
Method Article
Ricostituzione della motilità a base di actina con proteine disponibili in commercio
In questo articolo
Riepilogo
Questo protocollo descrive come produrre comete di actina sulle superfici delle perle utilizzando ingredienti proteici disponibili in commercio. Tali sistemi imitano le strutture protrusive presenti nelle cellule e possono essere utilizzati per esaminare i meccanismi fisiologici della produzione di forza in modo semplificato.
Abstract
Molti movimenti cellulari e cambiamenti di forma e alcuni tipi di motilità batterica e organelli intracellulare sono guidati dall'actina biopolimero che forma una rete dinamica sulla superficie della cellula, dell'organello o del batterio. Le basi biochimiche e meccaniche della produzione di forza durante questo processo possono essere studiate riproducendo il movimento basato sull'actina in modo acellulare su superfici inerti come perline che vengono funzionalizzate e incubate con un insieme controllato di componenti. Nelle condizioni appropriate, una rete elastica di actina si assembla sulla superficie del tallone e si apre a causa dello stress generato dalla crescita della rete, formando una "cometa di actina" che spinge la perla in avanti. Tuttavia, tali esperimenti richiedono la purificazione di una serie di diverse proteine leganti l'actina, spesso mettendole al di fuori della portata dei non specialisti. Questo articolo descrive un protocollo per ottenere in modo riproducibile comete di actina e motilità di perle utilizzando reagenti disponibili in commercio. Il rivestimento delle perline, la dimensione del tallone e la miscela di motilità possono essere modificati per osservare l'effetto sulla velocità del tallone, sulle traiettorie e su altri parametri. Questo test può essere utilizzato per testare le attività biochimiche di diverse proteine leganti l'actina e per eseguire misurazioni fisiche quantitative che fanno luce sulle proprietà della materia attiva delle reti di actina. Questo sarà uno strumento utile per la comunità, consentendo lo studio della motilità in vitro a base di actina senza conoscenze specialistiche nella purificazione delle proteine leganti l'actina.
Introduzione
La polimerizzazione dell'actina nelle cellule è controllata spazialmente e temporalmente da una stretta regolazione della nucleazione del filamento di actina a valle della segnalazione cellulare1. La nucleazione avviene attraverso la formazione di un trimero di actina, e quindi entrambe le estremità del filamento nascente polimerizzano spontaneamente, sebbene un'estremità sia più dinamica (l'estremità spinata) dell'altra (l'estremità appuntita)2. Quando la nucleazione e la polimerizzazione dell'estremità spinata sono dirette verso una superficie, producono una forza sufficiente (nell'intervallo pico-nano Newton) per spingere fuori la membrana cellulare per il movimento e per spostare oggetti di dimensioni micron all'interno della cellula con l'idrolisi ATP come fonte di energia3. Alcuni esempi includono i batteri Listeria monocytogenes che usano comete di actina per diffondersi da cellula a cellula e mitocondri, dove il movimento basato sulla cometa actina è importante per l'ereditarietà randomizzata durante la mitosi 4,5. Le comete di actina sugli endosomi e altre vescicole intracellulari sono implicate nel distacco dalle membrane donatrici 6,7,8.
Con il metodo qui presentato, gli aspetti di segnalazione della polimerizzazione cellulare dell'actina vengono bypassati e la polimerizzazione dell'actina viene prodotta su perle di polistirene micrometriche rivestendole con attivatori della nucleazione di actina ramificata, in particolare il dominio attivo della proteina WASP umana, VCA (chiamato anche WA o WCA)1. Le perle rivestite vengono quindi incubate in una miscela contenente gli ingredienti necessari per la polimerizzazione dell'actina, incluso il principale nucleatore di polimerizzazione dell'actina nelle cellule, il complesso Arp2/3, che viene attivato da VCA sulla superficie del cordone per formare nuovi filamenti come rami dai lati dei filamenti figli1. L'actina inizialmente polimerizza uniformemente attorno alla perla, ma poi rompe spontaneamente la simmetria per creare una cometa di actina che spinge la perla in avanti, ricreando così reti protrusive simili a cellule e comete in modo controllato. Approcci simili con perline e altre superfici rivestite sono stati usati in passato da noi e da altri per studiare la biochimica e la biofisica della polimerizzazione dell'actina 9,10,11,12, ma per questi esperimenti era necessaria una vasta esperienza nelle proteine leganti l'actina. Il protocollo qui presentato descrive come creare in modo robusto comete di actina e motilità interamente con reagenti disponibili in commercio (o presto disponibili), rendendo questo approccio accessibile a chiunque, anche in un contesto educativo per l'insegnamento di concetti biofisici. Le caratteristiche principali includono l'importanza di un pipettaggio delicato e affidabile, l'uso di monomeri complessati con profilina come fonte di actina e l'essenzialità dell'utilizzo di un attivatore complesso Apr2/3 altamente attivo come reagente di rivestimento delle perle.
Protocollo
1. Preparazione dei buffer
NOTA: utilizzare H2O ultrapuro per tutti i buffer. Non deve essere sterile. Filtrare tutte le soluzioni descritte nei punti 1.1-1.4 con un filtro a siringa da 0,2 μm, aliquote in porzioni da 500 μL-2 mL per tubo a seconda dell'uso, e conservare a -20 °C.
- Preparare il 10% di BSA pesando 2 g di BSA in un tubo conico da 50 mL e riempiendo fino al segno di 20 mL con H2O. Mescolare fino a quando la BSA è sciolta (circa 30 min), quindi portare il volume a 20 mL.
NOTA: Utilizzare BSA di alta qualità (vedere la tabella dei materiali). La soluzione di BSA al 10% viene utilizzata sia nella preparazione delle perle che nella miscela di motilità. - Per la preparazione delle perle, preparare il tampone Xb (10 mM HEPES, 0,1 M KCl, 1 mM MgCl 2 e 0,1 mM CaCl 2, pH 7,5) come soluzione 10x e diluire prima dell'uso (100 μL di soluzione madre 10x Xb + 900 μL di H2 O). Preparare Xb/1% BSA miscelando 100 μL di soluzione madre 10x Xb + 100 μL di BSA al 10% + 800 μL di H2O.
- Preparare G-buffer (2 mM Tris, 0,2 mM CaCl 2, 0,2 mM DTT,2 mM ATP), che è il tampone utilizzato per diluire l'actina monomerica (G-actina). Regolare a pH 7, non a pH 8 come tradizionalmente usato (vedi discussione).
- Preparare il tampone di motilità MB13 (10 mM HEPES, 1,5 mM ATP, 3 mM DTT, 1,5 mM MgCl2, 1 mM EGTA, 50 mM KCl, 1% BSA, pH 7,5). Per alcune applicazioni, 10x MB13 è utile. Tuttavia, preparare 10x MB13 senza BSA, poiché porta a problemi durante la regolazione del pH. Aggiungere BSA dalla soluzione madre al 10% (preparata al punto 1.1) quando si ricostituisce 1x MB13 da 10x MB13.
2. Preparazione di soluzioni proteiche
NOTA: Utilizzare H2O ultrapuro per tutte le risospensioni. Non deve essere sterile. Manipolare tutte le proteine su ghiaccio e aliquote in tubi pre-refrigerati. Manipolare delicatamente in modo da non produrre bolle e mai soluzioni proteiche a vortice. Per lo stoccaggio delle scorte a -80°C, non è necessario il congelamento istantaneo in azoto liquido. Adattare le dimensioni delle aliquote per evitare più di cinque cicli di congelamento-scongelamento, poiché ciò non sembra influenzare l'attività di nessuna delle proteine. Le aliquote di lavoro possono essere conservate a -20 °C per alcune settimane.
- Preparare la soluzione di G-actina (muscolo scheletrico di coniglio) come descritto di seguito.
- Centrifugare a impulsi la polvere di actina (vedi tabella dei materiali) a 4 °C per raccogliere il solido sul fondo del tubo.
- Aggiungere H 2 O secondo le istruzioni del produttore (1 mg di proteine in 100 μL di H 2 O freddo per actinanon marcata, 100 μg di proteine in 100 μL di H2O freddo per actina marcata con ATTO).
- Lasciare riposare sul ghiaccio per almeno 15 minuti. Mescolare delicatamente pipettando su e giù, lasciare riposare sul ghiaccio almeno altri 15 minuti e mescolare di nuovo. Centrifugare a impulsi a 4 °C per raccogliere la soluzione sul fondo del tubo e rimescolare.
- Preparare 10-50 μL di aliquote di actina non marcata a seconda dell'uso e 20 μL di aliquote di actina marcata con ATTO. Conservare le aliquote a -80 °C.
- Per depolimerizzare gli oligomeri di actina che si formano durante la liofilizzazione e il congelamento, diluire un'aliquota di actina risospesa ~8 volte in G-buffer con aggiunta di ATP e DTT (ad esempio, a 20 μL di soluzione di actina risospesa dal punto 2.1.4, aggiungere 134 μL di G-buffer, 0,32 μL di 0,2 mM ATP e 0,16 μL di 1 M DTT). Per l'etichettatura fluorescente, aggiungere circa il 10% di actina marcata; ad esempio, aggiungere 5 μL di actina marcata con ATTO a 40 μL di actina diluita non marcata. Lasciare depolimerizzare su ghiaccio con miscelazione occasionale (pipettaggio) almeno da alcuni giorni a una settimana prima di misurare la concentrazione proteica mediante il saggio Bradford come descritto al punto 3.
NOTA: Conservare l'actina diluita senza etichetta e fluorescente su ghiaccio in una cella frigorifera o in frigorifero; Non congelare o lasciare mai scaldare. La preparazione continuerà a depolimerizzare nel tempo e, se maneggiata correttamente, può essere utilizzata per almeno 6 mesi.
- Risospendere il complesso Arp2/3 (cervello suino) (vedi tabella dei materiali) seguendo le istruzioni del produttore (20 μg di proteine in 20 μL di H 2 O freddo), con la sequenza di centrifugazione a impulsi, miscelazione, ecc. su ghiaccio come descritto per l'actina al punto2.1. Combinare le soluzioni proteiche dalla sospensione di due provette di polvere per avere uno stock più grande per esperimenti riproducibili. Preparare 2 μL di aliquote e conservare a -80 °C.
- Risospendere la profilina (ricombinante umano) (vedi tabella dei materiali) ad una concentrazione 4 volte superiore a quella prevista nelle istruzioni del fabbricante (100 μg di proteine in 25 μL di H 2 O freddo), con la sequenza di centrifugazione a impulsi, miscelazione, ecc. su ghiaccio come per l'actina nella fase2.1. Combinare le soluzioni proteiche dalla sospensione di due provette di polvere prima di determinare la concentrazione proteica per avere uno stock più grande per esperimenti riproducibili.
NOTA: Conservare in frigorifero o in frigorifero; Non congelare o lasciare mai scaldare. Se maneggiata correttamente, la profilina risospesa è buona per almeno 6 mesi a 1 anno. - Risospendere la proteina di tappatura (α1β2, ricombinante umano) (cfr. tabella dei materiali) seguendo le istruzioni del fabbricante (50 μg di proteine in 50 μL di H 2 O freddo), con la sequenza di centrifugazione a impulsi, miscelazione ecc. su ghiaccio come descritto per l'actina al punto2.1. Raffreddare 50 μL di glicerolo sul ghiaccio e aggiungere i 50 μL di proteina di copertura risospesa; Mescolare delicatamente. Conservare a -20 °C.
NOTA: La soluzione non si congela e l'attività è robusta, quindi la soluzione può essere conservata come singola aliquota per mesi, o addirittura anni, se maneggiata con cura. La proteina di tappatura ricombinante del topo, quella più comunemente usata in passato negli esperimenti in vitro13, sarà presto disponibile in commercio. - Risospendere il gelsolin (ricombinante umano, con tag His) (vedi tabella dei materiali) seguendo le istruzioni del produttore (20 μg di proteine in 20 μL di H 2 O freddo), con la sequenza di centrifugazione ad impulsi, miscelazione, ecc. su ghiaccio come descritto per l'actina al punto2.1. Circa 2 μL di gelsolin vengono utilizzati al giorno di esperimenti; preparare quindi grandi aliquote (5-10 μL) e conservare a -80 °C.
NOTA: Il protocollo con gelsolin è fornito come alternativa. Si raccomanda l'uso di proteine tappanti al posto della gelsolina, acquistate o purificate come in13. - Risospendere VCA (WASP-VCA umana, marcata GST) (vedi tabella dei materiali) a una concentrazione 2 volte superiore a quella prevista nelle istruzioni del produttore (500 μg di proteine in 250 μL di H2O freddo), con la sequenza di centrifugazione a impulsi, miscelazione, ecc. su ghiaccio come per l'actina nella fase 2.1. Produrre aliquote da 10 μL e conservare a -80 °C.
NOTA: Una volta che SpVCA (pVCA umano, streptavidina e His-tagged) è commercializzato o se la purificazione delle proteine14 è possibile, si raccomanda l'uso di SpVCA invece di VCA. VCA non fornisce comete riproducibili nelle condizioni qui descritte.
3. Misurazione delle concentrazioni proteiche
- Costruire una curva standard di Bradford composta da due diluizioni seriali sovrapposte di BSA.
NOTA: La curva standard deve essere costruita solo ogni due mesi (o anche meno frequentemente) finché lo spettrofotometro non cambia.- Nella fila 1 di un rack per microtubi, posizionare i tubi per la serie di diluizione BSA #1: quattro tubi da 2 ml seguiti da quattro tubi da 1,5 ml. Nella fila 3 del rack, posizionare i tubi per la serie di diluizione BSA #2 come per la serie #1. Nella riga 5 del rack, posizionare una provetta da 2 ml per ciascun campione da misurare insieme a due provette da 1,5 ml. Aggiungere un singolo tubo da 1,5 mL nella riga 5 per il vuoto.
- Misurare il reagente Bradford (vedere la tabella dei materiali) nelle provette da 1,5 ml.
- Riempire un tubo conico da 15 mL verso l'alto con Bradford Reagent per facilitare il pipettaggio (l'eccesso verrà restituito al flacone di riserva nel frigorifero) sul ghiaccio. Assumere 200 μL di reagente Bradford ed espellerlo nuovamente nel tubo conico per bagnare la punta della pipetta. Poiché la soluzione è viscosa, pipettare lentamente per consentire alla soluzione di entrare e uscire completamente dalla punta senza formare bolle.
- Con la punta "pre-bagnata", pipettare lentamente 200 μL di Bradford Reagent in ciascuna delle provette da 1,5 mL nel rack (quattro per ogni diluizione BSA, una per il bianco e due per ogni campione da misurare). Farlo prima per consentire al reagente Bradford di riscaldarsi completamente a temperatura ambiente prima di miscelarlo con soluzioni proteiche. Riportare il contenuto rimanente del tubo conico da 15 mL nel flacone.
- Misurare H 2 O nei tubi da2ml. Nella riga 1 del rack, aggiungere 1.990 μL H2O al primo tubo e 900 μL agli altri tre tubi. Per la riga 3, aggiungere 1.992,5 μL al primo tubo e 900 μL agli altri tre tubi. Aggiungere 2.000 μL di H2O a ciascuna delle provette della riga 5.
NOTA: per tutti i volumi superiori a 1.000 μL, utilizzare una pipetta da 1.000 μL, ma somministrare l'intera quantità mediante pipettaggio due volte. È importante preparare tutto prima di iniziare i passaggi successivi, al fine di evitare di lasciare proteine altamente diluite in H2O per lunghi periodi di tempo. - Per preparare la serie di diluizione BSA #1, mescolare 10 μL di 2 mg/mL di BSA calibrato (vedere Tabella dei materiali) nel tubo con 1.990 μL di H2O per ottenere una soluzione da 10 μg/ml. Da questo, fare tre diluizioni seriali (5 μg/mL, 2,5 μg/mL e 1,25 μg/mL BSA) trasferendo 900 μL di ciascuna soluzione nella provetta successiva (contenente 900 μL H2O).
- Per preparare la serie di diluizione BSA #2, mescolare 7,5 μL di 2 mg/mL di BSA calibrato nel tubo con 1.992,5 μL di H2O per ottenere una soluzione da 7,5 μg/ml. Da questo, fare tre diluizioni seriali (3,75 μg/mL, 1,875 μg/mL e 0,9375 μg/mL BSA) trasferendo 900 μL di ciascuna soluzione nella provetta successiva (contenente 900 μL H2O).
- Mescolare e leggere l'assorbanza per generare la curva standard. Aggiungere 800 μL H2O al tubo del reagente Bradford per il pezzo grezzo e avviare il timer. Nel modo più efficiente possibile senza creare bolle, mescolare 800 μL di ciascun standard BSA con 200 μL di reagente Bradford nelle provette preparate. Una volta che tutti gli standard sono stati miscelati con il reagente Bradford (< 5 min), versare ogni standard in una cuvetta monouso e leggere l'assorbanza a 600 nm nello spettrofotometro dopo aver oscurato la macchina.
NOTA: La stessa cuvetta può essere utilizzata per leggere l'intera serie se viene letto prima lo standard meno concentrato e la cuvetta è ben svuotata tra una lettura e l'altra. Ripetete la curva standard finché l'adattamento lineare non ha un valore R di almeno 0,99. Solo dopo aver padroneggiato il pipettaggio e la miscelazione delicata, procedere alla lettura dei campioni.
- Misurare le concentrazioni di actina e proteine leganti l'actina
- Nelle provette da 2 mL contenenti 2.000 μL di H 2 O (preparate al punto 3.1.3), mescolare delicatamente quanto segue:2μL ciascuno di complesso Arp2/3 e profilina, 4 μL di G-actina marcata, 5 μL ciascuno di gelsolina e VCA e 8 μL di proteina di tappatura (ricombinante umano). Prendere immediatamente 800 μL della soluzione e mescolare con il reagente Bradford già preparato, quindi ripetere per avere due letture entro il 5% -10% l'una dall'altra per ciascun campione. Una differenza maggiore indica un problema con la risospensione o la maneggevolezza. Leggere entro pochi minuti dalla miscelazione con il reagente Bradford.
NOTA: se si utilizza una curva standard di un altro giorno, oltre ai campioni deve essere preparato solo il bianco del punto 3.1.6.
- Nelle provette da 2 mL contenenti 2.000 μL di H 2 O (preparate al punto 3.1.3), mescolare delicatamente quanto segue:2μL ciascuno di complesso Arp2/3 e profilina, 4 μL di G-actina marcata, 5 μL ciascuno di gelsolina e VCA e 8 μL di proteina di tappatura (ricombinante umano). Prendere immediatamente 800 μL della soluzione e mescolare con il reagente Bradford già preparato, quindi ripetere per avere due letture entro il 5% -10% l'una dall'altra per ciascun campione. Una differenza maggiore indica un problema con la risospensione o la maneggevolezza. Leggere entro pochi minuti dalla miscelazione con il reagente Bradford.
- Calcolare le concentrazioni di actina e proteine leganti l'actina utilizzando la curva standard e il fattore di diluizione. Rifare la misurazione per ogni nuova risospensione. I pesi molecolari per convertire le letture mg/mL ottenute tramite il saggio di Bradford in μM sono: actina 43 kD, complesso Arp2/3 224 kD, profilina 15 kD, gelsolina 95 kD, proteina capping 68 kD (ricombinante umano) o 63,5 kD (topo ricombinante), VCA 43 kD e SpVCA 54 kD (peso molecolare monomero).
4. Rivestimento di perline
- Pre-raffreddare la centrifuga a 4 °C e impostare il blocco secco di agitazione (vedere Tabella dei materiali) a 18 °C.
- Lavare le perline: pipettare 50 μL di tampone Xb in una provetta da microcentrifuga da 1,5 ml, aggiungere 9 μL di sospensione del tallone da 4,5 μm di diametro o 2 μL di sospensione del tallone da 1 μm di diametro (sospensione 2,5 % p/v) (vedere tabella dei materiali). Mescolare accuratamente e centrifugare i campioni a 20.000 x g per 10 minuti a 4 °C.
NOTA: La superficie totale delle perline di entrambe le dimensioni è di 3 cm2:
Derivato calcolando il numero di perline e quindi la loro superficie totale utilizzando equazioni classiche per il volume e la superficie delle sfere. Altre dimensioni di perline possono essere utilizzate se le quantità sono regolate per mantenere costante la superficie totale a 3 cm2. - Rivestire le perline: rimuovere con cura il surnatante senza disturbare le perle e risospendere il pellet di perline in 40 μL di 2 μM SpVCA (o 7 μM VCA) in tampone Xb mediante pipettaggio delicato. Agitare a 18 °C, 1.000 giri/min per 20 min.
- Lavare le perline rivestite: centrifugare la miscela (20.000 x g per 10 minuti a 4 °C) e rimuovere con cura il surnatante. Risospendere le sfere in 50 μL di Xb/1% BSA freddo e centrifugare a 20.000 x g per 10 minuti a 4 °C. Rimuovere il surnatante e ripetere il passaggio di lavaggio 1x.
- Risospendere il pellet di perline rivestito dal punto 4.4 in 120 μL di Xb freddo / 1% BSA per entrambe le dimensioni di perle in modo che la quantità di superficie del tallone / μL di soluzione di perline sia la stessa. Conservare in frigorifero o in cella frigorifera. Le perle rivestite continueranno a funzionare normalmente per almeno diverse settimane.
5. Preparazione del mix di motilità e delle diapositive per l'osservazione
NOTA: Il volume totale della miscela di motilità è di 8,4 μL per consentire uno spazio di circa 25 μm tra il vetrino e il coprivetrino da 18 x 18 mm in modo che le perline di tutte le dimensioni (fino a 10 μm di diametro) non vengano schiacciate. La miscela di motilità di base è di circa 5 μM di G-actina (actina fluorescente marcata al 10%) con 5 μM di profilina, 50 nM di complesso Arp2/3 e 25 nM di proteina capping (o 240 nM di gelsolina).
- Preparare la miscela di reazione alla motilità. Le quantità esatte di actina (e quindi di profilina) dipendono dalla concentrazione calcolata al punto 3.3, ma una reazione rappresentativa è la seguente. Mescolare su ghiaccio, in questo ordine: 3,2 μL di MB13, 1,5 μL di profilina a 30 μM diluita in MB13, 1 μL di proteina di tappatura a 0,21 μM diluita in MB13 o gelsolina diluita a 2 μM in MB13, 1 μL di complesso Arp2/3 a 0,47 μM diluito in MB13, 0,2 μL di sospensione di perline (vortice appena prima dell'uso), e 1,5 μL di actina a 30 μM in G-buffer. Mescolare bene ma rapidamente e avviare il timer.
- Individua l'intera miscela di reazione alla motilità su un vetrino. Coprire con un coprivetrino di 18 mm x 18 mm e sigillare il coprislip con VALAP fuso usando un piccolo pennello. VALAP è una miscela di lanolina, paraffina e vaselina (vedi Tabella dei materiali) 1:1:1 in peso, fusa e mescolata insieme.
6. Osservazione al microscopio
- Osservare immediatamente le reazioni di motilità utilizzando un obiettivo 100x su un microscopio verticale o invertito (vedi Tabella dei materiali), dotato di microscopia a contrasto di fase e / o epifluorescenza (cubo GFP, vedi Tabella dei materiali). Le osservazioni vengono effettuate a temperatura ambiente (23-25 °C).
- Per ottenere velocità di spostamento medie per un'intera popolazione di perline, registrare il contrasto di fase o le immagini fisse fluorescenti nel tempo scansionando l'intera diapositiva. Misura la lunghezza della cometa a mano e traccia il tempo. La pendenza dell'adattamento lineare è la velocità di crescita media.
- Per valutare la velocità delle singole perline, raccogliere filmati time-lapse al microscopio a contrasto di fase. A seconda della velocità del tallone e della risoluzione richiesta, prendere fotogrammi ogni 1-10 s. Utilizza lo strumento di tracciamento di qualsiasi programma di elaborazione delle immagini per ottenere velocità e traiettorie delle perline.
Risultati
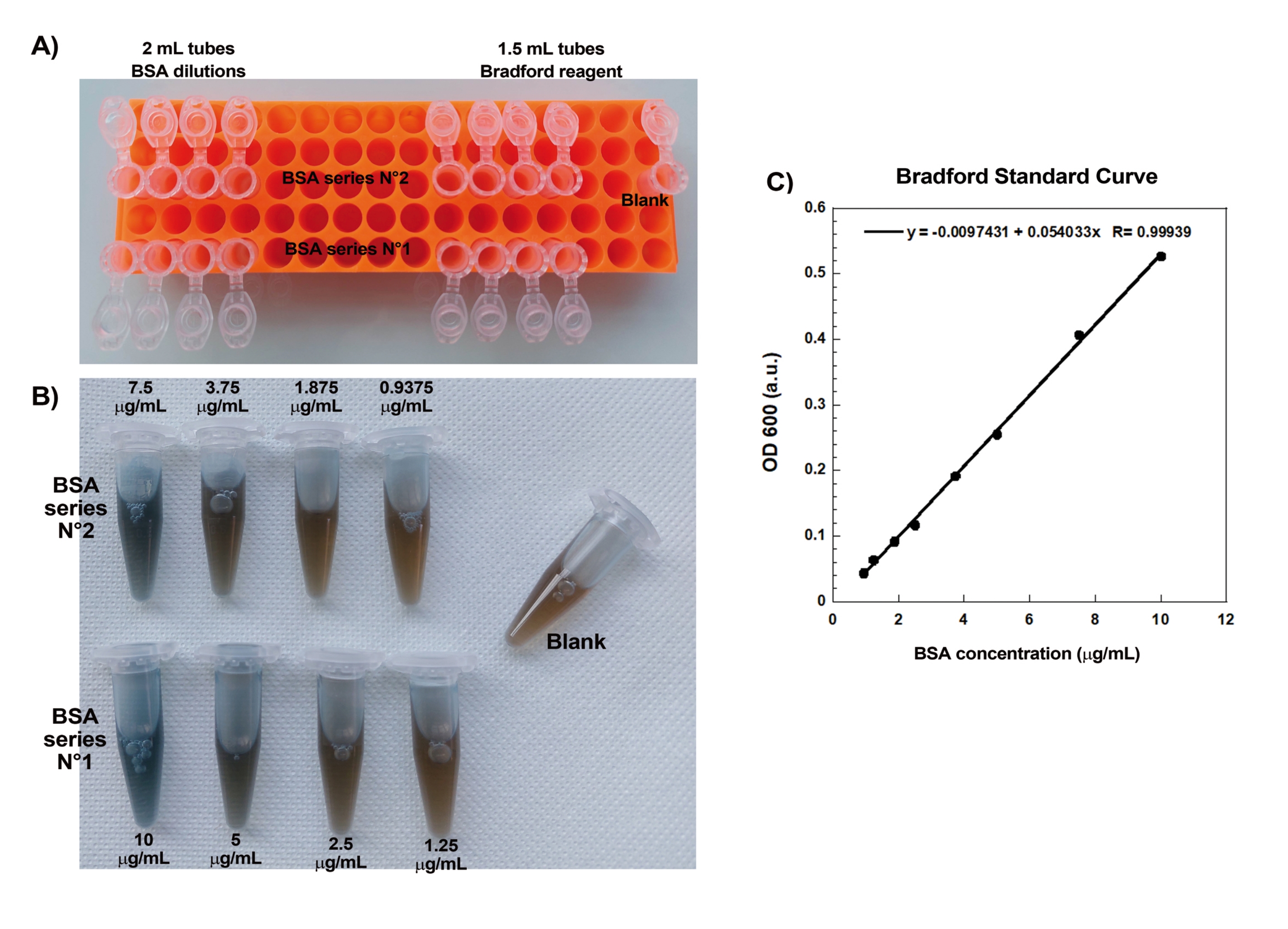
Uno degli aspetti chiave della creazione riproducibile di comete di actina su perline è il pipettaggio delicato e preciso di delicate proteine leganti l'actina. La generazione di una curva standard di Bradford è un buon modo per valutare le capacità di pipettaggio. La figura 1A,B mostra i tubi per la curva standard e un esempio di come appaiono le due diluizioni seriali di BSA una volta miscelate con il reagente Bradford. Si noti la tonalità blu graduata (una maggiore concentrazione proteica dà una soluzione più blu). Se lette nello spettrofotometro e tracciate, queste soluzioni forniscono una curva standard come mostrato nella Figura 1C. Per praticare un pipettaggio accurato, il test deve essere ripetuto fino a quando il fattore di correlazione lineare è 0,999, come mostrato.
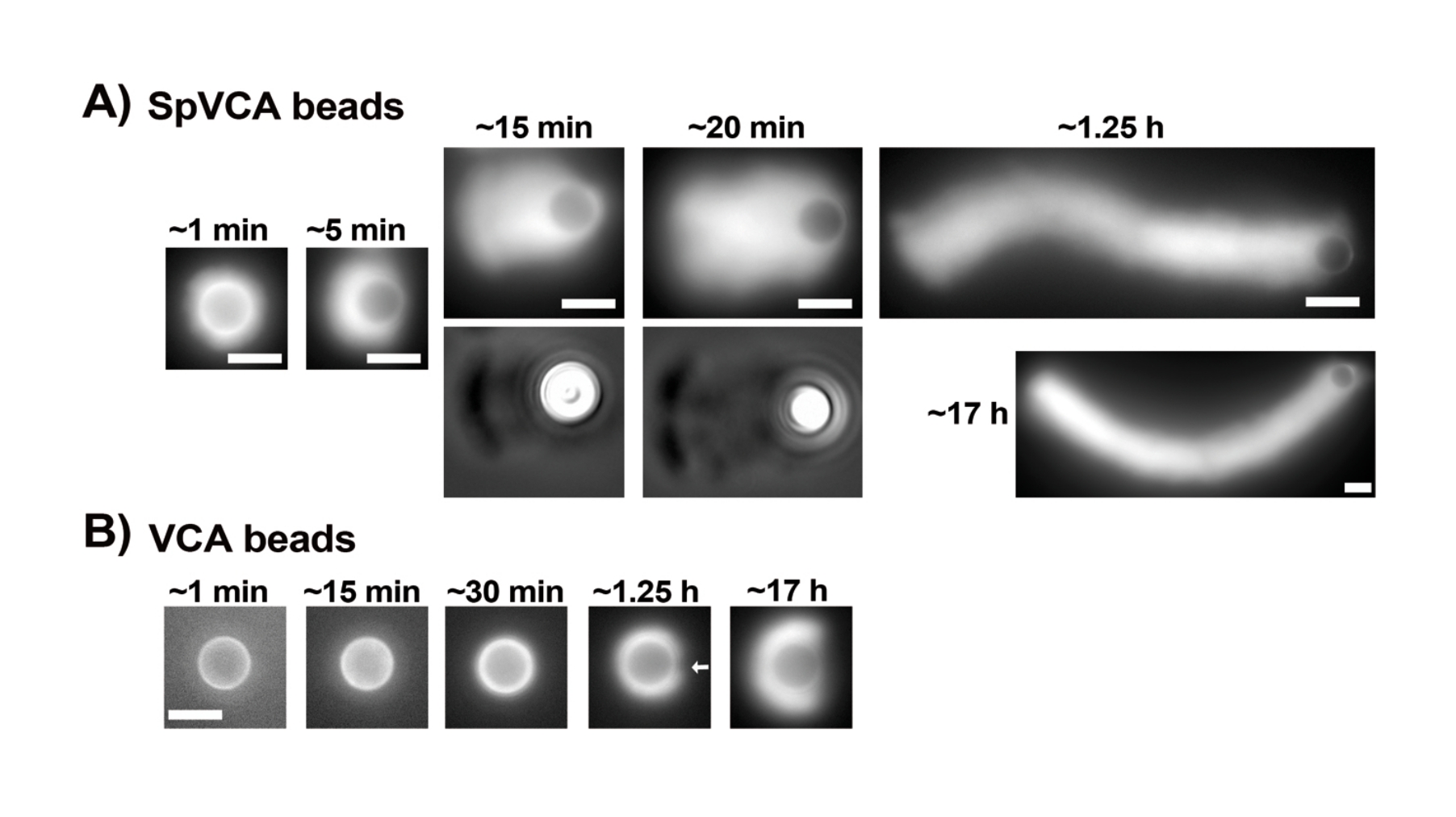
Una volta che le concentrazioni di proteine risospese commerciali sono state attentamente valutate tramite il test Bradford, le perle rivestite e il mix di motilità vengono preparati e miscelati insieme. La figura 2A mostra immagini rappresentative dei diversi stadi di formazione della cometa: le nubi di actina si formano entro pochi minuti dalla miscelazione delle perle rivestite di SpVCA e del mezzo di motilità; La polarizzazione delle nubi avviene a ~ 5 minuti e la produzione di comete a 15-20 minuti. Le comete di actina, visibili sia con l'epifluorescenza che con la microscopia a contrasto di fase (Figura 2A), continuano ad allungarsi per molte ore, ma non viene mantenuta una velocità costante, quindi la motilità delle perle viene normalmente valutata entro 1 ora. D'altra parte, ci vogliono 30 minuti con perle rivestite di VCA per ottenere nubi di actina luminose (Figura 2B), e non si formano comete, anche se la simmetria inizia a rompersi a 1-2 h (freccia nella Figura 2B) e le nuvole mostrano polarizzazione dopo l'incubazione notturna.
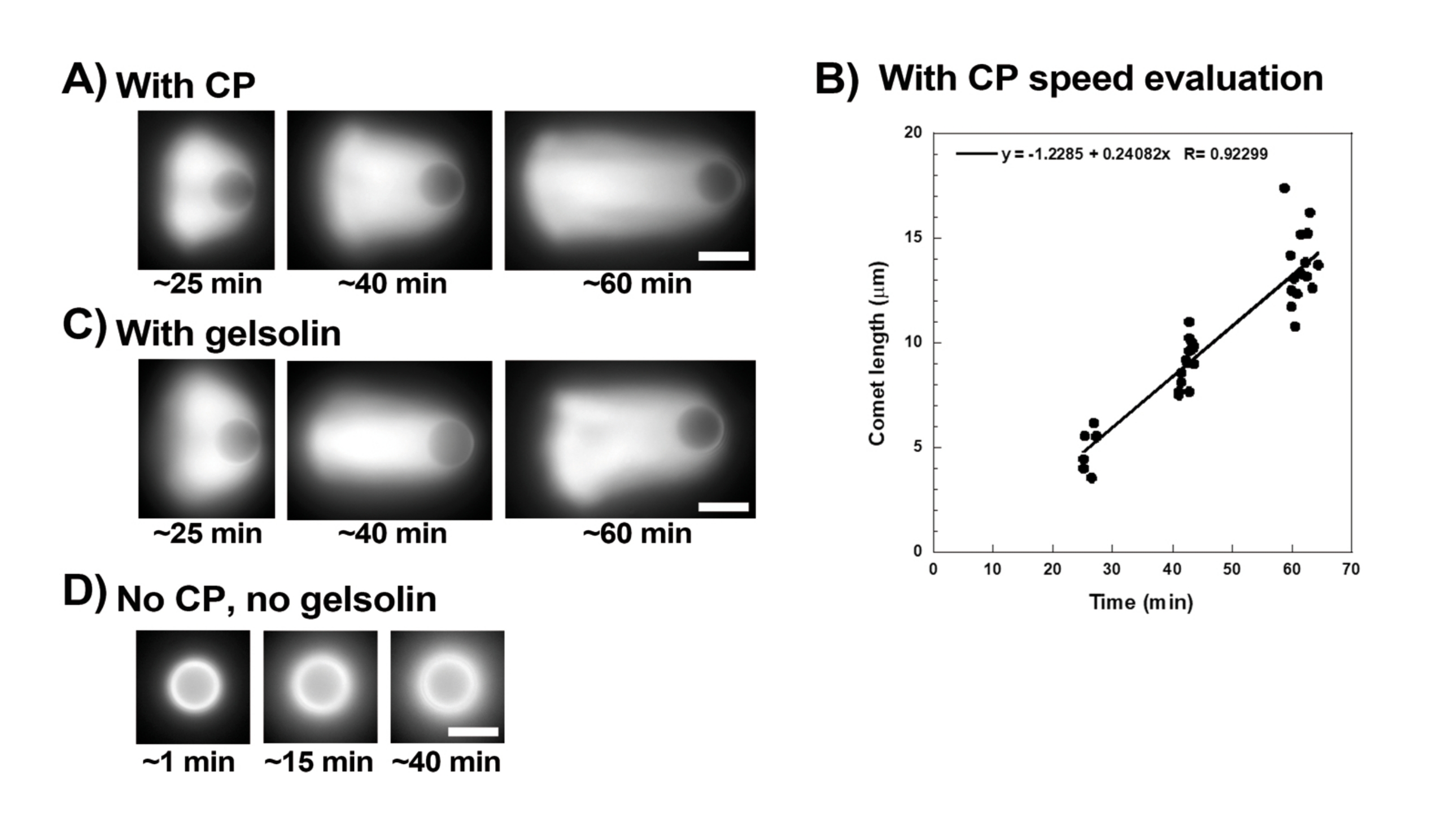
La Figura 3 mostra un esempio di valutazione della velocità delle perle in presenza di proteine capping. Poiché tutte le perle rompono la simmetria all'incirca nello stesso momento, vengono eseguite registrazioni "pseudo-time-lapse" in cui viene scansionata la diapositiva e le immagini dell'intera popolazione di comete vengono scattate nel tempo (Figura 3A). Le comete non depolimerizzano; pertanto, l'aumento delle lunghezze delle comete misurato nel tempo può essere utilizzato per calcolare la velocità di spostamento (Figura 3B). La gelsolina può essere utilizzata al posto della proteina di tappatura per la formazione della cometa se viene aggiunta 10 volte più gelsolina per compensare la sua ridotta attività di capping. Le comete formate in presenza di gelsolin sono qualitativamente le stesse e si muovono approssimativamente alle stesse velocità delle perle con la proteina di copertura (Figura 3C). L'attività di tappatura è fondamentale per concentrare la polimerizzazione sulla superficie della perla, e quando né la proteina di tappatura né la gelsolina sono incluse nel mix di motilità, le nubi di actina non si polarizzano mai per formare comete, sebbene nubi di actina luminose si formino attorno alle perle (Figura 3D). Le comete su perline possono essere utilizzate per misurare la produzione di forza basata sull'actina in diversi contesti biochimici alterando il mix di motilità e osservando il risultato sulla motilità utilizzando diverse tecniche di micromanipolazione, ad esempio15.

Figura 1: Curva standard di Bradford. (A) Immagine di come impostare i tubi per realizzare la curva standard di Bradford. Le provette campione non sono mostrate. (B) Immagine delle due diluizioni seriali BSA sovrapposte una volta miscelate con il reagente Bradford. (C) Le assorbanze a 600 nm delle soluzioni indicate al punto (B) sono misurate nello spettrofotometro e sono tracciate in funzione delle concentrazioni proteiche delle soluzioni BSA. L'adattamento lineare viene utilizzato per calcolare la concentrazione del campione. Il fattore di correlazione, R, dell'adattamento lineare è 0,999. Fare clic qui per visualizzare una versione ingrandita di questa figura.
{kind=link}

Figura 2: Formazione di comete su perle rivestite di SpVCA rispetto alle perle rivestite di VCA. (A) Perle rappresentative rivestite di SpVCA (perline diverse in ogni immagine) mostrate nel tempo. Viene indicato il tempo dal momento della miscelazione. Le nubi di actina si formano immediatamente e la polarizzazione delle nubi dà comete, che continuano ad allungarsi per ore. (B) Perle rappresentative rivestite di VCA (perline diverse in ogni immagine) mostrate nel tempo. Più di 1 ora è necessaria per vedere l'inizio della polarizzazione della nube di actina (freccia) e anche lunghe incubazioni non producono comete. Tutte le immagini sono di perle di 4,5 μm di diametro, epifluorescenza di actina fluorescente abbinata alla visualizzazione del contrasto di fase per i punti temporali di 15 e 20 minuti in (A), barre di scala = 5 μm. Fare clic qui per visualizzare una versione ingrandita di questa figura.
{kind=link}

Figura 3: Analisi delle comete e della velocità delle perline. (A) e (C) In presenza della proteina di tappatura (CP) o della gelsolina, le nubi di actina si polarizzano per formare comete nei primi 20 minuti di reazione e le comete si allungano nel tempo. Tempo di miscelazione indicato per ogni immagine; Ogni immagine è una perlina diversa. C'è una certa variabilità tra le perle e la preparazione, ma in media le perle si muovono a velocità micron / sub-micron al minuto (0,2-1 μm / min) nelle condizioni standard qui descritte. (B) Grafico rappresentativo che mostra la valutazione della lunghezza della cometa (intera popolazione di perle) nel tempo. La pendenza della correlazione lineare corrisponde alla velocità media di spostamento, in questo caso 0,24 μm/min. (D) In assenza di attività di tappatura (nessuna proteina di tappatura o gelsolina), le nubi di actina si formano attorno alle perle, ma le comete non si formano. Tutte le immagini sono di perle di 4,5 μm di diametro, epifluorescenza di actina fluorescente, barre di scala = 5 μm. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
Discussione
Il protocollo qui descritto descrive come ottenere la crescita della rete di actina sulle superfici delle perle, la formazione della cometa e la motilità delle perle utilizzando proteine disponibili in commercio. Tuttavia, a volte le comete non sono osservate in modo riproducibile o sono disomogenee tra il vetrino e il coprifoglio. La seguente discussione sottolinea alcuni punti chiave del protocollo e suggerisce alcuni parametri che possono essere regolati. Un fattore da tenere a mente è che la formazione delle comete e la velocità delle perle sono influenzate dalla temperatura, con temperature molto superiori a 25 ° C o molto inferiori a 23 ° C che hanno un impatto negativo sulla formazione delle comete e forniscono dati irriproducibili. Si consiglia vivamente l'uso di un microscopio a temperatura controllata o di un microscopio in una stanza climatizzata. Sebbene l'actina marcata con fluorescenza sia spesso inclusa nel mix di motilità per osservare le comete mediante microscopia a fluorescenza, una volta che le comete sono lunghe più di un diametro di perle, sono anche visibili dalla microscopia a contrasto di fase come uno striscio scuro accanto al tallone. La visualizzazione del contrasto di fase è più appropriata per l'imaging time-lapse poiché una certa fototossicità è associata all'imaging a fluorescenza anche tramite disco rotante. Poiché le perline si depositano nel tempo, un microscopio invertito produce meno deriva orizzontale delle perline rispetto a uno verticale ed è più appropriato per i film. L'uso di VALAP fuso per sigillare i vetrini è importante in quanto sostanze come lo smalto per unghie interferiscono con la formazione della cometa. Grandi quantità di VALAP possono essere prodotte in un becher e quindi raccolte per riempire becher più piccoli più suscettibili di fusione rapida. VALAP è buono per anni a temperatura ambiente.
Un altro aspetto tecnico chiave è la meticolosa preparazione del buffer e del mix di motilità. Prestare attenzione quando si prepara MB13, in particolare nella fase di regolazione del pH. Il pH di MB13 deve essere regolato rapidamente a neutro con NaOH per evitare l'idrolisi dell'ATP, ma non troppo rapidamente poiché l'EGTA si solubilizza quando il pH si avvicina al neutro. L'EGTA è un ingrediente chiave perché complessano il calcio legato all'actina, dando nel mix di motilità il magnesio più attivo forma16. MB13 preparato troppo velocemente o troppo lentamente dà una formazione di cometa non ottimale o addirittura nessuna. Un ulteriore punto chiave è quello di tenere traccia attenta della concentrazione di KCl nel mix di motilità quando si gioca con le condizioni. Ad esempio, quando si utilizza 1x MB13 nel mix di reazione e si diluisce la profilina, la proteina di tappatura e il complesso Arp2/3 in MB13, la concentrazione finale di KCl nella reazione di motilità è di circa 40-50 mM a causa della diluizione da parte del G-buffer. Questa concentrazione dà i migliori risultati nel saggio della cometa, e più di 60 mM KCl diminuisce l'attività nucleante complessa Arp2/3.
Dal punto di vista delle proteine, un aspetto tecnico critico per ottenere comete di actina è la corretta gestione delle proteine leganti l'actina commerciali, in particolare il pipettaggio preciso di quantità di microlitri. La linearità della curva standard di Bradford è un buon test di pipettaggio e la curva può quindi essere utilizzata per misurazioni di routine delle concentrazioni proteiche. Infatti, quando si utilizzano proteine commerciali risospese per la procedura cometa, è importante verificare sempre le concentrazioni proteiche, poiché la variabilità del lotto e l'errore dell'utente durante la risospensione possono portare a differenze tra concentrazioni reali e attese. A volte piccole differenze nelle concentrazioni proteiche possono portare alla completa assenza di comete.
Un altro aspetto importante del metodo qui presentato è l'uso della G-actina complessata con profilina come combustibile per la polimerizzazione. Storicamente, i sistemi in vitro utilizzavano actina filamentosa pre-polimerizzata (F-actina) come fonte di actina: depolimerizzazione nella polimerizzazione bulk alimentata sulla superficie10,17. Ciò aveva il vantaggio di controllare i livelli di G-actina, ma aggiungeva uno strato di complessità che richiedeva componenti aggiuntivi per catalizzare la depolimerizzazione. Poiché il turnover della rete di actina non è necessario per la produzione di forza e la motilità, che sono alimentate dalla nucleazione e dalla polimerizzazione sulla superficie della perla, mentre i fattori di depolimerizzazione dell'actina come ADF / cofilina agiscono sulle reti invecchiate lontane dalla superficie18, la maggior parte della ricostituzione in vitro della motilità basata sull'actina viene ora eseguita senza turnover per semplicità. Tuttavia, ci sono alcuni inconvenienti nell'utilizzo di G-actina. In primo luogo, quando si utilizza l'actina commerciale, che è stata liofilizzata, sono presenti oligomeri. Le fasi di depolimerizzazione qui descritte sono molto importanti per ottenere risultati riproducibili. In particolare, sebbene il G-buffer sia tradizionalmente regolato a pH 8, un pH più basso (pH 7, per esempio) sembra funzionare meglio nei saggi descritti in questo articolo, probabilmente perché un pH basso migliora la depolimerizzazione19. Un altro svantaggio dell'uso di G-actina è che una volta posto in condizioni saline permissive alla polimerizzazione, si verifica una nucleazione spontanea e si forma F-actina nella massa e sulla superficie del tallone. Il complessamento della G-actina con la profilina sopprime la nucleazione spontanea nella massa e la polimerizzazione dell'estremità appuntita, concentrando così sia la nucleazione che la polimerizzazione dell'estremità spinata sulla superficie20. La profilina-G-actina è fisiologicamente rilevante, poiché gran parte dell'actina nella cellula è presente in questa forma21. Qui viene utilizzato un rapporto 1: 1 di profilina: actina; tuttavia, rapporti più elevati (ad esempio 3:1) inibiscono più accuratamente la polimerizzazione nella massa, sebbene rapporti più alti inibiscano anche il complesso Arp2/3 e l'allungamento dell'estremità spinata in una certa misura22,23.
L'attività di capping è anche fondamentale per la formazione di comete poiché assicura l'inserimento di nuova actina in superficie attraverso cicli di nucleazione da parte del complesso Arp2/324,25 attivato dalla superficie. Senza tappo, le nubi di actina non rompono la simmetria per formare comete perché la polimerizzazione sulla superficie non accumula abbastanza tensione per aprire la nube26. In passato, abbiamo usato la proteina di tappatura del topo ricombinante purificata a casa 13, ma i test eseguiti per questo articolo indicano che la proteina di tappatura umana ricombinante disponibile in commercio è ugualmente efficace, così come la gelsolina disponibile in commercio, sebbene debba essere utilizzata10 volte più gelsolina e, per alcune applicazioni, potrebbe non essere appropriata in quanto ha attività di recisione dell'actina e tappatura27.
Infine la robustezza di questo metodo risiede nell'uso di un attivatore complesso Arp2/3 molto attivo, la streptavidina-pVCA (SpVCA)28. SpVCA include il dominio di legame profilina-G-actina di WASP (il dominio p) oltre al dominio di legame complesso Arp2/3 in quanto questo è risultato essere più efficiente in condizioni di profilina-G-actina29. Ancora più importante, l'uso del tag streptavidina, originariamente introdotto per consentire la funzionalizzazione superficiale attraverso il legame biotina-streptavidina, ha l'effetto aggiuntivo di aumentare l'attivazione del complesso Arp2/3, presumibilmente dovuto al fatto che la streptavidina è un tetramero e quindi raggruppa l'attivatore, noto per aumentare l'attività complessa Arp2/330 . SpVCA prodotto commercialmente è attualmente in fase di sviluppo e sarà presto disponibile per l'acquisto. Va inoltre notato che, sebbene 40 μL di 2 μM SpVCA siano abitualmente utilizzati per rivestire 3 cm2 di superficie del tallone, anche altre concentrazioni di rivestimento (più alte e più basse) funzionano, e giocando con queste condizioni si ottengono diverse velocità di crescita e morfologie della cometa. Infatti, quando le comete non si formano o la dimensione della cometa non è omogenea sul vetrino, dovrebbero essere testate diverse condizioni di rivestimento, nonché diverse concentrazioni di KCl e profilina nel mix di motilità. Le concentrazioni di actina, complesso Arp2/3 e proteina di tappatura nel mix di motilità possono anche essere alterate per ottimizzare la formazione della cometa, ma nelle nostre mani, cambiare queste proporzioni spesso dà risultati confusi.
Per concludere, i metodi qui descritti producono assemblaggio di actina su superfici di perline e motilità, ma è possibile utilizzare qualsiasi superficie che può essere funzionalizzata con SpVCA. Nei casi in cui l'adsorbimento come descritto qui non funziona, la porzione di streptavidina può essere utilizzata per attaccare SpVCA alla superficie di interesse dopo la biotinilazione. Le strutture di actina così formate, comete o altro, possono essere utilizzate per testare diversi aspetti biochimici e biofisici delle reti di actina e sono particolarmente appropriate per manipolazioni fisiche con micropipette, pinzette ottiche e ablazioni laser 15,26,31,32. Oltre ai suoi usi per la comunità di ricerca, l'approccio qui descritto è appropriato come strumento didattico per gli studenti universitari di biofisica per studiare concetti di materia attiva come la rottura della simmetria e l'auto-organizzazione.
Divulgazioni
Gli autori dichiarano di non avere conflitti di interesse con i contenuti di questo articolo.
Riconoscimenti
Riconosciamo sinceramente i membri della nostra nuova casa a LPENS per il loro caloroso benvenuto e, in particolare, il team ABCDJ per tutto il loro aiuto e supporto. J.P. riconosce il sostegno finanziario della Fondazione ARC (Grant PJA 20191209604) e C.S. riconosce il sostegno finanziario della Human Frontiers Science Program Organization (Grant RGP0026/2020).
Materiali
| Name | Company | Catalog Number | Comments |
| Actin, rabbit muscle, Alexa Fluor 488 conjugate | Invitrogen (ThermoFisher Scientific) | A12373 (recently discontinued) | This product can be replaced with ATTO-488 actin from Hypermol. |
| Actin, rabbit muscle, ATTO-488 | Hypermol | 8153 | |
| Actin, rabbit skeletal muscle | Cytoskeleton | AKL99 | |
| Arp2/3 complex | Cytoskeleton | RP01P | |
| ATP | Sigma | A7699 | |
| BioSpectrometer, basic | Eppendorf | 035739 | |
| Bradford Reagent | Bio-Rad | 500-0006 | |
| BSA, high quality | Sigma | A3059 | |
| BSA standard 2 mg/mL (Pierce) | Thermo Scientific | 23209 | |
| Capping protein (a1b2, mouse recombinant) | Home-purified (Reference 13) | This product will soon be commercially available from Cytoskeleton. | |
| Capping protein (a1b2, human recombinant) | Hypermol | 8322 | |
| Cube, GFP: U-MNIBA3 or U-MWB2 | Olympus | discontinued | Any GFP cube, adapted to the microscope being used, can be used. |
| Dry block, agitating: ThermoMixer C (refrigerated) | Eppendorf | 035963 | |
| ** with SmartBlock, 24 microtubes 2 mL | Eppendorf | 035969 | |
| Gelsolin (human recombinant, His-tagged) | Cytoskeleton | HPG6 | |
| Lanolin | Sigma | 49909 | |
| Microcentrifuge 5427R + rotor | Eppendorf | 934126 | |
| Microscope, upright: BX51 | Olympus | discontinued | Any epifluorescence upright microscope equipped with phase contrast optics can be used. |
| Microscope, inverted: IX70 | Olympus | discontinued | Any epifluorescence inverted microscope equipped with phase contrast optics can be used. |
| Paraffin | Sigma | 76244 | |
| Petroleum jelly: Vaseline | Sigma | 16415 | |
| Pipettes Research Plus | Eppendorf | Gilson pipettes don't work as well for delivery of very small volumes (0.5 µL for example). | |
| **10 µL | 933954 | ||
| **2.5 µL | 933953 | These two sizes are essential, but the use of high-quality pipettes (a full Research Plus set for example) is recommended. | |
| Polystyrene carboxylate beads | Polysciences | ||
| **approx. 1 µm diameter | 08226 | ||
| **approx. 4.5 µm diameter | 17140-5 | ||
| Profilin 1 (human recombinant, untagged) | Cytoskeleton | PR02 | |
| SpVCA (human WASP pVCA domain, N-ter His-tag, C-ter Streptavidin tag) | Home-purified (Reference 14) | This product will soon be commercially available from Cytoskeleton. | |
| VCA (human WASP VCA domain, GST-tagged) | Cytoskeleton | VCG03 |
Riferimenti
- Campellone, K. G., Welch, M. D. A nucleator arms race: cellular control of actin assembly. Nature Reviews Molecular Cell Biology. 11 (4), 237-251 (2010).
- Pollard, T. D. Rate constants for the reactions of ATP- and ADP-actin with the ends of actin filaments. Journal of Cell Biology. 103, 2747-2754 (1986).
- Blanchoin, L., Boujemaa-Paterski, R., Sykes, C., Plastino, J. Actin dynamics, architecture and mechanics in cell motility. Physiological Reviews. 94 (1), 235-263 (2014).
- Tilney, L. G., Tilney, M. S. The wily ways of a parasite: induction of actin assembly by Listeria. Trends in Microbiology. 1 (1), 25-31 (1993).
- Moore, A. S., et al. Actin cables and comet tails organize mitochondrial networks in mitosis. Nature. 591 (7851), 659-664 (2021).
- Taunton, J., et al. Actin-dependent propulsion of endosomes and lysosomes by recruitment of N-WASP. Journal of Cell Biology. 148 (3), 519-530 (2000).
- Velarde, N., Gunsalus, K. C., Piano, F. Diverse roles of actin in C. elegans early embryogenesis. BMC Developmental Biology. 7, 142(2007).
- Merrifield, C. J., et al. Endocytic vesicles move at the tips of actin tails in cultured mast cells. Nature Cell Biology. 1 (1), 72-74 (1999).
- Samarin, S., et al. How VASP enhances actin-based motility. Journal of Cell Biology. 163 (1), 131-142 (2003).
- Bernheim-Groswasser, A., Wiesner, S., Golsteyn, R. M., Carlier, M. -F., Sykes, C. The dynamics of actin-based motility depend on surface parameters. Nature. 417 (6886), 308-311 (2002).
- Boujemaa-Paterski, R., et al. Network heterogeneity regulates steering in actin-based motility. Nature Communications. 8 (1), 655(2017).
- Akin, O., Mullins, R. D. Capping protein increases the rate of actin-based motility by promoting filament nucleation by the Arp2/3 complex. Cell. 133 (5), 841-851 (2008).
- Palmgren, S., Ojala, P. J., Wear, M. A., Cooper, J. A., Lappalainen, P. Interactions with PIP2, ADP-actin monomers, and capping protein regulate the activity and localization of yeast twinfilin. Journal of Cell Biology. 155 (2), 251-260 (2001).
- Carvalho, K., et al. Actin polymerization or myosin contraction: two ways to build up cortical tension for symmetry breaking. Philosophical Transactions of the Royal Society B. 368 (1629), 20130005(2013).
- Marcy, Y., Prost, J., Carlier, M. -F., Sykes, C. Forces generated during actin-based propulsion: a direct measurement by micromanipulation. Proceedings of the National Academy of Sciences of the United States of America. 101 (16), 5992-5997 (2004).
- Carlier, M. -F. Actin: protein structure and filament dynamics. Journal of Biological Chemistry. 266 (1), 1-4 (1991).
- Loisel, T. P., Boujemaa, R., Pantaloni, D., Carlier, M. F. Reconstitution of actin-based motility of Listeria and Shigella using pure proteins. Nature. 401 (6753), 613-616 (1999).
- Reymann, A. -C., et al. Turnover of branched actin filament networks by stochastic fragmentation with ADF/cofilin. Molecular Biology of the Cell. 22 (14), 2541-2550 (2011).
- Wioland, H., Jegou, A., Romet-Lemonne, G. Quantitative variations with pH of Actin Depolymerizing Factor/Cofilin's multiple actions on actin filaments. Biochemistry. 58 (1), 40-47 (2019).
- Plastino, J., Blanchoin, L. Dynamic stability of the actin ecosystem. Journal of Cell Science. 132 (4), 219832(2019).
- Pollard, T. D., Blanchoin, L., Mullins, R. D. Molecular mechanisms controlling actin filament dynamics in nonmuscle cells. Annual Review of Biophysics and Biomolecular Structure. 29, 545-576 (2000).
- Suarez, C., et al. Profilin regulates F-actin network homeostasis by favoring formin over Arp2/3 complex. Developmental Cell. 32 (1), 43-53 (2015).
- Courtemanche, N., Pollard, T. D. Interaction of profilin with the barbed end of actin filaments. Biochemistry. 52 (37), 6456-6466 (2013).
- Achard, V., et al. A "primer"-based mechanism underlies branched actin filament network formation and motility. Current Biology. 20 (5), 423-428 (2010).
- Sykes, C., Plastino, J. Actin filaments up against a wall. Nature. 464 (7287), 365-366 (2010).
- vander Gucht, J., Paluch, E., Plastino, J., Sykes, C. Stress release drives symmetry breaking for actin-based movement. Proceedings of the National Academy of Sciences of the United States of America. 102 (22), 7847-7852 (2005).
- McGough, A. M., Staiger, C. J., Min, J. -K., Simonetti, K. D. The gelsolin family of actin regulatory proteins: modular structures, versatile functions. FEBS Letters. 552 (2-3), 75-81 (2003).
- Abou-Ghali, M., et al. Capping protein is not necessary for polarized actin network growth and actin based motility. Journal of Biological Chemistry. 295, 15366-15375 (2020).
- Yarar, D., D'Alessio, J. A., Jeng, R. L., Welch, M. D. Motility determinants in WASP family proteins. Molecular Biology of the Cell. 13 (11), 4045-4059 (2002).
- Padrick, S. B., et al. Hierarchical regulation of WASP/WAVE proteins. Molecular Cell. 32 (3), 426-438 (2008).
- Bussonier, M., et al. Mechanical detection of a long-range actin network emanating from a biomimetic cortex. Biophysical Journal. 107 (4), 854-862 (2014).
- Paluch, E., vander Gucht, J., Joanny, J. -F., Sykes, C. Deformations in actin comets from rocketing beads. Biophysical Journal. 91 (8), 3113-3122 (2006).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneEsplora altri articoli
This article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati