. D. Rate constants for the reactions of ATP- and ADP-actin with the ends of actin filaments. Journal of Cell Biology. 103, 2747-2754 (1986).
- Blanchoin, L., Boujemaa-Paterski, R., Sykes, C., Plastino, J. Actin dynamics, architecture and mechanics in cell motility. Physiological Reviews. 94 (1), 235-263 (2014).
- Tilney, L. G., Tilney, M. S. The wily ways of a parasite: induction of actin assembly by Listeria. Trends in Microbiology. 1 (1), 25-31 (1993).
- Moore, A. S., et al. Actin cables and comet tails organize mitochondrial networks in mitosis. Nature. 591 (7851), 659-664 (2021).
- Taunton, J., et al. Actin-dependent propulsion of endosomes and lysosomes by recruitment of N-WASP. Journal of Cell Biology. 148 (3), 519-530 (2000).
- Velarde, N., Gunsalus, K. C., Piano, F. Diverse roles of actin in C. elegans early embryogenesis. BMC Developmental Biology. 7, 142(2007).
- Merrifield, C. J., et al. Endocytic vesicles move at the tips of actin tails in cultured mast cells. Nature Cell Biology. 1 (1), 72-74 (1999).
- Samarin, S., et al. How VASP enhances actin-based motility. Journal of Cell Biology. 163 (1), 131-142 (2003).
- Bernheim-Groswasser, A., Wiesner, S., Golsteyn, R. M., Carlier, M. -F., Sykes, C. The dynamics of actin-based motility depend on surface parameters. Nature. 417 (6886), 308-311 (2002).
- Boujemaa-Paterski, R., et al. Network heterogeneity regulates steering in actin-based motility. Nature Communications. 8 (1), 655(2017).
- Akin, O., Mullins, R. D. Capping protein increases the rate of actin-based motility by promoting filament nucleation by the Arp2/3 complex. Cell. 133 (5), 841-851 (2008).
- Palmgren, S., Ojala, P. J., Wear, M. A., Cooper, J. A., Lappalainen, P. Interactions with PIP2, ADP-actin monomers, and capping protein regulate the activity and localization of yeast twinfilin. Journal of Cell Biology. 155 (2), 251-260 (2001).
- Carvalho, K., et al. Actin polymerization or myosin contraction: two ways to build up cortical tension for symmetry breaking. Philosophical Transactions of the Royal Society B. 368 (1629), 20130005(2013).
- Marcy, Y., Prost, J., Carlier, M. -F., Sykes, C. Forces generated during actin-based propulsion: a direct measurement by micromanipulation. Proceedings of the National Academy of Sciences of the United States of America. 101 (16), 5992-5997 (2004).
- Carlier, M. -F. Actin: protein structure and filament dynamics. Journal of Biological Chemistry. 266 (1), 1-4 (1991).
- Loisel, T. P., Boujemaa, R., Pantaloni, D., Carlier, M. F. Reconstitution of actin-based motility of Listeria and Shigella using pure proteins. Nature. 401 (6753), 613-616 (1999).
- Reymann, A. -C., et al. Turnover of branched actin filament networks by stochastic fragmentation with ADF/cofilin. Molecular Biology of the Cell. 22 (14), 2541-2550 (2011).
- Wioland, H., Jegou, A., Romet-Lemonne, G. Quantitative variations with pH of Actin Depolymerizing Factor/Cofilin's multiple actions on actin filaments. Biochemistry. 58 (1), 40-47 (2019).
- Plastino, J., Blanchoin, L. Dynamic stability of the actin ecosystem. Journal of Cell Science. 132 (4), 219832(2019).
- Pollard, T. D., Blanchoin, L., Mullins, R. D. Molecular mechanisms controlling actin filament dynamics in nonmuscle cells. Annual Review of Biophysics and Biomolecular Structure. 29, 545-576 (2000).
- Suarez, C., et al. Profilin regulates F-actin network homeostasis by favoring formin over Arp2/3 complex. Developmental Cell. 32 (1), 43-53 (2015).
- Courtemanche, N., Pollard, T. D. Interaction of profilin with the barbed end of actin filaments. Biochemistry. 52 (37), 6456-6466 (2013).
- Achard, V., et al. A "primer"-based mechanism underlies branched actin filament network formation and motility. Current Biology. 20 (5), 423-428 (2010).
- Sykes, C., Plastino, J. Actin filaments up against a wall. Nature. 464 (7287), 365-366 (2010).
- vander Gucht, J., Paluch, E., Plastino, J., Sykes, C. Stress release drives symmetry breaking for actin-based movement. Proceedings of the National Academy of Sciences of the United States of America. 102 (22), 7847-7852 (2005).
- McGough, A. M., Staiger, C. J., Min, J. -K., Simonetti, K. D. The gelsolin family of actin regulatory proteins: modular structures, versatile functions. FEBS Letters. 552 (2-3), 75-81 (2003).
- Abou-Ghali, M., et al. Capping protein is not necessary for polarized actin network growth and actin based motility. Journal of Biological Chemistry. 295, 15366-15375 (2020).
- Yarar, D., D'Alessio, J. A., Jeng, R. L., Welch, M. D. Motility determinants in WASP family proteins. Molecular Biology of the Cell. 13 (11), 4045-4059 (2002).
- Padrick, S. B., et al. Hierarchical regulation of WASP/WAVE proteins. Molecular Cell. 32 (3), 426-438 (2008).
- Bussonier, M., et al. Mechanical detection of a long-range actin network emanating from a biomimetic cortex. Biophysical Journal. 107 (4), 854-862 (2014).
- Paluch, E., vander Gucht, J., Joanny, J. -F., Sykes, C. Deformations in actin comets from rocketing beads. Biophysical Journal. 91 (8), 3113-3122 (2006).
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenWeitere Artikel entdecken
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten