
Method Article
Microscopie 4D: Démêler le développement embryonnaire de Caenorhabditis elegans à l’aide de la microscopie Nomarski
Dans cet article
Résumé
Nous présentons ici un protocole pour la préparation et le montage des embryons de Caenorhabditis elegans , l’enregistrement du développement sous un microscope 4D et le traçage de la lignée cellulaire.
Résumé
La microscopie 4D est un outil inestimable pour démêler le processus de développement embryonnaire chez différents animaux. Au cours des dernières décennies, Caenorhabditis elegans est devenu l’un des meilleurs modèles pour étudier le développement. D’un point de vue optique, sa taille et son corps transparent font de ce nématode un spécimen idéal pour la microscopie DIC (Differential Interference Contrast ou Nomarski). Cet article illustre un protocole pour la culture des nématodes C. elegans , la préparation et le montage de leurs embryons, la microscopie 4D et le traçage de la lignée cellulaire. La méthode est basée sur des enregistrements time-lapse multifocaux d’images Nomarski et d’analyses avec un logiciel spécifique. Cette technique révèle la dynamique du développement embryonnaire au niveau cellulaire. Tout défaut embryonnaire chez les mutants, tel que des problèmes d’orientation du fuseau, de migration cellulaire, d’apoptose ou de spécification du devenir cellulaire, peut être efficacement détecté et noté. Pratiquement chaque cellule de l’embryon peut être suivie jusqu’au moment où l’embryon commence à bouger. Tracer la lignée cellulaire complète d’un embryon de C. elegans par microscopie DIC 4D est laborieux, mais l’utilisation d’un logiciel spécifique facilite grandement cette tâche. De plus, cette technique est facile à mettre en œuvre en laboratoire. La microscopie 4D est un outil polyvalent et ouvre la possibilité d’effectuer une analyse inégalée du développement embryonnaire.
Introduction
La microscopie 4D est un système d’enregistrement time-lapse multifocal qui permet aux chercheurs d’enregistrer et de quantifier la dynamique cellulaire d’un échantillon biologique à la fois spatialement et au fil du temps. Les cultures cellulaires, les levures ou les tissus vivants peuvent être soumis à une analyse 4D mais cette technique est particulièrement adaptée à l’analyse du développement d’embryons vivants. La résolution de cette analyse atteint le niveau de chaque cellule de l’embryon. Chaque division cellulaire peut être détectée et les mouvements cellulaires peuvent être tracés au fil du temps. Les destins cellulaires sont évalués en fonction de la position et de la forme que les cellules acquièrent. L’utilisation de l’optique Nomarski améliore le contraste des échantillons transparents non colorés en utilisant des faisceaux lumineux polarisés orthogonalement qui interfèrent au niveau du plan focal. Les images résultantes apparaissent en trois dimensions, éclairées d’un côté.
D’autres méthodes basées sur l’utilisation de la microscopie confocale et des animaux transgéniques GFP pour la détection automatique des noyaux et la génération de lignées cellulaires ont été développées 1,2. L’avantage de ces systèmes est évident : le logiciel l’emporte largement sur la nécessité de marquer manuellement chaque noyau sur une période de temps (bien qu’une certaine supervision manuelle soit nécessaire pendant les dernières étapes). Cependant, les processus cellulaires impliquant des changements dans la forme cellulaire ou la dynamique de la membrane, tels que ceux qui se produisent lors de la différenciation cellulaire, de la migration, de l’apoptose ou de l’engloutissement des cadavres, restent cachés sous forme de fond noir dans les images de noyaux marqués par fluorescence.
En revanche, la microscopie Nomarski 4D (également appelée microscopie DIC, microscopie à contraste d’interférence différentielle) montre à la fois les changements de forme des noyaux et des cellules qui se produisent au cours du développement d’animaux de type sauvage ou mutants. Cela permet de tracer la lignée cellulaire à l’aide de microscopes standard, en utilisant uniquement la lumière transmise. Il n’est généralement pas nécessaire d’utiliser des animaux transgéniques, sauf pour montrer des modèles d’expression spécifiques, auquel cas les scans fluorescents peuvent être intercalés. Par conséquent, cela pourrait être l’approche optimale pour de nombreux laboratoires travaillant sur des processus cellulaires dynamiques tels que l’embryogenèse ou l’apoptose qui peuvent être mis en évidence sous microscopie DIC 3,4,5,6,7.
Plusieurs programmes flexibles et conviviaux sont disponibles pour capturer des images microscopiques et reconstruire des lignées cellulaires, des modèles 3D, des chemins de migration cellulaire, etc. dans l’échantillon enregistré. Dans une expérience standard, les images sont acquises dans une série de plans focaux, à une distance constante, dont le nombre dépend de l’épaisseur de l’échantillon. La résolution temporelle de l’analyse peut être optimisée en augmentant la fréquence de balayage. Il n’y a pratiquement aucune limite pour la durée de l’enregistrement autre que la capacité de stockage de l’ordinateur. Par exemple, pour une analyse du développement embryonnaire de C. elegans , nous acquérons régulièrement des images sur 30 plans focaux (1 micron-pas chacun), toutes les 30 secondes pendant 12 heures.
Ces systèmes ont été appliqués à l’analyse de plusieurs embryons animaux tels que Caenorhabditis elegans 8,9,10, Drosophila melanogaster11, d’autres embryons de nématodes12,13, tardigrades14,15 et même des embryons de souris précoces16. La seule exigence est d’avoir un embryon transparent capable de se développer sur la préparation de la lame au microscope.
En résumé, la microscopie 4D basée sur DIC est particulièrement utile pour 1) analyser le développement embryonnaire de petits animaux transparents: traçage de la lignée cellulaire, chemins de migration cellulaire, génération de modèles 3D, etc.; 2) définir les modèles d’expression des gènes; 3) étudier la dynamique de la culture cellulaire, de la levure aux cellules humaines; 4) l’analyse de la dynamique tissulaire ou des fragments d’embryons; 5) quantifier la cinétique de mort cellulaire et l’engloutissement des cadavres; et 6) effectuer une analyse phylogénie comparative basée sur les caractéristiques du développement embryonnaire. S’il y a un intérêt pour l’un de ces sujets (ou des sujets similaires), la microscopie 4D peut être utilisée.
Protocole
1. Cultivez C. elegans sur des boîtes de Pétri
- Préparez les assiettes de NGM et ensemencez-les avec E. coli OP50 comme source de nourriture (Figure 1). Cultiver et entretenir C. elegans comme décrit17. Conserver les assiettes ensemencées à 4 °C jusqu’à un mois.
- Ajustez les plaques à la température souhaitée avant d’ajouter les vers.
- Pour transférer les vers, retirez un morceau de gélose d’une vieille assiette et placez-le sur une assiette fraîche.
- Alternativement, capturez des animaux célibataires avec un cueilleur de vers stérile (un morceau de 1 pouce de fil de platine de calibre 32 avec une pointe aplatie, montée sur la pointe d’une pipette Pasteur) et placez-les sur la nouvelle plaque.
- Cultivez les vers à la température souhaitée.
- Cultivez les vers C. elegans à 20 °C, la température standard. Cependant, pour analyser le développement de mutants thermosensibles, effectuez une incubation de nuit, généralement à 25 °C. Ajustez la durée et la température de cette incubation au besoin, en fonction du mutant spécifique.
2. Préparer l’enregistrement de la microscopie 4D avant de monter les embryons (Figure 2)
- Installez le microscope et les contrôles de température avant de préparer l’embryon. Les embryons de C. elegans se divisent très rapidement. Préparez-vous à commencer l’enregistrement immédiatement après le montage des embryons.
- Réglez la température d’enregistrement à 15 °C, 20 °C ou 25 °C.
- Enregistrez régulièrement les embryons à 25 °C. Enregistrez les mutants thermosensibles à la température restrictive pour montrer leurs phénotypes. Enregistrez un contrôle WT (s’il est effectué dans une préparation différente) à la même température que les mutants.
REMARQUE: Les embryons WT se développent plus rapidement à 25 ° C et plus lentement à 15 ° C sans différences supplémentaires dans la lignée cellulaire. Placez les microscopes dans une pièce à température contrôlée. Un contrôle supplémentaire par refroidissement ou chauffage de la glissière est hautement souhaitable. Ceci peut être réalisé en faisant circuler de l’eau à une température spécifique à travers un anneau métallique autour de l’objectif du microscope et du condensateur. L’objectif et la préparation sont en contact direct grâce à l’huile d’immersion et le transfert de température est efficace. Ce système permet un contrôle précis de la température d’enregistrement et des changements de température pendant le développement de l’embryon.
- Enregistrez régulièrement les embryons à 25 °C. Enregistrez les mutants thermosensibles à la température restrictive pour montrer leurs phénotypes. Enregistrez un contrôle WT (s’il est effectué dans une préparation différente) à la même température que les mutants.
- Définissez les paramètres d’enregistrement dans le logiciel de microscopie.
- Pour un enregistrement C. elegans standard (sans balayage fluorescent), sélectionnez :
z-piles de 30 plans focaux, à une distance de 1 micron chacun.
Intervalles de 30 secondes entre le début de chaque pile z.
1500 piles z (12,5 heures d’enregistrement).
REMARQUE: Les programmes de contrôle de microscope commerciaux et open source peuvent être utilisés pour définir ce flux de travail pour la capture d’images. Maintenant, le microscope est prêt à enregistrer.
- Pour un enregistrement C. elegans standard (sans balayage fluorescent), sélectionnez :
3. Préparer et monter les embryons
- Préparez un tampon de gélose mince et homogène comme première étape pour obtenir une belle image (Figure 3).
- Préparer 50 ml d’une solution de gélose à 4,5% dans de l’eau désionisée. Chauffer à ébullition au micro-ondes et verser 0,5 à 1 ml dans des tubes en verre de 3 ml.
- Scellez soigneusement les tubes avec un film de cire pour éviter la dessiccation. Les tubes de gélose scellés peuvent être conservés à température ambiante jusqu’à deux mois.
- Sur le banc de laboratoire, avoir un bloc chauffant à 80 °C avec :
un tube à essai de vaseline pure (fondue), avec un pinceau fin à l’intérieur.
un tube à essai avec de l’eau distillée contenant une pipette Pasteur.
REMARQUE: Cela garantit que tous les matériaux requis seront chauds et que la gélose ne se solidifiera pas lors du processus de fabrication du tampon. - Retirez le film de cire du haut de l’un des tubes d’agar et chauffez-le soigneusement sur un brûleur à alcool pour faire fondre la gélose. Faites preuve de prudence car la gélose chaude expulsée du tube de verre pourrait causer des brûlures.
- Une fois la gélose fondue, placez le tube dans le bloc thermique pour maintenir la gélose sous forme liquide.
- Alternativement, placez les tubes de gélose dans le bloc chauffant 1h avant l’expérience pour les faire fondre sans utiliser de brûleur. La gélose fondue doit être jetée après un jour.
- Placez une lame de microscope (diapositive A) entre deux autres sur un morceau de plastique.
- Prenez une autre diapositive (diapositive B) et tenez-la avec vos doigts dans une main.
- D’autre part, placez une petite goutte de gélose fondue au centre de la diapositive A à l’aide de la pipette Pasteur chaude.
- Appuyez immédiatement sur la glissière B sur la goutte d’agar pour créer un tampon très fin entre les diapositives A et B. Gardez ces diapositives prises en sandwich ensemble jusqu’à l’étape 3.2.3.
- Montez les embryons.
- Collectez 5 à 10 hermaphrodites gravides avec le cueilleur et placez-les dans un verre horloger rempli d’eau.
- Utilisez un scalpel pour ouvrir les nématodes hermaphrodites et extraire les ovules précoces (1-4 cellules) de l’utérus, sous le stéréomicroscope.
- Retirez la diapositive de l’étape 3.1.10 et glissez doucement la diapositive B pour exposer le coussinet de gélose sur la diapositive A.
- Placez un œuf précoce au centre du tampon de gélose en pipetant avec un tube capillaire.
- Alternativement, pipettez une goutte contenant un ensemble d’embryons sur le tampon de gélose, puis recherchez des embryons à un stade précoce. Effectuez cette étape sous le stéréomicroscope.
- Si nécessaire, déplacez l’œuf en le poussant avec un cil collé à l’extrémité d’un cure-dent.
- Enlevez l’excès d’eau avec la pipette capillaire.
REMARQUE: Une pipette capillaire peut facilement être préparée en chauffant une pipette Pasteur sur un brûleur à alcool et en la tirant des deux extrémités. - Couvrez soigneusement la préparation avec un couvercle. Pour éviter les bulles d’air, placez un bord du couvercle sur la glissière et faites glisser doucement un scalpel le long du bord adjacent pour draper lentement et obliquement le couvercle sur la préparation.
- Utilisez une pipette pour remplir les 3/4 de l’espace entourant le tampon de gélose avec de l’eau. Laissez 1/4 de l’espace avec de l’air.
- Scellez le couvercle avec de la vaseline pour éviter la dessiccation pendant de longues périodes d’enregistrement.
- Utilisez la brosse fine pour étendre une fine couche de vaseline fondue autour du bord de la lèvre de couverture.
REMARQUE: Maintenant, la préparation est prête pour l’enregistrement.
4. Ajustez le DIC et démarrez l’enregistrement de la microscopie 4D
- Placez la lame sur la scène du microscope. Focalisez l’embryon à l’aide de l’objectif de faible grossissement (5x ou 10x).
- Passez à l’objectif d’immersion 100x.
- Ajustez les composants optiques du microscope pour obtenir une image Nomarski.
- Concentrez le condenseur.
- Ouvrez complètement l’ouverture du condenseur et fermez le diaphragme de champ (cela fournira une ouverture numérique plus élevée et donc une plus grande résolution).
- Vérifiez que les deux polariseurs du microscope sont orientés pour provoquer l’extinction maximale de la lumière.
- Tournez le prisme de Wollaston pour obtenir une belle image tridimensionnelle de l’embryon, illuminée d’un côté. Tournez le prisme dans l’autre sens pour obtenir l’effet d’avoir l’embryon illuminé de l’autre côté.
REMARQUE: Ces étapes peuvent être effectuées sur un échantillon d’essai avant l’enregistrement, de sorte que seul un réglage fin est nécessaire sur l’échantillon analysé.
- Lancez la capture d’image dans le microscope.
5. Analysez le film 4D (Figure 4).
REMARQUE: Une fois l’enregistrement terminé, utilisez un logiciel de traçage de lignée cellulaire pour reconstruire et analyser la lignée cellulaire.
Le logiciel de traçage de lignée cellulaire est un outil puissant pour effectuer des analyses détaillées du développement embryonnaire ou de la dynamique dans des cultures cellulaires ou des fragments de tissus. Le programme extrait et quantifie plusieurs ensembles de données sur la dynamique cellulaire de l’échantillon qui incluent la génération de la lignée cellulaire complète de chaque cellule enregistrée, y compris les divisions cellulaires, la longueur du cycle cellulaire, la migration ou l’apoptose ainsi que sa cinétique. En outre, la différenciation cellulaire peut être notée par les changements morphologiques de la cellule ou par l’expression de marqueurs spécifiques. Fondamentalement, l’écran du logiciel affiche deux fenêtres: dans la fenêtre de gauche, le film 4D peut être lu en avant et en arrière ou de haut en bas jusqu’aux niveaux supérieur ou inférieur afin que chaque cellule puisse être suivie dans le temps et l’espace tout au long de l’enregistrement. Sur la veuve de droite, la lignée cellulaire est générée. Cliquer sur un noyau de cellule dans le film 4D génère un point dans la fenêtre de lignée qui stocke les informations du nom de la cellule, du destin et des coordonnées spatiales. La lignée cellulaire d’une cellule spécifique est générée en lisant le film 4D vers l’avant et en cliquant périodiquement sur le noyau pour marquer la mitose de cette cellule spécifique au fil du temps. La répétition de ce processus pour chacune des cellules enregistrées génère la lignée cellulaire complète de l’embryon ou de l’échantillon. Les informations stockées pour les coordonnées spatiales de chaque cellule sont ensuite utilisées pour reconstruire des modèles embryonnaires 3D et des voies de migration cellulaire.
- Ouvrez le logiciel de suivi de lignée et créez un nouveau projet en accédant au menu de la barre supérieure et en sélectionnant :
| de fichiers Nouveau projet. - Sélectionnez le modèle de lignée cellulaire en fonction de la température d’enregistrement : DB08 pour l’enregistrement à 25 °C, DB10 pour l’enregistrement à 20 °C et DB12 pour l’enregistrement à 15 °C.
- Définissez les paramètres d’enregistrement dans la fenêtre émergente : nombre de scans (généralement 1500), temps entre les scans (30 secondes), nombre de niveaux (30) et distance entre les niveaux (1 micron).
- Sélectionnez le fichier image et le format.
- Sélectionnez le répertoire d’images dans lequel les images ont été enregistrées.
- Choisissez si les images doivent être enregistrées en tant qu’images uniques (une image par niveau et par heure) ou en tant que z-stacks multi-images.
- Déterminez le nom du fichier et le format d’image. Régulièrement, les images uniques sont enregistrées sous les noms suivants :
X0000L00C1 (pour l’analyse 0, niveau 0, canal 1)
X0000L01C1 (pour l’analyse 0, niveau 1, canal 1)
X0000L02C1 (pour l’analyse 0, niveau 2, canal 1)
...
X0001L00C1 (pour l’analyse 1, niveau 0, canal 1)
X0001L01C1 (pour l’analyse 1, niveau 1, canal 1)
...
X0300L04C1 (pour numérisation 300, niveau 4, canal 1)
...
REMARQUE: L’enregistrement des images dans un format compressé permet d’économiser de l’espace sur votre disque dur.
- Définissez les canaux lumineux : 1 pour l’optique DIC, 2 pour GFP, 3 pour l’appel d’offres, etc. Ajoutez ceux qui ont été utilisés dans l’enregistrement 4D. Cliquez sur « traitement des canaux activé » pour les détecter.
- Commencez à tracer la lignée cellulaire de l’embryon. L’écran contient maintenant deux fenêtres principales: la fenêtre vidéo et la fenêtre de lignée cellulaire.
- Dans la fenêtre de lignée, sélectionnez une branche de lignée et utilisez la souris pour cliquer sur le noyau de cellule correspondant à cette cellule dans la fenêtre vidéo.
- Suivez la cellule spatialement et au fil du temps en lisant le film 4D vers l’avant, vers l’arrière ou vers le haut ou vers le bas d’un niveau, à l’aide des touches du curseur.
- Cliquez périodiquement sur le noyau cellulaire. Cela génère un point dans la branche de la lignée et enregistre les coordonnées spatiales de la cellule à ce moment-là. En conséquence, la lignée cellulaire progresse et des reconstructions 3D de l’embryon sont possibles.
- Marquez la mitose en cliquant sur la touche de retour. Sélectionnez ensuite l’une des cellules filles et suivez-la comme précédemment.
- Répétez le processus (étapes 5.6.1 à 5.6.4) pour le reste des cellules embryonnaires afin de retracer la lignée cellulaire complète ou de suivre des cellules spécifiques d’intérêt telles que celles qui subissent l’apoptose.
- Comparez la lignée mutante avec la lignée cellulaire stéréotypée WT C. elegans .
Résultats
Pour caractériser le développement embryonnaire d’un mutant de C. elegans pour le gène gsr-1, qui code pour l’enzyme glutathion réductase, nécessaire à la régénération du glutathion réduit (GSH) et impliqué dans le maintien de l’homéostasie redox chez le nématode, nous avons effectué une microscopie 4D d’un mutant de délétion gsr-1 (tm3574) qui est un allèle de perte de fonction provoquant un phénotype d’arrêt embryonnaire précoce18. Les nématodes mutants WT et équilibrés gsr-1 (tm3574) C. elegans ont été cultivés sur des plaques de NGM ensemencées avec E. coli OP50 comme source de nourriture17. Les vers gsr-1 (tm3574) ont été cultivés comme hétérozygotes à 20 °C pendant deux générations, puis les vers homozygotes ségrégés (qui sont capables de grandir jusqu’à l’âge adulte grâce à la charge maternelle) ont été déplacés à 25 °C pour une incubation de nuit avant l’analyse embryonnaire. Les plaques à vis sans fin ont été incubées dans des boîtes en carton pour éviter la condensation (Figure 1). Les nématodes gravides ont été ouverts pour extraire de jeunes embryons.
Pour comparer le développement embryonnaire du mutant par rapport au WT stéréotypé dans des conditions identiques, un WT (comme témoin) et un embryon gsr-1 (tm3574) ont été placés sur la même préparation l’un à côté de l’autre. Le flux de travail de microscopie 4D a été exécuté sur un microscope vertical motorisé standard équipé d’une optique DIC. Les paramètres d’enregistrement sélectionnés sur le programme de contrôle du microscope étaient: des piles z de 30 plans focaux à une distance de 1 micron chacun, des intervalles de 30 secondes entre le début de chaque pile z et 1500 piles z (12,5 heures d’enregistrement). La température d’enregistrement a été ajustée à 25 °C (à la fois dans la pièce et sur la scène du microscope) (Figure 2).
Une fois l’enregistrement terminé, le fichier d’images a été ouvert et la lignée cellulaire a été reconstruite à l’aide d’un logiciel de traçage de lignée en cliquant sur les noyaux cellulaires affichés dans la fenêtre vidéo (Figure 4). La lignée cellulaire embryonnaire mutante gsr-1 (tm3574) tracée a été comparée à la lignée C. elegans WT représentée à l’arrière-plan. Un résultat majeur a été la détection d’un retard progressif du cycle cellulaire au cours du développement embryonnaire. En conséquence, les embryons mutants se sont arrêtés à des stades intermédiaires tandis que les embryons WT ont progressé et ont finalement éclos sous forme de larves.
La préparation et l’observation directe d’embryons au microscope ou l’immunocoloration avec des anticorps contre des marqueurs embryonnaires tardifs pourraient révéler la présence d’un pourcentage élevé d’embryons jeunes dans le mutant par rapport au WT. L’arrêt d’embryons pourrait alors être déduit comme l’explication la plus plausible. Cependant, la preuve directe et la quantification exacte du retard du cycle cellulaire ne peuvent être montrées et quantifiées que de manière élégante et facile par le biais d’une expérience de microscopie 4D. D’autres caractéristiques importantes du développement embryonnaire telles que la différenciation cellulaire ou l’apoptose (Figure 5) peuvent également être visualisées de manière dynamique à l’aide de la microscopie 4D qui offre une analyse détaillée de plusieurs aspects du développement dans une seule expérience.

Figure 1 : Nématodes de C. elegans se développant dans des conditions de laboratoire. Les nématodes sont cultivés sur des plaques de NGM ensemencées par E. coli, stockées dans des boîtes en carton et incubées à 15 °C, 20 °C ou 25 °C. Veuillez cliquer ici pour voir une version plus grande de cette figure.
{kind=link}

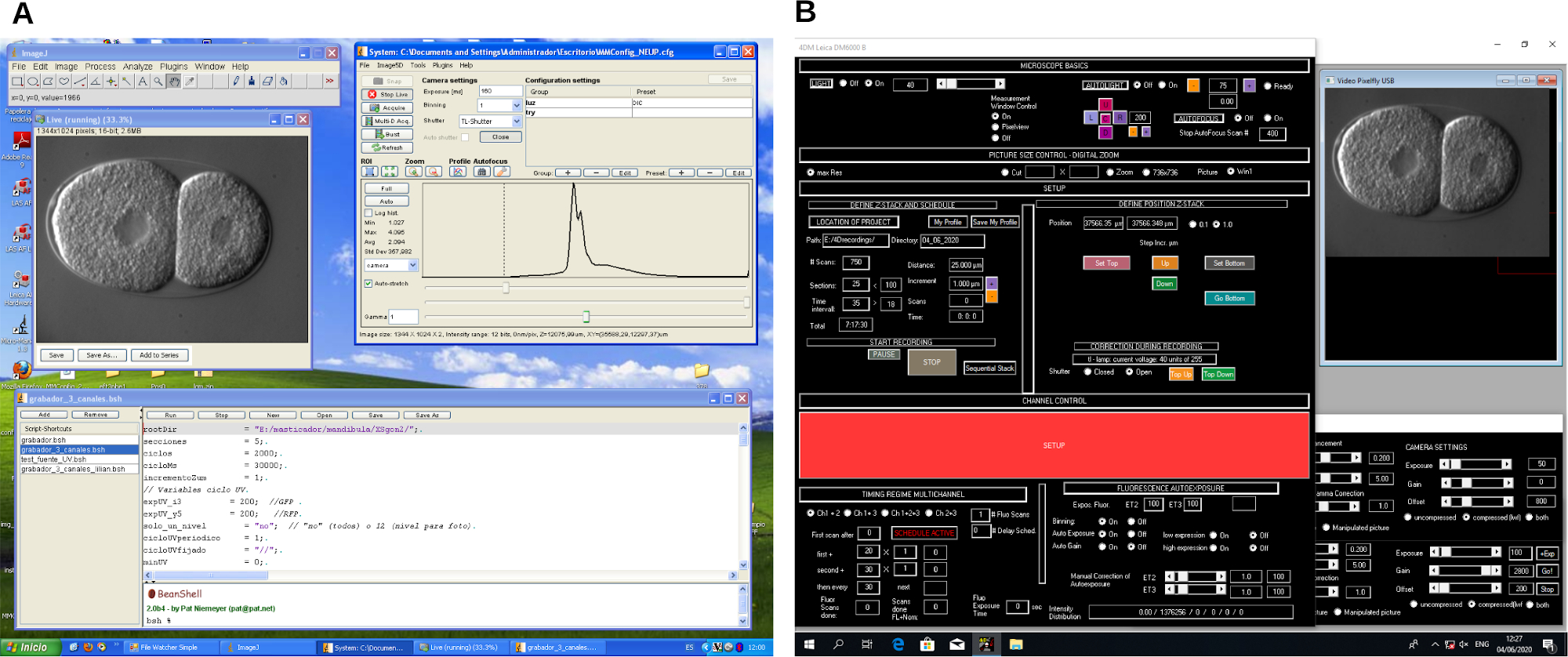
Figure 2 : Capture d’écran du logiciel d’enregistrement de microscopie 4D. Exemple de deux logiciels de contrôle de microscope différents (A et B). Ces programmes créent des flux de travail pour contrôler le microscope et la capture d’images pendant l’enregistrement de la microscopie 4D. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 3 : Photographies en série de la préparation et du montage de l’embryon de C. elegans . A. Tubes de gélose préparés. B-C. Préparation du tampon de gélose. D. Toboggan partiellement rempli d’eau. E. Sceller la lame avec de la vaseline. F. Préparation finale. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

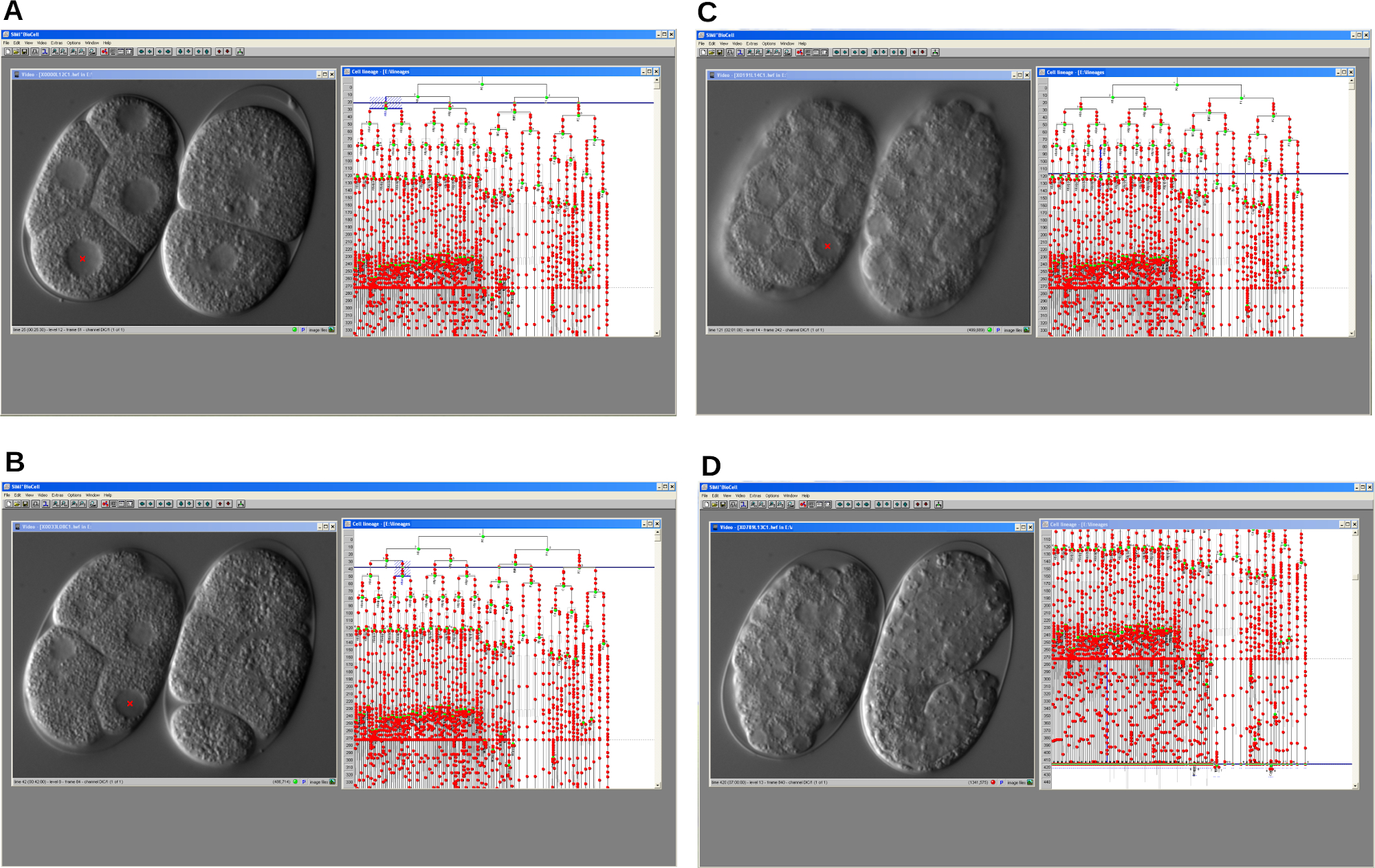
Figure 4 : Captures d’écran série du logiciel de traçage de lignée cellulaire. Le programme permet la reconstruction de la lignée cellulaire embryonnaire d’un mutant retardateur du cycle cellulaire (à gauche) et d’un embryon WT (à droite) de C. elegans . Un. Une première étape du développement. B-C. Le développement des deux embryons progresse au fil du temps. D. L’embryon WT se développe correctement et commence à s’allonger pendant que le mutant s’arrête. Dans tous les cas, le programme affiche la fenêtre vidéo et la fenêtre de lignage. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

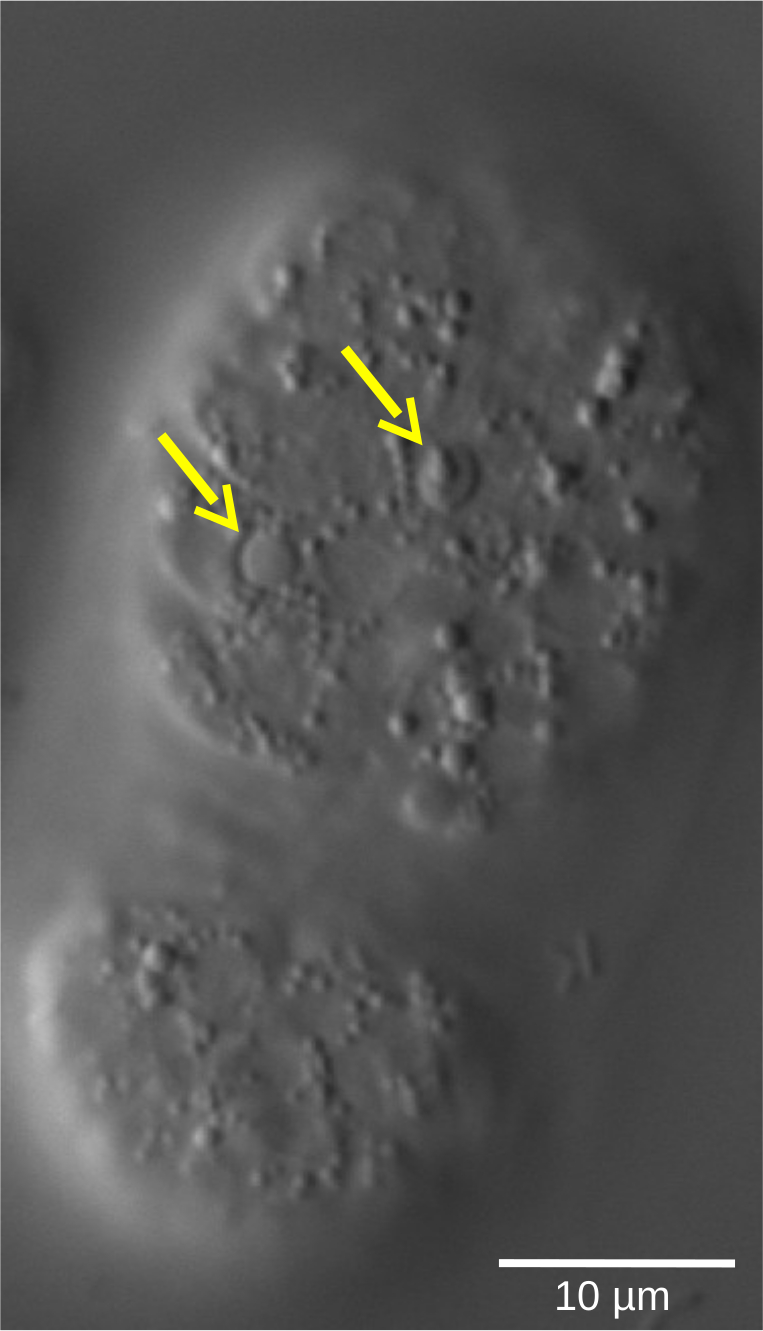
Figure 5 : Forme réfractaire des lentilles des cellules apoptotiques dans un embryon de C. elegans WT. Le devenir cellulaire, défini par les caractéristiques morphologiques, peut être évalué par microscopie 4D. L’image montre un embryon de C. elegans au stade du haricot. Les cellules vivantes montrent des noyaux de forme lisse entourés d’un cytoplasme granulaire. En revanche, les cellules apoptotiques (flèches jaunes) se condensent et adoptent une forme réfractaire semblable à celle d’une lentille. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Discussion
L’un des défis majeurs de la biologie moderne est de comprendre le développement des organismes multicellulaires. C. elegans est devenu l’un des modèles les mieux adaptés pour étudier la coordination fine entre la prolifération cellulaire et la différenciation cellulaire dans l’embryon en développement. D’un point de vue optique, son corps transparent et sa petite taille font de ce nématode un spécimen idéal pour la microscopie DIC. D’autres organismes présentant des caractéristiques similaires ont également été soumis à une analyse microscopique 4D 11,12,13,14,15,16.
Pour ces études développementales, l’inactivation des gènes par la génétique avant ou arrière fournit un indice de son implication dans l’embryogenèse. Une fois qu’il a été prouvé qu’un gène joue un rôle dans le développement, l’étape suivante consiste à définir son rôle exact dans l’établissement du plan corporel correct. L’immunocoloration est l’approche choisie pour la plupart des modèles. Cette technique élucide les problèmes de différenciation cellulaire ou d’expression de marqueurs spécifiques. Cependant, une limitation majeure de cette approche est qu’elle ne fournit qu’une vue statique de l’expression d’un ou plusieurs marqueurs à un point fixe du développement. Une vue dynamique de ces marqueurs tout au long du développement ne peut être obtenue qu’en colorant différents embryons à différents moments. En outre, la reconstruction de la lignée cellulaire n’est pas possible dans de tels échantillons fixes.
La microscopie 4D est une approche complémentaire pour étudier le développement embryonnaire. Cette technique révèle la dynamique de développement à une résolution au niveau de la cellule. Tout défaut dans l’embryon tel que des problèmes d’orientation du fuseau, de migration cellulaire, d’apoptose, de spécification du destin cellulaire, etc. apparaîtra dans un film 4D qui peut être visualisé en avant et en arrière, quantifié et noté par le chercheur. En utilisant cette technique, pratiquement chaque cellule de l’embryon peut être suivie jusqu’au moment où l’embryon commence à bouger. Les embryons soumis à la microscopie 4D avec seulement la lumière visible et l’optique Nomarski n’encourent pas de photodommages. Les scans fluorescents peuvent également être intercalés dans l’enregistrement pour détecter quand et où un gène est exprimé. Les embryons qui souffrent de photodommages importants sont identifiés par l’extension du cycle cellulaire qui provoque une forte irradiation UV par rapport à un embryon de lignée WT standard. Dans ce cas, les dommages photo peuvent être réduits en abaissant l’intensité de la lampe UV et en augmentant la sensibilité ou le temps d’exposition de l’appareil photo. Les caractéristiques morphologiques et les marqueurs moléculaires peuvent aider à clarifier le développement embryonnaire de tout mutant.
La mise en place d’un système de microscopie 4D est facile à mettre en œuvre en laboratoire et, après un peu de pratique, permet une analyse inégalée de la dynamique cellulaire et du traçage de la lignée des cultures cellulaires et des spécimens transparents vivants à un niveau de résolution de chaque cellule dans le champ du microscope. Le traçage de la lignée cellulaire sur les images DIC est toujours traité à la main. Cela prend du temps et, bien que le logiciel détecte les erreurs de lignée telles que différentes branches de lignée marquant la même cellule, des erreurs sont possibles. Alors que la détection automatique des cellules marquées GFP est bien développée2, un logiciel complémentaire de traçage de lignée basé sur des cellules non marquées et des images en lumière visible en est encore à ses débuts et n’est pas vraiment utile pour une analyse complète de l’embryon. Sans aucun doute, l’application des systèmes de reconnaissance d’images au domaine de la microscopie à lumière visible apportera une grande avancée dans ce domaine.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
Les auteurs souhaitent saluer le soutien de la Fondation Rioja Salud (Fondos FEDER) et du Ministerio de Ciencia, Innovación y Universidades (MCIU) espagnol (Subvention PGC2018-094276-B-I00). Cristina Romero Aranda est financée par une bourse de l’AECC (Asociación Española Contra el Cáncer).
matériels
| Name | Company | Catalog Number | Comments |
| Caenorhabditis elegans (N2) | GCG (Caenorhabditis Genetics Center) | N2 | WT C. elegans strain. Can be requested at GCG (Caenorhabditis Genetics Center): https://cgc.umn.edu/ |
| Caenorhabditis elegans (VZ454) | GCG (Caenorhabditis Genetics Center) | VZ454 | gsr-1(tm3574) C. elegans mutant strain. Can be requested at GCG (Caenorhabditis Genetics Center): https://cgc.umn.edu/ |
| Cell Lineage Tracing software | SIMI | Simi BioCell | This is the software to reconstruct the embryo cell lineage. For a detailed explanation check at: http://www.simi.com/en/products/cell-research/simi-biocell.html |
| Microscope camera | Hamamatsu | Orca-R2 | Miscroscope camera for both transmitted and UV light |
| Microscope control software | Caenotec | Time to Live | This software controls the microscope to perform the 4D image capture. Can be requested at: Caenotec Prof. Ralf Schnabel Kleine Dorfstr. 9 38312 Börßum, Germany, Ph: ++49 151 11653356 r.schnabel(at)tu-bs.de |
| Microscope control software | Micro-manager | Micro-manager | This software controls the microscope to perform the 4D image capture. Can be downloaded at: https://micro-manager.org/ |
| Motorized microscope | Leica | Leica DM6000 | Motorized upright microscope to perform 4D microscopy |
| Standard equipment in a Molecular Biology lab. | |||
| Stereomicroscope | Leica | MZ16FA | Steromicroscope to manipulate nematodes and prepare embryos. |
Références
- Hardin, J. Imaging embryonic morphogenesis in C. elegans. Methods in Cell Biology. 106, 377-412 (2011).
- Mace, D. L., Weisdepp, P., Gevirtzman, L., Boyle, T., Waterston, R. H. A high-fidelity cell lineage tracing method for obtaining systematic spatiotemporal gene expression patterns in Caenorhabditis elegans. G3: Genes, Genomes, Genetics (Bethesda). 3 (5), 851-863 (2013).
- Nieto, C., et al. ccz-1 mediates the digestion of apoptotic corpses in C. elegans. Journal of Cell Science. 123 (12), 2001-2007 (2010).
- Cabello, J., et al. PDR-1/hParkin negatively regulates the phagocytosis of apoptotic cell corpses in Caenorhabditis elegans. Cell Death & Disease. 5, 1120 (2014).
- Pinto, S. M., Almendinger, J., Cabello, J., Hengartner, M. O. Loss of Acetylcholine Signaling Reduces Cell Clearance Deficiencies in Caenorhabditis elegans. PLOS One. 11 (2), 0149274 (2016).
- Sáenz-Narciso, B., Gómez-Orte, E., Zheleva, A., Gastaca, I., Cabello, J. Control of developmental networks by Rac/Rho small GTPases: How cytoskeletal changes during embryogenesis are orchestrated. Bioessays. 38 (12), 1246-1254 (2016).
- Zheleva, A. Reduction of mRNA export unmasks different tissue sensitivities to low mRNA levels during Caenorhabditis elegans development. PLOS Genetics. 15 (9), 1008338 (2019).
- Schnabel, R., Hutter, H., Moerman, D., Schnabel, H. Assessing normal embryogenesis in Caenorhabditis elegans using a 4D microscope: variability of development and regional specification. Developmental Biology. 184 (2), 234-265 (1997).
- Verbrugghe, K. J. C., Chan, R. C. Imaging C. elegans Embryos using an Epifluorescent Microscope and Open Source Software. Journal of Visualized Experiments. (49), e2625 (2011).
- Boyd, L., Hajjar, C., O'Connell, K. Time-lapse Microscopy of Early Embryogenesis in Caenorhabditis elegans. Journal of Visualized Experiments. (54), e2852 (2011).
- Urbach, R., Schnabel, R., Technau, G. M. The pattern of neuroblast formation, mitotic domains and proneural gene expression during early brain development in Drosophila. Development. 130 (16), 3589-3606 (2003).
- Dolinski, C., Borgonie, G., Schnabel, R., Baldwin, J. G. Buccal capsule development as a consideration for phylogenetic analysis of Rhabditida (Nemata). Development Genes and Evolution. 208 (9), 495-503 (1998).
- Houthoofd, W., Jacobsen, K., Mertens, C., Vangestel, S., Coomans, A., Borgonie, G. Embryonic cell lineage of the marine nematode Pellioditis marina. Developmental Biology. 258 (1), 57-69 (2003).
- Hejnol, A., Schnabel, R. The eutardigrade Thulinia stephaniae has an indeterminate development and the potential to regulate early blastomere ablations. Development. 132 (6), 1349-1361 (2005).
- Hejnol, A., Schnabel, R. What a couple of dimensions can do for you: Comparative developmental studies using 4D microscopy-examples from tardigrade development. Integrative and Comparative Biology. 46 (2), 151-161 (2006).
- Bischoff, M., Parfitt, D. E., Zernicka-Goetz, M. Formation of the embryonic-abembryonic axis of the mouse blastocyst: relationships between orientation of early cleavage divisions and pattern of symmetric/asymmetric divisions. Development. 135 (5), 953-962 (2008).
- Mora-Lorca, J. A. Glutathione reductase gsr-1 is an essential gene required for Caenorhabditis elegans early embryonic development. Free Radical Biology and Medicine. 96, 446-461 (2016).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.