
JoVE 비디오를 활용하시려면 도서관을 통한 기관 구독이 필요합니다. 전체 비디오를 보시려면 로그인하거나 무료 트라이얼을 시작하세요.
Method Article
신경 발달 도중 ROS 살아있는 세포 화상 진찰
요약
이 프로토콜은 신경계 발달 중H2 O2의생리적 신호 역할을 평가하기 위한 배양 된 제브라피시 뉴런 및 애벌레에서 유전으로 인코딩 된 과산화수소 (H2O2)-바이오센서의 사용을 설명합니다. 그것은 다른 세포 유형에 적용 하 고 일반적인 개발에 반응성 산소 종 (ROS)를 공부 하는 실험 치료와 함께 수정 될 수 있습니다.
초록
반응성 산소 종 (ROS)은 정상적인 발달, 항상성 및 생리학에서 중요한 잘 확립 된 신호 분자입니다. 상이한 ROS 중, 과산화수소(H2 O2)는세포 신호의 역할에 대하여 가장 잘 특징지었습니다. H2O2는 여러 종의 발달 중에 연루되었습니다. 예를 들어, H 2O2의일시적인 증가는 수정 후 첫 날 동안 제브라피시 배아에서 검출되었다. 더욱이, 중요한 세포H2O2 소스, NADPH 산화효소(NOX)를 고갈시키는 것은 생체내 및 체외 에서 모두 분화, 축축한 성장 및 망막 신경절 세포(RGC)의 지도와 같은 신경계 발달을손상시합니다. 여기서, 우리는 유전자 인코딩 된 H 2O 2-특정 바이오 센서, roGFP2-Orp1을 사용하여 개발 중에 배양 된 제브라피시 뉴런 및 전체 애벌레에서 세포 내 H2O2수준을 이미징하는 방법을 설명합니다. 이 프로브는 제브라피시 애벌레에서 일시적으로 또는 안정적으로 표현될 수 있다. 더욱이, 비율 측정 판독은 차등 유전자 발현 또는 부피 효과로 인해 유물을 검출할 확률을 감소시다. 첫째, 우리는 일시적으로 roGFP2-Orp1을 표현하는 제브라피시 배아에서 파생된 RG를 분리하고 배양하는 방법을 보여줍니다. 그런 다음 전체 애벌레를 사용하여 조직 수준에서 H2O2 수준을 모니터링합니다. 센서는 H 2O2의첨가에 의해 검증되었습니다. 추가적으로, 이 방법론은 조직 특정 biosensor 발현을 가진 형질전환 동물을 생성해서 특정 세포 모형 및 조직에 있는 H2O2 수준을 측정하기 위하여 이용될 수 있었습니다. Zebrafish가 유전 및 발달 조작을 용이하게 하기 때문에, 여기에서 입증된 접근은 척추동물에 있는 신경 및 일반적인 배아 발달 도중 H2O2의 역할을 시험하는 파이프라인 역할을 할 수 있었습니다.
서문
반응성 산소 종(ROS) 신호는 신경계1의개발 및 기능을 조절한다. 중요한 세포 ROS 공급원은 NADPH 산화효소(NOX)로, 이는 과산화수소와 과산화수소(H2O2)2를생성하는 막 단백질이다. NOX 효소는 중추 신경계(CNS)를 통해 발견되며, NOX 유래 ROS는 뉴런 개발에기여3,4,5,6. 신경 줄기 세포의 유지 보수 및 분화, 신경 극성 확립, 중성염 아웃성장 및 시냅스 가소성은 ROS7,8,9,10,11의적절한 수준을 요구하는 것으로 나타났다. 한편, NOXes에 의한 ROS의 통제되지 않은 생산은 알츠하이머병, 다발성 경화증 및 외상성 뇌 손상을 포함한 신경 퇴행성 질환에 기여한다12,13,14. 따라서 생리학적으로 관련된 ROS의 생산은 건강한 상태를 유지하는 데 중요합니다.
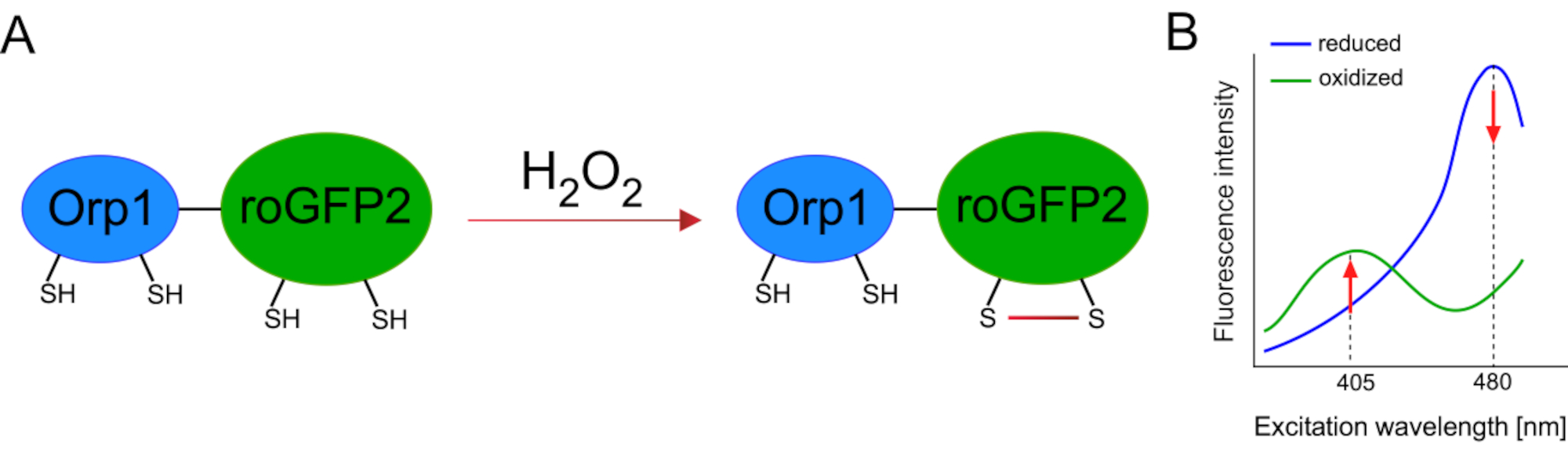
유전자 인코딩 된 바이오 센서의 개발은 세포 ROS의 검출을 크게 촉진시켰습니다. 유전자 인코딩 된 바이오 센서의 한 가지 중요한 장점은 이러한 센서가 특히 뚜렷한 위치를 대상으로 할 수 있기 때문에 ROS 신호의 증가 된 시간적 및 공간 해상도입니다. 레독스에 민감한 GFP(roGFP)는 이러한 ROS 바이오 센서의 한 유형입니다. roGFP2-Orp1 변이체는 효모15,16에서글루타티온 peroxiredoxin 가족 단백질인 Orp1 도메인을 통해 H2O2를 구체적으로 검출한다. Orp1 단백질의 산화는 그것의 형성을 바꾸기 위하여 roGFP2로 옮겨져(도 1A). 이 프로브는 405nm와 480nm 근처의 두 개의 여기 봉우리와 515 nm에서 단일 방출 피크를 전시합니다. 산화 시, 흥분 피크 주위 형광 강도 변경: 동안 405 nm 흥분 증가, 480 nm 흥분 감소. 따라서, roGFP2-Orp1은 비율인식 생체센서이며,H2O-2-레벨은두 개의 상이한 파장(도1B)에서흥분되는 형광 강도의 비율에 의해 검출된다. 전반적으로 roGFP2-Orp1은 생체 내에서효율적으로 활용할 수 있는 ROS 이미징을 위한 다목적 도구입니다.

그림 1: roGFP2-Orp1의 회로도 표현 및 여기 스펙트럼. (A)Oxidant 전송은 H 2 O2에대한 응답으로 Orp1과 roGFP2 사이에 발생하여 roGFP2의 형성적 변화를 초래한다. (B)roGFP2-Orp1의 흥분 스펙트럼은 405 nm와 480 nm및 515 nm에서 단일 방출 피크에서 두 개의 여기 피크를 나타낸다. H2O2에의해 산화시, 405 nm 흥분 증가 하는 동안 480 nm 흥분 감소. 이렇게 하면 H2 O2존재에 대한 비율 측정 판독이 발생합니다. 이 수치는 빌란과 벨로소프(2017)16및 모건 외(2011)25에서수정되었다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
Danio rerio (제브라피시) 모델 시스템은 유전자 인코딩 된 바이오 센서를 적용하는 데 몇 가지 장점이 있습니다. 배아와 애벌레의 광학적 투명성은 비침습적 생체 내 이미징을 가능하게 합니다. 새로운 이미징 도구는 더 높은 해상도와 더 깊은 침투를 달성하기 위해 개발되고있다17. 또한, 유전자 조작을 위한 확립된 도구(ectopic mRNA 발현, 톨2 전세대 등) 및 게놈 편집(TALENs, CRISPR/Cas9 등) 및 게놈 편집(TALEN, CRISPR/Cas9 등)이 있어 트랜스제닉 동물의 생성을 촉진하는18. 제브라피쉬 배아가 어머니 밖에서 발전함에 따라 이 시스템은 배아의 접근과 조작을 더욱 쉽게 할 수 있습니다. 예를 들어, 1 세포 단계 주사 및 약물 치료는 쉽게 할 수 있습니다.
여기서, 우리는 체외 전사 mRNA에 주입하여 H2O2-특정바이오 센서 roGFP2-Orp1을 일시적으로 표현하기 위해 제브라피쉬를 사용했습니다. 이러한 배아는 배양된 뉴런의 체외 이미징과 생체 내이미징(도 2)에 모두 사용될 수 있다. 우리는 제브라피시 배아에서 망막 신경절 세포 (RGC)를 해부하고 도금하기위한 프로토콜을 설명하고 배양 뉴런에서 H2O2-수준을 평가합니다. 그런 다음, 우리는 공초점 현미경 검사를 사용하여 roGFP2-Orp1 표현 배아 및 애벌레의 생체 내 이미징을 위한 방법을 제시합니다. 이 접근은 생리적인 H2O2-수준을 결정할 뿐만 아니라 다른 발달 단계 또는 조건에서 생기는 잠재적인 변경을 결정할 수 있습니다. 전반적으로, 이 시스템은 살아있는 세포및 동물에서H2O2를 검출하는 신뢰할 수 있는 방법을 제공하여 개발, 건강 및 질병에서 H 2 O2의역할을 연구한다.

그림 2. 실험적 접근 방식의 개요입니다. 간단히, 배아 수집 후, roGFP2-Orp1 mRNA는 1 세포 단계 제브라피시 배아의 노른자로 주입된다. 개발 배아는 생체 외에서 (A)및(B)생체 내 이미징 모두에 사용할 수 있습니다. (A)GFP 양성 배아는 34hpf에서 RGC 수집을 위한 망막을 해부하는 데 사용된다. 해리된 RG는 ZFCM(+) 미디어의 PDL/라미닌 코팅 커버립에 도금됩니다. RGCs가 도금 6-24 h 후 축축을 확장함에 따라 성장 콘 이미징을 수행할 수 있습니다. 세포는 H2O 2-level의 잠재적인 변화를 측정하기 위해 다른 치료를 받을 수 있습니다. 여기서, 우리는 RGC (빨간색)의 성장 콘에서 H2O2-수준을 측정했습니다. (B)GFP 양성 배아는 생체 내 이미징에 사용된다. 원하는 나이에, 태아는 공초점 화상 진찰을 위한 35 mm 유리 바닥 접시에 마취하고 장착될 수 있습니다. 여기서 배아는 망막 이미징을 위해 통풍구로 장착됩니다. 회로도는 제브라피시의 망막 발달을 보여줍니다. RGC는 망막에서 가장 안쪽 층인 신경절 세포 층(GCL)을 형성합니다. RGC 축축은 시신경으로 발전하여 중선을 넘어 시구를 형성합니다. 그런 다음 RGC 축축은 중뇌의 광학 텍텀에서 시냅스를 만들기 위해 등등으로 자랍니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
프로토콜
모든 동물 실험은 2020년 07/24/24에 승인된 프로토콜을 2006002050 가진 NIH 지침에 따라 Purdue 동물 관리 및 사용 위원회(PACUC)에 의해 윤리적으로 검토되고 승인되었습니다.
1. 솔루션 준비
- E2 미디어 (1x)
- 표1에 표시된 모든 구성 요소를 결합하여 100x E2A(500mL), 500x E2B(100mL) 및 500x E2C(100mL) 솔루션을 준비한다. 오토클레이브 E2A, E2B 및 E2C 솔루션. 4 °C에 보관하십시오.
- 1x E2 미디어의 경우: 100x E2A 5mL, 500x E2B 1mL, 500x E2C 1mL을 결합합니다. 부피를 멸균물로 500mL로 가져와보세요. pH를 7.0-7.5로 조정합니다.
- 장기 보관을 위해 -20°C에서 1x E2 미디어 스토어의 50mL 알리쿼트를 준비합니다. 그러나 해동 시 강수량이 발생할 수 있습니다. 스톡 솔루션을 사용하기 전에 강수량이 완전히 용해되었는지 확인하십시오.
| 용액 | 구성 요소 | 분량 | 농도 |
| 100X E2A (500mL) | |||
| 나Cl | 43.8g | 1500 mM | |
| KCl | 1.88 g | 50mM | |
| MgSO4 | 6 g | 100mM | |
| KH2PO4 | 1.03 g | 15 mM | |
| Na2HPO4 | 0.34 g | 5 mM | |
| 500X E2B (100mL) | |||
| 카Cl2 | 5.5 g | 500 mM | |
| 500X E2C (100mL) | |||
| 나코3 | 3 g | 350 mM | |
| 1X E2 (500mL) | |||
| 100X E2A | 5 mL | 1X | |
| 500X E2B | 1 mL | 1X | |
| 500X E2C | 1 mL | 1X |
표 1: 제브라피시 세포 배양을 위한 1x E2 배지의 성분.
- E3 미디어 (1x)
- 표 2에 표시된 대로 1L 멸균 물에 부품을 녹여 100배 의 재고를 만듭니다. 1x E3 미디어를 만들기 위해 멸균물에 육수를 희석합니다.
- 0.2% 메틸렌 블루를 추가합니다. 1x E3 미디어의 20mL의 경우 메틸렌 블루 40 μL을 추가합니다.
- 형광 화상 진찰을 위해 메틸렌 블루없이 또 다른 배치를 합니다.
| 구성 요소 | 양(g) | 100X 재고(mM)의 농도 |
| 나Cl | 29.22 | 500 |
| KCl | 1.26 | 17 |
| CaCl2 2H2O | 4.85 | 33 |
| MgSO4 7H2O | 8.13 | 33 |
표 2: 제브라피시 배아 유지를 위한 100x E3 매체의 성분.
- 80배 식염수 스톡 솔루션
- 표 3에표시된 모든 구성 요소를 결합합니다. 물을 추가하여 100mL 용액을 만듭니다. 모든 구성 요소가 용해될 때까지 섞습니다. 용액을 4°C로 저장합니다.
| 구성 요소 | 양(g) | 재고 농도 (mM) |
| 포도당 | 1.44 | 80 |
| 피루바테 나트륨 | 0.44 | 40 |
| CaCl2 2H2O | 0.148 | 10 |
| 헤페스 | 6.1 | 256 |
표 3: 제브라피시 세포 배양 매체용 80배 식염수 용액의 성분.
- 제브라피쉬 세포 배양 배지(ZFCM+)
- 표 4에 표시된 모든 구성 요소를 결합하여 250mL 미디어를 만듭니다. pH를 7.5로 조정합니다. 0.22 μm 필터를 사용하여 미디어를 필터링하고 4 °C에 저장합니다.
| 구성 요소 | 금액(mL) | 볼륨 % |
| L-15 미디엄(페놀 레드) | 212.75 | 85.1 |
| 태아 소 세럼 (FBS) | 5 | 2 |
| 페니실린/연쇄절제술 | 1 | 0.4 |
| 80X 식염수 솔루션 | 3.125 | 1.25 |
| 물 | 28.125 | 11.25 |
표 4: 세럼과 항생제를 함유한 제브라피쉬 세포 배양 배지의 성분.
- 이미징을 위한 제브라피쉬 세포 배양 배지(ZFCM-)
- 표 5에 표시된 모든 구성 요소를 결합하여 250mL 미디어를 만듭니다. pH를 7.5로 조정합니다. 0.22 μm 필터를 사용하여 미디어를 필터링합니다.
- 오염을 방지하기 위해 단일 사용 알리쿼트(50mL 배치)를 만듭니다. 4 °C에서 유지하십시오.
| 구성 요소 | 금액(mL) | 볼륨 % |
| L-15 미디엄(페놀 레드 없음) | 212.75 | 85.1 |
| 80X 식염수 솔루션 | 3.125 | 1.25 |
| 물 | 34.125 | 13.65 |
표 5: 체외 이미징용 혈청 및 항생제가 없는 제브라피시 세포 배양 배지의 성분.
- 사출 금형
- E3 미디어에서 1.5% 아가로즈를 녹입니다. 100 x 15mm 페트리 접시에 아가로즈 25mL를 붓습니다.
- 표면에 대하여 45° 각도로 아가로즈 위에 금형을 놓고 슬로우 모션으로 아가로즈 위에 떠 있게 하십시오. 이것은 거품을 피하는 데 도움이됩니다. 아가로즈를 식히고 벤치 탑에 굳어지게 하십시오.
- 완전히 고화되면, 아가로즈의 파손을 방지하기 위해 천천히 금형을 제거합니다. 신선한 E3 미디어를 추가하고, 파라핀 필름을 접시 주위에 추가하여 유출을 방지하고, 4°C(그림 3)에보관합니다.

그림 3: 사출 금형 이미지. (A)사출 판을 만드는 데 사용되는 플라스틱 금형. 금형에는 6개의 경사로, 1개의 90°및 1개의 45° 양면이 있습니다. (B)아가로즈 후 사출판이 고화되고 곰팡이가 제거된다. 스케일 바 = 1cm. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
2. roGFP2-Orp1 mRNA의 준비 및 주입
참고: roGFP2-Orp1 구조는 독일 DKFZ 의 토비아스 딕 박사로부터 수득되었습니다. 그것은 박사 청덩의 실험실에서 pCS2 + 벡터로 하위 복제되었다, 퍼듀 대학. RNase에 의한 분해를 방지하기 위해 몇 가지 예방 조치를 취해야 합니다. RNase 가약 및 튜브는 항상 사용되어야하며, 장갑은 모든 단계에 착용해야하며, 또는 RNase 제거를위한 세정제로 재료와 표면을 닦을 수 있습니다.
- NotI를 통해 pCS2+/roGFP2-Orp1 벡터의 3-10 μg를 선형화합니다.
- PCR 정리 키트로 선형화된 플라스미드를 정화합니다.
- 시험관 내 전사로 roGFP2-Orp1 mRNA에 체외 전사 키트가 제조업체가 제공한 지침에 따라 전사한다.
- 캡된 전사 반응 어셈블리
- RNA 폴리머라제와 선형화/정제 된 DNA를 얼음위에 놓습니다. 소용돌이 10 x 반응 버퍼와 2배 NTP/CAP가 완전히 용액이 될 때까지. NTP/CAP를 얼음에 저장하지만 반응을 조립하는 동안 상온(RT)에서 버퍼를 유지합니다. 오염을 방지하기 위해 튜브를 열기 전에 모든 시약을 터치 스핀합니다.
- RNase-free 0.5 mL 원심분리기 튜브에서 RT에서 아래 표시된 순서로 RNA 합성 반응을 설정합니다. 반응의 최종 부피는 20 μL. 반응 설정이 표 6에도시된다.
- 튜브에 필요한 경우 리보뉴클레오티드 믹스, 2배 NTP/CAP 및 뉴클레아제 프리 워터 10 μL을 추가합니다. 2 μL 10X 반응 버퍼를 추가합니다. 선형 DNA의 1-1.5 μg를 추가합니다(최대 6μL). 필요한 경우 핵이 없는 물을 추가하여 20 μL 반응 부피를 구성합니다.
- 튜브를 닫고, 소용돌이를 짧게 닫고, 마이크로퍼지를 터치스핀합니다. 10x SP6 효소 믹스의 2 μL을 추가합니다. 마이크로퍼지에서 손가락 클릭과 터치 스핀으로 닫습니다.
- 2-2.5 h (18 h까지 갈 수 있음)에 37 °C에 놓습니다.
참고: 제시된 실험에서 하룻밤 사이에 16-18h 인큐베이션에서 37°C에서 최상의 결과를 위해 수행하였다. - DNA 템플릿, 손가락 클릭, 터치 스핀 및 인큐베이션을 15 분 동안 37 °C에서 제거하기 위해 DNase 1 μL을 추가합니다.
- 캡된 전사 반응 어셈블리
| 시약 | 부피(μL) | 반응량 |
| NTP/캡 2배 | 10 | 1X |
| 10X 반응 버퍼 | 2 | 1X |
| 템플릿 DNA | 최대 6개 | 1-1.5 μg |
| 뉴클레아제 없는 물 | 20 μL을 만들기 위해 추가 | |
| 10X SP6 효소 믹스 | 2 | 1X |
표 6: 체외 전사에서 roGFP2-Orp1 mRNA에 대한 반응 설정.
- RNA 복구
- 시험관 내 전사 키트에 공급되는 염화물 리튬(LiCl)의 25μL을 추가합니다. -20°C에 비서리 냉동고에 30분 이상 놓습니다.
- 4°C에서 탁상 원심분리기에서 최대 속도로 25분 간 회전하십시오. 펠릿을 방해하지 않도록 상체를 조심스럽게 제거하고 폐기하십시오. 차가운 75% 에탄올 25 μL을 넣고 4°C에서 5분 동안 회전합니다.
- 신중하게 제거하고 상체를 폐기하십시오. RT에서 적어도 5 분 동안 펠릿 공기를 건조시키십시오. 건조하게 하지 마십시오. 뉴클레아제 프리 트라이-EDTA(TE) 버퍼(pH 7.0)의 12μL을 추가하고 샘플을 얼음 위에 보관합니다.
- 분광계로 RNA 농도를 측정합니다. 0.5- 1 μg/μL은 일반적으로 얻어진다.
- 페놀 레드 솔루션(덜벡코의 인산염 완충식염-DPBS에서 0.5% 페놀 레드)와 알리쿼트(3-5 μL)를 준비한다. - 80 °C에서 mRNA 알리쿼트저장.
- mRNA의 미세 주입
- 주사 당일, mRNA 알리쿼트 중 하나를 사용하고 제브라피쉬 배아 주입 프로토콜을 따라 노른자19를통해 1세포 단계 배아에 mRNA 1nL을 주입한다. 간략한 설명은 아래에 있습니다.
- 이전에 설명된 바와 같이 성인 물고기를 사육하고 배아를수집20.
- 파이펫 풀러로 미세 주입 바늘을 당깁니다. 10 μm 팁 개구부를 만들기 위해 집게로 바늘의 끝을 잘라.
- 1.6단계에서 설명된 사출 몰드에 배아를 정렬합니다.
- 유리 미세 주입 파이펫으로 페놀 레드에 mRNA 1 nL을 주입합니다.
- 배아를 수집하고 E3 미디어에 보관하십시오.
- 원하는 발달 단계가 달성될 때까지 E3 매체에서 27°C 인큐베이터에 배아를 보관하십시오. 주입된 배아는 체외(섹션 3 및 섹션 4) 및 생체 내 이미징(섹션 5)에 모두 사용할 수 있다. roGFP2-Orp1을 발현하는 배아는 형광광과 청색/녹색 필터 세트를 갖춘 정기적인 해부 현미경으로 실험하기 전에 미리 선택할 수 있다.
3. 제브라피시 배아에서 유래한 1차 망막 신경절 세포 배양
참고: 이 프로토콜은 이전에 게시된 메서드(21)에서적용됩니다. 라미나르 플로우 후드에서 3.1 및 3.2 단계를 수행합니다.
- 커버립 준비
- 각 실험에서 4-6 배양 판을 준비합니다. 100% 에탄올에 보관되는 산성 세척 커버립(22 x 22mm 정사각형, 0.16-0.19mm 두께)을 사용합니다.
- 집게를 사용하여 보관 용기에서 덮개 슬립 1개를 제거하고 화염을 제거하여 잔류 에탄올을 제거합니다.
- 35mm 배양 접시 안에 비스듬히 배치하여 커버슬립을 완전히 건조시합니다.
- 멸균 물에 10배 스톡(5 mg/mL)을 희석하여 폴리-D-Lysine(PDL) 작업 용액(1x)을 준비합니다.
- 각 커버슬립의 중앙에 0.5 mg/mL PDL의 100 μL을 적용하고 가장자리에 용액이 확산되는 것을 피하십시오.
- 실온(RT)에서 20-30분 동안 커버립에 PDL을 배양합니다. PDL이 건조되지 않았는지 확인합니다.
- PDL을 0.5mL 멸균물로 세 번 세척합니다. 접시를 완전히 건조시키십시오.
- 1x PBS에서 50배 스톡(1 mg/mL)을 희석하여 라미닌 작업 용액(1x)을 준비합니다.
- PBS에 20 μg/mL 라미닌의 100 μL을 각 커버슬립의 중앙에 적용하고 가장자리에 용액이 확산되는 것을 피하십시오.
- 2-6 h에 대한 가습 인큐베이터에서 플레이트를 37 °C에서 배양한다. 라미닌 용액의 건조를 피하십시오.
- 배아 해부 및 도금 RG
- 35mm 조직 배양 식기 4개에 35mm의 접시를 준비하고 라벨을 부착하고 4mL로 채우세요: 70% 에탄올, "E2 미디어 1", "E2 미디어 2", "E2 미디어 3" 해부 당일. 해부될 때까지 냉장고에 접시를 보관하십시오.
- 제브라피쉬 배아는 34시간 후 수정(hpf)인 경우, 인큐베이터에서 라미닌으로 코팅된 배양 요리를 1x PBS0.5mL로 세 번 씻어낸다.
- 마지막 세척 후 각 배양 접시에 ZFCM(+) 배지 4mL을 추가하고 접시건조를 피하십시오.
- 3.2.1 단계에서 준비된 문화 요리를 검색합니다. 그들이 RT에 평형하자.
- ZFCM(+) 매체의 15μL로 4-6PCR 튜브를 채웁니다. 하나의 커버슬립에 도금하기 위해 4 눈에서 RG를 준비하는 데 하나의 튜브가 필요합니다.
- 인큐베이터에서 제브라피쉬 배아를 회수하고 배아를 5-10s에 70% 함유하는 35mm 조직 배양 접시에 배아를 담급하여 살균합니다.
- 이송 파이펫을 사용하여 배아를 멸균 E2 매체를 함유한 E2 Media 1 접시로 이송하여 과도한 에탄올을 세척합니다.
- 배아를 E2 Media 2 접시로 옮기고 날카로운 집게로 축을 제거합니다.
- 배아를 최종 E2 Media 3 접시로 이송하여 해부를 수행합니다.
- 미세 한 쌍의 집게를 사용하여, 이전에 설명 된 바와 같이 망막을 해부 22.
- 포셉 중 하나를 사용하여 배아를 노른자에 배치하고 다른 포셉으로 노른자 낭에 꼬리 후방을 제거합니다.
- 집게로 목을 잡고 머리를 떼어 뇌와 눈을 E2 미디어에 노출시하십시오. 노른자 주머니를 자르지 마십시오.
- 미세 한 집게의 끝으로, 부드럽게 두 번째 집게와 함께 두개골 조직을 잡고, 머리에서 눈을 굴려. 인접한 조직 파편으로부터 눈을 분리하십시오.
- ZFCM(+)이 포함된 이전에 준비된 튜브 중 하나에 4개의 눈을 전달합니다.
- P20 파이펫과 노란 팁으로 약 45번 위아래로 부드럽게 적어 세포를 분리합니다. 기포를 피하십시오.
- 분리된 셀을 커버슬립의 중심으로 ZFCM(+)을 전송합니다. 추가 커버립을 위해 10-12 단계를 반복합니다.
- 진동을 흡수하기 위해 폴리스티렌 폼 랙에서 22°C의 벤치탑에서 배양을 유지합니다.
- 도금 후 6-24h 이미징을 수행합니다.
참고: 배아를 다른 배양 요리로 옮기기 위해 이송 파이펫을 사용합니다. 각 솔루션의 파이펫을 변경하여 에탄올(6-8단계)을 운반하지 못하게 합니다.
4. 배양 된 RGC 뉴런의 체외 ROS 이미징
- 이미징 의 날에 (일반적으로 6-24 세포 도금 후 h), RGC 축축의 성장을 검증 하기 위해 현미경에서 세포를 확인.
- 라이브 셀 이미징의 경우, 커버립을 문화 접시에서 라이브 세포 이미징 챔버로 옮기. 이 경우 이전에 설명된 맞춤형 열린 챔버가23을사용하였더라.
- 이미징을 위한 현미경 설정. 차동 간섭 콘트라스트(DIC) 목표, OG590 롱패스 레드 필터 및 EM-CCD 카메라를 장착한 반전 현미경을 사용합니다.
- 이미징하기 전에 ZFCM(+) 매체를 ZFCM(-)으로 바꿉니다.
- 세포가 10배 의 목표로 배치되면 높은 NA 오일 침지 목표를 사용하여 60배배율로 이미지를 획득합니다. 추가 1.5 배 의 배율을 사용합니다.
- 첫째, DIC 이미지를 획득합니다. 그런 다음 적절한 필터 세트를 사용하여 이미지 roGFP2-Orp1을 사용한다. 405/20 및 480/30 nm 난여기 필터를 장착한 roGFP2-Orp1을 순차적으로 발췌하고 방출 등이 이색 거울 505DCXR를 통과한 후 535/30 nm 방출 필터로 이미지를 획득한다.
- 이미지의 첫 번째 세트를 촬영 한 후, 다른 치료 솔루션을 포함하는 미디어와 미디어를 교환. 미디어는 pH 및 진동 변화를 피하기 위해 이미징의 매 30 분마다 변경해야합니다.
5. 배아 발달의 생체 내 ROS 이미징
- 생체 내 이미징의 경우, 22-24 hpf에서 메틸렌 블루없이 0.003 % 페닐티오레아 (PTU)를 포함하는 E3 매체에서 배아를 유지합니다. 미디어를 교환하고 매일 죽은 배아를 제거하십시오.
- 원하는 나이에, 0.016% 트리카인에 있는 태아를 마취합니다. 35mm 유리 바닥 배양 접시에 1% 저용 아가로즈에 마취 배아를 마운트. 배아는 화상 진찰을 위한 관심의 지구에 따라서, 통풍창 또는 측면으로, 등대, 환기 또는 측면에 지향될 수 있습니다.
- 아가로즈가 고성화된 후, 메틸렌 블루/0.016% 트리카인없이 E3 미디어로 요리를 채우게 하십시오.
- 이미징을 위한 현미경을 설정합니다. 반전 된 레이저 스캐닝 공초점 현미경을 사용합니다. 또는, 아가로즈 드롭 위에 장착 된 배아를 이미지에 물 침지 렌즈가 장착 된 똑바로 공초점 현미경을 사용합니다.
- 405nm 및 488 nm 난관 필터를 순차적으로 흥분시키고 515-535 nm 범위에서 방출 필터로 해당 이미지를 획득합니다.
- 배아의 원하는 부분을 통해 5 μm 단면 두께로 z 스택을 획득합니다. 배아는 발달의 후반 단계에서 화상 진찰을 위해 보관될 수 있습니다.
- 이미징 후 미세한 집게로 아가로즈에서 배아를 제거하고 PTU를 사용하여 메틸렌 블루 프리 미디어에서 원하는 나이가 될 때까지 인큐베이터에 보관하십시오.
6. 이미지 분석 및 처리
- 405/480 비율 값을 기준으로 H2O2 레벨 측정
- 이미지 분석에 적합한 소프트웨어를 사용합니다. ImageJ 소프트웨어는 이미지 분석 및 처리를 위해 여기에 사용되었습니다.
- 파일을 드래그하거나 파일을 클릭하여 ImageJ 소프트웨어에서 DIC, 405/535 및 480/535 이미지를 | 열립니다. 아직 완료되지 않은 경우 이미지를 클릭하여 이미지를 32비트로 변환| | 유형 32 비트.
- 제어 바(세포체, 성장 콘, 망막 등)로부터 무료 핸드 툴로 관심 영역(ROI)을 정의합니다. 분석 | 클릭하여 ROI 관리자를 엽니다. 도구 | ROI 매니저. ROI 관리자 탭에서 추가를 클릭하여 정의된 ROI를 추가합니다.
- ROI에 가까운 영역을 그리고 배경 ROI로 추가합니다. ROI를 선택하고 ROI 관리자 탭에서 측정값을 클릭하여 평균 배경 값을 측정합니다.
- 측정값의 평균 강도 값을 기록합니다. 프로세스 | 클릭하여 형광 이미지에서 평균 배경값을 뺍니다. 수학 | 빼기. 405/535 및 480/535 이미지모두에 대해 이 단계를 수행합니다.
- 프로세스 | 클릭하여 "0" 값을 제거하기 위해 480/535 형광 이미지에 "1"의 값을 추가합니다. 수학 | 비율 계산 에 앞서 함수를 추가합니다.
- 프로세스 | 클릭 이미지 계산기 | ImageJ에서 함수를 분할하여 405/535 이미지를 480/535 이미지 픽셀단위로 나눕니다. 480/535 이미지로 나눌 405/535 이미지를 선택합니다. 32비트 출력 이미지를 선택합니다.
- ROI 관리자 탭에서 먼저 비율 이미지를 클릭한 다음 ROI 관리자 탭에서 ROI대 비율 이미지를 적용합니다.
- ROI 관리자 탭에서 측정값을 클릭하여 405/535 이미지의 평균 비율 값을 480/535 이미지로 측정합니다.
- 적절한 통계 분석을 수행하기 위해 가능한 한 많은 샘플에 대해 6.1.2-6.1.9 단계를 수행하십시오.
- 비율 이미지 표시
참고: 이 절차는 표본 외부의 배경을 빼고 이미지에 색상 조회 테이블을 적용하는 것입니다.- 6.1.7단계에서 ImageJ에서 비율 이미지가 만들어지면 파일 | 클릭하여 32비트 검은색 이미지를 만듭니다. 새로운 | 이미지.
- 먼저 새 이미지를 클릭한 다음 ROI 관리자 탭에서 ROI를 클릭하여 H2O2 레벨을 새 이미지에 표시하려는 ROI를 적용합니다.
- 편집 | 클릭하여 마스크 만들기 선택 | 마스크 만들기.
- 마스크 이미지를 255로 나누어 ROI 값을 "1"으로 조정하고 배경 값은 "0"이 됩니다. 프로세스 | 클릭 수학 | 분할 및 입력 255.
- 프로세스 | 클릭하여 비율 이미지와 함께 마스크를 곱합니다. 이미지 계산기 | 기능을 곱합니다. 이렇게 하면 ROI만 표시하는 회색 배율 이미지가 생성됩니다.
- 이미지 | 클릭하여 조회 테이블을 "불"으로 변경합니다. 테이블 | 조회 화재.
참고: 프로세스 | 클릭하여 비율을 더 잘 시각화하기 위해 곱셈 계수를 모든 이미지에 적용할 수 있습니다. 이미지 | 곱합니다. - 이미지 | 클릭하여 비율 이미지를 8비트로 변환합니다. | 유형 8 비트.
- 분석 | 클릭하여 교정 표시줄 추가 도구 | 교정 표시줄.
결과
배양 된 제브라피시 RG는 1d 내에서 축축을 확장합니다. H2 O2-바이오센서의대표적인 405/480 비율 이미지가 도 4A에도시된다. 세포 몸, 축 사, 그리고 성장 콘은 명확 하 게 개별 뉴런에서 볼 수 있습니다. 이러한 뉴런은 H2O2 변화를 모니터링하기 위해 시간이 지남에 따라 다른 치료를 받을 수 있습니다. 앞서 문화매체에 100μMH2 O2를첨가하?...
토론
이 프로토콜 전체에 주의를 기울여야 하는 몇 가지 중요한 단계가 있습니다. 우리는 이러한 점을 고려하면 실험 흐름을 향상시킬 것이라고 믿습니다. 1 차적인 RGC 배양에 대 한, ZFCM의 멸균 (-) 매우 중요 한, 이 배양 매체 항생제를 포함 하지 않기 때문에 오염 이미징 전후 또는 동안 발생할 수 있습니다. 이를 방지하기 위해 생물 안전 캐비닛 내에서만 ZFCM(-)을 준비하고 사용하는 것이 좋습니다. 또?...
공개
저자는 이해 상충이 없다고 선언합니다.
감사의 말
이 작품은 국립 보건원 (그랜트 R01NS117701), 국립 과학 재단 (그랜트 1146944-IOS), 인디애나 외상척수및 뇌 손상 연구 기금 (그랜트 20000289), Purdue 연구 재단 (그랜트 209911), Purdue 대학의 연구 및 파트너십 담당 수석 부사장 (그랜트 210362)에 의해 지원되었다. 우리는 제브라피시 RGC 문화 프로토콜을 수립한 코리 J. 위버 박사와 헤일리 로더 박사에게 감사드립니다. 그림 4의 데이터를 제공해 주신 헤일리 로더에게 감사드립니다. 우리는 RGC 문화에 대한 도움레아 바이어시와 케니 응우옌에게 감사드립니다. 글편집에 감사드립니다. 우리는 roGFP2-Orp1 및 roGFP2-Orp1을 포함하는 pCS2+ 벡터에 대한 박사 칭 덩을 제공 토비아스 딕 박사에게 감사드립니다. 그림 2는 Biorender.com 함께 만들어집니다.
자료
| Name | Company | Catalog Number | Comments |
| 35-mm culture dish | Sarstedt | 83-3900 | |
| 35-mm glass bottom dish | MatTek | P35G-1.5-10-C | |
| Agarose | Fisher Scientific | BP160-500 | |
| Borosilicate Glass Capillary Tubes | Sutter/Fisher Scientific | NC9029378 | |
| Calcium Chloride Dihydrate | Fisher Scientific | C79-500 | |
| Cover glass | Corning | 2850-22 | |
| Disposable Petri Dishes (100 x 15 mm) | VWR | 25384-094 | |
| Fetal Bovine Serum | ThermoFisher Scientific | 26140087 | |
| Glucose | Sigma Aldich | G7528 | |
| HEPES | Sigma Aldich | H4034 | |
| Injection Mold | Adaptive Science Tools | TU-1 | |
| Inverted Microscope | Nikon | TE2000 | |
| Laminin | ThermoFisher Scientific | 23017-015 | |
| Laser Scanning Confocal Microscopy | Zeiss | 710 | |
| Leibovitz's L-15 Medium with phenol red | Gibco/Fisher Scientific | 11-415-064 | |
| Leibovitz's L-15 Medium without phenol red | Gibco/Fisher Scientific | 21-083-027 | |
| Low melting agarose | Promega | V2111 | |
| mMessage mMachine SP6 Transcription Kit | Invitrogen | AM1340 | |
| NotI | New England Biolabs | R0189S | |
| PBS | Hyclone/Fisher Scientific | SH3025601 | |
| Penicillin/streptomycin | ThermoFisher Scientific | 15140122 | |
| Phenol Red | Sigma Aldich | P0290 | |
| Phenylthiourea (PTU) | Sigma Aldich | P7629 | |
| Pneumatic PicoPump | World Precision Instruments | PV820 | |
| Poly-D-Lysine (PDL) | Sigma Aldich | P7280 | |
| QiaQUICK PCR Purification Kit | QIAGEN | 28104 | |
| Recombinant mouse slit2 | R&D Systems | 5444-SL-050 | |
| Sodium Pyruvate | Sigma Aldich | P5280 | |
| Steritop 0.22 µm filter | Millipore | S2GPT05RE | |
| TE Buffer | Ambion | AM9860 | |
| Tricaine Methanesulfonate | Sigma Aldich | E10521 | |
| Vertical Pipette Puller | David Kopf Instruments | 700C |
참고문헌
- Bórquez, D. A., et al. Dissecting the role of redox signaling in neuronal development. Journal of Neurochemistry. 137 (4), 506-517 (2016).
- Bedard, K., Krause, K. -. H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiological Reviews. 87 (1), 245-313 (2007).
- Weaver, C. J., Leung, Y. F., Suter, D. M. Expression dynamics of NADPH oxidases during early zebrafish development. Journal of Comparative Neurology. 524 (10), 2130-2141 (2016).
- Terzi, A., Suter, D. M. The role of NADPH oxidases in neuronal development. Free Radical Biology and Medicine. 154, 33-47 (2020).
- Infanger, D. W., Sharma, R. V., Davisson, R. L. NADPH oxidases of the brain: Distribution, regulation, and function. Antioxidants & Redox Signaling. 8 (9-10), 1583-1596 (2006).
- Coyoy, A., Olguin-Albuerne, M., Martinez-Briseno, P., Moran, J. Role of reactive oxygen species and NADPH-oxidase in the development of rat cerebellum. Neurochemistry International. 62 (7), 998-1011 (2013).
- Le Belle, J. E., et al. Proliferative neural stem cells have high endogenous ROS levels that regulate self-renewal and neurogenesis in a PI3K/Akt-dependant manner. Cell Stem Cell. 8 (1), 59-71 (2011).
- Nayernia, Z., et al. Decreased neural precursor cell pool in NADPH oxidase 2-deficiency: from mouse brain to neural differentiation of patient derived iPSC. Redox Biology. 13, 82-93 (2017).
- Wilson, C., Nunez, M. T., González-Billault, C. Contribution of NADPH oxidase to the establishment of hippocampal neuronal polarity in culture. Journal of Cell Science. 128 (16), 2989-2995 (2015).
- Munnamalai, V., et al. Bidirectional interactions between Nox2-type NADPH oxidase and the F-actin cytoskeleton in neuronal growth cones. Journal of Neurochemistry. 130 (4), 526-540 (2014).
- Kishida, K. T., et al. Synaptic plasticity deficits and mild memory impairments in mouse models of chronic granulomatous disease. Molecular and Cellular Biology. 26 (15), 5908-5920 (2006).
- Ravelli, K. G., et al. Nox2-dependent neuroinflammation in an EAE model of multiple sclerosis. Translational Neuroscience. 10 (1), 1-9 (2019).
- Park, L., et al. Nox2-derived radicals contribute to neurovascular and behavioral dysfunction in mice overexpressing the amyloid precursor protein. Proceedings of the National Academy of Sciences. 105 (4), 1347-1352 (2008).
- Schiavone, S., Neri, M., Trabace, L., Turillazzi, E. The NADPH oxidase NOX2 mediates loss of parvalbumin interneurons in traumatic brain injury: Human autoptic immunohistochemical evidence. Scientific Reports. 7 (1), 8752 (2017).
- Gutscher, M., et al. Proximity-based protein thiol oxidation by H2O2-scavenging peroxidases. Journal of Biological Chemistry. 284 (46), 31532-31540 (2009).
- Bilan, D. S., Belousov, V. V. New tools for redox biology: from imaging to manipulation. Free Radical Biology and Medicine. 109, 167-188 (2016).
- Abu-Siniyeh, A., Al-Zyoud, W. Highlights on selected microscopy techniques to study zebrafish developmental biology. Laboratory Animal Research. 36 (1), 12 (2020).
- Sassen, W. A., Koster, R. W. A molecular toolbox for genetic manipulation of zebrafish. Advances in Genomics and Genetics. 5, 151-163 (2015).
- Rosen, J. N., Sweeney, M. F., Mably, J. D. Microinjection of zebrafish embryos to analyze gene function. Journal of Visualized Experiments. (25), e1115 (2009).
- Avdesh, A., et al. Regular care and maintenance of a Zebrafish (Danio rerio) laboratory: An introduction. Journal of Visualized Experiments. (69), e4196 (2012).
- Chen, Z., et al. Primary neuron culture for nerve growth and axon guidance studies in Zebrafish (Danio rerio). PLoS One. 8 (3), 57539 (2013).
- Zhang, L., Leung, Y. F. Microdissection of zebrafish embryonic eye tissues. Journal of Visualized Experiments. (40), e2028 (2010).
- Suter, D. M. Live cell imaging of neuronal growth cone motility and duidance in vitro. Cell Migration: Methods in Molecular Biology. , 65-86 (2011).
- Weaver, C. J., et al. nox2/cybb deficiency affects zebrafish retinotectal connectivity. Journal of Neuroscience. 38 (26), 5854-5871 (2018).
- Morgan, B., Sobotta, M. C., Dick, T. P. Measuring EGSH and H2O2 with roGFP2-based redox probes. Free Radical Biology and Medicine. 51, 1943-1951 (2011).
- Li, Z., et al. Phenylthiourea specifically reduces zebrafish eye size. PLoS One. 7 (6), 40132 (2012).
- Ermakova, Y. G., et al. Red fluorescent genetically encoded indicator for intracellular hydrogen peroxide. Nature Communications. 5, 5222 (2014).
- Oparka, M., et al. Quantifying ROS levels using CM-H2DCFDA and HyPer. Methods. 109, 3-11 (2016).
- Dickinson, B. C., Peltier, J., Stone, D., Schaffer, D. V., Chang, C. J. Nox2 redox signaling maintains essential cell populations in the brain. Nature Chemical Biology. 7 (2), 106-112 (2011).
- Cannon, M. B., Remington, S. J. Redox-sensitive green fluorescent protein: Probes for dynamic intracellular redox responses. A review. Methods in Molecular Biology. 476, 51-65 (2009).
- Meyer, A. J., Dick, T. P. Fluorescent protein-based redox probes. Antioxidants & Redox Signaling. 13 (5), 621-650 (2010).
- Panieri, E., Millia, C., Santoro, M. M. Real-time quantification of subcellular H2O2 and glutathione redox potential in living cardiovascular tissues. Free Radical Biology and Medicine. 109, 189-200 (2017).
- Breus, O., Dickmeis, T. Genetically encoded thiol redox-sensors in the zebrafish model: Lessons for embryonic development and regeneration. Biological Chemistry. , (2020).
- Morgan, B., et al. Real-time monitoring of basal H2O2 levels with peroxiredoxin-based probes. Nature Chemical Biology. 12 (6), 437-443 (2016).
- Terzi, A., Roeder, H., Weaver, C. J., Suter, D. M. Neuronal NADPH oxidase 2 regulates growth cone guidance downstream of slit2/robo2. Developmental Neurobiology. , (2020).
- Bilan, D. S., Belousov, V. V. HyPer family probes: State of the art. Antioxidants & Redox Signaling. 24 (13), 731-751 (2016).
- Ermankova, Y. G., et al. SypHer3s: a genetically encoded fluorescent ratiometric probe with enhanced brightness and an improved dynamic range. Chemistry Communications. 54 (23), 2898-2901 (2018).
- Pak, V. V., et al. Ultrasensitive genetically encoded indicator for hydrogen peroxide identifies roles for the oxidant in cell migration and mitochondrial function. Cell Metabolism. 31 (3), 642-653 (2020).
- Kwan, K. M., et al. The Tol2kit: A multisite gateway-based construction Kit for Tol2 transposon transgenesis constructs. Developmental Dynamics. 236 (11), 3088-3099 (2007).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기더 많은 기사 탐색
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유