
Method Article
Nachweis von DNA-Doppelstrangbrüchen in Maus-Eizellen
In diesem Artikel
Zusammenfassung
Die Aufrechterhaltung der Integrität des Eizellgenoms ist notwendig, um die genetische Genauigkeit des resultierenden Embryos zu gewährleisten. Hier stellen wir ein genaues Protokoll zum Nachweis von DNA-Doppelstrangbrüchen in weiblichen Keimzellen von Säugetieren vor.
Zusammenfassung
Eizellen gehören zu den größten und langlebigsten Zellen im weiblichen Körper. Sie werden während der Embryonalentwicklung in den Eierstöcken gebildet und bleiben in der Prophase der Meiose I verhaftet. Der Ruhezustand kann Jahre andauern, bis die Eizellen einen Stimulus zum Wachstum erhalten und die Fähigkeit erlangen, die Meiose wieder aufzunehmen. Dieser langwierige Stillstand macht sie extrem anfällig für die Anhäufung von DNA-schädigenden Beleidigungen, die die genetische Integrität der weiblichen Keimzellen und damit die genetische Integrität des zukünftigen Embryos beeinträchtigen.
Folglich ist die Entwicklung einer genauen Methode zum Nachweis von DNA-Schäden, die der erste Schritt zur Etablierung von DNA-Schadensreaktionsmechanismen ist, von entscheidender Bedeutung. Diese Arbeit beschreibt ein gängiges Protokoll, um das Vorhandensein und Fortschreiten von DNA-Schäden in Prophase-Arrest-Eizellen während eines Zeitraums von 20 Stunden zu testen. Konkret sezieren wir die Eierstöcke der Maus, entnehmen die Cumulus-Eizell-Komplexe (COCs), entfernen die Cumuluszellen aus den COCs und kultivieren die Eizellen in einem μ2-Medium, das 3-Isobutyl-1-methylxanthin enthält, um den Zustand des Stillstands aufrechtzuerhalten. Danach werden die Eizellen mit dem zytotoxischen, antineoplasmatischen Medikament Etoposid behandelt, um Doppelstrangbrüche (DSBs) zu erzeugen.
Mit Hilfe von Immunfluoreszenz und konfokaler Mikroskopie können wir die Konzentrationen des Kernproteins γH2AX, der phosphorylierten Form des Histons H2AX, nachweisen und quantifizieren. H2AX wird nach DNA-Schäden an den Stellen von DSBs phosphoryliert. Die Unfähigkeit, die DNA-Integrität nach DNA-Schäden in Eizellen wiederherzustellen, kann zu Unfruchtbarkeit, Geburtsfehlern und erhöhten Raten von Spontanaborten führen. Daher ist das Verständnis der Reaktionsmechanismen von DNA-Schäden und gleichzeitig die Etablierung einer intakten Methode zur Untersuchung dieser Mechanismen für die reproduktionsbiologische Forschung unerlässlich.
Einleitung
Der Prozess der Meiose in weiblichen Keimzellen von Säugetieren wird in den Eierstöcken vor der Geburt eingeleitet. Die Gesamtzahl der Eizellen wird in den Eierstöcken vor allem während der Embryogenese gebildet. Die Eizellen treten in die Meiose ein und bleiben in der Prophase I1 verriegelt. Nach dem Einsetzen der Pubertät und der Produktion und endokrinen Wirkung des follikelstimulierenden Hormons (FSH) und des luteinisierenden Hormons (LH) können die Eizellen die Meiose wieder einleiten und abschließen2. Beim Menschen kann der Prophasenstillstand bis zu 50 Jahre andauern3. Die Zellteilungen nach dem Eintritt in die Meiose I sind asymmetrisch, was zur Produktion eines kleinen Polkörpers und einer Eizelle führt, die ihre Größe beibehält. Daher werden die meisten zytoplasmatischen Bestandteile während der frühen Embryogenese im Ooplasma gespeichert4. Dann treten die Eizellen in die Meiose II ein, ohne ihren Zellkern zu reformieren oder ihre Chromosomen zu dekondensieren, und bleiben bis zur Befruchtung in der Metaphase II gefangen5.
Ein einzigartiges Merkmal, das Eizellen von somatischen Zellen unterscheidet, ist der Zustand des Stillstands in der Prophase I, wenn die Eizelle einen intakten Zellkern besitzt (Keimvesikel [GV]-Arrest), der als GV-Stadium6 bezeichnet wird. Basierend auf der Chromatinorganisation werden die Eizellen im GV-Stadium in zwei Kategorien eingeteilt: nicht-umgebener Nukleolus (NSN) und umschlossener Nukleolus (SN)7,8. In NSN-GV-Oozyten breitet sich das Chromatin in der gesamten Kernregion aus, und die Transkription ist aktiv, während das Chromatin in SN-Oozyten einen kompakten Ring bildet, der den Nukleolus umgibt, und die Transkription schweigt9. Beide Arten von Eizellen im GV-Stadium zeigen meiotische Kompetenz; Sie treten mit der gleichen Geschwindigkeit in die Meiose ein, aber die NSN-Eizellen weisen eine geringe Entwicklungskapazität auf und können sich nicht über das Zweizellstadium hinaus entwickeln10.
Der langwierige Zustand des Prophase-I-Stillstands erhöht die Inzidenz der Akkumulation von DNA-Schäden11. Daher sind die Reaktionsmechanismen auf DNA-Schäden in Eizellen unerlässlich, um die Produktion von Keimzellen mit genetischer Integrität zu ermöglichen und sicherzustellen, dass der resultierende Embryo einen physiologischen Chromosomengehalt hat.
Ein zentraler Aspekt der DNA-Schadensantwort ist die DNA-Reparatur. Zu den wichtigsten Wegen für die DSB-Reparatur in eukaryotischen Zellen gehören die nicht-homologe Endverbindung (NHEJ), die homologe Rekombination (HR) und die alternative NHEJ12,13,14,15. NHEJ ist ein schnellerer, aber fehleranfälligerer Mechanismus, während HR mehr Zeit benötigt, um abgeschlossen zu werden, aber eine hohe Genauigkeit aufweist16.
Es gibt nicht genügend Wissen über die Mechanismen, die Eizellen für die Reparatur von DNA-Schäden nutzen. Studien haben gezeigt, dass DNA-Schäden, die in ausgewachsenen Säugetier-Eizellen durch den Einsatz von genotoxischen Substanzen wie Etoposid, Doxorubicin, UVB oder ionisierender Strahlung induziert werden, den Zeitpunkt und die Geschwindigkeit des Austritts aus dem Prophase-I-Arrest nicht beeinflussen17. Eizellen können auch bei erhöhter Schädigung einen GV-Abbau (GVBD) durchlaufen. Diese Schädigung kann durch die Beobachtung von γH2AX bestimmt werden. Diese phosphorylierte Form von H2AX (γΗ2ΑΧ) ist ein DSB-Marker, der sich an der Stelle von Brüchen befindet und als Gerüst fungiert, um die Reparatur von Faktoren und Proteinen zu unterstützen, die sich an gebrochenen Enden ansammeln18.
Das Fehlen eines Zellzyklusstillstands nach DNA-Schäden ist auf einen unzureichenden DNA-Schadens-Checkpoint zurückzuführen, der es Eizellen mit unreparierter DNA ermöglicht, wieder in die Meiose einzutreten. Nach einer hohen DNA-Schädigung kann ein Checkpoint den Prophase-Arrest durch die Aktivierung eines ATM/Chk1-abhängigen Signalwegs aufrechterhalten. Die eingeschränkte Checkpoint-Reaktion auf DSBs ist auf die begrenzte Aktivierung von ATM17,19 zurückzuführen. In der M-Phase der Meiose I hat die Forschung gezeigt, dass DNA-Schäden einen Spindle Assembly Checkpoint (SAC)-induzierten Meiose I-Checkpoint aktivieren können, der die Aktivierung des E3-Ubiquitin-Ligase-Anaphase-fördernden Komplexes/Zyklosoms (APC/C) und damit den M-Phasen-Austritt verhindert. Darüber hinaus überwindet die Ablation von SAC-Proteinen den Zustand des M-Phasen-Arrests, was die Bedeutung des SAC für die Etablierung des Meiose-I-Chekpoints20 unterstreicht.
Wie frühere Forschungen deutlich zeigen, können DSBs keinen robusten Prophase-Checkpoint in Maus-Eizellen induzieren. Wenn solche Schäden nicht repariert werden, kann dies dazu führen, dass Embryonen Chromosomenanomalien tragen. Daher ist es wichtig, die Reaktion auf DNA-Schäden in verschiedenen Stadien der weiblichen Gametogenese zu untersuchen, um die einzigartigen Wege besser zu verstehen, die Eizellen nutzen, um mit potenziellen genetischen Beleidigungen fertig zu werden.
Protokoll
Alle Mäuseversuche wurden von den lokalen Behörden (Region Ioannina, Griechenland) genehmigt und in Übereinstimmung mit den Richtlinien 2010/63/EU des Rates der Europäischen Gemeinschaften durchgeführt. Es wurden Experimente unter Berücksichtigung der Prinzipien der 3R durchgeführt. Alle CD-1-Mäuse, die für die Experimente verwendet wurden, wurden in der Tierhauseinrichtung der Universität von Ioannina, Griechenland, in einem Raum mit kontrollierter Temperatur (22 °C) und Luftfeuchtigkeit (60 %) gehalten und ad libitum gefüttert. Das Tierhaus verfügt über eine Lizenz zum Betrieb einer Anlage für Zucht (EL33-BIObr01), Lieferung (EL33-BIOsup01) und Versuche (EL33BIO-exp01).
1. Herstellung der Reagenzien
- 3-Isobutyl-1-methylxanthin (IBMX)-Pulver (siehe Materialtabelle) in Dimethylsulfoxid (DMSO) (siehe Materialtabelle) auf eine Endkonzentration von 200 mM verdünnen. 10 μl aliquotieren und bei −20 °C lagern. Verwenden Sie die Lösung innerhalb von 1 Monat.
HINWEIS: IBMX-Pulver wird bei −20 °C gelagert. - Bereiten Sie alle Immunfluoreszenzpuffer vor und lagern Sie sie bei 4 °C.
- Sterile phosphatgepufferte Kochsalzlösung (PBS) wird hergestellt, indem eine PBS-Tablette (siehe Materialtabelle) in 200 ml ddH2Ο verdünnt wird.
- Stellen Sie PHEM-Puffer her, indem Sie 80 ml ddH2 Ο, 0,59575 g HEPES, 1,81422 g PIPES, 0,38035 g EGTA und 0,04066 g MgCl2 (siehe Materialtabelle) unter Rühren mit einem Magnetrührer (siehe Materialtabelle) hinzufügen und gleichzeitig NaOH (siehe Materialtabelle) hinzufügen, bis der pH-Wert 6,9erreicht (überprüfen Sie dies mit einem pH/Redox-Messgerät [siehe Materialtabelle]). Dann wird ddH2Ο zu einem Endvolumen von 100 ml zugegeben.
- Bereiten Sie den Paraformaldehyd-Triton-X-100 (PFA-Tx-100)-Puffer vor, indem Sie PFA-Pulver (siehe Materialtabelle) in PHEM-Puffer verdünnen, während Sie mit einem Magnetrührer unter Erhitzung bei einer Endkonzentration von 4 % PFA gerührt werden. Filtern Sie dann den Puffer mit einer Spritze und einem 0,2-μm-Filter (siehe Materialtabelle) und fügen Sie 0,5 % Tx-100 hinzu (siehe Materialtabelle). Bereiten Sie ca. 10 ml PFA-Tx-100 (0,4 g PFA, 50 μl Tx-100) vor, was für ein Experiment ausreicht. Maximal 1 Woche bei 4 °C lagern.
VORSICHT: Tragen Sie Handschuhe, um PFA zu handhaben, und vermeiden Sie den Kontakt mit Haut und Augen. - Waschpuffer durch Zugabe von Rinderserumalbumin (Endkonzentration: 0,5 Gew.-% BSA) (siehe Materialtabelle) in PBS vorbereiten und mechanisch rühren. Fügen Sie 10 % w/v NaN3-Puffer (Natriumazid) in einer Verdünnung von 1:1.000 hinzu, um das Risiko einer Pilz- und Bakterienkontamination zu minimieren. Stellen Sie einen 10%igen w/v NaN3-Puffer her, indem Sie 1 gNaN3-Pulver (siehe Materialtabelle) zu 10 ml ddH2Ohinzufügen. Lagern Sie denNaN3-Puffer bei Raumtemperatur.
- Den Sperrpuffer durch Zugabe von BSA (Endkonzentration: 3 Gew.-%) in PBS und mechanisches Rühren herstellen. Fügen Sie 10 %NaN3-Puffer in einer Verdünnung von 1:1.000 hinzu.
2. Entnahme von GV-Eizellen aus präparierten Eierstöcken und Induktion von DSBs
HINWEIS: Alle Werkzeuge und Lösungen sollten steril sein. Die Handhabung der Eizellen erfolgt mit einer Mundpipette unter einem Stereomikroskop (siehe Materialtabelle), und alle Tropfen sind mit Mineralöl bedeckt (siehe Materialtabelle und Abbildung 1E).
- Intraperitoneal injizieren Sie Mäusen 46-48 Stunden vor der Keulung der Mäuse durch Zervixluxation 7 internationale Einheiten (IE) Serumgonadotropin (PMSG) der trächtigen Stute (siehe Materialtabelle).
HINWEIS: Alle verwendeten Mäuse sollten 8-12 Wochen alt sein. - Das M2-Nährmedium (siehe Materialtabelle) mit einer Spritze und einem 0,2-μm-Filter filtrieren und IBMX 200 mM zu einer Endkonzentration von 200 μM in ein 14-ml-Rundbodenröhrchen (siehe Materialtabelle) geben, um die Eizellen in der Prophase I zu halten. Bereiten Sie dann Tropfen des M2-IBMX-Mediums in einer Kunststoff-Gewebekulturschale vor (siehe Materialtabelle) und legen Sie sie bei 37 °C für mindestens 30 Minuten auf einen heißen Block (siehe Materialtabelle), bevor Sie die Eizellen isolieren. Lagern Sie den M2 bei 4 °C.
- Die Mäuse werden durch Zervixluxation geopfert, die Eierstöcke präpariert und mit M2-IBMX in ein 5-ml-Röhrchen mit rundem Boden (siehe Materialtabelle) gegeben.
- Die Eierstöcke werden auf einen Kunststoffdeckel mit 1,5 ml M2-IBMX übertragen, das perivarielle Fettgewebe oder die Eileitersegmente entfernt und die Koks durch mechanische Perforation der Eierstöcke mit einer 27-G-Nadel freigesetzt (siehe Materialtabelle und Abbildung 1A-C).
- Die Koks werden in eine Kulturschale mit Tropfen M2-IBMX (jeweils ca. 25-30 μl) überführt und die Kumuluszellen durch wiederholtes Pipettieren mit einer Pasteurpipette aus Glas mit schmaler Bohrung entnommen (siehe Materialtabelle und Abbildung 1D).
- Wählen Sie Oozyten im SN-GV-Stadium aus und übertragen Sie sie in einem Tropfen (25 μl) M2-IBMX-Medium auf einen heißen Block bei 37 °C, der vor Licht geschützt ist (Abbildung 1F).
- Suchen Sie nach SN-Oozyten aufgrund ihrer größeren Größe und ihrer zentral platzierten Zellkerne im Gegensatz zu NSN-Oozyten, bei denen die Kerne peripher positioniert sind21. Beobachten Sie in jedem Fall die DNA-Konfiguration unter einem konfokalen Mikroskop, bevor Sie die endgültige Entscheidung über die Art der GV-Eizelle (SN oder NSN) treffen.
- Induzieren Sie DSBs mit Etoposid (siehe Materialtabelle). Die Eizellen im GV-Stadium werden in Tropfen (je 25 μl) des genotoxischen Mittels 1 h lang bei 37 °C im Dunkeln auf den heißen Block gelegt.
HINWEIS: Etoposid ist ein Topoisomerase-II-Inhibitor, der DSBs in die DNAeinführt 22. Bewahren Sie das Etoposid in 10-μl-Aliquoten von 20 mg/ml bei Raumtemperatur vor Licht geschützt auf. Die getesteten Konzentrationen betragen 5 μg/ml, 20 μg/ml und 50 μg/ml. - Um die Eizellen im GV-Stadium über einen längeren Zeitraum zu halten, werden die Eizellen in Tropfen des M16-Kulturmediums (siehe Materialtabelle), ergänzt mit 400 μM IBMX, in einen Inkubator (siehe Materialtabelle) bei 37 °C und 5 % CO2 gelegt. Lagern Sie das M16 bei 4 °C, filtrieren Sie das Medium mit einer Spritze und einem 0,2 μm-Filter und inkubieren Sie es vor Gebrauch mindestens 1 h.

Abbildung 1: Prozess der Eizellenisolierung . (A) Entfernung von perivariellem Fettgewebe und übrig gebliebenen Eileitersegmenten aus Ovarien in M2-Medium mit IBMX. Das Foto wurde mit den Stereomikroskop-Okularen aufgenommen. Maßstabsbalken = 1 mm. (B) Isolierte Ovarien in M2-Medium mit IBMX. Das Bild wurde durch die Stereomikroskop-Okulare aufgenommen. Maßstabsbalken = 1 mm. (C) Mechanische Perforation der Eierstöcke mit einer 27-G-Nadel in M2-Medium mit IBMX. Das Bild wurde durch die Stereomikroskop-Okulare aufgenommen. Maßstabsbalken = 1 mm. (D) Koks, die nach Perforation in M2-Medium mit IBMX aus den Eierstöcken freigesetzt werden. Das Bild wurde durch die Stereomikroskop-Okulare aufgenommen. Skalenbalken = 100 μm. (E) Eizellentnahme mit einer Mundpipette. (F) Entblößte Eizellen nach Entfernung der umgebenden Kumuluszellen in M2-Medium mit IBMX. Das Bild wurde durch die Stereomikroskop-Okulare aufgenommen. Maßstabsleiste = 100 μm. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}
3. Eizellfixierung und Immunfluoreszenz
HINWEIS: Die Handhabung der Eizellen erfolgt mit einer Mundpipette unter einem Stereomikroskop, und alle Tropfen sind mit Mineralöl bedeckt.
- Geben Sie die Kontroll- und Etoposid-behandelten GV-Eizellen für 40 Minuten bei Raumtemperatur in verschiedene Kunststoff-Gewebekulturschalen mit PFA-Tx-100-Puffer.
- Waschen Sie die Eizellen in drei verschiedenen Tropfen Waschpuffer (je 50 μl) bei Raumtemperatur. Lassen Sie die Eizellen in jedem Tropfen 5 Minuten einwirken.
- Legen Sie die Eizellen in Tropfen des Blockierungspuffers (je 25 μl) für 1 h auf einen heißen Block bei 37 °C.
- Bereiten Sie den primären Antikörper vor, der γH2AX (Kaninchenphospho-Η2ΑΧ) erkennt (Ser139) (siehe Materialtabelle) (Stammlösung: 1 mg/ml). Verwenden Sie eine Verdünnung von 1:200 in Blocking-Puffer und geben Sie die Eizellen in Tropfen des primären Antikörpers (je 15 μl) bei 4 °C über Nacht.
HINWEIS: Phospho-Η2ΑΧ (γH2AX) ist ein gängiger Marker für den Nachweis von DSBs sowohl in somatischen Zellen als auch in GV-Oozyten18,23. - Am nächsten Tag waschen Sie die Eizellen in drei verschiedenen Tropfen Waschpuffer (je 50 μl) bei Raumtemperatur. Lassen Sie die Eizellen in jedem Tropfen 5 Minuten einwirken.
- Bereiten Sie den sekundären Antikörper, Alexa Fluor 488-konjugiertes Ziegen-Anti-Kaninchen (siehe Materialtabelle) vor (Stammlösung: 2 mg/ml). Verwenden Sie eine Verdünnung von 1:200 in Blocking-Puffer und legen Sie die Eizellen in Tropfen des sekundären Antikörpers (je 15 μl) für 1 h auf einen heißen Block bei 37 °C, der vor Licht geschützt ist.
- Übertragen Sie die Eizellen in Tropfen von DRAQ7 (je 25 μl) (Stammlösung: 0,3 mM; siehe Materialtabelle), einem dunkelrot fluoreszierenden DNA-Farbstoff, der DNA nur in permeabilisierten Zellen färbt. Verwenden Sie eine Verdünnung von 1:250 im Waschpuffer für 10 Minuten bei Raumtemperatur unter dunklen Bedingungen.
- Waschen Sie die Eizellen in drei verschiedenen Tropfen Waschpuffer (je 50 μl) bei Raumtemperatur. Lassen Sie sie 5 Minuten in jedem Tropfen stehen und geben Sie sie dann in kleine Tropfen (jeweils ca. 5 μl) Waschpuffer in einer 35-mm-Petrischale mit Glasboden (siehe Materialtabelle) für die konfokale Mikroskopie (Abbildung 2A).
HINWEIS: Das Waschen der DNA-Färbung und des sekundären Antikörpers wird gleichzeitig durchgeführt.
4. Konfokale Mikroskopie
HINWEIS: Die konfokale Mikroskopie sollte sofort durchgeführt werden, um eine Verringerung der Fluoreszenzintensität nach der Platzierung der Eizellen in Glasbodenschalen zu vermeiden. Der Zugang zu einem konfokalen Mikroskop (siehe Materialtabelle) mit motorisiertem Tisch ist erforderlich.
- Mikroskop einrichten
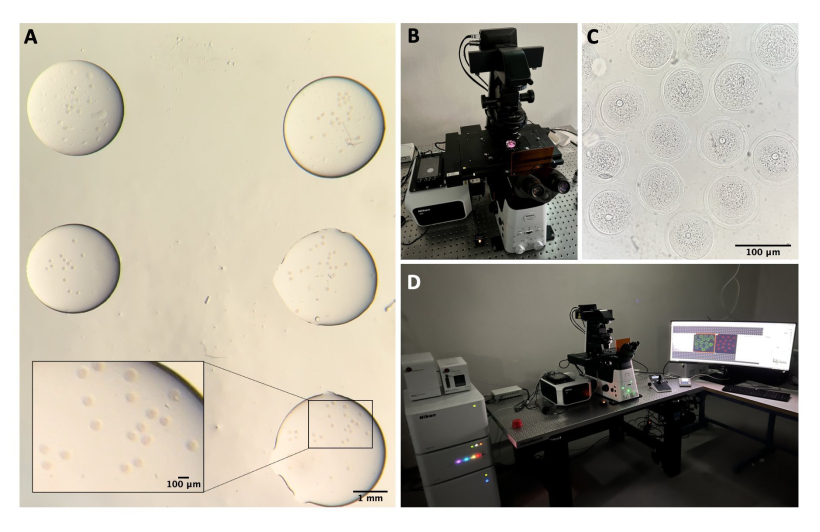
- Schalten Sie im konfokalen System den Lasercontroller, die Laser, den Mikroskopcontroller, die Lampen für das Durchlicht und den PC ein (Abbildung 2B, D).
- Öffnen Sie die konfokale Software und wählen Sie die 40-fache Öllinse.
- Legen Sie die Schale in den Probenhalter und versuchen Sie, sich auf die Eizellen zu konzentrieren, indem Sie den Tisch mit dem Joystick auf der XY- und Z-Achse bewegen (Abbildung 2C).
- Scannen der Eizellen
- Stellen Sie die Laserleistung, die Verstärkung und die Lochblendengröße für jedes Experiment unabhängig voneinander ein, um die Sättigung zu minimieren.
- Legen Sie für jede Eizelle den interessierenden Bereich fest, insbesondere im Zellkern im DNA-Bereich. Definieren Sie die Grenzen des DNA-Bereichs und stellen Sie die z-Schrittweite auf 3 μm ein. Starten Sie dann den Scanvorgang.
- Speichern Sie die Bilder für jede Zelle im ausgewählten Ordner.
- Wenn der Scanvorgang abgeschlossen ist, beenden Sie die Software, fahren Sie den Computer herunter und schalten Sie den Lasercontroller, die Laser, den Mikroskopcontroller und die Lampen für das Durchlicht aus.

Abbildung 2: Konfokale Mikroskopie . (A) Fixierte Eizellen nach Durchführung des Immunfluoreszenzprotokolls und der DNA-Färbung, die sich in separaten Tropfen des Waschpuffers befinden, mit Mineralöl bedeckt, in eine Glasbodenschale gelegt und für die konfokale mikroskopische Bildgebung vorbereitet werden. Jeder Tropfen enthält eine andere experimentelle Kategorie. Das Bild wurde durch die Stereomikroskop-Okulare aufgenommen. Maßstabsleiste = 1 mm/100 μm für den vergrößerten Teil. (B) Glasbodenplatte auf dem konfokalen Mikroskoptisch. (C) Hellfeldbild von Eizellen, aufgenommen durch konfokale Mikroskopie. Maßstabsbalken = 100 μm. (D) Das konfokale Mikroskopiesystem. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}
5. Bildgebende Analyse
- Laden Sie Fiji ImageJ-win64 im Browser herunter (https://imagej.net/software/fiji/downloads), öffnen Sie es und importieren Sie die Daten als TIFF-Stack-Dateien.
HINWEIS: Öffnen Sie jede Eizelldatei separat. - Klicken Sie auf Bild | Farbe | Kanäle teilen , um alle Kanäle aufzuteilen.
- Klicken Sie auf LUT (Look-up-Tabelle) und wählen Sie die bevorzugten Farben für jeden Kanal.
- Klicken Sie auf Bild | Farbe | Merge Channels , um die Kanäle für γΗ2ΑΧ und DNA zusammenzuführen. Lassen Sie den Hellfeldkanal nicht zusammengeführt.
- In NSN-Oozyten und in SN-Oozyten mit geringer DNA-Schädigung wird γΗ2ΑΧ als Herde in der DNA-Region nachgewiesen. Klicken Sie in diesem Fall auf den Befehl "Mehrpunkt" oder "Punkt" und wählen Sie jeden γΗ2ΑΧ-Fokus aus, der mit der DNA übereinstimmt. Wiederholen Sie diesen Schritt für alle Stapel.
- In SN-Oozyten mit hoher DNA-Schädigung ist das γΗ2ΑΧ-Signal über die gesamte DNA-Region verteilt. Klicken Sie in diesem Fall auf Bild | Stapel | Z-Projekt, und wählen Sie mit dem Befehl Freihandauswahl den gesamten DNA-Bereich aus.
- Um die γΗ2ΑΧ-Fluoreszenz zu messen, klicken Sie auf Analysieren | Messen Sie, und kopieren Sie die Messungen in eine .xlsx-Datei. Berechnen Sie dann die mittlere Fluoreszenz, normalisieren Sie die Werte und zählen Sie die Anzahl der Brennpunkte, bevor Sie Diagramme erstellen.
- Klicken Sie auf Analysieren | Legen Sie Maßstab fest, um den Maßstab festzulegen, und klicken Sie dann auf Analysieren | Werkzeuge | Maßstabsleiste , um den Kanälen eine Maßstabsleiste hinzuzufügen.
Ergebnisse
Mit dem hier gezeigten Verfahren wurden die Eierstöcke der Maus präpariert, das Fett entfernt und ausgewachsene Eizellen im GV-Stadium entnommen. Anschließend wurden die Kumuluszellen durch wiederholtes Pipettieren mit einer schmalen Pipette entnommen und in frische Tropfen M2-IBMX-Medium gegeben und mit Mineralöl auf einem heißen Block (37 °C) bedeckt (Abbildung 1A-F). Drei verschiedene Etoposidkonzentrationen (5 μg/ml, 20 μg/ml und 50 μg/ml) wurden unter Verwendung einer Stamm-Etoposidkonzentration von 20 mg/ml hergestellt. Die Eizellen im GV-Stadium wurden in drei verschiedenen Etoposid-Konzentrationen für 1 h in mit Mineralöl bedeckten und lichtgeschützten Tropfen bei 37 °C gegeben. Anschließend wurde das Immunfluoreszenzprotokoll befolgt, wie im Protokollabschnitt ausführlich beschrieben, und die Eizellen wurden in Glasbodenschalen gelegt und durch konfokale Mikroskopie beobachtet (Abbildung 2).
In den Oozyten des SN-GV-Stadiums stieg das Vorhandensein von γH2AX unmittelbar nach der DNA-Schädigung bei allen Etoposidkonzentrationen (5 μg/ml, 20 μg/ml und 50 μg/ml) an, und das γH2AX war über die gesamte DNA-Region verteilt (Abbildung 3). Die DSB-Quantifizierung und -Schätzung erfolgte durch die Beobachtung der γH2AX-Fluoreszenzintensität an DNA-Stellen. Die γH2AX-Fluoreszenz verstärkte sich proportional mit steigender Etoposidkonzentration. Darüber hinaus zeigten die Oozyten im GV-Stadium nach einem langwierigen Prophase-Arrest (20 Stunden nach der Etoposid-Behandlung) die Fähigkeit, die Anzahl und Intensität der γH2AX-Foci zu reduzieren, was auf das Vorhandensein aktiver Reparaturprozesse in den GV-Stadium-Arrest-Oozyten hindeutet (Abbildung 3E).
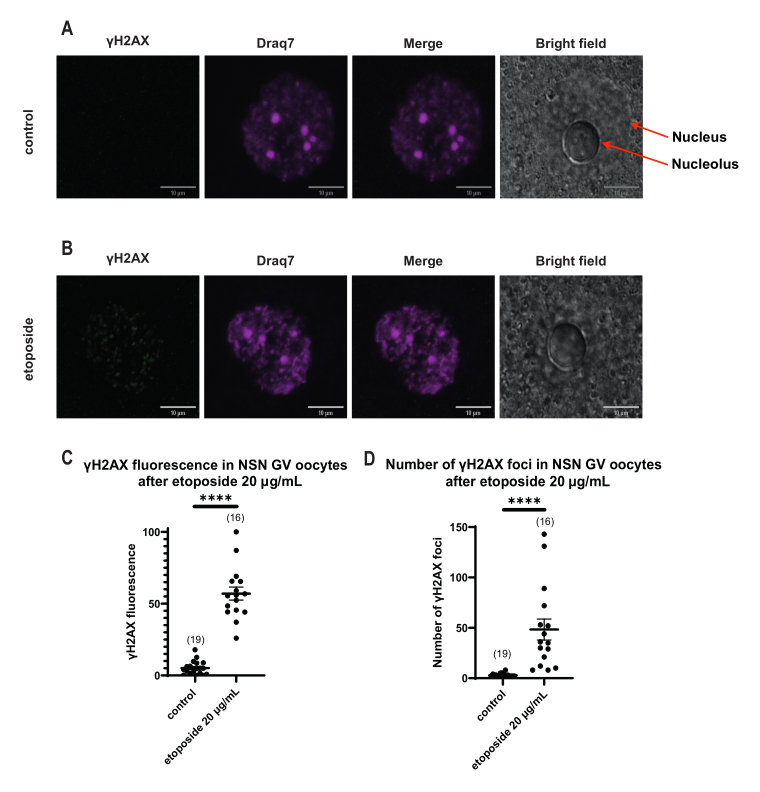
Im Gegensatz zu den SN-Oozyten, bei denen die γH2AX-Fluoreszenz über die DNA verteilt war, zeigte sich in den NSN-Oozyten γH2AX in den Herden unmittelbar nach der Behandlung mit Etoposid bei 20 μg/ml. Wir schätzten die Anzahl der Herde, die mit dem DNA-Bereich übereinstimmten, berechneten die Fluoreszenz jedes Schwerpunkts und präsentierten die mittlere Fluoreszenz aller Eizellen. Sowohl die Fluoreszenz als auch die Anzahl der Herde zeigten statistisch signifikante Unterschiede zwischen den beiden Eizellkategorien (Abbildung 4).
Die konfokale Mikroskopie liefert Informationen über die Anzahl und Intensität von Herden in verschiedenen Z-Stapeln und hilft so, das Vorhandensein von DNA-Schäden und die Reparaturdynamik zu bestimmten Zeitpunkten zu identifizieren. Das Galvano-Scannen bietet präzises Scannen mit geringem Hintergrund und eine bessere Analyse der Scanbilder.

Abbildung 3: Reduktion von γH2AX in Oozyten im SN-GV-Stadium, die mit drei verschiedenen Etoposid-Konzentrationen nach langwierigem GV-Arrest behandelt wurden. (A) γH2AX-Fluoreszenz in SN-GV-Stadien 0 h nach Etoposid-Behandlung. Der γH2AX-Wert steigt unmittelbar nach der Exposition bei allen Etoposid-Konzentrationen an, und der Anstieg ist konzentrationsabhängig (grün: γΗ2ΑΧ, magenta: DNA). Bei den Bildern handelt es sich um Z-Stack-Projektionen, und die Helligkeit/der Kontrast wurden für jeden Kanal mit Fiji / ImageJ angepasst. Skalenbalken = 10 μm. (B) Diagramm der γH2AX-Fluoreszenz in Oozyten im SN-GV-Stadium 0 h nach Behandlung mit unterschiedlichen Etoposidkonzentrationen. Die Daten stellen den Mittelwert ± SEM dar. Jeder Punkt steht für eine Eizelle (die Anzahl der Eizellen ist in der Grafik dargestellt) (ns = nicht signifikant, ** p < 0,005, **** p < 0,0001, Einweg-ANOVA mit dem Tukey-Mehrfachvergleichstest). (C) γH2AX-Fluoreszenz in Oozyten im SN-GV-Stadium 20 h nach Etoposidbehandlung. γH2AX reduziert sich 20 h nach der Exposition bei allen Etoposid-Konzentrationen (grün: γΗ2ΑΧ, magenta: DNA). Bei den Bildern handelt es sich um Z-Stack-Projektionen, und die Helligkeit/der Kontrast wurden für jeden Kanal mit Fiji/ImageJ angepasst. Maßstabsbalken = 10 μm. (D) Diagramm der γH2AX-Fluoreszenz in Oozyten im SN-GV-Stadium 20 h nach Behandlung mit unterschiedlichen Etoposidkonzentrationen. Die Daten stellen den Mittelwert ± SEM dar. Jeder Punkt steht für eine Eizelle (die Anzahl der Eizellen ist in der Grafik dargestellt), (ns = nicht signifikant, * p < 0,05, ** p < 0,005, *** p < 0,0005 , **** p < 0,0001, Einweg-ANOVA mit Tukey-Mehrfachvergleichstest). (E) Balkendiagramm der γH2AX-Fluoreszenzreduktion in Oozyten im SN GV-Stadium nach Prophasenarrest in mit Etoposid behandelten Oozyten. Die Zahl über jeder Spalte gibt den prozentualen Rückgang der γH2AX-Fluoreszenz an. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 4: Phosphorylierung von Η2ΑΧ in Oozyten im NSN-GV-Stadium nach Behandlung mit Etoposid bei 20 μg/ml. (A) Repräsentative konfokale Bilder einer Kontroll-NSN-Oozyte im GV-Stadium (grün: γΗ2ΑΧ, magenta: DNA). Bei den Bildern handelt es sich um Z-Stack-Projektionen, und die Helligkeit/der Kontrast wurden für jeden Kanal mit Fiji/ImageJ angepasst. Maßstabsbalken = 10 μm. (B) Repräsentative konfokale Bilder einer mit Etoposid behandelten NSN-Oozyte im GV-Stadium (grün: γΗ2ΑΧ, magenta: DNA). Die Eizellen wurden 0 h nach der Etoposidbehandlung fixiert. Bei den Bildern handelt es sich um Z-Stack-Projektionen, und Helligkeit/Kontrast wurden für jeden Kanal mit Fiji/ImageJ angepasst. Maßstabsbalken = 10 μm. (C) Die normalisierte γΗ2ΑΧ-Fluoreszenz in NSN-Eizellen im GV-Stadium nach 20 μg/ml Etoposidbehandlung. Die Daten stellen den Mittelwert ± SEM dar. Jeder Punkt steht für eine Eizelle (die Anzahl der Eizellen ist in der Grafik dargestellt), die aus zwei unabhängigen Experimenten stammt (**** p < 0,0001, ungepaarter nicht-parametrischer t-Test, Mann-Whitney-U-Test). (D) Anzahl der γΗ2ΑΧ-Herde in Oozyten im NSN-GV-Stadium nach 20 μg/ml Etoposidbehandlung. Die Daten stellen den Mittelwert ± SEM dar. Jeder Punkt steht für eine Eizelle (die Anzahl der Eizellen ist in der Grafik dargestellt), die aus zwei unabhängigen Experimenten stammt (**** p < 0,0001, ungepaarter nicht-parametrischer t-Test, Mann-Whitney-U-Test). Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}
Diskussion
Mit der hier beschriebenen Methode konnten wir DSBs in Säugetieroozyten nachweisen. Diese Methode ermöglicht den Nachweis und die Untersuchung des DNA-Reparaturprozesses in Eizellen. Das gleiche Protokoll könnte auch für die Analyse anderer Proteine verwendet werden, die an physiologischen Prozessen in Säugetiereizellen beteiligt sind. Es ist wichtig zu untersuchen, wie Eizellen auf mögliche DNA-Schäden reagieren, um die Ursache der weiblichen Subfertilität beim Menschen besser zu verstehen.
Die Untersuchung der DNA-Schadensantwort in Säugetier-Eizellen kann aufgrund der Empfindlichkeit der Eizellen eine Herausforderung darstellen. Der Umgang mit Eizellen erfordert bestimmte Temperaturen und CO2- und O2-Konzentrationen. Gleichzeitig müssen die Eizellen vor Licht geschützt werden. Die Handhabung sollte mit Glaspipetten erfolgen, die nicht zu schmal sind, da dies für die Eizellen schädlich sein könnte, aber auch nicht zu breit, da dies zu einer Verdünnung des Mediums führen und somit den Fixierungsvorgang negativ beeinflussen könnte. Bei jedem Schritt der Fixierung werden mehrere Tropfen Puffer verwendet, um den Verdünnungseffekt zu minimieren. Eine alternative Methode zur Beobachtung von DSBs ist der Comet-Assay24. Auch wenn diese Technik empfindlicher ist, ist sie komplizierter. Gleichzeitig ist es durch die Verwendung des Comet-Assays nicht möglich, die genaue DNA-Region zu erkennen, in der die Schädigung auftritt, und in Zellen mit reichlich RNA-Molekülen, wie z. B. Eizellen im GV-Stadium25, könnte der Hintergrund erhöht werden, was zu einem falschen DNA-Schadenssignal26 führt.
Durch die Verwendung des hier beschriebenen Immunfluoreszenzprotokolls können wir DSBs mit Genauigkeit nachweisen und den Reparaturfortschritt in Eizellen im GV-Stadium abschätzen, wie durch die Verringerung der γH2AX-Fluoreszenz im Laufe der Zeit angezeigt. Eine Einschränkung dieser Methode besteht jedoch darin, dass bestimmte Antikörper eine unspezifische Verteilung im gesamten Ooplasma aufweisen können, was zu Bildern mit hoher Hintergrundfluoreszenz führt. Der PFA-Tx-100-Puffer wird anstelle von sequentiellem PFA und Tx-100 verwendet, da wir beobachtet haben, dass er den Fixierungsprozess verbessert, indem er die Detektion von weniger Hintergrund- und unspezifischer Fluoreszenz ermöglicht. Eine zweite Einschränkung bei der Verwendung von γH2AX für die DSB-Detektion besteht darin, dass die Schädigung nach GVBD aufgrund der spontanen Phosphorylierung von γH2AX in der Meiose23 nicht abgeschätzt werden kann.
Bei diesem Immunfluoreszenzprotokoll verbleiben die Eizellen in einem flüssigen Puffer und können nicht in Objektträgern gespeichert werden. Diese Tatsache macht es schwierig, die fixierten Zellen für Tage nach der Zugabe des sekundären Antikörpers zu konservieren. Um eine gute Bildqualität zu erhalten und das Signal nicht zu verlieren, ist es vorzuziehen, die Bildgebung innerhalb weniger Stunden nach der Zugabe des sekundären Antikörpers durchzuführen. Es sollte auch beachtet werden, dass das Abtasten der Kerne durch die Z-Achse dazu führen kann, dass das Signal durch Überbelichtung schwächer wird. Aus diesem Grund ist es vorzuziehen, die Laserleistung zu verringern und die Geschwindigkeit des Scannens zu erhöhen.
Eine weitere Einschränkung des Immunfluoreszenzprotokolls besteht darin, dass es nur für fixierte/nicht lebende Zellen verwendet werden kann. Daher können wir nur das Vorhandensein und Fehlen von Faktoren zu bestimmten Zeitpunkten schätzen, ohne zu wissen, ob es Schwankungen in ihrer Konzentration oder Veränderungen in ihrem Verhalten im Laufe der Zeit gibt. Dieses Problem konnte durch den Einsatz von Lebendzell-Bildgebung und fluoreszenzmarkierten Markern überwunden werden.
Offenlegungen
Die Autoren haben keine Interessenkonflikte.
Danksagungen
Wir danken für die Unterstützung dieser Arbeit durch das Projekt "Etablierung von 'Capacity Building'-Infrastrukturen in der biomedizinischen Forschung (BIOMED-20)" (MIS 5047236), das im Rahmen der Aktion "Stärkung der Forschungs- und Innovationsinfrastruktur" durchgeführt wird, die durch das operationelle Programm "Wettbewerbsfähigkeit, Unternehmertum und Innovation" (NSRP 2014-2020) finanziert und von Griechenland und der Europäischen Union (Europäischer Fonds für regionale Entwicklung) kofinanziert wird.
Materialien
| Name | Company | Catalog Number | Comments |
| 3 mL Pasteur pipettes in LDPE, graduated | APTACA | 1502 | |
| 10 cc syringes | SoftCare | 114.104.21 | |
| Alexa Fluor 488-conjugated goat anti-rabbit Secondary Ab | Biotium | 20012 | |
| Anti-phospho-H2A.X (Ser139) | Merck Millipore | 07-164 | |
| ARE Heating Magnetic Stirrer | VELP Scientifica | F20500162 | |
| BD FALCON 5 mL Polystyrene Round-Bottom Tubes | BD Biosciences | 352054 | |
| BD Microlance 3 Needles 27 G - 0.40 x 13 mm | Becton Dickinson | 300635 | |
| Bovine Serum Albumin Fraction V | Roche | 10735078001 | |
| DMSO Anhydrous | Biotium | 90082 | |
| DRAQ7 DNA dye | BioStatus | DR71000 | |
| EGTA | Sigma-Aldrich | E4378-25G | |
| EMSURE MgCl2. 6H2O | Merck Millipore | 1058330250 | |
| Etoposide | CHEMIPHARM | L01CB01 | |
| FALCON 14 mL Polystyrene Round-Bottom Tubes | Corning Science | 532057 | |
| FALCON Tissue Culture Dishes, Easy-Grip, 35 x 10 mm Style | Corning Science | 353001 | |
| Glass Bottom Culture Dishes (35 mm Petri dish/ 14 mm Microwell, No. 0 coverglass) | MatTek Corporation | P35G-0-14-C | |
| HEPES | Sigma-Aldrich | H6147-25G | |
| HERACELL 150i CO2 Incubator | ThermoFisher Scientific | 50116048 | |
| IBMX powder | Sigma-Aldrich | I5879-100MG | |
| Leica M125 Stereo Microscope | Leica Microsystems | ||
| M16 Medium | Sigma-Aldrich | M7292 | |
| M2 Medium | Sigma-Aldrich | M7167 | |
| Mineral Oil | Sigma-Aldrich | M5310 | |
| NaN3 | Honeywell | 13412H | |
| NaOH | Merck Millipore | 1064981000 | |
| Nikon AX ECLIPSE Ti2 Confocal Microscope | Nikon Corporation | ||
| Nikon SMZ800N Stereo Microscope | Nikon Corporation | ||
| Paraformaldehyde | Sigma-Aldrich | 158127 | |
| Pasteur pippettes, glass, long form 230 mm | DURAN WHEATON KIMBLE | 357335 | |
| pH/ORP meter | Hanna Instruments Ltd | HI2211 | |
| Phosphate buffered saline tablets | Sigma-Aldrich | P4417-100TAB | |
| PIPES | Sigma-Aldrich | P1851 | |
| PMSG Protein Lyophilised | Genway Biotech (now AVIVA Systems Biology) | GWB-2AE30A (now OPPA01037) | |
| QBD4 Dry block heater | Grant Instruments (Cambridge) Ltd | A25218 | |
| Triton X-100 | Sigma-Aldrich | T8787 | |
| Whatman Puradisc 25 mm 0.2 μm filters | GE Healthcare | 6780-2502 |
Referenzen
- Wang, X., Pepling, M. E. Regulation of meiotic prophase one in mammalian oocytes. Frontiers in Cell and Developmental Biology. 9, 667306(2021).
- Filatov, M., Khramova, Y., Semenova, M. Molecular mechanisms of prophase I meiotic arrest maintenance and meiotic resumption in mammalian oocytes. Reproductive Sciences. 26 (11), 1519-1537 (2019).
- Adhikari, D., et al. Inhibitory phosphorylation of Cdk1 mediates prolonged prophase I arrest in female germ cells and is essential for female reproductive lifespan. Cell Research. 26 (11), 1212-1225 (2016).
- Sun, S. C., Kim, N. H. Molecular mechanisms of asymmetric division in oocytes. Microscopy and Microanalysis. 19 (4), 883-897 (2013).
- Jones, K. T. Mammalian egg activation: From Ca2+ spiking to cell cycle progression. Reproduction. 130 (6), 813-823 (2005).
- Solc, P., Schultz, R. M., Motlik, J. Prophase I arrest and progression to metaphase I in mouse oocytes: comparison of resumption of meiosis and recovery from G2-arrest in somatic cells. Molecular Human Reproduction. 16 (9), 654-664 (2010).
- Parfenov, V., Potchukalina, G., Dudina, L., Kostyuchek, D., Gruzova, M. Human antral follicles: oocyte nucleus and the karyosphere formation (electron microscopic and autoradiographic data). Gamete Research. 22 (2), 219-231 (1989).
- Zuccotti, M., Piccinelli, A., Giorgi Rossi, P., Garagna, S., Redi, C. A. Chromatin organization during mouse oocyte growth. Molecular Reproduction and Development. 41 (4), 479-485 (1995).
- Sun, X., et al. Comprehensive analysis of nonsurrounded nucleolus and surrounded nucleolus oocytes on chromatin accessibility using ATAC-seq. Molecular Reproduction and Development. 90 (2), 87-97 (2023).
- Zuccotti, M., Bellone, M., Longo, F., Redi, C. A., Garagna, S. Fully-mature antral mouse oocytes are transcriptionally silent but their heterochromatin maintains a transcriptional permissive histone acetylation profile. Journal of Assisted Reproduction and Genetics. 28 (12), 1193-1196 (2011).
- Winship, A. L., Stringer, J. M., Liew, S. H., Hutt, K. J. The importance of DNA repair for maintaining oocyte quality in response to anti-cancer treatments, environmental toxins and maternal ageing. Human Reproduction Update. 24 (2), 119-134 (2018).
- Lieber, M. R. The mechanism of human nonhomologous DNA end joining. The Journal of Biological Chemistry. 283 (1), 1-5 (2008).
- Chiruvella, K. K., Liang, Z., Wilson, T. E. Repair of double-strand breaks by end joining. Cold Spring Harbor Perspectives in Biology. 5 (5), 012757(2013).
- Chang, H. H. Y., Pannunzio, N. R., Adachi, N., Lieber, M. R. Non-homologous DNA end joining and alternative pathways to double-strand break repair. Nature Reviews. Molecular Cell Biology. 18 (8), 495-506 (2017).
- Shibata, A., Jeggo, P. A. Roles for the DNA-PK complex and 53BP1 in protecting ends from resection during DNA double-strand break repair. Journal of Radiation Research. 61 (5), 718-726 (2020).
- Mohiuddin, I. S., Kang, M. H. DNA-PK as an emerging therapeutic target in cancer. Frontiers in Oncology. 9, 635(2019).
- Marangos, P., Carroll, J. Oocytes progress beyond prophase in the presence of DNA damage. Current Biology. 22 (11), 989-994 (2012).
- Paull, T. T., et al. A critical role for histone H2AX in recruitment of repair factors to nuclear foci after DNA damage. Current Biology. 10 (15), 886-895 (2000).
- Bakkenist, C. J., Kastan, M. B. DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation. Nature. 421 (6922), 499-506 (2003).
- Marangos, P., et al. DNA damage-induced metaphase I arrest is mediated by the spindle assembly checkpoint and maternal age. Nature Communications. 6, 8706(2015).
- Lavrentyeva, E. A., Shishova, K. V., Zatsepina, O. V. Differences in nuclear dynamics in mouse GV oocytes with a diverse chromatin configuration. Biology Bulletin Russian Academy of Sciences. 46, 332-341 (2019).
- Montecucco, A., Zanetta, F., Biamonti, G. Molecular mechanisms of etoposide. EXCLI Journal. 14, 95-108 (1998).
- Mayer, A., et al. DNA damage response during mouse oocyte maturation. Cell Cycle. 15 (4), 546-558 (2016).
- Olive, P., Banáth, J. The comet assay: A method to measure DNA damage in individual cells. Nature Protocols. 1, 23-29 (2006).
- Wu, D., Dean, J. EXOSC10 sculpts the transcriptome during the growth-to-maturation transition in mouse oocytes. Nucleic Acids Research. 48 (10), 5349-5365 (2020).
- Simon, L., Emery, B., Carrell, D. DNA damage: COMET assay. Manual of Sperm Function Testing in Human Assisted Reproduction. Agarwal, A., Henkel, R., Majzoub, A. , 202-212 (2021).
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten