
Method Article
Cristallisation ABCG5/G8 dans un environnement bicellaire lipidique pour la cristallographie aux rayons X
Dans cet article
Résumé
Ce protocole décrit une configuration pour la cristallisation du transporteur de stérols ABCG5/G8. ABCG5/G8 est reconstitué en bicelles pour la cristallisation en gouttes suspendues. Le protocole ne nécessite pas de matériaux ou de substrats spécialisés, ce qui le rend accessible et facile à adapter dans n’importe quel laboratoire pour déterminer la structure de la protéine par cristallographie aux rayons X.
Résumé
Les transporteurs de cassettes de liaison à l’ATP (ABC) constituent des protéines membranaires enrobées de lipides. L’extraction de ces protéines membranaires de la bicouche lipidique vers un environnement aqueux est généralement réalisée à l’aide de détergents. Ces détergents désintègrent la bicouche lipidique et solubilisent les protéines. L’habitat intrinsèque des protéines membranaires au sein de la bicouche lipidique pose un défi dans le maintien de leur stabilité et de leur uniformité en solution pour la caractérisation structurale. Les bicelles, qui comprennent un mélange de phospholipides et de détergents à chaîne longue et courte, reproduisent la structure lipidique naturelle. L’utilisation de bicelles lipidiques et de détergents sert de système modèle approprié pour obtenir des cristaux de diffraction de haute qualité, en particulier pour déterminer la structure à haute résolution des protéines membranaires. Grâce à ces microenvironnements synthétiques, les protéines membranaires préservent leur conformation et leur fonctionnalité natives, facilitant la formation de cristaux tridimensionnels. Dans cette approche, l’hétérodimérique ABCG5/G8 solubilisé par le détergent a été réintégré dans les bicelles DMPC/CHAPSO, complété par du cholestérol. Cette configuration a été utilisée dans la procédure expérimentale de diffusion de vapeur pour la cristallisation des protéines.
Introduction
Les transporteurs de cassettes de liaison à l’ATP (ABC) constituent une superfamille de protéines membranaires responsables de divers processus de transport dépendants de l’ATP à travers les membranes biologiques 1,2,3,4,5. Ces protéines transporteuses sont impliquées dans les maladies cardiovasculaires et jouent un rôle important dans la facilitation de l’efflux de cholestérol vers la bile pour une excrétion ultérieure dans le foie. Par conséquent, le métabolisme et l’équilibre du cholestérol ont suscité un intérêt considérable au fil des ans6. Un mécanisme spécifique impliqué dans l’élimination du cholestérol et d’autres stérols de l’organisme implique des membres de la sous-famille des ABCG humains, notamment l’ABCG5/G8 hétérodimérique 7,8,9,10. Des mutations dans l’un ou l’autre de ces gènes perturbent l’hétérodimère, entraînant une perte de fonction et provoquant une sitostérolémie, un trouble affectant le trafic des stérols11,12,13. Compte tenu de la pertinence de la maladie et de leur rôle dans la promotion de l’efflux de cholestérol, les transporteurs de stérols ont attiré une attention importante. Néanmoins, les détails complexes de leur mécanisme moléculaire et de leur sélectivité du substrat restent largement non divulgués. Ainsi, l’élucidation de la structure cristalline d’ABCG5/G8 est une étape cruciale vers la compréhension des mécanismes et des fonctions en aval du transport du cholestérol.
Les protéines membranaires ont besoin d’un ancrage à l’intérieur des membranes pour se replier et fonctionner correctement. Par conséquent, l’extraction des protéines membranaires de leur environnement naturel entraîne souvent une instabilité des protéines, un mauvais repliement et une perte de fonction14,15. Ces défis soulignent les principaux obstacles rencontrés dans la cristallisation des protéines membranaires. Cependant, la reconstitution des protéines en bicouches de détergents synthétiques, comme les bicelles, est apparue comme une solution à cette situation difficile, permettant le maintien des protéines membranaires dans un milieu de bicouche de type natif16. Les bicelles sont des assemblages de phospholipides et de détergents synthétiques en suspension et solubilisés dans l’eau. Notamment, ils adoptent une structure bicouche qui imite les membranes biologiques16,17,18. Les bicelles peuvent passer de la phase liquide à la phase gel en fonction de la température et de la viscosité. La cristallisation bicellaire tire parti des petits disques bicouches et de la faible viscosité à des températures réduites, ce qui facilite le mélange complet des protéines et des solutions bicellaires. La taille des bicelles dépend du rapport détergent/lipide lors de la préparation19,20. Les détergents les plus répandus pour la formation de bicelles comprennent le 3-[(3-cholamidopropyl)diméthylammonio]-2-hydroxy-1-propanesulfonate (CHAPSO), ainsi que le 3-[(3-cholamidopropyl)diméthylammonio]-1-propanesulfonate (CHAPS) et le 1,2-ditridecanoyl-sn-glycérol-3-phosphocholine (DHPC)21. Ces détergents sont utilisés en conjonction avec des lipides tels que la di-myristoyl-phosphatidylcholine (DMPC) et la 1-palmitoyl-2-oléoyl-phosphatidylcholine (POPC). De plus, des études récentes ont démontré la pleine fonctionnalité des protéines membranaires dans les bicelles dans des conditions physiologiques. Par exemple, Lee et ses collègues ont réussi à cristalliser et à rapporter la structure cristalline d’ABCG5/ABCG8 sur la base d’une bicouche lipidique22,23. Dans le processus de cristallisation, les mélanges protéine-bicelle peuvent être adaptés à l’aide d’équipements standard, y compris des robots de cristallisation à haut débit24. La faisabilité de l’utilisation des bicelles, cependant, dépend de la thermostabilité des protéines en raison des conditions de cristallisation à des températures plus élevées. Néanmoins, par rapport à d’autres techniques, les conditions de cristallisation requises pour les protéines membranaires restent généralement douces, impliquant de faibles concentrations de précipitant, de sel et de tampon. Cela rend à la fois les mélanges protéine-bicellaire et la diffusion de vapeur des outils efficaces et faciles à mettre en œuvre pour les études structurales des protéines membranaires.
Ce protocole décrit les étapes essentielles de la préparation des protéines et de la cristallisation bicellaire pour déterminer la structure cristalline en rayons X d’ABCG5/G8 à haute résolution (Figure 1).
Protocole
1. Clonage et expression des protéines
- Cloner le gène humain ABCG5/G8 dans la levure Pichia pastoris en suivant les protocoles précédents25,26. Brièvement, dérivez les vecteurs d’expression pSGP18 et pLIC à partir de pPICZB. Ajoutez une balise qui code pour un site de protéase 3C de rhinovirus suivi d’un peptide de liaison à la calmoduline (CBP) à l’extrémité C de l’ADNc ABCG8 (pSGP18-G8-3C-CBP).
- Ajouter une étiquette à six histidines séparée par une glycine (His 6 GlyHis6) à l’extrémité C de l’ADNc ABCG5 (pLIC-G5-H12). Co-transformer les plasmides en souche KM71H de Pichia par électroporation.
REMARQUE : Veuillez consulter le tableau des matériaux pour plus de détails sur les plasmides, les milieux et les tampons utilisés. - Cultiver des cellules de levure transformées sur des plaques de gélose MD à 28 °C.
- Ajouter une étiquette à six histidines séparée par une glycine (His 6 GlyHis6) à l’extrémité C de l’ADNc ABCG5 (pLIC-G5-H12). Co-transformer les plasmides en souche KM71H de Pichia par électroporation.
- Après 1 à 2 jours, sélectionnez 10 à 12 colonies et inoculez-les dans 10 mL de milieu à base azotée de levure (MGY) à base de glycérol minimal à l’aide de tubes à centrifuger de 50 ml pour la culture à petite échelle.
- Faites trois petits trous dans le couvercle du tube de centrifugation pour une meilleure aération. Laisser les cellules se développer à 28 °C avec des secousses constantes à 250 tr/min jusqu’à ce que la densité optique à 600 nm (OD600) atteigne 10, ce qui prend généralement 1 à 2 nuits.
REMARQUE : La croissance cellulaire prend généralement entre 12 et 24 heures.
- Faites trois petits trous dans le couvercle du tube de centrifugation pour une meilleure aération. Laisser les cellules se développer à 28 °C avec des secousses constantes à 250 tr/min jusqu’à ce que la densité optique à 600 nm (OD600) atteigne 10, ce qui prend généralement 1 à 2 nuits.
- Le lendemain, prélever 1 L de milieu MGY stérile et l’inoculer avec la culture primaire dans un flacon de 2,4 L. Incuber le ballon à 28 °C dans un incubateur à agitateur à 250 tr/min pendant 24 h.
- Pour maintenir le pH entre 5 et 6, ajoutez 10 % d’hydroxyde d’ammonium (NH4OH) jusqu’à ce que le pH se stabilise.
- Ajustez le pH et induisez l’expression des protéines en ajoutant 1 mL de méthanol pur par culture de 1 L (0,1 % (v/v) de méthanol).
REMARQUE : Alimenter ensuite les cellules avec 5 mL de méthanol pur par litre de culture (0,5 % (v/v) de méthanol) toutes les 12 h pendant une durée totale de 36 à 48 h. - Récolter les cellules par centrifugation à 15 000 x g pendant 30 min à 4 °C.
- Recueillir les pastilles cellulaires et les remettre en suspension dans un tampon de lyse (0,33 M de saccharose, 0,3 M de Tris-Cl pH 7,5, 0,1 M d’acide aminohexanoïque, 1 mM d’EDTA et 1 mM d’EGTA) à une concentration de 0,5 g/mL. Conserver la suspension à -80 °C. En règle générale, on peut récupérer 30 ± 5 g de masse cellulaire à partir de 1 L de cellules en culture.
REMARQUE : Conservez les granulés cellulaires directement dans le congélateur ou effectuez une remise en suspension immédiate dans un tampon de lyse pour les préparations membranaires.
2. Préparation de la membrane microsomale
- Décongeler les cellules et ajouter des inhibiteurs de la protéase (2 μg/mL de leupeptine, 2 μg/mL de pepstatine A, 2 mM de PMSF, voir le tableau des matériaux).
- Pour lyser davantage les cellules, utilisez un émulsifiant ou un microfluidiseur refroidi à l’eau glacée (voir le tableau des matériaux) à 25 000-30 000 psi. Répétez ce processus 3 à 4 fois.
- Centrifugeuse pour éliminer les débris cellulaires : essorage à 3 500-4 000 x g pendant 15 min, suivi d’un deuxième essorage à 15 000 x g pendant 30 min. Maintenez les deux essors à 4 °C.
- Pour isoler les vésicules membranaires microsomales, transférer le surnageant dans des tubes à ultracentrifuger et le soumettre à une ultracentrifugation à 2 00 000 x g pendant 1,5 h à 4 °C.
- Remettre en suspension la pastille membranaire dans 50 mL de tampon A (50 mM de Tris-Cl pH 8,0, 100 mM de NaCl et 10 % de glycérol) à l’aide d’un homogénéisateur à dounce. Conserver la suspension à -80 °C.
3. Préparation-purification des hétérodimères des protéines
- Décongeler les membranes microsomiques congelées et ajuster la concentration à 4-6 mg/mL à l’aide d’un tampon de solubilisation. Le tampon doit contenir 50 mM de Tris-HCl, pH 8,0, 100 mM de NaCl, 10 % de glycérol, 1 % (p/v) de β-dodécyl maltoside (β-DDM), 0,5 % (p/v) de cholate, 0,1 % (p/v) d’hémisuccinate de cholestéryle (CHS), 5 mM d’imidazole, 5 mM de β-mercaptoéthanol (β-ME), 2 μg/mL de leupeptine, 2 μg/mL de pepstatine A et 2 mM de PMSF (voir le tableau des matériaux).
REMARQUE : On peut mélanger des volumes égaux de la préparation membranaire et du tampon de solubilisation, ou utiliser un tampon de solubilisation 2x sans inhibiteurs de protéase ni agents réducteurs. L’ébullition courte du tampon aide à dissoudre efficacement le SHC. Pour la purification, utilisez uniquement le tampon refroidi à 4 °C.- Remuez le mélange à vitesse moyenne pendant 1 h à 4 °C. Continuez ensuite à remuer à température ambiante (RT) pendant 20 à 30 minutes.
- Centrifuger le mélange à 1,00,000 x g pendant 30 min à 4 °C pour éliminer les membranes insolubles. Prélever le surnageant solubilisé et ajouter 20 mM d’imidazole et 0,1 mM de TCEP.
- Effectuer la chromatographie sur colonne d’affinité26 : lier le surnageant solubilisé à des billes de Ni-NTA pré-équilibrées (10-15 mL) (voir le tableau des matériaux) dans le tampon A (étape 2.2.1) pendant la nuit.
REMARQUE : Évitez d’utiliser du glycérol dans les tampons d’exécution à partir de ce point.- Laver la colonne deux fois avec 10 volumes de colonne de tampon B (50 mM HEPES, pH 7,5, 100 mM NaCl, 0,1 % (p/v) β-DDM, 0,05 % (p/v) cholate, 0,01 % (p/v) CHS, 0,1 mM TCEP) contenant 25 mM d’imidazole.
- Laver la colonne avec 10 volumes de colonne de tampon B contenant 50 mM d’imidazole.
- Éluer la protéine à l’aide du tampon C (tampon B avec 200 mM d’imidazole).
- Ajouter 1 mM de TCEP (voir le tableau des matériaux) et 10 mM de MgCl2 aux protéines éluées.
- Validez les fractions éluées sur un gel SDS-PAGE pour confirmer la taille correcte de la protéine26.
REMARQUE : Rendement protéique typique (1 er Ni-NTA) : 10à 20 mg de protéines par culture de 6 L. Utiliser le DDM à 10 ou 5 fois sa concentration micellaire critique (CMC), soit environ 0,01 %. Ce protocole utilise 0,1 % de DDM. - Diluer les fractions maximales de l’élution Ni-NTA avec un volume égal de tampon D1 (tampon B avec 1 mM de CaCl 2, 1 mM de MgCl2), mélanger et charger les fractions protéiques sur une colonne CBP (3-5 mL) (voir tableau des matériaux) qui a été pré-équilibrée avec le tampon de lavage CBP D1.
- Effectuer des lavages séquentiels sur la colonne CBP pour échanger les détergents à l’aide des tampons D1 et D2 (tampon B avec 1 mM de CaCl 2, 1 mM de MgCl2, 0,1 % (p/v) de décyl-maltose néopentyl glycol (DMNG), sans β-DDM) : tout d’abord, laver avec 3 volumes de colonne de D1 ; deuxièmement, laver avec 3 volumes de colonne de D1 :D2 (3 :1, v/v) ; troisièmement, laver avec 3 volumes de colonne de D1 :D2 (1 :1, v/v) ; quatrième étape, laver avec 3 volumes de colonne de D1 :D2 (1 :3, v/v), suivi de 6 à 10 volumes de colonne de D2.
- Éluer la protéine à l’aide du tampon de lavage CBP D2 avec 300 mM de NaCl en fractions de 1 mL à partir de la colonne CBP (total 10 mL). Concentrer les fractions éluées à 1-2 mL.
REMARQUE : Le rendement protéique typique (1 er CBP) est de 5à 15 mg de protéines par culture de 6 L. Les détergents à base de maltose néopentyl glycol (MNG) améliorent le stockage des protéines purifiées à 4 °C. Le DMNG et le Lauryl MNG (LMNG) ont tous deux été utilisés, le DMNG produisant de meilleurs cristaux diffractant les rayons X. Utilisez le DMNG à 10-20 fois sa concentration micellaire critique (CMC), soit environ 0,003 %. Ce protocole utilisait 0,1 % de DMNG. Une fraction de l’éluat de CBP peut être purifiée par chromatographie par filtration sur gel (étape 4.4.) afin d’analyser l’activité ATPase des protéines ou d’évaluer la monodispersité par microscopie électronique à transmission (MET).
4. Traitement de préparation des protéines - pré-cristallisation
- Cliver les glycanes liés à l’azote et les marqueurs CBP à l’aide de l’endoglycosidase H (Endo H, ~0,2 mg par 10-15 mg de protéine purifiée) et de la protéase HRV-3C (~2 mg par 10-15 mg de protéine purifiée), respectivement (voir le tableau des matériaux). Incuber toute la nuit à 4 °C.
- Pendant l’incubation des protéases Endo H et 3C, effectuer une alkylation réductrice sur les protéines regroupées. Commencer par incuber avec 20 mM d’iodoacétamide (voir tableau des matériaux) pendant une nuit à 4 °C. Poursuivez avec une incubation de 1 h avec 2 mM supplémentaires d’iodoacétamide sur glace.
REMARQUE : Cette étape stabilise davantage le stockage des protéines jusqu’à un mois à 4 °C. - Utilisez une deuxième colonne CBP (1 à 2 mL) pour séparer l’étiquette CBP clivée. Utilisez le tampon D2 pour ce processus.
REMARQUE : Le rendement typique en protéines (2 e CBP) est de 5à 10 mg de protéines par culture de 6 L. - Purifier la protéine sans marqueur CBP à l’aide de la chromatographie par filtration sur gel. Le tampon doit contenir 10 mM de HEPES, pH 7,5, 100 mM de NaCl, 0,1 % (p/v) de DMNG, 0,05 % (p/v) de cholate et 0,01 % (p/v) de CHS.
REMARQUE : Le rendement typique en protéines (filtration sur gel) est de 2 à 8 mg de protéines par culture de 6 L. Au cours de cette étape, l’absence d’un pic de DDM (~65 kD) lors de la filtration sur gel indique un échange réussi de détergent en DMNG. - Modifier les fractions protéiques regroupées par méthylation réductrice : ajouter 20 mM de diméthylamine borane (DMAB, voir Tableau des matériaux) et 40 mM de formaldéhyde à la protéine. Incuber pendant 2 h à 4 °C sur un agitateur oscillant. Ajoutez 10 mM de DMAB.
- Répétez l’étape 4.5, y compris l’ajout de 10 mM de DMAB, et incubez pendant la nuit (12-18 h) à 4 °C.
- Arrêtez la réaction en ajoutant 100 mM de Tris-Cl, pH 7,5.
- Chargez la protéine méthylée sur une colonne de Ni-NTA de 2 mL prééquilibrée avec 100 mM de Tris-Cl, pH 8,0 et 100 mM de NaCl.
- Laver la colonne à l’aide de 10 volumes de tampon de lavage (10 mM HEPES, pH 7,5, 100 mM NaCl, avec 0,5 mg/mL DE DOPC : DOPE (3 :1, p/p), 0,1 % (p/v) de DMNG, 0,05 % (p/v) de cholate, 0,01 % (p/v) de CHS).
- Éluer la protéine relipidée à l’aide d’un tampon d’élution (10 mM HEPES, pH 7,5, 100 mM NaCl, 200 mM d’imidazole, 0,5 mg/mL DOPC : DOPE (1 :1, p/p), 0,1 % (p/v) DMNG, 0,05 % (p/v) cholate, 0,01 % (p/v) CHS).
REMARQUE : Le rendement typique en protéines (2e Ni-NTA) est de 1 à 5 mg de protéines par culture de 6 L. - Faire passer les éluats de protéines à travers une colonne de dessalage-10 pré-équilibrée avec le tampon utilisé à l’étape 4.4.
- Incuber la protéine dessalée et relipidée pendant une nuit avec du cholestérol (préparé dans de l’isopropanol ou de l’éthanol) jusqu’à une concentration finale de ~20 μM.
- Le lendemain matin, retirer le précipitant par ultracentrifugation à 1,50 000 x g pendant 10 min à 4 °C. Récupérez le surnageant.
- Concentrer la protéine jusqu’à une concentration finale de 30 à 50 mg/mL à l’aide d’un concentrateur centrifuge de coupure de 100 kDa.
- Retirer le précipitant à l’aide d’une centrifugeuse réfrigérée de paillasse à la vitesse maximale pendant 30 min à 4 °C.
- Maintenir le surnageant sur la glace à 4 °C et établir les conditions de cristallisation dans les bicelles.
REMARQUE : Les protéines concentrées doivent être utilisées pour la croissance des cristaux dans un délai d’une semaine. Ne congelez pas les protéines.
5. Cristallisation des protéines dans les bicelles
- Préparer une solution mère bicellaire à 10 % avec des lipides DMPC et un détergent CHAPSO dans un rapport de 3 :1 (p/p) (voir le tableau des matériaux).
REMARQUE : Utiliser CHAPSO à 5 fois sa concentration micellaire critique (CMC), soit environ 0,5 %. Cela permet de maintenir la concentration de détergent autour de son CMC dans le mélange protéine-bice (étape 5.2).- Ajouter du détergent dissous H2O déminéralisé (CHAPSO) aux lipides pré-séchés (mélange de 5 % mol de cholestérol et de 95 mol de DMPC).
REMARQUE : Préparez diverses compositions lipidiques dans le chloroforme et séchez-les dans un tube à essai en verre à l’aide d’un flux d’azote gazeux à RT. Éliminez les solvants résiduels en les plaçant dans une chambre à vide pendant la nuit, formant une fine couche lipidique. - Remettre en suspension les lipides et le détergent à l’aide d’un sonicateur au bain-marie.
- Sonicer le mélange bicellaire dans de l’eau glacée en utilisant une puissance continue jusqu’à ce que la solution devienne transparente.
REMARQUE : Utilisez une protection auditive et assurez-vous d’une alimentation suffisante en glace pour maintenir le mélange en phase liquide. - Éliminez les composants non dissous à l’aide d’un filtre centrifuge de 0,2 μm (voir le tableau des matériaux).
REMARQUE : Conserver la solution bicellaire aliquotenée à -80 °C.
- Ajouter du détergent dissous H2O déminéralisé (CHAPSO) aux lipides pré-séchés (mélange de 5 % mol de cholestérol et de 95 mol de DMPC).
- Créer un mélange de protéines et de bicelles sur de la glace en combinant délicatement 10 % de bicelles (étape 5.1.4) et de protéines (étape 4.7.4) dans un rapport de 1 :4 (v/v), jusqu’à obtenir une concentration finale de protéines entre 5 et 10 mg/mL.
- Incuber le mélange de protéines et de bicelles sur de la glace pendant 30 min.
- Mettre en place des conditions de cristallisation dans un format de diffusion de vapeur de goutte suspendue à l’aide de plaques à 48 puits.
- Mélanger des volumes égaux (0,5 ou 1 μL) de mélange protéine/bicellaire et une solution de réservoir de cristallisation contenant 1,6 M-2,0 M de sulfate d’ammonium, 100 mM de MES (pH 6,5), 0 % à 4 % de PEG 400 et 1 mM de TCEP (voir le tableau des matériaux).
REMARQUE : Créer une matrice de la solution du réservoir avant chaque expérience, en ajustant le sulfate d’ammonium (1,6-2,0 M) et le PEG 400 (0 %-4 %). - Incuber pour cristallisation à 20 °C.
- Vérifiez les plateaux de cristallisation le lendemain pour vous assurer que le verre de couverture est bien scellé.
- Surveillez la croissance des cristaux au moins une fois par jour. Les cristaux de haute qualité apparaissent généralement dans les 1 à 2 semaines, mesurant 50-150 μm x 20-50 μm x 2-5 μm.
REMARQUE : Les cristaux peuvent prendre plus de temps à se former à des concentrations de protéines plus faibles. Les cristaux matures doivent être récoltés dans un délai d’un mois. - Trempez les cristaux de protéines dans du malonate de sodium de 0,2 M et congelez-les dans de l’azote liquide à l’aide de cryo-boucles de 50 ou 100 μm.
REMARQUE : Si un diffractomètre à rayons X est disponible, testez quelques cristaux avec une exposition de 15 à 30 min au faisceau de rayons X pour révéler une diffraction jusqu’à 5 Å. La diffraction à plus haute résolution nécessite une source de lumière synchrotron. En utilisant du malonate de sodium de 0,2 M comme cryoprotecteur, un cristal mesurant 100 μm x 50 μm x 2 μm peut fournir environ 90 images de diffraction avec des rayons X synchrotron.
- Mélanger des volumes égaux (0,5 ou 1 μL) de mélange protéine/bicellaire et une solution de réservoir de cristallisation contenant 1,6 M-2,0 M de sulfate d’ammonium, 100 mM de MES (pH 6,5), 0 % à 4 % de PEG 400 et 1 mM de TCEP (voir le tableau des matériaux).
Résultats
Les demi-transporteurs ABC recombinants, ABCG5 et ABCG8 humains, sont co-exprimés dans la levure Pichia pastoris . La fraction membranaire de la levure est ensuite fractionnée par centrifugation. Comme indiqué dans ce protocole, les protéines hétérodimériques sont extraites par chromatographie sur colonne tandem. Par la suite, les protéines prétraitées chimiquement sont cristallisées en les incubant avec des bicelles phospholipides/cholestérol. Des aperçus schématiques des procédés de purification et de cristallisation sont présentés à la figure 1.
Pour évaluer la monodispersité des protéines purifiées, des échantillons contenant 0,01 à 0,05 mg/mL de protéines sont colorés avec de l’acétate d’uranyle de 1 % à 2 %. Ces échantillons sont ensuite examinés à l’aide d’un MET à coloration négative (figure 2A). Afin d’évaluer la stabilité des protéines sans subir de cycles de congélation-décongélation, la chromatographie analytique par filtration sur gel est utilisée. Cette analyse consiste à surveiller l’entreposage dans le temps des protéines purifiées à l’aide de petites aliquotes de volume égal (Figure 2B). Il peut y avoir une légère perte de protéines aux fractions maximales après une semaine d’incubation à 4 °C, peut-être en raison d’agrégats de protéines solubles résiduelles. Néanmoins, le rendement global en protéines reste suffisant pour la croissance des cristaux. L’utilisation de la MET à coloration négative et de la chromatographie analytique par filtration sur gel est une pratique courante pour évaluer l’aptitude des protéines à la cristallisation, en particulier à partir de différentes constructions techniques.
Pour l’évaluation de la qualité des protéines à chaque étape du processus de chromatographie sur colonne, ainsi qu’après le traitement chimique de pré-cristallisation, des aliquotes de fractions correspondant à deux colonnes Ni-NTA, deux colonnes CBP, une filtration sur gel et une alkylation réductrice sont chargées sur un gel SDS-PAGE à 10 % (Figure 3). De plus, le même environnement réactionnel que celui utilisé pour l’alkylation peut être appliqué pour le marquage du mercure avec de l’éthylmercure (EMTS), bien que cela dépasse le cadre de la présente étude.
La croissance des cristaux est surveillée quotidiennement à l’aide d’un stéréomicroscope de table équipé d’un polariseur. Les cristaux matures et adaptés à la collecte de données atteignent généralement des dimensions de 50 μm x 100 μm x 2 μm (Figure 4). Pendant le processus de récolte des cristaux, les cristaux ou les amas plus petits sont délibérément évités.

Figure 1 : Aperçus schématiques de la purification (A) et de la cristallisation bicellaire (B) de l’ABCG5/G8 hétérodimérique. Les constructions d’ABCG5 (hG5) et d’ABCG8 (hG8) humains recombinants portent respectivement les marqueurs RGS-H 6-G-H6 et 3C-CBP (A, en haut). Chromatographie sur colonne d’affinité en tandem, suivie d’une chromatographie par filtration sur gel pour obtenir une purification hétérodimérique (A, en bas). Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 2 : Évaluation de la mono-dispersité (A) et de la stabilité (B) des protéines purifiées. (A) Micrographie électronique d’hétérodimères ABCG5/G8 (G5G8) colorés négativement à l’aide de la MET. Les particules représentatives sont mises en évidence par des cercles blancs pleins. Barre d’échelle = 100 nm. (B) Protéines alkylées stockées à 4 °C analysées par chromatographie analytique sur filtration sur gel pendant un mois avec une légère perte de protéines au bout d’une semaine. Veuillez cliquer ici pour voir une version agrandie de cette figure.
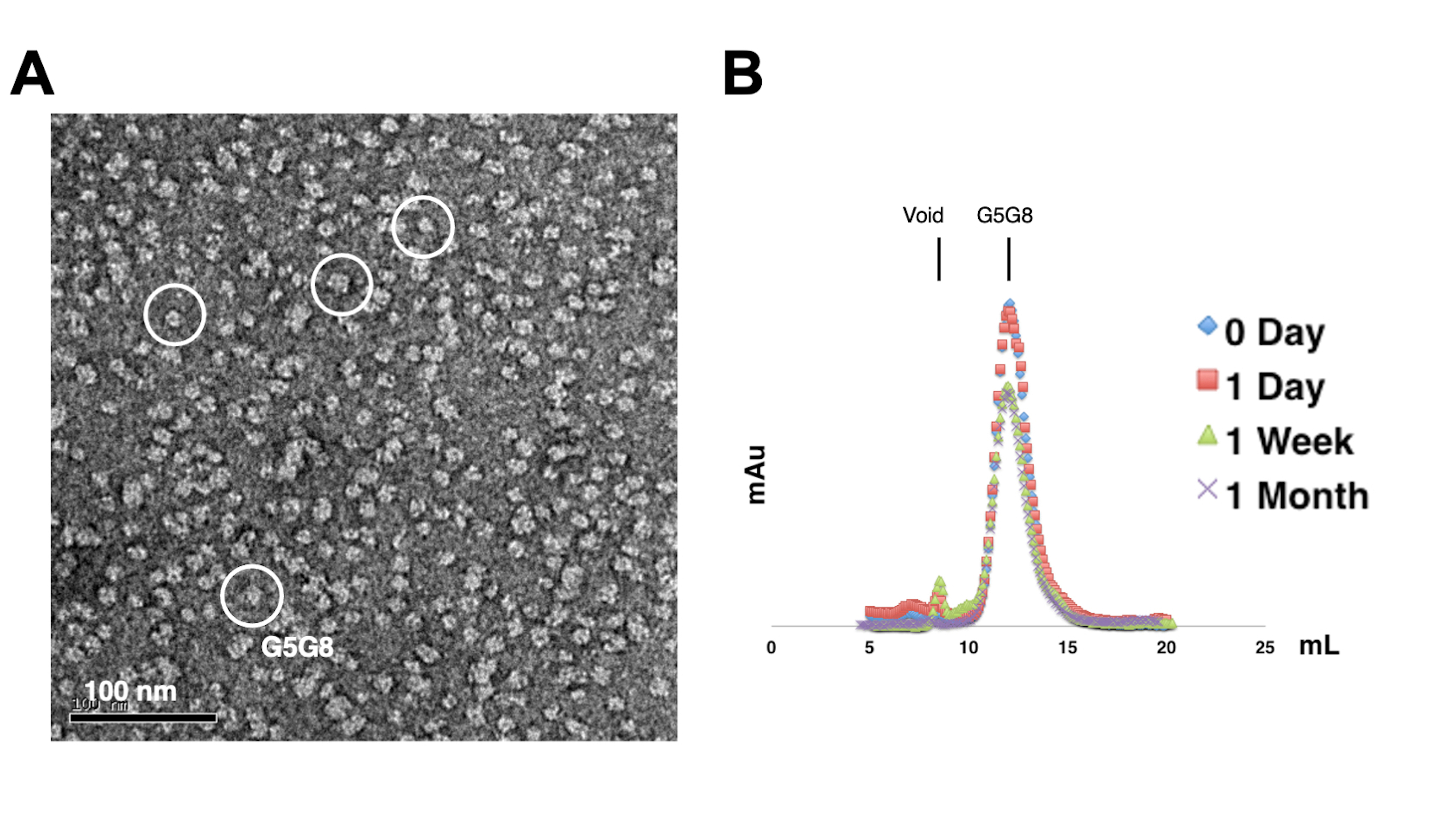{kind=link}

Figure 3 : Analyse SDS-PAGE des éluats protéiques de la chromatographie sur colonne et de l’alkylation réductrice. Différents volumes (1-10 μL) de fractions protéiques ont été chargés sur un gel Tris/Glycine à 10 % et ont fonctionné pendant 45 min à une tension constante de 200 V. Le gel a été coloré avec du bleu de Coomassie, décoloré, séché à l’air et scanné par un scanner de table. 1° & 2° Ni : première et deuxième colonnes Ni-NTA ; 1° & 2° CBP : première et deuxième colonnes CBP ; Ligne continue des fractions de crête : fractions regroupées pour la cristallisation ; Ligne pointillée des fractions maximales : fractions d’épaule ; EMTS : thiosalicynate d’éthylmercure ; IA : iodoacédamide. Veuillez cliquer ici pour voir une version agrandie de cette figure.
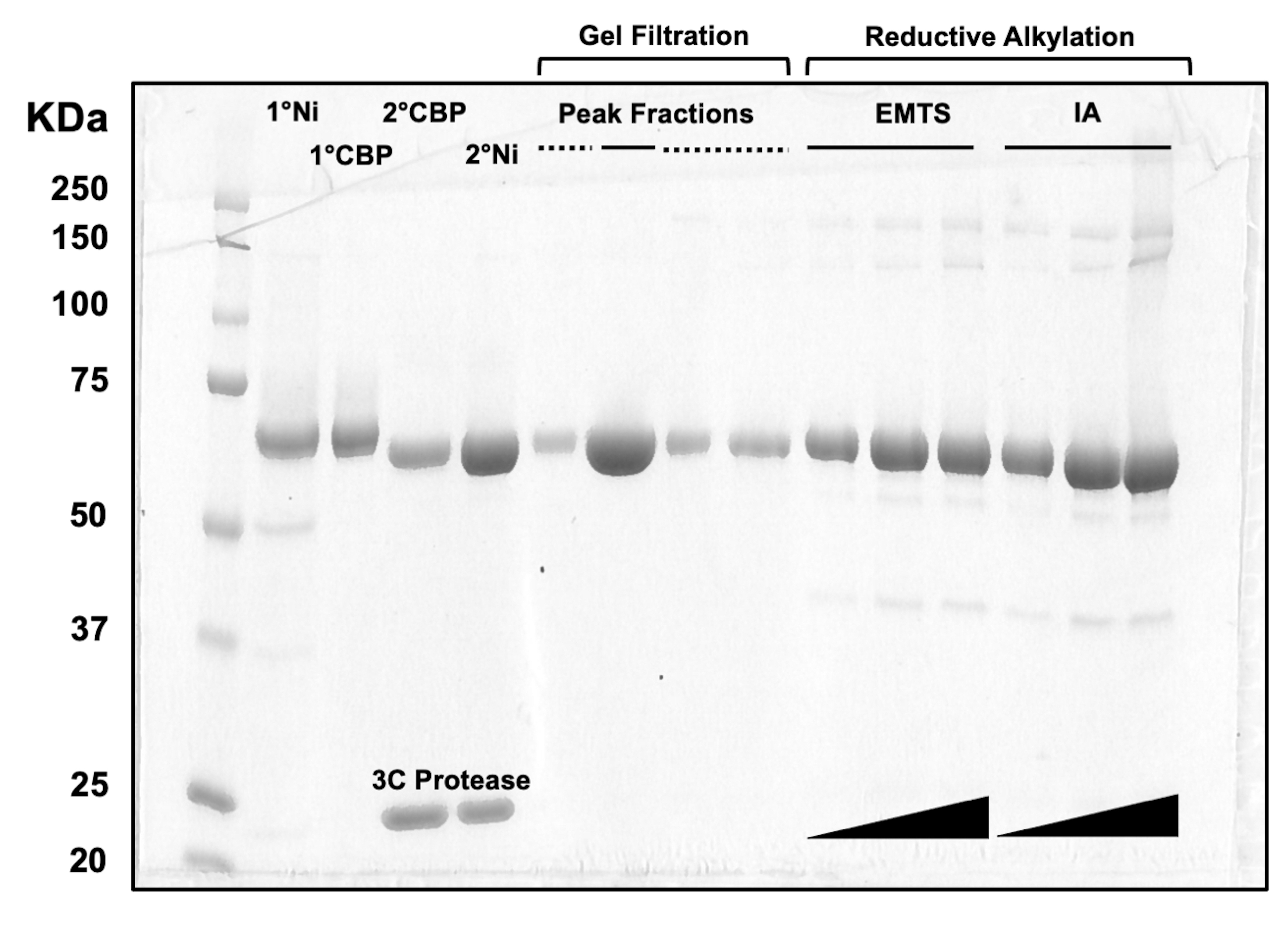{kind=link}

Figure 4 : Évaluation de la maturation des cristaux de protéines par microscopie optique. Des cristaux matures d’ABCG5/G8 provenant d’une goutte de cristallisation ont été visualisés sous un stéréomicroscope de table et équipé d’un polariseur. Barre d’échelle = 100 μm. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Discussion
Les défis associés à la cristallisation des protéines membranaires ont incité à la mise au point de méthodes de cristallisation basées sur les bicouches lipidiques, telles que les approches bicellaires27 ou en phase cubique lipidique (LCP)14 . Cependant, la réalisation d’une cristallisation réussie des protéines membranaires dépend toujours de l’étape critique et parfois goulot d’étranglement de la préparation des protéines. Notamment, les transporteurs ABC représentent un obstacle redoutable dans la croissance de cristaux adaptés à la cristallographie aux rayons X. Ce protocole fournit des conseils pratiques complets pour rationaliser la préparation du transporteur de stérols ABCG5/G8 humain et favoriser la croissance cristalline grâce à l’approche de cristallisation bicellaire.
L’un des principaux facteurs à prendre en compte lors de l’élaboration de ce protocole était l’impératif d’un rendement protéique substantiel dans les phases initiales de la purification des protéines, ce qui permet un certain degré de perte de protéines pendant le traitement de pré-cristallisation (Figure 3). Les stratégies courantes pour relever ce défi impliquent une ingénierie protéique approfondie, l’utilisation d’hôtes d’expression diversifiés et l’exploration d’orthologues ou d’homologues, entre autres approches. Néanmoins, avec cette procédure apparemment complexe, un certain nombre d’étapes cruciales ont été identifiées qui sous-tendent le succès du protocole et fournissent également des informations sur les limitations potentielles qui peuvent survenir lors de l’étude d’autres transporteurs ABC ou des protéines membranaires en général.
Tout d’abord, ce protocole utilise une centrifugation approfondie à chaque étape pour minimiser l’agrégation des protéines. De plus, une surveillance continue de la thermostabilité des protéines purifiées est cruciale. La microscopie électronique est utilisée pour vérifier la monodispersité des protéines, tandis que la filtration analytique sur gel permet de suivre la stabilité des protéines au fil du temps (Figure 2). Des techniques alternatives telles que le dichroïsme circulaire (CD) ou la calorimétrie différentielle à balayage (DSC) pourraient également être intégrées. De plus, l’incorporation de lipides à des stades spécifiques est essentielle pour maximiser à la fois l’activité et la cristallogenèse de l’ABCG5/G8 purifié. Par exemple, le cholate et le CHS sont nécessaires pour présenter une hydrolyse mesurable de l’ATP ; les phospholipides sont indispensables au maintien de la stabilité des protéines méthylées ; et le cholestérol est un composant nécessaire de la solution bicellaire, favorisant la croissance cristalline adaptée à la diffraction des rayons X à haute résolution (Figure 4).
Essentiellement, l’ensemble de la procédure peut être accompli en une semaine d’effort. Contrairement au LCP, la récupération des cristaux à partir de plateaux de cristallisation à gouttes suspendues est simple. À l’avenir, avec un rendement protéique substantiel (environ 10 mg), ce protocole est facilement adaptable pour le développement d’études cristallographiques impliquant des mutants ABCG5/G8 ou d’autres protéines de transport. Ceci est particulièrement pertinent pour les cas qui échappent actuellement à la visualisation par microscopie électronique.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
Ces travaux sont financés par une subvention à la découverte du Conseil de recherches en sciences naturelles et en génie du Canada (RGPIN 2018-04070) et une subvention de projet des Instituts de recherche en santé du Canada (PJT-180640) à JYL. Ce protocole est basé sur les rapports originaux sur les structures cristallines ABCG5/G8 rapportés précédemment par Farhat et al.22 et Lee et al.23.
matériels
| Name | Company | Catalog Number | Comments |
| ABCG5 | National Institute of Health collection | NCBI accession number NM_022436 | |
| ABCG8 | National Institute of Health collection | NCBI accession number NM_022437 | |
| ÄKTA FPLC system | Cytiva (formerly GE Healthcare Life Sciences) | ||
| CaCl2 | Wisent | 600-024-CG | Anhydrous |
| CBP | Agilent | 214303 | Calmodulin binding peptide affinity resin |
| Centrifugal concentrators (Vivaspin) | Sartorius | ||
| CHAPSO | Anatrace | C317 | Anagrade |
| Cholesterol | Anatrace | CH200 | |
| CHS | Steraloids | C6823-000 | |
| DMAB | MilliporeSigma | 180238 | 97% |
| DMNG | Anatrace | NG322 | |
| DMPC | Anatrace | D514 | |
| DOPC | Avanti | 850375 | |
| DOPC | Anatrace | D518 | |
| DOPE | Avanti | 850725 | |
| DTT | Fisher | BP172 | |
| Dual Thickness MicroLoops | MiTeGen | ||
| EDTA | BioShop | EDT003 | Disodium salt, dihydrate |
| EGTA | MilliporeSigma | 324626 | |
| Emulsifier (EmulsiFex-C3) | Avestin | ||
| Endo H | New England Biolabs | P0702 | |
| Ethanol | Greenfield | P016EAAN | Ethyl Alcohol Anhydrous |
| Formaldehyde | MilliporeSigma | 252549 | ACS Reagent |
| Glycerol | BioShop | GLY004 | |
| HEPES | BioShop | HEP001 | |
| HRV-3C protease | Homemade | ||
| Imidazole | BioShop | IMD510 | Reagent grade |
| Iodoacetamide | MilliporeSigma | I1149 | BioUltra |
| Isopropanol | Fisher | BP2618212 | |
| Leupeptin | BioShop | LEU001 | |
| MES | MilliporeSigma | 69892 | BioUltra |
| Methanol | Fisher | A412P | |
| MgCl2 | Wisent | 800-070-CG | Hydrated |
| microfluidizer (LM 20) | Microfluidics | ||
| NaCl | BioShop | SOD002 | |
| NH4OH | Fisher | A669-212 | ACS Reagent |
| Ni-NTA superflow | Qiagen | 30430 | Nickel-charged resins |
| PEG 400 | MilliporeSigma | 202398 | |
| Pepstatin | BioShop | PEP605 | |
| PMSF | MilliporeSigma | P7626 | |
| pSGP18 and pLIC | Homemade (derived from pPICZ, Invitrogen) | ||
| SDS | BioShop | SDS003 | |
| Sodium cholate | Fisher | 229101 | |
| Sodium malonate | MilliporeSigma | 63409 | |
| Sucrose | Wisent | 800-081-WG | Ultra pure |
| Superdex 200 30/100 GL | Cytiva (formerly GE Healthcare Life Sciences) | 28990944 | Prepacked gel-filtration column |
| TCEP | |||
| TEM | FEI, Technai | ||
| Tris Base | Fisher | BP152 | |
| β-DDM | Anatrace | D310S | Sol Grade |
| β-mercaptoethanol | MilliporeSigma | ||
| ε-aminocaproic acid | Fisher | AAA1471936 |
Références
- Hamada, H., Tsuruo, T. Purification of the 170- to 180-kilodalton membrane glycoprotein associated with multidrug resistance. 170- to 180-kilodalton membrane glycoprotein is an ATPase. Journal of Biological Chemistry. 263 (3), 1454-1458 (1988).
- Higgins, F., Hiles, D., Whalley, K., Jamieson, J. Nucleotide binding by membrane components of bacterial periplasmic binding protein-dependent transport systems. The EMBO Journal. 4 (4), 1033-1039 (1985).
- Higgins, F., et al. A family of related ATP-binding subunits coupled to many distinct biological processes in bacteria. Nature. 323 (6087), 448-450 (1986).
- Horio, M., Gottesman, M., Pastan, I. ATP-dependent transport of vinblastine in vesicles from human multidrug-resistant cells. Proceedings of the National Academy of Sciences of the United States of America. 85 (10), 3580-3584 (1988).
- Mimmack, L., et al. Energy coupling to periplasmic binding protein-dependent transport systems: stoichiometry of ATP hydrolysis during transport in vivo. Proceedings of the National Academy of Sciences of the United States of America. 86 (21), 8257-8261 (1989).
- Grundy, M. Absorption and Metabolism of Dietary Cholesterol. Annual Review of Nutrition. 3 (1), 71-96 (1983).
- Berge, E., et al. Accumulation of dietary cholesterol in sitosterolemia caused by mutations in adjacent ABC transporters. Science. 290 (5497), 1771-1775 (2000).
- Repa, J., et al. Regulation of ATP-binding cassette sterol transporters ABCG5 and ABCG8 by the Liver X receptors α and β. Journal of Biological Chemistry. 277 (21), 18793-18800 (2002).
- Yu, L., et al. Stimulation of cholesterol excretion by the Liver X receptor agonist requires ATP-binding cassette transporters G5 and G8. Journal of Biological Chemistry. 278 (18), 15565-15570 (2003).
- Yu, L., et al. Expression of ABCG5 and ABCG8 is required for regulation of biliary cholesterol secretion. Journal of Biological Chemistry. 280 (10), 8742-8747 (2005).
- Lütjohann, D., Björkhem, I., Beil, F., von Bergmann, K. Sterol absorption and sterol balance in phytosterolemia evaluated by deuterium-labeled sterols: effect of sitostanol treatment. Journal of Lipid Research. 36 (8), 1763-1773 (1995).
- Miettinen, A. Phytosterolaemia, xanthomatosis and premature atherosclerotic arterial disease: a case with high plant sterol absorption, impaired sterol elimination and low cholesterol synthesis. European Journal of Clinical Investigation. 10 (1), 27-35 (1980).
- Salen, G., et al. Sitosterolemia. Journal of Lipid Research. 33 (7), 945-955 (1992).
- Caffrey, M. Membrane protein crystallization. Journal of Structural Biology. 142 (1), 108-132 (2003).
- Michel, H. Crystallization of membrane proteins. Trends in Biochemical Sciences. 8 (2), 56-59 (1983).
- Dürr, N., Gildenberg, M., Ramamoorthy, A. The magic of bicelleslights up membrane protein structure. Chemical Reviews. 112 (11), 6054 (2012).
- Dürr, N., Soong, R., Ramamoorthy, A. When detergent meets bilayer: Birth and coming of age of lipid bicelles. Progress in nuclear magnetic resonance spectroscopy. 69 (1), 1-22 (2013).
- Dufourc, J. Bicelles and nanodiscs for biophysical chemistry. Biochimica et Biophysica Acta (BBA) - Biomembranes. 1863 (1), 183478 (2021).
- Beaugrand, M., et al. Lipid concentration and molar ratio boundaries for the use of isotropic bicelles. Langmuir. 30 (21), 6162-6170 (2014).
- Sanders, R., Schwonek, P. Characterization of magnetically orientable bilayers in mixtures of dihexanoylphosphatidylcholine and dimyristoylphosphatidylcholine by solid-state NMR. Biochemistry. 31 (37), 8898-8905 (1992).
- Seddon, M., Curnow, P., Booth, J. Membrane proteins, lipids and detergents: not just a soap opera. Biochimica et Biophysica Acta (BBA) - Biomembranes. 1666 (1-2), 105-117 (2004).
- Farhat, D., et al. Structural analysis of cholesterol binding and sterol selectivity by ABCG5/G8. Journal of Molecular Biology. 434 (20), 167795 (2022).
- Lee, J. Y., et al. Crystal structure of the human sterol transporter ABCG5/ABCG8. Nature. 533 (7604), 561-564 (2016).
- Ujwal, R., Bowie, U. Crystallizing membrane proteins using lipidic bicelles. Methods. 55 (4), 337-341 (2011).
- Johnson, H., Lee, J. Y., Pickert, A., Urbatsch, L. Bile acids stimulate ATP hydrolysis in the purified cholesterol transporter ABCG5/G8. Biochemistry. 49 (16), 3403-3411 (2010).
- Wang, Z., et al. Purification and ATP hydrolysis of the putative cholesterol transporters ABCG5 and ABCG8. Biochemistry. 45 (32), 9929-9939 (2006).
- Faham, S. Crystallization of bacteriorhodopsin from bicelle formulations at room temperature. Protein Science. 14 (3), 836-840 (2005).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationExplorer plus d’articles
This article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.