
Method Article
גישות בינה מלאכותית להערכת ריסים ראשוניים
In This Article
Summary
השימוש בבינה מלאכותית (Ai) לניתוח תמונות מתגלה כגישה חזקה, פחות מוטה ומהירה בהשוואה לשיטות נפוצות. כאן אימנו את Ai לזהות אברון תאי, ריסים ראשוניים, ולנתח תכונות כגון אורך ועוצמת הכתמים באופן קפדני ושחזורי.
Abstract
Cilia הם נספחים תאיים מבוססי מיקרוטובול המתפקדים כמרכזי איתות למגוון מסלולי איתות בסוגי תאי יונקים רבים. אורך Cilia נשמר מאוד, מוסדר היטב, ומשתנה בין סוגי תאים ורקמות שונים והיה מעורב בהשפעה ישירה על יכולת האיתות שלהם. לדוגמה, הוכח כי cilia משנה את אורךם בתגובה להפעלת קולטנים מצמידי חלבון G. עם זאת, מדידה מדויקת ורבייה של ריסים רבים היא הליך גוזל זמן רב ועבודה אינטנסיבית. גישות נוכחיות הן גם שגיאה והטיה נוטה. ניתן להשתמש בתוכניות בינה מלאכותית (Ai) כדי להתגבר על אתגרים רבים אלה בשל יכולות המאפשרות התבוללות, מניפולציה ואופטימיזציה של ערכות נתונים נרחבות. כאן, אנו מדגימים כי מודול Ai ניתן לאמן לזהות cilia בתמונות הן בדגימות vivo והן במבחנה. לאחר שימוש ב- Ai מאומן לזיהוי ריסים, אנו מסוגלים לעצב ולנצל במהירות יישומים המנתחים מאות ריסים במדגם יחיד לאורך, עוצמת פלואורסצנטיות ולוקליזציה משותפת. גישה בלתי משוחדת זו הגבירה את הביטחון והקפדנות שלנו כאשר משווים דגימות מהקדמה עצבית ראשונית שונה במבחנה, כמו גם על פני אזורי מוח שונים בתוך בעל חיים ובין בעלי חיים. יתר על כן, טכניקה זו יכולה לשמש כדי לנתח באופן אמין את הדינמיקה cilia מכל סוג תא ורקמות באופן תפוקה גבוהה על פני דגימות מרובות וקבוצות טיפול. בסופו של דבר, גישות מבוססות Ai צפויות להפוך לסטנדרט ככל שרוב השדות נעים לעבר גישות פחות מוטות ויותר ניתנות לשחזור לרכישת וניתוח תמונות.
Introduction
ריסים עיקריים הם אברונים חושיים הבולטים מרוב סוגי תאי היונקים1,2,3,4. הם בדרך כלל נספחים בודדים קריטי לתיאום מסלולי איתות תאים מגוונים על ידי שילוב אותות חוץ תאיים5,6,7. ריסים ראשוניים ממלאים תפקידים חשובים במהלך התפתחות עוברית והומאוסטזיס של רקמות בוגרות, ושיבוש תפקודם או המורפולוגיה שלהם קשור למספר הפרעות גנטיות, הנקראות יחד ciliopathies. בשל האופי הכמעט בכל מקום של ריסים, ciliopathies קשורים עם מגוון רחב של תכונות קליניות שיכולות להשפיע על כל מערכות האיברים8,9,10,11,12. במודלים בעלי חיים של ciliopathies, אובדן מבנה ciliary או יכולת איתות מתבטא במספר פנוטיפים רלוונטיים קלינית כולל השמנת יתר הקשורים היפרפגיה3,13,14,15. במערכות מודל רבות, שינויים באורך ריסים הוכחו להשפיע על יכולת האיתות שלהםפונקציות 16,17,18,19. עם זאת, ישנם מספר אתגרים ארוכי זמן וטכניים הקשורים להערכה מדויקת ורבייה של אורך הריסים והרכב.
מערכת העצבים המרכזית של היונקים הבוגרים (CNS) היא הקשר ביולוגי אחד שהציב אתגר להבנת מורפולוגיה ותפקוד ריסים. אמנם נראה כי נוירונים ותאים ברחבי מערכת רה"מ מחזיקים ריסים, בשל הכלים המוגבלים והיכולות להתבונן ולנתח את הריסים האלה הבנה של הפונקציות שלהם נשאר חמקמק20. לדוגמה, סמן הריסים הרב-טיפוס, אצטילאט α-טובולין, אינו מתייג ריסים עצביים20. הקושי לחקור את הריסים האלה נפתר בחלקו עם גילוי של מספר קולטנים מצמידי חלבון G (GPCR), מכונות איתות וחלבונים הקשורים לממברנה המועשרים על הממברנה של ריסים עצביים21,22. כל התצפיות הבסיסיות הפשוטות הללו מרמזות על החשיבות והמגוון של ריסים של CNS, אשר עד כה נראה שאין שני לו על ידי רקמות אחרות. לדוגמה, וריאציה באורך סיליה ולוקליזציה GPCR ניתן לראות בכל המוח, עם אורכים בגרעין עצבי מסוים להיות שונה בהשוואה גרעין אחרים19,23. באופן דומה, התוכן GPCR שלהם מחמאה מכונות איתות להראות גיוון המבוסס על מיקום נוירואנטומי וסוג עצבי2,24,25,26,27,28,29. תצפיות פשוטות אלה ממחישות כי אורך הריסים של היונקים והרכבם מוסדרים היטב, בדיוק כמו באורגניזמים מודל, כמו Chlamydomonas reinhardtii, אבל ההשפעה של הבדלי אורך אלה על תפקוד cilia, איתות ובסופו של דבר התנהגות עדיין לא ברור16,30,31,32.
מדידה מדויקת של אורך הריסים והרכבה מתגלה כאתגר טכני שנוטה לשגיאת משתמש וחוסר תוכחות. כיום cilia ב vivo ו במבחנה מזוהים לרוב באמצעות גישות אימונופלואורסצנטיות המגדירות חלבונים סיליאריים או כתב פלואורסצנטי מועשר ריסים אללס33,34,35. האורכים של ריסים מתויגים פלואורסצנטיים אלה נמדדים לאחר מכן מתמונה דו-ממדית (דו-ממדית) באמצעות כלי מדידת קו בתוכניות ניתוח תמונה כגון ImageJ36. תהליך זה הוא לא רק מייגע ועבודה אינטנסיבית, אלא גם נוטה הטיה וטעייה. אותם מכשולים מתעוררים בעת מדידת עוצמות ריסים, המסייעים להצביע על שינויים במבנהריסים 37. כדי למזער את חוסר העקביות בסוגים אלה של ניתוחי תמונה, תוכניות בינה מלאכותית (Ai) הופכות נפוץ יותר ובמחיר סביר אפשרויות38.
Ai הוא קידום של מערכות מחשב המשתמשות ביתרון של אלגוריתמים ממוחשבים ותכנות לביצוע משימות שבדרך כלל דורשות אינטליגנציה אנושית39. מכשירי Ai לומדים לתפוס דפוסים, פרמטרים ומאפיינים חוזרים ולנקוט פעולות כדי למקסם את הסיכויים ליצירת תוצאות מוצלחות. Ai הוא רב-תכליתי וניתן לאמן אותו לזהות אובייקטים או מבנים ספציפיים בעלי עניין, כגון cilia, ולאחר מכן להיות מתוכנת להפעיל מגוון של ניתוחים על האובייקטים המזוהים. לכן, נתוני תמונה מורכבים יכולים להיווצר במהירות ובשחזור על ידי Ai38. אוטומציה וניתוח Ai של תמונות שנתפסו יגבירו את היעילות והיעילות תוך הגבלת כל טעות אנוש פוטנציאלית והטיה39. יצירת מתודולוגיה מבוססת Ai לזיהוי ריסים יוצרת דרך עקבית עבור כל קבוצות המחקר לנתח ולפרש נתוני ריסים.
כאן אנו משתמשים במודול Ai כדי לזהות את הריסים הן ב- vivo והן במבחנה בתמונות 2D. באמצעות קבוצה של תמונות לדוגמה, ה- Ai מאומן לזהות ריסים. לאחר השלמת האימון, Ai המיועד משמש להחלת מסיכה בינארית על ריסים המזוהים על Ai בתמונה. הקבצים הבינאריים המוחלים על ידי Ai ניתנים לשינוי, במידת הצורך, כדי להבטיח שכל הריסים בתמונות מזוהים כראוי וזיהוי לא ספציפי מסולק. לאחר ניצול Ai כדי לזהות cilia, תוכניות ניתוח כללי שנבנה בהתאמה אישית (GA) משמשים לביצוע ניתוחים שונים כגון מדידת אורך cilia ועוצמת פלואורסצנטיות. הנתונים הנאספים מיוצאים לטבלה שניתן לקרוא, לפרש ולהשתמש בה בקלות עבור ניתוחים סטטיסטיים (איור 1). השימוש בטכנולוגיה אוטומטית ובבינה מלאכותית לזיהוי ריסים וקבלת מדידות ספציפיות בין קבוצות ניסוי יסייע במחקרים עתידיים שמטרתם להבין את ההשפעה של תפקוד ריסים של מערכת הסביבה ומורפולוגיה על תקשורת והתנהגות תאים-תאים.
Protocol
1. לרכוש תמונות גולמיות
- לתקן ודגימות אימונובל לפי הצורך20.
- ריסים של תמונה באמצעות מיקרוסקופ קונפוקלי בעומק סיביות מרבי באמצעות אותו גודל פיקסל עם רזולוציית Nyquist.
- יצא תמונות כקבצים בתבנית תמונה מתויגת בשחור-לבן (.tif).
הערה: פרוטוקול זה מתאר כיצד להשתמש במודול AI במיוחד בתוך תוכנת רכיבי NIS. אם תמונות נרכשו כקבצי .nd2, ייצוא תמונות כקבצי .tif אינו נחוץ והמשתמש יכול להמשיך ישירות לשלב 2.3. אם נרכשו תמונות במערכת אחרת, ניתן לרכוש רישיון רכיבי NIS בנפרד וניתן להמיר קבצי .tif כמתואר בשלבים הבאים.
2. לאמן Ai לזהות cilia
- פתח את ערכת נתוני האימון.
- בחר כ- 50 תמונות לדוגמה עם לפחותסיליום אחד לכל מסגרת כדי לאמן את התוכנה ולהעתיק אותן בתיקיה אחת. תיקיה זו משמשת לכוון את התוכנה בעת פתיחת התמונות. פתח 50 מסגרות אלה במסמך ND2 יחיד עם לפחותסיליום אחד לכל מסגרת. בחר קובץ > ייבוא/ייצוא > יצירת קובץ ND מרצף קבצים.
- בחר את התיקיה המכילה את ערכת נתוני האימון. פעולה זו תפתח את רשימת הקבצים במרכז חלון הדו-שיח. הגדר באופן ידני את ארגון הקבצים באמצעות אפשרות אחת לפחות בתפריט הנפתח לעיל. האפשרויות הן ריבוי נקודות (עבור קבצי הקרנה מרביים מרובים), סדרת Z (לתמונת מחסנית z), זמן (לתמונה של זמן-לשגות) ואורך גל (עבור קבצים מערוצים מרובים).
- הזן את הערכים המספריים המתאימים תחת כל אפשרות שנבחרה. בחר ללא בכל מקום שבו לא נבחרו אפשרויות. לחץ על המר כדי לפתוח את מסמך ND.
- כייל תמונות.
- הזנת גודל הפיקסלים בפינה השמאלית התחתונה של התמונה: לחץ באמצעות לחצן העכבר הימני על מסמך > לכייל מסמך > גודל פיקסל.
- זהה ריסים.
- זהה את הריסים באופן ידני על-ידי מעקב מדויק אחר מבני סילארי בודדים בכל המסגרות הפתוחות באמצעות זיהוי אוטומטי או ציור אובייקט בסרגל הכלים הבינארי. פעולה זו תמשוך מסיכות בינאריות על אובייקטי העניין. קבצים הבינאריים האלה ישמשו כאובייקטים לדוגמה לאימון התוכנה לזיהוי ריסים על מאפיינים מבוססי פיקסלים בניתוח תמונה ניסיוני עתידי. בחר הצג פקדי ניתוח > > סרגל הכלים הבינארי > צייר אובייקט.
הערה: הסר כל מסגרת שאין בה מסגרות הקבצים הבינאריים מכיוון שהתוכנה לא תתחיל להתאמן אלא אם כן היא מסוגלת לזהות מסגרות ינריות בכל המסגרות הפתוחות.
- זהה את הריסים באופן ידני על-ידי מעקב מדויק אחר מבני סילארי בודדים בכל המסגרות הפתוחות באמצעות זיהוי אוטומטי או ציור אובייקט בסרגל הכלים הבינארי. פעולה זו תמשוך מסיכות בינאריות על אובייקטי העניין. קבצים הבינאריים האלה ישמשו כאובייקטים לדוגמה לאימון התוכנה לזיהוי ריסים על מאפיינים מבוססי פיקסלים בניתוח תמונה ניסיוני עתידי. בחר הצג פקדי ניתוח > > סרגל הכלים הבינארי > צייר אובייקט.
- רכבת Ai.
- התחל לאמן את התוכנה. זה יפתח את תיבת Segment.ai הרכבת. בחר Segment.ai רכבת NIS.ai >.
- בתיבה Segment.ai רכבת, בחר את ערוץ המקור שישמש להכשרה. אם קבצים מערוצים מרובים פתוחים, בחר ערוץ אחד בלבד כערוץ המקור. לאחר מכן בחר את הקבצים הבינאריים המתאימים של האמת הקרקעית שעליהם יש לאמן את הבינה מלאכותית. לבסוף בחר את מספר האיטרציות הדרושות כדי לאמן את ה- Ai בהתאם לגודל ולהתפלגות של הקבצים הבינאריים.
הערה: אם הקבצים הבינאריים ניתנים לזיהוי בקלות מהסביבה ומופצים היטב ברחבי התמונה, ייתכן שהתוכנה תזדקק לפחות מ-1000 איטרציות כדי לעבור הכשרה לזיהוי התמונות. אם לתמונות יש יחס אות לרעש נמוך, מומלץ להפעיל לפחות 1000 איטרציות תוך כדי אימון כדי לאפשר ל- Ai לזהות ריסים בדגימות בדיקה בביטחון גבוה. - בחר את תיקיית היעד כדי לשמור את קובץ ה- Ai המאומן ( .sai) ולחץ על התאמן כדי לאמן את התוכנה. התוכנה תמשיך כעת לאמן את עצמה לזהות ריסים בהתבסס על הקבצים הבינאריים במעקב. תהליך זה אורך מספר שעות.
הערה: בעת אימון, התוכנה תציג גרף המציג אובדן אימון. הגרף יציג בתחילה עלייה חדה לפני התחדדות באופן אידיאלי סביב 1% הפסד שבו הוא מישורים למשך שארית האימונים. שמור את הגרף לעיון עתידי על-ידי סימון התיבה עבור שמור גרף צילום מסך בתיבה Segment.ai רכבת(איור משלים 1). - אם נדרש שיפור נוסף בהכשרה, המשך להתאמן באותה ערכת נתונים. לחלופין, התאמן על ערכת נתונים חדשה עם אותם פרמטרים בדיוק. לא מומלץ להכשיר Ai שכבר הוכשר על ערכת נתונים חדשה עם פרמטרים שונים או אובייקטים שונים של עניין. בחר segment.ai > התאמן המשך להתאמן > בחר קובץ Ai מאומן.
3. לזהות ריסים באמצעות Ai מאומן
- פתח את ערכת הנתונים הניסיונית.
- פתח את תמונות הקונפוקליות הניסיוניות של cilia בתוכנה על-ידי המרת קבצי .tif לדוגמה לקבצי .nd2, בדומה לשלב 2.1. בחר קובץ > ייבוא/ייצוא > יצירת קובץ ND מרצף קבצים.
הערה: התמונות צריכות להיות באותו גודל פיקסל כמו אלה המשמשות לאימון ה- Ai. אם התמונות כבר בתבנית ND2, דלג לשלב 3.3.
- פתח את תמונות הקונפוקליות הניסיוניות של cilia בתוכנה על-ידי המרת קבצי .tif לדוגמה לקבצי .nd2, בדומה לשלב 2.1. בחר קובץ > ייבוא/ייצוא > יצירת קובץ ND מרצף קבצים.
- כייל תמונות.
- הזינו את גודל הפיקסלים בפינה השמאלית התחתונה של התמונה. לחץ באמצעות לחצן העכבר הימני על מסמך > לכייל > גודל פיקסל.
- הפעל את ה- Ai המאומן בקבצים הפתוחים.
- התחל לזהות את הריסים באמצעות Ai. התוכנה תצייר כעת binaries על cilia בהתבסס על ההכשרה שהיא קיבלה בשלב הקודם. תהליך זה ייקח כמה שניות. בחר NIS.ai > Segment.ai.
הערה: התוכנה תתבקש לבחור את הערוץ אם ערוצים מרובים פתוחים. הערוצים רשומים בשמותיהם כאן. אם לא, התיבה המסומנת 'מונו' תיבחר באופן אוטומטי.
- התחל לזהות את הריסים באמצעות Ai. התוכנה תצייר כעת binaries על cilia בהתבסס על ההכשרה שהיא קיבלה בשלב הקודם. תהליך זה ייקח כמה שניות. בחר NIS.ai > Segment.ai.
- בדוק תמונות עבור עבירות עבירות מזוהות שגויות.
- לאחר שה- Ai זיהה ריסים ומסמכים binaries מצוירים, בדוק את התמונות עבור כל אובייקטים שזוהו בטעות. אם תרצה, מחק באופן ידני כל הקבצים הבינאריים שלא זוהו באופן שגוי. בחר הצג פקדי ניתוח > > סרגל הכלים הבינארי > מחק אובייקט.
4. מדידת אורך ועוצמה של ריסים
- צור מתכון חדש לניתוח כללי 3 (GA3).
- כעת, לאחר שהריסים זוהו ומפולחים, המשיכו לנתח פרמטרים שונים של ריסים כגון אורכים ועוצמות באמצעות הכלי GA3. פעולה זו תפתח חלון חדש עם שטח ריק במרכז שבו יוגדר הניתוח. בחר תמונה > מתכון GA3 חדש.
- בחר את הקבצים הבינאריים לניתוח.
- מכיוון שהריסים כבר מפולחים באמצעות Segment.ai, GA3 יזהה באופן אוטומטי את הקבצים הבינאריים המסומנים כראוי בהתאם ל- Ai ויכלול את הצומת. בחר'מסגרות הקבצים הבינאריים > Detect_AI אוטומטית' או 'מסגרות בין-אריות > צייר Object_AI'.
- בחר את הערוצים הדרושים לניתוח. GA3 גם יזהה באופן אוטומטי את הערוצים בתמונות ויציג את הכרטיסיות שלהם תחת ערוצים.
- הסר אובייקטים הנוגעים בגבול המסגרת.
- מאז Ai יהיה לפלח את כל cilia כמו אובייקטים במסגרת, זה גם יזהה cilia שלם לאורך הקצוות של המסגרת. ניתן להסיר אובייקטים אלה באופן ידני בשלב 3.4 או להסיר אותם באופן אוטומטי ב- GA3. בחר עיבוד בינארי > הסר אובייקטים > גבולות נוגעים ללב.
- בחר פרמטרים למדידת ריסים.
- גרור ושחרר את הפרמטרים כדי למדוד כגון אורך ריסים (אורך) ועוצמות (סיכום עוצמת אובייקט). חבר את הצמתים לצומת הבינארי המתאים (חיבור A) ולצומת הערוצים (חיבור B). רחף מעל חיבור הצומת עבור תיאור כלי כדי להציג לאיזה חיבור שייך הצומת. בחרו 'מדידה' > גודל עצם > אורך ומדידה > עוצמת העצם > 'עוצמת Obj של Sum Obj' .
הערה: אורך הצומת מתחבר רק לצומת הבינארי, ואילו עוצמת Sum Obj מתחברת הן לצמתי הבינארי והן לצמתי הערוץ.
- גרור ושחרר את הפרמטרים כדי למדוד כגון אורך ריסים (אורך) ועוצמות (סיכום עוצמת אובייקט). חבר את הצמתים לצומת הבינארי המתאים (חיבור A) ולצומת הערוצים (חיבור B). רחף מעל חיבור הצומת עבור תיאור כלי כדי להציג לאיזה חיבור שייך הצומת. בחרו 'מדידה' > גודל עצם > אורך ומדידה > עוצמת העצם > 'עוצמת Obj של Sum Obj' .
- צרף את המידות לטבלה אחת.
- שלב את כל המידות בטבלת פלט בודדת על-ידי גרירה ושחרור של הצומת הוסף עמודה לתרשים זרימת הניתוח וחבר אותה לצמתי המדידה, אורך ועוצמת Obj Sum. בחר > > בסיסית .
- למדוד ריסים.
- מדוד ריסים על-ידי לחיצה על הפעל. תהליך זה לוקח כמה רגעים כדי למדוד את כל cilia בתמונות ניסיוניות. האורכים והמעוצמות יופיעו בחלון תוצאות ניתוח חדש.
הערה: הטבלה יכולה לפעמים לכלול נתונים ממסכות ש- Ai זיהה כריסים אך היו קטנות מכדי להתגלות על ידי העין האנושית וחוסלו בשלב 3.4. ניתן להסיר אובייקטים אלה מערכת הנתונים באמצעות מסנן לפני ניתוח סטטיסטי. כאן, מסנן של 1 מיקרומטר שימש למדידות אורך במבחנה בסיליה באיור 2 ו-2 מיקרומטר עבור cilia in vivo. ניתן לעשות זאת לפני ייצוא נתונים באמצעות המסלול שלהלן. בחר חלון תוצאות ניתוח > הגדר מסנן > הזן ערך > השתמש במסנן.
- מדוד ריסים על-ידי לחיצה על הפעל. תהליך זה לוקח כמה רגעים כדי למדוד את כל cilia בתמונות ניסיוניות. האורכים והמעוצמות יופיעו בחלון תוצאות ניתוח חדש.
- יצא נתונים לניתוח סטטיסטי.
5. לימודי קולוקליזציה
הערה: ניתוח Colocalization יכול להיכלל באותו מתכון GA3 המשמש למדידות של אורך cilia וניתוח אינטנסיביות. אם משתמשים באותו מתכון, פתחו קבצים כמתואר להלן ומדדו את האורכים והמשעי של שני הערוצים יחד עם מקדמי הלוקסלציה באותו צינור ניתוח.
- פתח את ערכת הנתונים הניסיונית.
- פתח את תמונות הקונפוקליות הניסיוניות של ריסים בתוכנה על-ידי המרת קבצי .tif לדוגמה לקבצי .nd2. בחר קובץ > ייבוא/ייצוא > יצירת קובץ ND מרצף קבצים.
- בחלון המוקפץ, בחר את קבצי המונוכרום בעומק של 16 סיביות מכל ערוצי העניין מסייר החלונות הממוקם בעמודה הראשונה של החלון המוקפץ. בחר ריבוי נקודות או סדרת Z מהתפריט הנפתח הראשון והזן ערך המתאים למספר הכולל של תמונות או ערימות, בהתאמה.
- בתיבה הנפתחת השניה, בחר אורך גל ושנה את הערך למספר הערוצים הכולל בתיקיה. התוכנה תבטל באופן אוטומטי חלון בחירה של Waveleleth הממוקם בקצה השמאלי התחתון של החלון המוקפץ. השתמש בתפריט הנפתח צבע כדי לבחור את הצבע של כל ערוץ. ספק לכל ערוץ שם אחר תחת העמודה שם. לאחר עדכון כל המידע, לחץ על המר. התוכנה תיצור באופן אוטומטי קובץ תמונה כל כיסוי עם כל התמונות הבודדות מכל הערוצים שנבחרו.
- כייל תמונות.
- הזינו את גודל הפיקסלים בפינה השמאלית התחתונה של התמונה. לחץ באמצעות לחצן העכבר הימני על מסמך > לכייל > גודל פיקסל.
- הפעל את Ai מאומן בערוץ הראשון.
- התחל לזהות את הריסים באחד הערוצים הפתוחים (למשל, ACIII; איור 5א) באמצעות Ai. התוכנה תצייר כעת binaries על ACIII שכותרתו cilia בהתבסס על ההכשרה שהיא קיבלה עבור ערוץ זה. תהליך זה ייקח כמה שניות. בחר NIS.ai > Segment.ai > ערוצי מקור > ACIII.
- הפעל את Ai מאומן בערוץ השני.
- התחל לזהות את הריסים בערוץ הפתוח האחר (למשל, MCHR1; איור 5B) באמצעות Ai. התוכנה תצייר כעת binaries על MCHR1 שכותרתו cilia בהתבסס על ההכשרה שהיא קיבלה עבור ערוץ זה. תהליך זה ייקח כמה רגעים. בחר NIS.ai > Segment.ai > ערוצי מקור > MCHR1.
- בדוק תמונות עבור עבירות עבירות מזוהות שגויות.
- לאחר שה- Ai יזהה ריסים ומסמכים עבירות עבירות, בדוק את התמונות אחר אובייקטים שזוהו באופן שגוי. מחק באופן ידני כל הקבצים הבינאריים שלא זוהו בטעות במידת הצורך. בחר הצג פקדי ניתוח > > סרגל הכלים הבינארי > מחק אובייקט.
- צור מתכון GA3 חדש.
- כעת, לאחר שהריסים זוהו ופולחו, המשיכו לניתוח ה-colocalization באמצעות הכלי GA3. פעולה זו תפתח חלון חדש עם שטח ריק במרכז שבו יוגדר הניתוח. חלון עם כל הקבצים הבינאריים הערוצים המזוהים ייווצר. ודא שכל הערוצים והקבצים הבינאריים הרצויים הדרושים לניתוח קיימים ונבחרים. בחר תמונה > מתכון GA3 חדש.
- הסר אובייקטים הנוגעים בגבול המסגרת.
- מכיוון שה- Ai יפלח את כל האובייקטים דמויי הסיליה במסגרת, הוא גם יזהה ריסים לא שלמים לאורך קצות המסגרת. ניתן להסיר אובייקטים אלה באופן ידני בשלב 5.5 או שניתן להסיר אותם באופן אוטומטי ב- GA3. בחר עיבוד בינארי > הסר אובייקטים > גבולות נוגעים ללב.
- הגדר את מסלול הקובקליזציה ב- GA3.
- כדי למדוד חפיפה של שני הערוצים בתוך ריסים, השתמש בקורלציה המקדם של מנדר. גרור ושחרר את צומת מקדם המנדרס לתוך השטח הריק של מתכון GA3 וחבר אותו בינארי מתאים ערוצים. כאן, 'חיבור A' מתחבר עם ACIII בינארי, 'חיבור B' עם ערוץ MCHR1 ו'חיבור C' עם ערוץ ACIII כדי לקבוע את החפיפה של MCHR1 בתוך ACIII בינארי. בחרו 'מדידה > יחס אובייקטים' > מקדם המנדרס.
הערה: התוכנה מאפשרת מדידת colocalization באמצעות מתאם מקדם פירסון באמצעות אותם שלבים כמתואר בפרוטוקולזה 40.
- כדי למדוד חפיפה של שני הערוצים בתוך ריסים, השתמש בקורלציה המקדם של מנדר. גרור ושחרר את צומת מקדם המנדרס לתוך השטח הריק של מתכון GA3 וחבר אותו בינארי מתאים ערוצים. כאן, 'חיבור A' מתחבר עם ACIII בינארי, 'חיבור B' עם ערוץ MCHR1 ו'חיבור C' עם ערוץ ACIII כדי לקבוע את החפיפה של MCHR1 בתוך ACIII בינארי. בחרו 'מדידה > יחס אובייקטים' > מקדם המנדרס.
- צרף את המידות לטבלה אחת.
- שלב את כל המידות בטבלת פלט אחת. בחר > > בסיסית .
- מדוד קולוקליזציה.
- מדוד ריסים על-ידי לחיצה על הפעל. תהליך זה לוקח כמה רגעים כדי למדוד את כל cilia בתמונות ניסיוניות. הנתונים יופיעו בחלון תוצאות ניתוח חדש.
- יצא נתונים לניתוח סטטיסטי.
תוצאות
הכשרת Ai לזיהוי ריסים
מדידה והערכה של אורך המבנה וההרכב של cilia יכולים להיות תהליך מייגע, גוזל זמן ונוטה לשגיאות. כאן, אנו משתמשים ב- Ai לפולוח מדויק של ריסים ממאגר גדול של תמונות ולנתח את האורכים והמאינטנסיביות שלהם באמצעות כלי ניתוח (איור 1). כל גישות ה- Ai דורשות צעדי הכשרה ליישום שלהן. הקמנו צינור הדרכה לזיהוי ריסים, שבוצע על ידי החלה ידנית של מסכות בינאריות על מבנים נוצצים. מידע זה משמש לאחר מכן לאימון הבינה מלאכותית בהתבסס על מאפייני פיקסלים תחת הקבצים הבינאריים המוחלים. כהנחיה כללית, ההכשרה כוללת את התוכנה העוברת מספר איטרציות, כ-1000, ונחשבת לאופטימלית אם שיעור אובדן האימונים או השגיאה נמוך מ-1%. עם זאת, מספר האיטרציות והשגיאות בתהליך האימון עשוי להשתנות בהתאם לתמונות לדוגמה המשמשות להכשרה. לדוגמה, לאחר האימונים שלנו באמצעות תמונות סיליה עצבית במבחנה, שיעור השגיאות היה 1.378% לעומת 3.36% עבור תמונות ב- vivo המוח סעיף (איור 1 משלים). לאחר השלמת האימון, Ai יכול לשמש כדי לפלח ריסים מתמונות ניסיוניות בתוך שניות ואת מסכות בינאריות התוצאה משמשים למדידת פרמטרים מבניים. פעולה זו מבטלת את הצורך לפלח אובייקטים באמצעות השיטה המסורתית של סף עוצמה אשר עשוי להיות קשה בתמונות עם רעש רקע גבוה או כאשר אובייקטים נמצאים בסמיכות זה לזה. Ai גם מפחית את הפוטנציאל לשגיאה ולהטיה על-ידי החלת אותו אלגוריתם על כל התמונות, ללא קשר למשתמש.
מדידת אורך ריסים באמצעות GA3
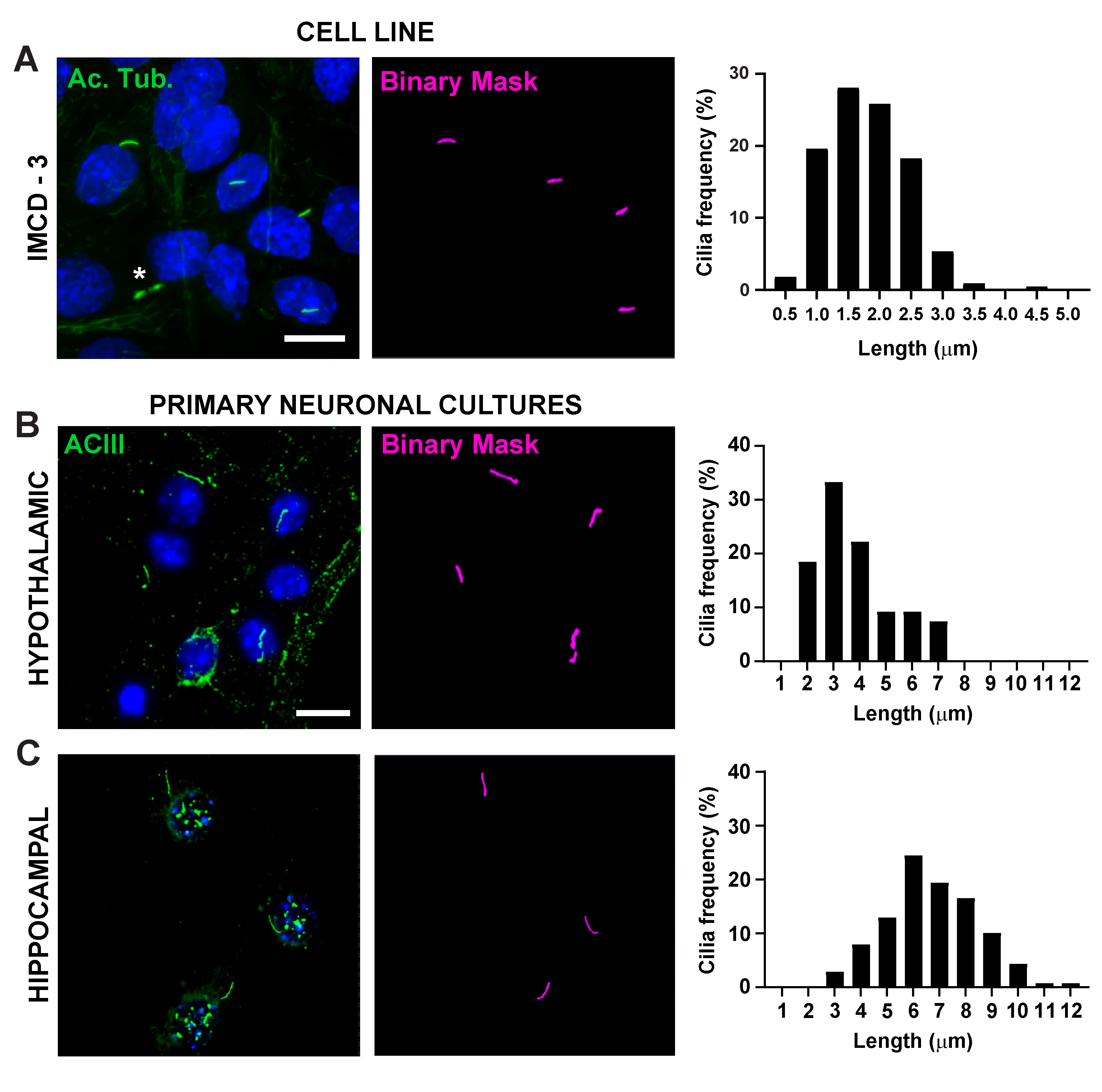
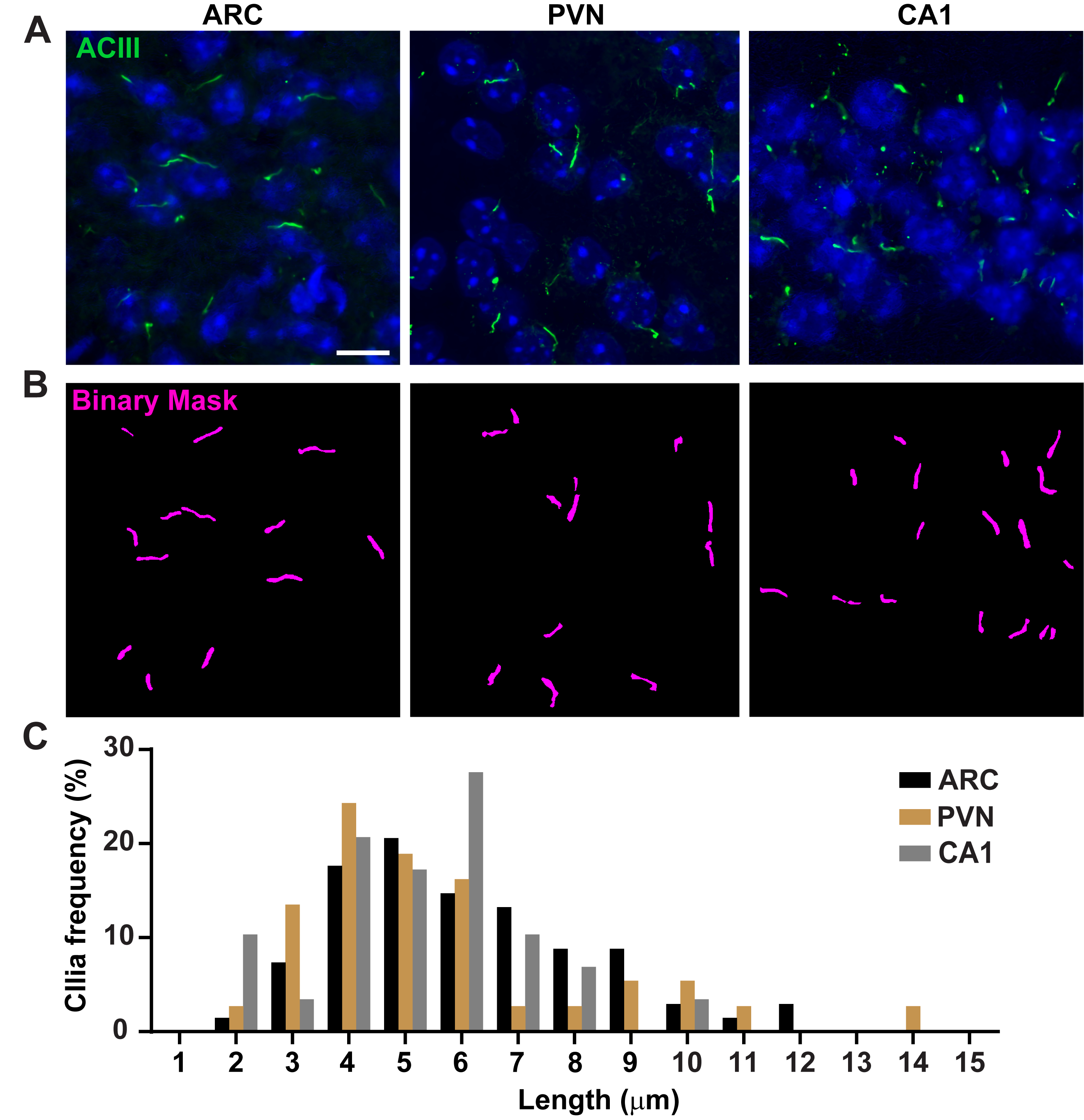
אורך Cilia מוסדר היטב והוא קשור עם השפעות פונקציונליות על איתות ciliary16,19. כאן מדדנו אורכי ריסים באמצעות צינור ניתוח בתוך תוכנת אלמנטים של NIS שנקראת ניתוח כללי 3 או GA3. GA3 מסייע בשילוב כלים מרובים בזרימת עבודה אחת כדי לבנות שגרות מותאמות אישית עבור כל ניסוי. התחלנו במדידת אורכי ריסים בקו תא. תאי צינור האיסוף הפנימיים של Cilia על העכבר (IMCD-3) היו מחוסנים עם טובולין אצטילציה וצוו באמצעות מיקרוסקופ קונפוקלי. מדדנו אורכי ריסים באמצעות GA3 לאחר פילוח עם segment.ai(איור משלים 3A). בעוד אצטילציה α-טובולין נמצא באופן מועדף בסיליום הראשי, הוא נמצא גם באזורים עשירים אחרים microtubule כגון ציטוסקלטון, כמו גם הגשר הציטוקינטי. Ai מאומן זיהה כראוי cilia בתמונה אבל לא מבנים אחרים שאינם ciliary, אצטילציה טובולין חיובי. Cilia על תאי IMCD נע בין 0.5 מיקרומטר ל 4.5 מיקרומטר עם אורך ממוצע של 1.8 ± 0.04 מיקרומטר(איור 2A). לאחר מכן בדקנו את היכולת של Ai למדוד אורכי ריסים בתרבויות עצביות ראשוניות. אנחנו תאינו נוירונים מההיפותלמוס וההיפוקמפוס של עכברי יילודים במשך 10 ימים ואימונולאבל אותם עם סמן הריסים אדנילט ציקלאז III (ACIII)21,41. בעת ניתוח תרביות עצביות, מצאנו את זה שימושי ליישם מסנן לפני ניתוח סטטיסטי של האורכים. בגלל יחס אות לרעש נמוך יותר, זוהו מספר עצמים הנמוכים מ- 1 מיקרומטר שלא היו ריסים. לכן, סיננו את הנתונים כדי לחסל את כל האובייקטים שהיו פחות מ 1 מיקרומטר אורך כדי להבטיח כי רק cilia נותחו. בתאי עצב היפותלמיים בתרבית, אורכי הריסים נעו בין 2 מיקרומטר ל-7 מיקרומטר עם אורך ממוצע של 3.8 ± 0.19 מיקרומטר(איור 2B). מעניין, ריסים עצביים בהיפוקמפוס בתרבית היו ארוכים יותר עם אורך ממוצע של 6.73 ±0.15μm(איור 2C). דווח כי גרעינים עצביים שונים בתוך ההיפותלמוס מראים אורכים ברורים של ריסים וכי ריסים אלה משנים את אורךם בתגובה לשינויים פיזיולוגיים באופן ספציפי לגרעין19,23. לכן, תייגנו גם מקטעי מוח היפותלמיים מעכברי C57BL/6J זכר בוגרים עם ACIII ודמיינו את גרעין הארקואט (ARC) ואת גרעין הפאבנטריקולרי (PVN). באמצעות GA3 כדי למדוד אורכי ריסים, ראינו כי הריסת ההיפותלמוס in vivo הופיעה זמן רב יותר מאשר במבחנה. באופן ספציפי, ריסים ההיפותלמיים ב-vivo נעים בין 1 מיקרומטר לכ-15 מיקרומטר(איור 3). לא היו הבדלים משמעותיים בין אורכי הריסים ב- PVN (5.54 ± 0.0.42 מיקרומטר) לבין אלה שב- ARC (6.16 ± 0.27 מיקרומטר) (איור 3C)23. באופן דומה, ריסים באזור קורנו אמוניס (CA1) בהיפוקמפוס מציגים טווח אורך צר יותר בין 1 מיקרומטר ל -10 מיקרומטר עם אורך ממוצע של 5.28 ± 0.33 מיקרומטר (איור 3). בהתאם למחקרים שפורסמו בעבר, הניתוח שלנו באמצעות כלי Ai ו- GA3 הראה כי cilia מאזורי מוח שונים מראים גיווןבאורך 19,23. יתר על כן, באמצעות גישה זו Ai אנו מסוגלים להעריך במהירות מספר רב של cilia.
מדידת הרכב ריסים באמצעות GA3
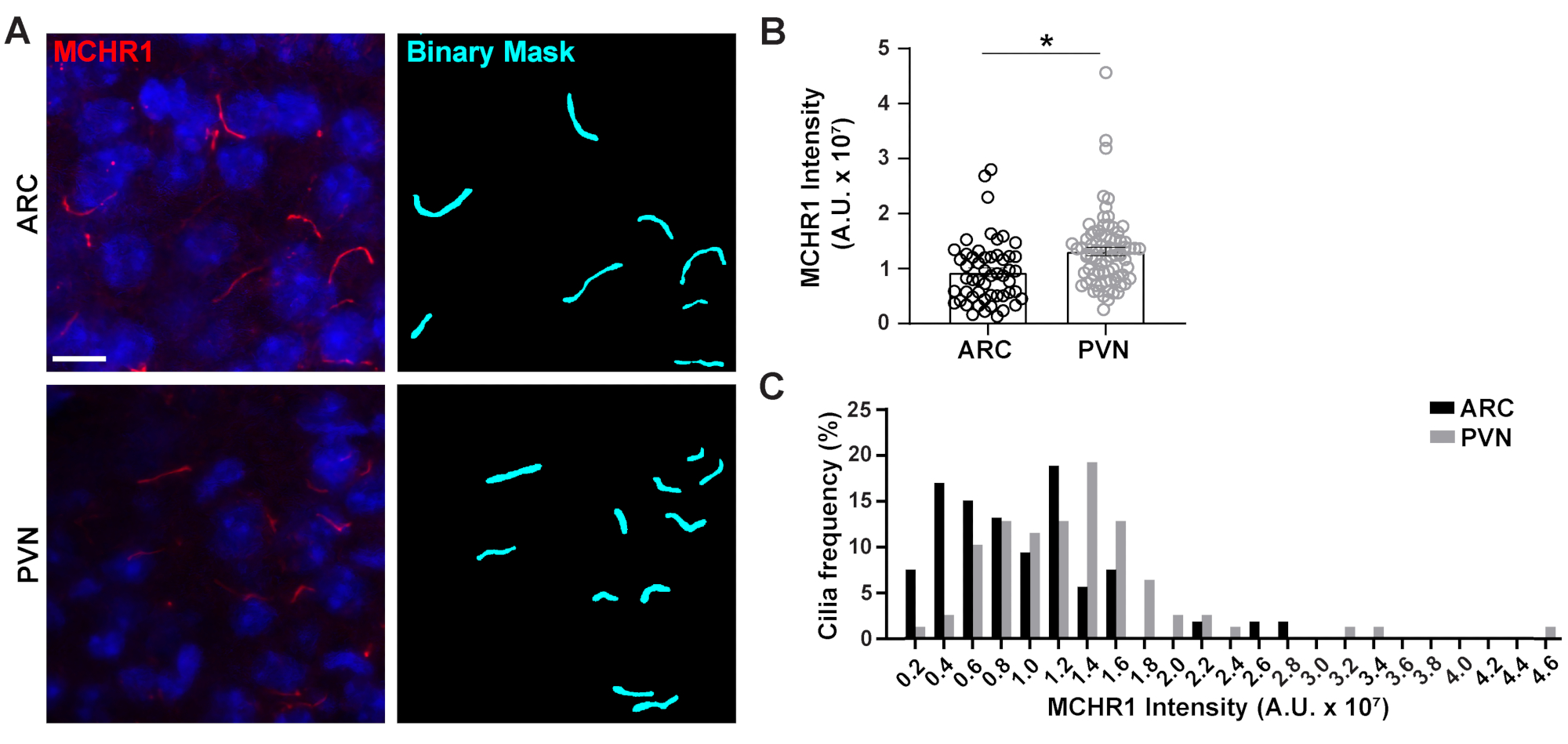
סיליום הראשי הוא רכזת איתות עבור מסלולים רבים המשתמשים בסוגים שונים של חלבונים כדי לבצע פונקציות ייחודיות כגון חלבונים מוטוריים, חלבוני הובלה תוך דליקתיים ו- GPCRs כדי להזכיר כמה3,24,42,43. שמירה על הרמות המתאימות של חלבונים אלה בתוךסיליום חשובה לתפקוד תקין ולעתים קרובות מופיעה תלויה בהקשר התא. תיוג פלואורסצנטי של חלבונים אלה לא רק אפשר לנו לדמיין אותם, אלא גם לכמת את עוצמותיהם כמדד לכמות החלבון המסומן בתוך התא הקטן יחסית20. לכן, ביקשנו לקבוע את העוצמות של GPCR ciliary, מלנין ריכוז קולטן הורמון 1 (MCHR1), ב vivo הן ARC והן PVN של ההיפותלמוס של עכברים זכרבוגרים 24,44. באמצעות Ai ו- GA3, מדדנו את האורכים של ריסים חיוביים MCHR1 יחד עם עוצמות כדי להבטיח את האובייקטים שנספרו היו cilia(איור משלים 3A). חיסלנו אובייקטים לאחר ניתוח שהיו באורך של פחות מ-2 מיקרומטר וניתחנו עוצמות של מסכות בינאריות שנותרו. מעניין, מצאנו כי עוצמת MCHR1 ciliary ב PVN הוא גבוה משמעותית מזה ב ARC המציין נוכחות חזקה יותר של MCHR1 ciliary ב PVN (איור 4). מחקרים נוספים נדרשים כדי לקבוע את המשמעות של MCHR1 ciliary במעגלים עצביים אלה. מדדנו גם את האינטנסיביות של MCHR1 ciliary בנוירונים בתרבית ראשונית של ההיפותלמוס וההיפוקמפוס. Cilia משתי התרבויות מציגה התפלגות רחבה של עוצמות MCHR1 המצביעות על נוכחות של אוכלוסיות עצביות הטרוגניות(איור 2 משלים). לכן, באמצעות כלים אנליטיים מתוחכמים כמו Ai ו- GA3 מאפשר הערכה של הטרוגניות ריסים בתוך אותה רקמה או בין רקמות מרובות. יהיה מעניין לראות אם GPCRs עצביים אחרים מראים הבדלים דומים לוקליזציה שלהם בתוך נוירונים של אותה רקמה ואם זה משתנה בתגובה לשינויים פיזיולוגיים.
Colocalization
בעוד שמדידת עוצמות פלואורסצנטיות בתוך שדה תמונה מלא יכולה ליצור רושם של חלבון, היא אינה מספקת מידע כגון הפצה מרחבית או קרבה לחלבונים ומבנים תאיים סמוכים אחרים. כאן מדדנו את החפיפה של MCHR1 עם ACIII כסמן ריסים על ידי התוויית האינטנסיביות של MCHR1 נגד זו של ACIII עבור כל מסכה בינארית(איור 5). הגרף מראה כי רוב הריסים חיוביים עבור שניהם, ACIII ו- MCHR1, אם כי כמה cilia להראות ביטוי חזק יותר של ערוץ אחד על פני השני. יתר על כן, ישנם כמה cilia המציגים את נוכחותם של ACIII או MCHR1 כפי שניתן לראות מן הנקודות הנמצאות ישירות על ציר x וציר y בהתאמה. כדי לכמת חפיפה זו, מדדנו את מקדם החפיפה של מנדר והשווינו את היקף ביטוי MCHR1 בריסים העצביים של ARC ו- PVN40. מעניין, הניתוח שלנו גילה כי חלה עלייה משמעותית מקדמי PVN (0.6382 ± 0.0151) מאשר אלה ב ARC (0.5430 ± 0.0181) (איור 5C). זה עולה בקנה אחד עם הנתונים הקודמים שלנו שבהם ראינו עוצמות MCHR1 גבוהות יותר ב- PVN בהשוואה ל- ARC (איור 4). נתונים אלה מצביעים על כך שכמו אורך הריסים, דפוס הביטוי של MCHR1 בתא הססני משתנה באזורים שונים של המוח. באמצעות אותו צינור ניתוח, ניתן יהיה לקבוע אם GPCRs ciliary אחרים כגון Neuropeptide Y קולטן סוג 2 (NPY2R) ו סומטוסטין קולטן סוג 3 (SSTR3) להראות כמויות דומות של גיוון.
פרופיל עוצמת מדידת לאורךסיליום
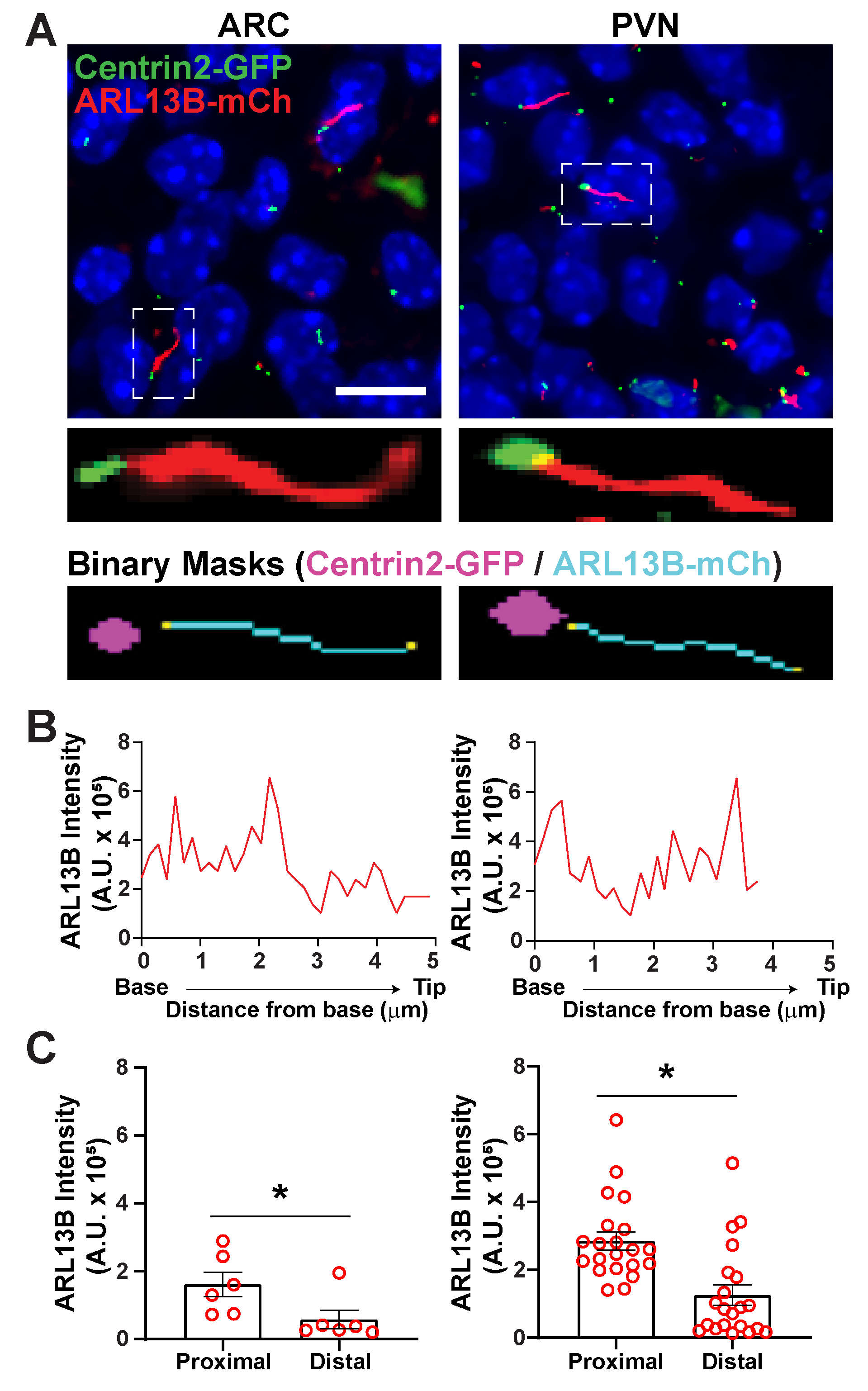
לאחר ריסים זוהו באמצעות segment.ai, המתכון GA3 ניתן לשנות לשלב ניתוח cilia עם זיהוי של מבנים אחרים של עניין בתמונה. לדוגמה, תיוג עם סמני גוף בזאליים שימושי לזיהוי קוטביות ריסים. כדי לבצע ניתוח זה, צילמנו מקטעי מוח היפותלמיים מעכברי P0 המבטאים ARL13B-mCherry ו- Centrin2-GFP ודמיינו את ARC ו- PVN34. כאן זוהו ריסים באמצעות Ai כמו קודם, אך כעת מתכון GA3 שונה כולל זיהוי של Centrin2-GFP, חלבון צנטריולר שנמצא בבסיס הריסים(איור 3B משלים). על ידי תיוג Centrin2-GFP, ניתן להבחין בין בסיס הריסים לבין קצות הריסים החיוביים ARL13B-mCherry(איור 6A). לאחר מכן, במקום למדוד את עוצמת הריסים כולה, אנו מסוגלים למדוד שינויים בעוצמת ARL13B לאורך הריסים (איור 6B). אנחנו יכולים גם להשוות הבדלים בעוצמת ARL13B בין הקצוות הפרוקסימליים לבין הקצוות הדיסטליים של הריסים. כדי לעשות זאת, חילקנו את אורך הסליום בפחים של מיקרון אחד החל מהבסיס וייעדנו את סל המיקרון הראשון כקצה הפרוקסימלי ואת סל המיקרון האחרון כקצה הדיסטלי. הניתוח שלנו גילה כי יש באופן משמעותי יותר ARL13B נוכח קרוב יותר לבסיס מאשר קצה של cilium הן ARC והן PVN, וזה עולה בקנה אחד עם מחקרים שפורסמו בעבר כונדרוציטים אנושיים45 (איור 6C). בסוג זה של ניתוח, במקום להחיל מסנן אורך כדי לא לכלול אובייקטים קטנים שאינם ciliary מניתוח, רק cilia המשויך Centrin2-GFP נותחים. זה יכול להיות יתרון במצבים שבהם מוטציות גנטיות להפוך ריסים קצרים מאוד, או אם שינויים subdomains cilia כמו אזור המעבר או טיפ כבר מעורב. זיהוי ריסים באמצעות ניתוח Ai ו- GA3 הוא מותאם מאוד וניתן להתאים אותו כך שיתאים למגוון שאלות מחקר מורכבות.

איור 1. זרימת עבודה למדידת אורך ועוצמה של ריסים באמצעות Ai. (A)כדי לאמן את הבינה האווירית, קבצים הבינאריים נמשכים סביב אובייקטי העניין (cilia) בתמונות האימון הגולמיות. באמצעות הקבצים הבינאריים המצוירים, Segment Ai מאומן לזהות את הצורה ואת עוצמות הפיקסלים של הריסים. (B)לאחר מכן, מקטע Ai מאומן מוחל על תמונות ניסיוניות גולמיות. הוא מצייר binaries על אובייקטים שהוא מזהה כמו cilia. ניתן לחדד הקבצים הבינאריים האלה כדי לוודא שכל הריסים רק מנותחים. (C)תוכנית GA3 בנויה כדי לנתח את העוצמה והאורך של אובייקטים המזוהים על ידי Ai. (D)הרשומות מיובאות לטבלה בתוכנה. לאחר מכן ניתן לייצא טבלה זו לניתוח נוסף. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 2. מדידות אורך של סיירת אין ויטרו. תמונות מייצגות של תאי IMCD (ירוק, אצטילאטאט טובולין) (B) תרביות היפותלמוס עיקריות (ירוק, ACIII) ו - (C) תרביות ההיפוקמפוס (ירוק, ACIII). Ai מאומן שימש לזיהוי ריסים כפי שמוצג במסכה הבינארית (מגנטה) ולאחר מכן GA3 שימש למדידת אורך ריסים. התפלגות אורך הריסים גרף כאחוז של cilia בפחים של 0.5 או 1.0 מיקרון. * מציין גשר ציטוקינטי כראוי לא מוכר על ידי Ai. n = 225 cilia בתאי IMCD מ 3 שכפולים, 54 ריסים בהיפותלמוס ו 139 ריסים בתרביות היפוקמפוס מ -3 בעלי חיים. סרגלי קנה מידה 10 מיקרומטר. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 3. במדידות אורך ריסים של ויוו. (A)תמונות מייצגות של ריסים (ירוק, ACIII) ב- ARC, PVN ו- CA1 של מקטעי מוח עכבר בוגרים. (ב)Ai מאומן באלמנטים שקלים שימש לזיהוי ריסים כפי שמוצג במסכה הבינארית (מגנטה) ולאחר מכן GA3 שימש למדידת אורך ריס. (C)התפלגות אורך הריסים גרף כאחוז של ריסים בפחים של מיקרון אחד. n = 68 ריסונים ב- ARC, 36 ב- PVN ו - 29 ב- CA1 מ -3 בעלי חיים. סרגלי קנה מידה 10 מיקרומטר. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 4. Ai סייעה לריסים מכתים מדידות עוצמת של ריסים עצביים היפותלמיים. (A)תמונות מייצגות של ריסים (MCHR1, אדום) ב- ARC ו- PVN של מקטעי מוח עכבר בוגר. Ai מאומן באלמנטים שקל שימש לזיהוי ריסים כפי שמוצג במסכה הבינארית (ציאן) ולאחר מכן GA3 שימש למדידת עוצמת מכתם MCHR1 בריסים. (B)עוצמות MCHR1 גרף כממוצע ± S.E.E.M. כל נקודה מייצגת סיליום. * p < 0.05, מבחן T של סטודנט. (C)התפלגות עוצמת MCHR1 גרף כאחוז cilia בפחים של 0.2 x 107 יחידות שרירותיות (A. U. ). n = 53 ריסים ב ARC, 78 ב PVN מ 3 בעלי חיים. סרגלי קנה מידה 10 מיקרומטר. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 5. Ai סייע ניתוח colocalization cilia. (א, ב) תמונות מייצגות של ריסים ב- ARC וב- PVN בהתאמה. Cilia מסומנים עם ACIII (ירוק) ו- MCHR1 (אדום). Ai מאומן באלמנטים של שקלים שימש לזיהוי ריסים כפי שמוצג במסכה הבינארית (מגנטה ל-ACIII המסומנת כריסים, ציאן ל-MCHR1 עם תווית ריסים). GA3 שימש לזיהוי ריסים שהכילו הן ACIII והן MCHR1. (C)ערכי מקדם החפיפה (MOC) של מנדרים גרף כממוצע ± S.E.E.M. כל נקודה מייצגת סיליום. * p < 0.05, מבחן T של סטודנט. (D)חלקת פיזור של עוצמת MCHR1 לעומת עוצמת ACIII ב- ARC ו- PVN. כל נקודה מייצגת סיליום. n = 72 ריסים ב ARC, 47 ב PVN מ 3 בעלי חיים. סרגלי קנה מידה 10 מיקרומטר. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 6. ניתוח גוף סיליה ובזל. (A)תמונות מייצגות של ריסים (אדום, ARL13B-mCherry) וסמן גוף בזאלי (ירוק, Centrin2-GFP) ב ARC ו PVN של עכברי P0. Ai מאומן שימש לזיהוי ריסים כפי שמוצג במסכה הבינארית (ציאן). המסכה הבינארית לגוף הבזל (מגנטה) צוירה על ידי סף במתכון GA3. (B)עוצמת סריקת קו מייצג שלסיליום. (C)עוצמות ARL13B בקצוות הפרוקסימליים והדיסטליים של Ai זיהו ריסים בגרף כממוצע ± S.E.E.M. הקצוות הפרוקסימליים והדיסטליים מוגדרים כאזור באורך 1 מיקרומטר הראשון ואורך 1 מיקרומטר האחרון בהתאמה מבסיססיליום. כל נקודה מייצגת סיליום. * p < 0.05. n = 6 ריסים ב- ARC מ-2 בעלי חיים ו-21 ריסים ב-PVN מ-3 בעלי חיים. סרגלי קנה מידה 10 מיקרומטר. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}
איור משלים 1. גרפי אובדן אימונים של באני. (A, B)גרפים המראים אובדן אימון של segment.ai על סיליה עצבית במבחנה ו in vivo, בהתאמה. נא לחץ כאן כדי להוריד קובץ זה.
איור משלים 2. Ai בסיוע סיליה מכתים מדידות עוצמת של vitro אורירוני . (א, ב) תמונות מייצגות של ריסים (MCHR1, אדום) בתרבויות ההיפותלאמיות וההיפוקמפוס העיקריות, בהתאמה. Ai מאומן באלמנטים שקל שימש לזיהוי ריסים כפי שמוצג במסכה הבינארית (ציאן) ולאחר מכן GA3 שימש למדידת עוצמת מכתם MCHR1 בריסים. התפלגות עוצמת MCHR1 גרף כאחוז של cilia ב 1000 A.U. פחים עבור ההיפותלמוס ו 2000 A. פחים U. עבור תרביות היפוקמפוס. n = 30 cilia בהיפותלמוס ו -106 ריסים בתרבויות ההיפוקמפוס מ -3 בעלי חיים. סרגלי קנה מידה 10 מיקרומטר. אנא לחץ כאן כדי להוריד קובץ זה.
איור משלים 3. ניתוח כללי 3 מתכונים לניתוח ריסים. (א)מתכון ניתוח כללי פשוט (GA3) למדידה של אורך ריסים, עוצמה ומקדם מנדר. (B)מתכון GA3 מורכב למדידת עוצמה לאורךסיליום באמצעות סמן לגוף הבזל. נא לחץ כאן כדי להוריד קובץ זה.
Discussion
מדידות אורך ועוצמה הן דרכים נפוצות שבהן מנתחים ריסים ראשוניים, עם זאת, אין שיטה קונבנציונלית סטנדרטית המשמשת בתחום. זיהוי וכימות של ריסים ראשיים באמצעות תוכנה כגון ImageJ הוא זמן רב נוטה הטיית משתמש וטעייה. הדבר מקשה על ניתוח מדויק של ערכות נתונים גדולות. כאן אנו מראים כי באמצעות תוכנית Ai יכול להתגבר על רבים מהאתגרים האלה מה שהופך ניתוח תפוקה גבוהה של cilia ראשוני בר השגה. כאן אנו מתארים את ההליך להכשרת יישום מבוסס Ai לזיהוי ריסים ראשוניים ומתארים את השלבים הנדרשים לניתוח אורך ועוצמת.
בעוד ההכשרה הראשונית של Ai לזהות cilia דורש זמן משמעותי מהמשתמש, לאחר השלמתו ניתן להשתמש בו על כל ערכת נתונים שנרכשה עם אותם פרמטרים. המסיכה הבינארית שנוצרת על-ידי ה- Ai ניתנת לשינוי כך שניתן יהיה לתקן שגיאות כלשהן. עם זאת, שגיאות בזיהוי cilia צריך לאותת למשתמש כי Ai צריך להיות מאומן עוד יותר עם תמונות נוספות. אחד היתרונות העיקריים של שיטה זו הוא כי Ai ניתן לאמן לזהות cilia בסוגי מדגם שונים הן 2D ו 3D. שיטות ניתוח קודמות שנוצרו בתוך מעבדות יש מגבלות שונות כולל דרישת סף ידני לזיהוי ובעיות בזיהוי cilia בתמונה מקטעי רקמות שבו צפיפות התא גבוהה36,46,47. שיטות אלה מתמחות גם בניתוח ריסים ואילו ניתוח באמצעות תוכנת אלמנטים של NIS יכול להעריך מספר היבטים של התמונות בו זמנית. מכיוון שה-Ai המתואר כאן הוא חלק מחבילת התוכנה של NIS Elements, ניתן להמשיך בקלות בתמונות שנרכשו באמצעות מיקרוסקופ ניקון עד לניתוח. עם זאת, הדמיה עם ניקון אינה נדרשת לשימוש בשיטה זו. ללא קשר לתבנית קובץ הנתונים הגולמיים שנתפסו, קבצי ".tif" יכולים להיפתח על ידי רכיבי NIS לשימוש ב- Ai.
יישום AI זה בתוך אלמנטים NIS זמין באופן נרחב ואולי כבר חלק מתוכנת ניתוח תמונה בשימוש על ידי מעבדות לומדות ריסים ראשוניים. עם ההתרחבות של טכנולוגיית הבינה מלאכותית, תוכנות הדמיה אחרות עשויות להרחיב את אפשרויות הניתוח שלהן כך שיכללו מודול AI דומה. החלת ניתוח Ai לזיהוי cilia יכול לשמש עבור מספר היבטים שונים של ניתוח cilia. בעוד שרטטנו שיטות למספר ניתוחים פשוטים כגוןאורך ( איור 2 ו- 3), עוצמה (איור 4) ואנלקציה (איור 5) ניתן להוסיף ניתוח מתוחכם יותר לזרימת העבודה של ניתוח GA3 כמו באיור 6. לדוגמה, במקום למדוד את עוצמתו שלסיליום שלם, הבדלים בעוצמה בתוך תת-אזור שלסיליום עשויים לעניין להעריך לוקליזציה תת-סיליארית. הבדלים בעוצמה בתוך תת-תחזית שלסיליום יכולים להצביע על כך שהחלבון מצטבר בקצה או בבסיס של הסליום, כגון האופן שבו חלבוני Gli מועשרים בקצה ריס48. בנוסף, יישום Ai זה יכול לשמש כדי לזהות בקלות הבדלים בין גנוטיפים או קבוצות טיפול. בעוד המעבדה שלנו משתמשת בעיקר בשיטה זו כדי לנתח ריסים שמצויים מחלקי מוח או מתרביות עצביות, ניתן להחיל אותה על תמונות שנרכשו מקווי תאים שונים או מסוגי רקמות אחרים. הגמישות של סוג המדגם כי יישום זה יכול לשמש על עושה שיטה זו של ניתוח בעל ערך עבור קבוצות רבות ושונות לומדות ריסים ראשוניים או כל אברון נפרד כי הוא מוערך כגון מיטוכונדריה, גרעין, או ER.
Disclosures
מחבר שותף ווסלי לואיס הוא עובד של ניקון. אין גילויים פיננסיים.
Acknowledgements
עבודה זו מומנה על ידי המכון הלאומי לסוכרת ומחלות עיכול וכליות R01 DK114008 ל- NFB ומענק מלגת איגוד הלב האמריקאי #18PRE34020122 ל- RB. אנו מודים לריץ' גרוסקין, המנהל הכללי של ניקון תוכנה, מליסה בנטלי, קורטני הייקראפט ותרזה מסלאצ'י על הערות תובנות על כתב היד.
Materials
| Name | Company | Catalog Number | Comments |
| Intel Xeon, 3.6 GHz, 32GB RAM | Intel Corporation | W-2123 | Processor used for running NIS Elements. |
| Nikon Elements Software | Nikon Instruments Inc. | - | Ai and GA3 software |
| Quadro RTX 4000 Graphics card | NVIDIA Corporation | Quadro RTX 4000 | |
| Windows 10 Professional 64-bit | Microsoft Inc. | - | Operating system used for running NIS Elements |
| Workstation | HP Development Company, L.P. | HP Z4G4 | Workstation used for running NIS Elements |
References
- Wang, L., et al. Ciliary gene RPGRIP1L is required for hypothalamic arcuate neuron development. JCI Insight. 4 (3), (2019).
- Siljee, J. E., et al. Subcellular localization of MC4R with ADCY3 at neuronal primary cilia underlies a common pathway for genetic predisposition to obesity. Nature Genetics. 50 (2), 180-185 (2018).
- Davenport, J. R., et al. Disruption of intraflagellar transport in adult mice leads to obesity and slow-onset cystic kidney disease. Current Biology. 17 (18), 1586-1594 (2007).
- Berbari, N. F., O'Connor, A. K., Haycraft, C. J., Yoder, B. K. The primary cilium as a complex signaling center. Current Biology. 19 (13), 526-535 (2009).
- Walz, G. Role of primary cilia in non-dividing and post-mitotic cells. Cell Tissue Research. 369 (1), 11-25 (2017).
- Nachury, M. V., Mick, D. U. Establishing and regulating the composition of cilia for signal transduction. Nature Reviews. Molecular Cell Biology. 20 (7), 389-405 (2019).
- Goetz, S. C., Anderson, K. V. The primary cilium: a signalling centre during vertebrate development. Nature Reviews Genetics. 11 (5), 331-344 (2010).
- Engle, S. E., Bansal, R., Antonellis, P. J., Berbari, N. F. Cilia signaling and obesity. Seminars in Cell and Developmental Biology. , (2020).
- Reiter, J. F., Leroux, M. R. Genes and molecular pathways underpinning ciliopathies. Nature reviews. Molecular Cell Biology. 18 (9), 533-547 (2017).
- Waters, A. M., Beales, P. L. Ciliopathies: an expanding disease spectrum. Pediatric Nephrology. 26 (7), 1039-1056 (2011).
- Hildebrandt, F., Benzing, T., Katsanis, N. Ciliopathies. New England Journal of Medicine. 364 (16), 1533-1543 (2011).
- Vaisse, C., Reiter, J. F., Berbari, N. F. Cilia and Obesity. Cold Spring Harbor Perspectives in Biology. 9 (7), (2017).
- Berbari, N. F., et al. Leptin resistance is a secondary consequence of the obesity in ciliopathy mutant mice. Proceedings of the National Academy of Sciences of the United States of America. 110 (19), 7796-7801 (2013).
- Jacobs, D. T., et al. Dysfunction of intraflagellar transport-A causes hyperphagia-induced obesity and metabolic syndrome. Disease Models & Mechanisms. 9 (7), 789-798 (2016).
- Arsov, T., et al. Fat aussie--a new Alström syndrome mouse showing a critical role for ALMS1 in obesity, diabetes, and spermatogenesis. Molecular Endocrinology. 20 (7), 1610-1622 (2006).
- Tam, L. W., Ranum, P. T., Lefebvre, P. A. CDKL5 regulates flagellar length and localizes to the base of the flagella in Chlamydomonas. Molecular Biology of the Cell. 24 (5), 588-600 (2013).
- Rajagopalan, V., Subramanian, A., Wilkes, D. E., Pennock, D. G., Asai, D. J. Dynein-2 affects the regulation of ciliary length but is not required for ciliogenesis in Tetrahymena thermophila. Molecular Biology of the Cell. 20 (2), 708-720 (2009).
- Bengs, F., Scholz, A., Kuhn, D., Wiese, M. LmxMPK9, a mitogen-activated protein kinase homologue affects flagellar length in Leishmania mexicana. Molecular Microbiology. 55 (5), 1606-1615 (2005).
- Han, Y. M., et al. Leptin-promoted cilia assembly is critical for normal energy balance. Journal of Clinical Investigation. 124 (5), 2193-2197 (2014).
- Caspary, T., Marazziti, D., Berbari, N. F., Satir, P., Tvorup Christensen, S. . Cilia: Methods and Protocols. , 203-214 (2016).
- Bishop, G. A., Berbari, N. F., Lewis, J., Mykytyn, K. Type III adenylyl cyclase localizes to primary cilia throughout the adult mouse brain. Journal of Comparative Neurology. 505 (5), 562-571 (2007).
- Domire, J. S., Mykytyn, K. Markers for neuronal cilia. Methods in Cell Biology. 91, 111-121 (2009).
- Sun, J. S., et al. Ventromedial hypothalamic primary cilia control energy and skeletal homeostasis. Journal of Clinical Investigation. 131 (1), (2021).
- Berbari, N. F., Johnson, A. D., Lewis, J. S., Askwith, C. C., Mykytyn, K. Identification of ciliary localization sequences within the third intracellular loop of G protein-coupled receptors. Molecular Biology of the Cell. 19 (4), 1540-1547 (2008).
- Brailov, I., et al. Localization of 5-HT(6) receptors at the plasma membrane of neuronal cilia in the rat brain. Brain Research. 872 (1-2), 271-275 (2000).
- Domire, J. S., et al. Dopamine receptor 1 localizes to neuronal cilia in a dynamic process that requires the Bardet-Biedl syndrome proteins. Cellular and Molecular Life Sciences. 68 (17), 2951-2960 (2011).
- Handel, M., et al. Selective targeting of somatostatin receptor 3 to neuronal cilia. Neuroscience. 89 (3), 909-926 (1999).
- Koemeter-Cox, A. I., et al. Primary cilia enhance kisspeptin receptor signaling on gonadotropin-releasing hormone neurons. Proceedings of the National Academy of Sciences of the United States of America. 111 (28), 10335-10340 (2014).
- Mukhopadhyay, S., et al. The ciliary G-protein-coupled receptor Gpr161 negatively regulates the Sonic hedgehog pathway via cAMP signaling. Cell. 152 (1-2), 210-223 (2013).
- Berman, S. A., Wilson, N. F., Haas, N. A., Lefebvre, P. A. A novel MAP kinase regulates flagellar length in Chlamydomonas. Current Biology. 13 (13), 1145-1149 (2003).
- Nguyen, R. L., Tam, L. W., Lefebvre, P. A. The LF1 gene of Chlamydomonas reinhardtii encodes a novel protein required for flagellar length control. Genetics. 169 (3), 1415-1424 (2005).
- Tam, L. W., Wilson, N. F., Lefebvre, P. A. A CDK-related kinase regulates the length and assembly of flagella in Chlamydomonas. Journal of Cell Biology. 176 (6), 819-829 (2007).
- O'Connor, A. K., et al. An inducible CiliaGFP mouse model for in vivo visualization and analysis of cilia in live tissue. Cilia. 2 (1), 8 (2013).
- Bangs, F. K., Schrode, N., Hadjantonakis, A. K., Anderson, K. V. Lineage specificity of primary cilia in the mouse embryo. Nature Cell Biology. 17 (2), 113-122 (2015).
- Delling, M., et al. Primary cilia are not calcium-responsive mechanosensors. Nature. 531 (7596), 656-660 (2016).
- Saggese, T., Young, A. A., Huang, C., Braeckmans, K., McGlashan, S. R. Development of a method for the measurement of primary cilia length in 3D. Cilia. 1 (1), 11 (2012).
- Kobayashi, Y., Hamamoto, A., Saito, Y. Analysis of ciliary status via G-protein-coupled receptors localized on primary cilia. Microscopy. 69 (5), 277-285 (2020).
- Zhou, L. Q., et al. Artificial intelligence in medical imaging of the liver. World Journal of Gastroenterology. 25 (6), 672-682 (2019).
- Naugler, C., Church, D. L. Automation and artificial intelligence in the clinical laboratory. Critical Reviews in Clinical Laboratory Sciences. 56 (2), 98-110 (2019).
- Dunn, K. W., Kamocka, M. M., McDonald, J. H. A practical guide to evaluating colocalization in biological microscopy. American Journal of Physiology-Cell Physiology. 300 (4), 723-742 (2011).
- Bansal, R., et al. Hedgehog Pathway Activation Alters Ciliary Signaling in Primary Hypothalamic Cultures. Frontiers in Cellular Neuroscience. 13, 266 (2019).
- Jin, H., et al. The conserved Bardet-Biedl syndrome proteins assemble a coat that traffics membrane proteins to cilia. Cell. 141 (7), 1208-1219 (2010).
- Liew, G. M., et al. The intraflagellar transport protein IFT27 promotes BBSome exit from cilia through the GTPase ARL6/BBS3. Developmental Cell. 31 (3), 265-278 (2014).
- Engle, S. E., et al. A CreER Mouse to Study Melanin Concentrating Hormone Signaling in the Developing Brain. Genesis. , (2018).
- Thorpe, S. D., et al. Reduced primary cilia length and altered Arl13b expression are associated with deregulated chondrocyte Hedgehog signaling in alkaptonuria. Journal of Cellular Physiology. 232 (9), 2407-2417 (2017).
- Lauring, M. C., et al. New software for automated cilia detection in cells (ACDC). Cilia. 8, 1 (2019).
- Dummer, A., Poelma, C., DeRuiter, M. C., Goumans, M. J., Hierck, B. P. Measuring the primary cilium length: improved method for unbiased high-throughput analysis. Cilia. 5, 7 (2016).
- Haycraft, C. J., et al. Gli2 and Gli3 localize to cilia and require the intraflagellar transport protein polaris for processing and function. PLoS Genetics. 1 (4), 53 (2005).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved