
Method Article
Detección de roturas de ADN bicatenario en ovocitos de ratón
En este artículo
Resumen
Mantener la integridad del genoma del ovocito es necesario para asegurar la fidelidad genética en el embrión resultante. Aquí, presentamos un protocolo preciso para detectar roturas de doble cadena de ADN en células germinales femeninas de mamíferos.
Resumen
Los ovocitos se encuentran entre las células más grandes y longevas del cuerpo femenino. Se forman en los ovarios durante el desarrollo embrionario y permanecen detenidas en la profase de la meiosis I. El estado de reposo puede durar años hasta que los ovocitos reciben un estímulo para crecer y obtener la competencia para reanudar la meiosis. Este prolongado estado de detención los hace extremadamente susceptibles a acumular agresiones dañinas para el ADN, que afectan a la integridad genética de los gametos femeninos y, por tanto, a la integridad genética del futuro embrión.
En consecuencia, el desarrollo de un método preciso para detectar el daño en el ADN, que es el primer paso para el establecimiento de mecanismos de respuesta al daño en el ADN, es de vital importancia. En este trabajo se describe un protocolo común para evaluar la presencia y el progreso del daño en el ADN en ovocitos detenidos en profase durante un período de 20 h. Específicamente, diseccionamos ovarios de ratón, recuperamos los complejos cúmulo-ovocito (AOC), eliminamos las células del cúmulo de los COC y cultivamos los ovocitos en medio Μ2 que contiene 3-isobutil-1-metilxantina para mantener el estado de detención. A partir de entonces, los ovocitos se tratan con el fármaco antineoplásico citotóxico etopósido para generar roturas de doble cadena (DSB).
Mediante el uso de inmunofluorescencia y microscopía confocal, detectamos y cuantificamos los niveles de la proteína central γH2AX, que es la forma fosforilada de la histona H2AX. El H2AX se fosforila en los sitios de los DSB después de un daño en el ADN. La incapacidad de restaurar la integridad del ADN después de un daño en el ADN de los ovocitos puede provocar infertilidad, defectos congénitos y un aumento de las tasas de abortos espontáneos. Por lo tanto, la comprensión de los mecanismos de respuesta al daño del ADN y, al mismo tiempo, el establecimiento de un método intacto para estudiar estos mecanismos son esenciales para la investigación en biología reproductiva.
Introducción
El proceso de meiosis en las células germinales femeninas de mamíferos se inicia en los ovarios antes del nacimiento. El número total de ovocitos se establece en los ovarios principalmente durante la embriogénesis. Los ovocitos entran en meiosis y permanecen detenidos en la profase I1. Tras el inicio de la pubertad y la producción y acción endocrina de la hormona foliculoestimulante (FSH) y la hormona luteinizante (LH), los ovocitos pueden reiniciar y completar la meiosis2. En los seres humanos, la detención de la profase puede durar hasta50 años. Las divisiones celulares tras la entrada en la meiosis I son asimétricas, lo que da lugar a la producción de un pequeño cuerpo polar y un ovocito que conserva su tamaño. Por lo tanto, la mayoría de los componentes citoplasmáticos se almacenan en el ooplasma durante la embriogénesis temprana4. A continuación, los ovocitos entran en la meiosis II, sin reformar su núcleo ni descondensar sus cromosomas, y permanecen detenidos en la metafase II hasta la fecundación5.
Una característica única que distingue a los ovocitos de las células somáticas es el estado de detención en la profase I, cuando el ovocito posee un núcleo intacto (detención de vesículas germinales [GV]), denominado estadio6 de GV. Con base en la organización de la cromatina, los ovocitos en estadio GV se clasifican en dos categorías: nucléolo no rodeado (NSN) y nucléolo rodeado (SN)7,8. En los ovocitos en estadio NSN GV, la cromatina se propaga por toda la región nuclear y la transcripción es activa, mientras que en los ovocitos SN, la cromatina forma un anillo compacto que rodea el nucléolo y la transcripción es silenciosa9. Ambos tipos de ovocitos en estadio GV muestran competencia meiótica; entran en meiosis al mismo ritmo, pero los ovocitos NSN presentan baja capacidad de desarrollo y no pueden desarrollarse más allá del embrión en estadio bicelular10.
El estado prolongado de detención de la profase I aumenta la incidencia de acumulación de daño en el ADN11. Por lo tanto, los mecanismos de respuesta al daño del ADN en los ovocitos son esenciales para permitir la producción de gametos con integridad genética y para asegurar que el embrión resultante tenga un contenido cromosómico fisiológico.
Un aspecto central de la respuesta al daño del ADN es la reparación del ADN. Las principales vías para la reparación de DSB en células eucariotas incluyen la unión de extremos no homólogos (NHEJ), la recombinación homóloga (HR) y la NHEJ alternativa12,13,14,15. El NHEJ es un mecanismo más rápido pero más propenso a errores, mientras que el HR requiere más tiempo para completarse, pero tiene una alta fidelidad16.
No se conoce lo suficiente sobre los mecanismos que utilizan los ovocitos para reparar el daño en el ADN. Los estudios han demostrado que el daño en el ADN inducido en ovocitos de mamíferos adultos por el uso de agentes genotóxicos, como etopósido, doxorrubicina o UVB o radiación ionizante, no afecta el momento y las tasas de salida de la detención de la profaseI 17. Los ovocitos pueden sufrir una descomposición del VG (GVBD) incluso en presencia de niveles elevados de daño. Este daño se puede determinar mediante la observación de γH2AX. Esta forma fosforilada de H2AX (γΗ2ΑΧ) es un marcador DSB, que se encuentra en el sitio de las roturas y funciona como un andamio para ayudar a reparar los factores y las proteínas que se acumulan en los extremos rotos18.
La ausencia de detención del ciclo celular después de un daño en el ADN se debe a un punto de control de daño en el ADN insuficiente que permite que los ovocitos con ADN no reparado vuelvan a entrar en meiosis. Después de altos niveles de daño en el ADN, un punto de control puede mantener la detención de la profase a través de la activación de una vía dependiente de ATM/Chk1. La respuesta limitada de los puntos de control a los OSD se debe a la activación limitada de ATM17,19. En la fase M de la meiosis I, la investigación ha demostrado que el daño en el ADN puede activar un punto de control de la meiosis I inducido por el punto de control del ensamblaje del huso (SAC), que impide la activación del complejo/ciclosoma promotor de la anafase de la ubiquitina ligasa E3 (APC/C) y, por lo tanto, la salida de la fase M. Además, la ablación de las proteínas SAC supera el estado de detención de la fase M, lo que subraya la importancia de la SAC en el establecimiento del chekpoint de meiosisI 20.
Como muestran claramente investigaciones anteriores, los DSB no pueden inducir un punto de control de profase robusto en los ovocitos de ratón. Si dicho daño no se repara, podría dar lugar a que los embriones porten anomalías cromosómicas. Por lo tanto, es importante estudiar la respuesta al daño del ADN en diferentes etapas de la gametogénesis femenina para comprender mejor las vías únicas que utilizan los ovocitos para hacer frente a posibles agresiones genéticas.
Protocolo
Todos los experimentos con ratones fueron aprobados por las autoridades locales (Región de Ioannina, Grecia) y se llevaron a cabo de acuerdo con las Directivas 2010/63/UE del Consejo de las Comunidades Europeas. Los experimentos se llevaron a cabo con respecto a los principios de las 3R. Todos los ratones CD-1 utilizados para los experimentos se mantuvieron en las instalaciones de la casa de animales de la Universidad de Ioannina, Grecia, en una habitación con temperatura controlada (22 °C) y humedad (60%) y fueron alimentados ad libitum. El animalario tiene una licencia para operar una instalación para la cría (EL33-BIObr01), el suministro (EL33-BIOsup01) y los experimentos (EL33BIO-exp01).
1. Preparación de reactivos
- Diluir 3-isobutil-1-metilxantina (IBMX) en polvo (ver la Tabla de Materiales) en dimetilsulfóxido (DMSO) (ver la Tabla de Materiales) hasta una concentración final de 200 mM. Μake 10 μL de alícuotas y almacenar a -20 °C. Utilice la solución en el plazo de 1 mes.
NOTA: El polvo de IBMX se mantiene a -20 °C. - Prepare todos los tampones de inmunofluorescencia y guárdelos a 4 °C.
- Prepare solución salina estéril tamponada con fosfato (PBS) diluyendo un comprimido de PBS (véase la Tabla de materiales) en 200 ml de ddH2Ο.
- Prepare el tampón PHEM agregando 80 ml de ddH 2 Ο, 0,59575 g de HEPES, 1,81422 g de PIPES, 0,38035 g de EGTA y 0,04066 g de MgCl2(ver la Tabla de Materiales) mientras agita con un agitador magnético (ver la Tabla de Materiales), y simultáneamente agregue NaOH (ver la Tabla de Materiales) hasta que el pH alcance 6.9 (verifique usando un medidor de pH/ORP [ver la Tabla de Materiales]). A continuación, añadir ddH2Ο a un volumen final de 100 mL.
- Prepare el tampón paraformaldehído-Triton-X-100 (PFA-Tx-100) diluyendo el polvo de PFA (consulte la Tabla de materiales) en tampón PHEM mientras agita con un agitador magnético bajo calentamiento a una concentración final de PFA al 4%. A continuación, filtre el tampón con una jeringa y un filtro de 0,2 μm (véase la Tabla de Materiales) y añada Tx-100 al 0,5% (véase la Tabla de Materiales). Prepare aproximadamente 10 ml de PFA-Tx-100 (0,4 g de PFA, 50 μl de Tx-100), lo que es suficiente para un experimento. Guárdelo a 4 °C durante 1 semana como máximo.
PRECAUCIÓN: Use guantes para manipular PFA y evite el contacto con la piel y los ojos. - Prepare el tampón de lavado añadiendo albúmina sérica bovina (concentración final: 0,5% p/v BSA) (véase la Tabla de Materiales) en PBS, y agite mecánicamente. Agregue un 10% p/v de tampón NaN3 (azida de sodio) en una dilución de 1:1,000 para minimizar el riesgo de contaminación fúngica y bacteriana. Hacer un tampón de NaN 3 al 10% p/v añadiendo 1 g de polvo de NaN3 (ver la Tabla de Materiales) a 10 mL de ddH2O; almacene el tampón NaN3 a temperatura ambiente.
- Prepare el tampón de bloqueo agregando BSA (concentración final: 3% p/v) en PBS y agitando mecánicamente. Añadir tampón NaN3 al 10% en una dilución de 1:1.000.
2. Recolección de ovocitos GV de ovarios disecados e inducción de DSB
NOTA: Todas las herramientas y soluciones deben ser estériles. La manipulación de los ovocitos se realiza mediante el uso de una pipeta bucal bajo un microscopio estereoscópico (ver la Tabla de Materiales), y todas las gotas se cubren con aceite mineral (ver la Tabla de Materiales y la Figura 1E).
- Inyectar a los ratones por vía intraperitoneal 7 unidades internacionales (UI) de gonadotropina sérica de yegua preñada (PMSG) (ver la Tabla de Materiales) 46-48 h antes de sacrificar a los ratones por luxación cervical.
NOTA: Todos los ratones utilizados deben tener entre 8 y 12 semanas de edad. - Filtrar el medio de cultivo M2 (ver la Tabla de Materiales) con una jeringa y un filtro de 0,2 μm, y añadir IBMX 200 mM a una concentración final de 200 μM en un tubo de fondo redondo de 14 mL (ver la Tabla de Materiales) para mantener los ovocitos detenidos en la profase I. A continuación, prepare gotas de medio M2-IBMX en una placa plástica de cultivo de tejidos (ver la Tabla de Materiales) y colóquela en un bloque caliente (ver la Tabla de Materiales) a 37 °C durante al menos 30 min antes del aislamiento de los ovocitos. Almacenar el M2 a 4 °C.
- Sacrificar los ratones por dislocación cervical, diseccionar los ovarios y colocarlos en un tubo de fondo redondo de 5 mL (ver la Tabla de Materiales) con M2-IBMX.
- Transfiera los ovarios a una tapa de plástico que contenga 1.5 mL de M2-IBMX, extraiga cualquier tejido adiposo periovárico o segmentos de trompas de Falopio y libere los AOC mediante perforación mecánica de los ovarios con una aguja de 27 G (consulte la Tabla de materiales y la Figura 1A-C).
- Transfiera los AOC a una placa de cultivo con gotas de M2-IBMX (aproximadamente 25-30 μL cada una) y elimine las células del cúmulo mediante pipeteo repetido con una pipeta Pasteur de vidrio de diámetro estrecho (consulte la Tabla de materiales y la Figura 1D).
- Seleccione ovocitos en estadio SN GV y transfiéralos en una gota (25 μL) de medio M2-IBMX en un bloque caliente a 37 °C protegido de la luz (Figura 1F).
- Busque ovocitos SN en función de su mayor tamaño y núcleos ubicados centralmente, en contraste con los ovocitos NSN, en los que los núcleos están colocados periféricamente21. En cualquier caso, observar la configuración del ADN bajo un microscopio confocal antes de tomar la decisión final sobre el tipo de ovocito GV (SN o NSN).
- Inducir DSB usando etopósido (ver la Tabla de Materiales). Colocar los ovocitos en estadio GV en gotas (25 μL cada una) del agente genotóxico durante 1 h en el bloque caliente a 37 °C en condiciones de oscuridad.
NOTA: El etopósido es un inhibidor de la topoisomerasa II que introduce DSB en el ADN22. Mantener el etopósido en alícuotas de 10 μL de 20 mg/ml a temperatura ambiente, protegido de la luz. Las concentraciones que se han probado son 5 μg/mL, 20 μg/mL y 50 μg/mL. - Para mantener los ovocitos en estadio GV detenidos durante un período prolongado, colocar los ovocitos en gotas de medio de cultivo M16 (ver la Tabla de Materiales) suplementado con 400 μM de IBMX en una incubadora (ver la Tabla de Materiales) a 37 °C y 5% de CO2. Almacenar el M16 a 4 °C, filtrar el medio con una jeringa y un filtro de 0,2 μm e incubarlo durante al menos 1 h antes de su uso.

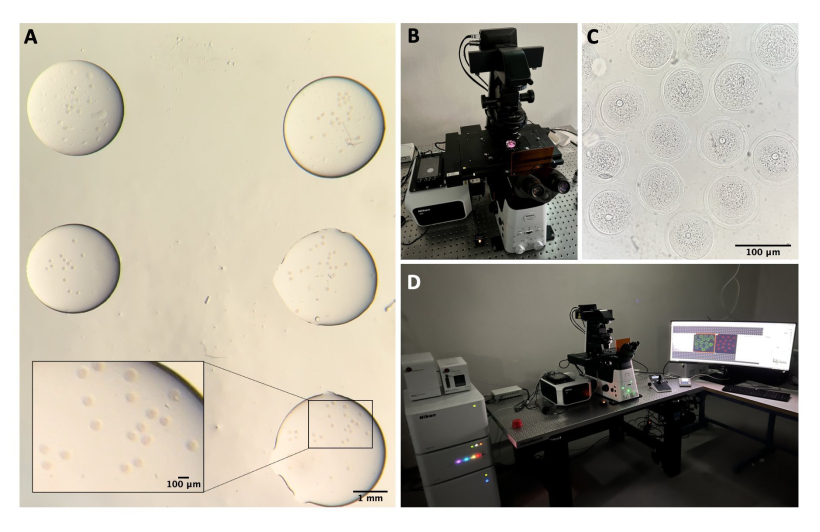
Figura 1: Proceso de aislamiento de ovocitos . (A) Extracción de tejido adiposo periovárico y restos de segmentos de trompas de Falopio de ovarios en medio M2 con IBMX. Fotografía obtenida a través de los oculares del microscopio estereoscópico. Barra de escala = 1 mm. (B) Ovarios aislados en medio M2 con IBMX. Imagen obtenida a través de los oculares del microscopio estereoscópico. Barra de escala = 1 mm. (C) Perforación mecánica de ovarios con aguja de 27 G en medio M2 con IBMX. Imagen obtenida a través de los oculares del microscopio estereoscópico. Barra de escala = 1 mm. (D) AOC liberados de los ovarios después de la perforación en medio M2 con IBMX. Imagen obtenida a través de los oculares del microscopio estereoscópico. Barra de escala = 100 μm. (E) Recolección de ovocitos con pipeta bucal. (F) Ovocitos desnudados, después de la eliminación de las células del cúmulo circundante, en medio M2 con IBMX. Imagen obtenida a través de los oculares del microscopio estereoscópico. Barra de escala = 100 μm. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
3. Fijación de ovocitos e inmunofluorescencia
NOTA: La manipulación de los ovocitos se realiza mediante el uso de una pipeta bucal bajo un microscopio estereoscópico, y todas las gotas se cubren con aceite mineral.
- Colocar los ovocitos GV control y tratados con etopósido en diferentes placas de cultivo de tejidos plásticos con tampón PFA-Tx-100 durante 40 min a temperatura ambiente.
- Lavar los ovocitos en tres gotas diferentes de tampón de lavado (50 μL cada una) a temperatura ambiente. Dejar actuar los ovocitos durante 5 min en cada gota.
- Colocar los ovocitos en gotas de tampón de bloqueo (25 μL cada una) durante 1 h sobre un bloque caliente a 37 °C.
- Preparar el anticuerpo primario que reconoce γH2AX (fosfo-Η2ΑΧ de conejo) (Ser139) (ver la Tabla de Materiales) (solución madre: 1 mg/mL). Utilizar una dilución 1:200 en tampón bloqueante y colocar los ovocitos en gotas de anticuerpo primario (15 μL cada una) a 4 °C durante la noche.
NOTA: El fosfo-Η2ΑΧ (γH2AX) es un marcador común para la detección de DSB tanto en células somáticas como en ovocitos GV18,23. - Al día siguiente, lavar los ovocitos en tres gotas diferentes de tampón de lavado (50 μL cada una) a temperatura ambiente. Dejar actuar los ovocitos durante 5 min en cada gota.
- Prepare el anticuerpo secundario, Alexa Fluor 488-conjugado cabra anti-conejo (ver la Tabla de Materiales) (solución madre: 2 mg/mL). Utilizar una dilución 1:200 en tampón bloqueante, y colocar los ovocitos en gotas de anticuerpo secundario (15 μL cada una) durante 1 h sobre un bloque caliente a 37 °C protegido de la luz.
- Transferir los ovocitos a gotas de DRAQ7 (25 μL cada una) (solución madre: 0,3 mM; ver la Tabla de Materiales), que es un colorante de ADN fluorescente de color rojo lejano que solo tiñe el ADN en células permeabilizadas. Utilice una dilución 1:250 en tampón de lavado durante 10 minutos a temperatura ambiente en condiciones de oscuridad.
- Lavar los ovocitos en tres gotas diferentes de tampón de lavado (50 μL cada una) a temperatura ambiente. Déjelos durante 5 minutos en cada gota y luego transfiéralos a pequeñas gotas (aproximadamente 5 μL cada una) de tampón de lavado en una placa de Petri con fondo de vidrio de 35 mm (consulte la Tabla de materiales) para microscopía confocal (Figura 2A).
NOTA: El lavado tanto de la tinción de ADN como del anticuerpo secundario se realiza al mismo tiempo.
4. Microscopía confocal
NOTA: La microscopía confocal debe realizarse inmediatamente para evitar la reducción de la intensidad de fluorescencia después de la colocación de los ovocitos en placas de fondo de vidrio. Se requiere acceso a un microscopio confocal (ver la Tabla de Materiales) con una platina motorizada.
- Configuración del microscopio
- En el sistema confocal, encienda el controlador láser, los láseres, el controlador del microscopio, las lámparas para la luz transmitida y la PC (Figura 2B, D).
- Abra el software confocal y elija la lente de aceite 40x.
- Coloque la placa en el portamuestras e intente enfocar los ovocitos moviendo la platina en los ejes XY y Z con el joystick (Figura 2C).
- Exploración de los ovocitos
- Ajuste la potencia del láser, la ganancia y el tamaño del agujero de alfiler de forma independiente para cada experimento con el fin de minimizar cualquier saturación.
- Para cada ovocito, establezca el área de interés, específicamente en el núcleo en el área del ADN. Defina los bordes del área de ADN y ajuste el tamaño del paso z a 3 μm. A continuación, inicie el escaneo.
- Guarde las imágenes de cada celda de la carpeta seleccionada.
- Cuando se complete el escaneo, salga del software, apague la computadora y apague el controlador láser, los láseres, el controlador del microscopio y las lámparas de la luz transmitida.

Figura 2: Microscopía confocal. (A) Ovocitos fijados después de realizar el protocolo de inmunofluorescencia y tinción de ADN, que se encuentran en gotas separadas de tampón de lavado, cubiertos con aceite mineral, colocados en un plato con fondo de vidrio y preparados para la obtención de imágenes de microscopía confocal. Cada gota contiene una categoría experimental diferente. Imagen obtenida a través de los oculares del microscopio estereoscópico. Barra de escala = 1 mm/100 μm para la parte ampliada. (B) Placa con fondo de vidrio colocada en la platina del microscopio confocal. (C) Imagen de campo claro de ovocitos obtenida mediante microscopía confocal. Barra de escala = 100 μm. (D) El sistema de microscopía confocal. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
5. Análisis de imágenes
- Descargue Fiji ImageJ-win64 en el navegador (https://imagej.net/software/fiji/downloads), ábralo e importe los datos como archivos de pila TIFF.
NOTA: Abra cada archivo de ovocitos por separado. - Haga clic en la imagen | Color | Dividir canales para dividir todos los canales.
- Haga clic en LUT (Tabla de búsqueda) y elija los colores preferidos para cada canal.
- Haga clic en la imagen | Color | Fusionar canales para fusionar los canales de γΗ2ΑΧ y ADN. Deje el canal de campo claro sin fusionar.
- En los ovocitos NSN y en los ovocitos SN con bajos niveles de daño en el ADN, γΗ2ΑΧ se detecta como focos en la región del ADN. En este caso, haga clic en el "Multipunto" o en el comando de punto, y seleccione cada foco γΗ2ΑΧ que coincida con el ADN. Repita este paso para todas las pilas.
- En los ovocitos SN con altos niveles de daño en el ADN, la señal γΗ2ΑΧ se distribuye por toda la región del ADN. En este caso, haga clic en Imagen | Pilas | Z y, con el comando Selecciones a mano alzada , seleccione toda el área de ADN.
- Para medir la fluorescencia γΗ2ΑΧ, haga clic en Analizar | Mida y copie las medidas en un archivo .xlsx. A continuación, calcule la fluorescencia media, normalice los valores y cuente el número de focos antes de crear cualquier gráfico.
- Haga clic en Analizar | Establecer escala para establecer la escala y, a continuación, en Analizar | Herramientas | Barra de escala para agregar una barra de escala a los canales.
Resultados
Utilizando el procedimiento que se muestra aquí, se diseccionaron los ovarios de ratón, se eliminó la grasa y se recolectaron ovocitos completamente desarrollados en etapa GV. A continuación, las células del cúmulo se extrajeron mediante pipeteo repetitivo con una pipeta estrecha y se colocaron en gotas frescas de medio M2-IBMX y se cubrieron con aceite mineral en un bloque caliente (37 °C) (Figura 1A-F). Se prepararon tres concentraciones diferentes de etopósido (5 μg/mL, 20 μg/mL y 50 μg/mL) utilizando una concentración de etopósido de 20 mg/mL. Los ovocitos en estadio GV se colocaron en tres concentraciones distintas de etopósido durante 1 h en gotas cubiertas con aceite mineral y protegidas de la luz a 37 °C. A continuación, se siguió el protocolo de inmunofluorescencia, tal y como se describe en detalle en el apartado de protocolo, y los ovocitos se colocaron en placas de fondo de cristal y se observaron mediante microscopía confocal (Figura 2).
En los ovocitos en estadio SN GV, inmediatamente después del daño en el ADN, la presencia de γH2AX aumentó en todas las concentraciones de etopósido (5 μg/mL, 20 μg/mL y 50 μg/mL), y el γH2AX se distribuyó por toda la región del ADN (Figura 3). La cuantificación y estimación de DSB se realizó mediante la observación de la intensidad de fluorescencia γH2AX en sitios de ADN. La fluorescencia γH2AX se intensificó proporcionalmente con el aumento de las concentraciones de etopósido. Además, tras una parada prolongada de la profase (20 h tras el tratamiento con etopósido), los ovocitos en estadio GV mostraron la capacidad de reducir el número y la intensidad de los focos γH2AX, lo que implica la presencia de procesos de reparación activa en los ovocitos detenidos en estadio GV (Figura 3E).
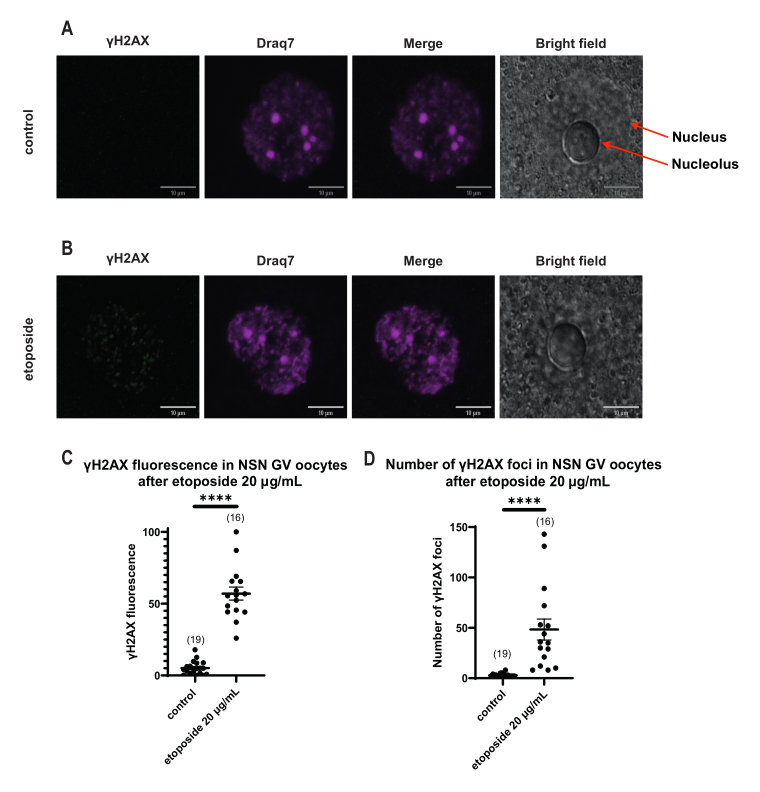
A diferencia de los ovocitos SN, en los que la fluorescencia γH2AX se distribuyó a través del ADN, en los ovocitos NSN, γH2AX se mostró en los focos inmediatamente después del tratamiento con etopósido a 20 μg/mL. Se estimó el número de focos que coincidían con el área de ADN, se calculó la fluorescencia de cada foco y se presentó la fluorescencia media de todos los ovocitos. Tanto la fluorescencia como el número de focos mostraron diferencias estadísticamente significativas entre las dos categorías de ovocitos (Figura 4).
La microscopía confocal proporciona información sobre el número y la intensidad de los focos en diferentes pilas Z, lo que ayuda a identificar la presencia de daños en el ADN y la dinámica de reparación en distintos puntos temporales. El escaneo Galvano proporciona un escaneo de precisión con un fondo bajo y un mejor análisis de las imágenes escaneadas.

Figura 3: Reducción de γH2AX en ovocitos en estadio SN GV tratados con tres concentraciones diferentes de etopósido después de un paro prolongado del GV. (A) Fluorescencia γH2AX en ovocitos en estadio SN GV 0 h después del tratamiento con etopósido. El γH2AX aumenta inmediatamente después de la exposición en todas las concentraciones de etopósido, y el aumento depende de la concentración (verde: γΗ2ΑΧ, magenta: ADN). Las imágenes son proyecciones Z-stack, y el brillo/contraste se ha ajustado para cada canal utilizando Fiji / ImageJ. Barra de escala = 10 μm. (B) Gráfico de la fluorescencia γH2AX en ovocitos en estadio SN GV 0 h después del tratamiento con distintas concentraciones de etopósido. Los datos representan la media ± SEM. Cada punto representa un ovocito (el número de ovocitos se muestra en el gráfico), (ns = no significativo, ** p < 0,005, **** p < 0,0001, ANOVA de un factor con la prueba de comparaciones múltiples de Tukey). (C) Fluorescencia γH2AX en ovocitos en estadio SN GV 20 h después del tratamiento con etopósido. γH2AX reduce 20 h después de la exposición a todas las concentraciones de etopósido (verde: γΗ2ΑΧ, magenta: ADN). Las imágenes son proyecciones de pila Z, y el brillo/contraste se ha ajustado para cada canal utilizando Fiji/ImageJ. Barra de escala = 10 μm. (D) Gráfico de la fluorescencia γH2AX en ovocitos en estadio SN GV 20 h después del tratamiento con distintas concentraciones de etopósido. Los datos representan la media ± SEM. Cada punto representa un ovocito (el número de ovocitos se muestra en el gráfico), (ns = no significativo, * p < 0,05, ** p < 0,005, *** p < 0,0005, **** p < 0,0001, ANOVA de un factor con la prueba de comparaciones múltiples de Tukey). (E) Gráfico de barras de la reducción de la fluorescencia γH2AX en ovocitos en estadio SN GV después de la detención de la profase en ovocitos tratados con etopósido. El número que aparece encima de cada columna indica el porcentaje de disminución de la fluorescencia γH2AX. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 4: Fosforilación de Η2ΑΧ en ovocitos en estadio NSN GV tras el tratamiento con etopósido a 20 μg/mL. (A) Imágenes confocales representativas de un ovocito de control en estadio NSN GV (verde: γΗ2ΑΧ, magenta: ADN). Las imágenes son proyecciones de pila Z, y el brillo/contraste se ha ajustado para cada canal utilizando Fiji/ImageJ. Barra de escala = 10 μm. (B) Imágenes confocales representativas de un ovocito en estadio GV de NSN tratado con etopósido (verde: γΗ2ΑΧ, magenta: ADN). Los ovocitos se fijaron 0 h después del tratamiento con etopósido. Las imágenes son proyecciones de pila Z, y el brillo/contraste se ha ajustado para cada canal utilizando Fiji/ImageJ. Barra de escala = 10 μm. (C) La fluorescencia normalizada γΗ2ΑΧ en ovocitos en estadio NSN GV después del tratamiento con etopósido de 20 μg/mL. Los datos representan la media ± SEM. Cada punto representa un ovocito (el número de ovocitos se muestra en el gráfico), tomado de dos experimentos independientes (**** p < 0,0001, prueba t no paramétrica no apareada, prueba U de Mann-Whitney). (D) Número de focos γΗ2ΑΧ en ovocitos en estadio NSN GV después de un tratamiento con etopósido de 20 μg/mL. Los datos representan la media ± SEM. Cada punto representa un ovocito (el número de ovocitos se muestra en el gráfico), tomado de dos experimentos independientes (**** p < 0,0001, prueba t no paramétrica no pareada, prueba U de Mann-Whitney). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Discusión
Mediante el método aquí descrito, detectamos DSB en ovocitos de mamíferos. Este método permite la detección y el estudio del proceso de reparación del ADN en los ovocitos. El mismo protocolo también podría utilizarse para analizar otras proteínas que participan en procesos fisiológicos en ovocitos de mamíferos. Es importante estudiar cómo responden los ovocitos a posibles daños en el ADN para comprender mejor la causa de la subfertilidad femenina en humanos.
El estudio de la respuesta al daño del ADN en ovocitos de mamíferos puede ser un desafío debido a la sensibilidad de los ovocitos. La manipulación de ovocitos requiere temperaturas específicas y concentraciones de CO2 yO2 . Al mismo tiempo, los ovocitos deben protegerse de la luz. Toda la manipulación debe realizarse utilizando pipetas de vidrio que no sean demasiado estrechas, ya que esto podría ser perjudicial para los ovocitos, pero tampoco demasiado anchas, ya que esto podría provocar la dilución del medio y, por lo tanto, afectar negativamente al procedimiento de fijación. En cada paso de la fijación, se utilizan varias gotas de tampones para minimizar el efecto de dilución. Una forma alternativa de observar los DSB es el ensayo del cometa24. Aunque esta técnica es más sensible, es más complicada. Al mismo tiempo, mediante el uso del ensayo Comet, no es posible detectar la región exacta del ADN donde se produce el daño, y en células con abundantes moléculas de ARN, como los ovocitos en estadio GV25, el fondo podría aumentar, lo que llevaría a una falsa señal de daño en el ADN26.
Mediante el uso del protocolo de inmunofluorescencia descrito aquí, podemos detectar DSB con precisión y estimar el progreso de la reparación en ovocitos en estadio GV, como lo indica la reducción de la fluorescencia γH2AX a lo largo del tiempo. Sin embargo, una limitación de este método es que ciertos anticuerpos pueden presentar una distribución inespecífica a lo largo del ooplasma, lo que da lugar a imágenes con alta fluorescencia de fondo. Se utiliza el tampón PFA-Tx-100 en lugar de PFA secuencial y Tx-100, ya que hemos observado que mejora el proceso de fijación al permitir la detección de menos fluorescencia de fondo e inespecífica. Una segunda limitación del uso de γH2AX para la detección de DSB es que el daño no se puede estimar después de la GVBD debido a la fosforilación espontánea de γH2AX en la meiosis23.
En este protocolo de inmunofluorescencia, los ovocitos permanecen en un tampón líquido y no pueden almacenarse dentro de portaobjetos. Este hecho dificulta la conservación de las células fijas durante días después de la adición del anticuerpo secundario. Para obtener imágenes de buena calidad y no perder la señal, es preferible realizar la imagen a las pocas horas de la adición del anticuerpo secundario. También hay que tener en cuenta que el escaneo de los núcleos a través del eje Z podría hacer que la señal se debilite debido a la sobreexposición. Por esa razón, es preferible reducir la potencia del láser y aumentar la velocidad de escaneo.
Por último, otra limitación del protocolo de inmunofluorescencia es que solo se puede utilizar para células fijas/no vivas. Por lo tanto, podemos estimar solo la presencia y ausencia de factores en puntos de tiempo específicos sin saber si hay fluctuaciones en su concentración o cambios en su comportamiento a lo largo del tiempo. Este problema podría superarse mediante el uso de imágenes de células vivas y marcadores marcados con fluorescencia.
Divulgaciones
Los autores no tienen conflictos de intereses.
Agradecimientos
Agradecemos el apoyo a este trabajo del proyecto "Establecimiento de infraestructuras de 'desarrollo de capacidades' en Investigación Biomédica (BIOMED-20)" (MIS 5047236), que se implementa en el marco de la Acción "Refuerzo de la Infraestructura de Investigación e Innovación", financiada por el Programa Operativo "Competitividad, Emprendimiento e Innovación" (NSRF 2014-2020), y cofinanciada por Grecia y la Unión Europea (Fondo Europeo de Desarrollo Regional).
Materiales
| Name | Company | Catalog Number | Comments |
| 3 mL Pasteur pipettes in LDPE, graduated | APTACA | 1502 | |
| 10 cc syringes | SoftCare | 114.104.21 | |
| Alexa Fluor 488-conjugated goat anti-rabbit Secondary Ab | Biotium | 20012 | |
| Anti-phospho-H2A.X (Ser139) | Merck Millipore | 07-164 | |
| ARE Heating Magnetic Stirrer | VELP Scientifica | F20500162 | |
| BD FALCON 5 mL Polystyrene Round-Bottom Tubes | BD Biosciences | 352054 | |
| BD Microlance 3 Needles 27 G - 0.40 x 13 mm | Becton Dickinson | 300635 | |
| Bovine Serum Albumin Fraction V | Roche | 10735078001 | |
| DMSO Anhydrous | Biotium | 90082 | |
| DRAQ7 DNA dye | BioStatus | DR71000 | |
| EGTA | Sigma-Aldrich | E4378-25G | |
| EMSURE MgCl2. 6H2O | Merck Millipore | 1058330250 | |
| Etoposide | CHEMIPHARM | L01CB01 | |
| FALCON 14 mL Polystyrene Round-Bottom Tubes | Corning Science | 532057 | |
| FALCON Tissue Culture Dishes, Easy-Grip, 35 x 10 mm Style | Corning Science | 353001 | |
| Glass Bottom Culture Dishes (35 mm Petri dish/ 14 mm Microwell, No. 0 coverglass) | MatTek Corporation | P35G-0-14-C | |
| HEPES | Sigma-Aldrich | H6147-25G | |
| HERACELL 150i CO2 Incubator | ThermoFisher Scientific | 50116048 | |
| IBMX powder | Sigma-Aldrich | I5879-100MG | |
| Leica M125 Stereo Microscope | Leica Microsystems | ||
| M16 Medium | Sigma-Aldrich | M7292 | |
| M2 Medium | Sigma-Aldrich | M7167 | |
| Mineral Oil | Sigma-Aldrich | M5310 | |
| NaN3 | Honeywell | 13412H | |
| NaOH | Merck Millipore | 1064981000 | |
| Nikon AX ECLIPSE Ti2 Confocal Microscope | Nikon Corporation | ||
| Nikon SMZ800N Stereo Microscope | Nikon Corporation | ||
| Paraformaldehyde | Sigma-Aldrich | 158127 | |
| Pasteur pippettes, glass, long form 230 mm | DURAN WHEATON KIMBLE | 357335 | |
| pH/ORP meter | Hanna Instruments Ltd | HI2211 | |
| Phosphate buffered saline tablets | Sigma-Aldrich | P4417-100TAB | |
| PIPES | Sigma-Aldrich | P1851 | |
| PMSG Protein Lyophilised | Genway Biotech (now AVIVA Systems Biology) | GWB-2AE30A (now OPPA01037) | |
| QBD4 Dry block heater | Grant Instruments (Cambridge) Ltd | A25218 | |
| Triton X-100 | Sigma-Aldrich | T8787 | |
| Whatman Puradisc 25 mm 0.2 μm filters | GE Healthcare | 6780-2502 |
Referencias
- Wang, X., Pepling, M. E. Regulation of meiotic prophase one in mammalian oocytes. Frontiers in Cell and Developmental Biology. 9, 667306(2021).
- Filatov, M., Khramova, Y., Semenova, M. Molecular mechanisms of prophase I meiotic arrest maintenance and meiotic resumption in mammalian oocytes. Reproductive Sciences. 26 (11), 1519-1537 (2019).
- Adhikari, D., et al. Inhibitory phosphorylation of Cdk1 mediates prolonged prophase I arrest in female germ cells and is essential for female reproductive lifespan. Cell Research. 26 (11), 1212-1225 (2016).
- Sun, S. C., Kim, N. H. Molecular mechanisms of asymmetric division in oocytes. Microscopy and Microanalysis. 19 (4), 883-897 (2013).
- Jones, K. T. Mammalian egg activation: From Ca2+ spiking to cell cycle progression. Reproduction. 130 (6), 813-823 (2005).
- Solc, P., Schultz, R. M., Motlik, J. Prophase I arrest and progression to metaphase I in mouse oocytes: comparison of resumption of meiosis and recovery from G2-arrest in somatic cells. Molecular Human Reproduction. 16 (9), 654-664 (2010).
- Parfenov, V., Potchukalina, G., Dudina, L., Kostyuchek, D., Gruzova, M. Human antral follicles: oocyte nucleus and the karyosphere formation (electron microscopic and autoradiographic data). Gamete Research. 22 (2), 219-231 (1989).
- Zuccotti, M., Piccinelli, A., Giorgi Rossi, P., Garagna, S., Redi, C. A. Chromatin organization during mouse oocyte growth. Molecular Reproduction and Development. 41 (4), 479-485 (1995).
- Sun, X., et al. Comprehensive analysis of nonsurrounded nucleolus and surrounded nucleolus oocytes on chromatin accessibility using ATAC-seq. Molecular Reproduction and Development. 90 (2), 87-97 (2023).
- Zuccotti, M., Bellone, M., Longo, F., Redi, C. A., Garagna, S. Fully-mature antral mouse oocytes are transcriptionally silent but their heterochromatin maintains a transcriptional permissive histone acetylation profile. Journal of Assisted Reproduction and Genetics. 28 (12), 1193-1196 (2011).
- Winship, A. L., Stringer, J. M., Liew, S. H., Hutt, K. J. The importance of DNA repair for maintaining oocyte quality in response to anti-cancer treatments, environmental toxins and maternal ageing. Human Reproduction Update. 24 (2), 119-134 (2018).
- Lieber, M. R. The mechanism of human nonhomologous DNA end joining. The Journal of Biological Chemistry. 283 (1), 1-5 (2008).
- Chiruvella, K. K., Liang, Z., Wilson, T. E. Repair of double-strand breaks by end joining. Cold Spring Harbor Perspectives in Biology. 5 (5), 012757(2013).
- Chang, H. H. Y., Pannunzio, N. R., Adachi, N., Lieber, M. R. Non-homologous DNA end joining and alternative pathways to double-strand break repair. Nature Reviews. Molecular Cell Biology. 18 (8), 495-506 (2017).
- Shibata, A., Jeggo, P. A. Roles for the DNA-PK complex and 53BP1 in protecting ends from resection during DNA double-strand break repair. Journal of Radiation Research. 61 (5), 718-726 (2020).
- Mohiuddin, I. S., Kang, M. H. DNA-PK as an emerging therapeutic target in cancer. Frontiers in Oncology. 9, 635(2019).
- Marangos, P., Carroll, J. Oocytes progress beyond prophase in the presence of DNA damage. Current Biology. 22 (11), 989-994 (2012).
- Paull, T. T., et al. A critical role for histone H2AX in recruitment of repair factors to nuclear foci after DNA damage. Current Biology. 10 (15), 886-895 (2000).
- Bakkenist, C. J., Kastan, M. B. DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation. Nature. 421 (6922), 499-506 (2003).
- Marangos, P., et al. DNA damage-induced metaphase I arrest is mediated by the spindle assembly checkpoint and maternal age. Nature Communications. 6, 8706(2015).
- Lavrentyeva, E. A., Shishova, K. V., Zatsepina, O. V. Differences in nuclear dynamics in mouse GV oocytes with a diverse chromatin configuration. Biology Bulletin Russian Academy of Sciences. 46, 332-341 (2019).
- Montecucco, A., Zanetta, F., Biamonti, G. Molecular mechanisms of etoposide. EXCLI Journal. 14, 95-108 (1998).
- Mayer, A., et al. DNA damage response during mouse oocyte maturation. Cell Cycle. 15 (4), 546-558 (2016).
- Olive, P., Banáth, J. The comet assay: A method to measure DNA damage in individual cells. Nature Protocols. 1, 23-29 (2006).
- Wu, D., Dean, J. EXOSC10 sculpts the transcriptome during the growth-to-maturation transition in mouse oocytes. Nucleic Acids Research. 48 (10), 5349-5365 (2020).
- Simon, L., Emery, B., Carrell, D. DNA damage: COMET assay. Manual of Sperm Function Testing in Human Assisted Reproduction. Agarwal, A., Henkel, R., Majzoub, A. , 202-212 (2021).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados