
Method Article
Подходы искусственного интеллекта к оценке первичных ресничек
В этой статье
Резюме
Использование искусственного интеллекта (ИИ) для анализа изображений становится мощным, менее предвзятым и быстрым подходом по сравнению с широко используемыми методами. Здесь мы обучили Ии распознавать клеточную органеллу, первичные реснички и анализировать такие свойства, как длина и интенсивность окрашивания, строгим и воспроизводимым образом.
Аннотация
Реснички представляют собой клеточные придатки на основе микротрубочек, которые функционируют как сигнальные центры для разнообразия сигнальных путей во многих типах клеток млекопитающих. Длина ресничек сильно сохраняется, жестко регулируется и варьируется между различными типами клеток и тканей и была вовлечена в непосредственное воздействие на их сигнальную способность. Например, было показано, что реснички изменяют свою длину в ответ на активацию цилиарных рецепторов, связанных с G-белком. Однако точное и воспроизводимое измерение длины многочисленных ресничек является трудоемкой и трудоемкой процедурой. Современные подходы также подвержены ошибкам и предвзятости. Программы искусственного интеллекта (Ai) могут быть использованы для преодоления многих из этих проблем из-за возможностей, которые позволяют ассимилировать, манипулировать и оптимизировать обширные наборы данных. Здесь мы демонстрируем, что модуль Ai может быть обучен распознавать реснички на изображениях как из образцов in vivo, так и in vitro. После использования обученного Ai для идентификации ресничек мы можем проектировать и быстро использовать приложения, которые анализируют сотни ресничек в одном образце на длину, интенсивность флуоресценции и колокализацию. Этот непредвзятый подход повысил нашу уверенность и строгость при сравнении образцов из разных первичных нейрональных препов in vitro, а также в разных областях мозга внутри животного и между животными. Кроме того, этот метод может быть использован для надежного анализа динамики ресничек из любого типа клеток и тканей с высокой пропускной способностью в нескольких образцах и группах лечения. В конечном счете, подходы на основе ИИ, вероятно, станут стандартными, поскольку большинство областей движутся к менее предвзятым и более воспроизводимым подходам к получению и анализу изображений.
Введение
Первичные реснички представляют собой сенсорные органеллы, выступающие из большинства клеток млекопитающих типов1,2,3,4. Они, как правило, являются одиночными придатками, критически важными для координации различных клеточных сигнальных путей путем интеграции внеклеточных сигналов5,6,7. Первичные реснички играют важную роль во время эмбрионального развития и гомеостаза взрослой ткани, а нарушение их функции или морфологии связано с несколькими генетическими нарушениями, которые в совокупности называются цилиопатиями. Из-за почти повсеместного характера ресничек цилиопатии связаны с широким спектром клинических признаков, которые могут влиять на все системы органов8,9,10,11,12. На животных моделях цилиопатий потеря цилиарной структуры или сигнальной способности проявляется в нескольких клинически значимых фенотипах, включая гиперфагию-ассоциированное ожирение3,13,14,15. Во многих модельных системах было показано, что изменения длины ресничек влияют на их сигнальную способность и функции16,17,18,19. Тем не менее, существует несколько трудоемких и технических проблем, связанных с точной и воспроизводимой оценкой длины и состава ресничек.
Центральная нервная система взрослых млекопитающих (ЦНС) является одним из биологических контекстов, который создает проблему для понимания морфологии и функции ресничек. Хотя кажется, что нейроны и клетки по всей ЦНС обладают ресничками, из-за ограниченных инструментов и способностей наблюдать и анализировать эти реснички понимание их функций остается неуловимым20. Например, прототипный маркер ресничек, ацетилированный α-тубулин, не маркирует нейронные реснички20. Трудность изучения этих ресничек была частично решена с открытием нескольких рецепторов, связанных с G-белком (GPCR), сигнальных механизмов и мембранно-ассоциированных белков, которые обогащаются на мембране нейрональных ресничек21,22. Все эти простые основные наблюдения намекают на важность и разнообразие ресничек ЦНС, которые до сих пор не имеют аналогов среди других тканей. Например, вариации длины ресничек и локализации GPCR могут наблюдаться по всему мозгу, причем длины в определенных нейронных ядрах отличаются по сравнению с другими ядрами19,23. Аналогичным образом, их содержание GPCR и комплимент сигнального механизма показывают разнообразие, основанное на нейроанатомическом расположении и нейронах типа2,24,25,26,27,28,29. Эти простые наблюдения показывают, что длина и состав ресничек ЦНС млекопитающих жестко регулируются, как и в модельных организмах, таких как Chlamydomonas reinhardtii,но влияние этих различий длины на функцию ресничек, передачу сигналов и, в конечном счете, поведение остается неясным16,30,31,32.
Точное измерение длины и состава ресничек оказывается технической проблемой, подверженной ошибкам пользователя и невоспроизводимости. В настоящее время реснички in vivo и in vitro чаще всего идентифицируются с использованием иммунофлуоресцентных подходов, которые маркируют цилиарные белки или обогащенные ресничками флуоресцентные репортерные аллели33,34,35. Длина этих флуоресцентно помеченных ресничек затем измеряется из 2-мерного (2D) изображения с использованием инструментов линейного измерения в программах анализа изображений, таких как ImageJ36. Этот процесс не только утомителен и трудоемкий, но и подвержен предвзятости и ошибкам. Эти же препятствия возникают при измерении интенсивности ресничек, которые помогают указать на изменения в структуре ресничек37. Чтобы свести к минимуму несоответствия в этих типах анализа изображений, программы искусственного интеллекта (Ai) становятся все более распространенными и доступными вариантами38.
ИИ - это развитие компьютерных систем, которые используют преимущества компьютерных алгоритмов и программирования для выполнения задач, которые обычно требуют человеческого интеллекта39. Устройства ИИ учат воспринимать повторяющиеся паттерны, параметры и характеристики и предпринимать действия, чтобы максимизировать шансы на создание успешных результатов. ИИ универсален и может быть обучен распознавать конкретные объекты или структуры, представляющие интерес, такие как реснички, а затем быть запрограммированным на выполнение различных анализов идентифицированных объектов. Таким образом, сложные данные изображения могут быть быстро и воспроизводимо сгенерированы Ai38. Автоматизация и ai-анализ захваченных изображений повысят эффективность и результативность, ограничивая при этом любую потенциальную человеческую ошибку и предвзятость39. Создание методологии идентификации ресничек на основе ИИ создает последовательный способ для всех исследовательских групп анализировать и интерпретировать данные ресничек.
Здесь мы используем модуль Ai для идентификации ресничек как in vivo, так и in vitro на 2D-изображениях. Используя набор образцов изображений, ИИ обучается идентифицировать реснички. После завершения обучения назначенный Ai используется для нанесения бинарной маски на идентифицированные Ai реснички на изображении. Двоичные файлы, применяемые Ai, при необходимости модифицируются, чтобы обеспечить правильную идентификацию всех ресничек на изображениях и устранение неспецифической идентификации. После использования Ai для идентификации ресничек используются специально разработанные программы общего анализа (GA) для выполнения различных анализов, таких как измерение длины ресничек и интенсивности флуоресценции. Собранные данные экспортируются в таблицу, которая может быть легко прочитана, интерпретирована и использована для статистического анализа(рисунок 1). Использование автоматизированных технологий и искусственного интеллекта для идентификации ресничек и получения конкретных измерений между экспериментальными группами поможет в будущих исследованиях, направленных на понимание влияния функции и морфологии ресничек ЦНС на связь и поведение клеток и клеток.
протокол
1. Получение необработанных изображений
- Фиксировать и иммуномаркировать образцы по мере необходимости20.
- Изображение ресничек с помощью конфокального микроскопа на максимальной битовой глубине с использованием того же размера пикселя с разрешением Найквиста.
- Экспортируйте изображения в виде монохромных файлов формата изображений с тегами (.tif).
ПРИМЕЧАНИЕ: Этот протокол описывает, как использовать модуль Ai конкретно в программном обеспечении NIS Elements. Если изображения были получены в виде файлов .nd2, экспорт изображений в виде .tif файлов не требуется, и пользователь может перейти непосредственно к шагу 2.3. Если изображения были получены в другой системе, лицензию NIS Elements можно приобрести отдельно, а файлы .tif можно преобразовать, как описано в следующих шагах.
2. Обучите Ai идентифицировать реснички
- Откройте набор обучающих данных.
- Выберите приблизительно 50 образцов изображений с по крайней мере одной ресничкой на кадр, чтобы обучить программное обеспечение и скопировать их в одну папку. Эта папка используется для направления программного обеспечения при открытии изображений. Откройте эти 50 кадров в одном документе ND2 с по крайней мере одной ресничкой на кадр. Выберите Файл > Импорт/Экспорт > Создать ND Файл из Последовательности Файлов.
- Выберите папку, содержащую набор обучающих данных. Откроется список файлов в центре диалогового окна. Определите организацию файлов вручную, используя хотя бы один параметр в раскрывающемся меню выше. Возможны следующие параметры: многоточечные (для нескольких максимальных файлов проекции), серия Z (для изображения стека z), время (для покадрового изображения) и длина волны (для файлов из нескольких каналов).
- Введите соответствующие числовые значения под каждым выбранным параметром. Выберите Нет там, где параметры не выбраны. Нажмите кнопку Преобразовать, чтобы открыть документ ND.
- Калибровка изображений.
- Введите размер пикселя в левом нижнем углу изображения: щелкните правой кнопкой мыши Некалиброванный > Калибровка документа > Размер пикселя.
- Определите реснички.
- Ручная идентификация ресничек путем точной трассировки отдельных цилиарных структур на всех открытых кадрах с помощью функции автоопределения или объекта рисования на двоичной панели инструментов. Это позволит нарисовать двоичные маски на интересующих объектах. Эти двоичные файлы будут служить образцами объектов для обучения программного обеспечения идентификации ресничек по пиксельным характеристикам в будущем экспериментальном анализе изображений. Выберите Просмотр элементов управления анализом > > двоичной панели инструментов > Рисование объекта.
ПРИМЕЧАНИЕ: Удалите любой кадр, который не имеет двоичных файлов, так как программное обеспечение не начнет обучение, если оно не сможет обнаружить двоичные файлы на всех открытых кадрах.
- Ручная идентификация ресничек путем точной трассировки отдельных цилиарных структур на всех открытых кадрах с помощью функции автоопределения или объекта рисования на двоичной панели инструментов. Это позволит нарисовать двоичные маски на интересующих объектах. Эти двоичные файлы будут служить образцами объектов для обучения программного обеспечения идентификации ресничек по пиксельным характеристикам в будущем экспериментальном анализе изображений. Выберите Просмотр элементов управления анализом > > двоичной панели инструментов > Рисование объекта.
- Поезд Ай.
- Начните обучение программному обеспечению. Это откроет ящик Segment.ai поезда. Выберите NIS.ai > Поезд Segment.ai.
- В поле Train Segment.ai выберите исходный канал, который будет использоваться для обучения. Если открыты файлы из нескольких каналов, выберите в качестве исходного канала только один канал. Затем выберите соответствующие основные двоичные файлы истины, на которых будет обучаться Ai. Наконец, выберите количество итераций, необходимых для обучения Ai в зависимости от размера и распределения двоичных файлов.
ПРИМЕЧАНИЕ: Если двоичные файлы легко обнаруживаются из окружения и хорошо распределены по всему изображению, программному обеспечению может потребоваться менее 1000 итераций для обучения идентификации изображений. Если изображения имеют низкое отношение сигнал/шум, идеально выполнить не менее 1000 итераций во время обучения, чтобы позволить Ai идентифицировать реснички в тестовых образцах с высокой степенью достоверности. - Выберите папку назначения, чтобы сохранить обученный ai-файл (.sai), и нажмите кнопку Train, чтобы обучить программное обеспечение. Теперь программное обеспечение приступит к обучению идентификации ресничек на основе отслеживаемых двоичных файлов. Этот процесс занимает несколько часов.
ПРИМЕЧАНИЕ: При обучении программное обеспечение будет отображать график, показывающий потери в обучении. График первоначально покажет всплеск, прежде чем сузиться до идеальной потери около 1%, где он плато для остальной части обучения. Сохраните график для дальнейшего использования, установив флажок Сохранить снимок экрана графика в поле Train Segment.ai(Дополнительный рисунок 1). - Если требуется дальнейшее уточнение обучения, продолжайте обучение на том же наборе данных. Кроме того, можно обучить новому набору данных с точно такими же параметрами. Не рекомендуется обучать уже обученный ИИ на новом наборе данных с другими параметрами или другими объектами, представляющими интерес. Выберите Обучать segment.ai > Продолжить обучение на > Выберите Обученный AI Файл.
3. Идентификация ресничек с помощью обученного Ai
- Откройте экспериментальный набор данных.
- Откройте экспериментальные конфокальные изображения ресничек в программном обеспечении, преобразовав образцы .tif файлов в файлы .nd2, аналогично шагу 2.1. Выберите Файл > Импорт/Экспорт > Создать ND Файл из Последовательности Файлов.
ПРИМЕЧАНИЕ: Изображения должны иметь тот же размер пикселя, что и те, которые используются для обучения Ai. Если изображения уже имеют формат ND2, перейдите к шагу 3.3.
- Откройте экспериментальные конфокальные изображения ресничек в программном обеспечении, преобразовав образцы .tif файлов в файлы .nd2, аналогично шагу 2.1. Выберите Файл > Импорт/Экспорт > Создать ND Файл из Последовательности Файлов.
- Калибровка изображений.
- Введите размер пикселя в левом нижнем углу изображения. Щелкните правой кнопкой мыши Некалиброванный > Калибровка документа > Размер пикселя.
- Запустите обученный Ai на открытых файлах.
- Начните идентифицировать реснички с помощью Ai. Теперь программное обеспечение будет рисовать двоичные файлы на ресничках на основе обучения, которое оно получило на предыдущем шаге. Этот процесс займет несколько секунд. Выберите NIS.ai > Segment.ai.
ПРИМЕЧАНИЕ: Программное обеспечение предложит выбрать канал, если открыто несколько каналов. Каналы перечислены здесь по их соответствующим названиям. Если нет, поле с пометкой «Моно» будет выбрано автоматически.
- Начните идентифицировать реснички с помощью Ai. Теперь программное обеспечение будет рисовать двоичные файлы на ресничках на основе обучения, которое оно получило на предыдущем шаге. Этот процесс займет несколько секунд. Выберите NIS.ai > Segment.ai.
- Проверьте изображения на наличие неправильно идентифицированных двоичных файлов.
- После того, как Ai идентифицировал реснички и нарисовал двоичные файлы, проверьте изображения на наличие любых ошибочно идентифицированных объектов. При желании вручную удалите все неправильно идентифицированные двоичные файлы. Выберите Просмотр элементов управления анализом > > двоичной панели инструментов > Удалить объект.
4. Измерение длины и интенсивности ресничек
- Создайте новый рецепт общего анализа 3 (GA3).
- Теперь, когда реснички идентифицированы и сегментированы, приступайте к анализу различных параметров ресничек, таких как длина и интенсивность, используя инструмент GA3. Откроется новое окно с пустым пространством в центре, где будет определяться анализ. Выберите Изображение > Новый рецепт GA3.
- Выберите двоичные файлы для анализа.
- Поскольку реснички уже сегментированы с помощью Segment.ai, GA3 автоматически обнаружит двоичные файлы, соответствующим образом помеченные в соответствии с Ai, и включит узел. Выберите'Двоичные файлы > авто Detect_AI' или 'Двоичные файлы > рисование Object_AI'.
- Выберите каналы, необходимые для анализа. GA3 также автоматически определяет каналы на изображениях и отображает их вкладки в разделе Каналы.
- Удалите объекты, касающиеся границы фрейма.
- Поскольку Ai будет сегментировать все реснички, подобные объектам в кадре, он также обнаружит неполные реснички по краям кадра. Эти объекты могут быть удалены вручную на шаге 3.4 или автоматически удалены в GA3. Выберите Двоичная обработка > Удалить объекты > Касание границ.
- Выберите параметры для измерения ресничек.
- Перетащите параметры для измерения, такие как длина ресничек(длина)и интенсивность(сумма интенсивности объекта). Подключите узлы к соответствующему двоичному узлу (соединение A) и узлам канала (соединение B). Наведите указатель мыши на подключение узла, чтобы получить всплывающую подсказку, чтобы показать, к какому соединению принадлежит узел. Выберите Измерение > Размер объекта > Длина и Измерение > Интенсивности объекта > Sum Obj Intensity.
ПРИМЕЧАНИЕ: Узел Length подключается только к двоичному узлу, тогда как Sum Obj Intensity подключается как к двоичному, так и к канальному узлам.
- Перетащите параметры для измерения, такие как длина ресничек(длина)и интенсивность(сумма интенсивности объекта). Подключите узлы к соответствующему двоичному узлу (соединение A) и узлам канала (соединение B). Наведите указатель мыши на подключение узла, чтобы получить всплывающую подсказку, чтобы показать, к какому соединению принадлежит узел. Выберите Измерение > Размер объекта > Длина и Измерение > Интенсивности объекта > Sum Obj Intensity.
- Добавьте измерения в одну таблицу.
- Объедините все измерения в одну выходную таблицу, перетащив узел Append Column на блок-схему анализа и соединив его с узлами измерения Length и Sum Obj Intensity. Выберите Управление данными > Базовый > Добавить столбец.
- Измерьте реснички.
- Измерьте реснички, щелкнув Выполнить. Этот процесс занимает несколько минут, чтобы измерить все реснички на экспериментальных изображениях. Длина и интенсивность появятся в новом окне Результаты анализа.
ПРИМЕЧАНИЕ: Таблица может иногда включать данные из масок, которые Ai распознал как реснички, но были слишком малы, чтобы их можно было обнаружить человеческим глазом и устранить на шаге 3.4. Эти объекты могут быть удалены из набора данных с помощью фильтра перед статистическим анализом. Здесь фильтр в 1 мкм использовался для измерения длины ресничек in vitro на рисунке 2 и 2 мкм для ресничек in vivo. Это можно сделать перед экспортом данных с помощью приведенного ниже пути. Окно Выбор результатов анализа > Определение фильтра > Ввод значения > Использование фильтра.
- Измерьте реснички, щелкнув Выполнить. Этот процесс занимает несколько минут, чтобы измерить все реснички на экспериментальных изображениях. Длина и интенсивность появятся в новом окне Результаты анализа.
- Экспорт данных для статистического анализа.
5. Исследования колокализации
ПРИМЕЧАНИЕ: Анализ колокализации может быть включен в тот же рецепт GA3, который используется для измерения длины ресничек и анализа интенсивности. Если используется один и тот же рецепт, откройте файлы, как описано ниже, и измерьте длину и интенсивность обоих каналов вместе с коэффициентами колокализации в одном и том же конвейере анализа.
- Откройте экспериментальный набор данных.
- Откройте экспериментальные конфокальные изображения ресничек в программном обеспечении, преобразовав пример .tif файлов в файлы .nd2. Выберите Файл > Импорт/Экспорт > Создать ND Файл из Последовательности Файлов.
- Во всплывающем окне выберите монохромные файлы глубиной 16 бит из всех интересующих каналов из проводника, расположенного в первом столбце всплывающего окна. Выберите Multipoint или Z Series в первом раскрывающемся меню и введите значение, соответствующее общему количеству изображений или стеков соответственно.
- Во втором раскрывающемся списке выберите Длина волны и измените значение на общее количество каналов в папке. Программное обеспечение автоматически разблокирует окно выбора длины волны, расположенное в правом нижнем углу всплывающего окна. Используйте раскрывающееся меню Цвет, чтобы выбрать цвет каждого канала. Укажите каждому каналу другое имя в столбце Имя. После обновления всей информации нажмите кнопку Преобразовать. Программа автоматически сгенерирует файл All image, наложенный на все отдельные изображения со всех выбранных каналов.
- Калибровка изображений.
- Введите размер пикселя в левом нижнем углу изображения. Щелкните правой кнопкой мыши Некалиброванный > Калибровка документа > Размер пикселя.
- Запустите обученный Ai на первом канале.
- Начать идентификацию ресничек по одному из открытых каналов (например, ACIII; Рисунок 5А) с помощью Ai. Теперь программное обеспечение будет рисовать двоичные файлы на ресничках, помеченных ACIII, на основе обучения, которое оно получило для этого канала. Этот процесс займет несколько секунд. Выберите исходные каналы NIS.ai > Segment.ai > > ACIII.
- Запустите обученный Ai на втором канале.
- Начать идентификацию ресничек на другом открытом канале (например, MCHR1; Рисунок 5B) с помощью Ai. Теперь программное обеспечение будет рисовать двоичные файлы на ресничках, помеченных MCHR1, на основе обучения, которое оно получило для этого канала. Этот процесс займет несколько минут. Выберите исходные каналы NIS.ai > Segment.ai > > MCHR1.
- Проверьте изображения на наличие неправильно идентифицированных двоичных файлов.
- После того, как Ai идентифицировал реснички и нарисовал двоичные файлы, проверьте изображения на наличие неправильно идентифицированных объектов. При необходимости вручную удалите все неправильно идентифицированные двоичные файлы. Выберите Просмотр элементов управления анализом > > двоичной панели инструментов > Удалить объект.
- Создайте новый рецепт GA3.
- Теперь, когда реснички идентифицированы и сегментированы, переходите к анализу колокализации с использованием инструмента GA3. Откроется новое окно с пустым пространством в центре, где будет определяться анализ. Будет сгенерировано окно со всеми идентифицированными двоичными файлами и каналами. Убедитесь, что все необходимые каналы и двоичные файлы, необходимые для анализа, присутствуют и выбраны. Выберите Изображение > Новый рецепт GA3.
- Удалите объекты, касающиеся границы фрейма.
- Поскольку Ai будет сегментировать все ресничкообразные объекты в кадре, он также обнаружит неполные реснички по краям кадра. Эти объекты могут быть удалены вручную на шаге 5.5 или автоматически удалены в GA3. Выберите Двоичная обработка > Удалить объекты > Касание границ.
- Настройте путь колокализации в GA3.
- Чтобы измерить перекрытие двух каналов внутри ресничек, используйте коэффициентную корреляцию Мандера. Перетащите узел Коэффициент Мандерса в пустое пространство рецепта GA3 и подключите его к соответствующему двоичному файлу и каналам. Здесь «соединение A» соединяется с двоичным файлом ACIII, «соединение B» с каналом MCHR1 и «соединение C» с каналом ACIII для определения перекрытия MCHR1 в двоичном коде ACIII. Выберите Измерение > Ратиометрия объектов > Коэффициент Мандерса.
ПРИМЕЧАНИЕ: Программное обеспечение позволяет измерять колокализацию с использованием корреляции коэффициентов Пирсона, используя те же шаги, которые описаны в этомпротоколе 40.
- Чтобы измерить перекрытие двух каналов внутри ресничек, используйте коэффициентную корреляцию Мандера. Перетащите узел Коэффициент Мандерса в пустое пространство рецепта GA3 и подключите его к соответствующему двоичному файлу и каналам. Здесь «соединение A» соединяется с двоичным файлом ACIII, «соединение B» с каналом MCHR1 и «соединение C» с каналом ACIII для определения перекрытия MCHR1 в двоичном коде ACIII. Выберите Измерение > Ратиометрия объектов > Коэффициент Мандерса.
- Добавьте измерения в одну таблицу.
- Объедините все измерения в одной выходной таблице. Выберите Управление данными > Базовый > Добавить столбец.
- Измерение колокализации.
- Измерьте реснички, щелкнув Выполнить. Этот процесс занимает несколько минут, чтобы измерить все реснички на экспериментальных изображениях. Данные появятся в новом окне Результаты анализа.
- Экспорт данных для статистического анализа.
Результаты
Обучение ИИ для идентификации ресничек
Измерение и оценка структурной длины и состава ресничек может быть утомительным, трудоемким и подверженным ошибкам процессом. Здесь мы используем Ai для точного сегментирования ресничек из большого пула изображений и анализа их длины и интенсивности с помощью инструмента анализа(рисунок 1). Все подходы КИ требуют учебных шагов для их реализации. Мы создали обучающий конвейер для распознавания ресничек, который выполнялся путем ручного нанесения бинарных масок на цилиарные структуры. Эта информация затем используется для обучения Ai на основе характеристик пикселей в примененных двоичных файлах. В качестве общего ориентира, обучение включает в себя программное обеспечение, проходящее через несколько итераций, примерно 1000, и считается оптимальным, если частота потерь или ошибок обучения составляет менее 1%. Однако количество итераций и ошибок в процессе обучения может варьироваться в зависимости от образцов изображений, используемых для обучения. Например, после наших тренировок с использованием изображений ресничек нейронов in vitro частота ошибок составила 1,378% по сравнению с 3,36% для изображений разреза мозга in vivo (дополнительный рисунок 1). После завершения обучения ИИ можно использовать для сегментирования ресничек из экспериментальных изображений в течение нескольких секунд, а результирующие бинарные маски используются для измерения структурных параметров. Это устраняет необходимость сегментировать объекты с использованием традиционного метода порогов интенсивности, который может быть затруднен на изображениях с высоким фоновым шумом или когда объекты находятся в непосредственной близости друг от друга. Ai также снижает вероятность ошибок и смещений, применяя один и тот же алгоритм ко всем изображениям, независимо от пользователя.
Измерение длины ресничек с помощью GA3
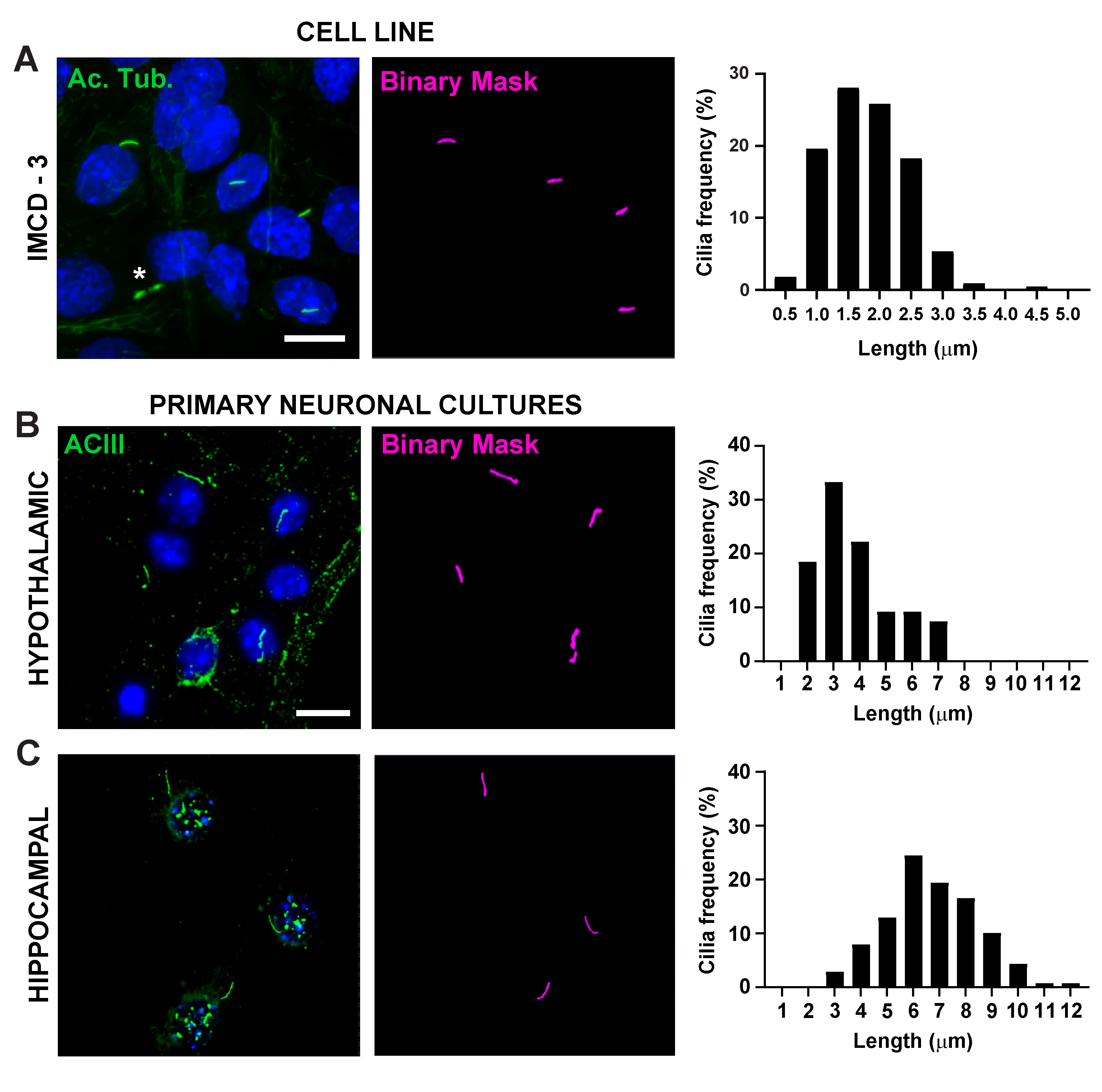
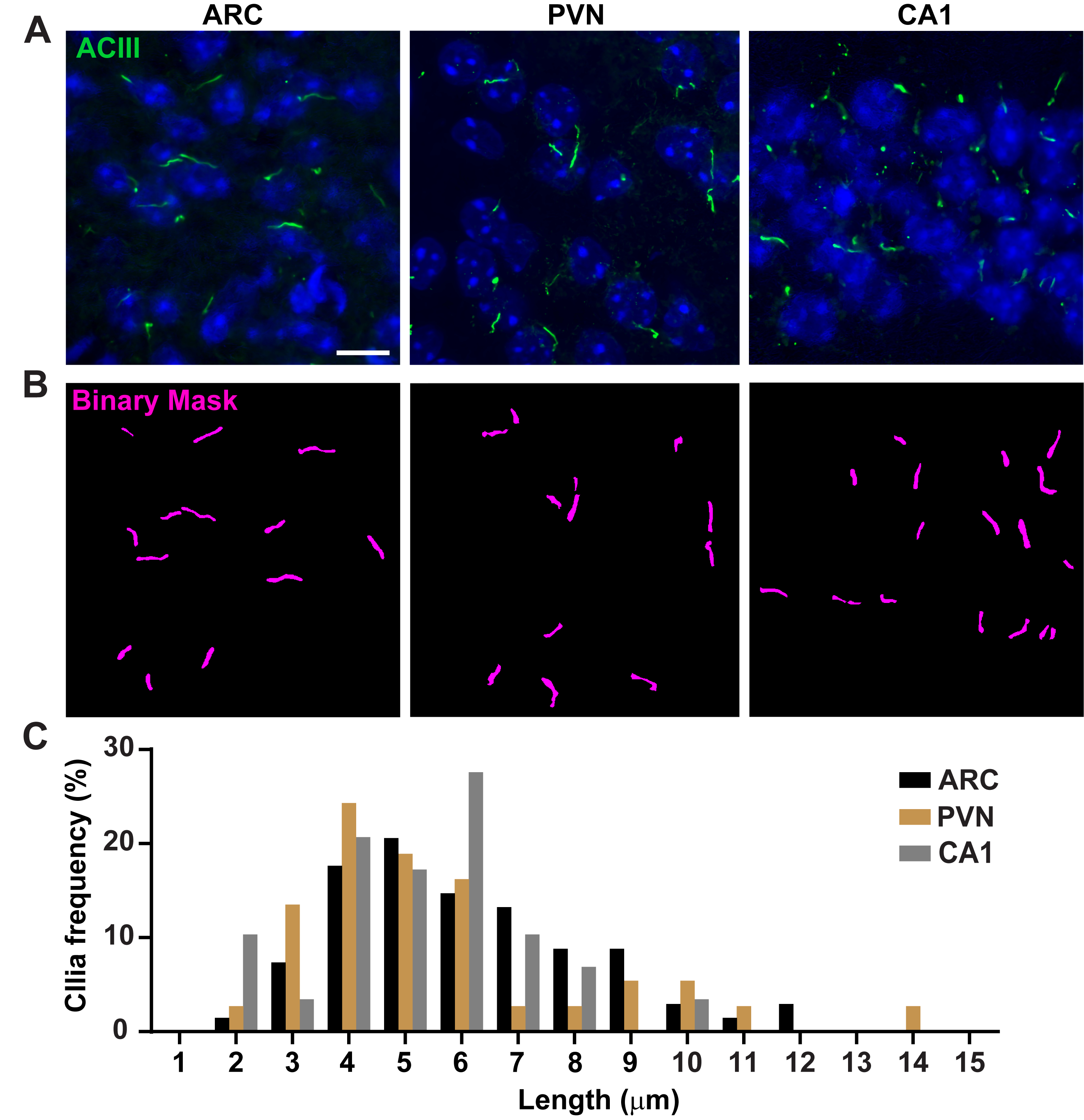
Длина ресничек жестко регулируется и связана с функциональным воздействием на цилиарную сигнализацию16,19. Здесь мы измерили длину ресничек с помощью конвейера анализа в программном обеспечении NIS Elements под названием General Analysis 3 или GA3. GA3 помогает объединить несколько инструментов в одном рабочем процессе для создания настраиваемых процедур для каждого эксперимента. Мы начали с измерения длины ресничек в клеточной линии. Реснички на клетках внутреннего медуллярного собирательного протока мыши (IMCD-3) были иммуномаркированы ацетилированным тубулином и визуализированы с помощью конфокального микроскопа. Мы измерили длину ресничек с помощью GA3 после сегментации с segment.ai(дополнительный рисунок 3A). В то время как ацетилированный α-тубулин предпочтительно находится в первичной ресничке, он также встречается в других областях, богатых микротрубочками, таких как цитоскелет, а также цитокинетический мост. Обученный Аи правильно идентифицировал реснички на изображении, но не другие нецилиарные, ацетилированные тубулиновые положительные структуры. Реснички на клетках IMCD варьировались от 0,5 мкм до 4,5 мкм со средней длиной 1,8 ± 0,04 мкм(рисунок 2А). Затем мы проверили способность ИИ измерять длину ресничек в первичных нейронных культурах. Мы культивировали нейроны из гипоталамуса и гиппокампа неонатальных мышей в течение 10 дней и иммуномаркировали их маркером ресничек аденилатциклазой III (ACIII)21,41. При анализе нейронных культур мы сочли полезным применить фильтр перед статистическим анализом длин. Из-за более низкого отношения сигнал/шум было идентифицировано несколько объектов размером менее 1 мкм, которые не были ресничками. Поэтому мы отфильтровали данные, чтобы исключить любые объекты длиной менее 1 мкм, чтобы гарантировать, что анализируются только реснички. В культивируемых гипоталамических нейронах длина ресничек варьировалась от 2 мкм до 7 мкм со средней длиной 3,8 ± 0,19 мкм(рисунок 2B). Интересно, что культивируемые реснички нейронов гиппокампа были длиннее со средней длиной 6,73 ±0,15 мкм(рисунок 2C). Сообщалось, что различные ядра нейронов в гипоталамусе имеют различную длину ресничек и что эти реснички изменяют свою длину в ответ на физиологические изменения определенным образом ядра19,23. Поэтому мы также пометили участки гипоталамического мозга у взрослых самцов мышей C57BL/6J с помощью ACIII и визуализировали дугообразное ядро (ARC) и паравентрикулярное ядро (PVN). Используя GA3 для измерения длины ресничек, мы заметили, что in vivo гипоталамические реснички оказались длиннее, чем реснички in vitro. В частности, гипоталамические реснички in vivo варьируются от 1 мкм до примерно 15 мкм(рисунок 3). Не было выявлено существенных различий между длинами ресничек в PVN (5,54 ± 0,0,42 мкм) и в ARC (6,16 ± 0,27 мкм)(рисунок 3C)23. Аналогичным образом, реснички в области cornu ammonis (CA1) гиппокампа имеют более узкий диапазон длины от 1 мкм до 10 мкм со средней длиной 5,28 ± 0,33 мкм(рисунок 3). В соответствии с ранее опубликованными исследованиями, наш анализ с использованием инструментов Ai и GA3 показал, что реснички из разных областей мозга показывают разнообразие по длине19,23. Более того, используя этот подход Ai, мы можем быстро оценить большое количество ресничек.
Измерение состава ресничек с помощью GA3
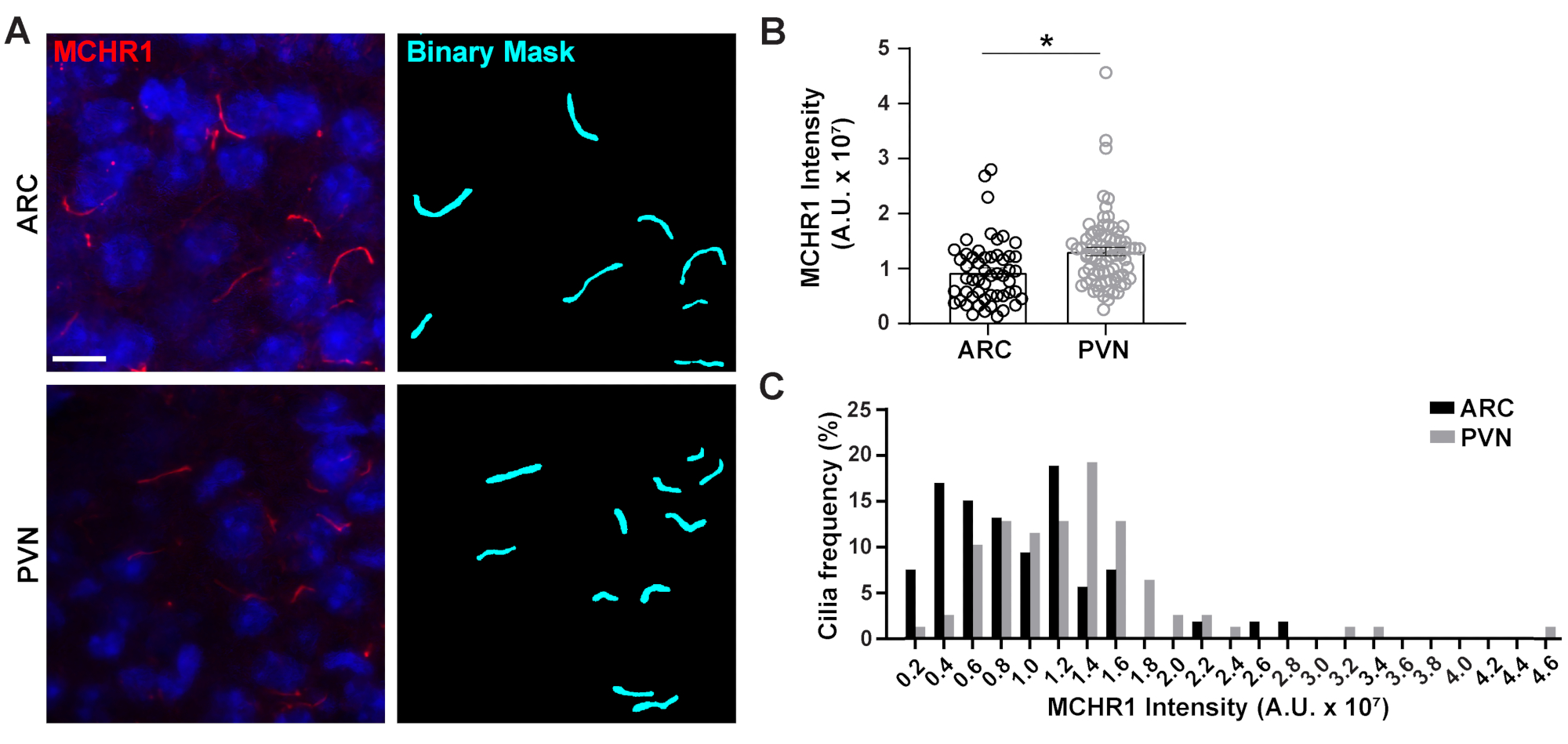
Первичная ресничка является сигнальным центром для многих путей, которые используют различные типы белков для выполнения уникальных функций, таких как моторные белки, внутрифлягеллярные транспортные белки иGPCR,чтобы назвать несколько3,24,42,43. Поддержание соответствующих уровней этих белков в ресничке важно для правильного функционирования и часто зависит от клеточного контекста. Флуоресцентная маркировка этих белков не только позволила нам визуализировать их, но и количественно оценить их интенсивность в качестве меры количества меченого белка в относительно небольшом отсеке20. Поэтому мы стремились определить интенсивность цилиарного GPCR, рецептора концентрационного гормона меланина 1 (MCHR1), in vivo как в ARC, так и в PVN гипоталамуса взрослых самцов мышей24,44. Используя Ai и GA3, мы измерили длину положительных ресничек MCHR1 вместе с интенсивностью, чтобы убедиться, что подсчитываемые объекты были ресничками(дополнительный рисунок 3A). Мы исключили объекты после анализа, которые были менее 2 мкм в длину, и проанализировали интенсивности оставшихся бинарных масок. Интересно, что мы обнаружили, что интенсивность цилиарного MCHR1 в PVN значительно выше, чем в ARC, что указывает на более сильное присутствие цилиарного MCHR1 в PVN(рисунок 4). Необходимы дальнейшие исследования для определения значимости цилиарного MCHR1 в этих нейронных цепях. Мы также измерили интенсивность цилиарного MCHR1 в первичных культивируемых нейронах гипоталамуса и гиппокампа. Реснички из обеих культур демонстрируют широкое распределение интенсивностей MCHR1, что свидетельствует о наличии гетерогенных нейронных популяций(дополнительный рисунок 2). Таким образом, использование сложных аналитических инструментов, таких как Ai и GA3, позволяет оценить гетерогенность ресничек в одной ткани или между несколькими тканями. Будет интересно посмотреть, показывают ли другие нейронные GPCR аналогичные различия в их локализации в нейронах той же ткани и изменяется ли это в ответ на физиологические изменения.
Колокализация
Хотя измерение интенсивности флуоресценции в полном поле изображения может создать впечатление белка, оно не может предоставить такую информацию, как пространственное распределение или близость к другим близлежащим белкам и клеточным структурам. Здесь мы измерили перекрытие MCHR1 с ACIII в качестве маркера ресничек, построив интенсивность MCHR1 против интенсивности ACIII для каждой двоичной маски(рисунок 5). График показывает, что большинство ресничек являются положительными как для ACIII, так и для MCHR1, хотя некоторые реснички показывают более сильную экспрессию одного канала над другим. Кроме того, есть некоторые реснички, которые показывают присутствие ACIII или MCHR1, что видно из точек, которые лежат непосредственно на оси x и оси Y соответственно. Чтобы количественно оценить это перекрытие, мы измерили коэффициент перекрытия Мандера и сравнили степень экспрессии MCHR1 в нейронных ресничках ARC и PVN40. Интересно, что наш анализ показал, что произошло значительное увеличение коэффициентов PVN (0,6382 ± 0,0151), чем в ARC (0,5430 ± 0,0181)(рисунок 5C). Это согласуется с нашими предыдущими данными, где мы наблюдали более высокие интенсивности MCHR1 в PVN по сравнению с ARC(рисунок 4). Эти данные свидетельствуют о том, что, как и длина ресничек, паттерн экспрессии MCHR1 в цилиарном компартменте варьируется в разных областях мозга. Используя тот же конвейер анализа, можно будет определить, показывают ли другие цилиарные GPCR, такие как нейропептидный рецептор Y типа 2 (NPY2R) и рецептор соматостатина типа 3 (SSTR3), аналогичные количества разнообразия.
Измерение профиля интенсивности вдоль реснички
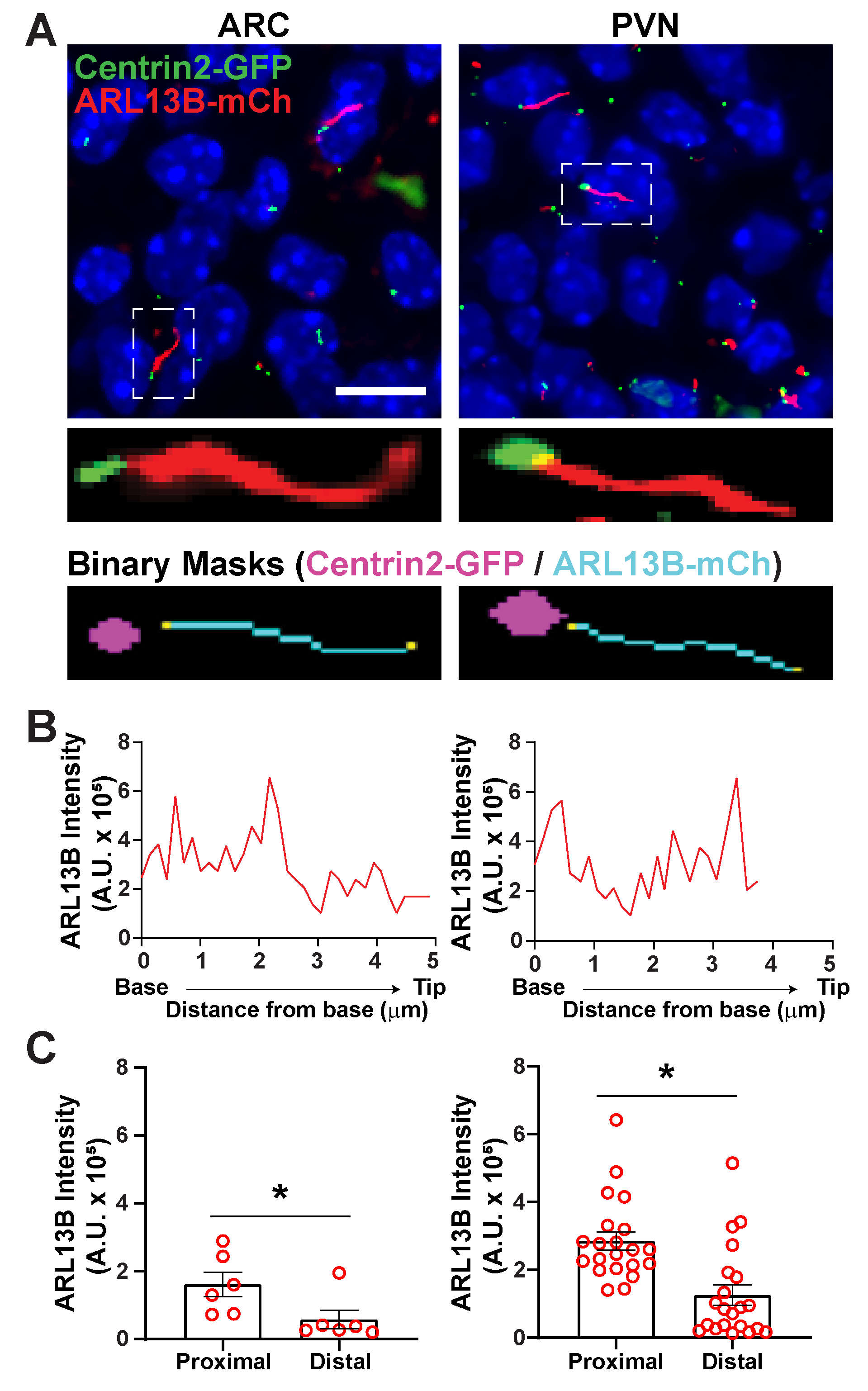
После того, как реснички были идентифицированы с использованием segment.ai, рецепт GA3 может быть изменен для объединения анализа ресничек с идентификацией других структур, представляющих интерес на изображении. Например, маркировка базальными маркерами тела полезна для идентификации полярности ресничек. Чтобы сделать этот анализ, мы изобразили участки гипоталамического мозга у мышей P0, которые экспрессируют ARL13B-mCherry и Centrin2-GFP, и изобразили ARC и PVN34. Здесь реснички были идентифицированы с использованием Ai, как и раньше, но теперь модифицированный рецепт GA3 включает идентификацию Centrin2-GFP, центриолярного белка, обнаруженного в основании ресничек(дополнительный рисунок 3B). По маркировке Centrin2-GFP основание ресничек можно отличить от кончиков ARL13B-mCherry положительных ресничек(рисунок 6A). Затем, вместо того, чтобы измерять интенсивность во всех ресничках, мы можем измерить изменения интенсивности ARL13B по длине ресничек(рисунок 6B). Мы также можем сравнить различия в интенсивности ARL13B между проксимальными концами и дистальными концами ресничек. Для этого мы разделили длину ресницы на 1-микронные бункеры, начиная с основания, и обозначили первый микронный бункер как проксимальный конец, а последний микронный бункер как дистальный конец. Наш анализ показал, что ближе к основанию присутствует значительно больше ARL13B, чем кончик ресничек как в ARC, так и в PVN, и это согласуется с ранее опубликованными исследованиями на хондроцитах человека45 (рисунок 6C). В этом типе анализа вместо применения фильтра длины для исключения небольших нецилярных объектов из анализа анализируются только реснички, связанные с маркировкой Centrin2-GFP. Это может быть выгодно в ситуациях, когда генетические мутации приводят к очень коротким ресничкам, или если были вовлечены изменения в поддоменах ресничек, таких как переходная зона или кончик. Идентификация ресничек с помощью анализа Ai и GA3 легко адаптируется и может быть адаптирована к различным сложным исследовательским вопросам.

Рисунок 1. Рабочий процесс для измерения длины и интенсивности ресничек с помощью Ai. (A) Для обучения Ai двоичные файлы рисуются вокруг интересующих объектов (ресничек) на необработанных обучающих изображениях. Используя нарисованные двоичные файлы, сегмент Ai обучается распознавать форму и интенсивность пикселей ресничек. (B) Затем обученный сегмент Ai применяется к необработанным экспериментальным изображениям. Он рисует двоичные файлы на объектах, которые он распознает как реснички. Эти двоичные файлы могут быть уточнены, чтобы убедиться, что анализируются все и только реснички. (C) Программа GA3 построена для анализа интенсивности и длины объектов, распознаваемых Ai. (D) Записи импортируются в таблицу в программном обеспечении. Затем эту таблицу можно экспортировать для дальнейшего анализа. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 2. Измерение длины ресничек in vitro. Репрезентативные изображения ресничек в (A)клетках IMCD (зеленый, ацетилированный тубулин)(B),первичных гипоталамических культурах (зеленый, ACIII) и(C)гиппокампальных культурах (зеленый, ACIII). Обученный Ai использовался для распознавания ресничек, как показано в бинарной маске (пурпурный), а затем GA3 использовался для измерения длины ресничек. Распределение длины ресничек отображается в процентах от ресничек в 0,5 или 1,0-микронных бункерах. * указывает на цитокинетический мост, должным образом не распознаваемый Аи. n=225 ресничек в клетках IMCD из 3 реплик, 54 ресничек в гипоталамике и 139 ресничек в культурах гиппокампа от 3 животных. Шкала 10 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 3. Измерение длины ресничек in vivo. (A) Репрезентативные изображения ресничек (зеленый, ACIII) в ARC, PVN и CA1 секций мозга взрослых мышей. (B) Обученный Ai в ЭЛЕМЕНТАХ NIS использовался для распознавания ресничек, как показано в бинарной маске (пурпурный), а затем GA3 использовался для измерения длины ресничек. (C) Распределение длины ресничек отображается как процент ресничек в одномикронных бункерах. n= 68 ресничек в ARC, 36 в PVN и 29 в CA1 у 3 животных. Шкала 10 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 4. Ai помог измерить интенсивность окрашивания ресничек гипоталамических нейронов. (A) Репрезентативные изображения ресничек (MCHR1, красный) в ARC и PVN секций мозга взрослых мышей. Обученный Ai в ЭЛЕМЕНТАХ NIS использовался для распознавания ресничек, как показано в бинарной маске (голубой), а затем GA3 использовался для измерения интенсивности окрашивания MCHR1 в ресничках. (B)Интенсивность MCHR1 отображается как средняя ± S.E.M. Каждая точка представляет собой ресничку. * p < 0,05, T-тест студента. (C)Распределение интенсивности MCHR1 отображается как процент ресничек в бункерах 0,2 x 107 произвольных единиц (A. U.). n= 53 ресничек в ARC, 78 в PVN у 3 животных. Шкала 10 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 5. Анализ колокализации ресничек с помощью искусственного интеллекта. (А, Б) Репрезентативные изображения ресничек в ARC и PVN соответственно. Реснички маркируются ACIII (зеленый) и MCHR1 (красный). Обученный Ai в NIS Elements использовался для распознавания ресничек, как показано в бинарной маске (пурпурный для меченых ACIII ресничек, голубой для меченых MCHR1 ресничек). GA3 использовался для распознавания ресничек, содержащих как ACIII, так и MCHR1. (C)Значения коэффициента перекрытия Мандерса (MOC) отображаются в виде среднего ± S.E.M. Каждая точка представляет собой ресничку. * p < 0,05, T-тест студента. (D)Диаграмма рассеяния интенсивности MCHR1 по сравнению с интенсивностью ACIII в ARC и PVN. Каждая точка представляет собой ресничку. n= 72 ресничек в ARC, 47 в PVN от 3 животных. Шкала 10 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 6. Анализ ресничек и базального тела. (A) Репрезентативные изображения ресничек (красный, ARL13B-mCherry) и базального маркера тела (зеленый, Centrin2-GFP) в ARC и PVN мышей P0. Обученный Ай использовался для распознавания ресничек, как показано в бинарной маске (голубой). Бинарная маска для базального тела (пурпурного) была нарисована путем порога в рецепте GA3. (B) Репрезентативная интенсивность сканирования реснички. (C)Интенсивность ARL13B на проксимальном и дистальном концах Ai идентифицировала реснички, изображенные как средние ± S.E.M. Проксимальный и дистальный концы определяются как область в пределах первой длины 1 мкм и последней длины 1 мкм соответственно от основания реснички. Каждая точка представляет собой ресничку. * p < 0,05. n = 6 ресничек в ARC от 2 животных и 21 ресничка в PVN от 3 животных. Шкала 10 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Дополнительный рисунок 1. Графики потерь обучения ai. (A, B) Графики, показывающие потерю тренировочных segment.ai на нейронных ресничках in vitro и in vivo соответственно. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный рисунок 2. Ai Assisted Cilia окрашивание интенсивности интенсивности in vitro нейрональных ресничек. (А, Б) Репрезентативные изображения ресничек (MCHR1, красный) в первичных гипоталамических и гиппокампальных культурах соответственно. Обученный Ai в ЭЛЕМЕНТАХ NIS использовался для распознавания ресничек, как показано в бинарной маске (голубой), а затем GA3 использовался для измерения интенсивности окрашивания MCHR1 в ресничках. Распределение интенсивности MCHR1 отображается как процент ресничек в 1000 A.U. bins для гипоталамических и 2000 A.U. bins для гиппокампальных культур. n= 30 ресничек в гипоталамике и 106 ресничек в культурах гиппокампа от 3 животных. Шкала баров 10 мкм. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный рисунок 3. Общий анализ 3 рецепта анализа ресничек. (A) Простой общий анализ (GA3) рецепт для измерения длины ресничек, интенсивности и коэффициента Мандера. (B) Комплексный рецепт GA3 для измерения интенсивности по длине реснички с использованием маркера для базального тела. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Обсуждение
Измерения длины и интенсивности являются распространенными способами анализа первичных ресничек, однако в полевых условиях не существует стандартизированного традиционного метода. Идентификация и количественная оценка первичных ресничек с помощью программного обеспечения, такого как ImageJ, занимает много времени и подвержена предвзятости и ошибкам пользователей. Это затрудняет точный анализ больших наборов данных. Здесь мы показываем, что использование программы ИИ может преодолеть многие из этих проблем, что делает достижимым высокопроизводительный анализ первичных ресничек. Здесь мы описываем процедуру обучения приложения на основе ИИ для распознавания первичных ресничек и описываем шаги, необходимые для анализа длины и интенсивности.
В то время как первоначальное обучение Ai распознаванию ресничек требует значительного времени от пользователя, после его завершения его можно использовать на любом наборе данных, полученных с теми же параметрами. Двоичная маска, генерируемая Ai, изменяется таким образом, что любые ошибки могут быть исправлены. Однако ошибки в идентификации ресничек должны сигнализировать пользователю о том, что ИИ нуждается в дальнейшем обучении с помощью дополнительных изображений. Одним из основных преимуществ этого метода является то, что ИИ можно обучить распознавать реснички в различных типах образцов как в 2D, так и в 3D. Предыдущие методы анализа, созданные в лабораториях, имеют различные ограничения, включая требование ручного порога для идентификации и проблемы идентификации ресничек, изображенных из участков ткани, где плотность клеток высока36,46,47. Эти методы также специализируются на анализе ресничек, тогда как анализ с использованием программного обеспечения NIS Elements может оценивать несколько аспектов изображений одновременно. Поскольку ai, описанный здесь, является частью программного пакета NIS Elements, изображения, полученные с помощью микроскопа Nikon, могут быть легко продолжены до анализа. Однако для использования этого метода визуализация с помощью Nikon не требуется. Независимо от формата захваченных файлов необработанных данных, файлы «.tif» могут быть открыты NIS Elements для использования в Ai.
Это приложение Ai в NIS Elements широко доступно и, возможно, уже является частью программного обеспечения для анализа изображений, используемого лабораториями, изучающими первичные реснички. С расширением распространенности технологии ИИ другое программное обеспечение для визуализации может расширить свои возможности анализа, включив в них аналогичный модуль ИИ. Применение анализа Ai к идентификации ресничек может быть использовано для нескольких различных аспектов анализа ресничек. В то время как мы изложили методы для нескольких простых анализов, таких как длина(рисунок 2 и 3),интенсивность(рисунок 4)и колокализация(рисунок 5),более сложный анализ может быть добавлен к рабочему процессу анализа GA3, как на рисунке 6. Например, вместо измерения интенсивности полной реснички различия в интенсивности в подобласти реснички могут представлять интерес для оценки субцилиарной локализации. Различия в интенсивности в субрегионе реснички могут указывать на то, что белок накапливается на кончике или основании реснички, например, как белки Gli обогащаются на кончике ресничек48. Кроме того, это приложение Ai может быть использовано для легкого выявления различий между генотипами или группами лечения. Хотя наша лаборатория в основном использует этот метод для анализа ресничек, изображенных из участков мозга или нейронных культур, он может быть применен к изображениям, полученным из различных клеточных линий или других типов тканей. Гибкость типа образца, для которого может быть использовано это приложение, делает этот метод анализа ценным для многих различных групп, изучающих первичные реснички или любую дискретную органеллу, которая оценивается, такую как митохондрии, ядро или ER.
Раскрытие информации
Соавтор Уэсли Льюис является сотрудником Nikon. Раскрытие финансовой информации отсутствует.
Благодарности
Эта работа финансировалась Национальным институтом диабета и заболеваний пищеварительной системы и почек R01 DK114008 для NFB и грантом Американской кардиологической ассоциации #18PRE34020122 RB. Мы благодарим Рича Грускина, генерального директора Nikon Software, Мелиссу Бентли, Кортни Хейкрафт и Терезу Мастраччи за проницательные комментарии к рукописи.
Материалы
| Name | Company | Catalog Number | Comments |
| Intel Xeon, 3.6 GHz, 32GB RAM | Intel Corporation | W-2123 | Processor used for running NIS Elements. |
| Nikon Elements Software | Nikon Instruments Inc. | - | Ai and GA3 software |
| Quadro RTX 4000 Graphics card | NVIDIA Corporation | Quadro RTX 4000 | |
| Windows 10 Professional 64-bit | Microsoft Inc. | - | Operating system used for running NIS Elements |
| Workstation | HP Development Company, L.P. | HP Z4G4 | Workstation used for running NIS Elements |
Ссылки
- Wang, L., et al. Ciliary gene RPGRIP1L is required for hypothalamic arcuate neuron development. JCI Insight. 4 (3), (2019).
- Siljee, J. E., et al. Subcellular localization of MC4R with ADCY3 at neuronal primary cilia underlies a common pathway for genetic predisposition to obesity. Nature Genetics. 50 (2), 180-185 (2018).
- Davenport, J. R., et al. Disruption of intraflagellar transport in adult mice leads to obesity and slow-onset cystic kidney disease. Current Biology. 17 (18), 1586-1594 (2007).
- Berbari, N. F., O'Connor, A. K., Haycraft, C. J., Yoder, B. K. The primary cilium as a complex signaling center. Current Biology. 19 (13), 526-535 (2009).
- Walz, G. Role of primary cilia in non-dividing and post-mitotic cells. Cell Tissue Research. 369 (1), 11-25 (2017).
- Nachury, M. V., Mick, D. U. Establishing and regulating the composition of cilia for signal transduction. Nature Reviews. Molecular Cell Biology. 20 (7), 389-405 (2019).
- Goetz, S. C., Anderson, K. V. The primary cilium: a signalling centre during vertebrate development. Nature Reviews Genetics. 11 (5), 331-344 (2010).
- Engle, S. E., Bansal, R., Antonellis, P. J., Berbari, N. F. Cilia signaling and obesity. Seminars in Cell and Developmental Biology. , (2020).
- Reiter, J. F., Leroux, M. R. Genes and molecular pathways underpinning ciliopathies. Nature reviews. Molecular Cell Biology. 18 (9), 533-547 (2017).
- Waters, A. M., Beales, P. L. Ciliopathies: an expanding disease spectrum. Pediatric Nephrology. 26 (7), 1039-1056 (2011).
- Hildebrandt, F., Benzing, T., Katsanis, N. Ciliopathies. New England Journal of Medicine. 364 (16), 1533-1543 (2011).
- Vaisse, C., Reiter, J. F., Berbari, N. F. Cilia and Obesity. Cold Spring Harbor Perspectives in Biology. 9 (7), (2017).
- Berbari, N. F., et al. Leptin resistance is a secondary consequence of the obesity in ciliopathy mutant mice. Proceedings of the National Academy of Sciences of the United States of America. 110 (19), 7796-7801 (2013).
- Jacobs, D. T., et al. Dysfunction of intraflagellar transport-A causes hyperphagia-induced obesity and metabolic syndrome. Disease Models & Mechanisms. 9 (7), 789-798 (2016).
- Arsov, T., et al. Fat aussie--a new Alström syndrome mouse showing a critical role for ALMS1 in obesity, diabetes, and spermatogenesis. Molecular Endocrinology. 20 (7), 1610-1622 (2006).
- Tam, L. W., Ranum, P. T., Lefebvre, P. A. CDKL5 regulates flagellar length and localizes to the base of the flagella in Chlamydomonas. Molecular Biology of the Cell. 24 (5), 588-600 (2013).
- Rajagopalan, V., Subramanian, A., Wilkes, D. E., Pennock, D. G., Asai, D. J. Dynein-2 affects the regulation of ciliary length but is not required for ciliogenesis in Tetrahymena thermophila. Molecular Biology of the Cell. 20 (2), 708-720 (2009).
- Bengs, F., Scholz, A., Kuhn, D., Wiese, M. LmxMPK9, a mitogen-activated protein kinase homologue affects flagellar length in Leishmania mexicana. Molecular Microbiology. 55 (5), 1606-1615 (2005).
- Han, Y. M., et al. Leptin-promoted cilia assembly is critical for normal energy balance. Journal of Clinical Investigation. 124 (5), 2193-2197 (2014).
- Caspary, T., Marazziti, D., Berbari, N. F., Satir, P., Tvorup Christensen, S. . Cilia: Methods and Protocols. , 203-214 (2016).
- Bishop, G. A., Berbari, N. F., Lewis, J., Mykytyn, K. Type III adenylyl cyclase localizes to primary cilia throughout the adult mouse brain. Journal of Comparative Neurology. 505 (5), 562-571 (2007).
- Domire, J. S., Mykytyn, K. Markers for neuronal cilia. Methods in Cell Biology. 91, 111-121 (2009).
- Sun, J. S., et al. Ventromedial hypothalamic primary cilia control energy and skeletal homeostasis. Journal of Clinical Investigation. 131 (1), (2021).
- Berbari, N. F., Johnson, A. D., Lewis, J. S., Askwith, C. C., Mykytyn, K. Identification of ciliary localization sequences within the third intracellular loop of G protein-coupled receptors. Molecular Biology of the Cell. 19 (4), 1540-1547 (2008).
- Brailov, I., et al. Localization of 5-HT(6) receptors at the plasma membrane of neuronal cilia in the rat brain. Brain Research. 872 (1-2), 271-275 (2000).
- Domire, J. S., et al. Dopamine receptor 1 localizes to neuronal cilia in a dynamic process that requires the Bardet-Biedl syndrome proteins. Cellular and Molecular Life Sciences. 68 (17), 2951-2960 (2011).
- Handel, M., et al. Selective targeting of somatostatin receptor 3 to neuronal cilia. Neuroscience. 89 (3), 909-926 (1999).
- Koemeter-Cox, A. I., et al. Primary cilia enhance kisspeptin receptor signaling on gonadotropin-releasing hormone neurons. Proceedings of the National Academy of Sciences of the United States of America. 111 (28), 10335-10340 (2014).
- Mukhopadhyay, S., et al. The ciliary G-protein-coupled receptor Gpr161 negatively regulates the Sonic hedgehog pathway via cAMP signaling. Cell. 152 (1-2), 210-223 (2013).
- Berman, S. A., Wilson, N. F., Haas, N. A., Lefebvre, P. A. A novel MAP kinase regulates flagellar length in Chlamydomonas. Current Biology. 13 (13), 1145-1149 (2003).
- Nguyen, R. L., Tam, L. W., Lefebvre, P. A. The LF1 gene of Chlamydomonas reinhardtii encodes a novel protein required for flagellar length control. Genetics. 169 (3), 1415-1424 (2005).
- Tam, L. W., Wilson, N. F., Lefebvre, P. A. A CDK-related kinase regulates the length and assembly of flagella in Chlamydomonas. Journal of Cell Biology. 176 (6), 819-829 (2007).
- O'Connor, A. K., et al. An inducible CiliaGFP mouse model for in vivo visualization and analysis of cilia in live tissue. Cilia. 2 (1), 8 (2013).
- Bangs, F. K., Schrode, N., Hadjantonakis, A. K., Anderson, K. V. Lineage specificity of primary cilia in the mouse embryo. Nature Cell Biology. 17 (2), 113-122 (2015).
- Delling, M., et al. Primary cilia are not calcium-responsive mechanosensors. Nature. 531 (7596), 656-660 (2016).
- Saggese, T., Young, A. A., Huang, C., Braeckmans, K., McGlashan, S. R. Development of a method for the measurement of primary cilia length in 3D. Cilia. 1 (1), 11 (2012).
- Kobayashi, Y., Hamamoto, A., Saito, Y. Analysis of ciliary status via G-protein-coupled receptors localized on primary cilia. Microscopy. 69 (5), 277-285 (2020).
- Zhou, L. Q., et al. Artificial intelligence in medical imaging of the liver. World Journal of Gastroenterology. 25 (6), 672-682 (2019).
- Naugler, C., Church, D. L. Automation and artificial intelligence in the clinical laboratory. Critical Reviews in Clinical Laboratory Sciences. 56 (2), 98-110 (2019).
- Dunn, K. W., Kamocka, M. M., McDonald, J. H. A practical guide to evaluating colocalization in biological microscopy. American Journal of Physiology-Cell Physiology. 300 (4), 723-742 (2011).
- Bansal, R., et al. Hedgehog Pathway Activation Alters Ciliary Signaling in Primary Hypothalamic Cultures. Frontiers in Cellular Neuroscience. 13, 266 (2019).
- Jin, H., et al. The conserved Bardet-Biedl syndrome proteins assemble a coat that traffics membrane proteins to cilia. Cell. 141 (7), 1208-1219 (2010).
- Liew, G. M., et al. The intraflagellar transport protein IFT27 promotes BBSome exit from cilia through the GTPase ARL6/BBS3. Developmental Cell. 31 (3), 265-278 (2014).
- Engle, S. E., et al. A CreER Mouse to Study Melanin Concentrating Hormone Signaling in the Developing Brain. Genesis. , (2018).
- Thorpe, S. D., et al. Reduced primary cilia length and altered Arl13b expression are associated with deregulated chondrocyte Hedgehog signaling in alkaptonuria. Journal of Cellular Physiology. 232 (9), 2407-2417 (2017).
- Lauring, M. C., et al. New software for automated cilia detection in cells (ACDC). Cilia. 8, 1 (2019).
- Dummer, A., Poelma, C., DeRuiter, M. C., Goumans, M. J., Hierck, B. P. Measuring the primary cilium length: improved method for unbiased high-throughput analysis. Cilia. 5, 7 (2016).
- Haycraft, C. J., et al. Gli2 and Gli3 localize to cilia and require the intraflagellar transport protein polaris for processing and function. PLoS Genetics. 1 (4), 53 (2005).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены