
Method Article
Sondare le proprietà strutturali e dinamiche delle nanostrutture subcellulari di traffico mediante spettroscopia di fluttuazione spaziotemporale
In questo articolo
Riepilogo
L'analisi iMSD (Imaging-derived mean square displacement) viene applicata ai macropinosomi per evidenziare la loro natura intrinseca in evoluzione temporale in termini di proprietà strutturali e dinamiche. I macropinosomi vengono quindi confrontati con i granuli secretori di insulina (ISG) come riferimento per strutture subcellulari con proprietà strutturali/dinamiche medie invarianti nel tempo.
Abstract
Lo spostamento quadrato medio derivato dall'imaging (iMSD) viene utilizzato per affrontare le proprietà strutturali e dinamiche delle nanostrutture subcellulari, come le vescicole coinvolte nel traffico endo/esocitotico di soluti e biomolecole. iMSD si basa sull'imaging time-lapse standard, è compatibile con qualsiasi configurazione ottica e non ha bisogno di soffermarsi su singoli oggetti per estrarre traiettorie. Da ogni traccia iMSD, una tripletta unica di parametri strutturali e dinamici medi (cioè dimensione, diffusività locale, coefficiente anomalo) viene calcolata e combinata per costruire la "firma iMSD" della nanostruttura in studio.
La potenza di questo approccio è dimostrata qui con il caso esemplare dei macropinosomi. Queste vescicole si evolvono nel tempo, cambiando la loro dimensione media, il numero e le proprietà dinamiche passando dalle fasi iniziali a quelle tardive del traffico intracellulare. Come controllo, i granuli secretori di insulina (ISG) sono usati come riferimento per le strutture subcellulari che vivono in uno stato stazionario in cui le proprietà strutturali e dinamiche medie dell'intera popolazione di oggetti sono invarianti nel tempo. L'analisi iMSD evidenzia quantitativamente queste caratteristiche peculiari e apre la strada ad applicazioni simili a livello subcellulare, sia nello stato fisiologico che patologico.
Introduzione
Le nanostrutture subcellulari (ad esempio, vescicole endocitiche/secretorie, organelli) svolgono un ruolo fondamentale nella regolazione della segnalazione cellulare1. Una corretta messa a punto delle loro caratteristiche strutturali (ad esempio, dimensioni) e / o dinamiche (ad esempio, diffusività) determina il modo in cui la cellula risponde agli stimoli interni o esterni 2,3,4. Sulla base di queste prove, non sorprende che le alterazioni di queste caratteristiche si trovino in molte condizioni patologiche. Gli esempi comprendono il ruolo dell'endocitosi mal regolata nel cancro 2,3, le alterazioni strutturali e dinamiche riscontrate a livello di ISG nelle cellule β esposte alle condizioni del diabete di tipo 25, la cattiva regolazione delle proprietà strutturali e di trasporto lisosomiale nella leucodistrofia a cellule globoidi o nella lipidosi galattosilceramide6 e disfunzionalità nella via endo-lisosomiale nelle malattie neurodegenerative (ad esempio il morbo di Alzheimer)7.
In questo contesto, i ricercatori hanno recentemente dimostrato che le prestazioni dei metodi di microscopia ottica standard possono essere migliorate regolando correttamente la risoluzione di campionamento spaziale e temporale8. Questo, a sua volta, può fornire ulteriori informazioni sui processi biologici di rilevanza. In pratica, ciò è reso possibile da un algoritmo di analisi della fluttuazione spaziotemporale, che estrae contemporaneamente le proprietà strutturali e dinamiche medie degli oggetti diffusi direttamente dalla pila standard di immagini al microscopio ottico senza alcuna necessità di conoscenze preliminari sull'oggetto biologico di interesse e di estrazione di traiettorie a singolo oggetto. Tutte queste informazioni sono racchiuse in un unico output del metodo: una traccia iMSD9 (i dettagli sulla derivazione e l'analisi della traccia iMSD sono forniti nel file supplementare 1).
Il protocollo sperimentale risultante consiste in pochi passaggi. In primo luogo, l'imaging della regione di interesse viene eseguito ad alta risoluzione temporale. Quindi, le funzioni medie di correlazione spazio-temporale vengono calcolate dalla pila di immagini. Infine, per adattamento gaussiano della serie di funzioni di correlazione, la "legge di diffusione" media viene ottenuta direttamente dall'imaging e analizzata per riconoscere la modalità di diffusione dell'oggetto. Il potenziale del metodo è già stato dimostrato per una varietà di oggetti biologici, che vanno dalle molecole alle nanoparticelle e persino interi organelli /strutture subcellulari 9,10,11,12,13,14,15.
Questo documento riporta l'applicazione dell'iMSD ai macropinosomi per evidenziare la loro natura intrinseca e irreversibile in evoluzione temporale in termini di proprietà strutturali e dinamiche medie (cioè a livello di intera popolazione). Inoltre, queste vescicole endocitiche vengono confrontate con le ISG come riferimento per le strutture subcellulari in uno "stato stazionario", cioè uno stato in cui le proprietà strutturali / dinamiche medie dell'intera popolazione di granuli rimangono costanti in qualsiasi momento. La macropinocitosi definisce una serie di eventi iniziati dall'estesa riorganizzazione (o arruffamento) della membrana plasmatica per formare una struttura macropinocitica esterna che viene poi internalizzata16. I macropinosomi in fase iniziale sono molto simili ai fagosomi. Allo stesso tempo, possono essere distinti da altre forme di vescicole endocitiche a causa delle loro caratteristiche di grandi dimensioni, eterogeneità morfologica e mancanza di strutture proteiche-mantello.
I saggi biochimici hanno rivelato che, dopo l'internalizzazione, i macropinosomi si arricchiscono progressivamente di marcatori proteici di altre vie endocitiche, a loro volta, suggerendo che le loro identità cambiano continuamente durante il traffico17. Utilizzando anticorpi contro marcatori noti della via endosomiale, è stato dimostrato che i macropinosomi adottano progressivamente caratteristiche endosomiali classiche: diminuiscono di dimensioni, si sviluppano in strutture endocitiche tardive (ad esempio, lisosomi) o alla fine perdono la loro identità attraverso il recupero mediato dalla membrana di specifici marcatori molecolari (ad esempio, l'ordinamento delle nexine)18,19 . Lo scenario generale è che ogni singolo macropinosoma all'interno della cellula cambia irreversibilmente la sua identità strutturale e dinamica (oltre che molecolare) durante il traffico dalla membrana plasmatica al suo destino intracellulare finale. Di conseguenza, anche le proprietà strutturali/dinamiche/molecolari dell'intera popolazione di macropinosomi stanno cambiando lungo lo stesso percorso temporale. Essendo intrinsecamente sensibile alle proprietà medie dell'intera popolazione di oggetti osservati, il metodo iMSD descrive quantitativamente la "natura evolutiva" mediante la quantificazione di parametri medi chiave, cioè la diffusività locale e il coefficiente anomalo (proprietà dinamiche) e la dimensione media dei macropinosomi (proprietà strutturali) in qualsiasi fase del loro traffico intracellulare.
Per confronto, misurazioni simili sono state eseguite su una ben nota struttura intracellulare racchiusa nella membrana, l'ISG, in un modello di cellule β. Come i macropinosomi, la regolazione delle proprietà strutturali e dinamiche degli ISG, dalla loro genesi alla Rete Trans Golgi (TGN) alla loro esocitosi alla membrana plasmatica, è fondamentale per la corretta esecuzione della funzione ISG20. Tuttavia, a differenza dei macropinosomi, gli ISG vivono in uno "stato stazionario" in cui, in qualsiasi momento, tutti gli stadi funzionali/strutturali/molecolari della durata della vita dell'ISG sono simultaneamente presenti all'interno della cellula, e ciascuno è rappresentato da una specifica sottopopolazione di ISG. Ciò significa che, sebbene ogni singolo granulo si evolva irreversibilmente dalla biogenesi alla secrezione, le proprietà strutturali/dinamiche medie dell'intera popolazione di granuli dovrebbero rimanere costanti in qualsiasi momento (a meno che le condizioni dello stato stazionario non vengano modificate, ad esempio, da stimoli esterni come glucosio, colesterolo e citochine13). Ciò è confermato dall'analisi iMSD.
Protocollo
1. Preparazione del campione
- Prima dell'esperimento di microscopia, le cellule di sottocoltura in piatti adatti per applicazioni di microscopia.
- Lavare due volte un piatto di 10 cm trattato con colture tissutali di cellule confluenti HeLa o INS 1E (insulinoma β-cell-like) con 0,01 M 1x PBS, aggiungere 1 mL di tripsina-EDTA allo 0,05% (1x) e metterlo in un incubatore a 37 °C, umidificato, 5% CO2 per 5 min.
- Risospesciare le celle staccate aggiungendo 9 mL di supporto DMEM completo (per celle HeLa) o RPMI 1640 (per celle INS-1E) e raccogliere i 10 mL finali in tubi centrifughi.
- Seminare circa 2 × 105 celle in ogni piatto di 35 mm x 10 mm in un volume finale di 1 mL del mezzo. Incubare le cellule per 24 ore a 37 °C e 5% di CO2.
- Per etichettare fluorescentmente i lisosomi, utilizzare LysoTracker Red DND-99.
- Diluire la soluzione madre in 1 mL di terreno preriscaldato fino ad una concentrazione finale di colorante di 70 nM.
- Sostituire il mezzo del piatto con un mezzo fresco contenente LysoTracker. Incubare le cellule in un mezzo contenente LysoTracker per 20 minuti a 37 °C in un'atmosfera di CO2 al 5% e lavarle due volte con mezzo fresco prima dell'esperimento.
- Per etichettare fluorescentemente i macropinosomi, utilizzare 70-kDa fluoresceina isotiocianato-destrano. Lavare le cellule in sottocoltura tre volte con 0,01 M 1x soluzione salina tamponata con fosfato (PBS), sostituirle con mezzo contenente destrano (1 mg/mL) e incubare a 37 °C per 30 min. Prima di procedere con l'esperimento di microscopia, lavare le cellule tre volte con mezzo fresco.
- Per etichettare fluorescentemente gli ISG nelle cellule INS-1E, trasfettare le cellule usando il reagente di trasfezione (vedi la Tabella dei materiali) e il plasmide13 della proteina fluorescente verde potenziata con peptide C (EGFP) secondo il protocollo del produttore e incubare a 37 ° C per 24 ore in un'atmosfera di CO2 al 5% prima dell'esperimento.
2. Acquisizione dati
- Per consentire al microscopio di equilibrarsi alla temperatura e all'atmosfera desiderate, accendere il sistema di controllo dell'incubatore del microscopio almeno 2 ore prima dell'esperimento.
NOTA: ogni acquisizione è una serie time-lapse. - Acquisisci le immagini utilizzando un microscopio confocale invertito dotato di un obiettivo ad immersione in acqua ad apertura numerica (NA) 60x, 1.2.
- Utilizzare un laser Argon a 488 nm per l'eccitazione di EGFP (cellule trasfettate) e macropinosomi marcati con fluoresceina. Raccogliere l'emissione di fluorescenza tra 500 e 600 nm utilizzando un rilevatore a tubi fotomoltiplicatore standard.
- Utilizzare un laser HeNe da 543 nm per eccitare Lysotracker e raccogliere la sua emissione di fluorescenza tra 555 e 655 nm.
- Impostare il diametro del foro stenopeico di rilevamento sulla dimensione di 1 Airy. Per ogni acquisizione, raccogli una serie di 1000 fotogrammi sequenziali. Impostare il tempo di permanenza dei pixel su 2 μs/pixel per un tempo di fotogramma di 129 ms.
NOTA: ogni fotogramma era composto da 256 x 256 pixel (16 bit/pixel) con una dimensione fisica di 69 nm/pixel, corrispondente approssimativamente a un'area di 17 μm x 17 μm.
3. iCalcolo MSD
NOTA: per eseguire correttamente il calcolo, utilizzare un software in grado di calcolare numericamente e programmare script. Lo script specifico (ad esempio, per il file di script 'iMSD.m', vedere File di supporto 1) deve essere presente nella stessa directory contenente la serie di immagini da elaborare. Ogni immagine della serie deve essere salvata come un file ".tif" distinto.
- Per inizializzare correttamente i parametri strumentali utilizzati per le acquisizioni, aprire iMSD.m con l'editor di testo del software e modificarne la prima sezione come segue.
- Impostare N come numero di fotogrammi nella serie temporale (ad esempio, 1000 in questo protocollo).
- Impostare px_size: dimensione dei pixel, espressa in μm (ad esempio, 0,069 in questo protocollo).
- Set f: risoluzione temporale di ogni fotogramma, espressa in secondi (ad esempio, 0,129 in questo protocollo).
- Imposta filtro: input binario per la correzione dello sfondo, imposta il valore su '0' per elaborare le immagini raw o imposta il valore su '1' per eseguire una sottrazione di sfondo basata su soglie.
- Impostare av_toll: soglia per la correzione dello sfondo; qualsiasi pixel con intensità inferiore a questo valore verrà impostato su 0 se Filtro=1.
- Impostare bit come numero intero che determina il campionamento dell'intensità (ad esempio, 8 bit, 16 bit).
- Salvare ed eseguire il file di script iMSD.m modificato.
- Controllare l'esecuzione dello script.
NOTA: lo stato di elaborazione del calcolo può essere controllato nella finestra di comando; se si verifica un problema irreversibile, il processo verrà interrotto e verrà visualizzato un messaggio di avviso per mostrare il tipo di errore e il codice riga correlato. In caso contrario, seguire i passaggi 3.3.1-3.3.3.- Importare la pila di immagini e sottrarre lo sfondo (se necessario).
- Calcolare la funzione di correlazione spaziotemporale G(ξ,η,τ) usando il metodo di Fourier.
- Adatta la funzione di correlazione spaziotemporale con una funzione gaussiana 2D.
NOTA: al termine del processo, i valori di uscita delle procedure di raccordo σ2(τ) verranno visualizzati nella finestra di comando: vengono riportate le medie, l'errore e la corrispondente bontà (R2) del raccordo.
- Controllare l'output grafico.
NOTA: la curva iMSD e le corrispondenti curve di raccordo sono mostrate in tre pannelli separati, ciascuno per un diverso tipo di equazione di raccordo utilizzata: diffusione browniana, diffusione anomala o diffusione confinata. Per ogni curva, il valore R2 è riportato nella legenda del grafico. - Controllare l'output del testo.
NOTA: al termine del processo, viene creato un file ".xls" con lo stesso nome del file ".tif" time-lapse originale. Il primo foglio contiene i valori di ξ, η, τ e σ2 calcolati per ogni ritardo temporale. I parametri di ingresso e i principali valori di uscita calcolati sono riportati nel secondo foglio, ovvero coefficiente di diffusione, coefficiente anomalo e intercetta σ20 dell'asse y.
Risultati
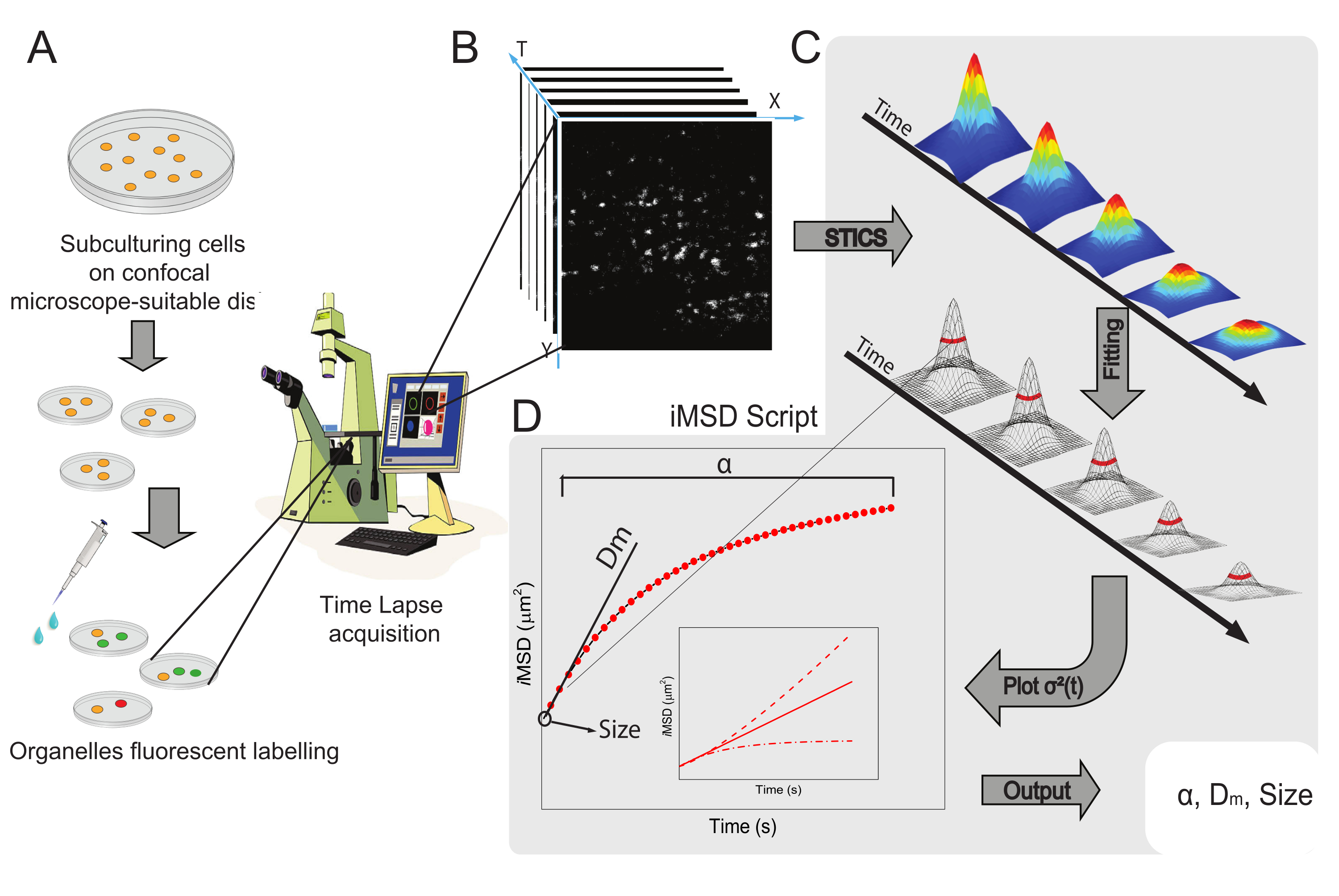
Il flusso di lavoro generale del metodo è illustrato nella Figura 1. Ricapitola le principali fasi presentate nella sezione protocollo, dalla preparazione del campione (Figura 1A) all'imaging time-lapse di nanostrutture intracellulari (Figura 1B), all'analisi della fluttuazione per il calcolo della serie di funzioni di correlazione spaziotemporale (Figura 1C), e all'adattamento per la derivazione delle proprietà strutturali/dinamiche medie dell'oggetto in studio (Figura 1D).
Un parametro critico è la risoluzione temporale adottata per l'imaging dell'oggetto subcellulare di interesse. Questo valore sperimentale imposterà la soglia temporale alla quale verrà misurato lo spostamento medio minimo degli oggetti di interesse. Tuttavia, la condizione preferita è l'impostazione di una risoluzione temporale dell'imaging in cui l'oggetto di interesse appare "immobile" all'interno del fotogramma catturato, cioè mostra una dimensione caratteristica che, in media, non è deformata a causa della velocità di imaging. Ciò è tecnicamente possibile se l'oggetto di interesse è una struttura subcellulare chiusa a membrana o un organello (come in questo caso). Tipicamente, le strutture subcellulari presentano coefficienti di diffusione locale (D, μm2/s, vedi Tabella 1) che sono di diversi ordini di grandezza inferiori a quelli delle singole molecole isolate nel citoplasma (ad esempio, GFP21). La convalida può essere eseguita immobilizzando artificialmente l'organello di interesse (ad esempio, mediante fissazione chimica). In effetti, questa condizione può servire come riferimento per determinare la dimensione effettiva dell'organello nelle condizioni sperimentali utilizzate (ad esempio, lunghezza d'onda di eccitazione, dimensione dei pixel, obiettivo).
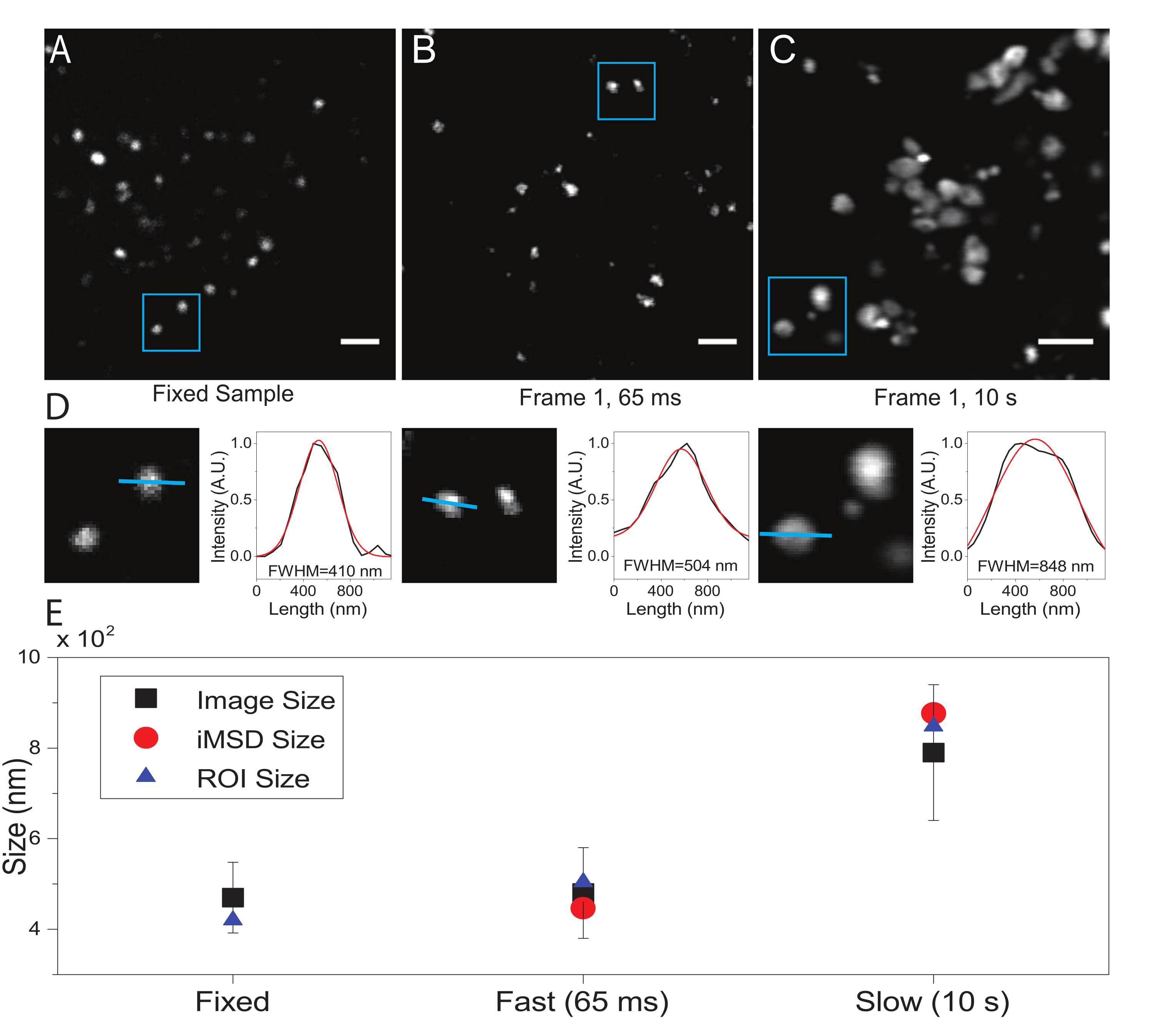
Qui, sebbene i lisosomi siano stati usati come organelli di prova per questa procedura, i risultati sono validi indipendentemente dalla struttura target. La Figura 2A mostra un'immagine di un lisosoma fisso (cioè immobile) insieme a un'acquisizione eseguita su cellule vive alla risoluzione temporale appropriata (cioè, tipicamente inferiore a 100 ms/frame; ad esempio, 65 ms/frame nell'esempio in Figura 2B) e un'acquisizione eseguita intenzionalmente a una risoluzione temporale molto bassa (ad esempio, 10 s/frame nell'esempio nella Figura 2C ). Per ogni condizione, la dimensione dell'oggetto diffusore viene estratta come segue: i) un profilo di intensità dello spot è derivato dallo strumento linea nel software ImageJ; ii) il profilo di intensità viene tracciato e interpolato da una funzione gaussiana per calcolare l'intera larghezza a metà massimo (FWHM) valore che, a sua volta, viene utilizzato come stima del diametro dello spot (Figura 2D). Come previsto e mostrato nel grafico della Figura 2E, l'acquisizione ad altissima risoluzione temporale (cioè 65 ms/frame) produce una dimensione media della struttura vicina a quella ottenuta nel campione fisso, sia utilizzando lo strumento standard sopra descritto sia estraendo l'intercetta dell'asse y iMSD. Invece, l'acquisizione a bassa velocità produce un aumento delle dimensioni apparenti della struttura, a causa della dinamica naturale della struttura durante l'imaging. In condizioni sperimentali non ottimali, le informazioni strutturali/dinamiche estratte non riflettono fedelmente le proprietà intrinseche dell'oggetto in studio.
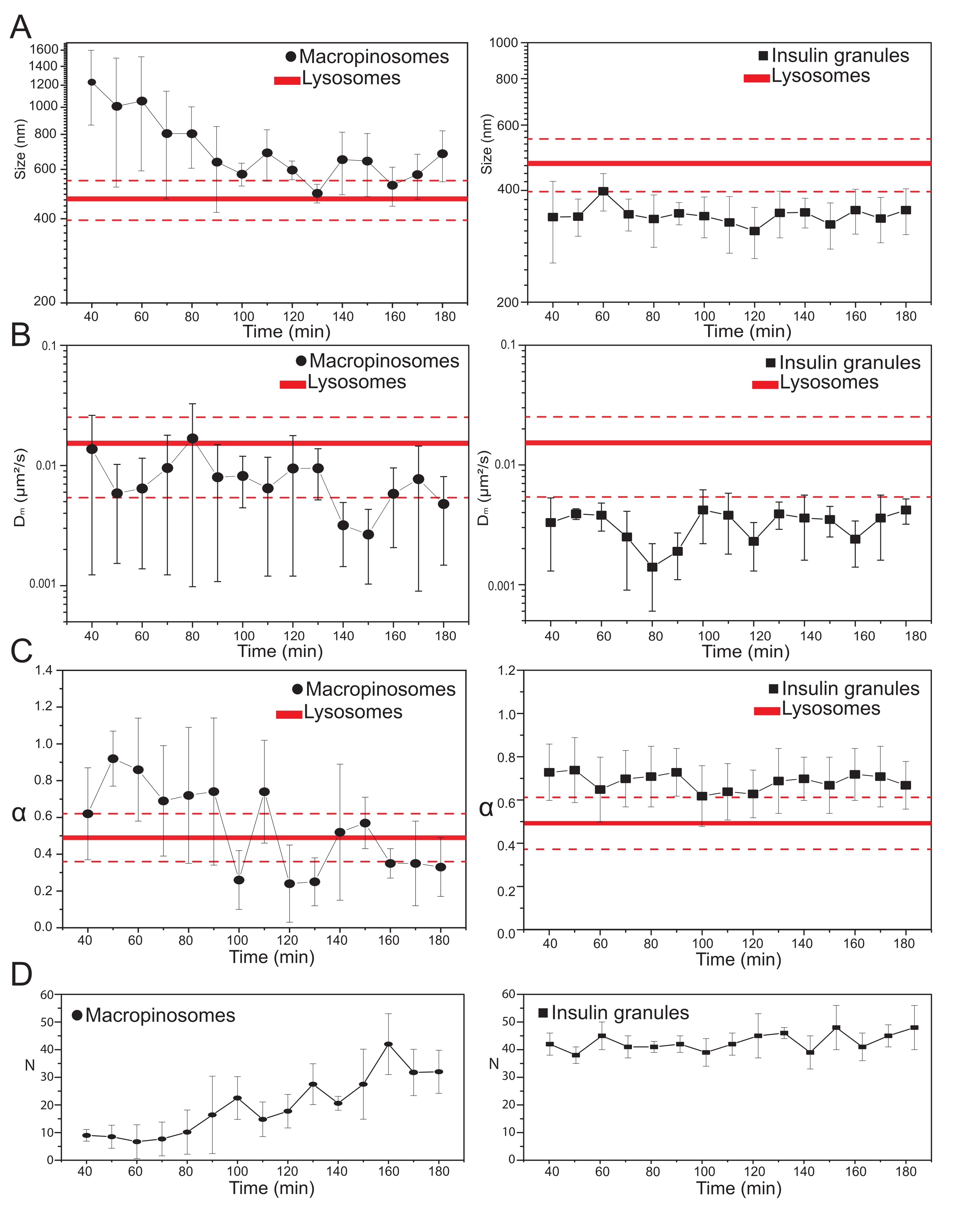
Una volta selezionati i principali parametri sperimentali, è possibile produrre set di dati per le strutture intracellulari target. Per i macropinosomi, dopo 20 minuti di incubazione delle cellule con 70 kDa dextrans, le serie temporali delle strutture intracellulari marcate sono state acquisite in diversi punti temporali dopo il trattamento, da 30 min fino a circa 180 min. È interessante notare che durante il traffico viene rilevato un graduale cambiamento nelle proprietà strutturali e dinamiche dei macropinosomi (Figura 3; le distribuzioni di σ02, Dm, α e N per i macropinosomi sono riportate nei grafici a sinistra). Mentre durante il traffico non vengono rilevati cambiamenti evidenti nella diffusività locale (Dm) dei macropinosomi, sia la dimensione caratteristica (σ02) che la modalità generale di movimento (α) si evolvono nel tempo.
Di particolare rilievo, durante il traffico si osserva una diminuzione della dimensione media dei macropinosomi (Figura 3A, a sinistra), insieme a un concomitante aumento della natura subdiffusiva del loro moto (cioè, indicato come una diminuzione dei valori α, Figura 3C, pannello di sinistra). Inoltre, il numero di macropinosomi è stato estratto da ogni acquisizione: i risultati, riportati nella Figura 3D, pannello di sinistra, rivelano chiaramente un aumento del numero di macropinosomi nel tempo. Tutti questi risultati sono in buon accordo con le aspettative perché si suppone che i macropinosomi marcati con destrano abbiano origine come vescicole isolate e di grandi dimensioni racchiuse nella membrana plasmatica (che sono anche competenti per il movimento lungo i componenti citoscheletrici) ma dovrebbero comunicare gradualmente con la via endo-lisosomiale costituita da una grande popolazione di strutture più piccole e casualmente diffuse.
Come anticipato sopra, i risultati per i macropinosomi sono in contrasto con misurazioni simili eseguite sugli ISG (Figura 3, colonna di destra). I granuli di insulina non mostrano un andamento in evoluzione temporale dei parametri strutturali/dinamici derivati dall'iMSD (e il loro numero medio all'interno della cellula) nella stessa finestra temporale osservata per i macropinosomi. Inoltre, i valori caratteristici di σ02, Dm e α sono molto diversi da quelli dei lisosomi, usati di nuovo come riferimento. Questo risultato conferma l'idea, come anticipato sopra, che i granuli siano sondati in uno "stato stazionario" in cui, in qualsiasi momento, le proprietà strutturali/dinamiche medie dell'intera popolazione di ISG sono invariate (cioè rimangono costanti, a meno che le condizioni dello stato stazionario non cambino, ad esempio, a causa di stimoli esterni).

Figura 1: Flusso di lavoro sperimentale. (A) Le cellule sono state placcate 24 ore (48 ore per esperimenti di trasfezione) prima di esperimenti confocali su piastre cellulari adatte per applicazioni microscopiche. Le cellule sono state poi opportunamente trattate secondo il metodo di etichettatura (vedi protocollo) per colorare l'organello citoplasmatico di interesse. (B) Una tipica acquisizione confocale consiste in una pila di immagini (time-lapse) di una porzione citoplasmatica di una cellula vivente, che descrive l'evoluzione temporale della dinamica degli organelli marcati. (C) Il filmato time-lapse viene analizzato con uno script Matlab personalizzato, calcolando prima la funzione di correlazione dell'immagine spaziotemporale e il fitting gaussiano per tracciare le curve iMSD (D) e i relativi parametri di fitting estratti che descrivono i parametri di dinamica strutturale degli organelli ripresi. Abbreviazioni: iMSD = spostamento quadrato medio derivato dall'imaging; STICS = spettroscopia di correlazione delle immagini spaziotemporali; α = coefficiente di diffusione anomalo; Dm = diffusività locale; σ2(τ) = varianza. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 2: Parametri sperimentali adeguati. (A) Immagine esemplare di lisosomi colorati in un campione fisso. Barra della scala = 2 μm. (B) Il primo fotogramma di una pila di immagini di lisosomi colorati in una cella vivente, acquisite con i parametri appropriati. Risoluzione temporale: 65 ms/frame. Barra di scala = 2 μm. (C) Il primo fotogramma di una pila di immagini di lisosomi colorati in una cellula vivente, acquisite a bassa velocità: è visibile la deformazione artefattuale della dimensione apparente del lisosoma dovuta al movimento degli organelli durante l'imaging. Risoluzione temporale: 10 s/frame. Barra di scala = 2 μm. (D) Esempio di calcolo delle dimensioni per i lisosomi ripresi in un ROI blu di (A), (B) e (C). Il profilo di intensità lungo la linea blu è stato dotato di una funzione gaussiana per recuperare il FWHM, cioè una stima delle dimensioni dello spot. I valori FWHM sono riportati per ciascun raccordo. (E) Rappresentazione grafica dei valori dimensionali ottenuti mediante l'analisi delle immagini descritta nel pannello (D) per tutti i lisosomi ripresi (quadrato nero, valore medio e deviazione standard), per i lisosomi racchiusi all'interno del ROI blu (triangolo blu) e recuperati dall'analisi iMSD (cerchio rosso). Questa cifra è da 22. Abbreviazioni: ROI = regione di interesse; FWHM = larghezza intera a metà massimo; iMSD = spostamento quadrato medio derivato dall'imaging. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 3: Evoluzione temporale degli organelli marcati. (A) Grafici dei valori dimensionali estratti da iMSD rispetto alla progressione temporale delle acquisizioni successive rappresentata come media dei valori misurati nelle acquisizioni eseguite su una finestra temporale di 10 minuti. A sinistra, progressiva riduzione della dimensione media dei macropinosomi (cerchi neri) e a destra, la dimensione invariante nel tempo dei granuli secretori di insulina (quadrati neri) rispetto ai lisosomi, rappresentata come valore medio di dimensione (linea rossa più spessa) ± deviazione standard (linee rosse tratteggiate). (B) e (C) Progressione temporale dei coefficienti Dm e α per i macropinosomi (a sinistra) e i granuli di insulina (a destra) estratti mediante analisi iMSD. (D) Evoluzione temporale del numero di macropinosomi marcati e granuli di insulina misurati nel primo fotogramma di ogni film time-lapse acquisito. Abbreviazioni: iMSD = spostamento quadrato medio derivato dall'imaging; α = coefficiente di diffusione anomalo; Dm = diffusività locale, N = numero. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
| Organello | Etichettatura | Linea cellulare | Dimensione (nm) | Dm ( × 10-3 μm2/s) | α | N | Ref. | ||
| Endosoma precoce (EE) | CellLight Endosoma precoce GFP | Hela | 395±74 | 3.0±2.4 | 1.02±0.20 | 40 | 10 | ||
| Endosoma tardivo (LE) | CellLight Endosoma tardivo GFP | Hela | 693±102 | 15.4±10.6 | 0.57±0.16 | 58 | 10 | ||
| Lisosoma (LY) | LysoTracker DND-99 | Hela | 471±76 | 15,3±9,0 | 0.49±0.13 | 143 | 10, 14 | ||
| Caveola (CAV) | Caveolin-EGFP | Hela | 405±49 | 3.1±1.8 | 1.00±0.22 | 15 | 10 | ||
| Vescicola rivestita di clatrina (CCV) | Transferrin-Alexa 488 | Hela | 513±62 | 16.2±9.9 | 0.48±0.17 | 33 | 10 | ||
| Granulo di insulina (IG) | C-peptide-EGFP | INS-1E · | 335±56 | 3.0±1.7 | 0,70±0.14 | 107 | 11 | ||
| Macropinosoma precoce (EMCR) | Fluoresceina-Destrano 70 kDa | Hela | 979±423 | 8.3±9 | 0.79±0.27 | 36 | 10 | ||
| Macropinosoma intermedio (IMCR) | Fluoresceina-Destrano 70 kDa | Hela | 702±180 | 13.7±19.9 | 0,60±0.38 | 29 | 10 | ||
| Macropinosoma tardivo (LMCR) | Fluoresceina-Destrano 70 kDa | Hela | 592±127 | 5.8±4.7 | 0.39±0.21 | 21 | 10 | ||
Tabella 1: Parametri strutturali e dinamiciestratti da MSD. La tabella mostra i valori di dimensione, Dm e coefficiente di α misurati per diversi organelli, specificando le strategie di etichettatura, la linea cellulare utilizzata e il numero di acquisizioni analizzate. I valori sono riportati come deviazione media ± standard. Abbreviazioni: iMSD = spostamento quadrato medio derivato dall'imaging; α = coefficiente di diffusione anomalo; Dm = diffusività locale, N = numero; GFP = proteina fluorescente verde; EGFP = proteina fluorescente verde potenziata.
File supplementare 1: Dettagli sulla derivazione e l'analisi delle tracce iMSD. Fare clic qui per scaricare questo file.
File di supporto 1: Fare clic qui per scaricare questo file.
Discussione
Le proprietà e i vantaggi dell'iMSD sono evidenti se confrontati con le tecniche disponibili per recuperare informazioni analoghe. Per le informazioni strutturali, la scelta preferita è l'analisi al microscopio elettronico a trasmissione (TEM). Con questo metodo, i dettagli ultrastrutturali a risoluzione molecolare e anche oltre possono essere recuperati, anche per le nanostrutture subcellulari. Tuttavia, la peculiare risoluzione spaziale di TEM è ottenuta a scapito delle informazioni nella dimensione temporale, che è di interesse qui. Per compensare questo, i recenti progressi nelle tecnologie di imaging delle cellule vive sono di particolare interesse. Questi includono nuovi marcatori fluorescenti con prestazioni migliorate (ad esempio, luminosità e fotostabilità), procedure di etichettatura ottimizzate e rilevatori più sensibili. Inoltre, sono disponibili strumenti analitici per affrontare sia la struttura (ad esempio, "dimensione" mediante analisi basata su fasor della spettroscopia di correlazione di immagini locali, PLICS23, aggregazione / oligomerizzazione mediante analisi Number&Brightness24) che dinamica (ad esempio, legge di diffusione mediante tracciamento di singole particelle, cioè (SPT)25,26,27,28 ) parametri su scala subcellulare. Il metodo SPT consente l'accesso diretto alla traiettoria dell'oggetto e al suo MSD. Tuttavia, lo svantaggio è la necessità di una bassa densità della sonda e di etichette molto luminose e di molte traiettorie a singolo oggetto da misurare per soddisfare criteri statistici. Per quanto riguarda la risoluzione temporale della misura, le sonde inorganiche e fotostabili (ad esempio, punti quantici o nanoparticelle metalliche) possono aumentare le prestazioni SPT ma a scapito di complesse procedure di produzione ed etichettatura.
Rispetto a questi standard, il metodo iMSD qui descritto mostra alcuni vantaggi chiave. In primo luogo, questo approccio può essere utilizzato in combinazione con tag fluorescenti relativamente deboli, come le proteine fluorescenti geneticamente codificate (ad esempio, l'applicazione agli ISG). Pertanto, rispetto all'SPT, si ottiene una risoluzione temporale più elevata (utilizzando la stessa etichetta) a causa della minore quantità di fotoni richiesta8. In secondo luogo, il metodo iMSD è limitato solo dalla risoluzione temporale ma non dalla diffrazione. Infatti, nonostante il setup ottico limitato alla diffrazione utilizzato, è possibile misurare spostamenti molecolari medi anche al di sotto del limite di diffrazione, come già dimostrato per i flussi molecolari utilizzando STICS29. La risoluzione effettiva nella misurazione degli spostamenti dipende da quanto accuratamente (in termini di segnale al rumore) può essere misurata la funzione di correlazione, spiegando così perché non è limitata dalla diffrazione. Pertanto, appare chiaro che lo spostamento minimo che può essere misurato dipende dalla diffusività dell'oggetto di interesse e dalla risoluzione temporale della configurazione dell'imaging.
A questo proposito, è importante considerare che l'applicazione a nanostrutture subcellulari, come macropinosomi o granuli di insulina, con un microscopio a scansione laser è ottimale: la velocità di scansione disponibile è significativamente superiore alla dinamica dell'oggetto di interesse. In tal caso, il movimento degli oggetti durante l'acquisizione è trascurabile e la funzione di correlazione può essere approssimata da una funzione gaussiana. Infine, l'approccio iMSD può essere facilmente applicato a una vasta gamma di configurazioni di microscopia ottica commerciale basate sulla scansione raster o sull'imaging basato su telecamere ad ampio campo, senza necessità di calibrazione del sistema (necessaria solo se è necessario ottenere una stima accurata delle dimensioni delle particelle). Un parametro importante per il funzionamento del metodo è il corretto campionamento spaziale. Come regola generale, per raggiungere una convergenza soddisfacente dell'algoritmo di fitting, la dimensione minima della regione di interesse per l'imaging dovrebbe essere almeno 3 volte maggiore dello spostamento massimo di interesse.
In conclusione, il metodo iMSD richiede solo un microscopio attrezzato per un'acquisizione rapida. La struttura di interesse può essere etichettata su qualsiasi fluoroforo geneticamente codificato o organico, consentendo così l'imaging multicanale. Si prevede che l'analisi cross-iMSD sarà utilizzata nel prossimo futuro per selezionare sottopopolazioni di nanostrutture subcellulari e rivelare le loro interazioni e co-diffusione all'interno della cellula, quest'ultimo è un argomento caldo nella biofisica cellulare. Se qualche dettaglio viene perso dall'analisi iMSD, questo è certamente correlato alla grande quantità di informazioni molecolari all'interno di nanostrutture subcellulari dinamiche. Tali informazioni sono inevitabilmente mediate durante la misurazione a causa della scarsa risoluzione temporale. Teoricamente, tuttavia, non esiste un limite tecnico dovuto alla possibilità di recuperare informazioni molecolari, a condizione che si possano raggiungere velocità di acquisizione sufficienti8. A causa dei continui miglioramenti nella velocità / sensibilità del rivelatore e nelle tecnologie di imaging, si prevede che le informazioni sull'intero compartimento subcellulare e sui suoi costituenti molecolari saranno estratte da un singolo set di dati.
Divulgazioni
Gli autori non hanno alcun conflitto di interessi da dichiarare.
Riconoscimenti
Questo lavoro ha ricevuto finanziamenti dal Consiglio europeo della ricerca (CER) nell'ambito del programma di ricerca e innovazione Horizon 2020 dell'Unione europea (convenzione di sovvenzione n. 866127, progetto CAPTUR3D).
Materiali
| Name | Company | Catalog Number | Comments |
| 100x Penicillin-Streptomycin-Glutamine | Gibco | 10378-016 | Cell medium supplement |
| C-peptide-EGFP | Plasmid | ||
| DMEM High Glucose | Gibco | 31053028 | Cell medium (HeLa) |
| FBS | Gibco | 10082147 | Cell medium supplement |
| Fluorescein isothiocyanate-dextran 70 kDa | Sigma Aldrich | 46945-100MG-F | Reagent |
| HeLa | ATCC | CCL-61 | Cell Line |
| Lipofectamine 2000 | TermoFisher | 11668019 | Trasfection reagent |
| Lysotracker Red DND-99 | Gibco | L7528 | Reagent |
| Matlab | MathWork | Software | |
| Microscope-suitable cell dishes | Willco | GWSt-3522 | Petri dishes |
| Olympus FV1000 | Olympus Japan | Confocal microscope | |
| RPMI 1640 | Gibco | 11835063 | Cell medium (INS-1E) |
Riferimenti
- Murphy, J. E., Padilla, B. E., Hasdemir, B., Cottrell, G. S., Bunnett, N. W. Endosomes: a legitimate platform for the signaling train. Proceedings of the National Academy of Sciences of the United States of America. 106 (42), 17615-17622 (2009).
- Mosesson, Y., Mills, G. B., Yarden, Y. Derailed endocytosis: an emerging feature of cancer. Nature Reviews Cancer. 8 (11), 835-850 (2008).
- Mellman, I., Yarden, Y. Endocytosis and cancer. Cold Spring Harbor Perspectives in Biology. 5 (12), 016949 (2013).
- Di Fiore, P. P. Endocytosis, signaling and cancer, much more than meets the eye. Preface. Molecular Oncology. 3 (4), 273-279 (2009).
- Bogan, J. S., Xu, Y., Hao, M. Cholesterol accumulation increases insulin granule size and impairs membrane trafficking. Traffic. 13 (11), 1466-1480 (2012).
- Ballabio, A., Gieselmann, V. Lysosomal disorders: From storage to cellular damage. Biochimica et Biophysica Acta. 1793 (4), 684-696 (2009).
- Hu, Y. -. B., Dammer, E. B., Ren, R. -. J., Wang, G. The endosomal-lysosomal system: from acidification and cargo sorting to neurodegeneration. Translational Neurodegeneration. 4, 18 (2015).
- Di Rienzo, C., Gratton, E., Beltram, F., Cardarelli, F. Spatiotemporal fluctuation analysis: a powerful tool for the future nanoscopy of molecular processes. Biophysical Journal. 111 (4), 679-685 (2016).
- Di Rienzo, C., Gratton, E., Beltram, F., Cardarelli, F. Fast spatiotemporal correlation spectroscopy to determine protein lateral diffusion laws in live cell membranes. Proceedings of the National Academy of Sciences of the United States of America. 110 (30), 12307-12312 (2013).
- Di Rienzo, C., Gratton, E., Beltram, F., Cardarelli, F. From fast fluorescence imaging to molecular diffusion law on live cell membranes in a commercial microscope. Journal of Visualized Experiments: JoVE. (92), e51994 (2014).
- Digiacomo, L., et al. Dynamic fingerprinting of sub-cellular nanostructures by image mean square displacement analysis. Scientific Reports. 7 (1), 14836 (2017).
- Durso, W., et al. Lysosome dynamic properties during neuronal stem cell differentiation studied by spatiotemporal fluctuation spectroscopy and organelle tracking. International Journal of Molecular Sciences. 21 (9), 3397 (2020).
- Ferri, G., et al. Insulin secretory granules labelled with phogrin-fluorescent proteins show alterations in size, mobility and responsiveness to glucose stimulation in living β-cells. Scientific Reports. 9 (1), 2890 (2019).
- Durso, W., D'Autilia, F., Amodeo, R., Marchetti, L., Cardarelli, F. Probing labeling-induced lysosome alterations in living cells by imaging-derived mean squared displacement analysis. Biochemical and Biophysical Research Communications. 503 (4), 2704-2709 (2018).
- Digiacomo, L., Digman, M. A., Gratton, E., Caracciolo, G. Development of an image Mean Square Displacement (iMSD)-based method as a novel approach to study the intracellular trafficking of nanoparticles. Acta Biomaterialia. 42, 189-198 (2016).
- Swanson, J. A., Watts, C. Macropinocytosis. Trends in Cell Biology. 5 (11), 424-428 (1995).
- Jones, A. T. Macropinocytosis: searching for an endocytic identity and role in the uptake of cell penetrating peptides. Journal of Cellular and Molecular Medicine. 11 (4), 670-684 (2007).
- Falcone, S., et al. Macropinocytosis: regulated coordination of endocytic and exocytic membrane traffic events. Journal of Cell Science. 119, 4758-4769 (2006).
- Kerr, M. C., et al. Visualisation of macropinosome maturation by the recruitment of sorting nexins. Journal of Cell Science. 119, 3967-3980 (2006).
- Rorsman, P., Renstrom, E. Insulin granule dynamics in pancreatic beta cells. Diabetologia. 46 (8), 1029-1045 (2003).
- Di Rienzo, C., Piazza, V., Gratton, E., Beltram, F., Cardarelli, F. Probing short-range protein Brownian motion in the cytoplasm of living cells. Nature Communications. 5 (1), 5891 (2014).
- Ferri, G., et al. Time-lapse confocal imaging datasets to assess structural and dynamic properties of subcellular nanostructures. Scientific Data. 5 (1), 180191 (2018).
- Scipioni, L., Gratton, E., Diaspro, A., Lanzanò, L. Phasor analysis of local ICS detects heterogeneity in size and number of intracellular vesicles. Biophysical Journal. 111 (3), 619-629 (2016).
- Digman, M. A., Dalal, R., Horwitz, A. F., Gratton, E. Mapping the number of molecules and brightness in the laser scanning microscope. Biophysical Journal. 94 (6), 2320-2332 (2008).
- Li, C. H., Bai, L., Li, D. D., Xia, S., Xu, T. Dynamic tracking and mobility analysis of single GLUT4 storage vesicle in live 3T3-L1 cells. Cell Research. 14 (6), 480-486 (2004).
- Donovan, K. W., Bretscher, A. Tracking individual secretory vesicles during exocytosis reveals an ordered and regulated process. Journal of Cell Biology. 210 (2), 181-189 (2015).
- Westphal, V., et al. Video-rate far-field optical nanoscopy dissects synaptic vesicle movement. Science. 320 (5873), 246-249 (2008).
- Tabei, S. M. A., et al. Intracellular transport of insulin granules is a subordinated random walk. Proceedings of the National Academy of Sciences of the United States of America. 110 (13), 4911-4916 (2013).
- Hebert, B., Costantino, S., Wiseman, P. W. Spatiotemporal image correlation spectroscopy (STICS) theory, verification, and application to protein velocity mapping in living CHO cells. Biophysical Journal. 88 (5), 3601-3614 (2005).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneEsplora altri articoli
This article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati