
Method Article
Зондирование структурно-динамических свойств субклеточных наноструктур трафика с помощью пространственно-временной флуктуационной спектроскопии
В этой статье
Резюме
Анализ среднего квадратного смещения (iMSD) применяется к макропиносомам, чтобы подчеркнуть их внутреннюю эволюционирующую во времени природу с точки зрения структурных и динамических свойств. Затем макропиносомы сравнивают с секреторными гранулами инсулина (ISG) в качестве эталона для субклеточных структур со средними структурными/динамическими свойствами, инвариантными во времени.
Аннотация
Среднее квадратное смещение, полученное из визуализации (iMSD), используется для решения структурных и динамических свойств субклеточных наноструктур, таких как везикулы, участвующие в эндо/экзоцитотическом трафике растворенных веществ и биомолекул. iMSD опирается на стандартную покадровую съемку, совместим с любой оптической установкой и не нуждается в остановке на отдельных объектах для извлечения траекторий. Из каждого следа iMSD рассчитывается уникальный триплет средних структурных и динамических параметров (т.е. размер, локальная диффузия, аномальный коэффициент) для построения «сигнатуры iMSD» исследуемой наноструктуры.
Эффективность этого подхода доказана здесь на примере макропиносом. Эти везикулы развиваются во времени, изменяя свой средний размер, количество и динамические свойства, переходя от ранних до поздних стадий внутриклеточного трафика. В качестве контроля в качестве ориентира для субклеточных структур, живущих в стационарном состоянии, при котором средние структурные и динамические свойства всей совокупности объектов инвариантны во времени. Анализ iMSD выделяет эти специфические особенности количественно и прокладывает путь к аналогичным применениям на субклеточном уровне, как в физиологических, так и в патологических состояниях.
Введение
Субклеточные наноструктуры (например, эндоцитарные/секреторные везикулы, органеллы) играют ключевую роль в клеточной сигнальной регуляции1. Правильная настройка их структурных (например, размер) и/или динамических (например, диффузионность) характеристик определяет, как клетка реагирует на внутренние или внешние раздражители 2,3,4. Исходя из этих доказательств, неудивительно, что изменения этих характеристик обнаруживаются при многих патологических состояниях. Примеры охватывают роль неправильно регулированного эндоцитоза при раке 2,3, структурные и динамические изменения, обнаруженные на уровне ISG в β-клетках, подвергшихся воздействию диабета 2 типа5, неправильную регуляцию лизосомальных структурных и транспортных свойств при глобоидно-клеточной лейкодистрофии или галактозилцерамидном липидозе6 и дисфункции в эндо-лизосомальном пути при нейродегенеративных расстройствах (например, болезнь Альцгеймера)7.
В этом контексте исследователи недавно доказали, что производительность стандартных методов оптической микроскопии может быть улучшена путем правильной настройки пространственного и временного разрешения выборки8. Это, в свою очередь, может дать дальнейшее понимание биологических процессов, имеющих отношение к делу. На практике это стало возможным благодаря алгоритму пространственно-временного флуктуационного анализа, который одновременно извлекает средние структурные и динамические свойства диффузных объектов непосредственно из стандартного стека изображений оптической микроскопии без необходимости предварительных знаний о интересующем биологическом объекте и извлечения однообъектных траекторий. Вся эта информация заключена в один вывод метода: iMSD trace9 (подробности о выводе и анализе трассировки iMSD приведены в дополнительном файле 1).
Полученный экспериментальный протокол состоит из нескольких шагов. Во-первых, визуализация интересующей области выполняется с высоким временным разрешением. Затем из стека изображений вычисляются средние пространственно-временные корреляционные функции. Наконец, путем гауссовского соответствия ряда корреляционных функций средний «закон диффузии» получается непосредственно из визуализации и анализируется для распознавания режима диффузии объекта. Потенциал метода уже был доказан для различных биологических объектов, начиная от молекул и заканчивая наночастицами и даже целыми субклеточными органеллами/структурами 9,10,11,12,13,14,15.
В данной статье сообщается о применении iMSD к макропиносомам, чтобы подчеркнуть их внутреннюю, необратимую эволюционирующую во времени природу с точки зрения их средних (т.е. на уровне всей популяции) структурных и динамических свойств. Кроме того, эти эндоцитарные везикулы сравниваются с ISG в качестве эталона для субклеточных структур в «стационарном состоянии», то есть состоянии, в котором средние структурные/динамические свойства всей популяции гранул остаются постоянными в любой момент времени. Макропиноцитоз определяет ряд событий, инициированных обширной реорганизацией (или взъерошением) плазматической мембраны с образованием внешней макропиноцитарной структуры, которая затеминтернализуется 16. Сформированные макропиносомы ранней стадии очень похожи на фагосомы. В то же время их можно отличить от других форм эндоцитарных пузырьков благодаря характерным большим размерам, морфологической неоднородности и отсутствию белково-покровных структур.
Биохимические анализы показали, что при интернализации макропиносомы постепенно обогащаются белковыми маркерами других эндоцитарных путей, что, в свою очередь, свидетельствует о том, что их идентичность постоянно меняется во время торговли17. Используя антитела против известных маркеров эндосомального пути, было продемонстрировано, что макропиносомы постепенно перенимают классические эндосомальные особенности: они уменьшаются в размерах, развиваются в поздние эндоцитарные структуры (например, лизосомы) или в конечном итоге теряют свою идентичность посредством мембранно-опосредованного извлечения специфических молекулярных маркеров (например, сортировка нексинов)18,19 . Общий сценарий заключается в том, что каждая макропиносома в клетке необратимо изменяет свою структурную и динамическую (а также молекулярную) идентичность во время транспортировки от плазматической мембраны к ее окончательной внутриклеточной судьбе. В результате структурные/динамические/молекулярные свойства всей популяции макропиносом также изменяются по тому же временному пути. Будучи внутренне чувствительным к средним свойствам всей совокупности наблюдаемых объектов, метод iMSD количественно изображает «эволюционирующую природу» путем количественной оценки ключевых средних параметров, т.е. локальной диффузии и аномального коэффициента (динамических свойств) и среднего размера макропиносом (структурного свойства) на любой стадии их внутриклеточного трафика.
Для сравнения, аналогичные измерения были выполнены на хорошо известной внутриклеточной мембранно-замкнутой структуре, ISG, в модели β-клеток. Как и макропиносомы, регуляция структурных и динамических свойств ISG, от их генезиса в Сети Транс Гольджи (TGN) до их экзоцитоза в плазматической мембране, имеет решающее значение для правильного выполнения функцииISG 20. Однако, в отличие от макропиносом, ISG живут в «стационарном состоянии», в котором в любое время все функциональные/структурные/молекулярные стадии продолжительности жизни ISG одновременно присутствуют внутри клетки, и каждая из них представлена определенной субпопуляцией ISG. Это означает, что, хотя каждая отдельная гранула необратимо эволюционирует от биогенеза к секреции, средние структурные/динамические свойства всей популяции гранул, как ожидается, останутся постоянными в любой момент времени (если условия стационарного состояния не изменяются, например, внешними стимулами, такими как глюкоза, холестерин и цитокины13). Это подтверждается анализом iMSD.
протокол
1. Пробоподготовка
- До эксперимента по микроскопии субкультурные клетки в чашках подходили для микроскопических применений.
- Дважды вымойте 10-сантиметровую посуду, обработанную культурой ткани, из сливающихся клеток HeLa или INS 1E (инсулинома β-клеточных) с 0,01 M 1x PBS, добавьте 1 мл 0,05% трипсина-ЭДТА (1x) и поместите его в 37 °C, увлажненный, 5% CO2 инкубатор в течение 5 мин.
- Повторно суспендируют отсоединенные ячейки путем добавления 9 мл полной среды DMEM (для клеток HeLa) или RPMI 1640 (для клеток INS-1E) и собирают конечные 10 мл в центрифужных трубках.
- Посейте примерно 2 × 105 ячеек в каждой тарелке размером 35 мм х 10 мм в конечном объеме 1 мл среды. Инкубируют клетки в течение 24 ч при 37 °C и 5% CO2.
- Чтобы флуоресцентно маркировать лизосомы, используйте LysoTracker Red DND-99.
- Раствор развести в 1 мл предварительно расплавленной среды до конечной концентрации красителя 70 нМ.
- Замените среду из блюда свежей средой, содержащей LysoTracker. Инкубируют клетки в среде, содержащей LysoTracker, в течение 20 мин при 37 °C в атмосфере 5% CO2 и промыть их дважды свежей средой перед экспериментом.
- Для флуоресцентной маркировки макропиносом используют 70-кДа флуоресцеина изотиоцианат-декстран. Промыть субкультурные клетки трижды 0,01 M 1x фосфатно-буферным физиологическим раствором (PBS), заменить декстрансодержащей средой (1 мг/мл) и инкубировать при 37 °C в течение 30 мин. Прежде чем приступить к микроскопическому эксперименту, трижды промыть клетки свежей средой.
- Для флуоресцентной маркировки ISG в клетках INS-1E трансфектируют клетки с помощью трансфекционного реагента (см. Таблицу материалов) и плазмиды13 зеленого флуоресцентного белка (EGFP) с улучшенным С-пептидом (EGFP) в соответствии с протоколом производителя и инкубируют при 37 °C в течение 24 ч в атмосфере 5%CO2 перед экспериментом.
2. Сбор данных
- Чтобы микроскоп уравновешивался при нужной температуре и атмосфере, включите систему управления инкубатором микроскопа не менее чем за 2 часа до начала эксперимента.
ПРИМЕЧАНИЕ: Каждое приобретение представляет собой покадровую серию. - Получайте изображения с помощью инвертированного конфокального микроскопа, оснащенного 60-кратным, 1,2-кратным объективом с числовой апертурой (NA).
- Используйте 488 нм аргоновый лазер для возбуждения EGFP (трансфектированных клеток) и флуоресцеин-меченых макропиносом. Соберите флуоресцентное излучение от 500 до 600 нм с помощью стандартного детектора фотоумножителя.
- Используйте 543-нм heNe-лазер для возбуждения Lysotracker и сбора его флуоресцентного излучения между 555 и 655 нм.
- Установите диаметр точечного отверстия обнаружения на размер 1 Airy. Для каждого приобретения соберите серию из 1000 последовательных кадров. Установите время выдержки пикселя равным 2 мкс/пиксель для времени кадра 129 мс.
ПРИМЕЧАНИЕ: Каждый кадр состоял из 256 x 256 пикселей (16 бит/пиксель) с физическим размером 69 нм/пиксель, что примерно соответствует площади 17 мкм x 17 мкм.
3. IРасчет MSD
ПРИМЕЧАНИЕ: Чтобы правильно выполнить расчет, используйте программное обеспечение, способное к числовым вычислениям и программированию сценариев. Конкретный сценарий (т.е. для файла сценария 'iMSD.m' см. Вспомогательный файл 1) должен присутствовать в том же каталоге, содержащем обрабатываемые серии изображений. Каждое изображение серии должно быть сохранено в виде отдельного файла «.tif».
- Чтобы правильно инициализировать инструментальные параметры, используемые для приобретений, откройте iMSD.m с помощью программного текстового редактора и измените его первый раздел следующим образом.
- Установите N в качестве числа кадров во временных рядах (например, 1000 в этом протоколе).
- Установить px_size: размер пикселя, выраженный в мкм (например, 0,069 в этом протоколе).
- Set f: временное разрешение каждого кадра, выраженное в секундах (например, 0,129 в настоящем протоколе).
- Установите фильтр: двоичный ввод для коррекции фона, установите значение '0' для обработки необработанных изображений или установите значение '1' для выполнения вычитания фона на основе пороговых значений.
- Установить av_toll: порог для коррекции фона; любой пиксель с интенсивностью ниже этого значения будет установлен как 0, если Filter=1.
- Установите бит в качестве целого числа, определяющего выборку интенсивности (например, 8 бит, 16 бит).
- Сохраните и запустите отредактированный файл сценария iMSD.m.
