Method Article
בשילוב optogenetic ולהקפיא-שבר Replica immunolabeling לבחינת הסדר ספציפי קלט של גלוטמט הקולטנים עכבר האמיגדלה
In This Article
Summary
This article illustrates how the expression of neurotransmitter receptors can be quantified and the pattern analyzed at synapses with identified pre and postsynaptic elements using a combination of viral transduction of optogenetic tools and the freeze-fracture replica immunolabeling technique.
Abstract
במיקרוסקופ אלקטרוני Freeze-שבר כבר טכניקת מרכזי מחקר ultrastructural במשך 40 שנים. עם זאת, עדר אמצעי יעיל כדי ללמוד את ההרכב המולקולרי של ממברנות פיק ירידה משמעותית את השימוש בו. לאחרונה, חלה תחייה מרכזית הודות במיקרוסקופ אלקטרונים להקפיא שבר לפיתוח דרכים יעילות כדי לחשוף חלבונים בממברנה נפרדים על ידי תיוג immunogold. אחת השיטות הללו שמכונה immunolabeling Replica Freeze-שבר דטרגנט-solubilized (FRIL).
השילוב של טכניקה FRIL עם optogenetics מאפשר ניתוח בקורלציה של נכסים מבניים ותפקודיים של סינפסות המרכזית. בגישה זו ניתן לזהות ולאפיין הוא נוירונים טרום postsynaptic על ידי הביטוי של כל אחד מהם של channelrhodopsin מתויג סמנים מולקולריים ספציפיים. המראה הייחודי של התמחות הממברנה הפוסט-סינפטי של glutamatergic הסינפסות נוסף מאפשר בעת תיוג של קולטני גלוטמט ionotropic, לכמת ולנתח את חלוקת intrasynaptic של קולטנים אלה. הנה, אנחנו נותנים צעד-אחר-צעד תיאור של ההליכים הנדרשים כדי להכין העתקים מותאמים ועל איך immunolabel אותם. נדונו גם האזהרות והמגבלות של טכניקת FRIL, במיוחד אלה הקשורים הטיות דגימה פוטנציאליות. השחזור הצדדי הגבוהים של טכניקת FRIL, בשילוב עם optogenetics, מציע גישה חזקה מאוד לאפיון של היבטים שונים של העברה סינפטית ב שבבים עצביים מזוהים במוח.
כאן, אנו מספקים דוגמא כיצד גישה זו שמשה להשיג תובנות יחסי מבנה-תפקוד של סינפסות מעוררות ב נוירונים של המוני תא intercalated של האמיגדלה העכבר. בפרט, אנו חקרנו את הביטוי של קולטני גלוטמט ionotropic ב תשומות מזוהות אוiginated מן intralaminar האחורי התלמוס וגרעיני ברך המדיאלי. סינפסות אלו הוצגו להעביר מידע חושי רלוונטי ללמידה פחד לעבור שינויי פלסטיק על מיזוג פחד.
Introduction
ההגדרה של אדריכלות פונקציונלית של בביו-ממברנות בקנה מידה ננומטרי כבר לערער בשנים האחרונות על ידי פיתוח של מספר טכניקות immunolabeling מתאים במיקרוסקופ אלקטרונים הילוכים. עם זאת, טכניקות אלה, למשל, לפני ואחרי הטמעת immunogold, יש מספר מגבלות חשובות, הכוללים איתור העניים של אנטיגנים ו / או הערכה כמותית מוגבלת של חלבונים קרום הנכנס. מגבלות אלה הופכים להיות קריטיות במיוחד בחקירה של המבנה העדין של מערכת העצבים, מאופיינת ברמה גבוהה של גיוון תא ההטרוגניות סינפסה. ההטרוגניות הזאת תוצאות משתי מגוון מבני ותפקודי מוכתב על ידי האלמנטים טרום postsynaptic ועל ידי ביטוי דיפרנציאלי, העשרה, או אינטראקציה של איתות חלבונים, כגון קולטנים, מובילים, ומולקולות מפעיל.
גישה חדשה עבור immunolabe ישירהלינג של חלבונים בממברנה נפרד או צולבים העתקים להקפיא שבר דטרגנט-solubilized (FRIL) הוצג לראשונה על ידי פוג'ימוטו לפני שני עשורים 1. שיטה מקורית זו הייתה, עם זאת, מספר מגבלות, כלומר, פיצול חמור של העתקים, אשר הקשו על קורלציות משמעותיות של מולקולות שכותרתו עם תאים ממופים בנפרד רקמות מורכבות כמו המוח. לפני כ 10 שנים, Shigemoto ו Fukazawa בהדרגה שיפר את הטכניקה 2. זה היה מקביל המאמצים מקבוצה אחרת של מדענים במעבדות Boulder של קולורדו סטייט, שגם שיפרה בצורה משמעותית את הטכניקה, בפרט לחקר צמתים הפער 3.
שיפור פרוטוקולים שבירים הקפאה ומכונות, כמו גם הכניסה של הקפאה מהירה (בלחץ גבוה), חברה מאפשרת לחוקרים כדי לייצר העתקים רצופים של דגימות של גודל גדול יחסית והיהתמונות באיכות GH של המרכיבים התאיים ביותר ללא מגבלות וממצאים היוצר על ידי קיבעונות כימיים חזקים.
טכניקת FRIL מציעה את היתרון הגדול של כמותי מאוד להציג תעודה מזהה עם באתרו של חלבונים אחד או יותר (בו זמנית) ב היסטולוגית מופתה cytologically תאים שזוהו בתוך רקמות מורכבות כמו המוח, עם היתרון הנוסף של נוף מישוריים של טרום postsynaptic גורמי העתק יחיד. לכן, טכניקת FRIL, למרות מכשולים טכניים רבים, ולפוטנציאל במשך מספר פריצות דרך מדעית מאוד משמעותיות, במיוחד עבור המתאם של נכסים מבניים ותפקודיים של סינפסות בודדות. במהלך העשורים האחרונים, הרבה מידע שהושג על המבנה מולקולרי לפצות, ותפקוד פיסיולוגי של סינפסות; עדיין סינפסות הן מורפולוגית מולקולרית מאוד מגוון תלוי הרשות מראש ו postsynapticלהשכרה נוירונים 4. רק בעבור חופן סוגי סינפסה היו מחקרי מבנה-תפקוד מושלם עד כה 5-7. זה היה בעיקר בשל אילוצים טכניים שמנעו זיהוי מדויק של מהות האלמנטים טרום postsynaptic.
ניתוח ultrastructural סיפקה תובנות קריטיות לתוך השתנות של התמחויות הממברנה הפוסט-סינפטי על פני קשרים סינפטיים ברורים הן מבחינת גודל הסינפטי ותוכן הנוירוטרנסמיטר קולטנים 6, אשר יש לו השפעה גדולה על חוזק הפלסטיות של העברה סינפטית. יתר על כן, מחקרים רבים עולים כי מספר ionotropic הקולטני גלוטמט הביע על סוגים שונים של סינפסה מוסדר בצורה afferent- ויעד תלויה 7-10.
כאן, שיטה המתואר המאפשר ניתוח של הרכב מבנה הקולטן של התמחויות הממברנה הפוסט-סינפטי עם להגדיראלמנטים ותפקוד presynaptic ד. גישה זו מנצלת ביטוי presynaptic של חלבוני אצות רגישים לאור שפותחו לאחרונה, כגון Channelrhodopsin2 (ChR2), ושל טכניקת FRIL לנתח את דפוס ביטוי postsynaptic של α-אמינו-3-הידרוקסי-5-מתיל-4-isoxazolepropionic חומצה (-R AMPA) ו-אספרטט N-methyl-D קולטני גלוטמט (-R NMDA). זו באה לידי ביטוי ב סינפסות הוקמה על ידי אקסונים שמקורם גרעינים התלמוס-מדיאלי ברך האחורי (PIN / MGN) על נוירונים של ההמונים תא intercalated של האמיגדלה (ITC). נוירונים ITC הם תאי GABAergic קוצניים קטנים מאורגנים באשכולות המקיפים את מבנה amygdaloid basolateral (BLA) 11, 12. נוירונים ITC ידועים לקבל תשומות מעוררות מתא העצב העיקרי BLA וכדי למקד את הגרעין המרכזי (CEA), ובכך לתפקד כשער מעכבות עבור זרימת המידע בין BLA ו- CEA 12-15.
לאחרונה, הראינו כי ה- ITנוירונים C הממוקמים באשכול מדיו-הגבו בין BLA ו- CEA גם לקבל תשומות מעוררות ישירות מתכנסות מאזורי קליפת מוח התלמוס ובזמן חושי, אשר שונו על למידת פחד במהלך מיזוג פחד שמיעתי הפבלובית 16. מיזוג פחד הוא אחת הצורות להבין בצורה הטובה ביותר של למידה אסוציאטיבית מבחינת מנגנוני מוח. בשנת מיזוג פחד, גירוי מותנה ניטראלי בתחילה (CS, למשל, בטון) מזווג עם גירוי מותנה מרתיע (ארה"ב, למשל, הלם ברגל קל) וכתוצאה מכך עמותת CS-US והתנה תגובת פחד 17, 18. מעורר תשומות משני אזורים התלמוס ואת neocortical, נושאות מידע המייצג את CS וארה"ב, בהתאמה, היו ידועות להתכנס על עצב פירמידליים של הגרעין לרוחב של האמיגדלה (LA) וכדי לעבור פלסטיות 19. העבודה הקודמת שלנו גילתה כי מידע קלט חושי הוא גם במקביל העביר נוירונים ITC 16.
כצעד ראשון לקראת ניתוח מולקולרי מכניסטית של הקלט החושי הפרט הסינפסות על נוירונים ITC, השתמשנו וירוס adeno הקשורים (AAV) להביע ChR2 מתויג עם חלבון פלואורסצנטי צהוב (YFP). AAV הוזרק מסופי PIN / MGN ואת האקסון זוהו על ידי הביטוי שלהם של ChR2-YFP. השתמשנו בשני הפניות שנוצרו על ידי טכניקת FRIL להעריך את הצפיפות של AMPA-R ו NMDA-R-סינפטי ב סינפסות הקים עם נוירונים ITC ידי מסופי האקסון PIN / MGN.
Protocol
הנהלים הקשורים בנושאים בעלי חיים אושרו על ידי טובינגן Regierungspraesidium, מדינת באדן-וירטמברג, גרמניה, ועל ידי מועצת האתיקה לניסויים בבע"ח האוסטרי, והיו בהתאם הדירקטיבה של האיחוד האירופי על שימוש בבעלי חיים למחקר.
1. זריקות stereotactic של AAV-Channelrhodopsin2-YFP
הערה: זריקות stereotactic בוצעו על פי פרוטוקול שפורסם בעבר 20.
- הכינו את כלי סטרילי על ידי חימום אותם ב 180 ºC עבור 1.5 שעות.
- משוך חדה (~ 50 מיקרומטר קוטר) פיפטות זכוכית זריקות באמצעות חולץ microelectrode אופקי להגדיר עם הפרמטרים הבאים: ערך Ramp = חום - 20, משוך = 0, מהירות = 100, זמן = 200, לחץ = 200.
הערה: ערך רמפה צריכה להיקבע לכל מנה של טפטפות זכוכית שנרכשו בהתאם להוראות שמספקות יצרן micropipette חולץ. - Premix 1 μl של פתרון וירוס ו -0.2 μl של פתרון ירוק מהיר 0.1% (ראות טובה יותר של הפתרון ב פיפטה זכוכית) ב חיץ מלוחים פוספט סטרילית (PBS; 25 מ"מ, 0.9% NaCl, pH 7.4). מלאו את פיפטה זכוכית באמצעות פיפטה 10 μl וטיפים ג'ל-fil. עבור מבנה ויראלי, השתמש rAAV-hSyn-ChR2 (H134R) -eYFP (סרוטיפ 2/9).
- להרדים עכבר באמצעות מכשיר הרדמת חיה קטן (3% isoflurane חמצן לזירוז). להשתמש. לצורך המחקר, השתמשו 3 עכברים wild-type בגילאי ~ 6 שבועות.
- לגלח את הראש בין האוזניים והעיניים ולחטא עם פתרון המבוסס povidone-יוד.
- החל משחת עין כדי למנוע התייבשות של עיניים במהלך הרדמה. תת עורי להזריק עכבר עם משכך כאבים (meloxicam מבוססי, 0.1 מ"ל של פתרון 5 מ"ג / מ"ל).
- העבר את העכבר בתוך מסגרת stereotactic ולשמור הרדמה באמצעות מכשיר גז הרדמה (2% isoflurane חמצן לצורך תחזוקה). בדוק עומק ההרדמה על ידי חוסר נסיגה איבררפלקס לפני שתמשיך.
- לשמור על תנאים סטריליים ככל האפשר במהלך ההליך הכירורגי כולו. ללבוש פן מסכה חד פעמית, גלימה וכפפות כירורגיות.
- לעשות חתך בעור של כ 1 ס"מ על החלק העליון של הראש באמצעות מספריים 20. משוך בעדינות את עור לצד באמצעות מלקחיים בוטים, לתקן עם מלחציים לחשוף פני גולגולת וגולגולת נקיה עם H 2 O 2.
- המוזרקים מארק על הגולגולת באמצעות סמן קבע טיפ נאה. לקדוח חור קטן (כ 1 מ"מ קוטר) לתוך הגולגולת באתר הניכר. עבור זריקת PIN / MGN חד-צדדי הרבעה זה, השתמש בכלי הקואורדינטות הבאים: מן גבחת (מ"מ): 3.0 האחוריות, לרוחב ± 1.8, 3.8 גחון.
- הר מילא את הכוס פיפטה על מסגרת stereotactic מחובר להתקן הזרקה בלחץ ולהביא פיפטה כדי גבחת עמדה.
- לשבור את קצה פיפטה הזכוכית באמצעות מלקחיים קצה ישר בסדר. ודא קצה פיפטה פתוח ידי applyinga בכמה פעימות לחץ שחול התבוננות של טיפות של תמיסת וירוס.
- עבור אל קואורדינטות הזרקה הרצויות ולהזריק מחצית תוכן פיפטה (~ 0.5 μl) באמצעות ההגדרות הבאות בהתקן ההזרקה בלחץ: לחץ 20 psi, אורך הפולס ממוצע 30 מילישניות, מספר ממוצע של פולסים 50.
- השאירו פיפטה במקום דקות ~ 1 לפני לאט (1 מ"מ / דקה) חוזרת בה זה.
- גולגולת נקיה עם PBS (pH 7.4) ולהסיר מלחציים. משוך בעדינות את העור יחד, תפר את החתך (3 - 4 קשרים). החל החיטוי (povidone יוד המבוססת) באזור הפצע.
- עצור ההרדמה ואל תשאירו את העכבר ללא השגחה עד ער לחלוטין. שמור עכברים חד שוכן. לאחר הניתוח, ממשיך לעקוב אחר מצב הבריאות; אם כאבי ניהול 'צורך.
- שמור חיות למשך 4 שבועות לפני קיבעון במוח על מנת להבטיח רמות מתאימות של הביטוי ויראלי.
2. הכנת הדגימה
- מוח קיבעון </ Strong>
- הכנה מקבע
- עבור 1 ליטר של מקבע, שוקל 10 גרם של paraformaldehyde ולהוסיף אותו ל -300 מ"ל של H 2 O. ללא יונים מחממים עד 55 - 60 ºC עבור ~ 10 דקות עם ערבוב מתמיד.
- כבו את מהאש ומוסיפים 7 - 8 טיפות של 4 N NaOH. הפתרון צריך להיות ברור ~ 10 דק '.
- תן לזה להתקרר עד RT, להוסיף 150 מ"ל של תמיסה רוויה של חומצה picric ולהביאה 500 מ"ל עם H 2 O. ללא יונים
- הוסף 500 מ"ל של 0.2 M חיץ פוספט (PB). מסנן עם נייר סינון. התאם ל- pH 7.4 עם NaOH.
- מצנן את מקבע עד 6 ºC, ולאחסן אותו בבקבוקי זכוכית כהים לא יותר מיום אחד ב -6 ºC.
- זלוף Transcardiac
- להרדים עכברים עם זריקה intraperitoneal של thiopental (120 מ"ג / ק"ג משקל גוף). ודא החיה בהרדמה עמוקה על ידי סימון רפלקטיבית נסיגת הדוושהלשעבר, אשר אמור להיעדר. מניח את החיה על גבית על שולחן זלוף עם ארבעת הגפיים הכבולים.
- פתח את דופן בטן longitudinally במספריים קהה-סוף ולעשות שני חתכים נוספים רוחבי לאורך גבול הזנב של כלוב הצלעות, לחשוף את הסרעפת. לחתוך את הסרעפת וחתך את קיר בית החזה בגבול osteocartilageneous משני הצדדים. הרם את סוף הזנב של הלוח המרכזי של קיר בית החזה המכיל את עצם החזה כדי לחשוף את הלב.
- הסר את קרום הלב, לעשות חתך מדויק קטן בקצה של החדר השמאלי להודות הצינורית של מנגנון זלוף. השתמש צינורית בוטה עם קוטר פנימי של 0.6 מ"מ. להעביר את הצינורית בעדינות דרך החדר עד הקצה מופיע לתוך האאורטה עולה ולאבטח את הצינורית עם מהדק. כדי לאפשר את הדם perfusates ליציאה מזרם הדם, לעשות חתך ב אטריום ימין.
- עכברים perfuse transcardially באמצעות משאבת peristaltic בקצב זרימה5 מ"ל / דקה בהתחלה עם PBS (25 מ"מ, 0.9% NaCl, pH 7.4) למשך כ 1 דקות, ואחריו מקבע קר כקרח במשך 7 דקות.
- לאחר קיבוע, לנתק את הראש העכבר עם זוג מספריים ולאחר מכן לחתוך את העור באמצעות קו האמצע מהצוואר עד האף. הסר את השריר כדי לחשוף את הגולגולת.
- בעזרת מספריים חדים, לעשות חתך אורכי דרך העורפית ועצמות interparietal החל מגנום foramen. בעזרת פינצטה בסדר להסיר העצמות האלה כדי לחשוף את המוח הקטן כולו. ואז לעשות עוד חתך אורכי דרך הקודקודית והעצמות חזיתית עד עצם האף ולהסיר אותם עם פינצטה כדי לחשוף את המוח כולו.
- בעזרת מרית להסיר את המוח מבלי לפגוע בו, ולמקם אותו קר כקרח 0.1 M PB.
- הכנה מקבע
- חתך זמירה של דגימות
- חותך בלוק עטרה של כ 5 - 6 מ"מ עם סכיני גילוח המכילים שטח של עניין. מדביקים אותו על המחזיקvibroslicer עם דבק cyanoacrylate. אוריינט לחסום רקמות כך הניאוקורטקס פונה להב רוטט. Slice חלקי עטרה, המכילים את האמיגדלה, ב 140 מיקרומטר עם vibroslicer (איור 1 א) ב 0.1 קר כקרח M PB, ולאסוף אותם בצלחת 6-היטב באותה החיץ.
- תחת סטראו, לקצץ את האזור של עניין (כאן, האשכול paracapsular מדיו-הגבי של ITC, ראה איור 1B) מן הפרוסה. האם זה בצלחת פטרי מצופה אלסטומר סיליקון מלא 0.1 M PB, באמצעות אזמל עיניים. ודא כי אבני גזוז להתאים לתוך החור של spacer (כ 1.5 מ"מ) (איור 1 ב).
- להזיז את אבני גזוז לתוך פתרון cryoprotection (גליצרול 30% ב 0.1 M PB) O / N ב -6 ºC.
3. הקפאה בלחץ גבוהה
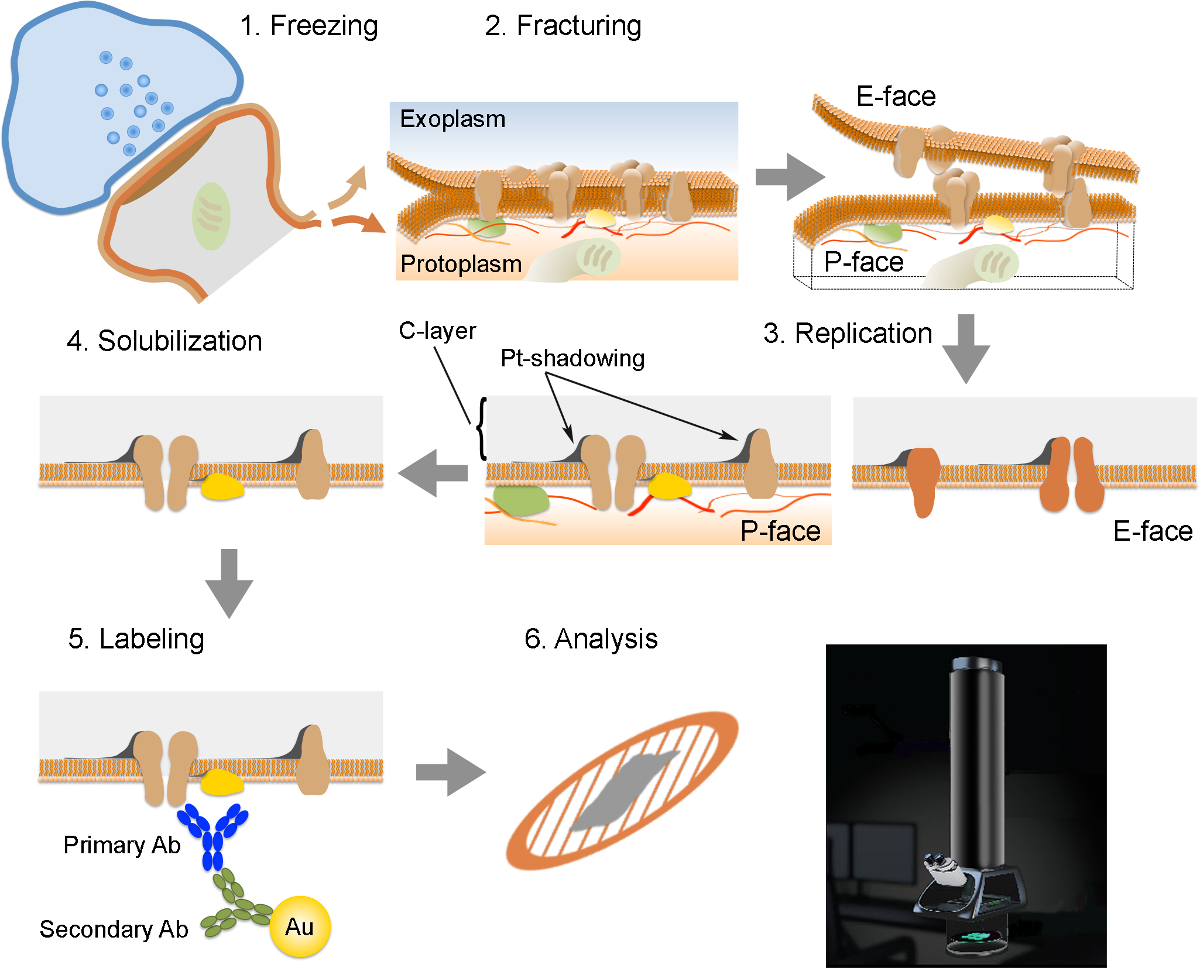
הערה: FRIL כוללת 6 צעדים חיוניים (איור 2): 1) להקפאה מהירה בלחץ גבוה (2,300 - בר 2,600) של הדגימה. 2) ושבר של הדגימה. המטוס שהבר בדרך כלל מלווה את הליבה הידרופובי המרכזית של ממברנות קפואות, פיצול אותם לשתי עלונים חצי קרום: וחצי שנמצא סמוך מפרוטופלסמה (P-הפנים) וחצי שנמצא בסמוך למרחב התאי או exoplasmic (E- פָּנִים). 3) שכפול של הדגימה ידי ואקום בתצהיר של פלטינה ופחמן. 4) דטרגנט-העיכול של הרקמה. 5) תיוג immunogold. 6) ניתוח של ההעתק באמצעות מיקרוסקופ אלקטרוני הילוכים.
- הכנת נושאות נחושות
הערה: כדי להיות מטופלים באמצעות רצף השלבים של הליך FRIL, דגימות צריכות להיות מותקנות על גבי מתכת (זהב או נחושת) נישא. ספקים אלה משתנים בגודלם ועיצוב בהתאם למצב של שבירה וסוג המכונות המשמשות. כאן השתמשנו נושאות נחושת (איור 1 ב ', ה) וכן צירים "שולחן העתק כפול"; (ראה איור 3 ב), אשר כאשר פתחו מייצר שבר מתיח באמצעות הדגימה קפואה (איור 2). זה מאפשר לשמור על ולשכפל שני הצדדים של הדגימה שבורה.- ספקים נחושים פולנים עם מסיר כתמים באמצעות גיליון של עור יעל.
- נישא מקום בתוך אגרטל זכוכית ונקי פעמים עם חומר ניקוי שאינו יוני (pH ~ 1.5) באמבט מי sonicating, ולאחר מכן לשטוף בהרחבה במים ברז ואחריו מים ללא יונים ולאחר מכן לשטוף פעמים עם אתנול.
- Sonicate נישא הנחוש אצטון במשך 15 דקות.
- מניח את נישא על נייר פילטר לייבוש.
- צרף טבעת דבק דו-צדדי על מפעיל נחושת (תרשים 1C), אשר ישמש מחזיק גם עבור בלוק גזוז (המוביל מחזיק).
- הקפאה של הדגימה
הערה: ידית חנקן נוזל בזהירות מרכיבת משקפים מתאימים.- הפעל את בלחץ גבוה חינםיחידת זיע (איור 1F) לפחות 1.5 שעות לפני תחילת עם הקפאת הדגימה.
- התחל חימום על ידי לחיצה על הכפתור "AIR המחמם", ואופה החוצה למשך 50 דקות. טמפרטורת האוויר נקבע ל -80 מעלות צלזיוס.
- חבר את מיכל חנקן ליחידת הקפאת בלחץ גבוה ולחץ על כפתור "חנקן" למלא את החלק הפנימי דיואר עם חנקן נוזלי. המנורה "LEVEL חנקן" אורות למעלה. התחל קירור על ידי לחיצה על הכפתור "קירור".
- לחצו על "דרייב" כפתור כאשר "LEVEL חנקן" יוצא. בדוק כי המערכת ההידראולית מעבירה את הבוכנה קדימה ואחורה 3 פעמים.
- לחץ על כפתור "AUTO", ואת הכפתור "חנקן". ברגע "מוכן" נדלק, היחידה הקפאת בלחץ גבוה מוכן להקפאה בלחץ גבוה.
- מניחים בלוק גזוז לתוך החור של נייר דו צדדית (איור 1B) באמצעות לולאה חוט פלטינה אשר כבר נמס לתוך AGפיפטה ילדה.
- הסר עודף הפתרון cryoprotectant באמצעות נייר סינון או מברשת.
הערה: הליך זה חשוב גם כדי להסיר בועות אוויר כי עלול להיווצר סביב הרקמה ואשר עלול לגרום לעיוות בצורת רקמות ו / או ultrastructure. - תחת סטראו, לכסות את הספק המחזיקים ביחד מוביל אחר, כך לחסום הרקמות הוא דחוק בין שני ספקים.
- הכנס את כריך המוביל לתוך מחזיק הדגימה של היחידה הקפאת בלחץ גבוה (איור 1D). הכנס את בעל הדגימה לתוך יחידת ההקפאה בלחץ גבוה (טיפ למטה) לאבטח אותו על ידי הברגה של בעל הדגימה.
- מפעיל את פעולת ההקפאה על ידי לחיצה על כפתור "ג'ט-אוטומטי". עבודה מהר ככל האפשר, הוצא את בעל הדגימה להטביע את הקצה עם חנקן נוזלי לתוך תיבה מבודדת. לטבול את קצות שני זוגות מלקחיים של החנקן הנוזלי כדי לקרר אותם.
- מוציאים בזהירות הדואר המוביל כריך מבעל הדגימה ומניחים אותו בתוך cryovial מראש צונן. ודא כי ספקי מטופלות רק עם מלקחי חנקן מקורר נוזל. Cryovials צריך להיות מחורר לאפשר חנקן לזרום החוצה מבקבוקון (איור 1G).
- חזור על שלבי 3.2.6 עד 3.2.11 עד שכל הדגימות הרצויות הוקפאו. -כריכים מובילים מרובים המכילים הסוג המדגם אותו ניתן לאחסן אותו הבקבוקון.
- אחסן את cryovials המכיל את הנושאים בתוך cryotank עד השכפול (1H איור).

באיור 1. הכנת רקמות והקפאה בלחץ גבוה. (א) Vibroslicer נהג סעיף הרקמה. (ב) סעיף מוח עטרת עכבר וכתוצאה המכיל בצד לראות האמיגדלה אל כתף עם acמנשא אופר מצויד עם טבעת דבק דו-צדדי. התיבה המקווקות מציינת את תחום העניין המכיל באשכול paracapsular המדיאלי של המרכז לטניס בישראל. הקוטר של החור דבק דו-צדדי הוא כ 1.5 מ"מ. (ג) כלים להכנת נושאות נחושת. מלמעלה-שמאל באופן כיוון השעון: דו צדדית קלטת, פינצטה, אגרופן, ספקים נחושת, ומספריים. (ד) קלטים המובילים-הכריך לתוך מחזיק הדגימה להקפאה בלחץ גבוה. המנשא-הכריך מושם לתוך החור של בעל הדגימה. (E) נושאות נחושות בלי ועם טבעת דבק דו-צדדי ואת "הכריך המוביל". (F) יחידת לחץ הקפאה גבוהה עם טנק בלחץ האכלת חנקן נוזלי אליו. (ז) cryovial להצטייד הרקמות קפואות. הערה החורה במצב הבינוני-הגבוה של הבקבוקון המאפשר גז חנקן לזרום החוצה של הבקבוקון (ראש החץ). (H) Cryotank לאחסון הרקמה הקפואה. אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
{kind=link}
Freeze-שבר שכפול 4.
- הכנת תותחי אלומת האלקטרונים
- לפני החדרת התותחים אלומת אלקטרונים, להסיר את המגן עם "צלחת מטה הטיה". מניח את "מד ההגדרה" למירכוז הנימה לתוך צ'אק קולט דרך מכסת קטודה הנמוכה.
הערה: הסוף בקוטר הגדול יותר של מד ההגדרה משמש את אקדח הפחם ואילו הקצה בקוטר הקטן יותר הוא על אקדח הפלטינה. - חלק את הנימה החדשה על המד עד "רבדי לחץ" יכול לצבוט את הקצוות של החוט, מה שמבטיח כי סליל חוט הלהט אינו טמון בזווית.
- הסר את מד ההגדרה והכנס את מוט הפחמן. תקנו ידי הידוק שיתוףצ'אק llet של בעל מוט המאייד, על מנת להבטיח כי גובה בקצה המוט נמצא באמצע הסליל השני מלמטה. עבור אקדח פלטינה, בעיצומה של סוף מוט פלטינה צריך להימצא במרכז של סליל חוט השני מלמעלה.
- החזר את הצלחת מטה ההטיה וכנס רובים ליחידה שבר הקפאה. נקו את הרובים עם Blaster-חול לאחר השימוש.
- לפני החדרת התותחים אלומת אלקטרונים, להסיר את המגן עם "צלחת מטה הטיה". מניח את "מד ההגדרה" למירכוז הנימה לתוך צ'אק קולט דרך מכסת קטודה הנמוכה.

איור 2. איור של השלבים העיקריים של טכניקת FRIL.
מתווה את השלבים השונים הנדרשים להכנה וניתוח של העתק. (1) הקפאת לחץ גבוה של רקמת גוף. (2) שביר. במהלך השבירה של רקמה קפוא, bilayer השומנים של קרום פלזמה מחולק לשני חצאים על הממשק הידרופובי. חלבונים בקרום פלזמה מוקצים על גם את exoplasmic (E-פנים) או פרוטוממברנות פלזמה (P-פנים). שכפול (3). אידוי פחמן (C) לוכד שומנים וחלבונים על פני השטח של הרקמה שבורה. החומר צופה 2 ננומטר פלטינה / פחמן הצללה בזווית 60 מעלות, ולאחר מכן שכבה נוספת פחמן 15 ננומטר מה שמחזק את המבנה של ההעתק (C השכבתי, Pt-הצללה). (4) solubilization. הרקמה לא לכוד על ידי קרום העתק הוא solubilized אז עם פתרון SDS. תיוג (5). חלבונים של עניין ניתן מדמיינים על העתק באמצעות קומפלקס עשוי נוגדנים ראשוניים ספציפיים (ראשי Ab) ונוגדנים משניים (Secondary Ab) מצומדות עם חלקיק זהב (Au). השימוש גדל שונה של חלקיקי זהב מאפשר זיהוי של יותר חלבון אחד על אותו ההעתק. (6) לאחר immunolabeling, העתקים נאספים על רשתות רשת נחושת ונותחו באמצעות מיקרוסקופ אלקטרונים הילוכים ב 80 -. 100 קילו וולט אנא Click כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
{kind=link}
- גדר של יחידת שבר הקפאה
- הפעל את יחידת שבר הקפאה (איור 3 א) על ידי סיבוב חשמל ל -1 לתיאור מפורט של הנהלים להקפיא שבר ושכפול, ראה הוראות הפעלה שספקו יצרן.
- לפני הקירור מכשיר להקפיא שבר, לאפות את מערכת הקירור כולה של היחידה עם אוויר חם. לחצו על כפתור ה "הפשרה" במכשיר 010 MTC (יחידת בקרת טמפרטורה) (איור 3 א) ולתת בטווח תהליך החוצה ואופים במשך 45 דקות.
- הפעל את תחנת ואקום. היחידה שבר הקפאת בדרך כלל פועלת במגוון ואקום של ~ 10 -6 - 10 -7 mbar.
- מלא את מכל חנקן ולחבר אותו ליחידת שבר הקפאה. בדוק כי בעל השסתום יבש גם לנקות את כניסת הטנק לפני ההחדרה בעל שסתום (לחות יכולה להפריע vacuuמ 'כל אינדיקציה לגבי N 2 מילוי המכל).
- התחל קירור על ידי קביעת הטמפרטורה ל -115 מעלות צלזיוס. קירור לוקח בערך 45 דקות.
- הכנס את רובי אלומת אלקטרונים ולהתאים זרם ומתח להגיע הפרמטרים הבאים לאידוי:
פחמן אקדח: סיבוב על, עמדה 90 °, שיעור צבירת פחמן 0.1 - 0.2 nm / sec
פחמן-פלטינה אקדח: סיבוב מעל, עמדה 60 °, שיעור הצבירה 0.06 - 0.1 ננומטר / sec
הערה: אם אקדח משמש בפעם הראשונה לאחר החליפין של מוט הפחמן או פלטינה, הדגה במשך 3 דקות לפני השימוש.
- שביר ושכפול
- כנס מוביל כריכים קפואים לתוך שולחן העתק כפול מוודאים כל המניפולציות נעשות בחנקן נוזלי.
- עבר שולחן העתק כפול לכלי דיואר ולתקן אותה למקלט בשלב הדגימה בזווית של 45 מעלות. רמת החנקן הנוזלית תמיד צריכה להיות מעל השולחן העתק הכפול.
- תרים את השולחן העתק הכפול עם מניפולטור השולחן ולהכניס אותו לתוך יחידת שהבר הקפאה לבמה הקרה. יש להמתין כ -20 דקות, כדי לאפשר הטמפרטורה של השולחן העתק הכפול להסתגל -115 מעלות צלזיוס.
- בדוק שהחלל הוא מתחת ל -10 -6 mbar והטמפרטורה -115 מעלות צלזיוס.
- שבר את הרקמה על ידי סיבוב נגד כיוון שעון הידני של הגלגל המחובר אל התכריכים הממוקמים מעל שולחן ההעתק הכפול. כאשר התכריך מסתובב, זה מכריח השולחן העתק פעמים כדי לפתוח, שבירת הרקמה.
- לחץ על כפתור ה "המתח גבוה" ב (יחידת בקרת אידוי אלומת אלקטרונים) EVM 030 התקן של יחידת שבר הקפאה (איור 3 א).
- לשכפל את המשטחים החשופים של הרקמה שבורה (איור 3 ג) על ידי אידוי של פחמן (סיבוב) באמצעות אקדח אלומת אלקטרונים ממוקם בזווית של 90 מעלות בעובי של 5 ננומטר, ואחריו שאדו חד כיווניכנף עם פחמן פלטינה בזווית 60 מעלות בעובי של 2 ננומטר. לבסוף, להחיל 15 שכבה עבה ננומטר של פחמן מזווית 90 מעלות (סיבוב).
- השתמש בפרמטרים הבאים לאידוי:
פחמן 1: סיבוב על, עמדה 90 °; מהירות 0.1 - 0.2 nm / sec; 5 ננומטר
פחמן-פלטינה 2: סך 60 °; מהירות 0.06 - 0.1 ננומטר / sec; 2 ננומטר
פחמן 3: סיבוב על, עמדה 90 °; מהירות 0.3 - 0.5 ננומטר / sec; 15 ננומטר - הסר את הדגימות לשכפל מיחידת שהבר הקפיא ולהעבירם קרמיקה 12-גם צלחת (איור 4 א) מלאים TBS (טריס שנאגר מלוח, pH 7.4).
- באמצעות מוט לולאת חוט פלטינה, להסיר את הרקמה המשוכפלת ממוביל הדגימה (איור 4 א).
- חזור על שלבי 4.3.1 עד 4.3.10 עד שכל הדגימות כבר משוכפלות.
 r /> איור 3. הקפאה-שביר ו שכפול.
r /> איור 3. הקפאה-שביר ו שכפול.
(א) יחידת שבר ההקפאה. המכונה כולל מספר יחידות מלאות צג. דוגמאות מוכנסות לתא דרך יציאה בצד השמאל של החדר. מיכל חנקן נוזלי בלחץ מחובר ליחידת שבר ההקפאה כדי לקרר את הבמה. תמונות מתחת נופים מוגדל הפגנת שתי היחידות (UPC 010 ו MDC 010) ואת הצג מוצגות פרמטרים במהלך האידוי של שכבת פחמן השני. (ב) נפתח (משמאל) וסגר (מימין) נופים של השולחן העתק הכפול. את "מוביל כריכים" מוכנסים לתוך החריצים של הטבלה (מסומן בחצים). הזרועות הקטנות למנוע "מובילים-כריכים" ינשור במהלך מניפולציה. (ג) מדגם מפולג משוכפל. רפליקות להופיע בסרטים שחורים דקים על גבי הרקמה שבורה.53 / 53853fig3large.jpg "target =" _ blank "> לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
- SDS-עיכול של ההעתק
- העברת העתק בקבוקון זכוכית 4 מ"ל מלא 1 מ"ל של חיץ SDS-עיכול (2.5% נתרן Lauryl Sulfate, 20% סוכרוז 15 מ"מ טריס, pH 8.3). תקציר עבור 18 שעות ב 80 ° C עם רועד (45 שבץ / min).
- העברת העתקים לצינור חדש מלא חיץ SDS-עיכול ולאחסן ב RT.
5. immunolabeling
הערה: כל incubations מבוצעת ב RT עם רעד עדין, למעט incubations עם נוגדנים.
- שטוף את ההעתק במשך 10 דקות במאגר SDS-עיכול טרי.
- שטפו את העתק פעם עם 2.5% BSA (אלבומין בסרום שור) ב TBS במשך 5 דקות, ולאחר מכן BSA 3 x 10 דקות עם 0.1% ב TBS.
- בלוק הלא ספציפי אתרי קישור ב TBS עם 5% BSA עבור שעה 1.
- החלת אנטי עיקרייםגופים מדוללים 2% BSA-TBS. בצע incubations בטיפת 30 μl (איור 4B) בתא לח ב 15 מעלות צלזיוס למשך 72 שעות (איור 4C).
- לשם כך המחקר, תהליך הן העתקים מן הרקמה שבורה. דגירה העתק אחד עם נוגדן polyclonal שרקן שהועלו כנגד חומצות אמינו 717 - 754 של העכבר GluR1 המשותף לכל יחידות משנה AMPA-R (דילול: 1: 200) או נוגדנים חד שבטיים עכבר שהועלו כנגד חלבון היתוך רקומביננטי כיסוי חומצות אמינו 660 - 811 של למקטע NR1 של NMDA-R (דילול: 1: 500), ו נוגדן polyclonal ארנב שהועלו כנגד חלבון פלואורסצנטי ירוק (דילול: 1: 300).
- דגירת ההעתק האחר עם נוגדן polyclonal ארנב שהועלה כנגד פפטיד סינטטי המתאים לחומצות אמינו 384 - 398 מתוך הקולטן μ-אופיואידים החולדה (דילול: 1: 500).
- לשטוף ב TBS עם 0.05% BSA (3 x 5 דקות.).
- החל נוגדנים משני. לקבלת u במחקר זה se זהב (5 ננומטר עבור ionotropic קולטני גלוטמט, 10 עבור μ-אופיואידים לקולטנים ו / או 15 ננומטר עבור ChR2-YFP) נוגדנים מצומדות מדולל TBS עם 2% BSA. מדולל נוגדנים משני 01:30 דגירה בטיפת 30 μl ב -15 ° CO / N.
- לשטוף דקות 3 x 5 אינץ '0.05% BSA-TBS ב RT.
- לשטוף דקות 2 x 5 במי ultrapure.
- העתק הר על רשת המקבילה formvar מצופה 100 אונליין (האיור 4D).
6. ניתוח Replica
- העתקי תמונה עם מיקרוסקופ אלקטרוני הילוכים (TEM) ב ק ו 80 או 100. לרכוש תמונות דיגיטליות באמצעות מצלמת (מכשיר טעון מצמיד) CCD.
- לא מקוון, למצוא מקביל אזורים על תמונות משני ההעתקים באמצעות ציוני דרך (איור 4E). ניתוח תמונות דיגיטליות באמצעות תמונת ג 'קבעה באזור postsynaptic ומספר החלקיקים שכותרתו immunogold מכוונים נגד קולטן המנותח.
page = "1"> 
איור 4. immunolabeling של Replica.
(א) כלי מניפולציה ושטיפת ההעתקים. צלחת קרמיקה 12 גם (למעלה מימין) ו -2 סוגים של טפטפות זכוכית (למעלה משמאל). פיפטה הזכוכית עם קצה עגול (משמאל למטה) משמשת להעברת העתק, ואת פיפטה עם מוט פלטינה (מרכז תחתון) משמשת כדי להתפתח העתקים. העתק חיץ כביסה (מימין למטה). (ב) immunolabeling של העתקים מתבצע בטיפות (30 μl) הניחו על חתיכה קטנה של parafilm בתוך באר של צלחת 6 באר בתרבית רקמה. שים לב העתק מכוסה הירידה של נוגדנים המכילים חיץ. כדי למנוע אידוי, חתיכת לחלח של נייר טישו מצויד מסביב לקצה הפנימי של הבאר. (C) חממת צעד immunolabeling. Incubations מתבצעות 15 ° C. (ד) העתק רכוב על formvar מצופה 100רשת קו מקביל. (E) micrographs בהגדלה נמוכה מזוג העתקים. הריבועים המנוקדים מציינים שלושה ציוני דרך טיפוסיות לזהות מיקום ההעתקים המקבילים. סרגל קנה מידה: 10 מיקרומטר. כל הנתונים מוצגים ± רשע כמו SEM אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
{kind=link}
תוצאות
טכניקת FRIL, בשילוב עם ביטוי של מפעילי optogenetic ממוצא מיקרוביאלי 21, כלומר, ערוצי משולב קרום הפלזמה והובלו ביעילות anterogradely לאורך אקסונים, מאפשרת לבחון את ביטוי postsynaptic כמותית של AMPA-R ו NMDA-R בכל תת-קבוצה מוגדרת של סינפסות. זו מוצגת כאן עבור אקסונים שמקורם גרעינים התלמוס מובהק (למשל, PIN / MGN) על נוירונים ITC באמיגדלה. גישה זו מאפשרת ניתוח מולקולרי של סינפסות הקלט החושי הפרט על נוירונים ITC, קבוצה של תאים שהיו מגיבים אפיון אנטומי ומולקולרית מפורט עד כה.
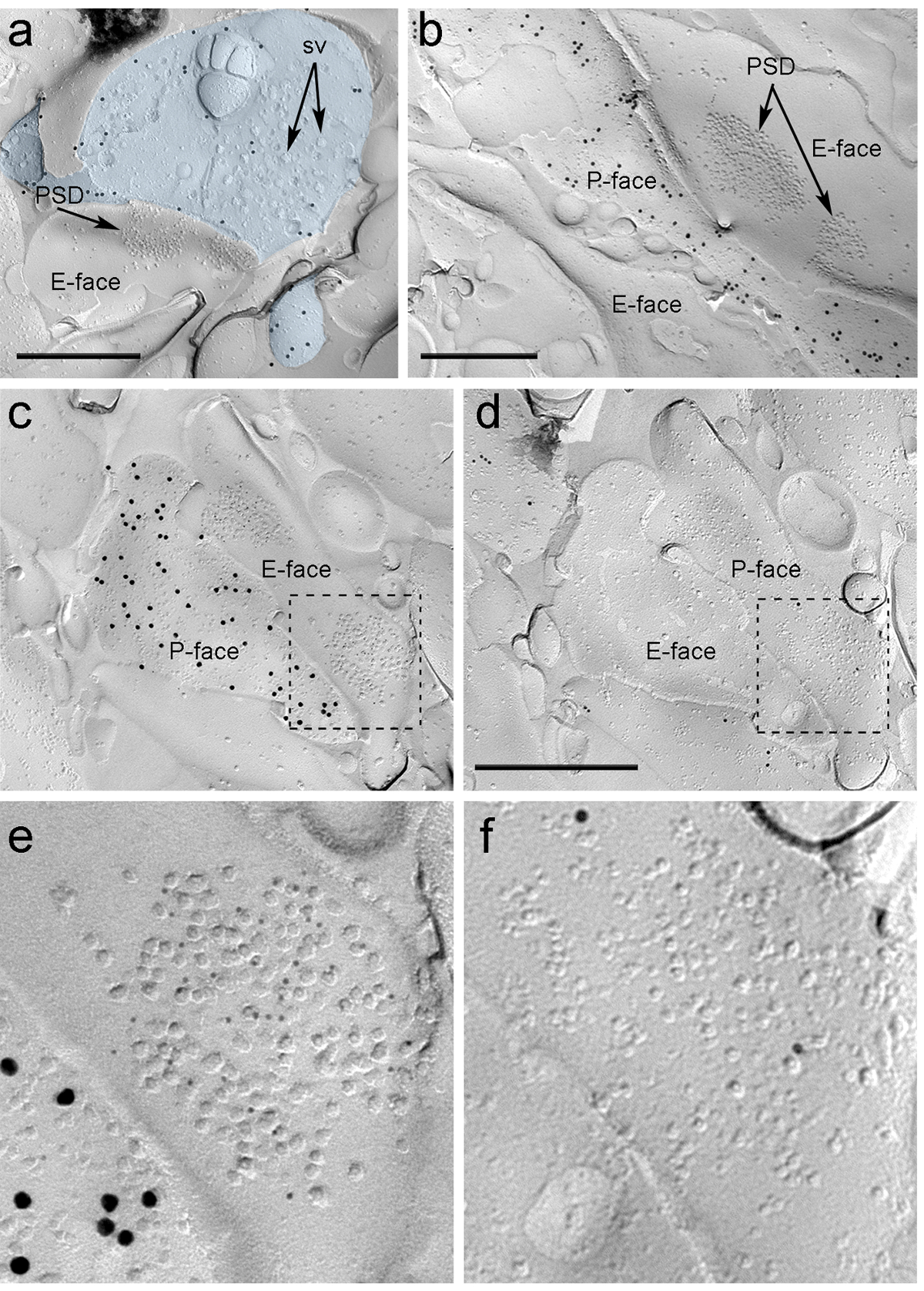
ארבעה שבועות לאחר ההזרקה stereotactic של rAAV-ChR2-YFP לתוך PIN / MGN, ChR2-YFP חיובי אקסונים בצפיפות מעוצבבים את הלוס אנג'לס, באזור המעבר amygdalostriatal (Astria) ואת ואני paracapsular המדיאלי אשכול TC באמיגדלה, דפוס עקבי לחלוטין עם העקיבה הקודם שלומד 16, 22. גם זיהינו זהב אינטנסיבי immunolabeling עבור ChR2-YFP על P פנים של אקסונים ומסופי ב העתק להקפיא שבר מן rAAV-ChR2-YFP- עכברים שהוזרק (איור 5 א), אך לא העתקים מעכברים הלא מוזרק. ההתמחות הממברנה הפוסט-סינפטי של סינפסות glutamatergic ב העתק ניתן לצפות כמו מקבץ של חלקיקים intramembrane (שדונים) על E-פני קרום התא 2, 23, והוא מלווה לעתים קרובות על ידי P-פני קרום התא presynaptic שלה 7 (איור 5 ב-ג). תכונות אלה אפשרו לזהות את התמחות postsynaptic של סינפסות glutamatergic הוקמה על ידי מסופי האקסון PIN / MGN (איור 5 ו -6). אנחנו שכותרתו AMPA-R עם נוגדן שמכיר כל ארבע יחידות משנה (GluA1-4), ואילו-R NMDA התגלה באמצעות נוגדן כנגד למקטע NR1 החיוני.
ntent "FO: keep-together.within-page =" 1 "> בגלל חוסר של כלים לגילוי על אותו העתק אם סינפסות אלה נעשו עם קוצים או פירים הדנדריטים של נוירונים ITC, אנו שכותרתו הפנים העתק המתאים עבור μ קולטנים -opioid, כמו נוירונים ITC מבטאים רמות גבוהות postsynaptically של קולטנים אלה 24. זה נדרש זיהוי של אותה פרופילים postsynaptic בשנתיים העתקים (איור 5 ג-F ואיור 6A-D) משתמש באסטרטגיה המעסיקה ציוני דרך (איור 4E) .
איתור איור 5. של ChR2-YFP ו ionotropic גלוטמט רצפטורים על Replica ידי immunogold חלקיקים. (א) מסוף האקסון צולב שבר (תכלת) ו מנות קטנות של P-הפנים שלה שכותרתו עם חלקיקי זהב 15 ננומטר גילוי ChR2-YFP. בתוך הטרמינל, קרום oשלפוחית סינפטית רבה f ניתן לצפייה (ד"ה). הערה הספציפית של immunolabeling במידה רבה מוגבלת קרום הפלזמה. תיוג עבור ChR2 מזהה את מסוף כנובעת ה- PIN / MGN. המסוף מהווה סינפסה סימטרית עם עמוד שדרה. התמחות הממברנה הפוסט-סינפטי (PSD) על E הפנים מציגה מקבץ מאפיין של חלקיקי intramembrane, והוא הנקרא עם 5 ננומטר חלקיקי זהב החושפים AMPA-R. (ב) P-פני האקסון להביע ChR2 (שכותרתו עם 15 חלקיקי זהב ננומטר) מוצג הגובלים שני דנדריטים, אחד מהם בעל שני PSDs שכותרתו עם חלקיקי זהב 5 ננומטר חושף NMDA-R. (CD) פרצופים השני של ממברנות טרום postsynaptic של סינפסה PIN / MGN-ITC. (ג) P-פני מסוף מבטא ChR2 (שכותרתו עם חלקיקי זהב 15 ננומטר) משתרע על פני E-פני שני פירים הדנדריטים, אחד מהם המכיל שני PSDs שכותרתו עבור AMPA-R (5 חלקיקי זהב ננומטר). ( ד) P פן המקבילים של שני דנדריטים מתויג עבור μ-אופיואידים לקולטנים (10 חלקיקי זהב ננומטר). (EF) נופים מוגדלים של האזורים שהתוו קווים מהקווקווים. ברי סולם:. 500 ננומטר אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
{kind=link}
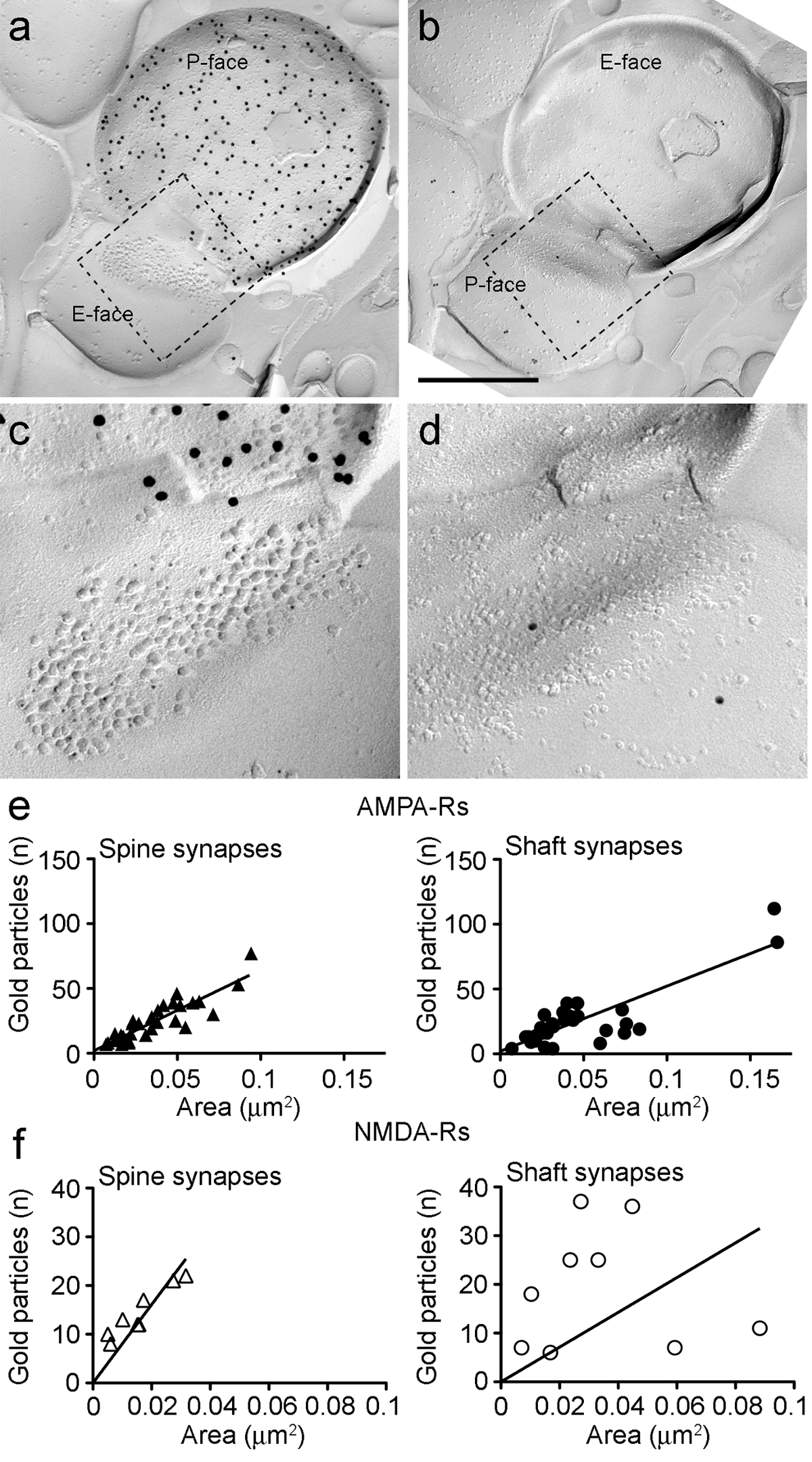
Immunoparticles עבור AMPA-R ב הסינפסות PIN / MGN-ITC נמצאו בכל רחבי אשכול IMP, דבר המצביע על חלוקה הומוגנית בתוך התמחות postsynaptic (איור 5E). גבוה משמעותית (p מבחן t מזווג <0.018) צפיפות של תיוג AMPA-R נצפתה PIN / MGN הסינפסות על קוצים ITC (715 ± 38 חלקיקי זהב / מיקרומטר 2, n = 32) לעומת סינפסות על דנדריטים ITC (590 ± 44 חלקיקי זהב / מיקרומטר 2, n = 32). בסך הכל, צפיפות-R AMPA ב PIN / סינפסות MGN-ITC הראו נמוך יחסיתשונה (מקדם שונה, CV = 0.37) עולה בקנה אחד עם חלוקה הומוגנית.
Immunoparticles עבור NMDA-R ב הסינפסות PIN / MGN-ITC לעתים קרובות נצפו התחלק באופן אחיד בתוך האשכולות IMP postsynaptic (איור 5). צפיפות תיוג NMDA-R היה דומה (עמ 'מבחן t מזווג = 0.39) בין PIN / MGN הסינפסות על קוצים ITC (חלקיקי זהב 1070 ± 153 / מיקרומטר 2, n = 8) דנדריטים ITC (812 ± 183 חלקיקי זהב / מיקרומטר 2, n = 9). בניגוד למה נצפה-R AMPA, הצפיפות של NMDA-R סינפסות PIN / MGN-ITC הייתה משתנית מאוד (CV = 0.54).

איור 6. AMPA-R ו NMDA-R immunolabeling ב Identified PIN / MGN-ITC סינפסות.
(AB) פרצופים שניים של ממברנות טרום postsynaptic של PIN / MGN-ITCסינפסה עשתה על גבי עמוד שדרה הדנדריטים שבו P הפנים של מסוף מבטא ChR2 (שכותרתו עם 15 חלקיקי זהב ננומטר) ואת PSD על גבי עמוד שדרה הדנדריטים מתויג עבור-R AMPA (5 חלקיקי זהב ננומטר). (CD) נופים מוגדלים של האזורים שמתווים הקו המקווקו. אזורים אלה כבר הסתובב כ 45 ° נגד כיוון השעון כדי לאפשר לראות טוב יותר של PSD. (E) Scatterplots של מספר החלקיקים AMPA-R לעומת שטח סינפטי קוצים דנדריטים ITC. בשני המבנים, מתאם חיובי נצפה. (F) Scatterplots של מספר החלקיקים NMDA-R נגד אזור סינפטי קוצים דנדריטים ITC. קשר חיובי משמעותי זוהה רק קוצים הדנדריטים. ברי סולם:. 500 ננומטר אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
{kind=link}
כיP-פני קרום התא presynaptic לעתים קרובות מעולף חלקית באשכול IMP-סינפטי, נוכל להעריך את אזור סינפטיים של רק 30% של סינפסות (קוצים: מתכוון באזור 0.032 מיקרומטר 2, רכס: 0.007 כדי 0,063 מיקרומטר 2, n = 8; דנדריטים: 0.047 מיקרומטר 2, רכס: 0.024 כדי 0.166 מיקרומטר 2, n = 11). אלה היו בסדר גודל דומה כמו glutamatergic telencephalic ניתח בעבר הסינפסות 25.
בשני קוצים דנדריטים, מספר immunoparticles זהב עבור AMPA-R סינפסות בודדות הייתה מתואמת חיובית עם השטח הסינפטי (ספירמן, קוצים: r = 0.88, דנדריטים: r = 0.60, p <0.0001) (איור 6E). לעומת זאת, מספר immunoparticles זהב-R NMDA נמצא בקורלציה עם שטח סינפטי קוצים (ספירמן, קוצים: r = 0.90, p <0.002), אך לא דנדריטים (r = 0.21, p = 0.29) (איור 6F ).
Discussion
במיקרוסקופ אלקטרוני Freeze-שבר כבר טכניקת מרכזי מחקר ultrastructural במשך 40 שנים. עם זאת, עדר אמצעי יעיל כדי ללמוד את ההרכב המולקולרי של ממברנות פיק ירידה משמעותית את השימוש בו. לאחרונה, חלה תחייה גדולה במיקרוסקופ אלקטרונים להקפיא שבר עקב ההתפתחות של דרכים יעילות כדי לחשוף חלבונים בממברנה אינטגרליים ידי immunogold תיוג 1, 2, כלומר את טכניקת FRIL.
טכניקת FRIL בעל מספר יתרונות על פני שיטות ultrastructural immunogold אחרות. ראשית, חלבונים נגישים בקלות לנוגדני הגדלת הרגישות. שנית, החשיפה של חלק גדול של התמחויות קרום הפלזמה, כגון הממברנה הפוסט-סינפטי, על משטח דו-ממדי של העתק מאפשר בדיקה של התפלגות מרחבית רצף פיזי של מולקולות של עניין ללא שיקום מייגע זמן רב של סריסעיפי ultrathin al. שלישית, על זמינותו של שני החצאים של קרום הפלזמה מגדילה את מספר חלבונים שיכולים להיות מתויג עבור כל אחד מהמבנים, נוגדנים מתאימים הניתנים זמינים. עם שבירה, הפנים ההידרופובי של הממברנה פיצול מצופה פלטינה פחמן כי משריש תחומים חלבון הנותרים על פני השטח שבורה. הדבר מונע גישה של נוגדנים לאנטיגנים בתחומים אלה. למשל על P-פני העתק רק אפיטופים וצפו בשטח protoplasmic יכולים להיות מזוהים על ידי נוגדנים, ואילו על E הפנים רק אפיטופים וצפו בשטח exoplasmic יכול להיות מחויבת על ידי נוגדנים (ראה איור 2).
מצד שני, את הטכניקה FRIL סובל גם ממגבלות מסוימות 2. כמו שברים להתרחש באופן אקראי, זה עלול להיות קשה למקד תאים או מבנים ספציפיים. זה גם יכול להוביל הטית דגימה, למשל, באוסף סינפסה, כתוצאה מההסתברות השונה של fracturing לאורך הממברנה של מבנים עם עקמומיות שונה (למשל, עמוד השדרה לעומת פירים). יתר על כן, הקצאת חלבונים בממברנה לאחד שני הפרצופים הוא בלתי צפויה. לפיכך, פיזורו של חלבון אל P-פנים או E-הפנים, בייחוד עבור מחקרים כמותיים, צריך להיות בזהירות בחן באמצעות נוגדנים תגובתי תחומים תאיים תאיים. לבסוף, זיהוי של העתק של מבנים מסוימים, כגון מסופי האקסון presynaptic, יכול להיות קשה כאשר רק על סמך תכונות מורפולוגיות. עם זאת, השימוש של נוגדנים ספציפיים עבור חלבוני סמן או התמרה של חלבונים בממברנה נפרדים מתויגים או ערוצים באמצעות וקטורים ויראליים מציע כלים נוספים כדי להקל על זיהוי של ממברנות השבורות. לדוגמה, במחקר זה ניצל התמרה של ChR2-YFP בנוירונים התלמוס לזהות efferents axonal שלהם באמיגדלה או תיוג לקולטני μ-אופיואידים לחשוף אותי postsynapticmbranes של נוירונים ITC.
על מנת לבצע את טכניקת FRIL בהצלחה, טיפול בפרט צריך לנקוט בנוגע קיבעון רקמות. קיבעון רקמות חזק (> 2% paraformaldehyde) יכול לגרום שיעור גבוה של שברי צלב וירידה ברגישות תיוג. מצד השני, קיבעונות חלשים להפוך את הטיפול ברקמה (למשל, חיתוך סעיפים) קשה. כמו כן, חשוב לשלוט כי העובי של אובניים הגזוז תואם את העובי של הקלטת דו צדדית. אם העובי של הדגימה נמוך מזה של הקלטת, המשטחים של הרקמה אולי לא לצרף אל פני השטח של שני נישאי המתכת, וכתוצאה מכך הדגימה קפואה אינה שבר. אם הרקמה עבה, זה יהיה דחוס עם עיוותים מבניות בלתי נמנעות כאשר הכריך של שתי הנושאות נחושת הוא עשה. הטמפרטורה שבה הדגימה הוא שבר (בפרוטוקול זה, -115 ° C) משחק גם תפקיד חשובעל המבנה של ההעתק. טמפרטורות גבוהות עשויות להניב שיעור גבוה יותר של חפצים כגון התעבות של אדי מים על פני השטח של הרקמה לפני או במהלך אידוי. בטמפרטורות נמוכות יותר (<-125 ° C) עלול להגביר את הסיכון של פיצול של החומר במהלך שבירה. חומר זה עלול ליפול על פני השטח של הדגימה או להישאר מחובר אליו. פתיתים אלה של חומרים הם גם מצופים בניגוד לייצר כתמים כהים בתמונה. שבירה בטמפרטורות נמוכות יכולה להשפיע גם על התדר של שברים צלבו במיוחד עבור מבני קנס קטנים כגון קוצים הדנדריטים. שלב קריטי נוסף בהכנת ההעתקים הוא עיכול חומר הניקוי. אם העיכול אינו שלם, הרקמה המעוכלת מופיעה כתמים כהים כמו על ההעתק, ערבבה בין הניתוח של המבנה על TEM. יתר על כן, הרקמה המעוכלת יכולה נוגדנים מלכודת או לאגד הלא ספציפית, הגדלת תיוג הרקע. מצד השני, השימוש של חומר ניקויים לעיכול רקמת יכול לפגל מולקולות השייכת לאוניברסיטה העתק שינוי מבני שניוני ושלישוני שלהם. לכן, עבור אנטיגנים מסוימים ייתכן שיהיה צורך בהדרגה לדלל את הריכוז של SDS עם צעדי כביסה נוספים.
לקבלת immunolabeling, הזמינות של גדלים שונים של חלקיקי זהב מצומדות כדי נוגדנים משני מאפשרת לזהות בעת ובעונה האחת, אבל רק מבחינה איכותית, חלבונים מרובים, אפילו microdomains הספציפי של קרום הפלזמה, כגון התמחות הפוסט-סינפטי. עם זאת, בשל הפרעה סטרית, מחקרים כמותיים מוגבלים בדרך כלל זיהוי של מולקולה אחת בלבד. הגודל של חלקיקי זהב יכול גם להשפיע על יעילות התיוג.
על הפרשנות של התיוג FRIL, זה צריך להישמר לזכור כי חלקיק immunogold יכול להיות ממוקם בכל מקום בתוך אונה עם רדיוס של 20-25 ננומטר מן אנטיגן בשל הצורה המורכבת הגמישהed ידי נוגדן ראשוני ומשני 26. למידע נוסף על התיאוריה והפרקטיקה של FRIL וטכנולוגיות רלוונטיות, אנו מפנים את הקורא גם למאמרים מתודולוגיות אחרות 27, 28.
טכניקת FRIL לאחרונה נעשתה שימוש עבור ניתוחים כמותיים ברזולוציה גבוהה של לוקליזציה קולטן הגלוטמט באוכלוסיות סינפסה מגוונות 29, 30. יתר על כן, את רגישות זיהוי של טכניקת FRIL עבור AMPA-R נאמדת גבוה ככל חלקיקי immunogold אחד לכל AMPA אחד הפונקציונלי -R ערוץ 29. לכן, גישה זו היא בסיכומו מאוד שימושי לכמת ולנתח את דפוס הביטוי postsynaptic של AMPA-R ו NMDA-R ב סינפסות המרכזי. הנה, הראינו את תחולתו ב סינפסות PIN / MGN-ITC, אתר והחשוב סביר להעברת ארה"ב מידע במהלך מיזוג פחד. באמצעות נוגדן העלה כנגד שאריות חומצת אמינו התאיות השמורות ביותר של היחידות משנה קולטן AMPA GluA1-GluA4, מצאנו חלוקה שווה של חלקיקי זהב בתוך האשכולות IMP המתאים התמחויות הממברנה הפוסט-סינפטי. הצפיפות-R AMPA ב קוצי ITC הייתה גבוהה באופן משמעותי לעומת סינפסות פירים כוונת של afferents התלמוס PIN / MGN. בשני סינפסות השדרה פיר, מתאם חיובי בין תיוג עבור AMPA-R ואזור postsynaptic זוהה, תכונה משותפת סינפסות glutamatergic אחרים 25. השונות הנמוכות בצפיפות של AMPA-R סינפסות PIN / MGN-ITC מראות התפלגות הומוגנית דומה סינפסות אחרים נוצרו על ידי efferents התלמוס 7, אבל שונה סינפסות קליפת מוח 25. לעומת זאת, צפיפות-R NMDA היה יותר משתנה ולא שונה בין הסינפסות השדרה פיר דבר המצביע על רגולציה שונה AMPA-R. בעתיד, השחזור הגבוה של טכניקת FRIL לא יאפשר רק להעריך את ההרכב המולקולרי הבסיסי של סינפסות מרכזית אך עשוי להקל זיהוי של גhanges במספרי קולטן ionotropic גלוטמט והפצת subsynaptic לאחר למידת פחד, המשלים הקלטות-vivo לשעבר של נכסים טרום postsynaptic של תשומות אלה.
לסיכום, גישה זו יכולה להיות בשימוש על ידי חוקרים אחרים כדי להשיג תובנות יחסי פונקציה-מבנה של סינפסות מעוררות ספציפי קלט רב מעגלים עצביים אחרים בם מתירים את מקורם של התשומות וכן אופי רכב אלמנטי postsynaptic הוא חיוני אך בעייתי .
Disclosures
The authors declare that they have no competing financial interests.
Acknowledgements
Funding was provided by the Austrian Science Fund FWF grant No. P-22969-B11 to F. Ferraguti, and by the Charitable Hertie Foundation and the Werner Reichardt Centre for Integrative Neuroscience and by the DFG (CIN-Exc. 307) to I. Ehrlich.
Materials
| Name | Company | Catalog Number | Comments |
| Surgery | |||
| Stereotactic frame | Stoelting, USA | 51670 | can be replaced by other stereotactic frame for mice |
| Steretoxic frame mouse adaptor | Stoelting, USA | 51625 | |
| Gas anesthesia mask for mice | Stoelting, USA | 50264 | no longer available, replaced by item no. 51609M |
| Pressure injection device, Toohey Spritzer | Toohey Company, USA | T25-2-900 | other pressure injection devices (e.g. Picospritzer) can be used |
| Kwik Fill glass capillaries | World Precision Instruments, Germany | 1B150F-4 | |
| Anesthesia machine, IsoFlo | Eickemeyer, Germany | 213261 | |
| DC Temperature Controler and heating pad | FHC, USA | 40-90-8D | |
| Horizontal Micropipette Puller Model P-1000 | Sutter Instruments, USA | P-1000 | |
| Surgical tool sterilizer, Sterilizator 75 | Melag, Germany | 08754200 | |
| rAAV-hSyn-ChR2(H134R)-eYFP (serotype 2/9) | Penn Vector Core, USA | AV-9-26973P | |
| fast green | Roth, Germany | 0301.1 | |
| Isoflurane Anesthetic, Isofuran CP (1ml/ml) | CP Pharma, Germany | ||
| Antiseptic, Betadine (providone-iodine) | Purdure Products, USA | BSOL32 | can be replaced by other disinfectants |
| Analgesic, Metacam Solution (5mg/ml meloxicam) | Boehringer Ingelheim, Germany | can be replaced by other analgesics | |
| Bepanthen eye ointment | Bayer, Germany | 0191 | can be replaced by other eye ointments |
| Drill NM3000 (SNKG1341 and SNIH1681) | Nouvag, Switzerland | ||
| Sutranox Suture Needle | Fine Science Tools, Germany | 12050-01 | |
| Braided Silk Suture | Fine Science Tools, Germany | 18020-60 | |
| Name | Company | Catalog Number | Comments |
| Tissue preparation | |||
| Paraformaldehyde EM grade | Agar Scientific Ltd., United Kingdom | AGR1018 | |
| Saturated picric acid solution | Sigma-Aldrich, USA | P6744-1GA | |
| Na2HPO4-2H20 | Merck Millipore, Germany | 1065860500 | |
| NaH2PO4-2H2O | Merck Millipore, Germany | 1063451000 | |
| NaCl | Merck Millipore, Germany | 1064041000 | |
| 4N NaOH | Carl Roth, Germany | T198.1 | |
| Thiopental | Sandoz, Austria | 5,133 | |
| Glycerol | Sigma-Aldrich, USA | G5516-500ML | |
| GenPure ultrapure water system | Thermo Fisher Scientific, USA | 50131235 | |
| Peristaltic pump | ISMATEC, Germany | ISM 930C | |
| Filter Paper | MACHEREY-NAGEL, Germany | MN 615 1/4 | |
| Vibroslicer, VT1000S | Leica Microsystems, Austria | ||
| Ophthalmic scalpel | Alcon Laboratories, USA | can be replaced by other ophthalmic scalpels | |
| Perfusion cannula | Vieweg, Germany | F560088-1 | can be replaced by similar items from other companies |
| Name | Company | Catalog Number | Comments |
| High-pressure Freezing | |||
| Copper carriers | Engineering Office M. Wohlwend, CH | 528 | |
| Sidol Polish | Henkel, Germany | can be replaced by same item from other companies | |
| Chamois skin | Household supply store | ||
| Hole punch, 1,5mm | Stubai, Austria | can be replaced by same item from other companies | |
| Denatured ethanol | Donauchem, Austria | can be replaced by same item from other companies | |
| Aceton | Roth, Germany | 9372.5 | CAUTION! |
| High Pressure Freezing Machine HPM 010 | BalTec, CH; now Leica Microsystems | HPM010 | not produced any more, substituted by LeicaEM HPM100 |
| Stereo-microscope | Olympus, Japan | SZX10 | |
| Liquid nitrogen | CAUTION! | ||
| Cryo-vials | Roth, Germany | E309.1 | can be replaced by same item from other companies |
| CryoCane | Nalge Nunc International,USA | 5015-0001 | can be replaced by same item from other companies |
| CryoSleeve | Nalge Nunc International,USA | 5016-0001 | can be replaced by same item from other companies |
| Liquid nitrogen storage vessel | Cryopal, France | GT38 | can be replaced by same item from other companies |
| Non-ionic detergent (Lavocid) | Werner & Mertz Professional, Germany | ||
| Name | Company | Catalog Number | Comments |
| Freeze-fracture and Replication | |||
| Sandblaster, Mikromat 200-1 | JOKE Joisten & Kettenbaum, Germany | SANDURET 2-K | can be replaced by same item from other companies |
| Siliciumcarbid SIC 360, grain size 25 - 21µ | JOKE Joisten & Kettenbaum, Germany | 955932 | |
| Freeze Fracture System BAF 060 | BalTec, CH; now Leica Microsystems | BAF060 | |
| Ceramic 12 well plate | Gröpel, Austria | 14511 | can be replaced by same item from other companies |
| Trizma base | SIGMA, USA | T1503 | can be replaced by same item from other companies |
| Trizma hydrochloride | SIGMA, USA | T3253 | can be replaced by same item from other companies |
| Sodium chloride | Merck, Germany | 1,064,041,000 | can be replaced by same item from other companies |
| SDS, Sodium lauryl sulfate | Roth, Germany | 5136.1 | CAUTION! ; can be replaced by same item from other companies |
| Sucrose | Merck, Germany | 1,076,871,000 | can be replaced by same item from other companies |
| TRIS | Roth, Germany | 5429.3 | can be replaced by same item from other companies |
| Universal Hybridization Oven | Binder, Germany | 7001-0050 | can be replaced by same item from other companies |
| Name | Company | Catalog Number | Comments |
| Immunolabelling | |||
| BSA | SIGMA, USA | A9647 | can be replaced by same item from other companies |
| Anti-GFP Antibody | Molecular Probes, USA | A11122 | |
| Anti-pan-AMPAR Antibody | Frontier Institute, Japan | pan AMPAR-GP-Af580-1 | |
| Anti-NMDAR1 Antibody, clone 54.1 | Merck Millipore, Germany | MAB363 | |
| Opioid Receptor-Mu (MOR) Antibody | ImmunoStar, USA | 24216 | |
| EM goat anti-guinea pig, 5nm; secondary antibody | BBInternational, | EM.GAG5 | |
| EM goat anti-rabbit, 15nm; secondary antibody | BBInternational, | EM.GAR15 | |
| Donkey anti-rabbit, 10nm, secondary antibody | AURION, Netherlands | DAR 10nm | |
| Copper grids, 100 Parallel Bar | Agar scientific, UK | G2012C | |
| Incubator | Major Science, USA | MO-RC | can be replaced by same item from other companies |
| Pioloform Powder | Agar scientific, UK | R1275 | |
| Chloroform | Roth, Germany | 3313.1 | CAUTION! ; can be replaced by same item from other companies |
| Name | Company | Catalog Number | Comments |
| EM analysis | |||
| Philips CM120 TEM | Philips/FEI | ||
| Morada CCD camera | Soft Imaging Systems, Germany | ||
| iTEM Ver. 5.2, imaging software | Soft Imaging Systems, Germany | ||
References
- Fujimoto, K. SDS-digested freeze-fracture replica labeling electron microscopy to study the two-dimensional distribution of integral membrane proteins and phospholipids in biomembranes: practical procedure, interpretation and application. Histochem Cell Biol. 107 (2), 87-96 (1997).
- Masugi-Tokita, M., Shigemoto, R. High-resolution quantitative visualization of glutamate and GABA receptors at central synapses. Curr Opin Neurobiol. 17 (3), 387-393 (2007).
- Rash, J. E., Yasumura, T. Direct immunogold labeling of connexins and aquaporin-4 in freeze-fracture replicas of liver, brain, and spinal cord: factors limiting quantitative analysis. Cell Tissue Res. 296 (2), 307-321 (1999).
- Emes, R. D., Grant, S. G. Evolution of synapse complexity and diversity. Annu Rev Neurosci. 35, 111-131 (2012).
- Matsuzaki, M., et al. Dendritic spine geometry is critical for AMPA receptor expression in hippocampal CA1 pyramidal neurons. Nat Neurosci. 4 (11), 1086-1092 (2001).
- Rollenhagen, A., Lübke, J. H. The morphology of excitatory central synapses: from structure to function. Cell Tissue Res. 326 (2), 221-237 (2006).
- Tarusawa, E., et al. Input-specific intrasynaptic arrangements of ionotropic glutamate receptors and their impact on postsynaptic responses. J Neurosci. 29 (41), 12896-12908 (2009).
- Nusser, Z., et al. Cell type and pathway dependence of synaptic AMPA receptor number and variability in the hippocampus. Neuron. 21 (3), 545-559 (1998).
- Nyìri, G., Stephenson, F. A., Freund, T. F., Somogyi, P. Large variability in synaptic N-methyl-D-aspartate receptor density on interneurons and a comparison with pyramidal-cell spines in the rat hippocampus. Neuroscience. 119 (2), 347-363 (2003).
- Nicholson, D. A., Geinisman, Y. Axospinous synaptic subtype-specific differences in structure, size, ionotropic receptor expression, and connectivity in apical dendritic regions of rat hippocampal CA1 pyramidal neurons. J Comp Neurol. 512 (3), 399-418 (2009).
- Millhouse, O. E. The intercalated cells of the amygdala. J Comp Neurol. 247 (2), 246-271 (1986).
- Busti, D., et al. Different fear states engage distinct networks within the intercalated cell clusters of the amygdala. J Neurosci. 31 (13), 5131-5144 (2011).
- Amano, T., Unal, C. T., Paré, D. Synaptic correlates of fear extinction in the amygdala. Nat Neurosci. 13 (4), 489-494 (2010).
- Duvarci, S., Pare, D. Amygdala microcircuits controlling learned fear. Neuron. 82 (5), 966-980 (2014).
- Jüngling, K., et al. Neuropeptide S-mediated control of fear expression and extinction: role of intercalated GABAergic neurons in the amygdala. Neuron. 59 (2), 298-310 (2008).
- Asede, D., Bosch, D., Lüthi, A., Ferraguti, F., Ehrlich, I. Sensory inputs to intercalated cells provide fear-learning modulated inhibition to the basolateral amygdala. Neuron. 86 (2), 541-554 (2015).
- Maren, S. Neurobiology of Pavlovian fear conditioning. Annu Rev Neurosci. 24, 897-931 (2001).
- Pape, H. C., Pare, D. Plastic synaptic networks of the amygdala for the acquisition, expression, and extinction of conditioned fear. Physiol Rev. 90 (2), 419-463 (2010).
- Sigurdsson, T., et al. Long-term potentiation in the amygdala: a cellular mechanism of fear learning and memory. Neuropharmacology. 52 (1), 215-227 (2007).
- Bosch, D., Asede, D., Ehrlich, I. Ex-vivo optogenetic dissection of fear circuits in brain slices. J. Vis. Exp. (110), e53628 (2016).
- Fenno, L., Yizhar, O., Deisseroth, K. The development and application of optogenetics. Annu Rev Neurosci. 34, 389-412 (2011).
- Bienvenu, T. C. M., et al. Large intercalated neurons of amygdala relay noxious sensory information. J. Neurosci. 35 (5), 2044-2057 (2015).
- Sandri, C., Akert, K., Livingston, R. B., Moor, H. Particle aggregations at specialized sites in freeze-etched postsynaptic membranes. Brain Res. 41 (1), 1-16 (1972).
- Likhtik, E., Popa, D., Apergis-Schoute, J., Fidacaro, G. A., Paré, D. Amygdala intercalated neurons are required for expression of fear extinction. Nature. 454 (7204), 642-645 (2008).
- Fukazawa, Y., Shigemoto, R. Intra-synapse-type and inter-synapse-type relationships between synaptic size and AMPAR expression. Curr Opin Neurobiol. 22 (3), 446-452 (2012).
- Amiry-Moghaddam, M., Ottersen, O. P. Immunogold cytochemistry in neuroscience. Nat Neurosci. 16 (7), 798-804 (2013).
- Fukazawa, Y., Masugi-Tokita, M., Tarusawa, E., Hagiwara, A., Shigemoto, R., Cavalier, A., et al. SDS-digested Freeze-fracture replica labelling (SDS-FRL). Handbook of Cryo-Preparation Methods for Electron Microscopy. , 567-586 (2007).
- Severs, N. J. Freeze-fracture electron microscopy. Nat Protoc. 2 (3), 547-576 (2007).
- Tanaka, J., et al. Number and density of AMPA receptors in single synapses in immature cerebellum. J Neurosci. 25 (4), 799-807 (2005).
- Mansouri, M., et al. Distinct subsynaptic localization of type 1 metabotropic glutamate receptors at glutamatergic and GABAergic synapses in the rodent cerebellar cortex. Eur J Neurosci. 41 (2), 157-167 (2015).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved