
Method Article
בדיקת תכונות מבניות ודינמיות של ננו-מבנים תת-תאיים של סחר בבני אדם על-ידי ספקטרוסקופיית תנודות מרחביות-טמפורליות
In This Article
Summary
ניתוח תזוזה ריבועית ממוצעת (iMSD) שמקורו בהדמיה מיושם על מקרופינוזומים כדי להדגיש את האופי הפנימי שלהם המתפתח בזמן במונחים של תכונות מבניות ודינמיות. לאחר מכן משווים את המקרופינוזומים לגרגירי הפרשת האינסולין (ISGs) כהתייחסות למבנים תת-תאיים בעלי תכונות מבניות/דינמיות ממוצעות של אינווריאנטים בזמן.
Abstract
תזוזה ריבועית ממוצעת שמקורה בהדמיה (iMSD) משמשת לטיפול בתכונות המבניות והדינמיות של ננו-מבנים תת-תאיים, כגון שלפוחיות המעורבות בסחר אנדו/אקסוציטוטי של מומסים וביו-מולקולות. iMSD מסתמך על הדמיה סטנדרטית של קיטועי זמן, תואם לכל הגדרה אופטית ואינו צריך להתעכב על עצמים בודדים כדי לחלץ מסלולים. מכל עקבות iMSD מחושבת ומשולבת שלישייה ייחודית של פרמטרים מבניים ודינמיים ממוצעים (כלומר, גודל, פיזור מקומי, מקדם חריג) כדי לבנות את "חתימת ה-iMSD" של הננו-מבנה הנחקר.
עוצמתה של גישה זו מוכחת כאן במקרה המופתי של מקרופינוזומים. בועיות אלה מתפתחות עם הזמן, ומשנות את הגודל, המספר והתכונות הדינמיות הממוצעות שלהן ועוברות משלבים מוקדמים למאוחרים של סחר תוך-תאי. כבקרה, גרגירי הפרשת אינסולין (ISGs) משמשים כהתייחסות למבנים תת-תאיים שחיים במצב נייח שבו התכונות המבניות והדינמיות הממוצעות של כל אוכלוסיית האובייקטים הן אינווריאנטיות בזמן. ניתוח iMSD מדגיש את התכונות המוזרות הללו באופן כמותי וסולל את הדרך ליישומים דומים ברמה התת-תאית, הן במצבים הפיזיולוגיים והן במצבים הפתולוגיים.
Introduction
ננו-מבנים תת-תאיים (למשל, בועיות אנדוציטיות/הפרשתיות, אברונים) ממלאים תפקיד מרכזי בוויסות איתות התא1. כוונון נכון של המאפיינים המבניים (למשל, הגודל) ו/או הדינמיים שלהם (למשל, פיזור) קובע כיצד התא מגיב לגירויים פנימיים או חיצוניים 2,3,4. בהתבסס על ראיות אלה, אין זה מפתיע כי שינויים של מאפיינים אלה נמצאים במצבים פתולוגיים רבים. דוגמאות לכך כוללות את תפקידה של אנדוציטוזה מוטעית בסרטן 2,3, את השינויים המבניים והדינמיים שנמצאו ברמה של ISGs בתאי β שנחשפו למצבי סוכרת מסוג 25, את הוויסות השגוי של תכונות מבניות ותחבורה ליזוזומיות בלויקודיסטרופיה של תאים כדוריים או גלקטוזילצרמיד ליפידוזיס6, וחוסר תפקוד במסלול האנדו-ליזוזומי בהפרעות נוירודגנרטיביות (למשל, מחלת אלצהיימר)7.
בהקשר זה, חוקרים הוכיחו לאחרונה כי ניתן לשפר את הביצועים של שיטות מיקרוסקופיה אופטית סטנדרטיות על ידי כוונון נכון של רזולוציה8 של דגימה מרחבית וטמפורלית. זה, בתורו, עשוי לספק תובנה נוספת על תהליכים ביולוגיים של רלוונטיות. בפועל, הדבר מתאפשר על ידי אלגוריתם של ניתוח תנודות מרחביות-טמפורליות, אשר מחלץ בו זמנית את התכונות המבניות והדינמיות הממוצעות של פיזור עצמים ישירות מתוך הערימה הסטנדרטית של תמונות מיקרוסקופיה אופטיות ללא כל צורך בידע ראשוני על האובייקט הביולוגי המעניין והוצאת מסלולים של אובייקט יחיד. כל המידע הזה מצורף בפלט יחיד של השיטה: מעקב iMSD9 (פרטים על הנגזרת והניתוח של עקבות iMSD ניתנים בקובץ משלים 1).
פרוטוקול הניסוי שנוצר מורכב מכמה שלבים. ראשית, הדמיה של אזור העניין מתבצעת ברזולוציה טמפורלית גבוהה. לאחר מכן, פונקציות מתאם מרחביות-טמפורליות ממוצעות מחושבות מתוך ערימת התמונות. לבסוף, על ידי התאמת גאוס של סדרת פונקציות המתאם, 'חוק הדיפוזיה' הממוצע מתקבל ישירות מהדמיה ומנותח כדי לזהות את מצב הדיפוזיה האובייקטית. הפוטנציאל של השיטה כבר הוכח עבור מגוון עצמים ביולוגיים, החל ממולקולות ועד ננו-חלקיקים ואפילו אברונים/מבנים תת-תאיים שלמים 9,10,11,12,13,14,15.
מאמר זה מדווח על יישום iMSD למקרופינוזומים כדי להדגיש את טבעם הפנימי והבלתי הפיך המתפתח בזמן במונחים של תכונותיהם המבניות והדינמיות הממוצעות (כלומר, ברמת האוכלוסייה כולה). יתר על כן, בועיות אנדוציטיות אלה מושוות ל-ISGs כהתייחסות למבנים תת-תאיים ב'מצב נייח', כלומר מצב שבו התכונות המבניות/דינמיות הממוצעות של כל אוכלוסיית הגרגירים נשארות קבועות בכל נקודת זמן. מקרופינוציטוזה מגדירה סדרה של אירועים שיזמו על ידי ארגון מחדש נרחב (או ראפלינג) של קרום הפלזמה ליצירת מבנה מקרופינוציטי חיצוני המופנם לאחר מכן16. המקרופינוזומים שנוצרו בשלב מוקדם דומים מאוד לפאגוזומים. יחד עם זאת, ניתן להבחין ביניהם לבין צורות אחרות של שלפוחיות אנדוציטיות בשל גודלן הגדול האופייני, ההטרוגניות המורפולוגית והיעדר מבנים של ציפוי חלבון.
בדיקות ביוכימיות גילו כי עם ההפנמה, המקרופינוזומים מתעשרים בהדרגה בסמני חלבונים של מסלולים אנדוציטיים אחרים, בתורם, מה שמרמז על כך שזהותם משתנה ללא הרף במהלך סחר בבניאדם 17. באמצעות שימוש בנוגדנים נגד סמנים ידועים של המסלול האנדוסומי, הוכח כי מקרופינוזומים מאמצים בהדרגה תכונות אנדוזומליות קלאסיות: הם פוחתים בגודלם, מתפתחים למבנים אנדוציטיים מאוחרים (למשל, ליזוזומים), או בסופו של דבר מאבדים את זהותם באמצעות שליפה בתיווך ממברנה של סמנים מולקולריים ספציפיים (למשל, מיון נקסינים)18,19 . התרחיש הכולל הוא שכל מקרופינוזום בודד בתוך התא משנה באופן בלתי הפיך את זהותו המבנית והדינמית (כמו גם המולקולרית) במהלך הסחר מממברנת הפלזמה לגורלו התוך-תאי הסופי. כתוצאה מכך, התכונות המבניות/דינמיות/מולקולריות של כל אוכלוסיית המקרופינוזומים משתנות גם הן באותו נתיב זמני. בהיותה רגישה במהותה לתכונות הממוצעות של כל אוכלוסיית העצמים הנצפים, שיטת iMSD מתארת באופן כמותי את 'האופי המתפתח' על ידי כימות של פרמטרים ממוצעים מרכזיים, כלומר, הדפיזור המקומי והמקדם החריג (תכונות דינמיות) והגודל הממוצע של המקרופינוזומים (תכונה מבנית) בכל שלב של הסחר התוך-תאי שלהם.
לשם השוואה, מדידות דומות בוצעו על מבנה ידוע של ממברנה תוך-תאית סגורה, ה-ISG, במודל של תאי β. בדומה למקרופינוזומים, ויסות התכונות המבניות והדינמיות של ISGs, החל מראשיתם ברשת Trans Golgi (TGN) ועד לאקסוציטוזה שלהם בקרום הפלזמה, הוא מרכזי לביצוע נכון של פונקציית ISG20. עם זאת, בניגוד למקרופינוזומים, ISGs חיים ב'מצב נייח' שבו, בכל עת, כל השלבים התפקודיים/מבניים/מולקולריים של תוחלת החיים של ISG נמצאים בו-זמנית בתוך התא, וכל אחד מהם מיוצג על-ידי תת-אוכלוסייה ספציפית של ISGs. משמעות הדבר היא שלמרות שכל גרגיר מתפתח באופן בלתי הפיך מביוגנזה להפרשה, התכונות המבניות/דינמיות הממוצעות של כל אוכלוסיית הגרגירים צפויות להישאר קבועות בכל נקודת זמן (אלא אם כן תנאי המצב הנייח משתנים, למשל, על ידי גירויים חיצוניים כגון גלוקוז, כולסטרול וציטוקינים13). זה אושר על ידי ניתוח iMSD.
Protocol
1. הכנה לדוגמה
- לפני ניסוי המיקרוסקופיה, תאים תת-תרבותיים במנות התאימו ליישומי מיקרוסקופיה.
- שטפו צלחת בגודל 10 ס"מ שטופלה בתרבית רקמה של תאי HeLa או INS 1E (אינסולינומה β דמויי תאים) באורך 10 ס"מ עם 0.01 M 1x PBS, הוסיפו 1 מ"ל של 0.05% טריפסין-EDTA (1x), והניחו אותה באינקובטור של 37 מעלות צלזיוס, לחות, 5% CO2 למשך 5 דקות.
- בצעו שימוש חוזר בתאים המנותקים על ידי הוספת 9 מ"ל של מדיום DMEM שלם (עבור תאי HeLa) או RPMI 1640 (עבור תאי INS-1E), ואסוף את 10 מ"ל האחרונים בצינורות צנטריפוגה.
- זרעים בערך 2 × 105 תאים בכל צלחת 35 מ"מ x 10 מ"מ בנפח סופי של 1 מ"ל של המדיום. הדגירה של התאים למשך 24 שעות בטמפרטורה של 37 מעלות צלזיוס ו-5% CO2.
- כדי לתייג באופן פלואורסצנטי ליזוזומים, השתמש ב- LysoTracker Red DND-99.
- דיללו את תמיסת המניות ב-1 מ"ל של בינוני מוכן מראש לריכוז צבע סופי של 70 ננומטר.
- החליפו את המדיום מהמנה במדיום טרי המכיל LysoTracker. דגירו את התאים במדיום המכיל LysoTracker למשך 20 דקות בטמפרטורה של 37 מעלות צלזיוס באטמוספרה של 5% CO2 , ושטפו אותם פעמיים במדיום טרי לפני הניסוי.
- כדי לתייג באופן פלואורסצנטי מקרופינוזומים, השתמש ב-70-kDa פלואורסציין איזותיוציאנט-דקסטרן. לשטוף את התאים התת-תרבותיים שלוש פעמים עם 0.01 M 1x 1x מלוחים ביציאת פוספט (PBS), להחליף עם תווך המכיל dextran (1 מ"ג / מ"ל), ולדגום ב 37 °C (37 °C) במשך 30 דקות. לפני שתמשיכו בניסוי המיקרוסקופיה, שטפו את התאים שלוש פעמים במדיום טרי.
- כדי לתייג באופן פלואורסצנטי ISGs בתאי INS-1E, לבצע טרנספקציה של התאים באמצעות ריאגנט טרנספקציה (ראו טבלת החומרים) וחלבון פלואורסצנטי ירוק משופר C-פפטיד (EGFP) פלסמיד13 על פי פרוטוקול היצרן, ולדגום בטמפרטורה של 37 מעלות צלזיוס למשך 24 שעות באטמוספרה של 5% CO2 לפני הניסוי.
2. רכישת נתונים
- כדי לאפשר למיקרוסקופ להשתוות בטמפרטורה ובאטמוספרה הרצויות, הפעילו את מערכת בקרת המיקרוסקופים לפחות שעתיים לפני הניסוי.
הערה: כל רכישה היא סדרה בהילוך מהיר. - קבל את התמונות באמצעות מיקרוסקופ קונפוקלי הפוך המצויד במטרת טבילה במים בגודל 60x, 1.2 צמצם מספרי (NA).
- השתמש בלייזר ארגון של 488 ננומטר לעירור של EGFP (תאים שעברו טרנספקטציה) ומקרופינוזומים המסומנים בפלואורסצין. אסוף את הפליטה הפלואורסצנטית בין 500 ל-600 ננומטר באמצעות גלאי צינורות פוטומולטיפלייר סטנדרטי.
- השתמש בלייזר HeNe של 543 ננומטר כדי לעורר את Lysotracker ולאסוף את פליטת הפלואורסצנציה שלו בין 555 ל-655 ננומטר.
- הגדר את קוטר חור הפין לגילוי לגודל של 1 אוורירי. עבור כל רכישה, לאסוף סדרה של 1000 מסגרות עוקבות. הגדר את זמן השהייה של הפיקסלים ל- 2 μs/pixel עבור זמן מסגרת של 129 אלפיות השנייה.
הערה: כל מסגרת כללה 256 x 256 פיקסלים (16 סיביות/פיקסלים) עם ממד פיזי של 69 ננומטר/פיקסל, המתאים בערך לשטח של 17 מיקרומטר x 17 מיקרומטר.
3. חישוב IMSD
הערה: כדי לבצע את החישוב כראוי, השתמש בתוכנה המסוגלת לחישוב מספרי ותכנות סקריפט. הסקריפט הספציפי (כלומר, עבור קובץ הסקריפט 'iMSD.m', ראה קובץ תומך 1) חייב להיות נוכח באותה ספרייה המכילה את סדרת התמונות לעיבוד. כל תמונה של הסדרה חייבת להישמר כקובץ '.tif' מובחן.
- כדי לאתחל כראוי את הפרמטרים האינסטרומנטליים המשמשים לרכישות, פתח את iMSD.m עם עורך הטקסט של התוכנה ושנה את החלק הראשון שלו באופן הבא.
- הגדר את N כמספר הפריימים בסדרת הזמן (לדוגמה, 1000 בפרוטוקול זה).
- הגדר px_size: גודל פיקסל, המבוטא ב- μm (לדוגמה, 0,069 בפרוטוקול זה).
- סט f: רזולוציה טמפורלית של כל מסגרת, המבוטאת בשניות (לדוגמה, 0,129 בפרוטוקול זה).
- הגדר מסנן: קלט בינארי לתיקון רקע, הגדר את הערך ל- '0' כדי לעבד תמונות גולמיות, או הגדר את הערך ל- '1' כדי לבצע חיסור רקע מבוסס סף.
- הגדר av_toll: סף לתיקון רקע; כל פיקסל בעוצמה נמוכה מערך זה יוגדר כ- 0 אם Filter=1.
- הגדר סיבית כמספר השלם הקובע את דגימת העוצמה (לדוגמה, 8 סיביות, 16 סיביות).
- שמור והפעל את קובץ הסקריפט iMSD.m הערוך.
- בדוק את ביצוע הסקריפט.
הערה: ניתן לבדוק את מצב עיבוד החישוב בחלון הפקודה; אם מתרחשת בעיה קטלנית כלשהי, התהליך יופרע, ותוצג הודעת אזהרה כדי להציג את סוג השגיאה ואת קוד השורה הקשורה. אחרת, בצע את השלבים 3.3.1-3.3.3.- יבא את ערימת התמונות וחיסר את הרקע (במידת הצורך).
- חישוב פונקציית המתאם הספטיוטמפוראלי G(ξ,η,τ) בשיטת פורייה.
- התאימו את פונקציית המתאם הספטיוטמפוראלי עם פונקציית גאוס דו-ממדית.
הערה: בסוף התהליך, ערכי הפלט של נהלי ההתאמה של σ2(τ) יוצגו בחלון הפקודה: ממוצעים, שגיאות וטובות מתאימות (R2) של התאמה מדווחים.
- בדוק את הפלט הגרפי.
הערה: עקומת iMSD ועקומות ההתאמה המתאימות מוצגות בשלושה לוחות נפרדים, כל אחד עבור סוג אחר של משוואה מתאימה המשמשת: דיפוזיה בראונית, דיפוזיה חריגה או דיפוזיה מוגבלת. עבור כל עקומה, הערך R2 מדווח במקרא התרשים. - בדוק את פלט הטקסט.
הערה: בסוף התהליך, נוצר קובץ '.xls' עם אותו שם של קובץ '.tif' המקורי עם חלוף הזמן. הגיליון הראשון מכיל את הערכים של ξ, η, τ ו- σ2 המחושבים עבור כל השהיית זמן. פרמטרי הקלט וערכי הפלט המחושבים העיקריים מדווחים בגיליון השני, כלומר, מקדם דיפוזיה, מקדם חריג וציר y σ20 יירוט.
תוצאות
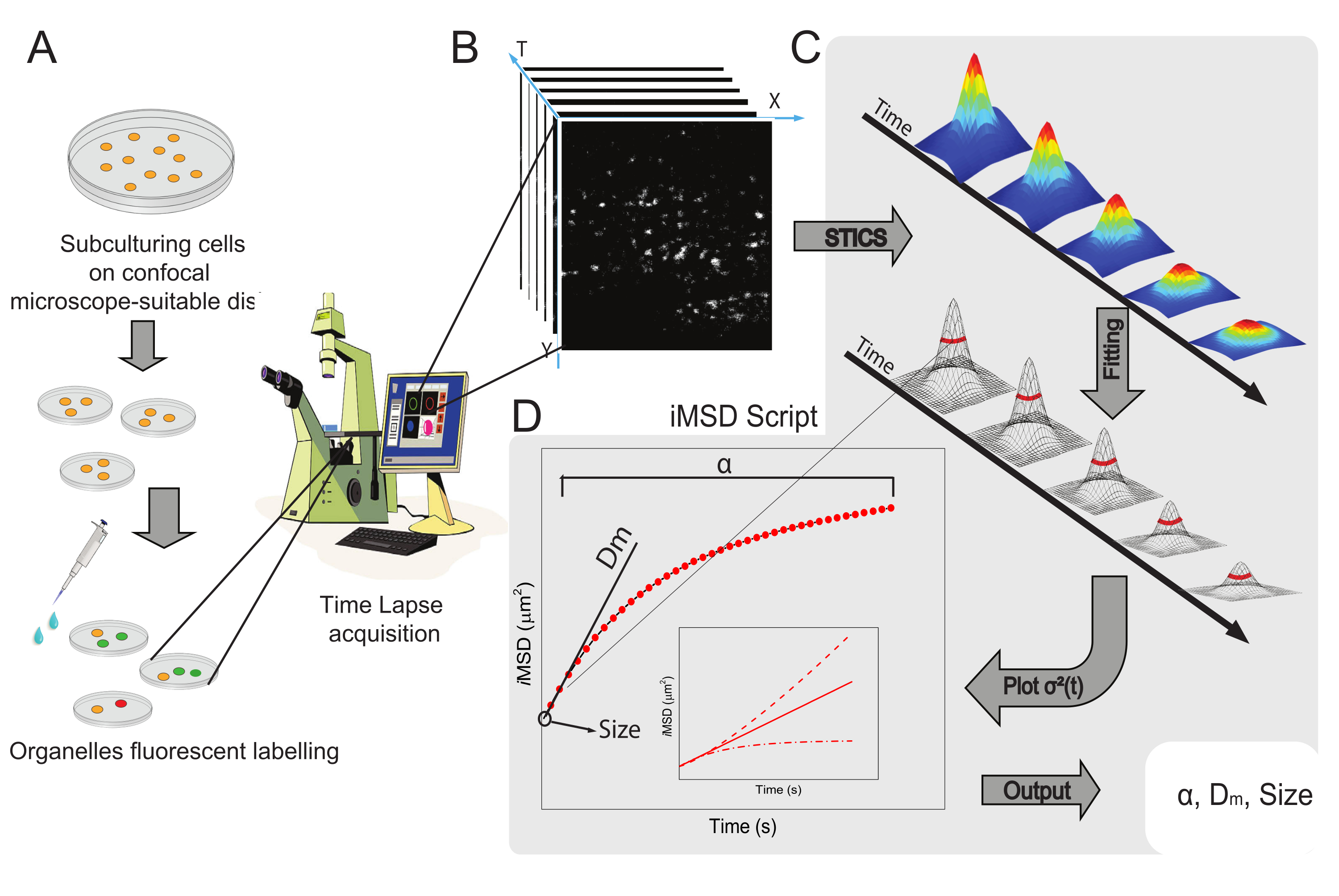
זרימת העבודה הכללית של השיטה מוצגת באיור 1. הוא משחזר את השלבים העיקריים המוצגים בסעיף הפרוטוקול, מהכנת דגימה (איור 1A) ועד הדמיה של ננו-מבנים תוך-תאיים (איור 1B), ניתוח תנודות לחישוב סדרת פונקציות המתאם הספטיוטמפוראלי (איור 1C), והתאמה לגזירת התכונות המבניות/דינמיות הממוצעות של האובייקט הנחקר (איור 1D).
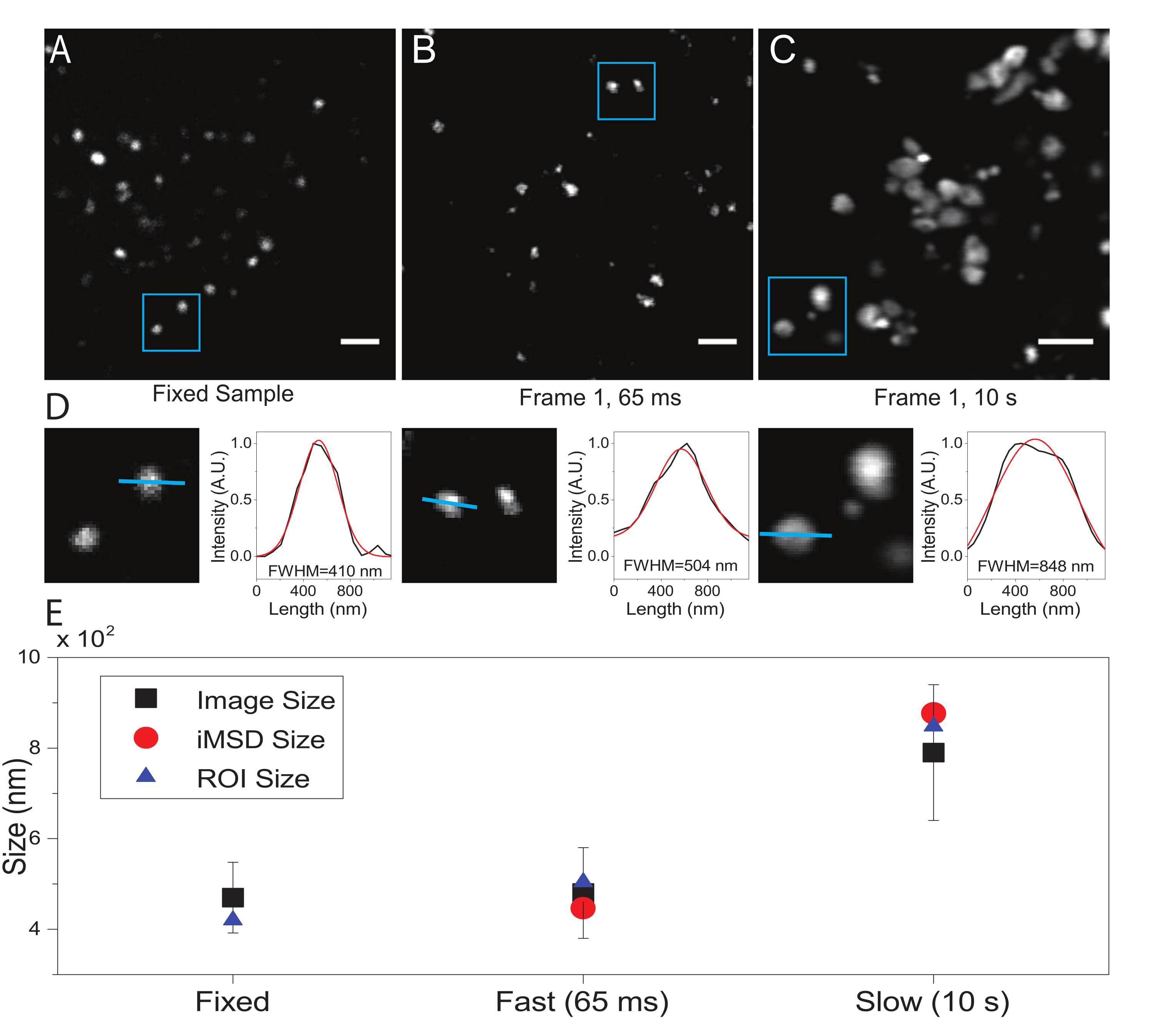
פרמטר קריטי הוא רזולוציית הזמן שאומצה להדמיית האובייקט התת-תאי המעניין. ערך ניסיוני זה יקבע את סף הזמן שבו תימדד התזוזה הממוצעת המינימלית של האובייקטים המעניינים. עם זאת, התנאי המועדף הוא קביעת רזולוציית זמן של הדמיה שבה מושא העניין נראה 'חסר תנועה' בתוך המסגרת שנתפסה, כלומר, הוא מציג גודל אופייני שבממוצע אינו מעוות בשל מהירות ההדמיה. זה אפשרי מבחינה טכנית אם מושא העניין הוא מבנה תת-תאי סגור קרום או אברון (כמו במקרה זה). בדרך כלל, מבנים תת-תאיים מציגים מקדמי דיפוזיה מקומיים (D, μm2/s, ראו טבלה 1) שהם נמוכים בכמה סדרי גודל מאלו של מולקולות בודדות מבודדות בציטופלסמה (למשל, GFP21). אימות יכול להתבצע על ידי שיתוק מלאכותי של אברון העניין (למשל, על ידי קיבוע כימי). ואכן, מצב זה יכול לשמש כהתייחסות לקביעת גודל האורגן בפועל בתנאי הניסוי שבהם נעשה שימוש (למשל, אורך גל של עירור, גודל פיקסל, מטרה).
כאן, למרות שליזוזומים שימשו כאברוני בדיקה להליך זה, התוצאות תקפות באופן עצמאי ממבנה המטרה. איור 2A מראה תמונה של ליזוזום קבוע (כלומר, חסר תנועה) יחד עם רכישה המבוצעת על תאים חיים ברזולוציה הזמנית המתאימה (כלומר, בדרך כלל מתחת ל-100 אלפיות השנייה/מסגרת; למשל, 65 אלפיות השנייה/מסגרת בדוגמה באיור 2B) ורכישה המבוצעת במכוון ברזולוציה טמפורלית נמוכה מאוד (למשל, 10 s/frame בדוגמה באיור 2C ). עבור כל תנאי, גודלו של האובייקט המתפזר מופק באופן הבא: 1) פרופיל עוצמה של הנקודה נגזר על ידי כלי הקו בתוכנת ImageJ; ii) פרופיל העוצמה משורטט ועושה אינטרפולציה על ידי פונקציית גאוס כדי לחשב את הרוחב המלא בחצי ערך מקסימלי (FWHM) שבתורו משמש כאומדן של קוטר הנקודה (איור 2D). כצפוי ומוצג בחלקה של איור 2E, הרכישה ברזולוציה טמפורלית גבוהה מאוד (כלומר, 65 אלפיות השנייה למסגרת) מניבה גודל ממוצע של המבנה קרוב לזה שהתקבל במדגם הקבוע, בין אם על ידי שימוש בכלי הסטנדרטי שתואר לעיל או על ידי חילוץ היירוט iMSD y-axis. במקום זאת, הרכישה במהירות איטית מניבה עלייה בגודלו הנראה לעין של המבנה, בשל הדינמיקה של המבנה הטבעי במהלך ההדמיה. בתנאים ניסיוניים לא אופטימליים, המידע המבני/דינמי שחולץ אינו משקף בנאמנות את התכונות הפנימיות של האובייקט הנחקר.
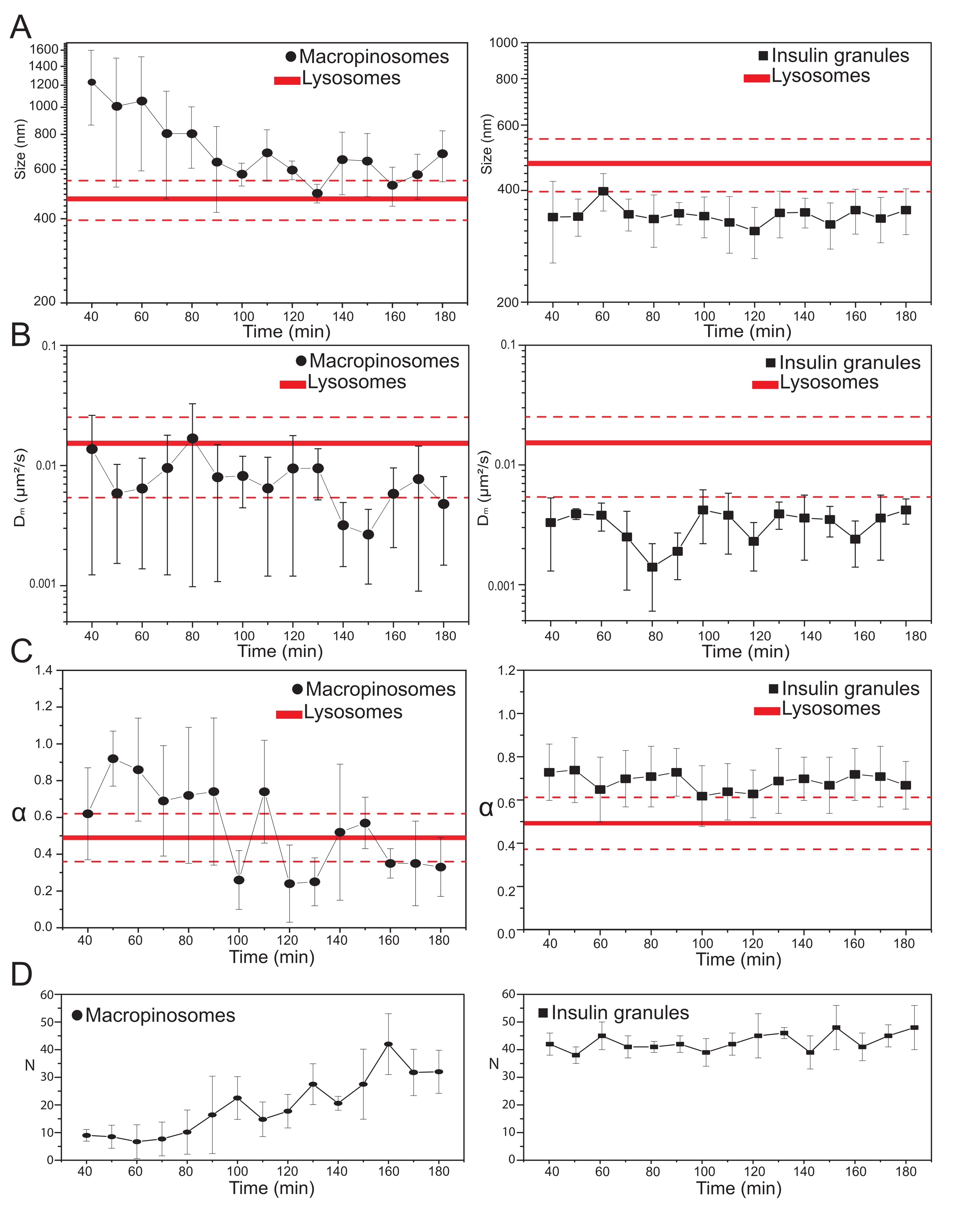
לאחר בחירת הפרמטרים הניסוייים העיקריים, ניתן לייצר מערכי נתונים עבור המבנים התוך-תאיים של המטרה. עבור מקרופינוזומים, לאחר 20 דקות של דגירה של התאים עם 70 kDa dextrans, סדרות זמן של המבנים התוך תאיים המסומנים נרכשו בנקודות זמן שונות לאחר הטיפול, מ 30 דקות עד כ 180 דקות. באופן מעניין, שינוי הדרגתי בתכונות המבניות והדינמיות של מקרופינוזומים מזוהה במהלך סחר בבני אדם (איור 3; התפלגויות של σ02, Dm, α ו-N עבור מקרופינוזומים מדווחות בחלקות משמאל). בעוד שלא זוהו שינויים ברורים בפיזור המקומי (Dm) של מקרופינוזומים במהלך הסחר, הן הגודל האופייני (σ0 2) והן אופן התנועה הכולל (α) מתפתחים בזמן.
שימו לב במיוחד, ירידה בגודל הממוצע של המקרופינוזומים נצפית במהלך סחר בבני אדם (איור 3A, משמאל), יחד עם עלייה מקבילה באופי התת-פיזורי של תנועתם (כלומר, מסומן כירידה בערכי α, איור 3C, לוח שמאלי). בנוסף, מספר המקרופינוזומים הוצא מכל רכישה: התוצאות, שדווחו באיור 3D, בלוח השמאלי, חושפות בבירור עלייה במספר המקרופינוזומים בזמן. כל התוצאות הללו תואמות היטב את הציפיות מכיוון שמקרופינוזומים המסומנים בדקסטרן אמורים להיווצר כשלפוחית מבודדת, סגורה בממברנה הגדולה, המוקפת בממברנת הפלזמה (שגם הם מוכשרים לתנועה לאורך רכיבי השלד הציטו-שלדיים) אך אמורים לתקשר בהדרגה עם המסלול האנדו-ליזוזומי המורכב מאוכלוסייה גדולה של מבנים קטנים יותר ומפזרים באופן אקראי.
כצפוי לעיל, התוצאות עבור מקרופינוזומים מנוגדות למדידות דומות שבוצעו ב-ISGs (איור 3, עמודה ימנית). גרגירי אינסולין אינם מראים מגמה מתפתחת בזמן של הפרמטרים המבניים/דינמיים שמקורם ב-iMSD (והמספר הממוצע שלהם בתוך התא) באותו חלון זמן שנצפה עבור מקרופינוזומים. יתר על כן, הערכים האופייניים של σ02, Dm ו- α שונים למדי מאלה של ליזוזומים, המשמשים שוב כהפניה. תוצאה זו מאששת את הרעיון, כצפוי לעיל, שגרגירים נבדקים ב'מצב נייח' שבו, בכל עת, התכונות המבניות/דינמיות הממוצעות של כל אוכלוסיית ה-ISGs אינן משתנות (כלומר, הן נשארות קבועות, אלא אם כן תנאי המצב הנייח משתנים, למשל, עקב גירויים חיצוניים).

איור 1: זרימת עבודה ניסיונית. (A) התאים צופו 24 שעות (48 שעות לניסויים בטרנספקציה) לפני ניסויים קונפוקליים על צלחות תאים המתאימות ליישומים מיקרוסקופיים. לאחר מכן טופלו התאים בהתאם לשיטת הסימון (ראו פרוטוקול) כדי להכתים את האברון הציטופלסמי בעל העניין. (B) רכישה קונפוקלית טיפוסית מורכבת מערימה של תמונות (קיטועי זמן) של חלק ציטופלסמי של תא חי, המתארות את התפתחות הזמן של דינמיקת אברונים מסומנים. (C) סרט קיטועי זמן מנותח באמצעות סקריפט Matlab מותאם אישית, תחילה חישוב פונקציית המתאם של תמונה מרחבית-טמפורלית והתאמה של גאוס להתוויית עקומות iMSD (D) ופרמטרים נלווים של התאמה שחולצו המתארים פרמטרים של דינמיקה מבנית של אברונים מדומים. קיצורים: iMSD = הדמיה נגזרת תזוזה ריבועית ממוצעת; STICS = ספקטרוסקופיית מתאם תמונה מרחבית-טמפורלית; α = מקדם דיפוזיה חריג; Dm = פיזור מקומי; σ2(τ) = שונות. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}

איור 2: פרמטרים ניסיוניים נכונים. (A) תמונה מופתית של ליזוזומים מוכתמים בדגימה קבועה. סרגל קנה מידה = 2 μm. (B) המסגרת הראשונה של ערימת תמונות של ליזוזומים מוכתמים בתא חי, שנרכשה בפרמטרים המתאימים. רזולוציה טמפורלית: 65 אלפיות השנייה למסגרת. סרגל קנה מידה = 2 μm. (C) המסגרת הראשונה של ערימת תמונות של ליזוזומים מוכתמים בתא חי, שנרכשו במהירות נמוכה: דפורמציה חרצית של גודל הליזוזומים לכאורה עקב תנועה אברונית במהלך ההדמיה נראית לעין. רזולוציה זמנית: 10 שניות למסגרת. סרגל קנה מידה = 2 μm. (D) דוגמה לחישוב גודל עבור ליזוזומים שצולמו בהחזר השקעה כחול של (A), (B) ו- (C). פרופיל העוצמה לאורך הקו הכחול צויד בפונקציה של גאוס כדי לאחזר את ה-FWHM, כלומר להעריך את גודל הנקודה. ערכי FWHM מדווחים עבור כל התאמה. (E) ייצוג גרפי של ערכי גודל המתקבלים על ידי ניתוח תמונה המתואר בפאנל (D) עבור כל הליזוזומים המצולמים (ריבוע שחור, ערך ממוצע וסטיית תקן), עבור ליזוזומים הסגורים בתוך החזר ההשקעה הכחול (משולש כחול), ומאוחזרים על ידי ניתוח iMSD (עיגול אדום). נתון זה הוא מתוך 22. קיצורים: ROI = אזור עניין; FWHM = רוחב מלא בחצי מקסימום; iMSD = תזוזה ריבועית ממוצעת שמקורה בהדמיה. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}

איור 3: אבולוציה טמפורלית של אברונים מסומנים. (A) חלקות של ערכי גודל שחולצו על ידי iMSD לעומת התקדמות בזמן של רכישות עוקבות המיוצגות כממוצע של ערכים מדודים ברכישות שבוצעו בחלון זמן של 10 דקות. משמאל, הפחתה הדרגתית בגודל הממוצע של המקרופינוזומים (עיגולים שחורים) ומימין, הגודל אינווריאנטי בזמן של גרגירי הפרשת אינסולין (ריבועים שחורים) בהשוואה לליסוזומים, המיוצגים כערך גודל ממוצע (קו אדום עבה יותר) ± סטיית תקן (קווים אדומים מקווקווים). (B) ו-(C) התקדמות בזמן של מקדמי Dm ו-α עבור מקרופינוזומים (משמאל) וגרגרי אינסולין (מימין) המופקים על ידי ניתוח iMSD. (D) התפתחות-זמן של מספרים של מקרופינוזומים מסומנים וגרגרי אינסולין שנמדדו בפריים הראשון של כל סרט קיטועי זמן נרכשים. קיצורים: iMSD = הדמיה נגזרת תזוזה ריבועית ממוצעת; α = מקדם דיפוזיה חריג; Dm = פיזור מקומי, N = מספר. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
{kind=link}
| אורגנל | תיוג | קו סלולרי | גודל (nm) | Dm ( × 10-3 מיקרומטר2/s) | α | N | ת.ז. | ||
| אנדוזום מוקדם (EE) | CellLight מוקדם אנדוזום GFP | הלה | 395±74 | 3.0±2.4 | 1.02±0.20 | 40 | 10 | ||
| אנדוזום מאוחר (LE) | CellLight מאוחר אנדוזום GFP | הלה | 693±102 | 15.4±10.6 | 0.57±0.16 | 58 | 10 | ||
| ליזוזום (LY) | LysoTracker DND-99 | הלה | 471±76 | 15.3±9.0 | 0.49±0.13 | 143 | 10, 14 | ||
| קייבולה (CAV) | קאוולין-EGFP | הלה | 405±49 | 3.1±1.8 | 1.00±0.22 | 15 | 10 | ||
| שלפוחית מצופה קלתרין (CCV) | טרנספרין-אלכסה 488 | הלה | 513±62 | 16.2±9.9 | 0.48±0.17 | 33 | 10 | ||
| גרגיר אינסולין (IG) | C-פפטיד-EGFP | INS-1E | 335±56 | 3.0±1.7 | 0.70±0.14 | 107 | 11 | ||
| מקרופינוסום מוקדם (EMCR) | Fluorescein-Dextran 70 kDa | הלה | 979±423 | 8.3±9 | 0.79±0.27 | 36 | 10 | ||
| מקרופינוזום ביניים (IMCR) | Fluorescein-Dextran 70 kDa | הלה | 702±180 | 13.7±19.9 | 0.60±0.38 | 29 | 10 | ||
| מקרופינוזום מאוחר (LMCR) | Fluorescein-Dextran 70 kDa | הלה | 592±127 | 5.8±4.7 | 0.39±0.21 | 21 | 10 | ||
טבלה 1: פרמטרים מבניים ודינמיים שחולצועל-ידי I MSD. הטבלה מציגה את ערכי הגודל, Dm ומקדם α שנמדדו עבור אברונים שונים, תוך ציון אסטרטגיות תיוג, קו התא שבו נעשה שימוש ומספר הרכישות שנותחו. הערכים מדווחים כסטיית תקן ממוצעת ±. קיצורים: iMSD = הדמיה נגזרת תזוזה ריבועית ממוצעת; α = מקדם דיפוזיה חריג; Dm = פיזור מקומי, N = מספר; GFP = חלבון פלואורסצנטי ירוק; EGFP = חלבון פלואורסצנטי ירוק משופר.
קובץ משלים 1: פרטים על הנגזרת והניתוח של מעקב iMSD. אנא לחץ כאן כדי להוריד קובץ זה.
קובץ תומך 1: אנא לחץ כאן כדי להוריד קובץ זה.
Discussion
המאפיינים והיתרונות של iMSD ניכרים בהשוואה לטכניקות הזמינות לאחזר מידע אנלוגי. עבור מידע מבני, הבחירה המועדפת היא ניתוח מיקרוסקופ אלקטרונים (TEM) של הולכה. בשיטה זו ניתן לאחזר פרטים אולטרה-סטרוקטורליים ברזולוציה מולקולרית ואף מעבר לכך, אפילו עבור ננו-מבנים תת-תאיים. עם זאת, הרזולוציה המרחבית המוזרה של TEM מושגת על חשבון המידע בממד הזמני, שמעניין כאן. כדי לפצות על כך, ההתקדמות האחרונה בטכנולוגיות הדמיה של תאים חיים מעניינת במיוחד. אלה כוללים סמנים פלואורסצנטיים חדשים עם ביצועים משופרים (למשל, בהירות ויציבות צילום), הליכי תיוג ממוטבים וגלאים רגישים יותר. בנוסף, כלים אנליטיים זמינים כדי לטפל הן במבנה (למשל, 'גודל' על ידי ניתוח מבוסס phasor של ספקטרוסקופיה מקומית של מתאם תמונה, PLICS23, צבירה/אוליגומריזציה על ידי ניתוח מספר ובהירות24) והן דינמיים (למשל, חוק דיפוזיה על ידי מעקב אחר חלקיקים בודדים, כלומר (SPT)25,26,27,28 ) פרמטרים בסולם התת-תאי. שיטת SPT מאפשרת גישה ישירה למסלול האובייקט ול-MSD שלו. עם זאת, החיסרון הוא הצורך בצפיפות נמוכה של הגשושית ותוויות בהירות מאוד ומסלולים רבים של עצם יחיד שיש למדוד כדי לעמוד בקריטריונים סטטיסטיים. ביחס לרזולוציה הטמפורלית של המדידה, גשושיות אי-אורגניות, הניתנות לצילום (למשל, נקודות קוונטיות או ננו-חלקיקי מתכת) יכולות לשפר את ביצועי ה-SPT אך על חשבון הליכי ייצור ותיוג מורכבים.
בהשוואה לתקנים אלה, שיטת iMSD המתוארת כאן מציגה כמה יתרונות מרכזיים. ראשית, ניתן להשתמש בגישה זו בשילוב עם תגים פלואורסצנטיים עמומים יחסית, כגון חלבונים פלואורסצנטיים המקודדים גנטית (למשל, היישום על ISGs). לפיכך, בהשוואה ל- SPT, מושגת רזולוציה טמפורלית גבוהה יותר (באמצעות אותה תווית) בשל הכמות הנמוכה יותר של פוטונים הנדרשת8. שנית, שיטת iMSD מוגבלת רק על ידי הרזולוציה הזמנית אך לא עקיפה. למעשה, למרות ההתקנה האופטית המוגבלת של עקיפה, ניתן למדוד תזוזות מולקולריות ממוצעות אפילו מתחת לגבול הדיפרקציה, כפי שכבר הודגם עבור זרימות מולקולריות באמצעות STICS29. הרזולוציה בפועל במדידת התזוזות תלויה במידת הדיוק (במונחים של אות לרעש) שניתן למדוד את פונקציית המתאם, ובכך להסביר מדוע היא אינה מוגבלת על ידי עקיפה. לפיכך, נראה ברור כי התזוזה המינימלית שניתן למדוד תלויה בפיזור של מושא העניין וברזולוציה הזמנית של מערך ההדמיה.
בהקשר זה, חשוב לקחת בחשבון כי היישום על ננו-מבנים תת-תאיים, כגון מקרופינוזומים או גרגירי אינסולין, עם מיקרוסקופ סריקת לייזר הוא אופטימלי: מהירות הסריקה הזמינה גבוהה משמעותית מהדינמיקה של מושא העניין. במקרה כזה, תנועת האובייקטים במהלך הרכישה היא זניחה, ופונקציית המתאם יכולה להיות משוערת על ידי פונקציית גאוס. לבסוף, ניתן ליישם בקלות את גישת iMSD על מגוון רחב של מערכי מיקרוסקופיה אופטית מסחריים המבוססים על סריקת רסטר או הדמיה מבוססת מצלמה בשדה רחב, ללא צורך בכיול המערכת (נדרש רק אם יש צורך בהערכה מדויקת של גודל החלקיקים). פרמטר חשוב לשיטה לעבודה הוא דגימה מרחבית נכונה. ככלל, כדי להגיע להתכנסות משביעת רצון של האלגוריתם המתאים, הגודל המינימלי של אזור העניין להדמיה צריך להיות לפחות פי 3 גדול יותר מהתזוזה המקסימלית של העניין.
לסיכום, שיטת iMSD דורשת רק מיקרוסקופ המצויד לרכישה מהירה. ניתן לתייג את מבנה העניין לכל פלואורופור מקודד גנטית או אורגני, ובכך לאפשר הדמיה רב-ערוצית. התחזית היא שניתוח חוצה iMSD ישמש בעתיד הקרוב כדי לבחור תת-אוכלוסיות של ננו-מבנים תת-תאיים ולחשוף את האינטראקציות שלהם ואת הדיפוזיה המשותפת שלהם בתוך התא, כאשר האחרון הוא נושא חם בביופיזיקה התאית. אם פרט כלשהו הולך לאיבוד על ידי ניתוח iMSD, זה בהחלט קשור לכמות הגדולה של מידע מולקולרי בתוך ננו-מבנים תת-תאיים דינמיים. מידע כזה הוא בהכרח ממוצע החוצה במהלך המדידה בשל רזולוציה טמפורלית ירודה. תיאורטית, עם זאת, אין מגבלה טכנית בשל האפשרות לאחזר מידע מולקולרי, בתנאי שניתן להשיג מהירויות רכישה מספיקות8. בשל השיפורים המתמשכים במהירות/רגישות הגלאים ובטכנולוגיות ההדמיה, התחזית היא שמידע על כל התא התת-תאי ועל מרכיביו המולקולריים יופק ממערך נתונים יחיד.
Disclosures
למחברים אין ניגוד עניינים להצהיר עליו.
Acknowledgements
עבודה זו קיבלה מימון ממועצת המחקר האירופית (ERC) במסגרת תוכנית המחקר והחדשנות Horizon 2020 של האיחוד האירופי (הסכם מענקים No 866127, project CAPTUR3D).
Materials
| Name | Company | Catalog Number | Comments |
| 100x Penicillin-Streptomycin-Glutamine | Gibco | 10378-016 | Cell medium supplement |
| C-peptide-EGFP | Plasmid | ||
| DMEM High Glucose | Gibco | 31053028 | Cell medium (HeLa) |
| FBS | Gibco | 10082147 | Cell medium supplement |
| Fluorescein isothiocyanate-dextran 70 kDa | Sigma Aldrich | 46945-100MG-F | Reagent |
| HeLa | ATCC | CCL-61 | Cell Line |
| Lipofectamine 2000 | TermoFisher | 11668019 | Trasfection reagent |
| Lysotracker Red DND-99 | Gibco | L7528 | Reagent |
| Matlab | MathWork | Software | |
| Microscope-suitable cell dishes | Willco | GWSt-3522 | Petri dishes |
| Olympus FV1000 | Olympus Japan | Confocal microscope | |
| RPMI 1640 | Gibco | 11835063 | Cell medium (INS-1E) |
References
- Murphy, J. E., Padilla, B. E., Hasdemir, B., Cottrell, G. S., Bunnett, N. W. Endosomes: a legitimate platform for the signaling train. Proceedings of the National Academy of Sciences of the United States of America. 106 (42), 17615-17622 (2009).
- Mosesson, Y., Mills, G. B., Yarden, Y. Derailed endocytosis: an emerging feature of cancer. Nature Reviews Cancer. 8 (11), 835-850 (2008).
- Mellman, I., Yarden, Y. Endocytosis and cancer. Cold Spring Harbor Perspectives in Biology. 5 (12), 016949 (2013).
- Di Fiore, P. P. Endocytosis, signaling and cancer, much more than meets the eye. Preface. Molecular Oncology. 3 (4), 273-279 (2009).
- Bogan, J. S., Xu, Y., Hao, M. Cholesterol accumulation increases insulin granule size and impairs membrane trafficking. Traffic. 13 (11), 1466-1480 (2012).
- Ballabio, A., Gieselmann, V. Lysosomal disorders: From storage to cellular damage. Biochimica et Biophysica Acta. 1793 (4), 684-696 (2009).
- Hu, Y. -. B., Dammer, E. B., Ren, R. -. J., Wang, G. The endosomal-lysosomal system: from acidification and cargo sorting to neurodegeneration. Translational Neurodegeneration. 4, 18 (2015).
- Di Rienzo, C., Gratton, E., Beltram, F., Cardarelli, F. Spatiotemporal fluctuation analysis: a powerful tool for the future nanoscopy of molecular processes. Biophysical Journal. 111 (4), 679-685 (2016).
- Di Rienzo, C., Gratton, E., Beltram, F., Cardarelli, F. Fast spatiotemporal correlation spectroscopy to determine protein lateral diffusion laws in live cell membranes. Proceedings of the National Academy of Sciences of the United States of America. 110 (30), 12307-12312 (2013).
- Di Rienzo, C., Gratton, E., Beltram, F., Cardarelli, F. From fast fluorescence imaging to molecular diffusion law on live cell membranes in a commercial microscope. Journal of Visualized Experiments: JoVE. (92), e51994 (2014).
- Digiacomo, L., et al. Dynamic fingerprinting of sub-cellular nanostructures by image mean square displacement analysis. Scientific Reports. 7 (1), 14836 (2017).
- Durso, W., et al. Lysosome dynamic properties during neuronal stem cell differentiation studied by spatiotemporal fluctuation spectroscopy and organelle tracking. International Journal of Molecular Sciences. 21 (9), 3397 (2020).
- Ferri, G., et al. Insulin secretory granules labelled with phogrin-fluorescent proteins show alterations in size, mobility and responsiveness to glucose stimulation in living β-cells. Scientific Reports. 9 (1), 2890 (2019).
- Durso, W., D'Autilia, F., Amodeo, R., Marchetti, L., Cardarelli, F. Probing labeling-induced lysosome alterations in living cells by imaging-derived mean squared displacement analysis. Biochemical and Biophysical Research Communications. 503 (4), 2704-2709 (2018).
- Digiacomo, L., Digman, M. A., Gratton, E., Caracciolo, G. Development of an image Mean Square Displacement (iMSD)-based method as a novel approach to study the intracellular trafficking of nanoparticles. Acta Biomaterialia. 42, 189-198 (2016).
- Swanson, J. A., Watts, C. Macropinocytosis. Trends in Cell Biology. 5 (11), 424-428 (1995).
- Jones, A. T. Macropinocytosis: searching for an endocytic identity and role in the uptake of cell penetrating peptides. Journal of Cellular and Molecular Medicine. 11 (4), 670-684 (2007).
- Falcone, S., et al. Macropinocytosis: regulated coordination of endocytic and exocytic membrane traffic events. Journal of Cell Science. 119, 4758-4769 (2006).
- Kerr, M. C., et al. Visualisation of macropinosome maturation by the recruitment of sorting nexins. Journal of Cell Science. 119, 3967-3980 (2006).
- Rorsman, P., Renstrom, E. Insulin granule dynamics in pancreatic beta cells. Diabetologia. 46 (8), 1029-1045 (2003).
- Di Rienzo, C., Piazza, V., Gratton, E., Beltram, F., Cardarelli, F. Probing short-range protein Brownian motion in the cytoplasm of living cells. Nature Communications. 5 (1), 5891 (2014).
- Ferri, G., et al. Time-lapse confocal imaging datasets to assess structural and dynamic properties of subcellular nanostructures. Scientific Data. 5 (1), 180191 (2018).
- Scipioni, L., Gratton, E., Diaspro, A., Lanzanò, L. Phasor analysis of local ICS detects heterogeneity in size and number of intracellular vesicles. Biophysical Journal. 111 (3), 619-629 (2016).
- Digman, M. A., Dalal, R., Horwitz, A. F., Gratton, E. Mapping the number of molecules and brightness in the laser scanning microscope. Biophysical Journal. 94 (6), 2320-2332 (2008).
- Li, C. H., Bai, L., Li, D. D., Xia, S., Xu, T. Dynamic tracking and mobility analysis of single GLUT4 storage vesicle in live 3T3-L1 cells. Cell Research. 14 (6), 480-486 (2004).
- Donovan, K. W., Bretscher, A. Tracking individual secretory vesicles during exocytosis reveals an ordered and regulated process. Journal of Cell Biology. 210 (2), 181-189 (2015).
- Westphal, V., et al. Video-rate far-field optical nanoscopy dissects synaptic vesicle movement. Science. 320 (5873), 246-249 (2008).
- Tabei, S. M. A., et al. Intracellular transport of insulin granules is a subordinated random walk. Proceedings of the National Academy of Sciences of the United States of America. 110 (13), 4911-4916 (2013).
- Hebert, B., Costantino, S., Wiseman, P. W. Spatiotemporal image correlation spectroscopy (STICS) theory, verification, and application to protein velocity mapping in living CHO cells. Biophysical Journal. 88 (5), 3601-3614 (2005).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved