
Method Article
Une méthode polyvalente pour le montage de Arabidopsis quitte pour l’imagerie Intravitale time-lapse
Dans cet article
Résumé
Nous présentons une méthode simple et souple pour l’exécution de vivre-imagerie fluorescente de Arabidopsis thaliana feuilles sur une longue période de temps. Nous utilisons une plante Arabidopsis transgénique exprimant un gène rapporteur fluorescent sous le contrôle d’un promoteur liées à l’immunité à titre d’exemple pour comprendre spatio-temporelle des réactions immunitaires des plantes.
Résumé
La réponse immunitaire de plantes associée à une reprogrammation transcription du génome est lancée sur le site de l’infection. Ainsi, la réponse immunitaire est réglementée dans le temps et l’espace. L’utilisation d’un gène fluorescent sous le contrôle d’un promoteur liées à l’immunité en combinaison avec un microscope de fluorescence automatisé est un moyen simple de comprendre le règlement spatio-temporelle de l’immunité de la plante. Par contraste avec les tissus de la racine qui ont été utilisés pour un certain nombre de différentes expériences d’imagerie fluorescentes intravitale, il existe peu d’exemples d’imagerie live fluorescent pour les tissus foliaires qui rencontrent un tableau d’infections microbiennes aéroportés. Par conséquent, nous avons développé une méthode simple pour monter les feuilles des plantes Arabidopsis thaliana pour l’imagerie de cellules vivantes sur une longue période de temps. Nous avons utilisé les plantes Arabidopsis transgéniques exprimant la protéine fluorescente jaune (YFP) de genefused pour le signal de localisation nucléaire (NLS) sous le contrôle du promoteur d’un gène marqueur liées à la défense, 1 Pathogenesis-Related ) PR1). Nous avons infiltré une feuille transgénique avec Pseudomonas syringae pv. tomate DC3000 (avrRpt2) (Pst_a2) de la souche et effectué l’imagerie in vivo de time-lapse du signal YFP pour un total de 40 h en utilisant un stéréomicroscope de fluorescence automatisé. Cette méthode peut être utilisée non seulement pour les études sur les réactions immunitaires des plantes, mais aussi pour les analyses de divers événements développementaux et les réponses environnementales survenant dans les tissus foliaires.
Introduction
La réponse immunitaire des plantes implique une reprogrammation transcriptionnelle dynamique relevant de la transcription de plusieurs facteurs ainsi que de phytohormones1. L’accumulation de données transcriptome offre des possibilités de collecte d’information sur le système immunitaire de végétaux : par exemple, la structure du réseau de signalisation cascades2. Cependant, notre connaissance de la dynamique spatiale et temporelle de l’immunité de la plante reste limitée3,4,5.
Dans des études antérieures, règlement spatio-temporelle de l’expression des gènes reliés à la défense a été surtout analysée à l’aide de l’hybridation in situ et un β-glucuronidase (GUS) journaliste dosage6,7,8. Ces méthodes permettent de visualiser l’activation transcriptionnelle des gènes différents d’intérêt in situ. Toutefois, ces procédures nécessitent une fixation chimique des échantillons et donc se traduire par la perte de toutes les informations temporelles. Événements biologiques, tels que l’immunité, des progrès au fil du temps. L’utilisation de la luciférase comme journaliste a permis la capture de la dynamique temporelle du promoteur d’intérêt3. Cependant, essai luciférase nécessite un substrat coûteux et détecteurs très sensibles. Accroître notre compréhension des aspects spatio-temporels de la réponse immunitaire de plante à l’aide d’une procédure simple, nous avons produit transgénique Arabidopsis thaliana plantes exprimant la protéine fluorescente jaune (YFP) gène fusionné à la signal de localisation nucléaire (YFP-NLS) sous le contrôle du promoteur du gène marqueur liées à la défense, Pathogenesis-Related 1 (PR1)9. Nous avons utilisé la chlorophylle autofluorescence, un marqueur des cellules vivantes, de saisir le processus de mort cellulaire programmée (PCD), qui se produit souvent au cours de l’effecteur déclenchée l’immunité (ETI), une forme d’immunité végétale induite par l’agent pathogène spécifique infection9 , 10. suivi de la dynamique temporelle de l’intensité de signal de fluorescence en se déplaçant librement les objets, tels que vivent les feuilles de l’Arabidopsis , nécessite une image complexe traitement logiciel interne intégré ou disponible dans le commerce. Alternativement, empêchant les spécimens de déménagement est une méthode simple pour résoudre le problème. Ici, nous avons développé une méthode simple et versatile pour montage vivant des feuilles d’une plante Arabidopsis transgénique pour l’observation à long terme sous un stéréomicroscope de fluorescence automatisé. La méthode permet de capturer le promoteur dynamique au sein de la plante intacte cultivés en sol laisse pendant quelques jours.
Protocole
Remarque : Il est important d’empêcher le sommeil circulation des feuilles vivantes des échantillons en time-lapse imagerie. Pour minimiser les contraintes mécaniques sur les feuilles, fixation douce des feuilles est nécessaire. Les échantillons de feuilles bien préparés uniquement produisent des images time-lapse adaptés aux diverses analyses d’image. Un protocole utilisant des plantes Arabidopsis transgéniques exprimant YFP-NLS fusion sous le contrôle du promoteur du gène PR1 (plantes pPR1-YFP-NLS) et Pseudomonas syringae pv. tomate DC3000 souche (avrRpt2) (Pst_a2) est décrit ci-dessous, à titre d’exemple.
1. préparation de plantes et pathogènes
- Remplir un germoir en plastique cellulaire (Table des matières) avec sol autoclavé.
- Semer une graine de Arabidopsis transgénique par cellule (Figure 1A).
- Le plateau de transfert à une chambre de croissance maintenue à 23 ° C et cultiver les plantes sous une lumière blanche continue pendant 2-3 semaines.
- Deux jours avant l’inoculation de l’agent pathogène, ensemencer Pseudomonas syringae pv. tomate DC3000 avrRpt2 (Pst_a2) de la souche provenant d’un stock de glycérol sur NYG moyen (peptone de 5 g/L, extrait de levure 3 g/L, glycérol 20 mL/L, agar bactériologique de 15 g/L, pH 7,0) contenant 100 mg/L rifampicine et 50 mg/L kanamycine et incuber à 28 ° C pendant 48 h.
- Récolter les cellules bactériennes qui apparaissent sur la surface du milieu à l’aide de pointes en plastique, transférez-les dans un tube en plastique contenant 10 mM MgCl2et remettre en suspension. Mesurer la densité optique (Do) de la solution à 600 nm (OD600). Ajuster la concentration finale de cellules des cellules bactériennes à 108 unités formation de colonies (UFC) / mL, ce qui correspond normalement à OD600 = 0,211.
2. l’inoculation pathogène
- Découper soigneusement une fiche de cellule contenant une plante âgée de 2-3 semaine sans endommager la plante. La valeur de la cellule dans un germoir de cellule vide (2 x 2 cellules suffisent) pour maintenir un bon équilibre (Figure 1B).
- Sélectionnez une feuille visiblement saine pour l’inoculation. En règle générale, la troisième, quatrième et cinquième laisse (#3, #4 et #5, respectivement, dans la Figure 1C) du bas de la plante sont faciles à manipuler. Utilisation laisse à la même position dans une série d’expériences pour la meilleure reproductibilité. Eau du sol, tenant la plante avant l’inoculation d’imagerie time-lapse à long terme.
- Éventuellement, dans le cas d’analyse des promoteurs sensibles aux stress tels que pPR1, pour s’assurer que les plantes ne sont pas naturellement soulignés, examinez les feuilles sous un stéréomicroscope de fluorescence avant l’inoculation de l’agent pathogène afin de vérifier l’absence de signal de la YFP. Exclure les feuilles signal YFP montrant de l’expérience.
- Porter des gants jetables en latex avant infiltration pour éviter un contact direct avec l’agent pathogène. À l’aide d’une seringue plastique sans aiguille de 1 mL, infiltrer soigneusement le côté abaxial de la feuille avec la suspension bactérienne (108 UFC/mL)11 (Figure 1D). L’inoculation d’une petite portion sur la moitié de la feuille permet une bonne visualisation de l’activité de pPR1 ; la zone infiltrée devient visible en vert plus foncé en couleur par rapport à la feuille restante. Être extrêmement attention à ne pas causer aucun dommage mécanique à la feuille lors de l’infiltration.
Remarque : Assurez-vous que tous les espaces intercellulaires, dans la région infiltrée sont complètement remplies (verticalement) avec la suspension de l’agent pathogène ; dans le cas contraire, le domaine PCD sera difficile à visualiser sous le stéréomicroscope fluorescent. Cela peut être simplement confirmé par le parachèvement d’écologisation sombre dans la zone infiltrée. - Absorber l’excès d’une suspension bactérienne de la zone qui entoure la section infiltrée de la feuille infiltrée avec une serviette en papier doux.
3. montage de la feuille inoculée
- Immédiatement après l’inoculation, fixer une lame de verre sur le plateau en plastique à l’aide de ruban chirurgical (Table des matières), telle que la feuille infiltrée se trouve au centre de la lame de verre. Veiller à ce que la lame feuille inoculée est complètement montée à l’intérieur de la lame de verre (Figure 2A).
- Préparer deux couches de ruban en plastique (dans le cas de bande épaisse de 0,2 mm, voir la Table des matières) et le couper en deux morceaux (Figure 2B; pièces 1 et 2) pour adapter les espaces le long du pétiole de la feuille infiltrée. Organiser la longueur de ces deux pièces, indiqué par une flèche à deux pointes dans la Figure 2B, pour s’adapter à la longueur de la flèche à deux pointes, illustrée à la Figure 2A.
Remarque : En découpant un corner de chacune des deux pièces permet d’éviter les dommages au Limbe (Figure 2B, pointes de flèches ; voir également l’étape 3.4 ci-dessous). N’importe quel genre de bande en plastique d’épaisseur similaire est adapté pour la fabrication d’un pont sur le pétiole. La rigidité de la bande en plastique est importante pour une manipulation aisée au cours de la procédure décrite ci-dessous. - À l’aide de pinces fines, coller morceaux de bandes 1 et 2 de chaque côté du pétiole tels que les coins coupés de chaque pièce s’aligner sur la base du limbe (Figure 2C). Veiller à ce que les morceaux de ruban adhésif ne pas touchent la lame pétiole ou feuille.
Remarque : Ces morceaux de ruban en plastique double couche à la base du limbe agissent comme entretoises et prévenir le stress physique sur le pétiole lors du montage. - Préparer un autre morceau de ruban en plastique double couche (Figure 2B, pièce 3) pour adapter la taille de la double-flèche dans la Figure 2B et le coller sur le dessus les morceaux de bandes 1 et 2 pour former un pont sur le pétiole ( Figure 2D, E). Être extrêmement attention à ne pas attraper la lame pétiole et feuilles directement entre les morceaux de ruban adhésif aux positions indiquées par des flèches dans la Figure 2D, E.
- Doucement de coller un petit morceau de ruban chirurgical (Figure 2F, pièce 4) sur la lame de verre au-dessus de l’extrémité du limbe afin que le limbe est fixé très doucement sur la lame de verre. Seulement appuyer fermement la partie du ruban chirurgical touchant directement la lame de verre (Figure 2F, région décrite avec une ligne rouge en pointillés), pas l’autre partie recouvrant les feuilles.
- Doucement s’en tenir un autre petit morceau de ruban chirurgical (Figure 2G, pièce 5) à la frontière du pétiole et ruban plastique pièces (1, 2 et 3) afin que le pétiole est fixé très doucement sur la lame de verre et de la bande en plastique pièces. Seulement fixer fermement la partie du ruban chirurgical touchant directement la lame de verre à la lame de verre et des morceaux de ruban en plastique (Figure 2G, zone décrite avec une ligne rouge en pointillés), pas l’autre partie recouvrant le pétiole.
- Empêcher les feuilles voisines de se déplacer dans le champ de vision du microscope à l’aide de pointes de pipette 200 µL (Figure 2H). Insérer les pointes de pipette dans le sol pour tenir doucement les feuilles voisines de la feuille infiltrée. Veillez à ne pas insérer les pointes trop profondément dans le sol pour éviter d’endommager la racine possible.
Remarque : L’usine de prêt est maintenant prêt pour l’imagerie de fluorescence stéréomicroscope.
4. microscopique observation time-lapse
- Allumez le stéréomicroscope fluorescent.
NOTE : Ici, un stéréomicroscope automatique équipé d’un monochrome hautement sensibles de 1,4 millions de pixels appareil photo numérique en mode 12 bits est utilisé (Table des matières). Le microscope doit être placé dans une pièce sombre, équipé d’un système de climatisation ou dans une armoire sombre avec une ventilation suffisante pour maintenir la température de la pièce à 23 ° C en time-lapse imagerie. - Mettre les plantes dans l’espace ci-dessus sous l’objectif du stéréomicroscope pour l’imagerie.
- Définir les paramètres pour l’imagerie time-lapse (voir exemples dans le tableau 1). Assurez-vous que programmation des étapes pour l’exposition à la lumière pendant la période d’intervalle de time-lapse imagerie puisque la lumière a un impact majeur sur les plantes l’immunité12.
- Utiliser le filtre YFP classique (excitation 500-520 nm ; émission 540-580 nm) pour visualiser le signal de la YFP.
- Utilisez le filtre de Texas Red (TXR) (excitation 540-580 nm ; longue passe de d’émission 610 nm) pour visualiser la chlorophylle autofluorescence afin que le domaine PCD est visible comme une zone sombre (aucune autofluorescence) entourée par YFP séropositifs cellule couches9 (Figure 3 ( A).
- Utilisez les paramètres de champ d’epi-lumineuses classiques pour les étapes de l’exposition à la lumière supplémentaire.
NOTE : Avant l’expérience, mesurer la puissance de la source lumineuse de l’epi-lumineux et l’ajuster au niveau de la condition de croissance propre usine.
- Exécutez le programme d’imagerie time-lapse. Pour l’observation à long terme sur plusieurs jours, envisager d’arroser la plante appropriée, par exemple, au cours des étapes de l’exposition à la lumière.
- Après l’acquisition d’images, omettre les canaux supplémentaires utilisés pour l’exposition à la lumière dans les intervalles (correspondant aux canaux 3-8 dans le tableau 1) de l’ensemble de données. Analyser les données selon différentes méthodes, telles que l’analyse de région d’intérêt (ROI), avec logiciel d’analyse différents d’image tels que Fidji9.
Résultats
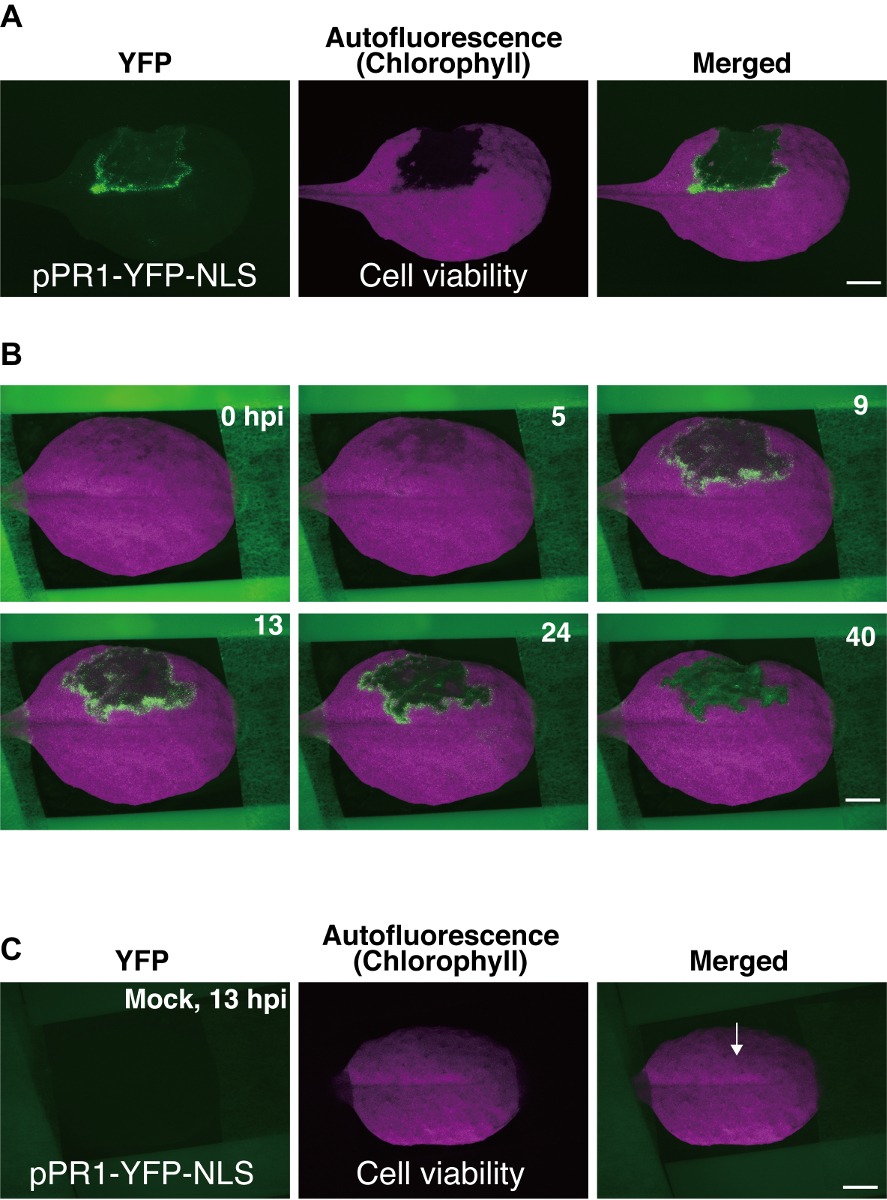
Ici, nous avons utilisé Pst_a2-induite des ETI à titre d’exemple d’imagerie time-lapse. Time-lapse données ont été obtenues comme une série d’images, dont quelques sont indiquées dans la Figure 3Bet comme un film time-lapse (supplémentaire 1 film)9. Dans les expériences réussies à l’aide de pPR1-YFP-NLS pendant Pst_a2-ETI induite, activation transitoire des pPR1 a été observée, comme le montre de YFP exprimant des foyers, dans plusieurs couches de cellules qui entourent le domaine PCD (Figure 3B) 9. l’activation des pPR1 dans les cellules qui entourent le domaine PCD habituellement commence à environ 5 heures après l’inoculation (hpi), pics à environ 12 hpi et dure jusqu'à 40 hpi (Figure 3B)9. Puisque les images ont été acquises grâce à un système d’épi-fluorescence, les signaux de la YFP fournis ici ont été générés de plusieurs couches de cellules adaxial, y compris les cellules du mésophylle épidermique comme supérieure.

Figure 1 : Méthode utilisée pour l’infiltration de la bactérie pathogène dans le Arabidopsis thaliana feuille. (A) deux à trois-semaines Arabidopsis plantes ont été cultivées dans un germoir de cellule. (B) unicellulaire fiche contenant la plante sélectionnée pour être utilisée pour l’inoculation et d’observation a été découpée et placée dans le germoir cellule vide pour maintenir l’équilibre. (C) La troisième, quatrième et cinquième laisse (#3, #4 et #5, respectivement) sont adaptés pour l’analyse de l’image. (D) l’Infiltration de la suspension de l’agent pathogène dans une feuille sélectionnée à l’aide d’une seringue sans aiguille. On peut reconnaître la région infiltrée basée sur sa couleur verte plus foncée que la feuille restante. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}

Figure 2 : Méthode utilisée pour le montage des infiltrés Arabidopsis feuille pour l’imagerie time-lapse. Photographie (A) de la plante Arabidopsis avec une lame de verre fixée sous la feuille inoculée. La feuille infiltrée a été placée au centre de la lame de verre. La double-flèche indique la longueur des entretoises en plastique ruban. (B) préparation des entretoises de ruban en plastique et pont. Double - couches de ruban en plastique a été coupé en deux pièces (1 et 2) pour les entretoises et un autre morceau (3, décrit avec une ligne en pointillés rouge) a été coupé pour préparer un pont. La longueur des flèches gauche-droite est presque identique à la longueur de la flèche à deux pointes (A). Un des quatre coins des pièces 1 et 2, a indiqué avec des flèches, ont été coupés. (C) deux morceaux de ruban en plastique double couche (numérotés 1 et 2) ont été soigneusement collées sur le verre de glisser le long du pétiole à l’aide de pinces fines, sans établir un contact direct avec la lame du pétiole et de la feuille. (D), un autre morceau de double couche en plastique ruban (numéroté 3), qui a été préparé à partir de (B), a été placé sur les deux entretoises basales (1 et 2) pour former un pont sur le pétiole, tout en assurant pour ne pas attraper le pétiole entre bandes 1 et 2 à positions indiquées par des flèches. (E) photographie montrant le pétiole collée. Il est important de ne pas attraper le tissu de la plante directement entre les morceaux de ruban en plastique aux positions indiquées par les flèches. Fun petit morceau de ruban chirurgical (numéroté 4) a été collé doucement sur la lame de verre autour de la pointe du limbe. Seule la zone décrite par la ligne pointillée rouge a été enfoncée fermement sur la lame de verre. (G), un autre petit morceau de ruban chirurgical (numéroté 5) a été légèrement collé à la frontière du pétiole et morceaux de ruban en plastique. Seule la zone décrite par la ligne pointillée rouge a été pressée pour la fixation du pétiole doucement sur le verre de la diapositive et morceaux de ruban en plastique. Pointes de pipette jetable (H), deux ont été utilisées pour empêcher les feuilles voisines de se déplacer dans le champ de vision de la stéréomicroscope. Les pointes de pipette furent insérées directement dans le sol aux positions appropriées. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}

Figure 3 : Représentant les résultats à l’aide de la Arabidopsis feuille de montage méthode. (A) Fluorescent des images d’une feuille d’Arabidopsis transgénique (usine de pPR1-YFP-NLS). Une partie de la feuille a été infiltrée avec Pst_a2 (108 UFC/mL) sur sa surface abaxiale, et les images ont été capturées à 22 heures après l’inoculation (hpi). Noyaux dans lequel a été activé le promoteur pPR1 ont été détectés en utilisant le filtre de la YFP, et la mort cellulaire programmée (PCD) a été détectée, fondée sur la perte de chlorophylle autofluorescence en utilisant le filtre de Texas Red (TXR). Echelle = 2,5 mm. (B) quelques images de time-lapse choisies parmi une série de 800 images obtenues en utilisant le protocole décrit ici. Ces données fournissent une vue d’ensemble in vivo de la dynamique spatio-temporelle de l’activité pPR1 pour 40 hpi. Une image fusionnée d’images YFP et TXR sont affichées. Echelle = Mock infiltration de 2,5 mm. (C) d’une feuille de transgénique (usine de pPR1-YFP-NLS). En infiltration simulacre, 10 mM MgCl2 a été infiltré dans le côté abaxial de la feuille, suivi par imagerie de time-lapse. Le traitement simulé n’a pas causé l’activité ectopique pPR1 et n’interfère pas avec la chlorophylle autofluorescence à 13 hpi. Une flèche indique la position d’infiltration simulée. Echelle = 2,5 mm. Ces images ont été modifiés depuis une précédente étude9. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}

Tableau 1 : Time-lapse programme d’imagerie utilisée dans cette étude.

Supplémentaire film 1 : time-lapse film montrant la dynamique spatio-temporelle des pPR1 activation en une transgéniques Arabidopsis feuilles suivante déclenchées effecteur l’immunité (ETI) induite par Pst_a2 inoculation. Une partie de la face abaxiale de feuille transgénique porteurs de la construction pPR1-YFP-NLS a été infiltrée avec Pst_a2 (108 UFC/mL). Les images ont été acquises à intervalles de 3 min pour un total de 40 h. L’horodatage indiqué est dans le format dd:hh:mm:ss.sss. Ce film a été modifié d’une précédente étude9. S’il vous plaît cliquez ici pour voir cette vidéo. (Clic droit pour télécharger.)
Discussion
Nous rapportons ici une méthode simple pour monter une vie Arabidopsis leaf exprimant un gène rapporteur fluorescent sous le contrôle d’un promoteur d’intérêt pour l’observation à long terme à l’aide d’un stéréomicroscope fluorescent automatisé. Time-lapse imagerie d’un reporter fluorescent a été fréquemment effectuée dans les tissus de la racine ; Cependant, seules quelques études similaires ont été menées dans les tissus foliaires. C’est probablement parce que les feuilles sont capables de se déplacer librement dans l’espace, tandis que les racines sont souvent enterrés et fixe solide gélosé.
Dans ce rapport, nous nous sommes concentrés sur la dynamique spatio-temporelle de l’activité pPR1 au cours de l’ETI induite par le Pst_a2. Outre la fixation douce de la feuille détaillée ci-dessus, il est important de visualiser clairement la dynamique spatio-temporelle des événements cellulaires tels que l’activation du promoteur et PCD. Si la distinction entre les cellules présentant l’activité pPR1 et PCD n’est pas forte, s’assurer que tous les espaces intercellulaires dans la région infiltrée sont complètement remplis avec la suspension de l’agent pathogène (voir étape 2,4). Ceci est crucial lorsque vous utilisez stéréomicroscopes grand-angulaire fluorescence puisque ces microscopes capturent tous les signaux détectables sur la même position verticale de l’échantillon. Chlorophylle autofluorescence des cellules survivantes au-dessus ou sous les cellules dans le domaine PCD masque facilement les cellules mortes ne présentant aucune autofluorescence. Cela vaut également pour le signal de la YFP.
Conditions pour l’imagerie time-lapse doivent être réalisés avec soin par le biais de plusieurs expériences préliminaires dans des conditions expérimentales différentes. Paramètres pour l’imagerie time-lapse dépendent de plusieurs facteurs tels que le système microscopique, des plantes transgéniques et pathogènes. Pour obtenir ces paramètres, nous avons tout d’abord analysé différentes durées d’exposition pour l’intensité du signal dans la feuille infiltrée à 7 hpi, qui coïncide presque avec l’activation initiale de pPR1YFP. Une exposition s 5 a été déterminée en fonction de capter le signal YFP avec le stéréomicroscope utilisé dans cette étude. Un test similaire a été réalisé pour l’imagerie chlorophylle autofluorescence. Exposition de l’échantillon à la lumière entre les intervalles de 3 min a été programmée dans le time-lapse programme d’imagerie comme l’imagerie, un champ lumineux normal avec durée maximale d’exposition. Notre système (Table des matières) nous a permis d’avoir 2,5 min en plus de la YFP, TXR et l’imagerie du champ lumineux. Cette contrainte a été la principale raison de choisir un intervalle de 3 min. Ensuite, nous avons confirmé que cette condition de time-lapse ne causé aucun dommage apparent aux échantillons de plantes et n’induit pas d’activation de liées au stress léger ectopique d' pPR1 (Figure 3B, C). Cela a conduit à l’élaboration du programme utilisé dans cette étude. Ainsi, des intervalles de 3 min de l’imagerie de fluorescence ont été jugés suffisants pour capturer pPR1 dynamique au cours de la Pst_a2-mediated ETI9.
Constructions de promoteur-journaliste, particulièrement avec le journaliste fluorescent fusionné à la NLS, ont été utilisées par de nombreux groupes et sont facilement accessibles depuis le milieu de la recherche ; Nous avons utilisé la construction publiée par Kubo et al. 13. par conséquent, le protocole décrit ici peut être utilisé dans toute étude de la biologie végétale examinant les tissus foliaires, si les plantes transgéniques appropriées sont disponibles. Notre protocole simple et facile fournit une excellente occasion pour les chercheurs désireux d’analyser la dynamique spatio-temporelle de n’importe quel évènement biologique qui se produisent dans les feuilles, comme la réponse immunitaire. Il est plausible que notre méthode en utilisant des morceaux de ruban induit un stress physique légère sur les spécimens. Toutefois, cette question peut être contrôlée en incluant des contrôles positifs et négatifs appropriés, tels que des simulacres soins, dans les expériences (Figure 3C). Les conditions expérimentales peuvent être modifiées et optimisées en analysant ces contrôles dans des conditions différentes.
Ces dernières années, le développement rapide des instruments et des techniques d’imagerie a stimulé l’intérêt des chercheurs pour les aspects spatio-temporels complexes d’événements biologiques. Dans toute analyse d’imagerie, approprié de montage et de fixation des spécimens sont parmi les questions les plus importantes. La méthode simple et souple de montage vivre Qu'arabidopsis laisse développée dans cette étude peut être appliquée et optimisée pour diverses expériences d’imagerie.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
Ce travail a été soutenu par la Japan Science and Technology Agency [PRESTO117665 à S.B., ERATOJPMJER1502 à n. n.] et par la société japonaise pour la Promotion des sciences (subventions pour le démarrage de l’activité de recherche [22880008 à S.B.] et pour les jeunes scientifiques (B) [ 23780040 à S.B.]). Nous remercions Senzaki A., Y. Suzuki, Y. Sugisawa et E. Betsuyaku pour assistance technique excellente, J. Parker pour prévoyant la fourniture le vecteur pBGYN de la souche Pst_a2 et T. Denys.
matériels
| Name | Company | Catalog Number | Comments |
| Bacto Agar | BD biosciences | 214010 | |
| Bacto Protease Peptone No. 3 | BD biosciences | 211693 | |
| Bacto Yeast Extract | BD biosciences | 212750 | |
| BM2 soil | Berger | ||
| Glycerol | nacalai tesque | 17017-35 | |
| Kanamycin sulfate | Wako | 113-00343 | |
| Leica M205FA with DFC365FX, LED_MCI and Leica EL6000 (Las X software-regulated) | Leica Microsystems | Yellow fluorescent protein (YFP) and Texas Red (TXR) filters installed | |
| Magnesium Chloride Hexahydrate | Wako | 135-00165 | |
| Micro Slide Glass (Size: 76 mm × 26 mm, Thickness: 1.0–1.2 mm) | MATSUNAMI | S1112 | |
| Micropore Surgical Tape (1.25 cm × 9 m) | 3M | 1530-0 | |
| Needleless 1 ml plastic syringe | Terumo | SS-01T | |
| Plastic cell plug tray (cell size: 44 mm × 44mm × 44 mm) | Tanaka sangyo, Japan | htray-bk72 | Any tray with similar sized cells can be used |
| Rifampicin | nacalai tesque | 30259-81 | |
| Plastic Tape (0.2 mm thick × 19 mm wide × 10 m long) | Yamato | No0200-19-1 | Any plastic/vinyl tape with similar thickness can be used |
Références
- Tsuda, K., Somssich, I. E. Transcriptional networks in plant immunity. New Phytologist. 206 (3), 932-947 (2015).
- Mine, A., Sato, M., Tsuda, K. Toward a systems understanding of plant-microbe interactions. Frontiers in Plant Science. 5, (2014).
- Murray, S. L., Thomson, C., Chini, A., Read, N. D., Loake, G. J. Characterization of a novel, defense-related Arabidopsis mutant, cir1, isolated by luciferase imaging. Molecular Plant-Microbe Interactions. 15 (6), 557-566 (2002).
- Spoel, S. H., Johnson, J. S., Dong, X. Regulation of tradeoffs between plant defenses against pathogens with different lifestyles. Proceedings of the National Academy of Sciences of the United States of America. 104 (47), 18842-18847 (2007).
- Asai, S., Shirasu, K. Plant cells under siege: plant immune system versus pathogen effectors. Current Opinion in Plant Biology. 28, 1-8 (2015).
- Schmelzer, E., Kruger-Lebus, S., Hahlbrock, K. Temporal and Spatial Patterns of Gene Expression around Sites of Attempted Fungal Infection in Parsley Leaves. The Plant Cell. 1 (10), 993-1001 (1989).
- Ohshima, M., Itoh, H., Matsuoka, M., Murakami, T., Ohashi, Y. Analysis of stress-induced or salicylic acid-induced expression of the pathogenesis-related 1a protein gene in transgenic tobacco. The Plant Cell. 2 (2), 95-106 (1990).
- Rushton, P. J., Reinstädler, A., Lipka, V., Lippok, B., Somssich, I. E. Synthetic plant promoters containing defined regulatory elements provide novel insights into pathogen- and wound-induced signaling. The Plant Cell. 14 (4), 749-762 (2002).
- Betsuyaku, S., et al. Salicylic Acid and Jasmonic Acid Pathways are Activated in Spatially Different Domains Around the Infection Site During Effector-Triggered Immunity in Arabidopsis thaliana. Plant & Cell Physiology. 59 (1), 8-16 (2018).
- Dodds, P. N., Rathjen, J. P. Plant immunity: towards an integrated view of plant-pathogen interactions. Nature Reviews Genetics. 11 (8), 539-548 (2010).
- Katagiri, F., Thilmony, R. The Arabidopsis thaliana-Pseudomonas syringae interaction. Arabidopsis Book. , (2002).
- Zeier, J., Pink, B., Mueller, M. J., Berger, S. Light conditions influence specific defence responses in incompatible plant-pathogen interactions: uncoupling systemic resistance from salicylic acid and PR-1 accumulation. Planta. 219 (4), 673-683 (2004).
- Kubo, M., et al. Transcription switches for protoxylem and metaxylem vessel formation. Genes & Development. 19 (16), 1855-1860 (2005).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.