- Проверьте выполнение скрипта.
ПРИМЕЧАНИЕ: Статус обработки вычислений можно проверить в командном окне; при возникновении какой-либо неустранимой проблемы процесс будет прерван, и появится предупреждающее сообщение, отображающее тип ошибки и соответствующий код строки. В противном случае выполните шаги 3.3.1-3.3.3.- Импортируйте стек изображений и вычтите фон (при необходимости).
- Вычислите пространственно-временную корреляционную функцию G(ξ,η,τ) методом Фурье.
- Подойдите к пространственно-временной корреляционной функции с 2D-функцией Гаусса.
ПРИМЕЧАНИЕ: В конце процесса в командном окне будут показаны выходные значения процедур установки σ2(τ): сообщаются средние значения, погрешность и соответствующая добротность (R2) подгонки.
- Проверьте графический вывод.
ПРИМЕЧАНИЕ: Кривая iMSD и соответствующие кривые подгонки показаны на трех отдельных панелях, каждая из которых предназначена для различных типов используемых уравнений подгонки: броуновская диффузия, аномальная диффузия или ограниченная диффузия. Для каждой кривой значение R2 указывается в легенде графика. - Проверьте вывод текста.
ПРИМЕЧАНИЕ: В конце процесса создается файл '.xls' с тем же именем, что и исходный файл time-lapse '.tif'. Первый лист содержит значения ξ, η, τ и σ2 , рассчитанные для каждой временной задержки. Входные параметры и основные расчетные выходные значения сообщаются во втором листе, т.е. коэффициент диффузии, аномальный коэффициент и ось Y σперехвата 20 .
Результаты
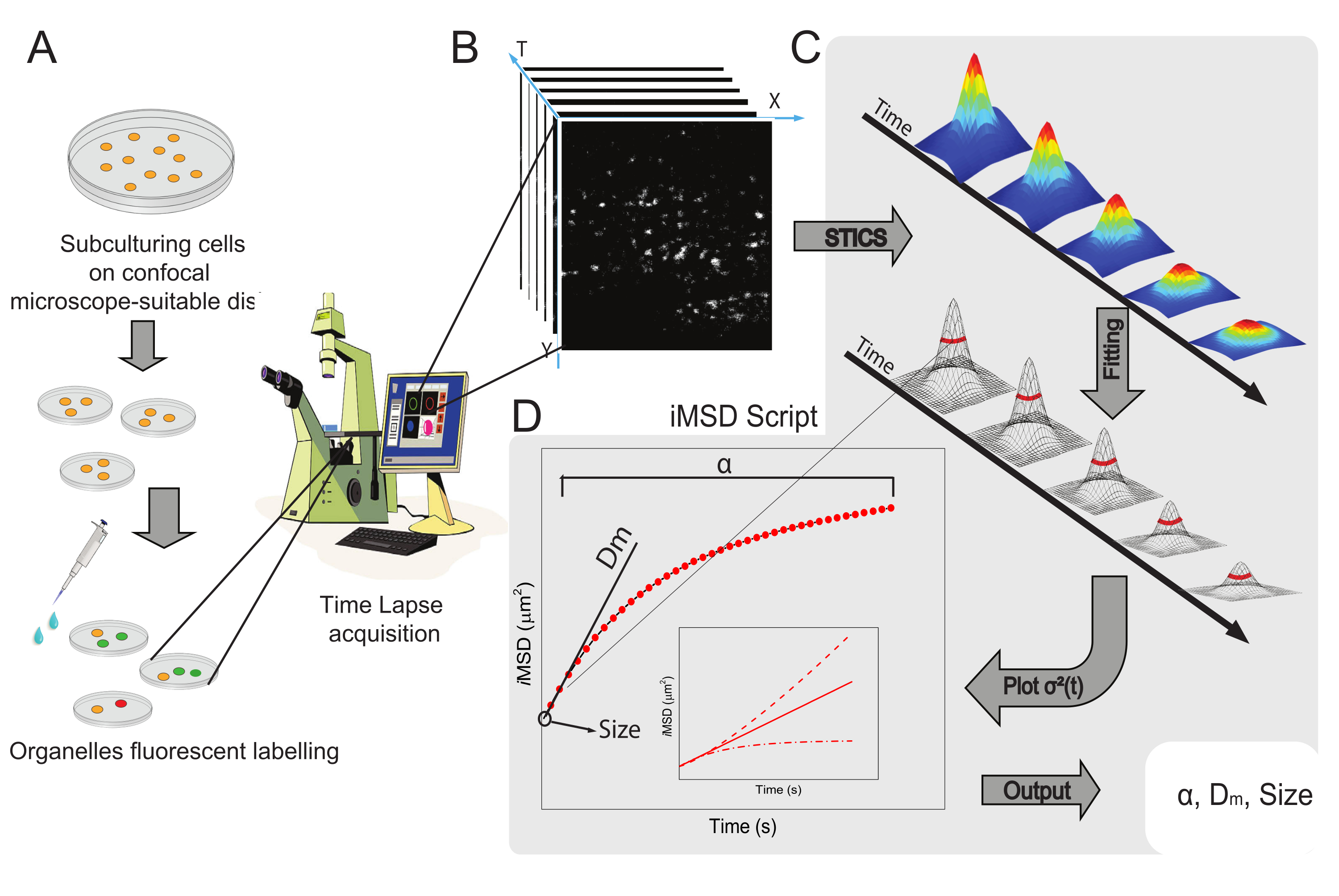
Общий рабочий процесс метода представлен на рисунке 1. В нем кратко излагаются основные этапы, представленные в разделе протокола, от пробоподготовки (рисунок 1A) до покадровой визуализации внутриклеточных наноструктур (рисунок 1B), флуктуационного анализа для расчета ряда пространственно-временных корреляционных функций (рисунок 1C) и подгонки для получения средних структурных/динамических свойств исследуемого объекта (рисунок 1D).
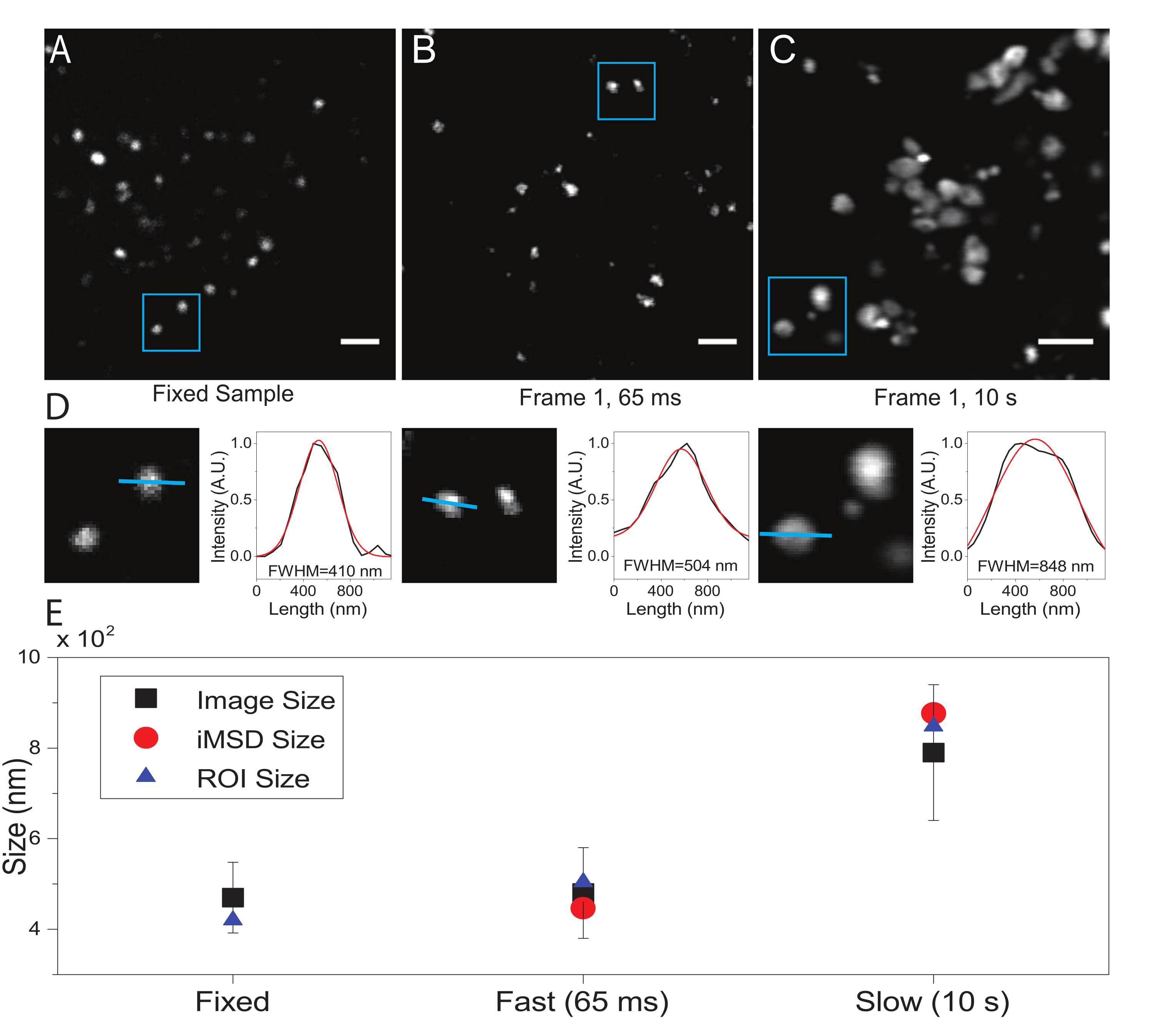
Критическим параметром является временное разрешение, принятое для визуализации субклеточного объекта, представляющего интерес. Это экспериментальное значение установит порог времени, при котором будет измеряться минимальное среднее смещение интересующих объектов. Однако предпочтительным условием является установка временного разрешения изображения, при котором интересующий объект кажется «неподвижным» в пределах захваченного кадра, то есть он отображает характерный размер, который в среднем не деформируется из-за скорости изображения. Это технически возможно, если объект интереса представляет собой мембранно-замкнутую субклеточную структуру или органеллу (как в данном случае). Как правило, субклеточные структуры демонстрируют локальные коэффициенты диффузии (D, мкм2/с, см. Таблицу 1), которые на несколько порядков ниже, чем у изолированных одиночных молекул в цитоплазме (например, GFP21). Валидация может быть выполнена путем искусственной иммобилизации интересующей органеллы (например, путем химической фиксации). Действительно, это условие может служить ориентиром для определения фактического размера органеллы в используемых экспериментальных условиях (например, длина волны возбуждения, размер пикселя, объектив).
Здесь, хотя лизосомы использовались в качестве тестовых органелл для этой процедуры, результаты действительны независимо от целевой структуры. На рисунке 2A показано изображение фиксированной (т.е. неподвижной) лизосомы вместе с захватом, выполненным на живых клетках с соответствующим временным разрешением (т.е. обычно ниже 100 мс/кадр; например, 65 мс/кадр в примере на рисунке 2B) и захватом, выполненным намеренно при очень низком временном разрешении (например, 10 с/кадр в примере на рисунке 2C). ). Для каждого условия размер рассеивающего объекта извлекается следующим образом: i) профиль интенсивности пятна выводится линейным инструментом в программном обеспечении ImageJ; ii) профиль интенсивности строится и интерполируется функцией Гаусса для вычисления полной ширины при половинном максимальном значении (FWHM), которое, в свою очередь, используется в качестве оценки диаметра пятна (рисунок 2D). Как и ожидалось и показано на графике рисунка 2E, получение при очень высоком временном разрешении (т.е. 65 мс/кадр) дает средний размер структуры, близкий к тому, который получен в фиксированном образце, либо с помощью стандартного инструмента, описанного выше, либо путем извлечения перехвата оси iMSD Y. Вместо этого захват на медленной скорости приводит к увеличению видимого размера структуры из-за естественной динамики структуры во время визуализации. В неоптимальных экспериментальных условиях извлеченная структурно-динамическая информация не отражает достоверно внутренние свойства исследуемого объекта.
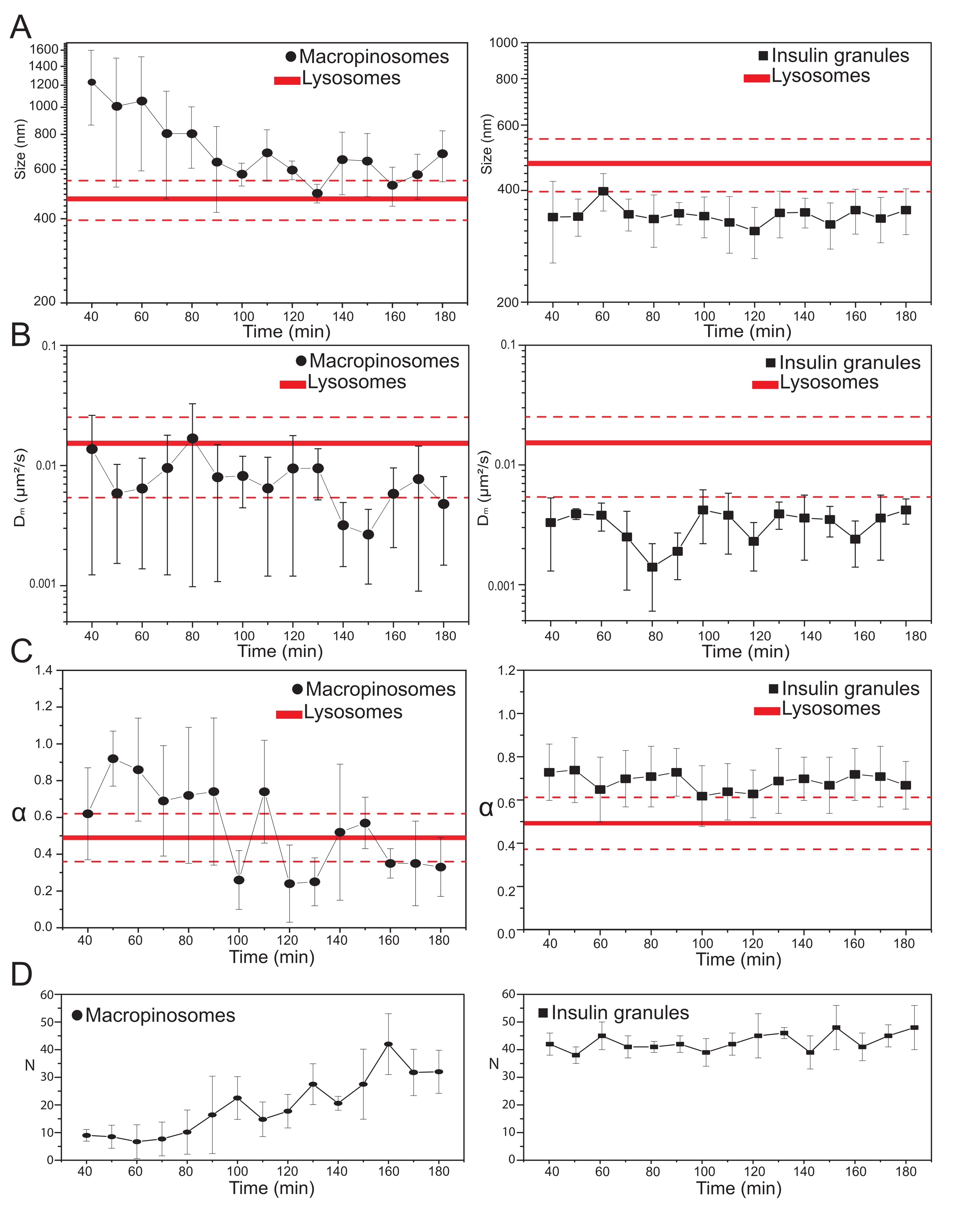
После выбора основных экспериментальных параметров могут быть получены наборы данных для целевых внутриклеточных структур. Для макропиносом после 20 мин инкубации клеток с 70 кДа декстрансом временные ряды меченых внутриклеточных структур приобретали в разные моменты времени после лечения, от 30 мин до примерно 180 мин. Интересно, что при незаконном обороте обнаруживается постепенное изменение структурно-динамических свойств макропиносом (рисунок 3; на участках слева сообщается о распределениях σ02, Dm, α и N для макропиносом). Хотя во время незаконного оборота не обнаруживается никаких очевидных изменений в локальной диффузии (Dm) макропиносом, как характерный размер (σ02), так и общий режим движения (α) со временем развиваются.
Особо следует отметить, что при незаконном обороте наблюдается уменьшение среднего размера макропиносом (рисунок 3А, слева) вместе с сопутствующим увеличением субдиффузного характера их движения (т.е. обозначается уменьшением значений α, рисунок 3С, левая панель). Кроме того, количество макропиносом было извлечено из каждого приобретения: результаты, представленные на рисунке 3D, левая панель, ясно показывают увеличение числа макропиносом во времени. Все эти результаты хорошо согласуются с ожиданиями, потому что макропиносомы, меченые декстраном, должны возникать как изолированные, большие мембранно-заключенные везикулы на плазматической мембране (которые также компетентны для движения вдоль цитоскелетных компонентов), но должны постепенно взаимодействовать с эндо-лизосомальным путем, состоящим из большой популяции меньших и случайно диффузных структур.
Как и предполагалось выше, результаты для макропиносом контрастируют с аналогичными измерениями, выполненными на ISG (рисунок 3, правая колонка). Гранулы инсулина не показывают временно развивающейся тенденции структурных/динамических параметров, полученных из iMSD (и их среднего числа в клетке) в том же временном окне, которое наблюдалось для макропиносом. Более того, характерные значения σ02, Dm и α сильно отличаются от значений лизосом, используемых снова в качестве эталона. Этот результат подтверждает идею, как и предполагалось выше, о том, что гранулы исследуются в «стационарном состоянии», в котором в любое время средние структурные/динамические свойства всей популяции ISG остаются неизменными (т. е. они остаются постоянными, если условия стационарного состояния не изменяются, например, из-за внешних раздражителей).

Рисунок 1: Экспериментальный рабочий процесс. (A) Клетки были покрыты 24 ч (48 ч для экспериментов по трансфекции) перед конфокальными экспериментами на клеточных чашках, пригодных для микроскопических применений. Затем клетки соответствующим образом обрабатывали в соответствии с методом маркировки (см. протокол) для окрашивания цитоплазматической органеллы, представляющей интерес. (B) Типичное конфокальное приобретение состоит из стопки изображений (time-lapse) цитоплазматической части живой клетки, описывающих временную эволюцию меченой динамики органелл. (C) Покадровый фильм анализируется с помощью специально созданного сценария Matlab, сначала вычисляя пространственно-временную корреляционную функцию изображения и гауссову подгонку для построения кривых iMSD (D) и связанных с ними извлеченных параметров подгонки, описывающих параметры структурной динамики изображенных органелл. Сокращения: iMSD = среднее квадратное смещение, полученное из визуализации; STICS = пространственно-временная корреляционная спектроскопия изображений; α = аномальный коэффициент диффузии; Dm = локальная диффузионная способность; σ2(τ) = дисперсия. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 2: Правильные экспериментальные параметры. (А) Примерное изображение окрашенных лизосом в неподвижном образце. Шкала = 2 мкм. (Б) Первый кадр стопки изображений окрашенных лизосом в живой клетке, полученных с соответствующими параметрами. Временное разрешение: 65 мс/кадр. Шкала бара = 2 мкм. (C) Первый кадр стопки изображений окрашенных лизосом в живой клетке, полученных на низкой скорости: видна артефактная деформация видимого размера лизосомы из-за движения органелл во время визуализации. Временное разрешение: 10 с/кадр. Шкала = 2 мкм. (D) Пример расчета размера для изображенных лизосом с синей рентабельностью инвестиций (A), (B) и (C). Профиль интенсивности вдоль синей линии был снабжен функцией Гаусса для извлечения FWHM, т.е. оценки размера пятна. Значения FWHM сообщаются для каждой установки. (E) Графическое представление значений размеров, полученных с помощью анализа изображений, описанного на панели (D) для всех изображенных лизосом (черный квадрат, среднее значение и стандартное отклонение), для лизосом, заключенных в синюю рентабельность инвестиций (синий треугольник), и полученное с помощью анализа iMSD (красный круг). Эта цифра из 22. Сокращения: ROI = интересующий регион; FWHM = полная ширина при половинном максимуме; iMSD = среднее квадратное смещение, полученное из визуализации. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 3: Временная эволюция меченых органелл. (A) Графики значений размера, извлеченных iMSD, по сравнению с временной прогрессией последующих приобретений, представленных в виде среднего значения измеренных значений в приобретениях, выполненных в 10-минутном временном окне. Слева прогрессирующее уменьшение среднего размера макропиносом (черные круги), а справа — инвариантный по времени размер секреторных гранул инсулина (черных квадратов) по сравнению с лизосомами, представленный как среднее значение размера (более толстая красная линия) ± стандартное отклонение (пунктирные красные линии). (B) и (C) Временная прогрессия коэффициентов Dm и α для макропиносом (слева) и гранул инсулина (справа), извлеченных методом анализа iMSD. (D) Временная эволюция числа меченых макропиносом и гранул инсулина, измеренных в первом кадре каждого приобретенного покадрового фильма. Сокращения: iMSD = среднее квадратное смещение, полученное из визуализации; α = аномальный коэффициент диффузии; Dm = локальная диффузионная способность, N = число. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
| Органелла | Этикетирование | Клеточная линия | Размер (нм) | Dm ( × 10-3 мкм2/с) | α | N | См. | ||
| Ранняя эндосома (EE) | CellLight Ранняя эндосома GFP | Хела | 395±74 | 3.0±2.4 | 1.02±0.20 | 40 | 10 | ||
| Поздняя эндосома (LE) | CellLight Поздняя эндосома GFP | Хела | 693±102 гг. | 15.4±10.6 | 0.57±0.16 | 58 | 10 | ||
| Лизосома (LY) | ЛисоТрекер ДНД-99 | Хела | 471±76 | 15.3±9.0 | 0.49±0.13 | 143 | 10, 14 | ||
| Кавеола (CAV) | Кавеолин-ЭГФП | Хела | 405±49 руб. | 3.1±1.8 | 1.00±0.22 | 15 | 10 | ||
| Весцикл с клатриновым покрытием (CCV) | Трансферрин-Алекса 488 | Хела | 513 62 ± гг. | 16.2±9.9 | 0.48±0.17 | 33 | 10 | ||
| Гранулы инсулина (IG) | С-пептид-ЭГФП | ИНС-1Е | 335±56 | 3.0±1.7 | 0.70±0.14 | 107 | 11 | ||
| Ранние макропиносомы (EMCR) | Флуоресцеин-Декстран 70 кДа | Хела | 979 423 ± гг. | 8.3±9 | 0.79±0.27 | 36 | 10 | ||
| Промежуточные макропиносомы (IMCR) | Флуоресцеин-Декстран 70 кДа | Хела | 702± 180 чел. | 13.7±19.9 | 0.60±0.38 | 29 | 10 | ||
| Поздние макропиносомы (LMCR) | Флуоресцеин-Декстран 70 кДа | Хела | 592±127 гг. | 5.8±4.7 | 0.39±0.21 | 21 | 10 | ||
Таблица 1: Структурные и динамические параметры,извлеченные из MSD. В таблице показаны значения размера, Dm и коэффициента α, измеренные для различных органелл, с указанием стратегий маркировки, используемой клеточной линии и количества анализируемых приобретений. Значения указываются как среднее ± стандартного отклонения. Сокращения: iMSD = среднее квадратное смещение, полученное из визуализации; α = аномальный коэффициент диффузии; Dm = локальная диффузионная способность, N = число; GFP = зеленый флуоресцентный белок; EGFP = улучшенный зеленый флуоресцентный белок.
Дополнительный файл 1: Подробная информация о выводе и анализе трассировки iMSD. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Вспомогательный файл 1: Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Обсуждение
Свойства и преимущества iMSD очевидны по сравнению с методами, доступными для получения аналогичной информации. Для структурной информации предпочтительным выбором является анализ просвечивающей электронной микроскопии (ТЭМ). С помощью этого метода ультраструктурные детали с молекулярным разрешением и даже за его пределами могут быть извлечены, даже для субклеточных наноструктур. Тем не менее, своеобразное пространственное разрешение ТЕА достигается за счет информации во временном измерении, что представляет здесь интерес. Чтобы компенсировать это, особый интерес представляют последние достижения в области технологий визуализации живых клеток. К ним относятся новые флуоресцентные маркеры с повышенными характеристиками (например, яркость и фотостабильность), оптимизированные процедуры маркировки и более чувствительные детекторы. Кроме того, доступны аналитические инструменты для решения как структурных (например, «размер» путем фазорного анализа локальной корреляционной спектроскопии изображений, PLICS23, агрегации/олигомеризации с помощью анализа чисел и яркости24), так и динамических (например, закон диффузии путем отслеживания одной частицы, т.е. (SPT)25,26,27,28 ) параметры по субклеточной шкале. Метод ППП обеспечивает прямой доступ к траектории объекта и его MSD. Однако недостатком является необходимость измерения низкой плотности зонда и очень ярких меток и множества траекторий одного объекта для удовлетворения статистических критериев. Что касается временного разрешения измерения, то неорганические, фотостабильные зонды (например, квантовые точки или наночастицы металлов) могут повысить производительность SPT, но за счет сложных процедур производства и маркировки.
По сравнению с этими стандартами метод iMSD, описанный здесь, показывает некоторые ключевые преимущества. Во-первых, этот подход может быть использован в сочетании с относительно тусклыми флуоресцентными метками, такими как генетически закодированные флуоресцентные белки (например, применение к ISG). Таким образом, по сравнению с SPT достигается более высокое временное разрешение (с использованием той же метки) за счет меньшего количества фотонов, требуемого8. Во-вторых, метод iMSD ограничен только временным разрешением, но не дифракцией. Фактически, несмотря на используемую оптическую установку с дифракционным ограничением, средние молекулярные смещения даже ниже дифракционного предела могут быть измерены, как уже было продемонстрировано для молекулярных потоков с помощью STICS29. Фактическое разрешение при измерении перемещений зависит от того, насколько точно (с точки зрения сигнал-шум) может быть измерена корреляционная функция, тем самым объясняя, почему она не ограничена дифракцией. Таким образом, представляется очевидным, что минимальное смещение, которое может быть измерено, зависит от диффузии интересующего объекта и временного разрешения установки визуализации.
В связи с этим важно учитывать, что применение к субклеточным наноструктурам, таким как макропиносомы или гранулы инсулина, с лазерным сканирующим микроскопом является оптимальным: доступная скорость сканирования значительно превышает динамику интересующего объекта. В таком случае движение объектов при приобретении ничтожно мало, а корреляционная функция может быть аппроксимирована гауссовской функцией. Наконец, подход iMSD может быть легко применен к широкому спектру коммерческих оптических микроскопических установок, основанных на растровом сканировании или широкоугольном изображении на основе камеры, без необходимости калибровки системы (требуется только в том случае, если необходимо достичь точной оценки размера частиц). Важным параметром для работы метода является правильная пространственная выборка. Как правило, для достижения удовлетворительной сходимости алгоритма подгонки минимальный размер области, представляющей интерес для визуализации, должен быть как минимум в 3 раза больше максимального смещения интереса.
В заключение, метод iMSD требует только микроскопа, оснащенного для быстрого получения. Интересующая структура может быть помечена любым генетически закодированным или органическим флуорофором, что обеспечивает многоканальную визуализацию. Предполагается, что кросс-iMSD-анализ будет использоваться в ближайшем будущем для отбора субпопуляций субклеточных наноструктур и выявления их взаимодействий и кодиффузии внутри клетки, причем последняя является горячей темой в клеточной биофизике. Если какие-либо детали теряются при анализе iMSD, это, безусловно, связано с большим количеством молекулярной информации в динамических субклеточных наноструктурах. Такая информация неизбежно уселяется во время измерения из-за плохого временного разрешения. Теоретически, однако, не существует технического ограничения из-за возможности извлечения молекулярной информации при условии, что могут быть достигнуты достаточные скорости сбора8. В связи с постоянным улучшением скорости/чувствительности детектора и технологий визуализации предполагается, что информация обо всем субклеточном компартменте и его молекулярных составляющих будет извлекаться из одного набора данных.
Раскрытие информации
У авторов нет конфликта интересов, о котором можно было бы заявить.
Благодарности
Эта работа получила финансирование от Европейского исследовательского совета (ERC) в рамках Программы исследований и инноваций Европейского союза Horizon 2020 (грантовое соглашение No 866127, проект CAPTUR3D).
Материалы
| Name | Company | Catalog Number | Comments |
| 100x Penicillin-Streptomycin-Glutamine | Gibco | 10378-016 | Cell medium supplement |
| C-peptide-EGFP | Plasmid | ||
| DMEM High Glucose | Gibco | 31053028 | Cell medium (HeLa) |
| FBS | Gibco | 10082147 | Cell medium supplement |
| Fluorescein isothiocyanate-dextran 70 kDa | Sigma Aldrich | 46945-100MG-F | Reagent |
| HeLa | ATCC | CCL-61 | Cell Line |
| Lipofectamine 2000 | TermoFisher | 11668019 | Trasfection reagent |
| Lysotracker Red DND-99 | Gibco | L7528 | Reagent |
| Matlab | MathWork | Software | |
| Microscope-suitable cell dishes | Willco | GWSt-3522 | Petri dishes |
| Olympus FV1000 | Olympus Japan | Confocal microscope | |
| RPMI 1640 | Gibco | 11835063 | Cell medium (INS-1E) |
Ссылки
- Murphy, J. E., Padilla, B. E., Hasdemir, B., Cottrell, G. S., Bunnett, N. W. Endosomes: a legitimate platform for the signaling train. Proceedings of the National Academy of Sciences of the United States of America. 106 (42), 17615-17622 (2009).
- Mosesson, Y., Mills, G. B., Yarden, Y. Derailed endocytosis: an emerging feature of cancer. Nature Reviews Cancer. 8 (11), 835-850 (2008).
- Mellman, I., Yarden, Y. Endocytosis and cancer. Cold Spring Harbor Perspectives in Biology. 5 (12), 016949 (2013).
- Di Fiore, P. P. Endocytosis, signaling and cancer, much more than meets the eye. Preface. Molecular Oncology. 3 (4), 273-279 (2009).
- Bogan, J. S., Xu, Y., Hao, M. Cholesterol accumulation increases insulin granule size and impairs membrane trafficking. Traffic. 13 (11), 1466-1480 (2012).
- Ballabio, A., Gieselmann, V. Lysosomal disorders: From storage to cellular damage. Biochimica et Biophysica Acta. 1793 (4), 684-696 (2009).
- Hu, Y. -. B., Dammer, E. B., Ren, R. -. J., Wang, G. The endosomal-lysosomal system: from acidification and cargo sorting to neurodegeneration. Translational Neurodegeneration. 4, 18 (2015).
- Di Rienzo, C., Gratton, E., Beltram, F., Cardarelli, F. Spatiotemporal fluctuation analysis: a powerful tool for the future nanoscopy of molecular processes. Biophysical Journal. 111 (4), 679-685 (2016).
- Di Rienzo, C., Gratton, E., Beltram, F., Cardarelli, F. Fast spatiotemporal correlation spectroscopy to determine protein lateral diffusion laws in live cell membranes. Proceedings of the National Academy of Sciences of the United States of America. 110 (30), 12307-12312 (2013).
- Di Rienzo, C., Gratton, E., Beltram, F., Cardarelli, F. From fast fluorescence imaging to molecular diffusion law on live cell membranes in a commercial microscope. Journal of Visualized Experiments: JoVE. (92), e51994 (2014).
- Digiacomo, L., et al. Dynamic fingerprinting of sub-cellular nanostructures by image mean square displacement analysis. Scientific Reports. 7 (1), 14836 (2017).
- Durso, W., et al. Lysosome dynamic properties during neuronal stem cell differentiation studied by spatiotemporal fluctuation spectroscopy and organelle tracking. International Journal of Molecular Sciences. 21 (9), 3397 (2020).
- Ferri, G., et al. Insulin secretory granules labelled with phogrin-fluorescent proteins show alterations in size, mobility and responsiveness to glucose stimulation in living β-cells. Scientific Reports. 9 (1), 2890 (2019).
- Durso, W., D'Autilia, F., Amodeo, R., Marchetti, L., Cardarelli, F. Probing labeling-induced lysosome alterations in living cells by imaging-derived mean squared displacement analysis. Biochemical and Biophysical Research Communications. 503 (4), 2704-2709 (2018).
- Digiacomo, L., Digman, M. A., Gratton, E., Caracciolo, G. Development of an image Mean Square Displacement (iMSD)-based method as a novel approach to study the intracellular trafficking of nanoparticles. Acta Biomaterialia. 42, 189-198 (2016).
- Swanson, J. A., Watts, C. Macropinocytosis. Trends in Cell Biology. 5 (11), 424-428 (1995).
- Jones, A. T. Macropinocytosis: searching for an endocytic identity and role in the uptake of cell penetrating peptides. Journal of Cellular and Molecular Medicine. 11 (4), 670-684 (2007).
- Falcone, S., et al. Macropinocytosis: regulated coordination of endocytic and exocytic membrane traffic events. Journal of Cell Science. 119, 4758-4769 (2006).
- Kerr, M. C., et al. Visualisation of macropinosome maturation by the recruitment of sorting nexins. Journal of Cell Science. 119, 3967-3980 (2006).
- Rorsman, P., Renstrom, E. Insulin granule dynamics in pancreatic beta cells. Diabetologia. 46 (8), 1029-1045 (2003).
- Di Rienzo, C., Piazza, V., Gratton, E., Beltram, F., Cardarelli, F. Probing short-range protein Brownian motion in the cytoplasm of living cells. Nature Communications. 5 (1), 5891 (2014).
- Ferri, G., et al. Time-lapse confocal imaging datasets to assess structural and dynamic properties of subcellular nanostructures. Scientific Data. 5 (1), 180191 (2018).
- Scipioni, L., Gratton, E., Diaspro, A., Lanzanò, L. Phasor analysis of local ICS detects heterogeneity in size and number of intracellular vesicles. Biophysical Journal. 111 (3), 619-629 (2016).
- Digman, M. A., Dalal, R., Horwitz, A. F., Gratton, E. Mapping the number of molecules and brightness in the laser scanning microscope. Biophysical Journal. 94 (6), 2320-2332 (2008).
- Li, C. H., Bai, L., Li, D. D., Xia, S., Xu, T. Dynamic tracking and mobility analysis of single GLUT4 storage vesicle in live 3T3-L1 cells. Cell Research. 14 (6), 480-486 (2004).
- Donovan, K. W., Bretscher, A. Tracking individual secretory vesicles during exocytosis reveals an ordered and regulated process. Journal of Cell Biology. 210 (2), 181-189 (2015).
- Westphal, V., et al. Video-rate far-field optical nanoscopy dissects synaptic vesicle movement. Science. 320 (5873), 246-249 (2008).
- Tabei, S. M. A., et al. Intracellular transport of insulin granules is a subordinated random walk. Proceedings of the National Academy of Sciences of the United States of America. 110 (13), 4911-4916 (2013).
- Hebert, B., Costantino, S., Wiseman, P. W. Spatiotemporal image correlation spectroscopy (STICS) theory, verification, and application to protein velocity mapping in living CHO cells. Biophysical Journal. 88 (5), 3601-3614 (2005).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены