
Un abonnement à JoVE est nécessaire pour voir ce contenu. Connectez-vous ou commencez votre essai gratuit.
Method Article
ROS Imagerie des cellules vivantes au cours du développement neuronal
Dans cet article
Résumé
Ce protocole décrit l’utilisation d’un peroxyde d’hydrogène génétiquement codé (H2O2)-biocapteur dans les neurones et les larves de poisson zèbre cultivés pour évaluer les rôles physiologiques de signalisation deH2O2pendant le développement du système nerveux. Il peut être appliqué à différents types de cellules et modifié avec des traitements expérimentaux pour étudier les espèces réactives de l’oxygène (ROS) dans le développement général.
Résumé
Les espèces réactives de l’oxygène (ROS) sont des molécules de signalisation bien établies, qui sont importantes dans le développement normal, l’homéostasie, et la physiologie. Parmi les différents ROS,le peroxyde d’hydrogène (H2O2) est le mieux caractérisé en ce qui concerne les rôles dans la signalisation cellulaire. H2O2 a été impliqué lors du développement chez plusieurs espèces. Par exemple, une augmentation transitoire deH2O2 a été détectée chez les embryons de poissons zèbres au cours des premiers jours suivant la fécondation. En outre, l’épuisement d’une source cellulaire importante H2O2, NADPH oxydase (NOX), altère le développement du système nerveux tel que la différenciation, la croissance axonale, et le guidage des cellules ganglionnaires rétiniennes (RGCs) in vivo et in vitro. Ici, nous décrivons une méthode d’imagerie des niveaux intracellulaires deH2O2 dans les neurones de poissons zèbres cultivés et les larves entières pendant le développement en utilisant le biocapteur génétiquement codéH2 O2spécifique, roGFP2-Orp1. Cette sonde peut être exprimée de manière transitoire ou stable chez les larves de poissons zèbres. En outre, la lecture ratiométrique diminue la probabilité de détecter des artefacts en raison de l’expression génique différentielle ou des effets de volume. Tout d’abord, nous démontrons comment isoler et mettre en culture des GRC dérivés d’embryons de poissons zèbres qui expriment transitoirement roGFP2-Orp1. Ensuite, nous utilisons des larves entières pour surveiller les niveauxde H2O2au niveau des tissus. Le capteur a été validé par l’ajout deH2O2. De plus, cette méthodologie pourrait être utilisée pour mesurer les niveauxde H2O2dans des types de cellules et des tissus spécifiques en générant des animaux transgéniques avec une expression de biocapteur spécifique aux tissus. Comme les poissons zèbres facilitent les manipulations génétiques et développementales, l’approche démontrée ici pourrait servir de pipeline pour tester le rôle deH2O2 au cours du développement neuronal et embryonnaire général chez les vertébrés.
Introduction
La signalisation réactive des espèces de l’oxygène (ROS) régule le développement et le fonctionnement du système nerveux1. Une source importante de ROS cellulaires sont les oxydases NADPH (NOX), quisont des protéines transmembranaires générant du superoxyde et du peroxyde d’hydrogène (H2O2)2. Les enzymes NOX se trouvent dans tout le système nerveux central (SNC), et les ROS dérivés du NOX contribuent au développement neuronal3,4,5,6. Il a été démontré que le maintien et la différenciation des cellules souches neurales, l’établissement de la polarité neuronale, l’excroissance des neurites et la plasticité synaptique nécessitent des niveaux adéquats de ROS7,8,9,10,11. D’autre part, la production incontrôlée de ROS par les NEX contribue aux troubles neurodégénératifs, y compris la maladie d’Alzheimer, la sclérose en plaques et les lésions cérébrales traumatiques12,13,14. Par conséquent, la production de ROS physiologiquement pertinents est essentielle au maintien de conditions saines.
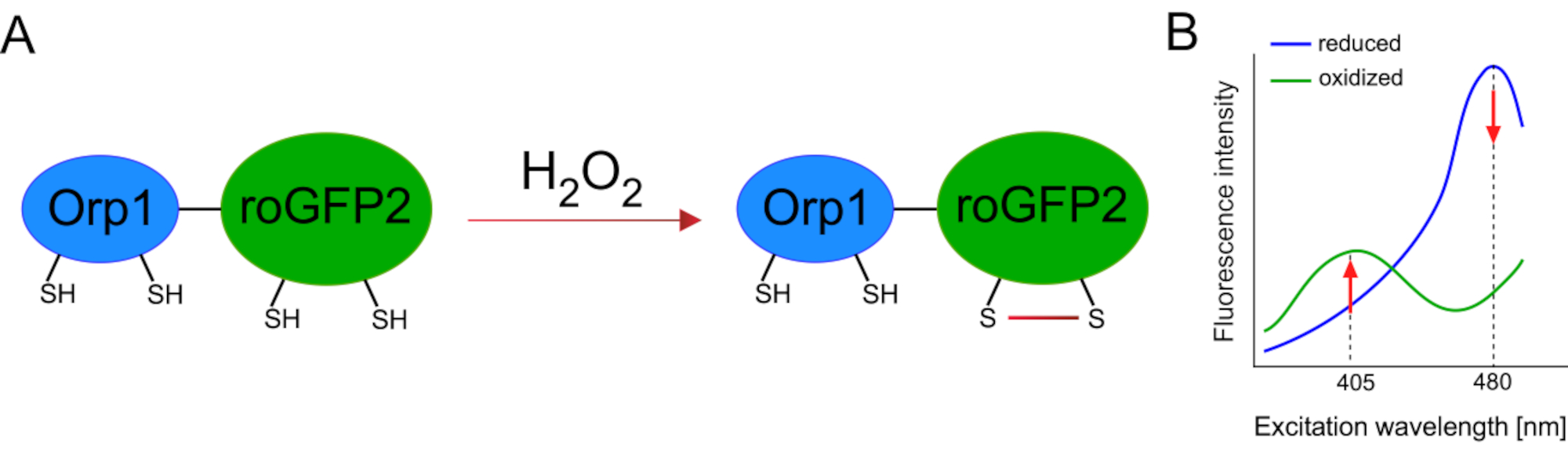
La mise au point de biocapteurs génétiquement codés a grandement facilité la détection des ROS cellulaires. Un avantage important des biocapteurs génétiquement codés est l’augmentation de la résolution temporelle et spatiale du signal ROS, car ces capteurs peuvent être spécifiquement ciblés à des endroits distincts. Le GFP sensible aux redox (roGFP) est l’un des types de ces biocapteurs ROS. La variante roGFP2-Orp1 détecte spécifiquement H2O2à travers son domaine Orp1, qui est une protéine de la famille des glutathion peroxiredoxin de levure15,16. L’oxydation de la protéine Orp1 est transférée dans roGFP2 pour modifier sa conformation(figure 1A). La sonde présente deux pics d’excitation proches de 405 nm et 480 nm, et un seul pic d’émission à 515 nm. Lors de l’oxydation, l’intensité de fluorescence autour des pics d’excitation change: alors que l’excitation de 405 nm augmente, l’excitation de 480 nm diminue. Ainsi, roGFP2-Orp1 est un biocapteur ratiométrique,et les niveauxH2O2sont détectés par le rapport des intensités de fluorescence excitées à deux longueurs d’onde différentes(figure 1B). Dans l’ensemble, roGFP2-Orp1 est un outil polyvalent pour l’imagerie ROS qui peut être utilisé efficacement in vivo.

Figure 1: Spectres schématiques de représentation et d’excitation de roGFP2-Orp1. (A) Le transfert d’oxydant se produit entre Orp1 et roGFP2 en réponse àH2O2,conduisant à des changements conformationnels dans roGFP2. (B) Les spectres d’excitation du roGFP2-Orp1 présentent deux pics d’excitation à 405 nm et 480 nm et un pic d’émission unique à 515 nm. Lors de l’oxydation parH2O2,l’excitation de 405 nm augmente tandis que l’excitation de 480 nm diminue. Il en résulte une lecture ratiométrique pour la présenceH2O2. Le chiffre a été modifié à partir de Bilan et Belousov (2017)16 et Morgan et coll. (2011)25. Veuillez cliquer ici pour voir une version plus grande de cette figure.
{kind=link}
Le système modèle Danio rerio (poisson zèbre) présente plusieurs avantages pour l’application de biocapteurs génétiquement codés. La transparence optique des embryons et des larves permet une imagerie in vivo non invasive. De nouveaux outils d’imagerie sont en cours de développement pour atteindre une résolution plus élevée et une pénétration plus profonde17. En outre, il existe des outils établis pour la manipulation génétique (expression ectopique de l’ARNm, transgénèse Tol2, etc.) et l’édition du génome (TALENs, CRISPR/Cas9, etc.), qui favorisent la génération d’animaux transgéniques18. Au fur et à mesure que les embryons de poisson zèbre se développent à l’extérieur de la mère, ce système facilite l’accès et la manipulation des embryons. Par exemple, les injections au stade unicellulaire et les traitements médicamenteux peuvent facilement être effectués.
Ici, nous avons utilisé le poissonzèbre pour exprimer transitoirement le biocapteur roGFP2-Orp1 spécifique àH2O2en injectant de l’ARNm transcrit in vitro. Ces embryons peuvent être utilisés à la fois pour l’imagerie in vitro de neurones cultivés et pour l’imagerie in vivo (figure 2). Nous décrivons un protocole pour disséquer et plaquer les cellules rétiniennes de ganglion (RGCs) des embryons de poisson zèbre suivis d’évaluer H2O2- niveaux dans les neurones cultivés. Ensuite, nous présentons une méthode d’imagerie in vivo d’embryons et de larves exprimant roGFP2-Orp1 à l’aide de la microscopie confocale. Cette approche permet non seulement de déterminer les niveaux physiologiques deH2O2-mais aussi les changements potentiels survenant à différents stades ou conditions de développement. Dans l’ensemble, ce système fournit une méthode fiable pour détecterH2O2dans les cellules vivantes et les animaux pour étudier le rôle de H2O2 dans le développement, la santé et la maladie.

Figure 2. Aperçu de l’approche expérimentale. En bref, après le prélèvement d’embryons, l’ARNm roGFP2-Orp1 est injecté dans le jaune d’embryons de poisson zèbre au stade cellulaire. Les embryons en développement peuvent être utilisés à la fois pour l’imagerie in vitro (A)et(B) in vivo. (A) Les embryons GFP-positifs sont utilisés pour disséquer des rétines pour la collecte de RGC à 34 hpf. Les RGCs dissociés sont plaqués sur des lamaux de couverture enduits de PDL/laminine dans des milieux ZFCM (+). L’imagerie du cône de croissance peut être effectuée car les RGCs étendent leurs axones après 6-24 h de placage. Les cellules peuvent être soumises à différents traitements pour mesurer les changements potentiels des niveaux deH2O2. Ici, nous avons mesuréles niveaux deH2O2dans les cônes de croissance des RGCs (rouge). (B) les embryons positifs à la GFP sont utilisés pour l’imagerie in vivo. À l’âge souhaité, les embryons peuvent être anesthésiés et montés sur des plats à fond de verre de 35 mm pour l’imagerie confocale. Ici, les embryons sont montés ventralement pour l’imagerie rétinienne. Le schéma montre le développement rétinien chez le poisson zèbre. Les RGCs forment la couche cellulaire ganglionnaire (GCL), qui est la couche la plus interne de la rétine. Les axones de RGC se développent en nerf optique pour croiser le midline, formant le chiasm optique. Ensuite, les axones RGC se développent dorsalement pour faire des synapses dans le tectum optique dans le mécérine. Veuillez cliquer ici pour voir une version plus grande de cette figure.
{kind=link}
Access restricted. Please log in or start a trial to view this content.
Protocole
Toutes les expériences sur les animaux ont été examinées et approuvées sur le plan éthique par le Purdue Animal Care and Use Committee (PACUC), conformément aux directives des NIH avec le protocole 2006002050 approuvé le 24/07/2020.
1. Préparation des solutions
- Support E2 (1x)
- Préparer des solutions 100x E2A (500 mL), 500x E2B (100 mL) et 500x E2C (100 mL) en combinant tous les composants indiqués dans le tableau 1. Solutions Autoclave E2A, E2B et E2C. Conserver à 4 °C.
- Pour 1x support E2 : Combinez 5 mL de 100x E2A, 1 mL de 500x E2B et 1 mL de 500x E2C. Porter le volume à 500 mL avec de l’eau stérile. Ajuster le pH à 7,0-7,5.
- Préparez des aliquotes de 50 mL de 1x magasin de médias E2 à -20 °C pour un stockage à long terme. Cependant, des précipitations peuvent se produire lors du dégel. Assurez-vous que la précipitation est complètement dissoute avant d’utiliser la solution mère.
| solution | composant | quantité | concentration |
| 100X E2A (500mL) | |||
| NaCl | 43,8 g | 1500 mM | |
| Kcl | 1,88 g | 50 mM | |
| MgSO4 | 6 g | 100 mM | |
| KH2PO4 | 1,03 g | 15 mM | |
| Na2HPO4 | 0,34 g | 5 mM | |
| 500X E2B (100 mL) | |||
| CaCl2 | 5,5 g | 500 mM | |
| E2C 500X (100 mL) | |||
| NaHCO3 | 3 g | 350 mM | |
| 1X E2 (500 mL) | |||
| 100X E2A | 5 mL | 1X | |
| E2B 500X | 1 mL | 1X | |
| E2C 500X | 1 mL | 1X |
Tableau 1 : Composants de 1x milieu E2 pour la culture cellulaire du poisson zèbre.
- Médias E3 (1x)
- Dissoudre les composants dans de l’eau stérile de 1 L comme indiqué dans le tableau 2 pour faire 100x stock. Diluer le stock dans de l’eau stérile pour faire 1x support E3.
- Ajouter 0,2 % de bleu de méthylène. Pour 20 mL de 1x média E3, ajouter 40 μL de bleu de méthylène.
- Faire un autre lot sans bleu de méthylène pour l’imagerie fluorescente.
| composant | Montant (g) | Concentration en stock 100X (mM) |
| NaCl | 29.22 | 500 |
| Kcl | 1.26 | 17 |
| CaCl2 2H2O | 4.85 | 33 |
| MgSO4 7H2O | 8.13 | 33 |
Tableau 2 : Composants des milieux 100x E3 pour le maintien des embryons de poissons zèbres.
- Solution mère saline 80x
- Combinez tous les composants indiqués dans le tableau 3. Ajouter de l’eau pour faire une solution de 100 mL. Mélanger jusqu’à ce que tous les composants soient dissous. Conserver la solution à 4 °C.
| composant | Montant (g) | Concentration en stock (mM) |
| glucose | 1.44 | 80 |
| Sodium Pyruvate | 0.44 | 40 |
| CaCl2 2H2O | 0.148 | 10 |
| HEPES | 6.1 | 256 |
Tableau 3 : Composants d’une solution saline 80x pour les milieux de culture cellulaire de poisson zèbre.
- Milieu de culture cellulaire du poisson zèbre (ZFCM+)
- Combinez tous les composants indiqués dans le tableau 4 pour créer un support de 250 mL. Régler le pH à 7,5. Filtrer les milieux à l’aide d’un filtre de 0,22 μm et les conserver à 4 °C.
| composant | Montant (mL) | Volume % |
| Milieu L-15 (avec du rouge de phénol) | 212.75 | 85.1 |
| Sérum fœtal bovin (SBF) | 5 | 2 |
| Pénicilline/streptomycine | 1 | 0.4 |
| Solution saline 80X | 3.125 | 1.25 |
| Eau | 28.125 | 11.25 |
Tableau 4 : Composants du milieu de culture cellulaire du poisson zèbre avec sérum et antibiotiques.
- Milieu de culture cellulaire de poisson zèbre pour l’imagerie (ZFCM-)
- Combinez tous les composants indiqués dans le tableau 5 pour créer un support de 250 mL. Régler le pH à 7,5. Média filtrant utilisant un filtre de 0,22 μm.
- Faire des aliquotes à usage unique (lots de 50 mL) pour prévenir la contamination. Conserver à 4 °C.
| composant | Montant (mL) | Volume % |
| Milieu L-15 (pas de rouge de phénol) | 212.75 | 85.1 |
| Solution saline 80X | 3.125 | 1.25 |
| Eau | 34.125 | 13.65 |
Tableau 5 : Composants du milieu de culture cellulaire du poisson zèbre sans sérum ni antibiotiques pour l’imagerie in vitro.
- Moules d’injection
- Dissoudre 1,5 % de l’agarose dans les milieux E3. Verser ~ 25 ml d’agarose dans une boîte de Pétri de 100 x 15 mm.
- Posez le moule sur le dessus de l’agarose avec un angle de 45 ° par rapport à la surface, et laissez-le flotter sur l’agarose au ralenti. Cela aidera à éviter les bulles. Laissez l’agarose refroidir et solidifier sur le dessus de la banquette.
- Une fois complètement solidifié, retirez le moule lentement pour éviter la rupture de l’agarose. Ajouter du support E3 frais, ajouter un film de paraffine autour du plat pour éviter les déversements et conserver à 4 °C (figure 3).

Figure 3: Images de moule d’injection. (A) Moule en plastique utilisé pour fabriquer des plaques d’injection. Le moule a six rampes, une à 90 ° et une côté biseauté à 45 ° pour maintenir les embryons en place. (B) La plaque d’injection après l’agarose solidifiée et le moule est retiré. Barres d’échelle = 1 cm. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
2. Préparation et injection d’ARNm roGFP2-Orp1
REMARQUE: la construction roGFP2-Orp1 a été obtenue auprès du Dr Tobias Dick, DKFZ, Allemagne. Il a été sous-cloné dans le vecteur pCS2+ dans le laboratoire du Dr Qing Deng de l’Université Purdue. Pour prévenir la dégradation par RNase, plusieurs précautions doivent être prises. Des réactifs et des tubes sans RNase doivent être utilisés en tout temps, des gants doivent être portés pour toutes les étapes et, alternativement, les matériaux et les surfaces peuvent être essuyés avec un agent nettoyant pour l’élimination RNase.
- Linéariser les 3-10 μg du vecteur pCS2+/roGFP2-Orp1 avec NotI.
- Purifier le plasmide linéarisé avec un kit de nettoyage PAR PCR.
- Transcrire in vitro l’ARNm roGFP2-Orp1 avec un kit de transcription in vitro selon les instructions fournies par le fabricant.
- Ensemble réaction de transcription plafonné
- Placer l’ARN polymérase et l’ADN linéarisé/purifié sur de la glace. Tampon de réaction Vortex 10x et 2x NTP/CAP jusqu’à ce qu’ils soient complètement en solution. Entreposer le NTP/CAP sur de la glace, mais conserver le tampon à température ambiante (RT) tout en assemblant la réaction. Touchez tous les réactifs avant d’ouvrir les tubes pour éviter la contamination.
- Mettre en place la réaction de synthèse de l’ARN dans l’ordre indiqué ci-dessous à RT dans un tube centrifuge de 0,5 mL sans RNase. Le volume final de la réaction est de 20 μL. La configuration de la réaction est indiquée dans le tableau 6.
- Ajouter 10 μL de mélange de ribonucléotide, 2x NTP/CAP et de l’eau sans nucléase, si nécessaire, dans le tube. Ajouter 2 μL de tampon de réaction 10X. Ajouter 1-1,5 μg d’ADN linéaire (jusqu’à 6 μL). Au besoin, ajouter de l’eau sans nucléase pour compléter le volume de réaction de 20 μL.
- Fermez le tube, le vortex brièvement et le microfuge touch-spin. Ajouter 2 μL de mélange d’enzymes 10x SP6. Fermez avec un clic de doigt et touchez-tournez dans un microfuge.
- Placer à 37 °C pendant 2-2,5 h (peut aller jusqu’à 18 h).
REMARQUE: Dans l’expérience présentée pendant la nuit 16-18 h incubation à 37 °C a été effectuée pour de meilleurs résultats. - Ajouter 1 μL de DNase pour enlever le gabarit d’ADN, cliquer sur le doigt, toucher la rotation et incuber à 37 °C pendant 15 min.
- Ensemble réaction de transcription plafonné
| Réactif | Volume (μL) | Montant en réaction |
| 2X NTP/CAP | 10 | 1X |
| Tampon de réaction 10X | 2 | 1X |
| ADN modèle | Jusqu’à 6 | 1-1,5 μg |
| Eau sans nucléase | Ajouter pour faire 20 μL | |
| Mélange d’enzymes 10X SP6 | 2 | 1X |
Tableau 6 : Configuration de la réaction pour la transcription in vitro de l’ARNm roGFP2-Orp1.
- Récupération de l’ARN
- Ajouter 25 μL de chlorure de lithium (LiCl) fourni dans la trousse de transcription in vitro. Placer à -20 °C dans un congélateur non gelé pendant au moins 30 min.
- Tourner pendant 25 min à la vitesse maximale dans une centrifugeuse de table à 4 °C. Retirez et jetez soigneusement le surnageant afin de ne pas déranger les pastilles. Ajouter 25 μL d’éthanol froid à 75 % et tourner pendant 5 min à 4 °C.
- Retirez soigneusement et jetez le surnageant. Laisser sécher à l’air pendant au moins 5 min à TA. Ne laissez pas sécher. Ajouter 12 μL de tampon Tris-EDTA (TE) exempt de nucléase (pH 7,0) et conserver l’échantillon sur la glace.
- Mesurer la concentration d’ARN avec un spectrophotomètre. On obtient habituellement 0,5 à 1 μg/μL.
- Préparer 100 ng/μL d’ARNm dans une solution rouge de phénol (0,5 % de rouge de phénol dans la solution saline tamponnée au phosphate -DPBS) de Dulbecco et une partie aliquote à usage unique (3-5 μL). Conserver les aliquotes de l’ARNm à - 80 °C.
- Microinjection de l’ARNm
- Le jour de l’injection, utilisez l’une des aliquotes d’ARNm et suivez le protocole d’injection d’embryons de poisson zèbre pour injecter 1 nL de l’ARNm dans les embryons au stade unicellulaire à travers leur jaune19. Une brève description est fournie ci-dessous.
- Élever des poissons adultes et recueillir des embryons comme décrit précédemment20.
- Tirez les aiguilles de microinjection avec un tireur de pipette. Coupez la pointe des aiguilles avec une pince pour créer une ouverture de pointe de 10 μm.
- Aligner les embryons dans un moule d’injection qui a été décrit à l’étape 1.6.
- Injecter 1 nL de l’ARNm dans du rouge de phénol avec une pipette de microinjection en verre.
- Recueillir des embryons et les conserver dans les milieux E3.
- Conservez les embryons dans l’incubateur à 27 °C en milieux E3 jusqu’à ce que le stade de développement souhaité soit atteint. Les embryons injectés peuvent être utilisés à la fois pour l’imagerie in vitro (section 3 et section 4) et in vivo (section 5). Les embryons exprimant roGFP2-Orp1 peuvent être présélectionnés avant les expériences avec un microscope de dissection régulier équipé de lumière fluorescente et d’un ensemble de filtres bleu/vert.
3. Culture cellulaire ganglionnaire rétinienne primaire dérivée d’embryons de poissons zèbres
Remarque : ce protocole est adapté d’une méthode précédemment publiée 21. Effectuez les étapes 3.1 et 3.2 dans une hotte à flux laminaire.
- Préparation des lamaux de couverture
- Préparer 4-6 plaques de culture dans chaque expérience. Utilisez des lamelle de couverture nettoyées à l’acide (22 x 22 mm carrés; épaisseur 0,16-0,19 mm) qui sont stockées dans de l’éthanol à 100 %.
- Retirez une lame de couverture du contenant d’entreposage à l’aide d’une pince et enflammez-la pour éliminer l’éthanol résiduel.
- Séchez complètement la lamelle à couvercle en la plaçant à un angle à l’intérieur d’un plat de culture de 35 mm.
- Préparer la solution de travail de poly-D-lysine (PDL) (1x) en diluant 10x stock (5 mg/mL) dans de l’eau stérile.
- Appliquer 100 μL de 0,5 mg/mL de PDL au centre de chaque lamelle et éviter l’étalement de la solution sur les bords.
- Incuber la LDO sur des lamelle pendant 20-30 min à température ambiante (RT). Assurez-vous que le PDL ne se dessèche pas.
- Lavez le PDL avec 0,5 mL d’eau stérile trois fois. Laissez les plaques sécher complètement.
- Préparer la solution de travail de laminine (1x) en diluant 50x stock (1 mg/mL) dans 1x PBS.
- Appliquer 100 μL de 20 μg/mL de laminine dans du PBS au centre de chaque lamelle et éviter la propagation de la solution sur les bords.
- Incuber les plaques à 37 °C dans un incubateur humidifié pendant 2-6 h. Évitez le séchage de la solution de laminine.
- Dissection embryonnaire et placage des GRC
- Préparer et étiqueter quatre plats de culture tissulaire de 35 mm et les remplir de 4 mL d’éthanol à 70 %, de « milieu E2 1 », « milieu E2 2 », « milieu E2 3 » le jour de la dissection. Conservez la vaisselle au réfrigérateur jusqu’aux dissections.
- Lorsque les embryons de poisson zèbre sont 34 heures après la fécondation (hpf), sortez les plats de culture recouverts de laminine de l’incubateur et lavez les lamaux trois fois avec 0,5 mL de PBS 1x.
- Après le lavage final, ajoutez 4 mL de milieu ZFCM(+) à chaque plat de culture et évitez de sécher la plaque.
- Récupérez les plats de culture préparés à l’étape 3.2.1. Qu’ils s’équilibrent à RT.
- Remplissez 4 à 6 tubes PCR avec 15 μL de milieu ZFCM(+). Un tube est nécessaire pour préparer les RGCs de 4 yeux à plaquer sur une lamelle de couverture.
- Récupérer des embryons de poisson zèbre de l’incubateur et immerger les embryons dans un plat de culture de tissus de 35 mm contenant 70% d’éthanol pendant 5-10 s pour stériliser.
- À l’aide d’une pipette de transfert, transférer les embryons dans un plat E2 Media 1 contenant un support E2 stérile pour laver l’excès d’éthanol.
- Transférer les embryons dans le plat E2 Media 2 et retirer leurs chorions avec une pince tranchante.
- Transférer les embryons dans la parabole finale E2 Media 3 pour effectuer des dissections.
- À l’aide d’une paire de pinces fines, disséquer les rétines comme décrit précédemment 22.
- Positionnez et maintenez les embryons antérieurs à leur jaune avec l’une des pinces et retirez la queue postérieure au sac vitellin avec les autres pinces.
- Prenez le cou avec une pince et enlevez la tête pour exposer le cerveau et les yeux aux médias E2. Évitez de couper le sac vitellin.
- Avec la pointe de la pince fine, rouler doucement les yeux de la tête, tout en tenant le tissu crânien vers le bas avec la deuxième pince. Gardez les yeux isolés des débris de tissu adjacents.
- Transférer quatre yeux sur l’un des tubes préalablement préparés contenant du ZFCM(+).
- Titrez doucement de haut en bas avec la pipette P20 et une pointe jaune environ 45 fois pour dissocier les cellules. Évitez les bulles d’air.
- Transférez le ZFCM(+) avec des cellules dissociées au centre de la lamelle. Répétez les étapes 10 à 12 pour des lamelle supplémentaires.
- Maintenir les cultures sur paillasse à 22 °C sur un support en mousse de polystyrène pour absorber les vibrations.
- Effectuer l’imagerie 6-24 h après le placage.
REMARQUE: Utilisez la pipette de transfert pour transférer des embryons vers différents plats de culture. Changer la pipette de chaque solution pour éviter le transport de l’éthanol (étapes 6 à 8).
4. Imagerie ROS in vitro des neurones RGC cultivés
- Le jour de l’imagerie (généralement 6-24 h après le placage cellulaire), vérifiez les cellules au microscope pour valider la croissance des axones RGC.
- Pour l’imagerie à cellules vivantes, transférez les lamaux de couverture de la parabole de culture vers une chambre d’imagerie de cellules vivantes. Dans ce cas, une chambre ouverte sur mesure, qui a été précédemment décrite, a été utilisée23.
- Configurez le microscope pour l’imagerie. Utilisez un microscope inversé équipé d’un objectif de contraste différentiel d’interférence (DIC), d’un filtre rouge OG590 à passe longue et d’une caméra EM-CCD.
- Avant l’imagerie, remplacez le milieu ZFCM(+) par ZFCM(-).
- Une fois que les cellules sont positionnées avec un objectif 10x, acquérir des images à un grossissement de 60x à l’aide d’un objectif d’immersion dans l’huile NA élevé. Utilisez un grossissement supplémentaire de 1,5x.
- Tout d’abord, acquérir des images DIC. Ensuite, image roGFP2-Orp1 à l’aide d’un jeu de filtres approprié. Exciter roGFP2-Orp1 avec des filtres d’excitation 405/20 et 480/30 nm séquentiellement et acquérir des images avec filtre d’émission 535/30 nm après que la lumière d’émission a passé le miroir dichroïque 505DCXR.
- Après avoir pris le premier ensemble d’images, échangez des médias avec des médias contenant différentes solutions de traitement. Les supports doivent être changés toutes les 30 minutes d’imagerie pour éviter les changements de pH et d’osmolarité.
5. Imagerie ROS in vivo d’embryons en développement
- Pour l’imagerie in vivo, conservez les embryons dans un milieu E3 contenant 0,003% de phénylthiourée (PTU) sans bleu de méthylène de 22 à 24 hpf. Échangez des médias et retirez quotidiennement les embryons morts.
- À l’âge souhaité, anesthésier les embryons dans 0,016% de tricaine. Monter des embryons anesthésiés dans de l’agarose à faible fusion à 1% sur des plats de culture de fond de verre de 35 mm. Les embryons peuvent être orientés dorsalement, ventralement ou latéralement, selon la région d’intérêt pour l’imagerie.
- Après solidifie l’agarose, remplissez les plats avec un support E3 sans bleu de méthylène / 0,016% de tricaine.
- Configurez le microscope pour l’imagerie. Utilisez un microscope confocal à balayage laser inversé. Vous pouvez également utiliser un microscope confocal vertical équipé d’une lentille d’immersion dans l’eau pour imager des embryons montés sur une goutte d’agarose.
- Exciter roGFP2-Orp1 avec des filtres d’excitation de 405 nm et 488 nm séquentiellement et acquérir des images correspondantes avec des filtres d’émission dans la gamme de 515-535 nm.
- Acquérir des z-stacks d’une épaisseur de section de 5 μm à travers la partie souhaitée des embryons. Les embryons peuvent être conservés pour l’imagerie à des stades ultérieurs de développement.
- Après l’imagerie, retirer les embryons de l’agarose avec une pince fine et les conserver dans l’incubateur jusqu’à l’âge souhaité dans des milieux sans bleu de méthylène avec PTU.
6. Analyse et traitement des images
- Mesure des niveaux deH2O2 sur la base des valeurs du rapport 405/480
- Utilisez un logiciel approprié pour l’analyse d’images. Le logiciel ImageJ a été utilisé ici pour l’analyse et le traitement d’images.
- Ouvrez les images DIC, 405/535 et 480/535 dans le logiciel ImageJ en faisant glisser les fichiers ou en cliquant sur Fichier | Ouvrez. Si ce n’est pas déjà fait, convertissez les images en 32 bits en cliquant sur Image | Type | 32 bits.
- Définissez la région d’intérêt (ROI) avec l’outil à main levée de la barre de contrôle (corps cellulaire, cône de croissance, rétine, etc.). Ouvrez le gestionnaire de retour sur investissement en cliquant sur Analyser | Outils | Gestionnaire de roi. Cliquez sur Ajouter dans l’onglet Gestionnaire de retour sur investissement pour ajouter le retour sur investissement défini.
- Dessinez une région proche du retour sur investissement et ajoutez-le en tant que retour sur investissement d’arrière-plan. Mesurez les valeurs d’arrière-plan moyennes en sélectionnant le retour sur investissement et en cliquant sur Mesurer dans l’onglet Gestionnaire de retour sur investissement.
- Notez les valeurs d’intensité moyenne de la mesure. Soustrayez la valeur de l’arrière-plan moyen des images fluorescentes en cliquant sur Traiter | | mathématiques Soustraire. Effectuez cette étape pour les images 405/535 et 480/535.
- Ajoutez la valeur de « 1 » à l’image fluorescente 480/535 pour éliminer les valeurs « 0 » en cliquant sur Traiter | | mathématiques Ajoutez une fonction avant le calcul du ratio.
- Cliquez sur Traiter | Calculatrice d’images | Diviser la fonction dans ImageJ pour diviser l’image 405/535 par 480/535 image pixel par pixel. Sélectionnez l’image 405/535 à diviser par l’image 480/535. Sélectionnez une image de sortie 32 bits.
- Appliquez le retour sur investissement à l’image de ratio en cliquant d’abord sur l’image de ratio, puis sur le retour sur investissement dans l’onglet Gestionnaire de retour sur investissement.
- Mesurez les valeurs de rapport moyennes de l’image 405/535 à l’image 480/535 en cliquant sur Mesurer dans l’onglet Gestionnaire de retour sur investissement.
- Effectuer les étapes 6.1.2 à 6.1.9 pour autant d’échantillons que possible afin d’effectuer l’analyse statistique appropriée.
- Affichage de l’image de ratio
REMARQUE : cette procédure consiste à soustraire l’arrière-plan à l’extérieur de l’échantillon et à appliquer une table de recherche de couleur à l’image.- Une fois que l’image de ratio a été créée dans ImageJ à l’étape 6.1.7., créez une image noire 32 bits en cliquant sur Fichier | Nouvelle| Image.
- Appliquez le retour sur investissement pour lequel vous souhaitez afficher les niveaux H2O2 à la nouvelle image en cliquant d’abord sur la nouvelle image, puis sur le retour sur investissement à partir de l’onglet Gestionnaire de retour sur investissement.
- Créer un masque en cliquant sur Modifier | | de sélection Créer un masque.
- Divisez l’image du masque par 255 pour ajuster la valeur du retour sur investissement à « 1 » et les valeurs d’arrière-plan seront « 0 ». Cliquez sur Traiter | | mathématiques Diviser et taper 255.
- Multipliez le masque avec l’image de ratio en cliquant sur Traiter | Calculatrice d’images | Fonction multiplier. Il en résultera une image en échelle de gris affichant uniquement le retour sur investissement.
- Remplacez la table de recherche par « Fire » en cliquant sur Image | Rechercher des tables | Feu.
REMARQUE : un facteur de multiplication peut être appliqué à toutes les images pour une meilleure visualisation du ratio en cliquant sur Traiter | | d’images Multiplier. - Convertissez l’image de ratio en 8 bits en cliquant sur Image | Type | 8 bits.
- Ajouter une barre d’étalonnage en cliquant sur Analyser | Outils | Barre d’étalonnage.
Access restricted. Please log in or start a trial to view this content.
Résultats
Les CRG de poissons zèbres d’élevage étendent les axones à l’intérieur de 1d. Une image représentative au rapport 405/480du biocapteurH2O2est représentée sur la figure 4A. Le corps cellulaire, axone, et les cônes de croissance sont clairement visibles dans les neurones individuels. Ces neurones peuvent être soumis à différents traitements au fil du temps pour surveiller les changementsH2O2. Nous avons précédemment constaté que l?...
Access restricted. Please log in or start a trial to view this content.
Discussion
Il y a plusieurs étapes critiques qui nécessitent une attention particulière tout au long de ce protocole. Nous pensons que la prise en compte de ces points améliorera le flux expérimental. Pour la culture primaire de RGC, la stérilité du ZFCM(-) est très importante, car ce milieu de culture ne contient pas d’antibiotiques et la contamination peut survenir avant ou pendant l’imagerie. Pour l’éviter, nous vous conseillons de préparer et d’utiliser ZFCM(-) uniquement à l’intérieur d’une armoire de b...
Access restricted. Please log in or start a trial to view this content.
Déclarations de divulgation
Les auteurs déclarent qu’ils n’ont pas de conflit d’intérêts.
Remerciements
Ce travail a été soutenu par les National Institutes of Health (Grant R01NS117701), la National Science Foundation (Grant 1146944-IOS), l’Indiana Traumatic Spinal Cord and Brain Injury Research Fund (Grant 20000289), la Purdue Research Foundation (Grant 209911) et le Bureau du vice-président exécutif pour la recherche et les partenariats à l’Université Purdue (Subvention 210362). Nous remercions le Dr Cory J. Weaver et Haley Roeder d’avoir établi le protocole d’élevage du poisson zèbre RGC. Nous remercions également Haley Roeder d’avoir fourni les données de la figure 4. Nous remercions Leah Biasi et Kenny Nguyen pour leur aide à la culture RGC. Nous remercions Gentry Lee d’avoir édité le texte. Nous remercions le Dr Tobias Dick d’avoir fourni roGFP2-Orp1 et le Dr Qing Deng pour le vecteur pCS2+ contenant roGFP2-Orp1. La figure 2 est créée avec Biorender.com.
Access restricted. Please log in or start a trial to view this content.
matériels
| Name | Company | Catalog Number | Comments |
| 35-mm culture dish | Sarstedt | 83-3900 | |
| 35-mm glass bottom dish | MatTek | P35G-1.5-10-C | |
| Agarose | Fisher Scientific | BP160-500 | |
| Borosilicate Glass Capillary Tubes | Sutter/Fisher Scientific | NC9029378 | |
| Calcium Chloride Dihydrate | Fisher Scientific | C79-500 | |
| Cover glass | Corning | 2850-22 | |
| Disposable Petri Dishes (100 x 15 mm) | VWR | 25384-094 | |
| Fetal Bovine Serum | ThermoFisher Scientific | 26140087 | |
| Glucose | Sigma Aldich | G7528 | |
| HEPES | Sigma Aldich | H4034 | |
| Injection Mold | Adaptive Science Tools | TU-1 | |
| Inverted Microscope | Nikon | TE2000 | |
| Laminin | ThermoFisher Scientific | 23017-015 | |
| Laser Scanning Confocal Microscopy | Zeiss | 710 | |
| Leibovitz's L-15 Medium with phenol red | Gibco/Fisher Scientific | 11-415-064 | |
| Leibovitz's L-15 Medium without phenol red | Gibco/Fisher Scientific | 21-083-027 | |
| Low melting agarose | Promega | V2111 | |
| mMessage mMachine SP6 Transcription Kit | Invitrogen | AM1340 | |
| NotI | New England Biolabs | R0189S | |
| PBS | Hyclone/Fisher Scientific | SH3025601 | |
| Penicillin/streptomycin | ThermoFisher Scientific | 15140122 | |
| Phenol Red | Sigma Aldich | P0290 | |
| Phenylthiourea (PTU) | Sigma Aldich | P7629 | |
| Pneumatic PicoPump | World Precision Instruments | PV820 | |
| Poly-D-Lysine (PDL) | Sigma Aldich | P7280 | |
| QiaQUICK PCR Purification Kit | QIAGEN | 28104 | |
| Recombinant mouse slit2 | R&D Systems | 5444-SL-050 | |
| Sodium Pyruvate | Sigma Aldich | P5280 | |
| Steritop 0.22 µm filter | Millipore | S2GPT05RE | |
| TE Buffer | Ambion | AM9860 | |
| Tricaine Methanesulfonate | Sigma Aldich | E10521 | |
| Vertical Pipette Puller | David Kopf Instruments | 700C |
Références
- Bórquez, D. A., et al. Dissecting the role of redox signaling in neuronal development. Journal of Neurochemistry. 137 (4), 506-517 (2016).
- Bedard, K., Krause, K. -H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiological Reviews. 87 (1), 245-313 (2007).
- Weaver, C. J., Leung, Y. F., Suter, D. M. Expression dynamics of NADPH oxidases during early zebrafish development. Journal of Comparative Neurology. 524 (10), 2130-2141 (2016).
- Terzi, A., Suter, D. M. The role of NADPH oxidases in neuronal development. Free Radical Biology and Medicine. 154, 33-47 (2020).
- Infanger, D. W., Sharma, R. V., Davisson, R. L. NADPH oxidases of the brain: Distribution, regulation, and function. Antioxidants & Redox Signaling. 8 (9-10), 1583-1596 (2006).
- Coyoy, A., Olguin-Albuerne, M., Martinez-Briseno, P., Moran, J. Role of reactive oxygen species and NADPH-oxidase in the development of rat cerebellum. Neurochemistry International. 62 (7), 998-1011 (2013).
- Le Belle, J. E., et al. Proliferative neural stem cells have high endogenous ROS levels that regulate self-renewal and neurogenesis in a PI3K/Akt-dependant manner. Cell Stem Cell. 8 (1), 59-71 (2011).
- Nayernia, Z., et al. Decreased neural precursor cell pool in NADPH oxidase 2-deficiency: from mouse brain to neural differentiation of patient derived iPSC. Redox Biology. 13, 82-93 (2017).
- Wilson, C., Nunez, M. T., González-Billault, C. Contribution of NADPH oxidase to the establishment of hippocampal neuronal polarity in culture. Journal of Cell Science. 128 (16), 2989-2995 (2015).
- Munnamalai, V., et al. Bidirectional interactions between Nox2-type NADPH oxidase and the F-actin cytoskeleton in neuronal growth cones. Journal of Neurochemistry. 130 (4), 526-540 (2014).
- Kishida, K. T., et al. Synaptic plasticity deficits and mild memory impairments in mouse models of chronic granulomatous disease. Molecular and Cellular Biology. 26 (15), 5908-5920 (2006).
- Ravelli, K. G., et al. Nox2-dependent neuroinflammation in an EAE model of multiple sclerosis. Translational Neuroscience. 10 (1), 1-9 (2019).
- Park, L., et al. Nox2-derived radicals contribute to neurovascular and behavioral dysfunction in mice overexpressing the amyloid precursor protein. Proceedings of the National Academy of Sciences. 105 (4), 1347-1352 (2008).
- Schiavone, S., Neri, M., Trabace, L., Turillazzi, E. The NADPH oxidase NOX2 mediates loss of parvalbumin interneurons in traumatic brain injury: Human autoptic immunohistochemical evidence. Scientific Reports. 7 (1), 8752(2017).
- Gutscher, M., et al. Proximity-based protein thiol oxidation by H2O2-scavenging peroxidases. Journal of Biological Chemistry. 284 (46), 31532-31540 (2009).
- Bilan, D. S., Belousov, V. V. New tools for redox biology: from imaging to manipulation. Free Radical Biology and Medicine. 109, 167-188 (2016).
- Abu-Siniyeh, A., Al-Zyoud, W. Highlights on selected microscopy techniques to study zebrafish developmental biology. Laboratory Animal Research. 36 (1), 12(2020).
- Sassen, W. A., Koster, R. W. A molecular toolbox for genetic manipulation of zebrafish. Advances in Genomics and Genetics. 5, 151-163 (2015).
- Rosen, J. N., Sweeney, M. F., Mably, J. D. Microinjection of zebrafish embryos to analyze gene function. Journal of Visualized Experiments. (25), e1115(2009).
- Avdesh, A., et al. Regular care and maintenance of a Zebrafish (Danio rerio) laboratory: An introduction. Journal of Visualized Experiments. (69), e4196(2012).
- Chen, Z., et al. Primary neuron culture for nerve growth and axon guidance studies in Zebrafish (Danio rerio). PLoS One. 8 (3), 57539(2013).
- Zhang, L., Leung, Y. F. Microdissection of zebrafish embryonic eye tissues. Journal of Visualized Experiments. (40), e2028(2010).
- Suter, D. M. Live cell imaging of neuronal growth cone motility and duidance in vitro. Cell Migration: Methods in Molecular Biology. , 65-86 (2011).
- Weaver, C. J., et al. nox2/cybb deficiency affects zebrafish retinotectal connectivity. Journal of Neuroscience. 38 (26), 5854-5871 (2018).
- Morgan, B., Sobotta, M. C., Dick, T. P. Measuring EGSH and H2O2 with roGFP2-based redox probes. Free Radical Biology and Medicine. 51, 1943-1951 (2011).
- Li, Z., et al. Phenylthiourea specifically reduces zebrafish eye size. PLoS One. 7 (6), 40132(2012).
- Ermakova, Y. G., et al. Red fluorescent genetically encoded indicator for intracellular hydrogen peroxide. Nature Communications. 5, 5222(2014).
- Oparka, M., et al. Quantifying ROS levels using CM-H2DCFDA and HyPer. Methods. 109, 3-11 (2016).
- Dickinson, B. C., Peltier, J., Stone, D., Schaffer, D. V., Chang, C. J. Nox2 redox signaling maintains essential cell populations in the brain. Nature Chemical Biology. 7 (2), 106-112 (2011).
- Cannon, M. B., Remington, S. J. Redox-sensitive green fluorescent protein: Probes for dynamic intracellular redox responses. A review. Methods in Molecular Biology. 476, 51-65 (2009).
- Meyer, A. J., Dick, T. P. Fluorescent protein-based redox probes. Antioxidants & Redox Signaling. 13 (5), 621-650 (2010).
- Panieri, E., Millia, C., Santoro, M. M. Real-time quantification of subcellular H2O2 and glutathione redox potential in living cardiovascular tissues. Free Radical Biology and Medicine. 109, 189-200 (2017).
- Breus, O., Dickmeis, T. Genetically encoded thiol redox-sensors in the zebrafish model: Lessons for embryonic development and regeneration. Biological Chemistry. , (2020).
- Morgan, B., et al. Real-time monitoring of basal H2O2 levels with peroxiredoxin-based probes. Nature Chemical Biology. 12 (6), 437-443 (2016).
- Terzi, A., Roeder, H., Weaver, C. J., Suter, D. M. Neuronal NADPH oxidase 2 regulates growth cone guidance downstream of slit2/robo2. Developmental Neurobiology. , (2020).
- Bilan, D. S., Belousov, V. V. HyPer family probes: State of the art. Antioxidants & Redox Signaling. 24 (13), 731-751 (2016).
- Ermankova, Y. G., et al. SypHer3s: a genetically encoded fluorescent ratiometric probe with enhanced brightness and an improved dynamic range. Chemistry Communications. 54 (23), 2898-2901 (2018).
- Pak, V. V., et al. Ultrasensitive genetically encoded indicator for hydrogen peroxide identifies roles for the oxidant in cell migration and mitochondrial function. Cell Metabolism. 31 (3), 642-653 (2020).
- Kwan, K. M., et al. The Tol2kit: A multisite gateway-based construction Kit for Tol2 transposon transgenesis constructs. Developmental Dynamics. 236 (11), 3088-3099 (2007).
Access restricted. Please log in or start a trial to view this content.
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.