
Method Article
Détection de cassures double brin de l’ADN dans des ovocytes de souris
Dans cet article
Résumé
Le maintien de l’intégrité du génome de l’ovocyte est nécessaire pour assurer la fidélité génétique de l’embryon résultant. Nous présentons ici un protocole précis pour détecter les cassures double brin de l’ADN dans les cellules germinales femelles de mammifères.
Résumé
Les ovocytes sont parmi les cellules les plus grandes et les plus durables du corps féminin. Ils se forment dans les ovaires au cours du développement embryonnaire et restent arrêtés à la prophase de la méiose I. L’état de quiétude peut durer des années jusqu’à ce que les ovocytes reçoivent un stimulus pour se développer et obtenir la compétence nécessaire pour reprendre la méiose. Cet état d’arrêt prolongé les rend extrêmement susceptibles d’accumuler des insultes préjudiciables à l’ADN, qui affectent l’intégrité génétique des gamètes femelles et, par conséquent, l’intégrité génétique du futur embryon.
Par conséquent, le développement d’une méthode précise pour détecter les dommages à l’ADN, qui est la première étape pour l’établissement de mécanismes de réponse aux dommages à l’ADN, est d’une importance vitale. Cet article décrit un protocole commun pour tester la présence et la progression des dommages à l’ADN dans les ovocytes arrêtés en prophase pendant une période de 20 h. Plus précisément, nous disséquons des ovaires de souris, récupérons les complexes cumulus-ovocytes (COC), retirons les cellules cumulus des COC et cultivons les ovocytes dans un milieu Μ2 contenant de la 3-isobutyl-1-méthylxanthine pour maintenir l’état d’arrêt. Par la suite, les ovocytes sont traités avec le médicament cytotoxique et antinéoplasmique, l’étoposide, pour engendrer des cassures double brin (DSB).
En utilisant l’immunofluorescence et la microscopie confocale, nous détectons et quantifions les niveaux de la protéine centrale γH2AX, qui est la forme phosphorylée de l’histone H2AX. H2AX devient phosphorylé sur les sites des DSB après des dommages à l’ADN. L’incapacité à restaurer l’intégrité de l’ADN à la suite d’une lésion de l’ADN dans les ovocytes peut entraîner l’infertilité, des malformations congénitales et une augmentation des taux d’avortements spontanés. Par conséquent, la compréhension des mécanismes de réponse aux dommages de l’ADN et, en même temps, la mise en place d’une méthode intacte pour étudier ces mécanismes sont essentielles pour la recherche en biologie de la reproduction.
Introduction
Le processus de méiose dans les cellules germinales femelles de mammifères est initié dans les ovaires avant la naissance. Le nombre total d’ovocytes est établi dans les ovaires principalement au cours de l’embryogenèse. Les ovocytes entrent en méiose et restent arrêtés à la prophaseI 1. Après le début de la puberté et la production et l’action endocrinienne de l’hormone folliculo-stimulante (FSH) et de l’hormone lutéinisante (LH), les ovocytes peuvent relancer et terminer la méiose2. Chez l’homme, l’arrêt de la prophase peut durer jusqu’à 50 ans3. Les divisions cellulaires qui suivent l’entrée en méiose I sont asymétriques, ce qui entraîne la production d’un petit corps polaire et d’un ovocyte qui conserve sa taille. Ainsi, la plupart des composants cytoplasmiques sont stockés dans l’ooplasme au début de l’embryogenèse4. Ensuite, les ovocytes entrent en méiose II, sans reformer leur noyau ni décondenser leurs chromosomes, et restent arrêtés à la métaphase II jusqu’à la fécondation5.
Une caractéristique unique qui distingue les ovocytes des cellules somatiques est l’état d’arrêt en prophase I, lorsque l’ovocyte possède un noyau intact (arrêt de la vésicule germinale [GV]), appelé stadeGV 6. Sur la base de l’organisation de la chromatine, les ovocytes de stade GV sont classés en deux catégories : le nucléole non entouré (NSN) et le nucléole entouré (SN)7,8. Dans les ovocytes au stade NSN GV, la chromatine se propage dans toute la région nucléaire et la transcription est active, tandis que dans les ovocytes SN, la chromatine forme un anneau compact qui entoure le nucléole, et la transcription est silencieuse9. Les deux types d’ovocytes au stade GV présentent une compétence méiotique ; ils entrent en méiose au même rythme, mais les ovocytes NSN présentent une faible capacité de développement et ne peuvent pas se développer au-delà du stade embryonnaire à deux cellules10.
L’état prolongé d’arrêt de la prophase I augmente l’incidence de l’accumulation de dommages à l’ADN11. Par conséquent, les mécanismes de réponse aux dommages de l’ADN dans les ovocytes sont essentiels pour permettre la production de gamètes avec une intégrité génétique et pour garantir que l’embryon résultant a un contenu chromosomique physiologique.
Un aspect central de la réponse aux dommages de l’ADN est la réparation de l’ADN. Les principales voies de réparation de la DSB dans les cellules eucaryotes comprennent la jonction d’extrémité non homologue (NHEJ), la recombinaison homologue (HR) et la NHEJalternative 12,13,14,15. Le NHEJ est un mécanisme plus rapide mais plus sujet aux erreurs, tandis que le HR nécessite plus de temps pour être complété mais a une haute fidélité16.
Il n’y a pas assez de connaissances sur les mécanismes que les ovocytes utilisent pour réparer les dommages à l’ADN. Des études ont montré que les dommages à l’ADN induits dans les ovocytes de mammifères adultes par l’utilisation d’agents génotoxiques, tels que l’étoposide, la doxorubicine, les UVB ou les rayonnements ionisants, n’affectent pas le moment et les taux de sortie de l’arrêt de la prophaseI 17. Les ovocytes peuvent subir une dégradation de la GV (GVBD) même en présence de niveaux élevés de dommages. Ces dommages peuvent être déterminés par l’observation de γH2AX. Cette forme phosphorylée de H2AX (γΗ2ΑΧ) est un marqueur DSB, qui est situé sur le site des cassures et fonctionne comme un échafaudage pour aider les facteurs de réparation et les protéines à s’accumuler aux extrémités cassées18.
L’absence d’arrêt du cycle cellulaire à la suite d’une lésion de l’ADN est due à un point de contrôle insuffisant des dommages à l’ADN qui permet aux ovocytes dont l’ADN n’est pas réparé de rentrer dans la méiose. À la suite de niveaux élevés de dommages à l’ADN, un point de contrôle peut maintenir l’arrêt de la prophase grâce à l’activation d’une voie dépendante de l’ATM/Chk1. La réponse limitée des points de contrôle aux ORD est due à l’activation limitée de l’ATM17,19. Dans la phase M de la méiose I, la recherche a montré que les dommages à l’ADN peuvent activer un point de contrôle de la méiose I induite par le point de contrôle de l’assemblage du fuseau (SAC), ce qui empêche l’activation du complexe promoteur de l’anaphase/cyclosome E3 ubiquitine ligase (APC/C) et, par conséquent, la sortie de la phase M. De plus, l’ablation des protéines SAC permet de surmonter l’état d’arrêt de la phase M, soulignant ainsi l’importance de la SAC dans l’établissement du point de chékpoint20 de la méiose I.
Comme le montrent clairement des recherches antérieures, les DSB ne peuvent pas induire un point de contrôle de prophase robuste dans les ovocytes de souris. Si ces dommages ne sont pas réparés, ils peuvent conduire à des embryons porteurs d’anomalies chromosomiques. Par conséquent, il est important d’étudier la réponse aux dommages de l’ADN à différents stades de la gamétogenèse féminine afin de mieux comprendre les voies uniques que les ovocytes utilisent pour faire face aux agressions génétiques potentielles.
Protocole
Toutes les expériences sur les souris ont été approuvées par les autorités locales (région de Ioannina, Grèce) et menées conformément aux directives 2010/63/UE du Conseil des Communautés européennes. Des expériences ont été menées dans le respect des principes des 3R. Toutes les souris CD-1 utilisées pour les expériences ont été gardées dans l’animalerie de l’Université de Ioannina, en Grèce, dans une pièce à température contrôlée (22 °C) et à humidité (60%) et ont été nourries ad libitum. L’animalerie dispose d’une licence pour l’exploitation d’une installation d’élevage (EL33-BIObr01), d’approvisionnement (EL33-BIOsup01) et d’expérimentations (EL33BIO-exp01).
1. Préparation des réactifs
- Diluer la poudre de 3-isobutyl-1-méthylxanthine (IBMX) (voir le tableau des matériaux) dans du diméthylsulfoxyde (DMSO) (voir le tableau des matériaux) jusqu’à une concentration finale de 200 mM. Μake 10 μL aliquotes, et conserver à −20 °C. Utilisez la solution dans un délai de 1 mois.
REMARQUE : La poudre IBMX est conservée à −20 °C. - Préparez tous les tampons d’immunofluorescence et conservez-les à 4 °C.
- Préparer une solution saline stérile tamponnée au phosphate (PBS) en diluant un comprimé de PBS (voir le tableau des matériaux) dans 200 mL de ddH2Ο.
- Fabriquer un tampon PHEM en ajoutant 80 mL de ddH 2 Ο,0,59575 g de HEPES, 1,81422 g de PIPES, 0,38035 g d’EGTA et 0,04066 g de MgCl2 (voir le tableau des matériaux) tout en agitant avec un agitateur magnétique (voir le tableau des matériaux), et ajouter simultanément du NaOH (voir le tableau des matériaux) jusqu’à ce que le pH atteigne 6,9 (vérifier à l’aide d’un pH-mètre/ORP [voir le tableau des matériaux]). Ensuite, ajouter ddH2Ο à un volume final de 100 mL.
- Préparer le tampon de paraformaldéhyde-Triton-X-100 (PFA-Tx-100) en diluant de la poudre de PFA (voir le tableau des matériaux) dans le tampon PHEM tout en agitant avec un agitateur magnétique sous chauffage à une concentration finale de 4 % de PFA. Ensuite, filtrez le tampon à l’aide d’une seringue et d’un filtre de 0,2 μm (voir le tableau des matériaux) et ajoutez 0,5 % de Tx-100 (voir le tableau des matériaux). Préparez environ 10 mL de PFA-Tx-100 (0,4 g de PFA, 50 μL de Tx-100), ce qui est suffisant pour une expérience. Conservez-le à 4 °C pendant 1 semaine maximum.
ATTENTION: Portez des gants pour manipuler le PFA et évitez tout contact avec la peau et les yeux. - Préparer le tampon de lavage en ajoutant de l’albumine sérique bovine (concentration finale : 0,5 % p/v de BSA) (voir le tableau des matériaux) dans du PBS et agiter mécaniquement. Ajouter 10 % p/v de tampon NaN3 (azoture de sodium) à une dilution de 1:1 000 pour minimiser le risque de contamination fongique et bactérienne. Obtenir un tampon de NaN 3 à 10 % p/v en ajoutant 1 g de poudre de NaN3 (voir le tableau des matériaux) à 10 mL de ddH2O; Conservez le tampon NaN3 à température ambiante.
- Préparer le tampon de blocage en ajoutant du BSA (concentration finale : 3 % p/v) dans du PBS et en agitant mécaniquement. Ajouter 10 % de tampon NaN3 à une dilution de 1:1 000.
2. Prélèvement d’ovocytes GV à partir d’ovaires disséqués et induction de DSB
REMARQUE: Tous les outils et solutions doivent être stériles. La manipulation des ovocytes est effectuée à l’aide d’une pipette buccale sous un microscope stéréoscopique (voir le tableau des matériaux), et toutes les gouttes sont recouvertes d’huile minérale (voir le tableau des matériaux et la figure 1E).
- Injecter à des souris par voie intrapéritonéale 7 unités internationales (UI) de gonadotrophine sérique (PMSG) de jument gestante (voir le tableau des matériaux) 46 à 48 h avant d’éliminer les souris par luxation cervicale.
REMARQUE: Toutes les souris utilisées doivent être âgées de 8 à 12 semaines. - Filtrer le milieu de culture M2 (voir le tableau des matériaux) à l’aide d’une seringue et d’un filtre de 0,2 μm, et ajouter IBMX 200 mM à une concentration finale de 200 μM dans un tube à fond rond de 14 mL (voir le tableau des matériaux) pour maintenir les ovocytes arrêtés à la prophase I. Ensuite, préparez des gouttes de milieu M2-IBMX dans une boîte de culture de tissus en plastique (voir le tableau des matériaux) et placez-les sur un bloc chaud (voir le tableau des matériaux) à 37 °C pendant au moins 30 minutes avant l’isolement des ovocytes. Conservez le M2 à 4 °C.
- Sacrifiez les souris par luxation cervicale, disséquez les ovaires et placez-les dans un tube à fond rond de 5 ml (voir le tableau des matériaux) avec M2-IBMX.
- Transférez les ovaires dans un couvercle en plastique contenant 1,5 mL de M2-IBMX, retirez tout tissu adipeux périovarien ou segment de trompe de Fallope et libérez les COC par perforation mécanique des ovaires avec une aiguille de 27 G (voir le tableau des matériaux et la figure 1A-C).
- Transvaser les COC dans une boîte de culture avec des gouttes de M2-IBMX (environ 25 à 30 μL chacune) et retirer les cellules de cumulus par pipetage répété à l’aide d’une pipette Pasteur en verre à alésage étroit (voir le tableau des matériaux et la figure 1D).
- Sélectionner les ovocytes de stade SN GV et les transférer dans une goutte (25 μL) de milieu M2-IBMX sur un bloc chaud à 37 °C à l’abri de la lumière (Figure 1F).
- Recherchez les ovocytes SN en raison de leur plus grande taille et de leur noyau central, contrairement aux ovocytes NSN, dans lesquels les noyaux sont positionnés en périphérie21. Dans tous les cas, observez la configuration de l’ADN au microscope confocal avant de prendre la décision finale sur le type d’ovocyte GV (SN ou NSN).
- Induire des DSB à l’aide de l’étoposide (voir le tableau des matériaux). Placer les ovocytes de stade GV en gouttes (25 μL chacune) de l’agent génotoxique pendant 1 h sur le bloc chaud à 37 °C dans des conditions d’obscurité.
REMARQUE : L’étoposide est un inhibiteur de la topoisomérase II qui introduit des DSB dans l’ADN22. Conserver l’étoposide dans 10 μL aliquotes de 20 mg/mL à température ambiante à l’abri de la lumière. Les concentrations testées sont de 5 μg/mL, 20 μg/mL et 50 μg/mL. - Pour maintenir les ovocytes au stade GV arrêtés pendant une période prolongée, placer les ovocytes dans des gouttes de milieu de culture M16 (voir le tableau des matériaux) additionnées de 400 μM IBMX dans un incubateur (voir le tableau des matériaux) à 37 °C et 5 % de CO2. Conservez le M16 à 4 °C, filtrez le milieu à l’aide d’une seringue et d’un filtre de 0,2 μm et incubez-le pendant au moins 1 h avant utilisation.

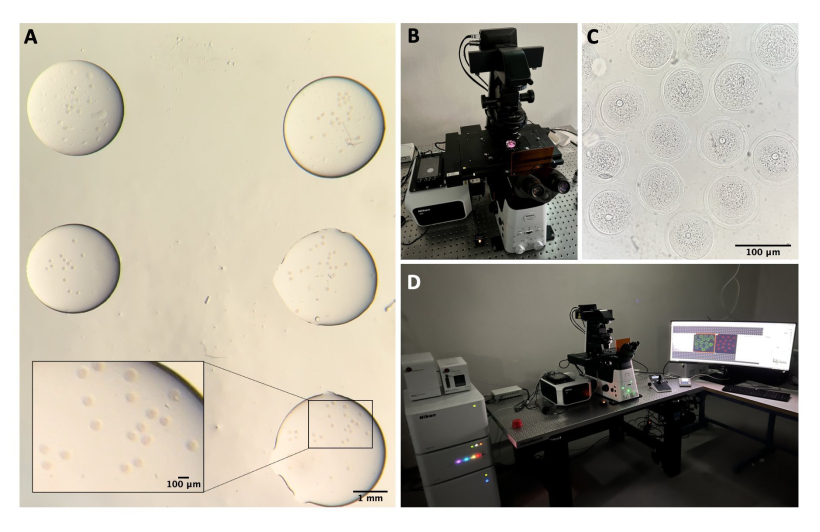
Figure 1 : Processus d’isolement des ovocytes. (A) Prélèvement du tissu adipeux péri-ovarien et des segments restants des trompes de Fallope des ovaires dans un milieu M2 avec IBMX. Photographie obtenue à l’aide des oculaires du stéréomicroscope. Barre d’échelle = 1 mm. (B) Ovaires isolés dans un milieu M2 avec IBMX. Image obtenue à l’aide des oculaires du stéréomicroscope. Barre d’échelle = 1 mm. (C) Perforation mécanique des ovaires à l’aide d’une aiguille de 27 G en milieu M2 avec IBMX. Image obtenue à l’aide des oculaires du stéréomicroscope. Barre d’échelle = 1 mm. (D) COC libérés par les ovaires après perforation dans un milieu M2 avec IBMX. Image obtenue à l’aide des oculaires du stéréomicroscope. Barre d’échelle = 100 μm. (E) Prélèvement d’ovocytes à l’aide d’une pipette buccale. (F) Ovocytes dénudés, après l’ablation des cellules cumulus environnantes, en milieu M2 avec IBMX. Image obtenue à l’aide des oculaires du stéréomicroscope. Barre d’échelle = 100 μm. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
3. Fixation des ovocytes et immunofluorescence
REMARQUE: La manipulation des ovocytes est effectuée à l’aide d’une pipette buccale sous un microscope stéréoscopique, et toutes les gouttes sont recouvertes d’huile minérale.
- Placer les ovocytes GV témoins et traités à l’étoposide dans différentes boîtes de culture tissulaire en plastique avec un tampon PFA-Tx-100 pendant 40 min à température ambiante.
- Lavez les ovocytes dans trois gouttes différentes de tampon de lavage (50 μL chacune) à température ambiante. Laissez les ovocytes pendant 5 min dans chaque goutte.
- Placer les ovocytes en gouttes de tampon bloquant (25 μL chacune) pendant 1 h sur un bloc chaud à 37 °C.
- Préparer l’anticorps primaire qui reconnaît γH2AX (phospho-Η2ΑΧ de lapin) (Ser139) (voir le tableau des matériaux) (solution mère : 1 mg/mL). Utiliser une dilution de 1:200 dans un tampon bloquant et placer les ovocytes dans des gouttes d’anticorps primaires (15 μL chacune) à 4 °C pendant la nuit.
NOTE : Le phospho-η2ΑΧ (γH2AX) est un marqueur courant pour la détection des DSB dans les cellules somatiques et les ovocytes GV18,23. - Le lendemain, lavez les ovocytes dans trois gouttes différentes de tampon de lavage (50 μL chacune) à température ambiante. Laissez les ovocytes pendant 5 min dans chaque goutte.
- Préparer l’anticorps secondaire, Alexa Fluor 488-conjugué chèvre anti-lapin (voir le tableau des matériaux) (solution mère : 2 mg/mL). Utiliser une dilution de 1:200 dans un tampon bloquant et placer les ovocytes dans des gouttes d’anticorps secondaires (15 μL chacune) pendant 1 h sur un bloc chaud à 37 °C à l’abri de la lumière.
- Transférer les ovocytes dans des gouttes de DRAQ7 (25 μL chacune) (solution mère : 0,3 mM ; voir le tableau des matériaux), qui est un colorant d’ADN fluorescent rouge lointain qui ne colore l’ADN que dans les cellules perméabilisées. Utiliser une dilution de 1:250 dans un tampon de lavage pendant 10 min à température ambiante dans des conditions sombres.
- Lavez les ovocytes dans trois gouttes différentes de tampon de lavage (50 μL chacune) à température ambiante. Laissez-les pendant 5 min dans chaque goutte, puis transférez-les sur de petites gouttes (environ 5 μL chacune) de tampon de lavage dans une boîte de Pétri à fond en verre de 35 mm (voir le tableau des matériaux) pour la microscopie confocale (Figure 2A).
REMARQUE: Le lavage de la coloration de l’ADN et de l’anticorps secondaire est effectué en même temps.
4. Microscopie confocale
REMARQUE: La microscopie confocale doit être effectuée immédiatement pour éviter la réduction de l’intensité de fluorescence après la mise en place des ovocytes dans des boîtes à fond en verre. L’accès à un microscope confocal (voir le tableau des matériaux) avec une platine motorisée est nécessaire.
- Configuration du microscope
- Dans le système confocal, allumez le contrôleur laser, les lasers, le contrôleur du microscope, les lampes pour la lumière transmise et le PC (Figure 2B, D).
- Ouvrez le logiciel confocal et choisissez la lentille à huile 40x.
- Placez la boîte dans le porte-échantillon et essayez de vous concentrer sur les ovocytes en déplaçant la platine sur les axes XY et Z à l’aide du joystick (Figure 2C).
- Scintigraphie des ovocytes
- Réglez la puissance du laser, le gain et la taille du sténopé indépendamment pour chaque expérience afin de minimiser toute saturation.
- Pour chaque ovocyte, définissez la zone d’intérêt, plus précisément dans le noyau au niveau de la zone de l’ADN. Définissez les limites de la zone de l’ADN et ajustez la taille du pas z à 3 μm. Ensuite, lancez l’analyse.
- Enregistrez les images de chaque cellule du dossier sélectionné.
- Une fois la numérisation terminée, quittez le logiciel, éteignez l’ordinateur et éteignez le contrôleur laser, les lasers, le contrôleur du microscope et les lampes pour la lumière transmise.

Figure 2 : Microscopie confocale. (A) Ovocytes fixés après avoir effectué le protocole d’immunofluorescence et la coloration de l’ADN, qui sont dans des gouttes séparées de tampon de lavage, recouvertes d’huile minérale, placées dans une boîte à fond en verre et préparées pour l’imagerie par microscopie confocale. Chaque goutte contient une catégorie expérimentale différente. Image obtenue à l’aide des oculaires du stéréomicroscope. Barre d’échelle = 1 mm/100 μm pour la partie agrandie. (B) Plaque à fond de verre placée sur la platine du microscope confocal. (C) Image en fond clair d’ovocytes obtenue par microscopie confocale. Barre d’échelle = 100 μm. (D) Le système de microscopie confocale. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
5. Analyse d’imagerie
- Téléchargez Fiji ImageJ-win64 dans le navigateur (https://imagej.net/software/fiji/downloads), ouvrez-le et importez les données sous forme de fichiers de pile TIFF.
REMARQUE: Ouvrez chaque fichier d’ovocytes séparément. - Cliquez sur l’image | Couleur | Diviser les canaux pour diviser tous les canaux.
- Cliquez sur LUT (Look up Table) et choisissez les couleurs préférées pour chaque canal.
- Cliquez sur l’image | Couleur | Fusionner les canaux pour fusionner les canaux de γΗ2ΑΧ et d’ADN. Laissez le canal de fond clair non fusionné.
- Dans les ovocytes NSN et dans les ovocytes SN présentant de faibles niveaux de dommages à l’ADN, γΗ2ΑΧ est détecté comme foyer dans la région de l’ADN. Dans ce cas, cliquez sur la commande « Multi-point » ou point , et sélectionnez chaque foyer γΗ2ΑΧ qui coïncide avec l’ADN. Répétez cette étape pour toutes les piles.
- Dans les ovocytes SN présentant des niveaux élevés de dommages à l’ADN, le signal γΗ2ΑΧ est distribué dans toute la région de l’ADN. Dans ce cas, cliquez sur Image | Piles | Z et, à l’aide de la commande Sélections à main levée , sélectionnez l’ensemble de la zone d’ADN.
- Pour mesurer la fluorescence γΗ2ΑΧ, cliquez sur Analyser | Mesurez et copiez les mesures dans un fichier .xlsx. Ensuite, calculez la fluorescence moyenne, normalisez les valeurs et comptez le nombre de foyers avant de créer des graphiques.
- Cliquez sur Analyser | Définir l’échelle pour définir l’échelle, puis sur Analyser | Outils | Barre d’échelle pour ajouter une barre d’échelle aux canaux.
Résultats
À l’aide de la procédure démontrée ici, les ovaires de souris ont été disséqués, la graisse a été retirée et des ovocytes adultes au stade GV ont été collectés. Ensuite, les cellules de cumulus ont été retirées par pipetage répétitif à l’aide d’une pipette étroite et ont été placées dans des gouttes fraîches de milieu M2-IBMX et recouvertes d’huile minérale sur un bloc chaud (37 °C) (Figure 1A-F). Trois concentrations différentes d’étoposide ont été préparées (5 μg/mL, 20 μg/mL et 50 μg/mL) en utilisant une concentration d’étoposide mère de 20 mg/mL. Les ovocytes de stade GV ont été placés à trois concentrations distinctes d’étoposide pendant 1 h dans des gouttes recouvertes d’huile minérale et protégées de la lumière à 37 °C. Le protocole d’immunofluorescence a ensuite été suivi, tel que décrit en détail dans la section du protocole, et les ovocytes ont été placés dans des boîtes à fond de verre et observés par microscopie confocale (Figure 2).
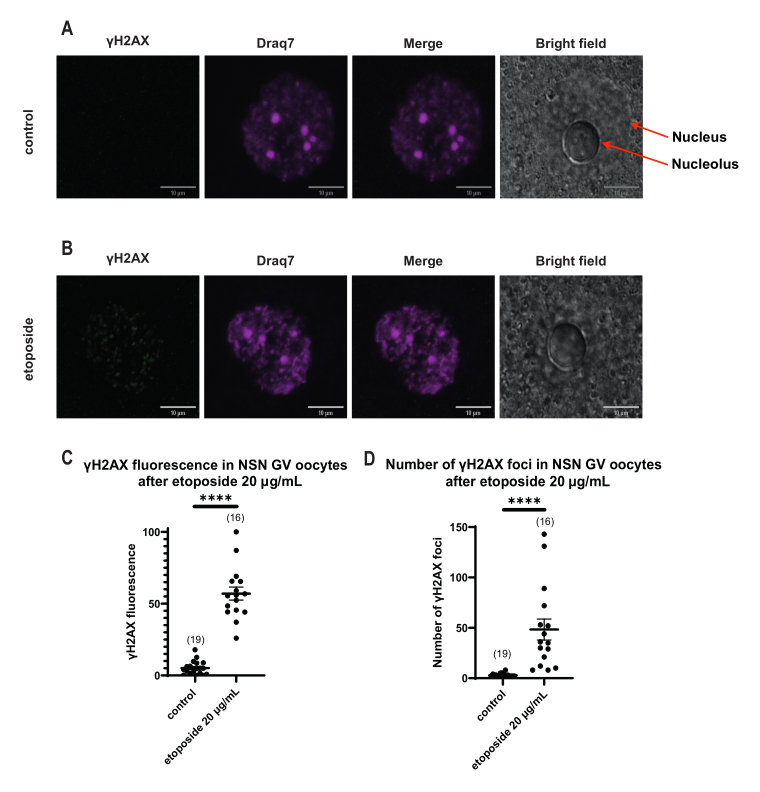
Dans les ovocytes au stade SN GV, immédiatement après une lésion de l’ADN, la présence de γH2AX a augmenté à toutes les concentrations d’étoposides (5 μg/mL, 20 μg/mL et 50 μg/mL), et le γH2AX a été distribué dans toute la région de l’ADN (Figure 3). La quantification et l’estimation de l’ORD ont été réalisées en observant l’intensité de fluorescence γH2AX sur les sites de l’ADN. La fluorescence γH2AX s’est intensifiée proportionnellement avec l’augmentation des concentrations d’étoposides. De plus, après un arrêt prolongé de la prophase (20 h après le traitement par l’étoposide), les ovocytes du stade GV ont montré la capacité de réduire le nombre et l’intensité des foyers γH2AX, ce qui implique la présence de processus de réparation actifs dans les ovocytes arrêtés au stade GV (Figure 3E).
Contrairement aux ovocytes SN, dans lesquels la fluorescence γH2AX était distribuée à travers l’ADN, dans les ovocytes NSN, γH2AX a été montré dans les foyers immédiatement après le traitement par l’étoposide à 20 μg/mL. Nous avons estimé le nombre de foyers qui coïncidaient avec la zone de l’ADN, calculé la fluorescence de chaque foyer et présenté la fluorescence moyenne de tous les ovocytes. La fluorescence et le nombre de foyers ont montré des différences statistiquement significatives entre les deux catégories d’ovocytes (Figure 4).
La microscopie confocale fournit des informations sur le nombre et l’intensité des foyers dans différentes piles Z, aidant ainsi à identifier la présence de dommages à l’ADN et la dynamique de réparation à des moments distincts. Le balayage Galvano permet un balayage de précision avec un faible bruit de fond et une meilleure analyse des images numérisées.

Figure 3 : Réduction de γH2AX dans les ovocytes de stade SN GV traités avec trois concentrations différentes d’étoposide après un arrêt prolongé de GV. (A) Fluorescence γH2AX dans les ovocytes de stade SN GV 0 h après le traitement par étoposide. Le γH2AX augmente immédiatement après l’exposition à toutes les concentrations d’étoposides, et l’augmentation dépend de la concentration (vert : γΗ2ΑΧ, magenta : ADN). Les images sont des projections Z-stack, et la luminosité/contraste ont été ajustés pour chaque canal à l’aide de Fiji / ImageJ. Barre d’échelle = 10 μm. (B) Graphique de la fluorescence γH2AX dans les ovocytes du stade SN GV 0 h après le traitement avec des concentrations distinctes d’étoposides. Les données représentent la moyenne ± SEM. Chaque point représente un ovocyte (le nombre d’ovocytes est indiqué dans le graphique), (ns = non significatif, ** p < 0,005, **** p < 0,0001, ANOVA à un facteur avec le test de comparaisons multiples de Tukey). (C) Fluorescence γH2AX dans les ovocytes de stade SN GV 20 h après le traitement par étoposide. γH2AX diminue 20 h après l’exposition à toutes les concentrations d’étoposides (vert : γΗ2ΑΧ, magenta : ADN). Les images sont des projections Z-stack, et la luminosité/contraste ont été ajustés pour chaque canal à l’aide de Fiji/ImageJ. Barre d’échelle = 10 μm. (D) Graphique de la fluorescence γH2AX dans les ovocytes de stade SN GV 20 h après le traitement avec des concentrations distinctes d’étoposides. Les données représentent la moyenne ± SEM. Chaque point représente un ovocyte (le nombre d’ovocytes est indiqué dans le graphique), (ns = non significatif, * p < 0,05, ** p < 0,005, *** p < 0,0005, **** p < 0,0001, ANOVA à un facteur avec le test de comparaisons multiples de Tukey). (E) Graphique à barres de la réduction de la fluorescence γH2AX dans les ovocytes de stade SN GV après arrêt de la prophase dans les ovocytes traités par étoposide. Le nombre au-dessus de chaque colonne indique le pourcentage de déclin de la fluorescence γH2AX. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 4 : Phosphorylation de Η2ΑΧ dans les ovocytes de stade NSN GV après traitement par l’étoposide à 20 μg/mL. (A) Images confocales représentatives d’un ovocyte témoin de stade NSN GV (vert : γΗ2ΑΧ, magenta : ADN). Les images sont des projections Z-stack, et la luminosité/contraste ont été ajustés pour chaque canal à l’aide de Fiji/ImageJ. Barre d’échelle = 10 μm. (B) Images confocales représentatives d’un ovocyte de stade NSN GV traité par étoposide (vert : γΗ2ΑΧ, magenta : ADN). Les ovocytes ont été fixés 0 h après le traitement par étoposide. Les images sont des projections Z-stack, et la luminosité/contraste ont été ajustés pour chaque canal à l’aide de Fiji/ImageJ. Barre d’échelle = 10 μm. (C) La fluorescence γΗ2ΑΧ normalisée dans les ovocytes de stade NSN GV après un traitement à l’étoposide à 20 μg/mL. Les données représentent la moyenne ± SEM. Chaque point représente un ovocyte (le nombre d’ovocytes est indiqué dans le graphique), tiré de deux expériences indépendantes (**** p < 0,0001, test t non paramétrique non apparié, test U de Mann-Whitney). (D) Nombre de foyers γΗ2ΑΧ dans les ovocytes du stade NSN GV après un traitement à l’étoposide à 20 μg/mL. Les données représentent la moyenne ± SEM. Chaque point représente un ovocyte (le nombre d’ovocytes est indiqué dans le graphique), tiré de deux expériences indépendantes (**** p < 0,0001, test t non paramétrique non apparié, test U de Mann-Whitney). Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Discussion
En utilisant la méthode décrite ici, nous avons détecté des DSB dans les ovocytes de mammifères. Cette méthode permet de détecter et d’étudier le processus de réparation de l’ADN dans les ovocytes. Le même protocole pourrait également être utilisé pour analyser d’autres protéines qui participent aux processus physiologiques dans les ovocytes de mammifères. Il est important d’étudier comment les ovocytes réagissent aux dommages potentiels de l’ADN afin de mieux comprendre la cause de l’hypofertilité féminine chez l’homme.
L’étude de la réponse aux dommages de l’ADN dans les ovocytes de mammifères peut être difficile en raison de la sensibilité des ovocytes. La manipulation des ovocytes nécessite des températures spécifiques et des concentrations de CO 2 et O2. Dans le même temps, les ovocytes doivent être protégés de la lumière. Toute manipulation doit être effectuée à l’aide de pipettes en verre qui ne sont pas trop étroites, car cela pourrait être nocif pour les ovocytes, mais aussi pas trop larges, car cela pourrait provoquer la dilution du milieu et, par conséquent, affecter négativement la procédure de fixation. À chaque étape de fixation, plusieurs gouttes de tampons sont utilisées pour minimiser l’effet de dilution. Une autre façon d’observer les DSB est le test Comet24. Même si cette technique est plus sensible, elle est plus compliquée. Dans le même temps, en utilisant le test Comet, il n’est pas possible de détecter la région exacte de l’ADN où les dommages se produisent, et dans les cellules avec des molécules d’ARN abondantes, comme les ovocytes au stade GV25, le bruit de fond pourrait être augmenté, conduisant à un faux signal de dommages à l’ADN26.
En utilisant le protocole d’immunofluorescence décrit ici, nous pouvons détecter les DSB avec précision et estimer les progrès de la réparation dans les ovocytes au stade GV, comme indiqué par la réduction de la fluorescence γH2AX au fil du temps. Néanmoins, l’une des limites de cette méthode est que certains anticorps peuvent présenter une distribution non spécifique dans tout l’ooplasme, conduisant ainsi à des images avec une fluorescence de fond élevée. Le tampon PFA-Tx-100 est utilisé à la place du PFA séquentiel et du Tx-100, car nous avons observé qu’il améliore le processus de fixation en permettant la détection de moins de bruit de fond et de fluorescence non spécifique. Une deuxième limite de l’utilisation de γH2AX pour la détection DSB est que les dommages ne peuvent pas être estimés après GVBD en raison de la phosphorylation spontanée de γH2AX dans la méiose23.
Dans ce protocole d’immunofluorescence, les ovocytes restent dans un tampon liquide et ne peuvent pas être stockés dans des lames. Ce fait rend difficile la conservation des cellules fixées pendant des jours après l’ajout de l’anticorps secondaire. Afin d’obtenir des images de bonne qualité et de ne pas perdre de signal, il est préférable d’effectuer l’imagerie dans les heures qui suivent l’ajout de l’anticorps secondaire. Il convient également de noter que le balayage des noyaux à travers l’axe Z pourrait affaiblir le signal en raison d’une surexposition. Pour cette raison, il est préférable de réduire la puissance du laser et d’augmenter la vitesse de numérisation.
Enfin, une autre limite du protocole d’immunofluorescence est qu’il ne peut être utilisé que pour les cellules fixes/non vivantes. Par conséquent, nous ne pouvons estimer que la présence et l’absence de facteurs à des moments précis sans savoir s’il y a des fluctuations dans leur concentration ou des changements dans leur comportement au fil du temps. Ce problème pourrait être résolu en utilisant l’imagerie de cellules vivantes et des marqueurs marqués par fluorescence.
Déclarations de divulgation
Les auteurs n’ont aucun conflit d’intérêts.
Remerciements
Nous reconnaissons le soutien apporté à ce travail par le projet « Mise en place d’infrastructures de renforcement des capacités en recherche biomédicale (BIOMED-20) » (MIS 5047236), mis en œuvre dans le cadre de l’action « Renforcement de l’infrastructure de recherche et d’innovation », financé par le programme opérationnel « Compétitivité, entrepreneuriat et innovation » (CRSN 2014-2020) et cofinancé par la Grèce et l’Union européenne (Fonds européen de développement régional).
matériels
| Name | Company | Catalog Number | Comments |
| 3 mL Pasteur pipettes in LDPE, graduated | APTACA | 1502 | |
| 10 cc syringes | SoftCare | 114.104.21 | |
| Alexa Fluor 488-conjugated goat anti-rabbit Secondary Ab | Biotium | 20012 | |
| Anti-phospho-H2A.X (Ser139) | Merck Millipore | 07-164 | |
| ARE Heating Magnetic Stirrer | VELP Scientifica | F20500162 | |
| BD FALCON 5 mL Polystyrene Round-Bottom Tubes | BD Biosciences | 352054 | |
| BD Microlance 3 Needles 27 G - 0.40 x 13 mm | Becton Dickinson | 300635 | |
| Bovine Serum Albumin Fraction V | Roche | 10735078001 | |
| DMSO Anhydrous | Biotium | 90082 | |
| DRAQ7 DNA dye | BioStatus | DR71000 | |
| EGTA | Sigma-Aldrich | E4378-25G | |
| EMSURE MgCl2. 6H2O | Merck Millipore | 1058330250 | |
| Etoposide | CHEMIPHARM | L01CB01 | |
| FALCON 14 mL Polystyrene Round-Bottom Tubes | Corning Science | 532057 | |
| FALCON Tissue Culture Dishes, Easy-Grip, 35 x 10 mm Style | Corning Science | 353001 | |
| Glass Bottom Culture Dishes (35 mm Petri dish/ 14 mm Microwell, No. 0 coverglass) | MatTek Corporation | P35G-0-14-C | |
| HEPES | Sigma-Aldrich | H6147-25G | |
| HERACELL 150i CO2 Incubator | ThermoFisher Scientific | 50116048 | |
| IBMX powder | Sigma-Aldrich | I5879-100MG | |
| Leica M125 Stereo Microscope | Leica Microsystems | ||
| M16 Medium | Sigma-Aldrich | M7292 | |
| M2 Medium | Sigma-Aldrich | M7167 | |
| Mineral Oil | Sigma-Aldrich | M5310 | |
| NaN3 | Honeywell | 13412H | |
| NaOH | Merck Millipore | 1064981000 | |
| Nikon AX ECLIPSE Ti2 Confocal Microscope | Nikon Corporation | ||
| Nikon SMZ800N Stereo Microscope | Nikon Corporation | ||
| Paraformaldehyde | Sigma-Aldrich | 158127 | |
| Pasteur pippettes, glass, long form 230 mm | DURAN WHEATON KIMBLE | 357335 | |
| pH/ORP meter | Hanna Instruments Ltd | HI2211 | |
| Phosphate buffered saline tablets | Sigma-Aldrich | P4417-100TAB | |
| PIPES | Sigma-Aldrich | P1851 | |
| PMSG Protein Lyophilised | Genway Biotech (now AVIVA Systems Biology) | GWB-2AE30A (now OPPA01037) | |
| QBD4 Dry block heater | Grant Instruments (Cambridge) Ltd | A25218 | |
| Triton X-100 | Sigma-Aldrich | T8787 | |
| Whatman Puradisc 25 mm 0.2 μm filters | GE Healthcare | 6780-2502 |
Références
- Wang, X., Pepling, M. E. Regulation of meiotic prophase one in mammalian oocytes. Frontiers in Cell and Developmental Biology. 9, 667306(2021).
- Filatov, M., Khramova, Y., Semenova, M. Molecular mechanisms of prophase I meiotic arrest maintenance and meiotic resumption in mammalian oocytes. Reproductive Sciences. 26 (11), 1519-1537 (2019).
- Adhikari, D., et al. Inhibitory phosphorylation of Cdk1 mediates prolonged prophase I arrest in female germ cells and is essential for female reproductive lifespan. Cell Research. 26 (11), 1212-1225 (2016).
- Sun, S. C., Kim, N. H. Molecular mechanisms of asymmetric division in oocytes. Microscopy and Microanalysis. 19 (4), 883-897 (2013).
- Jones, K. T. Mammalian egg activation: From Ca2+ spiking to cell cycle progression. Reproduction. 130 (6), 813-823 (2005).
- Solc, P., Schultz, R. M., Motlik, J. Prophase I arrest and progression to metaphase I in mouse oocytes: comparison of resumption of meiosis and recovery from G2-arrest in somatic cells. Molecular Human Reproduction. 16 (9), 654-664 (2010).
- Parfenov, V., Potchukalina, G., Dudina, L., Kostyuchek, D., Gruzova, M. Human antral follicles: oocyte nucleus and the karyosphere formation (electron microscopic and autoradiographic data). Gamete Research. 22 (2), 219-231 (1989).
- Zuccotti, M., Piccinelli, A., Giorgi Rossi, P., Garagna, S., Redi, C. A. Chromatin organization during mouse oocyte growth. Molecular Reproduction and Development. 41 (4), 479-485 (1995).
- Sun, X., et al. Comprehensive analysis of nonsurrounded nucleolus and surrounded nucleolus oocytes on chromatin accessibility using ATAC-seq. Molecular Reproduction and Development. 90 (2), 87-97 (2023).
- Zuccotti, M., Bellone, M., Longo, F., Redi, C. A., Garagna, S. Fully-mature antral mouse oocytes are transcriptionally silent but their heterochromatin maintains a transcriptional permissive histone acetylation profile. Journal of Assisted Reproduction and Genetics. 28 (12), 1193-1196 (2011).
- Winship, A. L., Stringer, J. M., Liew, S. H., Hutt, K. J. The importance of DNA repair for maintaining oocyte quality in response to anti-cancer treatments, environmental toxins and maternal ageing. Human Reproduction Update. 24 (2), 119-134 (2018).
- Lieber, M. R. The mechanism of human nonhomologous DNA end joining. The Journal of Biological Chemistry. 283 (1), 1-5 (2008).
- Chiruvella, K. K., Liang, Z., Wilson, T. E. Repair of double-strand breaks by end joining. Cold Spring Harbor Perspectives in Biology. 5 (5), 012757(2013).
- Chang, H. H. Y., Pannunzio, N. R., Adachi, N., Lieber, M. R. Non-homologous DNA end joining and alternative pathways to double-strand break repair. Nature Reviews. Molecular Cell Biology. 18 (8), 495-506 (2017).
- Shibata, A., Jeggo, P. A. Roles for the DNA-PK complex and 53BP1 in protecting ends from resection during DNA double-strand break repair. Journal of Radiation Research. 61 (5), 718-726 (2020).
- Mohiuddin, I. S., Kang, M. H. DNA-PK as an emerging therapeutic target in cancer. Frontiers in Oncology. 9, 635(2019).
- Marangos, P., Carroll, J. Oocytes progress beyond prophase in the presence of DNA damage. Current Biology. 22 (11), 989-994 (2012).
- Paull, T. T., et al. A critical role for histone H2AX in recruitment of repair factors to nuclear foci after DNA damage. Current Biology. 10 (15), 886-895 (2000).
- Bakkenist, C. J., Kastan, M. B. DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation. Nature. 421 (6922), 499-506 (2003).
- Marangos, P., et al. DNA damage-induced metaphase I arrest is mediated by the spindle assembly checkpoint and maternal age. Nature Communications. 6, 8706(2015).
- Lavrentyeva, E. A., Shishova, K. V., Zatsepina, O. V. Differences in nuclear dynamics in mouse GV oocytes with a diverse chromatin configuration. Biology Bulletin Russian Academy of Sciences. 46, 332-341 (2019).
- Montecucco, A., Zanetta, F., Biamonti, G. Molecular mechanisms of etoposide. EXCLI Journal. 14, 95-108 (1998).
- Mayer, A., et al. DNA damage response during mouse oocyte maturation. Cell Cycle. 15 (4), 546-558 (2016).
- Olive, P., Banáth, J. The comet assay: A method to measure DNA damage in individual cells. Nature Protocols. 1, 23-29 (2006).
- Wu, D., Dean, J. EXOSC10 sculpts the transcriptome during the growth-to-maturation transition in mouse oocytes. Nucleic Acids Research. 48 (10), 5349-5365 (2020).
- Simon, L., Emery, B., Carrell, D. DNA damage: COMET assay. Manual of Sperm Function Testing in Human Assisted Reproduction. Agarwal, A., Henkel, R., Majzoub, A. , 202-212 (2021).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.