
Method Article
משימת תכנון חזותי-מרחבי בשילוב עם מערכות מעקב עיניים ואלקטרואנצפלוגרמה
In This Article
Summary
חקר התכנון הקוגניטיבי המשלב EEG ומערכות מעקב עיניים מספק גישה רב-מודאלית לחקירת המנגנונים העצביים המתווכים שליטה קוגניטיבית והתנהגות מכוונת מטרה בבני אדם. כאן, אנו מתארים פרוטוקול לחקירת התפקיד של תנודות מוח ותנועות עיניים בתכנון ביצועים.
Abstract
תהליך התכנון, המאופיין ביכולת לגבש תוכנית מסודרת להשגת מטרה, חיוני להתנהגות אנושית מוכוונת מטרה. מכיוון שהתכנון נפגע במספר הפרעות נוירו-פסיכיאטריות, יישום בדיקות קליניות וניסיוניות מתאימות לבחינת התכנון הוא קריטי. בשל אופי פריסת התכנון, בה משתתפים מספר תחומים קוגניטיביים, הערכת התכנון ועיצוב פרדיגמות התנהגותיות בשילוב עם שיטות הדמיה מוחית הם אתגרים עכשוויים במדעי המוח הקוגניטיביים. מטלת תכנון הוערכה בשילוב עם מערכת אלקטרואנצפלוגרמה (EEG) ורישומי תנועות עיניים ב-27 משתתפים בוגרים בריאים. ניתן להפריד את התכנון לשני שלבים: שלב תכנון מנטלי בו מיוצג רצף שלבים פנימי ושלב ביצוע בו משתמשים בפעולה מוטורית להשגת מטרה שתוכננה בעבר. הפרוטוקול שלנו כלל משימת תכנון ומשימת בקרה. משימת התכנון כללה פתרון של 36 ניסויי מבוך, שכל אחד מהם מייצג מפה של גן חיות. למשימה היו ארבע תקופות: 1) תכנון, שבו הנבדקים הונחו לתכנן נתיב לביקור במיקומים של ארבעה בעלי חיים על פי מערכת כללים; 2) תחזוקה, שבה הנבדקים היו צריכים לשמור את הנתיב המתוכנן בזיכרון העבודה שלהם; 3) ביצוע, שבו הנבדקים השתמשו בתנועות עיניים כדי לעקוב אחר הנתיב שתוכנן מראש כפי שצוין על ידי מערכת מעקב העיניים; ו-4) תגובה, שבה הנבדקים דיווחו על סדר בעלי החיים שביקרו בהם. למשימת הבקרה היה מבנה דומה, אך מרכיב התכנון הקוגניטיבי הוסר על ידי שינוי מטרת המשימה. הדפוסים המרחביים והזמניים של ה-EEG גילו כי תכנון גורם לעלייה הדרגתית ומתמשכת בפעילות התטא הקדמית-קו האמצע (FMθ) לאורך זמן. מקור הפעילות הזו זוהה בקליפת המוח הקדם-מצחית על ידי ניתוחי מקורות. התוצאות שלנו הצביעו על כך שהפרדיגמה הניסויית המשלבת EEG ומערכות מעקב עיניים הייתה אופטימלית להערכת תכנון קוגניטיבי.
Introduction
במהלך 10 השנים האחרונות נערך מחקר מקיף לבחינת תפקידה של דינמיקה עצבית תנודתית הן על קוגניציה והן על התנהגות. מחקרים אלה קבעו כי אינטראקציות ספציפיות לתדר בין אזורים מיוחדים ונרחבים בקליפת המוח ממלאות תפקיד מכריע בקוגניציה ובשליטה קוגניטיבית 1,2,3. גישה זו מדגישה את האופי הקצבי של פעילות המוח, המסייעת לתאם דינמיקה קליפת המוח בקנה מידה גדול ועומדת בבסיס עיבוד קוגניטיבי והתנהגות מוכוונת מטרה 4,5. ישנן ראיות משמעותיות המראות כי תנודות קצביות במוח מעורבות בתהליכים קוגניטיביים שונים, כולל תפיסה6, קשב 7,8,9, קבלת החלטות10, הפעלה מחדש של זיכרון11, זיכרון עבודה12 ושליטה קוגניטיבית13. מנגנוני תנודה שונים הוצעו להנחיית התנהגות מוכוונת מטרה, כאשר רשתות חולפות בקנה מידה גדול ספציפיות לתדר מספקות מסגרת לעיבוד קוגניטיבי 1,14,15. לדוגמה, ממצאים אחרונים מצביעים על כך שרצועות תדרים ספציפיות במוח עשויות לשקף מנגנון משוב המווסת את פעילות הזינוק, ומספק מסגרת ייחוס זמנית לתיאום ריגוש קליפת המוח ותזמון ספייק לייצור התנהגות 16,17,18. סקירה מסופקת על ידי הלפריך ונייט19.
גוף הראיות הזה מעלה שאלות לגבי האופן שבו קליפת המוח הקדם-מצחית (PFC) מקודדת הקשרים של משימות תכנון וכללים רלוונטיים מבחינה התנהגותית. קליפת המוח הקדם-מצחית נחשבת זה מכבר כתומכת בשליטה קוגניטיבית ובהתנהגות מוכוונת מטרה באמצעות דפוסי התנודות של הפעילות העצבית שהיא יוצרת, הטיה סלקטיבית של הפעילות העצבית באזורי מוח מרוחקים ושליטה בזרימת המידע ברשתות עצביות בקנה מידה גדול20. בנוסף, הוצע כי אזורים המציגים סנכרון מקומי נוטים יותר להשתתף בפעילות בין-אזורית 21,22,23. בפרט, תנודות תטא-פס קליפת המוח (4-8 הרץ), כפי שנמדדו על ידי אלקטרואנצפלוגרמה של הקרקפת (EEG), הוצעו כמנגנון פוטנציאלי להעברת שליטה מלמעלה למטה על פני רשתות רחבות13. באופן ספציפי, פעילות תטא-בנד בבני אדם משקפת תהליכים קוגניטיביים ברמה גבוהה, כגון קידוד ושליפה של זיכרון, שימור זיכרון עבודה, זיהוי חידושים, קבלת החלטות ושליטה מלמעלהלמטה 12,24,25,26.
בהקשר זה, קוואנה ופרנק13 הציעו שני מנגנונים עוקבים של תהליכי בקרה: ההכרה בצורך בשליטה ויצירת שליטה. ההכרה בצורך בבקרה עשויה להיות מסומנת על ידי פעילות תטה קו האמצע הקדמי (FMθ) שמקורה בקליפת המוח הקדם-מצחית האמצעית (mPFC), שתוארה במונחים של רכיבי פוטנציאל הקשורים לאירוע (ERP) המשקפים תהליכי בקרה הקשורים ל-mPFC בתגובה למצבים שונים, כגון מידע חדש27,28,29, דרישות גירוי-תגובה סותרות30, משוב שגיאה31, וזיהוי שגיאות32. רכיבי ERP אלה, המשקפים את הצורך בשליטה קוגניטיבית מוגברת בנוכחות חידוש, קונפליקט, עונש או טעות, מציגים חתימה ספקטרלית משותפת בפס התטא שנרשם באלקטרודות קו האמצע הקדמיות 26,27,33,34,35,36,37,38,39,40, 41,42,43,44.
תגובות ה-EEG של פעילות FMθ מציגות דפוס של איפוס פאזה ושיפור הספק בפס תדרי התטא26. למרות המגבלות של שיטת ה-EEG מבחינת הרזולוציה המרחבית שלה, נאספו מקורות שונים של ראיות כדי להוכיח שפעילות FMθ נוצרת על ידי קליפת המוח האמצעית (MCC)13. מאמינים כי דינמיקת תטא זו משמשת כמסגרות זמניות המווסתות את התהליכים העצביים של ה-mPFC, אשר מוגברים לאחר מכן בתגובה לאירועים הדורשים שליטה מוגברת26. זה הוכח באמצעות ניתוח מקורות 31,33,45,46,47, הקלטות EEG והדמיית תהודה מגנטית תפקודית (fMRI) במקביל48,49, והקלטות EEG פולשניות בבני אדם50 וקופים 51,52,53.
על סמך תצפיות אלה, תטה קו האמצע הקדמי נחשבת למשמשת כמנגנון אוניברסלי, שפה משותפת, לביצוע בקרה אדפטיבית במצבים שונים בהם יש חוסר ודאות לגבי הפעולות והתוצאות, כמו במהלך התכנון. הפרדיגמה ההתנהגותית שאנו מציעים בפרוטוקול זה שימשה לחקר תכנון קוגניטיבי ומאפייניו הזמניים והעצביים. למרות שמנגנונים שונים לשליטה קוגניטיבית דווחו בתרחישים אחרים, הפרוטוקול הנוכחי איפשר תיאור עדכני של תכנון ותכונות עצביות וזמניות הקשורות אליו54. התהליך הקוגניטיבי של התכנון מורכב משני שלבים נפרדים: שלב התכנון המנטלי, שבמהלכו מפותח ייצוג פנימי של רצף תוכניות55, ושלב ביצוע התכנון, בו מבוצעות קבוצה של פעולות מוטוריות להשגת המטרה שתוכננהקודם לכן 56. ידוע כי תכנון דורש שילוב של מרכיבים שונים של תפקודים ניהוליים, כולל זיכרון עבודה, בקרת קשב ועיכוב תגובה, מה שהופך את המניפולציה הניסיונית והמדידה המבודדת של תהליכים אלה למאתגרים57,58.
מחקרי הדמיה מוחית על תכנון קוגניטיבי השתמשו בדרך כלל בפרדיגמות התנהגותיות כגון מצודת לונדון 59,60,61; עם זאת, על מנת לשלוט בגורמים המבלבלים, המשימות המשמשות לחקר תכנון קוגניטיבי יכולות להיות מוגבלות ומלאכותיות, מה שמוביל לתוקף פחות ניבוי ואקולוגי 62,63,64,65. כדי להתגבר על בעיה זו בתחום הנוירופסיכולוגיה, הוצעו מצבי תכנון בעולם האמיתי כמשימות אקולוגיות62,63. תת-מבחן המשימה של מפת גן החיות בסוללת ההערכה ההתנהגותית של תסמונת דיסאקזקיוטיב מודד מיומנויות תכנון וארגון בצורה טבעית ורלוונטית יותר64,66. מבחן זה הוא מבחן עיפרון ונייר הכולל תכנון מסלול לביקור ב-6 מתוך 12 מיקומים על מפת גן החיות. המיקומים הם מקומות נפוצים שניתן למצוא בגן חיות רגיל, כמו בית פילים, כלוב אריות, אזור מנוחה, בית קפה וכו'. ישנם שני תנאים המעריכים רמות שונות של תכנון: א) תנאי הניסוח, שבו הנבדקים מונחים לתכנן מסלול לביקור בשישה מקומות לפי סדר בחירתם אך על פי מערכת כללים; ו-2) תנאי הביצוע, שבו הנבדקים מונחים לבקר בשישה מקומות בסדר מסוים ובהתאם למערכת כללים. שני תנאים אלה מספקים מידע על מיומנויות תכנון בבעיות לא מובנות (ניסוח) ומובנות היטב (ביצוע)67. הראשונה מוצגת כמשימה קוגניטיבית תובענית יותר במצב פתוח מכיוון שהיא דורשת מהנבדקים לפתח אסטרטגיה הגיונית להשגת המטרה. לפני מעקב אחר נתיב, יש להמציא רצף של אופרטורים; אחרת, סביר להניח שיתרחשו שגיאות. מצד שני, תנאי הביצוע דורש דרישה קוגניטיבית נמוכה יותר מכיוון שפתרון משימה הכרוכה בביצוע אסטרטגיה כפויה ספציפית רק דורש מהנבדק לעקוב אחר יישום התוכנית שנוסחה להשגת המטרה66. מצד שני, מבוך פורטאוס הוא משימה ידועה בתחום הפסיכולוגיה, במיוחד בתחומי הפסיכולוגיה הקוגניטיבית והנוירופסיכולוגיה, והוא נמצא בשימוש נרחב ככלי להערכת היבטים שונים של קוגניציה, כגון פתרון בעיות ותכנון 68,69. משימת מבוך פורטאוס היא משימת עיפרון ונייר שמתחילה בניתוח גירויים חזותיים פשוט והופכת לקשה יותר ויותר. על הנבדק למצוא ולעקוב אחר הנתיב הנכון מנקודת התחלה ליציאה (בין מספר אפשרויות) תוך שמירה על כללים, כגון הימנעות מנתיבים מצטלבים ומבוי סתום, ולפעול במהירות האפשרית68. בכל פעם שמופיע מזלג בזמן שרטוט השביל, הנבדקים מקבלים החלטות כדי להגיע למטרה ולהימנע משבירת הכללים הנתונים69.
בהתחשב במגבלות ובחוזקות של המשימות הנפוצות והאקולוגיות, עיצבנו את הפרדיגמה ההתנהגותית שלנו בעיקר על סמך משימת מפת גן החיות66 ומשימת מבוך פורטאוס68. הפרדיגמה ההתנהגותית מורכבת מארבעה שלבים נפרדים המקיפים את התהליך הקוגניטיבי של תכנון בתרחיש חיי היומיום. שלבים אלה הם כדלקמן: שלב 1, תכנון, שבו המשתתפים נדרשים ליצור מסלול לביקור במקומות שונים על המפה, תוך הקפדה על הכללים שנקבעו; שלב 2, תחזוקה, שבו המשתתפים נדרשים לשמור את המסלול המתוכנן בזיכרון העבודה שלהם; שלב 3, ביצוע, שבו המשתתפים מבצעים את המסלול שתוכנן מראש על ידי שרטוט ומעקב צמוד אחר דיוקו; ושלב 4, תגובה, שבו המשתתפים מדווחים על רצף בעלי החיים שביקרו על פי מסלול54 המתוכנן שלהם. הפרדיגמה שלנו מאפשרת מדידה של פרמטרים שונים של יכולת תכנון באמצעות שלבים שונים, המשקפים את מרכיבי התכנון השונים (כגון זיכרון עבודה, קשב ניהולי ומיומנויות חזותיות-מרחביות) בצורה מציאותית יותר שכן מיפוי מסלולים הוא תופעה שכיחה בחיי היומיום. בנוסף, כדי לשלוט בגורמים מבלבלים, הפרדיגמה כוללת משימת בקרה עם מבנה משימת תכנון וגירויים שווי ערך, המעסיקה את המרכיבים הקוגניטיביים הניהוליים המעורבים גם בתכנון אך אינה כוללת את מרכיב תהליך התכנון. זה מאפשר הפרדה של מרכיב תהליך התכנון להשוואה הן של סמנים אלקטרופיזיולוגיים והן פרמטרים התנהגותיים54.
יתר על כן, מעקב עיניים תרם תרומה משמעותית למחקרים במדעי המוח הקוגניטיביים על ידי מתן שיטה לא פולשנית למדידה וניתוח של תנועות עיניים, שיכולה לספק תובנות חשובות לגבי התהליכים הקוגניטיביים והמנגנונים העצביים העומדים בבסיס התפיסה, הקשב והתפקודים הקוגניטיביים. מדידת סוגים שונים של תנועות עיניים עם מערכת מעקב עיניים יכולה לספק מידע רב ערך על התהליכים הקוגניטיביים והמנגנונים העצביים המעורבים בתכנון. לדוגמה, ניתן למדוד את ההיבטים הבאים: קיבעונות, שהם תקופות המבט היציב שבמהלכן נרכש מידע חזותי70; סקאדות, שהן תנועות העיניים המהירות המשמשות להעברת המבט ממקום אחד למשנהו71; מרדף חלק, שהוא סוג של תנועת עיניים המאפשרת לעיניים לעקוב אחר אובייקט נע בצורה חלקה72; מיקרוסקאדות, שהן תנועות עיניים קטנות ומהירות המתרחשות גם במהלך קיבועים73; ומצמוצים, שהם פעולת רפלקס המסייעת לשמור על העיניים משומנות ולהגן עליהן מפני עצמים זרים74. תנועות עיניים אלו יכולות לספק תובנות לגבי התהליכים הקוגניטיביים המעורבים בחיפוש חזותי, הקצאת קשב70, מעקב חזותי72, תפיסה73 וזיכרון עבודה74, שהם מרכיבים חשובים לתכנון ושליטה קוגניטיבית.
מצד שני, מחקרים אחרונים על מערכת locus coeruleus-norepinephrine (LC-NE) הראו את תפקידה הרלוונטי בשליטה קוגניטיבית75. ה-locus coeruleus (LC) מקרין למספר אזורי מוח, כגון קליפת המוח, ההיפוקמפוס, התלמוס, המוח האמצעי, גזע המוח, המוח הקטן וחוט השדרה 76,77,61. עצבוב LC-NE צפוף במיוחד מקבל אזורי מוח PFC הקשורים לשליטה קוגניטיבית75. חוץ מזה, כמה מחקרים מצביעים על כך שהיפראקטיביות כרונית של מערכת ה-LC עשויה לתרום לתסמינים של הפרעת מאניה-דפרסיה, כגון אימפולסיביות ונדודי שינה. לעומת זאת, ירידה כרונית בתפקוד LC נקשרה לירידה בביטוי רגשי, מאפיין שכיח בקרב חולים הסובלים מדיכאון78. תגובה פעילה יתר של ה-locus coeruleus לגירויים עלולה להוביל לתגובה מוגזמת אצל אנשים עם הפרעות מתח או חרדה79. לכן, שינויים במערכת LC-NE עשויים לתרום לתסמינים של חוסר ויסות קוגניטיבי ו/או רגשי. ניתן להשתמש בטכניקות לא פולשניות כדי לבחון את פעילות הלוקוס קורולאוס, אחת מהן היא שינויים בקוטר האישון, הנשלטים בעיקר על ידי נוראדרנלין המשתחרר מהלוקוס קורולאוס. נוראדרנלין פועל על שריר מרחיב הקשתית על ידי גירוי קולטני האלפא-אדרנוצפטורים ועל גרעין אדינגר-ווסטפאל, השולח אותות לגנגליון הריסי ושולט בהתרחבות הקשתית באמצעות הפעלת אדרנוצפטורים אלפא-2 פוסט-סינפטיים 66,80,81,82. הקלטות עצביות ישירות של LC מקופים אישרו את הקשר בין פעילות LC-NE, קוטר האישון וביצועים קוגניטיביים83. הרחבת אישונים נצפתה שוב ושוב בתגובה לדרישות עיבוד מוגברות במספר משימות קוגניטיביות 71,84,85,86,87.
סמנים אלקטרופיזיולוגיים של שליטה קוגניטיבית בשילוב עם מעקב עיניים והקלטות אישונים עשויים לנתק שאלות מכריעות לגבי האופן שבו שליטה ותכנון קוגניטיביים מיושמים במוח. החשיבות של שימוש בפרוטוקול שלנו המשלב EEG ומערכות מעקב עיניים היא כפולה. מצד אחד, נראה כי שליטה קוגניטיבית דורשת השתתפות של פעילות מוחית מבוזרת ביחסים זמניים מדויקים, המהווים מועמדים אידיאליים לחקר תפקוד רשת המוח. מצד שני, לחריגות בכל אחת מהיכולות הללו יש השפעה חמורה על התנהגות נורמלית, כפי שעשוי להיות במקרה של מגוון הפרעות קוגניטיביות ונוירו-פסיכיאטריות, כגון הפרעת קשב וריכוז88,89, הפרעת דיכאון מג'ורי 90,91, הפרעה דו קוטבית 91, סכיזופרניה92, דמנציה פרונטו-טמפורלית93, כמו גם הפרעות עקב נגעים קדמיים94. בנוסף, הפרוטוקול הנוכחי מאפשר להשתמש באישון כפרמטר להשוואת פעילות LC-NE ותנודות באמצעות מעקב עיניים ואלקטרואנצפלוגרפיה. זה עשוי לא רק לספק ראיות לקשר התיאורטי בין LC-NE, אישון וסמנים עצביים בבני אדם, אלא גם יכול לאפשר מעקב אחר מסלול ההתפתחות של מאפיינים הקשורים למערכת LC-NE במהלך תכנון קוגניטיבי. עם זאת, במודל שלנו, התמקדנו בבדיקה אם יש דפוס ספציפי של סקאדות במהלך התכנון שעלול לגרום לשינויי תנודה ספציפיים95. בנוסף, השתמשנו במערכת מעקב עיניים כחלק חשוב בבחינת הביצוע ההתנהגותי של תוכנית בשלב הביצוע של הפרדיגמה ההתנהגותית שלנו.
לסיכום, פרוטוקול זה עשוי לייצר מודלים ניתנים לבדיקה של דינמיקה של רשת המוח שיכולים לשמש פלטפורמה הן למחקר בסיסי נוסף והן ליישומים קליניים וטיפוליים בסופו של דבר.
Protocol
כל ההליכים בפרוטוקול זה אושרו על ידי הוועדה הביו-אתית של הפקולטה לרפואה של האוניברסיטה הקתולית של צ'ילה, וכל המשתתפים חתמו על טופס הסכמה מדעת לפני תחילת המחקר (מספר פרויקט מחקר: 16-251).
1. גיוס משתתפים
- לגייס מבוגרים בריאים ימניים (גברים ונשים) עם ראייה תקינה או מתוקנת לנורמה, ולסנן אותם על פי קריטריוני הכללה/אי הכללה.
הערה: במחקר זה גויסו 27 אנשים בריאים בגילאי 19 עד 38 ודוברים שוטף. גודל המדגם יכול להשתנות בהתאם לרמת העוצמה הסטטיסטית הרצויה, וטווח הגילאים עשוי להשתנות בהתאם לשאלת המחקר הספציפית שיש לטפל בה. בפרוטוקול שלנו חישבנו את גודל המדגם על ידי התחשבות במבחן הדירוג הסטטיסטי של Wilcoxon, גודל אפקט של 0.7, רמת אלפא של 0.05 ועוצמה של 0.95, כפי שמתואר ב-Faul et al.96. השתמשנו בראיון נוירו-פסיכיאטרי בינלאומי97, שיושם על ידי פסיכולוג מיומן, כדי להעריך את המשתתפים במונחים של קריטריוני הכללה/אי הכללה. גייסו רק נבדקים ימניים כדי להפחית את השונות באות ה-EEG מכיוון שאנשים שמאליים עשויים להציג התפלגות טופוגרפית שונה של פעילות EEG 98,99,100.
2. הכנת גירויים
- עבור משימת התכנון, צור סט גירויים באמצעות תוכנת עורך גרפי וקטורי (ראה טבלת חומרים). עבור כל גירוי, עצב מבוך בגווני אפור המייצג מפת גן חיות. בתוך המבוך, צרו שער וכמה שבילים שמובילים למיקומי החיות (לדוגמה, במחקר הזה היו ארבעה מיקומים של חיות, ראו איור 1).
הערה: במחקר הזה יצרנו 36 מבוכים שבהם כל גירוי מורכב ממפת גן חיות עם שער התחלה, ארבע תמונות של חיות שממוקמות במבוך וכמה שבילים. השבילים במבוך עשויים להוביל או לא להוביל למיקומי בעלי החיים. שימוש בגירויים בגווני אפור עם ניגודיות מופחתת מועדף לרוב עבור אישון מכיוון שהוא מפחית את הגירוי של קונוסי הרשתית, האחראים על ראיית צבעים. שינוי זה בדגש הגירוי מאפשר מדידה רגישה יותר של שינויים בגודל האישון, הנחשבים כמשקפים שינויים במצב העוררות או הקשב. בנוסף, השימוש בגירויים בגווני אפור מפחית את השונות במדידה עקב הבדלים בראיית צבעים בין אנשים101. - על מנת להשיג רמות שונות של מורכבות במשימה הסופית, חלקו את הגירויים על פי מספר הפתרונות התקפים בהתאם למטרה ולכללים שנקבעו (במיוחד, המטרה היא לתכנן מסלול לביקור במקומות בעלי חיים). מספר הפתרונות התקפים מתייחס למספר הנתיבים האפשריים לתכנון בהתאם לכללים (ראה כללים באיור 2 ובשלב 5.12.1.). סווג גירויים עם יותר מחמישה פתרונות אפשריים כ"קלים" ואלה עם חמישה פתרונות אפשריים או פחות כ"קשים". לאחר מכן, צור מספר שווה של גרסאות ניסיון עבור כל קטגוריה.
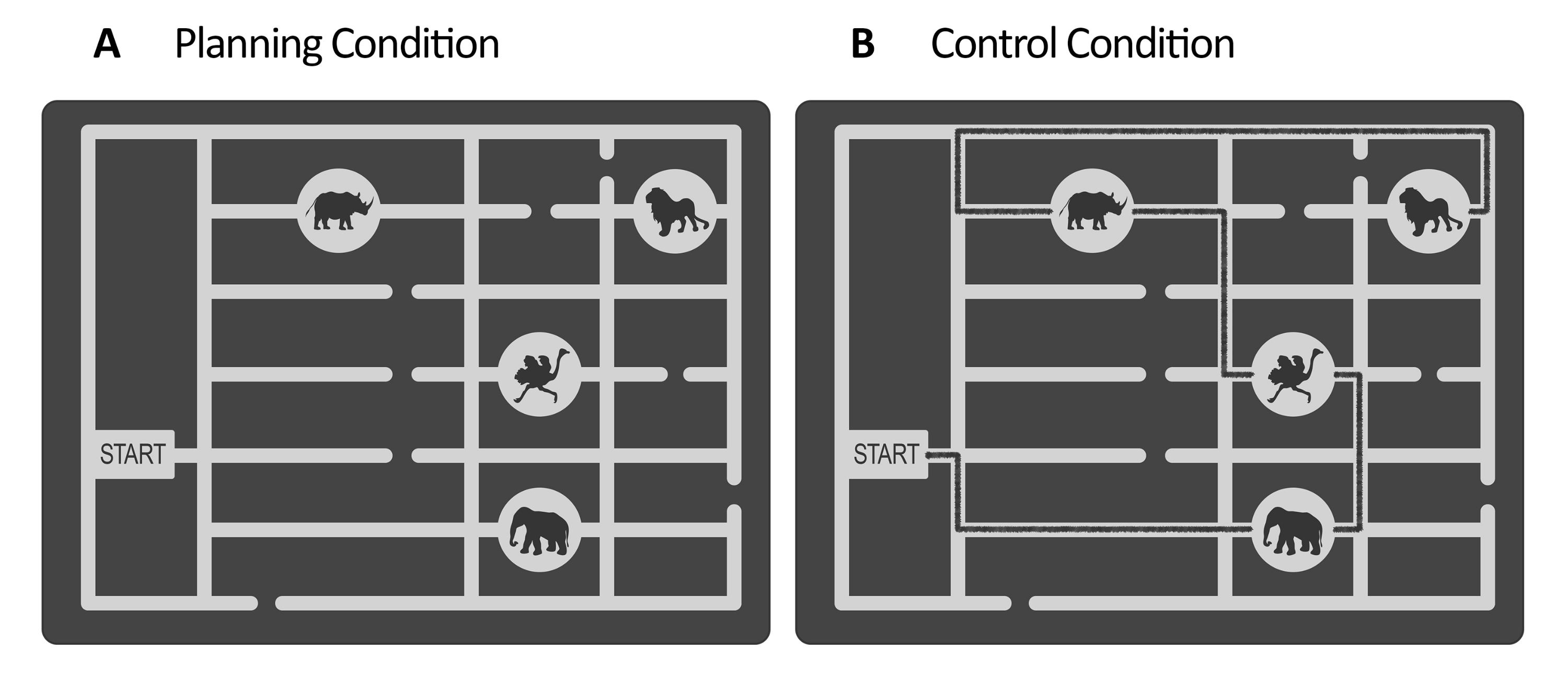
הערה: לחלופין, בקש מהמחברים את הגירויים שנוצרו עבור Domic-Siede et al.54 , מכיוון שהגירויים הללו נוצרו בעקבות הוראות אלה. קחו בחשבון שכל החומרים זמינים לפי בקשה, אך המפרט מפורט כאן. במחקר זה, יצרנו 18 ניסויים קלים ו-18 ניסויים קשים. הערכת ההבדלים ברמות הקושי ברמה ההתנהגותית והאלקטרופיזיולוגית חשובה מכיוון שהיא תעזור לקבוע אם אתה מודד ביקוש קוגניטיבי / מאמץ קוגניטיבי / קושי או היבטים מהותיים של תכנון קוגניטיבי (ראה תוצאות מייצגות וסעיפי דיון). - עבור משימת הבקרה, השתמשו באותו מבנה כמו משימת התכנון (תקופת הערכה, תחזוקה, ביצוע, תגובה, ראו איור 2), והשתמשו באותם גירויים שנוצרו עבור משימת התכנון, אך הוסיפו קו משורטט המייצג נתיב מסומן לביקור ברצף מיקומי בעלי החיים החל מהשער ועד למיקום האחרון (ראו איור 1B). הפוך את השביל המסומן לצבע מעט כהה יותר מהנתיבים הראשיים של המבוך, עם ניגודיות נמוכה המוערכת באמצעות מד לוקס (ראה שלב 2.4).
הערה: הרעיון מאחורי זה הוא לשמור על המאפיינים הפסיכופיזיים של שני המצבים (משימות תכנון ובקרה) דומים ככל האפשר. הנתיבים המסומנים של המבוכים יכלו לעקוב אחר הכללים או לא (ראה סעיף 5 של הפרוטוקול לפרטים נוספים לגבי ההוראות שניתנו למשתתפים). במחקר זה, למחצית מהגירויים היה רצף ביקור נכון בהתאם לכללים, בעוד שהמחצית השנייה הציגה שגיאות (כגון שימוש באותו נתיב פעמיים או חציית מבוי סתום, ראה שלב 5.12.1 ושלב 5.12.3 ואיור 2). - העריכו את עוצמת הבהירות של הגירויים באמצעות מד לוקס הממוקם במשענת הסנטר שבה ישתמשו הנבדקים (ראו שלב 4.5 ושלב 4.6) ובאותו מרחק מהמסך. כל גירוי של כל מצב מייצר ערך לוקס. רשום כל ערך באופן ידני לניתוחים נוספים.
הערה: לא צפויים הבדלים בתאורה בין התנאים (ראה שלב 4.6). אחרת, בדוק את הניגודיות של הגירויים. זה רלוונטי אם קוטר האישון יימדד102. - צרו גירוי אחד שמייצג משוב נכון (אגודל למעלה כשהוא נכון) וגירוי אחר שמייצג משוב שגוי (אגודל למטה כשהוא לא נכון) באמצעות עורך גרפי וקטורי (ראו טבלת חומרים) גם כן (איור 2).

איור 1: גירויים של מטלת הניסוי והבקרה. מוצגות דוגמאות להמחשה של גירוי (A) תכנון ו-(B) משימת בקרה. הגירויים מייצגים מפת גן חיות המורכבת משער, ארבעה מיקומים של בעלי חיים במקומות שונים ומספר שבילים. הגירויים לשני המצבים היו דומים; ההבדל היחיד היה שעבור מטלת הבקרה, (B) לגירויים היה קו מסומן המציין נתיב שכבר קיים (קו שחור כאן למטרות המחשה). קו זה בגירוי הבקרה האמיתי היה מעט כהה יותר, עם ניגודיות נמוכה שנשלטה על ידי הארה (ראה שלב 2.4). נתון זה שונה מ-Domic-Siede et al.54. אנא לחץ כאן לצפייה בגרסה גדולה יותר של איור זה.
{kind=link}

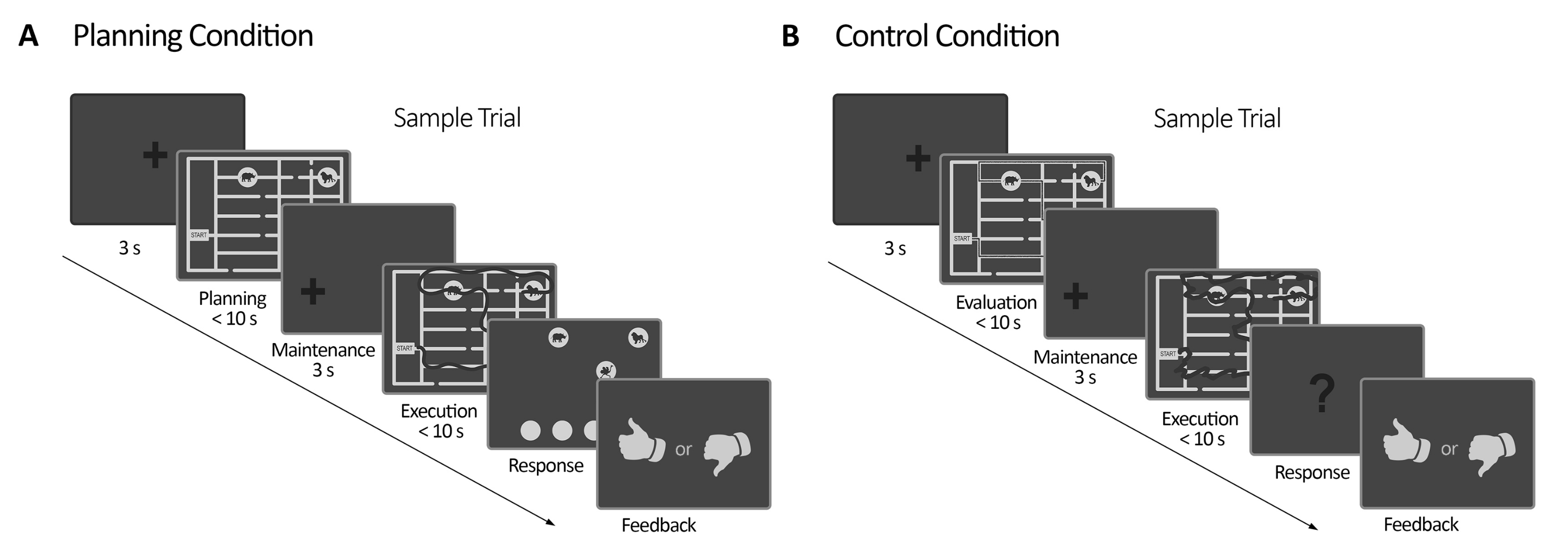
איור 2: תכנון ניסוי. (A) תכנון ניסוי מטלות. ניסויים במצב זה התחילו עם צלב קיבוע של 3 שניות. לאחר מכן, המשתתפים הונחו לתכנן נתיב לביקור בכל ארבעת האתרים של בעלי החיים בהתאם למערכת כללים (מקסימום 10 שניות). לאחר מכן, הופיע צלב קיבוע מוסט (3 שניות), ואחריו הופיע שוב המבוך. בתקופה זו (ביצוע), הנבדקים היו צריכים לבצע את העקבות שתוכננו בתקופת התכנון הקודמת באמצעות מבטם באמצעות משוב חזותי מקוון (שניתן על ידי מערכת מעקב העין), אשר תיאר את תנועת המבט שלהם בזמן אמת (קו כהה) (מקסימום 10 שניות). לאחר מכן, בתקופת התגובה, הנבדקים היו צריכים לדווח על הרצף שנעשה במהלך ההוצאה להורג על ידי הזמנת ביקור בעלי החיים. על פי תשובותיהם, נמסר משוב. (ב) ניסוי משימת בקרה. ניסויים במצב זה התחילו עם צלב קיבוע של 3 שניות. לאחר מכן, המשתתפים הונחו להעריך אם נתיב מסומן (קו כהה) עוקב אחר הכללים או לא. לאחר מכן, הופיע צלב קיבוע מוסט (3 שניות), ואחריו הופיע שוב המבוך. בתקופה זו, הנבדקים נאלצו לשרטט מחדש את הנתיב שכבר אותר עם משוב ויזואלי מקוון, כמו בתקופת ביצוע התכנון (מקסימום 10 שניות). לאחר מכן, בתקופת התגובה, הנבדקים היו צריכים לענות (כן או לא) האם הרצף העוקב פעל לפי הכללים שצוינו קודם לכן על פי תשובותיהם, נמסר משוב. נתון זה שונה מ-Domic-Siede et al.54. אנא לחץ כאן לצפייה בגרסה גדולה יותר של איור זה.
{kind=link}
3. תכנון ובקרה של תכנות משימות
- כתוב סקריפט המקודד פרדיגמת משימת תכנון המבוססת על משימת מפת גן החיות66 ומבוך פורטאוס68באמצעות תוכנת הצגת גירויים/ניסויים התנהגותיים (ראה טבלת החומרים והקובץ המשלים).
- קודד את המשימה בהתחשב בשני תנאים (תנאי תכנון ותנאי בקרה) עם מבנה דומה לזה שהוסבר בסעיף 2 ובסעיף 4 (ראה איור 2 והקובץ המשלים).
הערה: חשוב להשתמש באותו מבנה בשני התנאים על מנת לשלוט בגורמים המבלבלים ובמרכיבי התפיסה המעורבים בתהליך פתרון דרישות המשימה (איור 2). שימוש באותו מבנה משפר את ההערכה של התהליך הקוגניטיבי הספציפי המעורב בתכנון קוגניטיבי. - סנכרן את התקשורת בין מחשב התצוגה, מחשב ה-EEG והמחשב המארח (מחשב עוקב עיניים) באמצעות תקשורת Ethernet ויציאה מקבילית השולחת פולסים לוגיים של טרנזיסטור-טרנזיסטור (TTL) ממחשב התצוגה (ראה איור 3).
- כתוב קוד לכיול תנועות העיניים עם מערכת מעקב העיניים בתחילת משימות התכנון והבקרה ולאחר כל חמישה ניסיונות שהושלמו מכיוון שמיקום המבט על המסך הוא קריטי לתקופת הביצוע (ראה שלב 3.2.3 ושלב 3.3.3 בפרוטוקול, פרק הדיון והקוד בקובץ המשלים).
הערה: ייתכנו עיכובים בתקשורת המחשב. ישנן מספר שיטות למדידת העיכוב בין פולסי TTL בשני מחשבים שונים, אך גישה נפוצה אחת היא להשתמש בהתקן חומרה כגון אוסצילוסקופ דיגיטלי או מנתח לוגיקה. גישה נוספת היא להשתמש בשיטות מבוססות תוכנה, כגון שליחת פולסי TTL על גבי חיבור רשת ושימוש בכלי ניתוח רשת כדי למדוד את העיכוב. גישה נוספת היא לסנכרן את השעונים של שני המחשבים, באמצעות מערכת מיקום גלובלית (GPS) או שרת פרוטוקול זמן רשת (NTP) או באמצעות פתרון סנכרון מבוסס חומרה, לחשב את העיכוב בין חותמת הזמן לזמן ההגעה של כל פולס, ולחשב ממוצע של התוצאות כדי לקבל את העיכוב הכולל בין שני המחשבים.
- קודד את המשימה בהתחשב בשני תנאים (תנאי תכנון ותנאי בקרה) עם מבנה דומה לזה שהוסבר בסעיף 2 ובסעיף 4 (ראה איור 2 והקובץ המשלים).
- כתוב קוד למשימת התכנון עם המבנה הבא: תקופת התכנון, תקופת התחזוקה, תקופת ביצוע התכנון, תקופת התגובה ומשוב (איור 2, קובץ משלים).
- תקופת התכנון: התחל את תנאי התכנון על-ידי הגדרת צלב קיבוע המוצג למשך 3 שניות כקו בסיס.
- הצג באופן אקראי את קבוצת המבוכים בזה אחר זה (36 במחקר זה).
הערה: בתקופת תכנון זו, המשתתפים מתבקשים לתכנן נתיב לביקור בארבעת האתרים של בעלי החיים, עם זמן מקסימלי של 10 שניות, בהתאם למערכת כללים (הכללים הוסברו להם בעבר; ראו סעיף 5 של הפרוטוקול כדי לראות את הכללים שניתנו, כמו גם איור 2). - כלול טריגר TTL בקוד המסמן את תחילת הצגת הגירוי באמצעות קוד תג, ושלח טריגר זה למחשב EEG ולמחשב המארח של עוקב העין לניתוחים צרים יותר וחלונות.
- כתוב בקוד שתקופת התכנון מגיעה לשיאה ברגע שלוחצים על כפתור מג'ויסטיק/מקלדת בכל פעם שהנושא מסיים לתכנן או אם חורג מהזמן המקסימלי. יש לרשום את זמן התגובה (RT) בקובץ היומן לצורך ניתוחים נוספים.
הערה: בתקופה זו, השתמשנו בקוד טריגר באמצעות המספר 1, אך מומלץ להשתמש בתגי מתארי אירועים היררכיים (HED) על פני קודים מספריים, מכיוון שתגי HED מספקים משמעות ומבנה לתוכן, ובכך מקלים על חוקרים או משתפי פעולה אחרים להבין את תוכן הנתונים. - תקופת התחזוקה: התחל תקופה זו באמצעות צלב קיבוע מוסט המוצג למשך 3 שניות. אתר את צלב הקיבוע המוסט במיקום המרחבי שבו ממוקם שער המבוך על מנת לצפות את מיקום ההתחלה (שער) של מפת גן החיות (ראה איור 2).
הערה: מטרת תקופה זו היא משולשת. ראשית, צלב הקיבוע המוסט מקל על ביצוע העקבות המייצגות את הנתיב המתוכנן לתקופה הבאה (ראה שלב 3.2.8). שנית, במהלך תקופה זו, המשתתפים שומרים בזיכרון העבודה שלהם את התוכנית שפותחה במהלך תקופת התכנון. לבסוף, תקופה זו משמשת כמרווח בין ניסיוני לתיחום סוף תקופת התכנון ותחילת התקופה הבאה - תקופת ביצוע התכנון. - תקופת ביצוע התכנון: לאחר הצלבת קיבוע מוסטת מוצגת למשך 3 שניות במהלך תקופת התחזוקה, הצג את המבוך שוב.
- שלח טריגר TTL ל-EEG ולמחשב עוקב העיניים המארח כדי לציין את תחילת התקופה הזו באמצעות קוד תג ספציפי.
- כתוב קוד כדי לתת משוב חזותי בזמן אמת (קו כהה, ראה תקופת הביצוע באיור 2) של מיקום המבט של הנבדק כ-992 אלפיות השנייה לאחר תחילת תקופה זו.
הערה: התחלת התיחום בעיכוב (1,000 אלפיות השנייה בערך) נותנת לנבדקים זמן להתמצא במבוך, ומאפשרת להם לשרטט את הנתיב המתוכנן הקודם שלהם (במהלך תקופת התכנון) בקו כהה. - רשום את הקואורדינטות של הנתיבים לשחזור נוסף של הנתיבים שבוצעו על ידי הנבדקים, ודרג את הביצועים במצב לא מקוון (ראה שלב 6.1.1, איור 4).
- הקפידו על זמן מקסימלי של 10 שניות למעקב אחר הנתיב המתוכנן, ואפשרו לנבדקים לסיים תקופה זו על ידי לחיצה על כפתור. בדרך זו, הנבדקים יכולים לשלוט מתי הם מסיימים את הנתיב המצויר שלהם.
- שמור את ה-RT בקובץ היומן לניתוחים נוספים.
- תקופת התגובה: כתבו קוד לתקופת התגובה, שמתחילה לאחר 10 שניות של ביצוע התכנון או בלחיצת כפתור בתום תקופת ביצוע התכנון, בה המבוך נעלם אך בעלי החיים ומיקומם המרחבי נשארים על המסך.
- מקם ארבעה עיגולים ריקים אופקית בתחתית המסך בתקופת התגובה.
הערה: מטרת תקופה זו היא לאפשר לנבדקים לציין את רצף בעלי החיים שביקרו במהלך תקופת ביצוע התכנון על ידי הכנסת בעלי החיים למעגלים באותו סדר שבו ביקרו אותם באמצעות ג'ויסטיק או מקלדת. - הגדירו את התוכנית/קוד כך שיאפשרו לנבדקים להשתמש בג'ויסטיק או במקלדת כדי לבחור כל אחת מהחיות (ארבע חיות במחקר זה) שהוצגו קודם לכן ולהכניס אותן לכל אחד מארבעת המעגלים (ראו קובץ משלים ואיור 2).
- משוב: כתוב קוד כדי לספק 3 שניות של משוב למשתתפים. יש להציג תמונת אגודל למעלה בתגובה לשילובים חוקיים של בעלי חיים שביקרו אם מקפידים על הכללים, בעוד שתמונת אגודל למטה צריכה להיות מוצגת אם השילוב המדווח אינו חוקי.
- שלח גורם מפעיל TTL, באמצעות קוד תג ספציפי למשוב נכון וקוד תג אחר שגוי, למחשבי EEG ומעקב עיניים.
הערה: הסיבה למתן משוב היא להקל על ניטור הביצועים ולשמור על מוטיבציה במהלך המשימה. מתן משוב בזמן אמת זה משפר את אפקט התגמול ומעודד ביצוע נכון של משימות103.
- כתוב קוד עבור משימת הבקרה עם מבנה זהה לתנאי התכנון: תקופת בקרה, תקופת תחזוקה, תקופת ביצוע בקרה, תקופת תגובה ומשוב (ראה קובץ משלים, איור 2).
- תקופת הבקרה: כתוב קוד עבור תקופת תנאי הבקרה כדי להפחית גורמים מבלבלים. הקוד לתקופה זו חייב להתחיל בצלב קיבוע המוצג למשך 3 שניות כקו בסיס.
הערה: מכיוון שמשימת התכנון דורשת בעיקר יישום של תכנון אך גם מגייסת תחומים קוגניטיביים אחרים כחלק מהתפקוד הניהולי, כגון תפקוד חזותי-מרחבי, זיכרון עבודה, בקרת קשב, שליטה מעכבת וכו'.66,88,104,105, משימת בקרה היא חיונית להפחתת גורמים מבלבלים. לפיכך, המטרה העיקרית של משימה זו היא לדרוש את כל הפונקציות הקוגניטיביות והתפיסתיות הדרושות כדי לפתור את משימת התכנון תוך הסרת היישום של תכנון קוגניטיבי54. - הצג באופן אקראי את מבוכי מצב הבקרה בזה אחר זה (מבוכים עם נתיב מסומן שכבר סומן). קידוד זמן מקסימלי של 10 שניות.
- כלול טריגר TTL בקוד המסמן את תחילת הצגת הגירוי באמצעות תג, ושלח טריגר זה למחשב EEG ולמחשב המארח של עוקב העיניים.
- כתוב בקוד שתקופת בקרה זו מגיעה לשיאה ברגע שלוחצים על כפתור מג'ויסטיק/מקלדת בכל פעם שהנושא מסתיים או אם חורג מהזמן המרבי.
הערה: הנבדקים מונחים להעריך את הנתיבים המסומנים (בין אם הם פועלים לפי הכללים ובין אם לאו, ראה שלב 5.12 לפרטים על ההוראות שניתנו למשתתפים). - שמור את זמן התגובה (RT) בקובץ היומן לניתוחים נוספים.
- תקופת התחזוקה: לאחר סיום תקופת הבקרה, הצג צלב קיבוע מוזז למשך 3 שניות.
- כתקופת ביצוע התכנון, מקם את צלב הקיבוע במקום בו נמצאת הכניסה לשער כדי להקל על משיכת המבט לתקופה הבאה.
- תקופת ביצוע הבקרה: הציגו שוב את המבוך, ובמקביל שלחו טריגר TTL ל-EEG ומארחים מחשבי מעקב עיניים עם תג המסמן את תחילת תקופת הביצוע.
- חזור על אותו קוד כמו בתקופת ביצוע התכנון כדי לתת משוב מקוון על מיקום המבט וכדי לתחום ולחפוף את מבטו עם הנתיב המעקב.
- הקפידו על זמן מקסימלי של 10 שניות למעקב אחר הנתיב, ואפשרו לנבדקים לסיים תקופה זו על ידי לחיצה על כפתור.
- שמור את ה-RT בקובץ היומן לניתוחים נוספים.
- תקופת תגובת הבקרה: לאחר סיום תקופת ביצוע הבקרה, הצג סימן שאלה המציין את תקופת התגובה.
- תכנת שני לחצנים, בהתאמה, כדי שהנבדקים יתנו תגובה באמצעות ג'ויסטיק או מקלדת.
הערה: כאן, הנבדקים מתבקשים לענות אם הרצף שסומן על ידי המעקב היה נכון או לא על ידי בחירת כפתור אחד עבור נכון/כן ואחר עבור שגוי/לא. - שמור את הדיוק בקובץ יומן הרישום.
- משוב: כתוב קוד כדי לספק 3 שניות של משוב נכון בכל פעם שהנבדקים מגיבים נכון (תמונת אגודל למעלה) וספק 3 שניות של משוב שגוי כאשר הנבדקים מגיבים בצורה לא נכונה (תמונת אגודל למטה).
- כמו בתנאי התכנון, שלח גורם מפעיל TTL ל-EEG וארח מחשבי מעקב עיניים עם תג עבור משוב נכון ותג נוסף עבור משוב שגוי.
- תקופת הבקרה: כתוב קוד עבור תקופת תנאי הבקרה כדי להפחית גורמים מבלבלים. הקוד לתקופה זו חייב להתחיל בצלב קיבוע המוצג למשך 3 שניות כקו בסיס.
- משימות הדרכה: צור גירויים, כתוב קוד והצג לפני משימות התכנון והבקרה הנ"ל מפגש הדרכה קצר של כשישה ניסויים/מבוכים לכל מצב (תכנון ובקרה)
הערה: הרעיון הוא להבטיח היכרות עם הגדרת המשימה. מומלץ לקבוע קריטריונים להמשך ההליך. במחקר זה, אם שלושת הניסויים האחרונים היו נכונים, והמשתתפים דיווחו על הבנת המטרה וההליך בסוף האימון, המשתתפים המשיכו למפגש הניסוי.

איור 3: דוגמה למערך מעבדה. ייצוג סכמטי של מערך מעבדה המציג שלושה מחשבים מחוברים זה לזה. המחשב המארח (מחשב מעקב עיניים) אחראי על מעקב ואחסון נתוני תנועת העיניים. מחשב ה-EEG רוכש ומאחסן את אותות ה-EEG. מחשב התצוגה שולט בניסוי ההתנהגותי, מציג את הגירויים לנבדקים ושולח טריגרים לאירועים למחשבי המארח וה-EEG דרך יציאות מקבילות וחיבורי LAN כדי לסנכרן את איסוף הנתונים. אנא לחץ כאן לצפייה בגרסה גדולה יותר של איור זה.
{kind=link}

איור 4: שחזור נתיב ממשוב ויזואלי מקוון שניתן על ידי מערכת מעקב העיניים. דוגמאות להמחשה של שחזור מסלול מביצוע מוטורי של תוכנית (A, בסגול, תקופת ביצוע תכנון) ותקופת ביצוע בקרה (B, קו בירוק) ועם נתוני מעקב עיניים. הנתיב המשוחזר בתקופת ביצוע התכנון משמש להערכת הדיוק של כל ניסיון של משימת תכנון. אנא לחץ כאן לצפייה בגרסה גדולה יותר של איור זה.
{kind=link}
4. הגדרת מעבדה וציוד
- השתמשו במערכת רכישת EEG כדי לתעד את פעילות ה-EEG מהקרקפת של המשתתף, כאשר אלקטרודות ה-EEG ממוקמות בהתאם למערכת הבינלאומית10-20 106. מקם שתי אלקטרודות על המסטואידים של המשתתף להפניה מחדש במצב לא מקוון. השתמש באלקטרודות אלקטרוקולוגרמה כדי לזהות אותות תנועת עיניים אנכיים, אופקיים וממצמצים במהלך בדיקה חזותית.
- השתמש בתוכנת רכישת EEG עבור קליטת נתוני EEG עם קצב דגימה של 2,048 הרץ או 1,024 הרץ ומסנן מעביר-פס בין 0.1-100 הרץ במחשב EEG.
הערה: קצבי דגימה של 1,024 הרץ ו-2,048 הרץ מספקים רזולוציה מספקת לניתוח תנודות בתדר נמוך. חשוב לרכוש אותות EEG עם קצב דגימה גבוה, כגון מעל 1,000 הרץ, בעת ניתוח תנודות בתדר נמוך על מנת להבטיח שהאותות בתדר נמוך אינם מכונים. - השתמש במחשב תצוגה, המחובר למחשב EEG ולמחשב עוקב העיניים המארח באמצעות יציאות מקבילות ו-Ethernet ויש לו פלטפורמה להפעלת ניסויים התנהגותיים המותקנת עליו, כדי להקרין את הגירויים על צג מורחב ברזולוציה מינימלית של 1,920 פיקסלים x 1,080 פיקסלים וקצב רענון של 60 הרץ. מקם צג זה במרחק של כ-82 ס"מ מהנושא.
הערה: השתמשנו בצג 24 אינץ' עם קצב רענון מוגדר ל-144 הרץ הממוקם במרחק של 82 ס"מ מהמשתתף. מומלץ להשתמש בצג בגודל מסך של לפחות 19 אינץ' לניסויים קוגניטיביים הכוללים רישום EEG ותנועות עיניים. בנוסף, המלצה כללית היא למקם את המוניטור במרחק המאפשר למשתתף לבצע את המשימה בנוחות ולשמור על מבט יציב על המסך תוך מתן אפשרות להקלטה מדויקת של EEG ותנועות עיניים. רצוי לבדוק ולהתאים את ההגדרה לפי הצורך כדי להבטיח את התוצאות הטובות ביותר. - השתמשו במערכת מעקב עיניים כדי לתת למשתתפים משוב בזמן אמת על תנועות העיניים שלהם במהלך תקופות הביצוע, ולתעד את גודל האישון. הגדר את קצב הדגימה ל-1,000 הרץ לרזולוציה זמנית נאותה.
- הימנע מתנועות ראש. יש צורך בהגבלות תנועת ראש שמאלה וימינה ולמעלה ולמטה כדי לשמור על העין בשדה הראייה של מצלמת הווידאו. יש צורך בהגבלת תנועה קדימה ואחורה כדי לשמור על העין בטווח המוקד של מצלמת הווידאו. השתמש בשילוב של משענות מצח/סנטר כדי לשמור על תנועות בטווח זה.
- העריכו את בהירות הגירויים באמצעות מד לוקס דיגיטלי או דומה כדי להשוות את ההבדלים בין גירוי התכנון לגירוי הבקרה.
הערה: ניתן להשתמש במבחן סטטיסטי כגון מבחן t או Wilcoxon כדי להעריך את ההבדלים בין הגירויים של שני המצבים. - השתמש בג'ויסטיק בקרה או במקלדת עם ארבעה לחצנים לפחות: שני לחצנים לשאלות כן/לא ממצב הבקרה; אחד משני הכפתורים הללו לסיום ניסויים; ועוד שני כפתורים לתקופת התגובה של תנאי התכנון לנוע קדימה או אחורה כדי להכניס את החיות לכל אחד מארבעת העיגולים בתחתית המסך.
5. מפגשי הקלטה של אלקטרואנצפלוגרפיה ומעקב עיניים
- לפני תחילת המחקר, בקשו מהמשתתפים להשלים הסכמה מדעת בכתב ובחתימה.
- לפני מפגש ההקלטה, בקשו מהמשתתפים לא להשתתף באיפור (מסקרה ואייליינר יכולים להיות מזוהים כאישון על ידי מערכת מעקב העיניים), לאחר נטילת סמים או קפאין107,108, או אם הם מרגישים עייפות חמורה109 (מתח, חוסר שינה וכו').
- בקשו מהמשתתפים להשלים סקר דמוגרפי כדי לספק מידע על המין, הגיל, הידיים, שפת האם וההיסטוריה הנוירו-פסיכיאטרית שלהם באמצעות הראיון הנוירו-פסיכיאטרי הבינלאומיMINI-International 97 המיושם על ידי פסיכולוג מיומן.
- נקו את המצח, הקרקפת, המסטואידים ותנוחת העור האלקטרוקולוגרמה (EOG) של הנבדק עם מגבון אלכוהול.
- הנח את כל האלקטרודות החיצוניות על המשתתף. הניחו את ה-EOG האופקי בצורה דו-קוטבית על הקנתי החיצוני של שתי העיניים ואת ה-EOG האנכי מעל ומתחת לעין ימין של המשתתף. שים שתי אלקטרודות חיצוניות על המסטואיד הימני והשמאלי להתייחסות חוזרת מאוחר יותר.
- מדדו את ראש הנבדק, והניחו את מכסה ה-EEG בגודל הנכון בהתאם למערכת הבינלאומית המורחבת 10-20. על מנת לעשות זאת, מצא ומקם את האלקטרודה Cz לפי השלבים הבאים:
- זהה את קו האמצע של הקרקפת על ידי בדיקה ויזואלית של קו השיער והחלק העליון של האף. זהה קו המחבר בין שתי הנקודות הללו כדי להגדיר את קו האמצע.
- אתר את ה-Cz. ה-Cz מוגדר בדרך כלל כנקודת האמצע בין שתי הנקודות הקדם-אוזניות (כלומר, הנקודות הממוקמות ממש לפני כל אוזן). אתר את הנקודות הללו, ולאחר מכן זהה קו המחבר ביניהן כדי לזהות את המיקום המשוער של ה-Cz.
- מדדו וסמנו את ה-Cz. מדדו את המרחק מהנזיון (כלומר, הבליטה הגרמית בחלק העליון של האף) ל-Cz. המרחק מהנסיון ל-Cz הוא בדרך כלל בסביבות 53% מהיקף הראש הכולל במערכת 10-20. סמן את מיקום ה-Cz באמצעות עט או כלי סימון אחר.
הערה: חשוב לעקוב אחר נוהל עקבי וסטנדרטי למיקום אלקטרודות על מנת למזער שגיאות ולהבטיח את תקפות נתוני ה-EEG. מומלץ לפתח נוהל השמה סטנדרטי. צור נוהל סטנדרטי להנחת האלקטרודות על הקרקפת של כל נבדק, וודא שנעשה שימוש באותו הליך עבור כל נבדק. במקרה שיש צוות או צוות שמבצע את ההקלטות, הכשירו את הטכנאים או עוזרי המחקר על נוהל המיקום הנכון כדי להבטיח שהם ממקמים את האלקטרודות באופן עקבי ומדויק. יתר על כן, דיגיטציה של מיקומי האלקטרודות עבור כל נושא תהיה גישה רצויה על מנת לבצע ניתוח מקור מאוחר יותר. במחקרי EEG, המיקום התלת-ממדי המדויק של כל אלקטרודה על ראשו של הנבדק הוא לעתים קרובות דרישה לביסוס מתאם בין נתוני ה-EEG לבין הפעילות המוחית המקבילה110. מידע זה הוא קריטי גם ליישור נכון של נתוני ה-EEG עם תמונות אנטומיות הנגזרות מטכניקות הדמיית MRI או CT111,112.
- הכנס ג'ל מוליך לכל חור במכסה באמצעות מזרק עם מחט קהה, והרחיק את שיער המשתתף עם הקצה. לאחר מכן, הניחו את כל האלקטרודות של הקרקפת על מכסה ה-EEG.
- בדוק את העכבות באמצעות תוכנת הקלטת EEG, וודא שהן מתחת לרמת ההתנגדות המומלצת על ידי מערכת EEG.
- בקשו מהמשתתף להישאר דומם ככל האפשר במהלך הניסוי. בדקו את אות ה-EEG ובדקו אותו על ידי כך שתבקשו מהמשתתף למצמץ, ליצור לסת ולהישאר כמה שניות בעיניים עצומות.
- הושיבו את המשתתף בחדר חשוך ומוחלש קול. השתמשו במשענת סנטר כדי לייצב את הראש ולמזער את התנועה, ובדקו שיש מרחק של כ-82 ס"מ בין משענת הסנטר למרכז מסך הגירוי.
- הניחו ג'ויסטיק או מקלדת לפני המשתתף לקבלת התגובות.
- הוראות: תן הוראות בעל פה באמצעות עזרים חזותיים לפני תחילת כל מצב (תכנון ובקרה). בהוראות יש לכלול דוגמאות חזותיות של הגירויים, ולהסביר כיצד לפתור את המבוכים בתנאי התכנון והבקרה, בהתאמה.
- לצורך משימת התכנון, הנחו את הנבדקים למצוא נתיב להשלמת רצף של ביקורים במקומות מסוימים של בעלי חיים (ארבעה מיקומים במחקר זה) במקומות שונים במבוך בכל סדר ובהתאם למערכת כללים: "(1) לתכנן את השביל מהר ככל האפשר תוך 10 שניות לכל היותר; (2) התחילו מהשער, וסיימו את השביל בבעל החיים הרביעי שביקרתם; (3) אל תעבור באותו שביל או באותו פינה פעמיים; (4) אל תחצו מבוי סתום; (5) אל תחצה שביל בניצב"54.
הערה: עבור כלל 2 עד כלל 5, אנו ממליצים להציג דוגמאות חזותיות למשתתף. - לאחר מכן, התחל את מפגש ההדרכה של משימות התכנון של שישה ניסויים.
הערה: הנחו את המשתתפים לדווח לפני כל תקופת כיול של עוקב עיניים אם הייתה בעיה כלשהי בביצוע המשימה, במיוחד במהלך תיחום הנתיב בתקופת הביצוע. רשום לעצמך אם היה ניסיון לבדיקה לא מקוונת/לאחר עיבוד (ראה שלב 6.1.1). - עבור תנאי הבקרה, הנחו את הנבדקים להעריך אם הנתיב שכבר סומן במבוך נעשה בצורה נכונה או לא נכונה בהתחשב בכללים שנלמדו בעבר.
הערה: תן דוגמאות עם תמיכה חזותית כיצד להעריך את המבוכים מבלי להשתמש באסטרטגיות תכנון, כגון לא לנסות לתכנן נתיב חדש כאשר מתגלות שגיאות (כגון שרטוטים המשתמשים באותו נתיב פעמיים, חציית מבוי סתום וכו'). כאשר נמצאה שגיאה, ההתמקדות צריכה להיות אך ורק בדיווח על זיהוי השגיאה ולא בתיקון הנתיב. לאחר כל ניסוי, שאלו את המשתתפים על האסטרטגיות שהם יישמו. לאחר מכן, ספק משוב בעל פה על ביצועיהם כדי לוודא שהם העריכו את הנתיבים ששורטטו ונמנעו מתכנון נתיבים חדשים. לאחר מכן, התחל את מפגש ההדרכה של משימות הבקרה של שישה ניסיונות.
- לצורך משימת התכנון, הנחו את הנבדקים למצוא נתיב להשלמת רצף של ביקורים במקומות מסוימים של בעלי חיים (ארבעה מיקומים במחקר זה) במקומות שונים במבוך בכל סדר ובהתאם למערכת כללים: "(1) לתכנן את השביל מהר ככל האפשר תוך 10 שניות לכל היותר; (2) התחילו מהשער, וסיימו את השביל בבעל החיים הרביעי שביקרתם; (3) אל תעבור באותו שביל או באותו פינה פעמיים; (4) אל תחצו מבוי סתום; (5) אל תחצה שביל בניצב"54.
- בדוק את אות ה-EEG כדי לוודא שכל הערוצים נרכשים כהלכה. התחל את הקלטת ה- EEG.
- כייל את עוקב העיניים.
הערה: ודא את היכולת של עוקב העיניים לקבוע את מיקום המבט כאשר המשתתף מפנה את מבטו לאזורים שונים במסך.- הודע למשתתף שעוקב העיניים יכויל ושהוא הולך לראות עיגול לבן (עם נקודה אפורה קטנה) נע באופן אקראי לארבע פינות המסך (הליך כיול של חמש נקודות). הנחו אותם לקבע את מבטם במעגל, והודיעו להם שכאשר הוא עובר למקום אחר, עליהם לעקוב אחר מיקום העיגול ולקבע את מבטם שוב במיקום החדש.
- הריצו את הניסוי, התחילו לשמור את תנועות העיניים על ידי לחיצה על פלט/הקלט, ובקשו מהמשתתף לעקוב אחר ההוראות שניתנו קודם לכן, ולהודיע לו שהניסוי יתחיל כעת.
- שמור את חדר המעבדה בסביבה חשוכה. השינויים הגדולים ביותר בהתרחבות האישונים מתרחשים בתגובה לשינויים בבהירות113. שמור על רמת אור עקבית בסביבת הניסוי.
6. ניתוח נתונים
- ניתוחים התנהגותיים
- לנתח את הנתונים ההתנהגותיים באמצעות תוכנה סטטיסטית (ראה טבלת חומרים). מדוד את הדיוק (אחוז התגובות המדויקות) כפרמטר התנהגותי כמותי הן בתנאי התכנון והן בתנאי הבקרה. עבור תנאי התכנון, השתמש בנתוני עוקב עיניים (קואורדינטות x ו-y של מיקום המבט) כדי לשחזר את הנתיבים שנלקחו במהלך תקופת הביצוע במצב לא מקוון, ולקבוע את הדיוק של הנתיבים המתוכננים בהשוואה לנתיבים שבוצעו בפועל (איור 4). לשם כך, בדוק באופן ידני את ההתאמה בין השילובים שבוצעו בצורה נכונה/שגויה בתקופת התגובה לבין המעקב שבוצע.
- חשב את ה-RT, שהוא הזמן הממוצע המושקע בפתרון המבוכים לתקופת התכנון והזמן הממוצע המושקע בהערכת השבילים המסומנים לתקופת הבקרה.
- חשב את ה-RT הממוצע של תקופות הביצוע עבור תנאי התכנון והבקרה. באופן ספציפי, השתמש ב- RT המתאים רק לגירסאות הניסיון הנכונות.
הערה: באופן משלים, ניתן להשתמש בציון דיוק המהירות המשולב הליניארי (LISAS)114,115 המתואר ב-Domic-Siede et al.54, המספק מדד משולב המתחשב בזמן תגובה ודיוק. מכיוון שזמן התגובה במהלך תקופת ביצוע התכנון ודיוק התכנון קשורים זה בזה, ניתן להשתמש ב-LISAS כדי לחשב מדד הלוקח בחשבון את זמן התגובה המתוקן למספר השגיאות שנעשו. בנוסף, ניתן להשתמש במדד LISAS כדי להעריך את המתאם בין אותות אלקטרופיזיולוגיים לביצועים התנהגותיים גם כן. הוא מחושב כשילוב ליניארי של זמן תגובה (RT) ויחס שגיאה (PE). - להעריך את ההומוססדסטיות באמצעות מבחן סטטיסטי כגון מבחן לוין116,117, ולבדוק את הנורמליות באמצעות מבחן התפלגות הנורמליות של ד'אגוסטינו ופירסון118 או מבחן שפירו וילק119 כדי לבחור את המבחן הסטטיסטי המתאים להשוואה (פרמטרית או לא פרמטרית).
- העריכו אם מרכיב התכנון בתנאי התכנון תובעני יותר מבחינה קוגניטיבית ממצב הבקרה באמצעות מבחן הדירוג החתום של Wilcoxon120 או מבחן ה-t של הזוג המותאם121 כדי להשוות את הפרמטרים ההתנהגותיים בין התנאים.
הערה: בדרך זו, תקף שהפרדיגמה ההתנהגותית היא אופטימלית להערכת תכנון קוגניטיבי. - הפרד את הניסויים בתנאי התכנון לקטגוריות "קל" ו"קשה" (עיין בשלב 2.2), ולאחר מכן השתמש במבחן t מותאם כדי להשוות את הדיוק וזמני התגובה בתקופות התכנון והביצוע בין הניסויים "הקלים" וה"קשים".
- עיבוד מקדים של EEG ותנועות עיניים
- בצע את צינור העיבוד המקדים של נתוני EEG המוסבר בנקודות הבאות באמצעות סקריפטים מתוצרת עצמית ו/או ארגזי כלים מבוססים, כגון אלה המתוארים ב-Delorme ו-Makeig122, ב-Dimigen et al.123, וב-Mognon et al.124, בתוכנת שפת תכנות (ראה טבלת חומרים).
- סנכרן את פעילות תנועת העיניים עם הקלטות ה-EEG כדי לייבא את הקיבוע, הסקאדות, ואירועי המצמוץ לבדיקה חזותית טובה יותר או ניתוחים נוספים (ראה שלב 3.1.2 והקובץ המשלים).
הערה: במחקר זה, השתמשנו בחותמות הזמן על נתוני מעקב העיניים ובחותמות הזמן על נתוני ה-EEG כפי שמתואר ב-Domic-Siede et al.54 וב-Dimigen et al.123 כדי לייבא את אירועי תנועת העיניים לנתוני EEG בתוכנת שפת תכנות. - דגימה למטה של הנתונים ל-1,024 הרץ למקרה שהם נרשמו ב-2,028 הרץ כדי להפחית את הדרישות החישוביות.
הערה: קצב דגימה של 1,024 הרץ מספיק בהתאם לטווח התדרים המעניין של 4-8 הרץ, רזולוציית התדרים הצפויה והדרישות החישוביות של הניתוח. - התייחס מחדש לאות ה-EEG לממוצע האלקטרודות על המסטואידים.
הערה: הפניות אחרות אפשריות. בחירת ההפניה יכולה להשפיע על תוצאות ניתוח ה-EEG ועל פרשנות הנתונים, ולכן חשוב לשקול היטב את היתרונות והחסרונות של אפשרויות הפניה שונות. התייחסות המסטואיד הממוצעת היא בחירה פופולרית למחקרי EEG מכיוון שהיא מספקת התייחסות יציבה שקל לחשב, והיא הוכחה כיעילה לניתוח אותות EEG רבים ושונים. התייחסות לנתוני ה-EEG לממוצע המסטואידים (המכונה התייחסות המסטואיד הממוצע) היא גישה נפוצה לניתוח פעילות תטה קדמית בנתוני EEG בקרקפת. אלקטרודות המסטואיד ממוקמות ליד האוזן ומספקות התייחסות לאותות ה-EEG. התייחסות לממוצע המסטואידים יכולה לעזור להפחית את ההשפעה של רעש וחפצים שאינם מעניינים תוך הימנעות מביטול אות העניין, מה שעוזר למשתמש לקבל ייצוג ברור יותר של אותות ה-EEG. - החל תגובת דחף סופית (FIR) אפס-פאזה עם תדר ניתוק מעביר-גבוה של 1 הרץ ותדר ניתוק מעביר-נמוכים של 40 הרץ על פני האות המורחב (ללא עידון) באמצעות תוכנת שפת תכנות.
הערה: במחקר זה, השתמשנו בארגז הכלים המתואר ב-Delorme ו-Makeig122. - עבור כל מצב, בהתחשב במספר הניסויים, חלקו את הנתונים לתקופות המתרכזות סביב תחילת תקופות התכנון והבקרה, בהתאמה. השתמש ב-1 שניות לפני תחילת הצגת המבוך כקו הבסיס וב-4 שניות לאחר תקופת התכנון או הבקרה כקטעי העניין. השתמש בתוכנת שפת תכנות.
הערה: במחקר זה, השתמשנו בארגז הכלים המתואר ב-Delorme ו-Makeig122ו-36 תקופות/ניסויים. - צור פילוח שני שבמרכזו סוף תקופות התכנון והבקרה תוך שימוש ב-4 שניות לפני הסוף ו-1 שניות לאחר מכן כתקופת התחזוקה.
הערה: הסיבה לבחירת 4 השניות הראשונות והאחרונות של תקופות התכנון והבקרה (שלב 6.2.6 ושלב 6.2.7) היא שמשך כל תקופה בשני התנאים יכול להשתנות, וניתוח השניות הראשונות והאחרונות של התכנון יכול לספק מבט מקיף יותר על תהליך התכנון. לפיכך, אורכי חלונות אלה מספיקים ומספיקים כדי לנתח את הדינמיקה התנודתית העומדת בבסיס התכנון. - מעל האות המפולח, הפעל את אלגוריתם ניתוח הרכיבים הבלתי תלויים של Logistic Infomax (ICA)125כדי לזהות ולהסיר רכיבים מלאכותיים.
- השתמש בקריטריון יחס השונות של סקדה לקיבוע המומלץ ב-Plöchl et al.126כדי לזהות אוטומטית רכיבים רועשים פוטנציאליים, והשתמש בגלאי חפצי EEG אוטומטי המבוסס על שימוש משותף בתכונות מרחביות וזמניות המומלצות ב-Mognon et al.124.
הערה: אנו ממליצים להשתמש בסיווג הרכיבים הבלתי תלוי המוצע ב-Pion-Tonachini et al.127, המעריך סיווגי רכיבים בלתי תלויים כווקטורים קומפוזיציוניים על פני שבע קטגוריות, ומסייע בזיהוי חפצים. - בדוק רכיבים מלאכותיים פוטנציאליים אחרים כגון EMG, תנועת אלקטרודות או רכיבים שאינם קשורים למוח. אמת את הדחייה של רכיבים אלה על ידי בדיקה חזותית של הטופוגרפיות, הספקטרום וההפעלות לאורך זמן.
- אינטרפולציה (אינטרפולציה כדורית) ערוצים רועשים על ידי דחיית ערוצים אוטומטית באמצעות קריטריון הקורטוזיס (עם ציון z של 5 כסף).
- ניתוחי תדירות זמן
- בצע התמרת פורייה מהירה לזמן קצר (FFT) (1 הרץ עד 40 הרץ) באמצעות אורך חלון של 250 אלפיות השנייה וצעד זמן של 5 אלפיות השנייה. השתמש בחלון Hanning. השתמש בציון z כדי לנרמל את תרשימי תדירות הזמן לקו הבסיס (-1 שניות עד -0.1 שניות).
הערה: ההדמיה של הספקטרום כפופה לפשרה בין גודל החלון לרזולוציה הזמנית. כדי להשיג תצוגה מקיפה של כל הספקטרום כולל טווח התטא של 4 הרץ עד 8 הרץ, אנו ממליצים להשתמש בגבול התחתון של גודל החלון, שהוא 250 אלפיות השנייה, כדי להבטיח רזולוציה זמנית גבוהה יותר במהלך כל ניסוי ומשימה. בנוסף, אנו ממליצים להשתמש בחלון Hanning, מכיוון שזו נחשבת לבחירה קונבנציונלית במקרים אלה. לרזולוציה טובה יותר בזמן ובתדירות, ראה את השלבים הנוספים. - בחר תרשים תדר זמן מאלקטרודה קדמית, כגון Fz, או קבוצה ממוצעת של אלקטרודות קדמיות.
הערה: קחו בחשבון את הראיות הרחבות לגבי הקשר בין שליטה קוגניטיבית לתטא קו האמצע הקדמי 12,128,129. - בחר תרשימי זמן-תדר בקרה לא חזיתית מאלקטרודות כגון אלקטרודות Pz ו-Oz כדי לקדם את ההשוואות.
- עבור האלקטרודות הקדמיות והבקרה, בצע בדיקת תמורות מבוססת אשכול לא פרמטרית עבור דגימות מזווגות, עם ערך p < 0.05 עבור ההשוואות ברמת הקבוצה של תרשימי תדירות הזמן משני התנאים. השתמש בשיטת מונטה קרלו עם 1,000 הגרלות אקראיות. השתמש בערך הסטטיסטי המרבי של האשכול כדי לבצע את בדיקת התמורות130.
- ממוצע של פס תדרי התטא (4-8 הרץ) מ-4 השניות הראשונות של התכנון והבקרה, בהתאמה, וגם מקטע 4 השניות האחרונות.
- השווה את פעילות התטא הממוצעת בין התנאים באמצעות מבחן t של זוג תואם או מבחן דירוג חתום של Wilcoxon.
- נתח את פרופיל הזמן של פעילות התטא. לשם כך, ממוצע טווח התדרים של 4-8 הרץ על פני הניסויים לפי נבדק.
- השווה את דינמיקת פעילות התטא בין התנאים באמצעות מבחן Wilcoxon חתום בהתאמה ותוקן עם שיעור גילוי השווא (FDR).
הערה: השתמשנו בשלבים של 88 אלפיות השנייה של חלונות לא חופפים במבחן Wilcoxon.
- בצע התמרת פורייה מהירה לזמן קצר (FFT) (1 הרץ עד 40 הרץ) באמצעות אורך חלון של 250 אלפיות השנייה וצעד זמן של 5 אלפיות השנייה. השתמש בחלון Hanning. השתמש בציון z כדי לנרמל את תרשימי תדירות הזמן לקו הבסיס (-1 שניות עד -0.1 שניות).
- שחזור מקור
- השתמש בארגז כלים לשחזור ניתוח מקורות, כגון ארגז הכלים של הגישה הפתוחה המתואר ב-Tadel et al.131 או ארגז כלים דומה אחר.
- חשב את מקורות אות ה-EEG המעובד מראש מ-4 השניות הראשונות של התכנון באמצעות אלגוריתם כגון טומוגרפיה אלקטרומגנטית סטנדרטית ברזולוציה נמוכה של המוח (sLORETA)132ושיטת הדמיה מינימלית-נורמה, כמו גם שיטת אלמנט הגבול הסימטרי (BEM סימטרי), בעזרת ארגז כלים כמו זה המתואר ב-Gramfort et al.133כדי לפתור את הבעיה ההפוכה.
- השתמשו באלגוריתם המקור (אלגוריתם sLORETA במחקר זה) על תבנית MNI אנטומית (השתמשנו בתבנית MNI בסיעור מוחות "Colin27") עם מיקומי האלקטרודות המוגדרים כברירת מחדל עבור כל משתתף למקרה שאין דיגיטציה תלת-ממדית של האלקטרודות (ראו שלב 5.6).
הערה: יש לציין כי שימוש במיקומי האלקטרודות המוגדרים כברירת מחדל אינו השיטה היעילה ביותר לקביעת מקורות הפעילות המוחית. עם זאת, זה עדיין יכול לספק הבנה כללית של מקור הפעילות. חשוב לזכור שמקורות הלוקליזציה המתקבלים בשיטות אלה הם קירובים גסים ויש לפרש אותם בזהירות במהלך ניתוח התוצאות. - החל מסנן פס פס של 4-8 הרץ על האות המעובד מראש.
- החל נורמליזציה של ציון z באמצעות התקופה -1,000 אלפיות השנייה עד -10 אלפיות השנייה לפני תחילת הניסוי כקו הבסיס.
- ממוצע של פעילות התטא באמצעות חלון זמן של עניין בין 1 שניות ל-4 שניות לאחר תחילת הניסוי.
- השווה את מקורות הרווח הממוצעים בין התנאים באמצעות מבחן סימן תמורות לא פרמטרי עם דגימת מונטה קרלו (1,000 אקראיות)131.
- על מנת לקבוע את אזורי העניין (ROI), סמנו את קליפת המוח באמצעות אטלס מוח.
הערה: השתמשנו ב-DestrieuxAtlas 134 המיושם בארגז הכלים המתואר ב-Tadel et al.131. - בחרו את אזורי העניין במוח (ROI).
הערה: שקלנו את הראיות המדווחות על כך שאזורי קליפת המוח הקדם-מצחית, כגון הג'ירי המצחי העליון הדו-צדדי (SF), גירי פרונטו-קוטבי רוחבי דו-צדדי (FP), ACC דו-צדדי, MCC דו-צדדי וקליפת המוח הקדם-מצחית הדורסולטרליתהדו-צדדית 137,138, מעורבים בתפקודי בקרה קוגניטיביים1 35,136. - בצע ניתוח רכיבים עיקריים (PCA) על פני אות ה-EEG הקודם שעובד מראש (טווח של 1-40 הרץ) עבור כל החזר ROI, וקח את המצב הראשון של פירוק ה-PCA עבור כל החזר ROI.
- בצע ניתוח ספקטרלי באמצעות התמרת פורייה מהירה לזמן קצר, והשווה את התוצאות בין אזורי העניין השמאלי והימני באמצעות מבחן תמורות לא פרמטרי מבוסס אשכול130.
- חלץ וייצג את החזר ה-ROI השמאלי והימני המראה שאין הבדלים כסדרת זמן דו-צדדית אחת: SF, ACC ו-MCC. לאחר מכן, התווה תרשימי תדירות זמן והשווה בין התנאים.
- השווה את תרשימי תדירות הזמן בהתאם לרמת המורכבות של משימת התכנון (ניסיונות קלים לעומת ניסויים קשים) עבור כל החזר ROI.
- שקף את קצה האות עבור כל אחת מ-512 הדגימות, ובצע מסנן פס פס בין 4 הרץ ל-8 הרץ עבור החזר ה-ROI שנבחר.
- החל התמרת הילברט כדי להשיג את המשרעת המיידית139באמצעות ארגז כלים לעיבוד אותות מתוכנת שפת תכנות (ראה טבלת חומרים).
- תקן את האות באמצעות נורמליזציה של ציון z (-1,000 עד -10 אלפיות השנייה כקו בסיס), וממוצע על פני הניסויים לפי נבדק.
- השווה כל פרופיל זמן של פס תטא בין התנאים באמצעות מבחן הדירוג החתום של Wilcoxon (זוגות תואמים, 1 שניות של חלונות לא חופפים), ותקן עם ה-FDR.
- מתאמים בין פעילות EEG לביצועים התנהגותיים
- נרמל את סדרות זמן המקור של החזר ה-ROI לקו הבסיס באמצעות ניקוד z. בחר חלון מ-1 שניות עד 4 שניות לאחר תחילת התכנון או הבקרה (כאשר נצפית פעילות תטא בולטת בתרשימי תדירות הזמן).
- כדי לקבוע את הגידול בפעילות התטא בתנאי התכנון בהשוואה לתנאי הבקרה, ראשית המירו את האות לתחום התדרים (1-40 הרץ) באמצעות שיטת multitaper באמצעות ארגז כלים כגון ארגז הכלים של Chronux140 עבור כל תנאי ומקור באזורי העניין.
- חשב את התדר הממוצע של פס התטא (4-8 הרץ), וחשב שני מדדים של הספק תטא: i) ההפרש בין כוח התטא במהלך תקופת התכנון (θ תכנון) לתקופת הבקרה (θ control), המסומן כ-Δ theta, ו-ii) הגידול היחסי בפעילות התטא, המבוטא כיחס בין Δ תטה (Δ θ) ופעילות התטא במהלך תקופת הבקרה (θ control), כמו ב-Domic-Siede et al.54:
 (1)
(1)

- חישוב שני פרמטרים התנהגותיים: 3) Δ תכנון LISAS, על ידי הפחתת בקרת LISAS מתכנון LISAS, ו-4) Δ ביצוע תכנון LISAS, על ידי הפחתת ביצוע בקרת LISAS מביצוע התכנון של LISAS, כמו ב-Domic-Siede et al.54:


- בצע את מתאמי ה-rho של ספירמן באמצעות הפרמטרים האלקטרופיזיולוגיים וההתנהגותיים המחושבים, ולאחר מכן תוקן על ידי ה-FDR.
- ניתוח תנועות עיניים: כדי לשלוט בהבדלים הפוטנציאליים בתנועות העיניים עבור כל מצב שעלול לגרום לדינמיקה תנודתית שונה, בצע את הניתוח הבא:
- קבע את משרעת הסקדה ואת מהירות השיא של הסקדה מכל הניסוי ומ-0 שניות עד 3.75 שניות במהלך תנאי התכנון והבקרה.
- השווה את התוצאות באמצעות מבחן הדירוג החתום של Wilcoxon או מבחן ה-t של הזוג המותאם, לפי המתאים.
הערה: ארגז כלים כמו זה המתואר ב-Dimigen et al.123יכול להועיל. - לחשב ולהעריך את הקוהרנטיות בין עוצמת ה-EEG של פורייה באלקטרודה קדמית אחת (למשל, ה-Fz או אלקטרודת ROI קדמית ממוצעת) לבין קצב ה-saccade כמתואר ב-Sato and Yamaguchi141.
- השתמש במבחן הדירוג החתום של Wilcoxon כדי להשוות את ערכי קצב ההספק-סקאד הקוהרנטי של 4 השניות הראשונות של כל ניסוי בין שני התנאים.
תוצאות
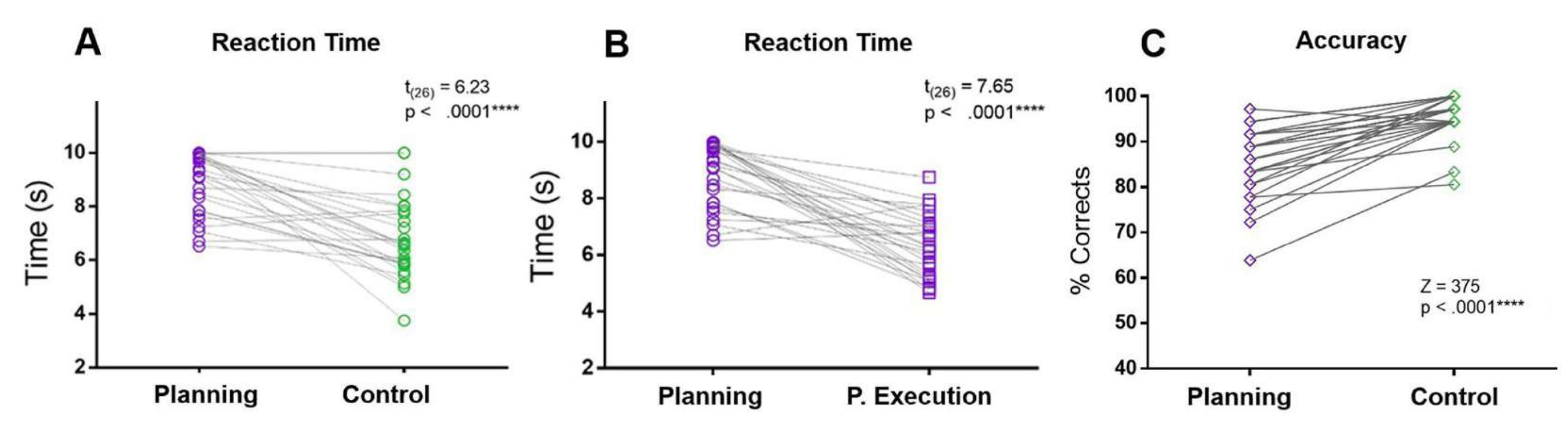
בפרוטוקול הנוכחי, ה-RT של תקופת התכנון הושווה ל-RTs של תקופת הבקרה ותקופת ביצוע התכנון. ה-RT התכנוני היה גדול יותר מה-RTs של הבקרה ותקופת הביצוע של התכנון. בנוסף, בהשוואה לתנאי הבקרה, המשתתפים עשו יותר טעויות והיו בעלי דיוק נמוך יותר במהלך תקופת התכנון (איור 5).

איור 5: זמן תגובה ודיוק למשימת התכנון. השוואה בין זמני התגובה (A) בתקופת התכנון (עיגולים סגולים) לתקופת הביקורת (עיגולים ירוקים) באמצעות מבחן t מותאם. (ב) השוואה בין זמני התגובה בתקופת התכנון (עיגולים סגולים) לתקופת ביצוע התכנון (ריבועים סגולים) באמצעות מבחן t מותאם. (ג) השוואה בין שיעור הדיוק בתנאי התכנון (יהלומים סגולים) לבין תנאי הבקרה (יהלומים ירוקים) באמצעות מבחן דירוג חתום של וילקוקסון. נתון זה שונה מ-Domic-Siede et al.54. אנא לחץ כאן לצפייה בגרסה גדולה יותר של איור זה.
{kind=link}
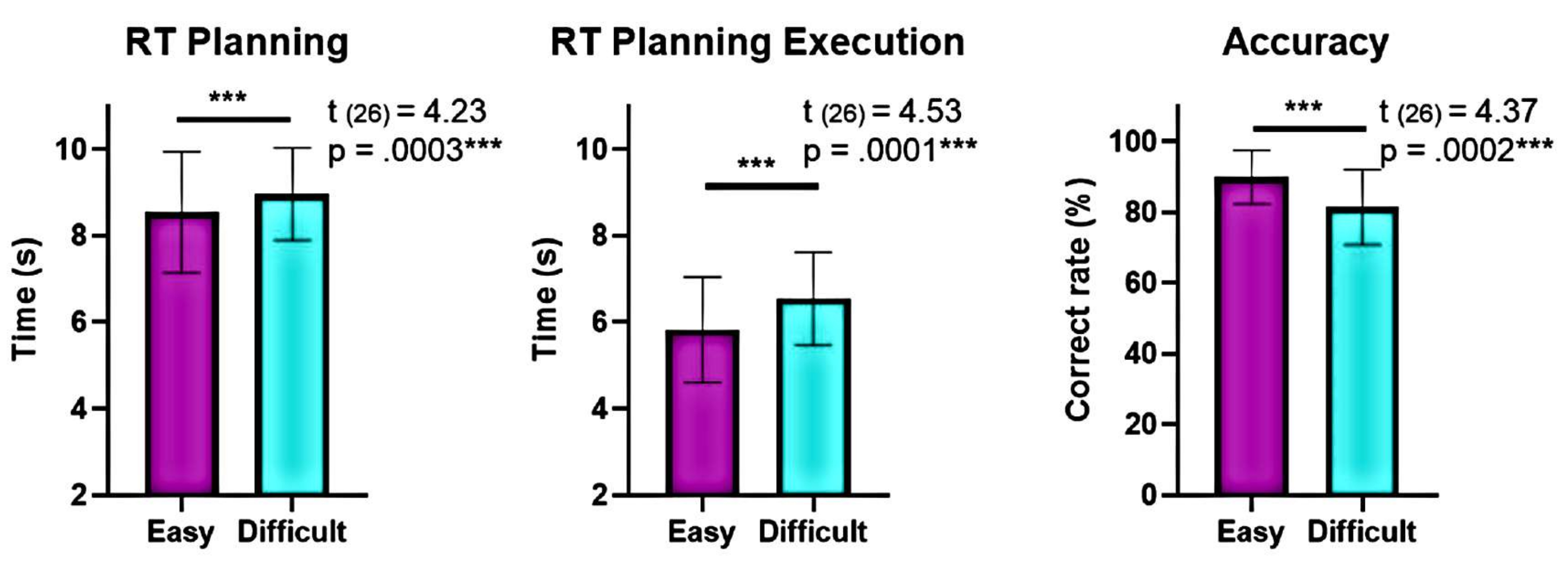
יתר על כן, ניתוח רמות המורכבות התכנונית הראה הבדלים משמעותיים ברמת הדיוק וזמני התגובה (RTs) בין הרמות ה"קשות" וה"קלות" במהלך התכנון והביצוע (איור 6). לרמה ה"קשה" היו RTs ארוכים יותר ודיוק נמוך יותר. ממצאים אלה מצביעים על כך שחלוקת הניסויים על סמך מספר הפתרונות התקפים יכולה להבחין בין ניסויים "קלים" ל"קשים".

איור 6: השוואה של ביצועים התנהגותיים ברמות המורכבות השונות. הבדלים משמעותיים בביצועים התנהגותיים ברמות המורכבות "הקלות" וה"קשות" זוהו באמצעות מבחן t של זוג תואם. זמני תגובה נמוכים יותר (RTs) נצפו במהלך התכנון והביצוע עבור הרמה ה"קלה" בהשוואה לרמה ה"קשה", והדיוק היה גבוה יותר עבור הרמה ה"קלה". קווי השגיאה מייצגים את ה- SEM (שגיאת תקן של הממוצע). נתון זה שונה מ-Domic-Siede et al.54. אנא לחץ כאן לצפייה בגרסה גדולה יותר של איור זה.
{kind=link}
תוצאות אלה הצביעו על כך שכאשר מרכיב התכנון הוסר בהצלחה ממצב הבקרה (באמצעות מניפולציה של הוראות), משימת התכנון הייתה מורכבת יותר מבחינה קוגניטיבית, תובענית, מאתגרת וגוזלת זמן. לפיכך, ניתן להשוות את המתאמים העצביים המושרים על ידי המשימות זה לזה.
כדי לנתח את פעילות התטא בקו האמצע הקדמי במהלך התכנון, פס תדר התטא הממוצע במהלך התכנון של אלקטרודת Fz הושווה לזה של תקופת הביקורת, ונמצאה עלייה משמעותית בתדר פס התטא במהלך התכנון (איור 7).

איור 7: פעילות תטא בקו האמצע הקדמי במהלך תכנון קוגניטיבי. (A) מפות טופוגרפיות שמייצגות את עוצמת פס התטא בכל הנבדקים מנורמלות לציוני z במהלך מטלת התכנון (משמאל), מטלת הבקרה (באמצע) ואפקט התכנון (מימין). במהלך התכנון הקוגניטיבי, הנבדקים הראו עלייה בפעילות תטה בקו האמצע הקדמי. סרגל הצבעים מציג את ערכי ה-z בין -0.5 ל-1.5. (B) תרשים כינור שמראה את ערכי המינימום, הרביעונים, החציון והמקסימום של ניקוד z של פעילות תטא בקרב הנבדקים במהלך התכנון (סגול) בהשוואה לתקופת הבקרה (ירוק) עבור האלקטרודות Fz (משמאל), Pz (באמצע) ו-Oz (מימין) באמצעות מבחן t של זוג תואם. נתון זה שונה מ-Domic-Siede et al.54. אנא לחץ כאן לצפייה בגרסה גדולה יותר של איור זה.
{kind=link}
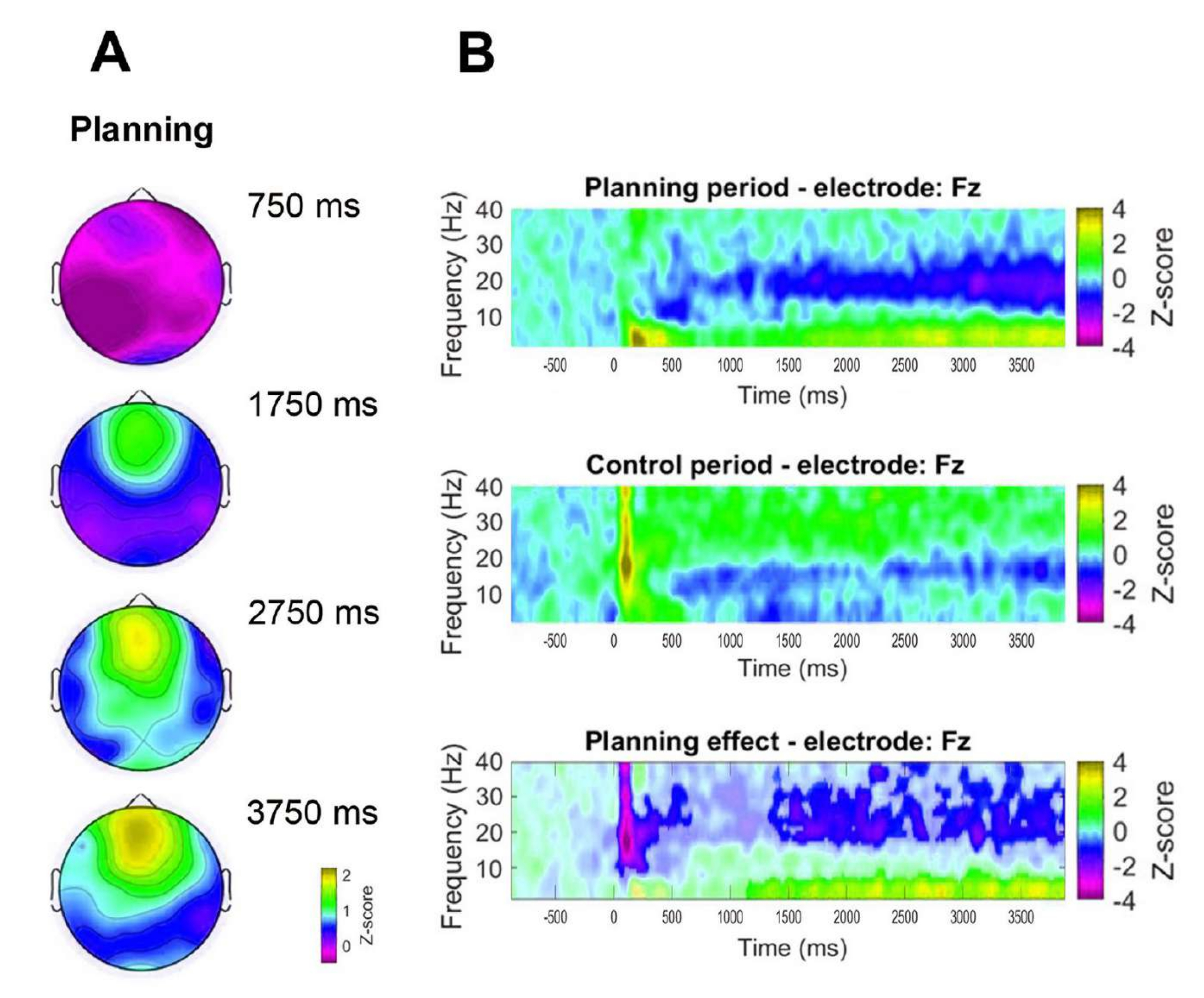
בנוסף, כדי להעריך את הדינמיקה הזמנית של פעילות התטא הקדמית שנצפתה, נוסחו מפות טופוגרפיות המתאימות לנקודות זמן ספציפיות של עוצמת פס תטא (750 ms, 1,750 ms, 2,750 ms ו-3,750 ms) (איור 8A). יתר על כן, בהשוואה לתקופת הביקורת, ניתוח תדירות הזמן הראה עלייה משמעותית, מתקדמת ומתמשכת בפעילות התטא החל משנייה אחת לאחר תחילת תקופת התכנון (איור 8B).

איור 8: דינמיקה טמפורלית של תטא בקו האמצע הקדמי. (A) פרוסות זמן טופוגרפיות של פעילות התטא. נצפתה עלייה הדרגתית בפעילות התטא של קו האמצע הקדמי לאורך זמן במהלך יישום התכנון (תקופת התכנון). סרגל הצבעים מציין את יחידות ציון z (-0.5 עד 2.2). (ב) תרשימי תדירות זמן לתקופת התכנון (למעלה), תקופת הבקרה (אמצעית) וההשפעה התכנונית, המחושבים על ידי הפחתת תקופת הבקרה מתקופת התכנון (למטה). פיקסלים לא משמעותיים, כפי שנקבעו באמצעות מבחן תמורות מבוסס אשכול לא פרמטרי עבור דגימות זוגיות, מוצגים קלים יותר בתרשים אפקט התכנון. סרגל הצבעים מציין את יחידות ניקוד z (-4 עד 4). נתון זה שונה מ-Domic-Siede et al.54. אנא לחץ כאן לצפייה בגרסה גדולה יותר של איור זה.
{kind=link}
עבור שחזור מקור של פעילות התטא, תבנית מודל מוחי הומחשה והושוותה בין מצבים, וזה הצביע על כך שפעילות התטא מקורה באזורי קליפת המוח הקדם-מצחית (קליפת המוח העליונה הקדמית, FS; פיתול החגורה הקדמי, ACC; וקליפת המוח האמצעית, MCC), כמו גם שהיו הבדלים משמעותיים בין התנאים (ב-SF הדו-צדדי, ה-ACC הדו-צדדי וה-MCC הדו-צדדי) (איור 9), עם פעילות תטא גבוהה יותר שנצפתה בתקופת התכנון (איור 9).

איור 9: שחזור מקורות. נעשה שימוש באלגוריתם sLORETA כדי להעריך את פעילות התטא ממקורות מוח שונים. פעילות התטא הייתה מסוננת של 4-8 הרץ, ציון z מנורמל, מתוקן לפי קו הבסיס, ממוצע בין 1 שניות ל-4 שניות לאחר תכנון או התחלת בקרה, בהתאמה, והושווה בין תנאים. עלייה משמעותית בפעילות התטא נמצאה באזור העליון המצחי הדו-צדדי, בקליפת המוח החגורה הקדמית הדו-צדדית ובקליפת המוח הדו-צדדית האמצעית. האיור מציג ערכי t משמעותיים ממבחן התמורה. קיצורים: FS = עליון פרונטלי; ACC = פיתול החגורה הקדמי; MCC = קליפת המוח האמצעית. נתון זה שונה מ-Domic-Siede et al.54. אנא לחץ כאן לצפייה בגרסה גדולה יותר של איור זה.
{kind=link}
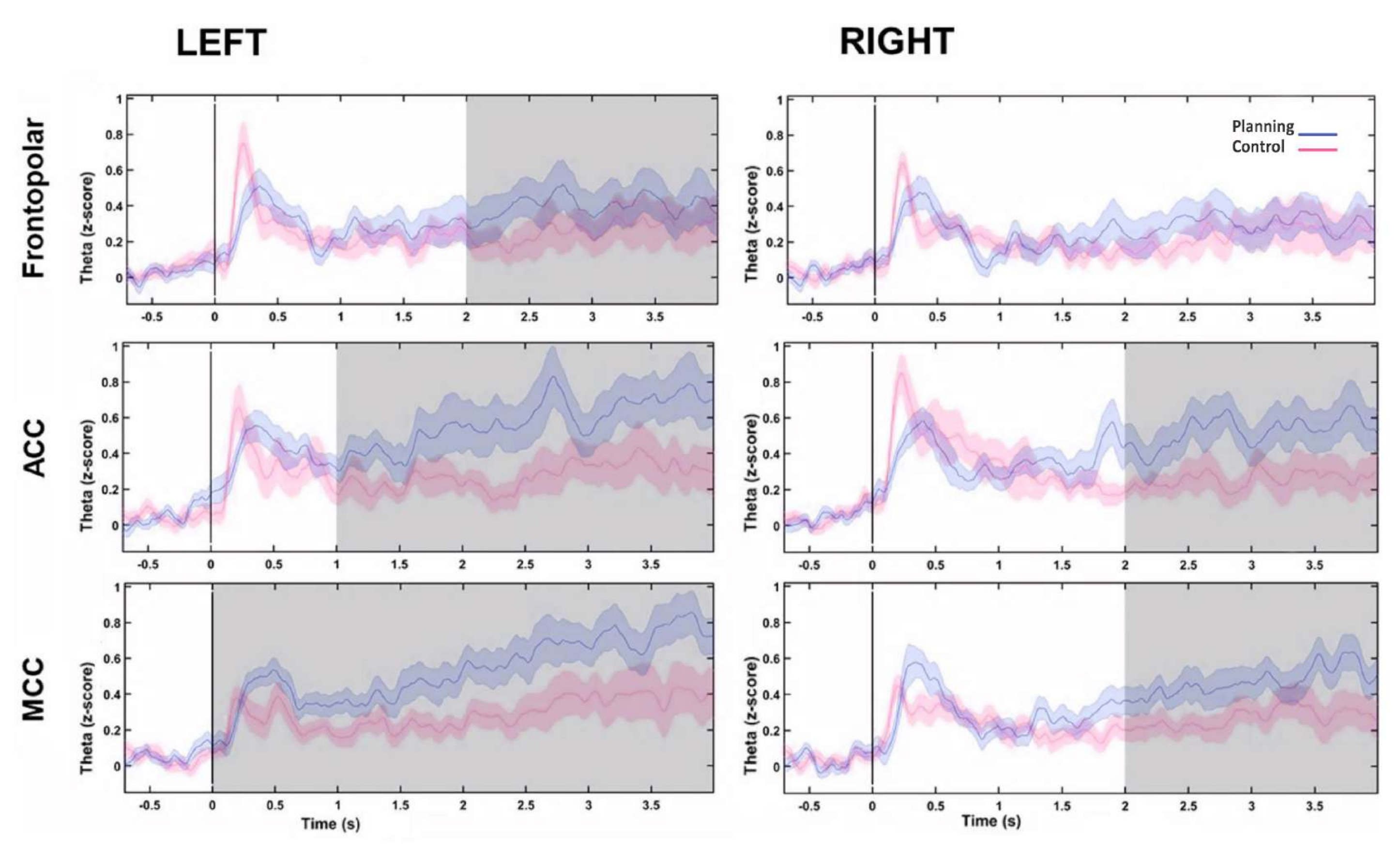
לאחר מכן, פרופיל הזמן של התטא משתנה לאורך זמן עבור כל מקור הוערך על ידי חישוב התמרת הילברט, ולאחר מכן השווינו את המשרעת המיידית של פעילות התטא בין התנאים. מצאנו שמקורות ה-ACC הקדמיים השמאליים, הדו-צדדיים וה-MCC הדו-צדדיים הציגו פעילות תטא גבוהה יותר לאחר תחילת תקופת התכנון (איור 10). תוצאות אלה הציעו כי פרדיגמת הניסוי שלנו הדורשת תכנון קוגניטיבי גרמה לפעילות תטא שמקורה באזורי קליפת המוח הקדם-מצחית.

איור 10: פרופיל זמן פעילות Theta של מקורות PFC. המשרעת המיידית שחושבה עם התמרת הילברט הוחלה על הרכיב הראשון של פירוק ה-PCA עבור כל מקור קדמי ושני התנאים וקו הבסיס נורמלו לציון z כדי להראות את פעילות התטה הקדמית לאורך זמן. האזורים האפורים המוצלים מראים הבדלים משמעותיים שנקבעו באמצעות חלון נע לא חופף עם שלבים של 1 שניות (מבחן Wilcoxon signed-rank) שתוקנו על ידי ה-FDR. האזורים המוצללים מייצגים רווחי בר-סמך של 95%. אזור ה-FP השמאלי, ה-ACC הדו-צדדי וה-MCC הדו-צדדי הראו עלייה בפעילות התטא לאחר תחילת התכנון. קיצורים: ACC = פיתול החגורה הקדמי; MCC = קליפת המוח האמצעית. התנאי התכנוני מוצג בסגול. מצב הבקרה מוצג באדום. נתון זה שונה מ-Domic-Siede et al.54. אנא לחץ כאן לצפייה בגרסה גדולה יותר של איור זה.
{kind=link}
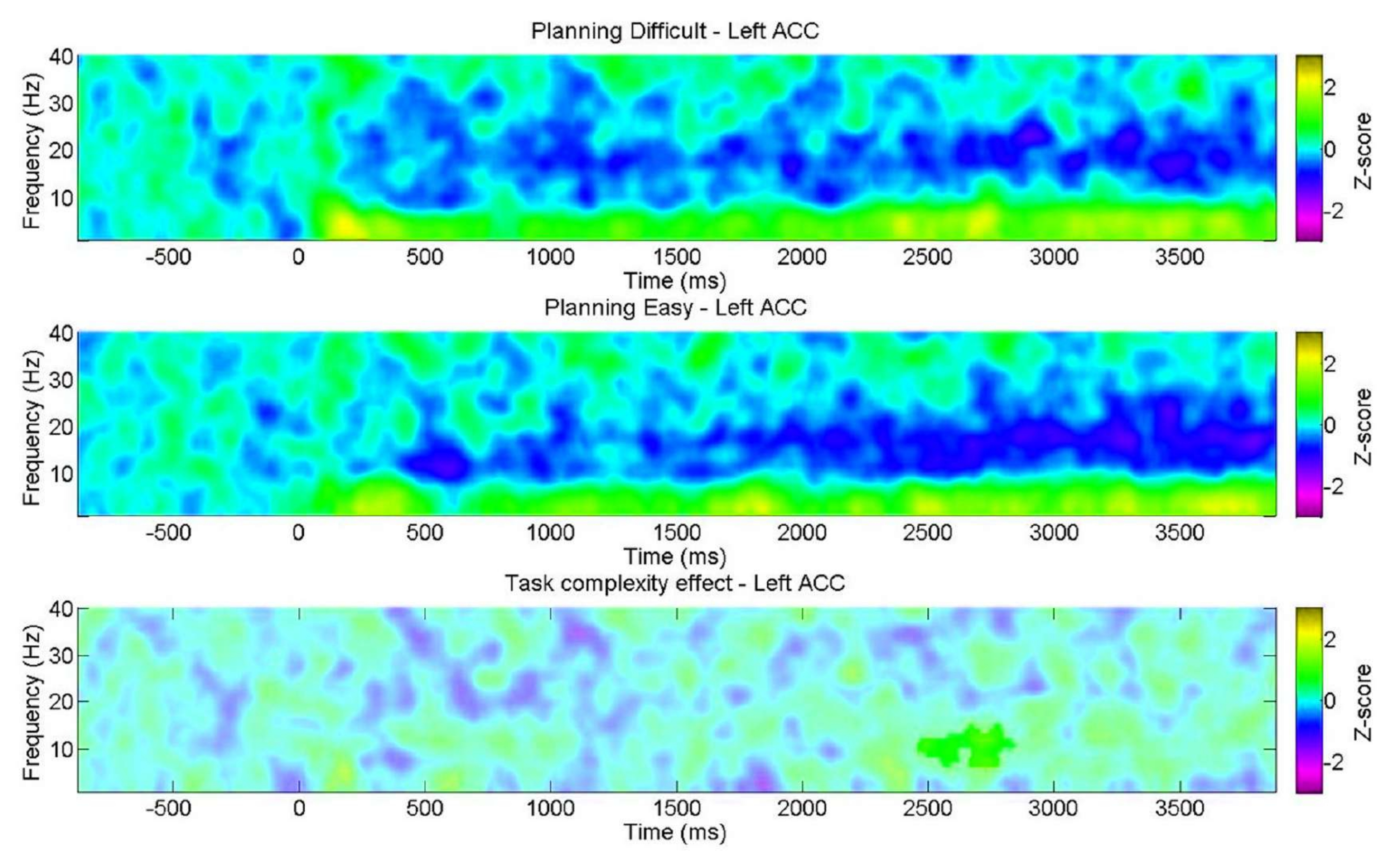
יתר על כן, מטרתנו הייתה לבחון את השונות בתכונות הספקטרליות במהלך התכנון ביחס לרמת המורכבות, כפי שעולה מהתוצאות ההתנהגותיות. יש לציין כי אי התאמה משמעותית נמצאה רק ב-ACC השמאלי בתוך רצועת האלפא. זה תומך ברעיון שמשימת התכנון שלנו מעריכה את ההיבטים הפנימיים של התכנון באמצעות שינויים בתנודות תטא במידה רבה יותר מאשר הדרישות הקוגניטיביות הכלליות (מאמץ) שבדרך כלל נתקלים בהן במשימות בקרה קוגניטיביות (איור 11).

איור 11: EEG על פני רמות המורכבות של התכנון. תרשימי תדירות הזמן של החזר ההשקעה הראו אשכול חיובי משמעותי ברצועת האלפא אך ורק בפיתול החגורה הקדמי השמאלי (ACC) לרמה "הקשה". פיקסלים לא משמעותיים, כפי שנקבעו באמצעות מבחן תמורות מבוסס אשכול לא פרמטרי עבור דגימות זוגיות, מוצגים בגוון בהיר יותר על העלילה, כאשר סרגל הצבעים מציין את יחידות ציון z מ-3 עד 3. נתון זה שונה מ-Domic-Siede et al.54. אנא לחץ כאן לצפייה בגרסה גדולה יותר של איור זה.
{kind=link}
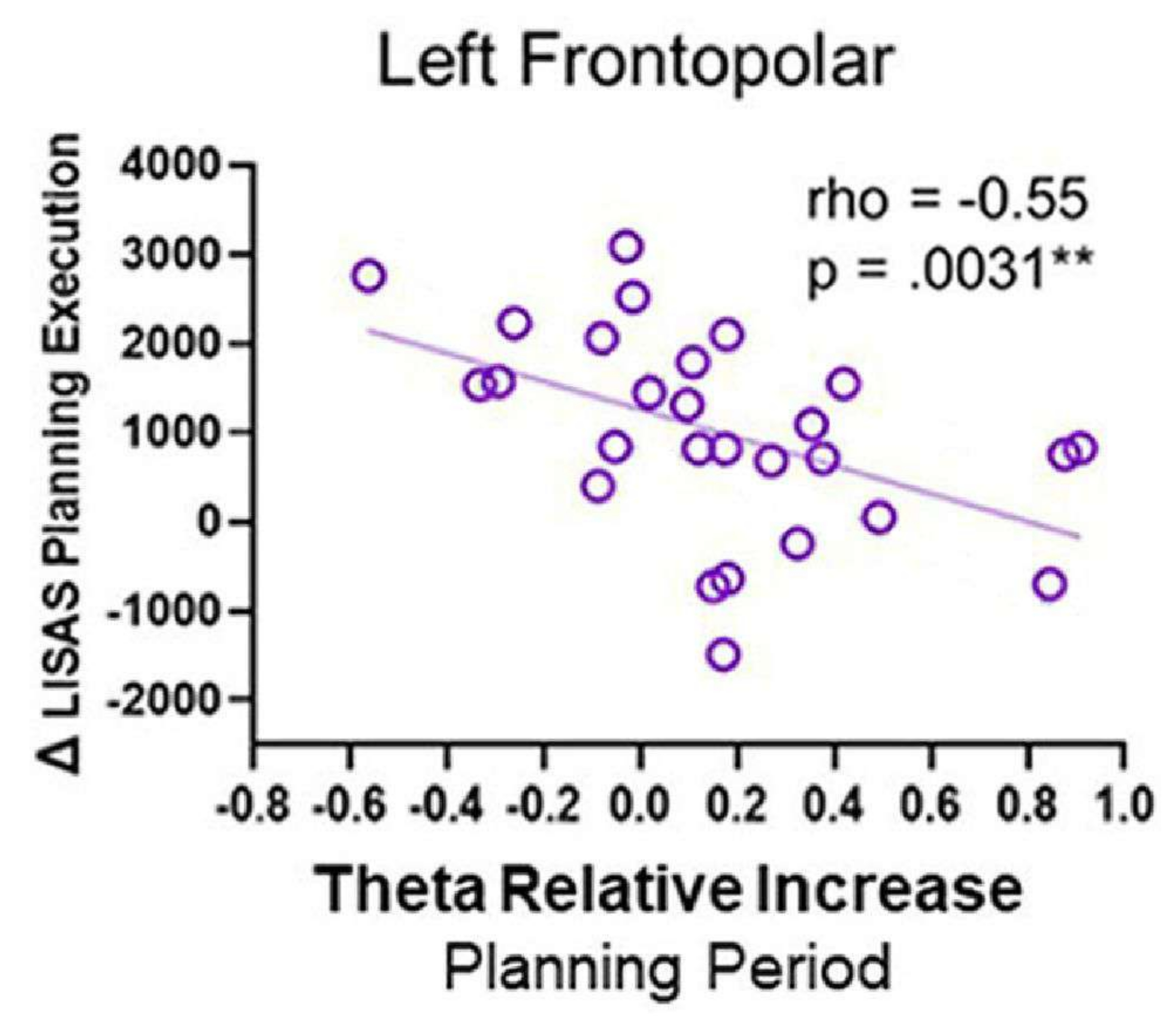
כאשר בוצעו מתאמים בין פעילות תטא לביצועים התנהגותיים, נצפה מתאם שלילי; באופן ספציפי, ככל שפעילות התטא באזור הקדמי השמאלי במהלך תקופת התכנון גדלה, ציון ביצוע התכנון של LISAS ירד (איור 12). דפוס זה עשוי לשקף שאזור ה-FP השמאלי עשוי להיות נחוץ במהלך פיתוח התכנון כדי לבצע תוכנית בהצלחה לאחר מכן ומציע תפקיד לפעילות תטא.

איור 12: פעילות תטא וביצועים התנהגותיים. מתאם ה-rho של ספירמן בין פעילות התטא מקליפת המוח הקדמית השמאלית לבין ביצוע התכנון של Δ LISAS הראה מתאם שלילי משמעותי. נתון זה שונה מ-Domic-Siede et al.54. אנא לחץ כאן לצפייה בגרסה גדולה יותר של איור זה.
{kind=link}
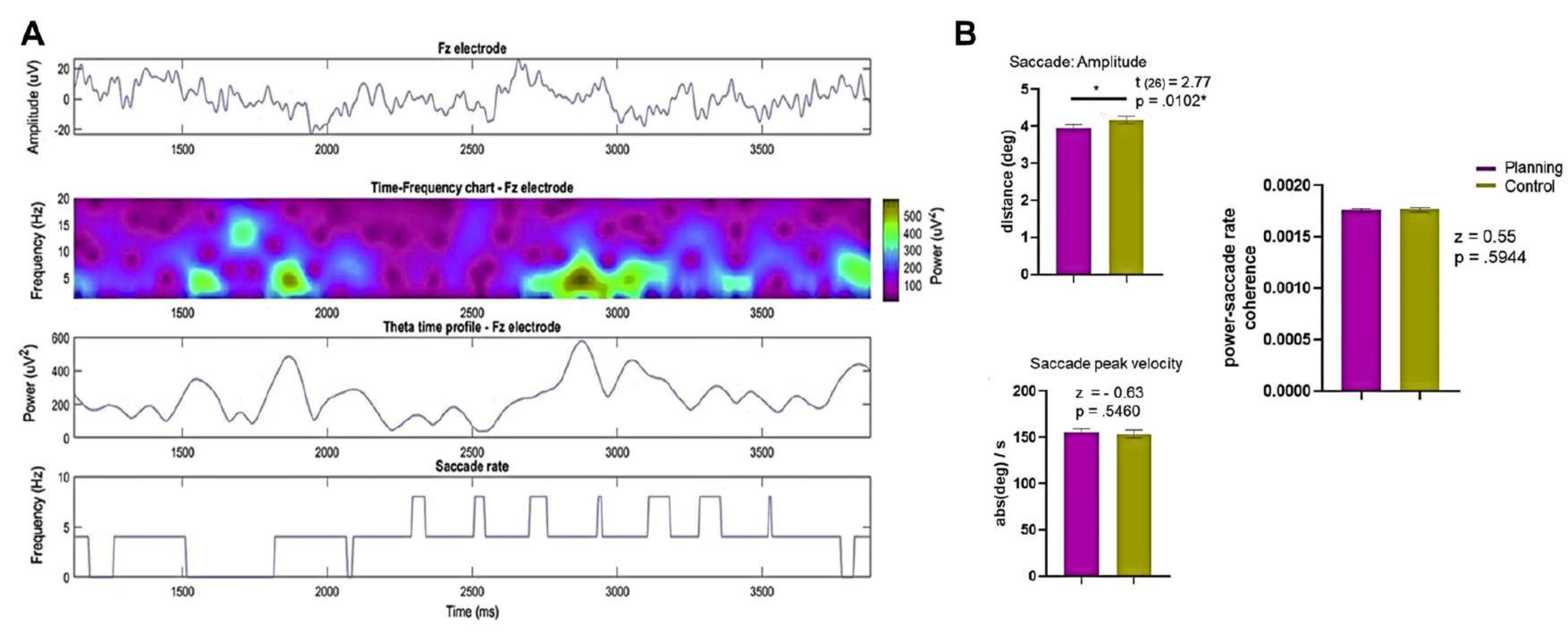
בנוסף, הדרישות והמטרות הקוגניטיביות המשתנות שנגרמו על ידי כל מצב עשויות לגרום לתנועות עיניים מנוגדות בין תנאי התכנון והבקרה, מה שהוביל לדפוסי פעילות תנודתיים שונים95. כדי לטפל בבעיה הנ"ל, ניתחנו את נתוני הנבדק היחיד והניסוי היחיד ברמות שונות. יש לציין כי נראה כי לסדרות הזמן של ערוץ Fz ולדינמיקת הזמן של פעילות התטא אין קשר לקצב הסקאדות לאורך זמן (איור 13A).

איור 13: תוצאות של רישומי EEG ותנועות עיניים. (A) השורות מציגות את ה-EEG (למעלה), את תרשים הזמן-תדר (למעלה האמצעי), את פרופיל זמן התטא מהאלקטרודה Fz (תחתון אמצעי), ואת קצב ה-saccade של נבדק 8 וניסוי 9 (למטה) בתנאי התכנון. (B) מוצגת השוואת מבחן בדרגה חתומה של Wilcoxon של משרעת ה-saccade, מהירות השיא של ה-saccade וקוהרנטיות קצב ההספק-saccade בין התנאים, וחושפת הבדלים סטטיסטיים משמעותיים במשרעת ה-saccade בין תנאי התכנון והבקרה. ה- SEM מיוצג על ידי קווי השגיאה. נתון זה שונה מ-Domic-Siede et al.54. אנא לחץ כאן לצפייה בגרסה גדולה יותר של איור זה.
{kind=link}
לאחר מכן, השגנו את משרעת הסקדה ומהירות השיא מכל הניסוי ומ-0 שניות ל-3.75 שניות להשוואה (איור 13B). גילינו שמשרעת הסקדה הייתה גדולה יותר בתנאי הביקורת. עם זאת, לא נמצאו הבדלים מובהקים סטטיסטית בין התנאים במדד הקוהרנטיות בין כוח התטה של פורייה באלקטרודה Fz לבין קצב הסקדה (איור 10B), מה שמצביע על כך שכל קשר פוטנציאלי בין סקאדות לפעילות תטא היה עקבי בין תנאים.
יחד, תוצאות אלה תומכות בכך שפרוטוקול הניסוי המתואר מתאים לחקר תכנון קוגניטיבי כפונקציית בקרה קוגניטיבית.
Discussion
הפרוטוקול המתואר כאן מציע גישה חדשנית להערכת תכנון קוגניטיבי ושליטה קוגניטיבית במהלך משימת תכנון אקולוגית חדשה בשילוב עם מדידות התנהגותיות ופיזיולוגיות רלוונטיות ומשלימות, כגון דינמיקה תנודתית ואישונים. במהלך הניסויים שלנו, פעילות EEG תועדה בזמן שהמשתתפים ביצעו את מטלת התכנון, שבה המשתתפים הונחו תחילה לפרט ולאחר מכן לבצע תוכנית. תנאי הבקרה, שכלל הערכת נתיב משורטט מראש על מפת גן החיות, נקבע כדי לבטל את היבט התכנון הקוגניטיבי תוך שמירה על סביבה ומבנה דומים. גישה זו מאפשרת להעריך האם תכנון קוגניטיבי, כפונקציית בקרה קוגניטיבית, מוביל ליצירת פעילות תטא פרונטלית מאזורי קליפת המוח הקדם-מצחית והאם מקורות תנודות תטה שונים של קליפת המוח הקדם-מצחית קשורים להיבטים שונים של ביצועי תכנון. היבט נוסף שניתן להעריך באמצעות פרוטוקול זה הוא ההבחנה בין התהליכים הקוגניטיביים השונים המעורבים במהלך שלבי התכנון, כגון פיתוח תוכנית, ביצוע תוכנית, השגת תוכנית ועיבוד משוב. מצאנו שתכנון גרם לפעילות תטא קדמית קאנונית הקשורה לשליטה קוגניטיבית, שתרמה להשגת מטרה ביעילות. תוצאות אלה מחזקות את העניין של פרוטוקול זה.
למרות ההתפתחויות הנרחבות במדעי המוח הקוגניטיביים, רוב ניסויי ההדמיה המוחית בוחנים תפקודים קוגניטיביים מבודדים באמצעות משימות מלאכותיות בסביבות חסרות תחושה מוטורית וגירויים פשוטים מדי כדי לשלוט בבלבולים. באופן בעייתי, ייתכן שניסויים אלה לא יוכלו לזהות את המנגנונים המוחיים האמיתיים המעורבים כאשר פונקציה קוגניטיבית מיושמת בסיטואציה יומיומית (במהלך מצבים אקולוגיים)138,140. בפרט, קשה להעריך את היכולות לניסוח מטרות, תכנון וביצוע תוכניות ביעילות מכיוון שהן דורשות פונקציות קוגניטיביות שונות (זיכרון עבודה, שליטה מעכבת, גמישות קוגניטיבית וכו').104,144. לפיכך, עיצוב משימה התנהגותית אקולוגית מעודד ומוצע על סמך המגמות הנוכחיות במדעי המוח הקוגניטיביים 142,143,145,146.
משימת התכנון שלנו, למרות שהתרחשה בסביבת מעבדה (בתוך חדר עם הגירויים המוצגים על מסך), נעשתה מרתקת ואינטראקטיבית עבור המשתתפים באמצעות שימוש בגירויים משמעותיים ומטרות שהם יכלו לתקשר איתם על המסך. בנוסף, המשימה דורשת מהמשתתפים לעסוק בתכנון מצב בחיים האמיתיים - נתיב לביקור במקומות שונים. כדי לעצב משימה אקולוגית, הפרדיגמה חייבת לאתגר את הנבדקים לבצע התנהגות או תפקוד קוגניטיבי ספציפי באופן דומה למה שהם יצטרכו לעשות בחיי היומיום 62,63,147. כדי לפתח תכנון משימה אקולוגית, משימת התכנון המשמשת כאן כוללת תכנון נתיב לביקור במקומות שונים בכמה שלבים56. השלב הראשון כולל את המשתתפים ביצירת תוכנית תוך הקפדה על עמידה במערכת כללים. השלב השני כולל שמירה על התוכנית בזיכרון העבודה, ואילו השלב השלישי כולל ביצוע התוכנית ומעקב אחר עמידתה בכללים. שלבים אלה מייצגים שלבים שונים של תכנון ותזמור של פונקציות ניהוליות אחרות, כגון גמישות קוגניטיבית, שליטה מעכבת וזיכרון עבודה. על מנת שיהיה תכנון משימה קוגניטיבי אקולוגי תקף, המשימה צריכה להיות מסוגלת לזהות ליקויים קוגניטיביים ספציפיים בחולים עם הפרעות פסיכיאטריות או קוגניטיביות המתקשים לבצע את הפונקציה הקוגניטיבית הספציפית הזובחיי היומיום שלהם. ניתן להשיג זאת באמצעות מחקר עתידי באמצעות פרוטוקול זה.
התוצאות ההתנהגותיות שהושגו באמצעות שימוש בפרוטוקול זה היו תואמות לתחזיות הניסוי. הבדל משמעותי בהתנהגות נצפה כאשר מרכיב התכנון הוסר ממשימת הבקרה כדי ליצור מצב בקרה, ובכך הקל על השוואות נוספות. התנאי התכנוני נמצא תובעני יותר מבחינה קוגניטיבית מתנאי הביקורת, כפי שמעידים פרמטרים כמו זמן התגובה והדיוק. זה עשוי לשקף את המעורבות המוגברת של תפקודים קוגניטיביים גבוהים ביישום התכנון 23,55,56,57,148,149,150.
בהתחשב בכך שתנאי הבקרה היה כרוך בקוגניציה פחות מורכבת, כפי שמעידים זמני התגובה המהירים יותר, הביצועים הטובים יותר והתהליכים הקוגניטיביים השונים הנדרשים (הערכת כללים), שינוי חלופי אפשרי הוא לנצל את רמות המורכבות הקיימות במשימת התכנון, לתמרן אותן ולנתח את פונקציית התכנון באופן פרמטרי על פי רמות מורכבות שונות (למשל, הגדלת מספר הניסיונות, ויצירת תנאי ניסוי קשים, בינוניים וקלים). עם זאת, תוצאות הפרוטוקול שלנו הראו כי בעוד שניתן היה להבחין בין ניסויים "קלים" ו"קשים" על סמך מדדים התנהגותיים, לא זוהו הבדלים במדדים אלקטרופיזיולוגיים. זה מצביע על כך שהתוצאות בפרוטוקול שלנו משקפות בצורה מדויקת יותר את המאפיינים הפנימיים של פונקציית התכנון ולא היבטים רחבים יותר של שליטה קוגניטיבית, כגון קשב, מאמץ מנטלי, רמת קושי או רמה גבוההשל דרישה קוגניטיבית. אף על פי כן, מחקר נוסף עשוי לשקול סוגים אחרים של תנאי בקרה, כגון מעקב אחר נתיב מסומן המבקר בארבעת המיקומים של בעלי החיים, אך גם לזכור את סדר הרצף. בדרך זו, ניתן היה לשלוט טוב יותר ברמת הקושי, ולהבחין בין תכנון לזיכרון עבודה, אך חסרון אפשרי אחד לכך הוא עייפות, מכיוון שהנבדקים יצטרכו לבצע שתי משימות תובעניות ביותר.
מספר מחקרים קישרו פרמטרים שונים של תנועת עיניים לאירועים קוגניטיביים ספציפיים. מצד אחד, מחקרים מסוימים מצאו מתאמים בין תנודות תטא לקוטר האישון במהלך מטלות קוגניטיביות, מה שמרמז על קשר בין שני המדדים הללו של תפקוד קוגניטיבי. לדוגמה, Lin et al.152 מצאו מתאם בין פעילות תטה אמצעית לשינויים בגודל האישון, המשקף דרגות שונות של קונפליקט סובייקטיבי. הממצאים שלהם מצביעים על כך שאותות אלה מייצגים עיבוד קונפליקטים, קשב מוגבר ותגובות התנהגותיות גמישות. לפיכך, נראה כי הקשר בין פעילות תטא אמצעית לתגובות התלמידים ממלא תפקיד בשקלול עלויות ותועלת במהלך תהליכי קבלת החלטות. במחקר אחר, Yu et al.153 בחנו כיצד מעורבות בזמן במשימה משפיעה באופן נוירופיזיולוגי על שליטה קוגניטיבית באמצעות משימת זיכרון עבודה המווסתת תגובות מעכבות שליטה. הם חקרו את הקשר בין נתוני קוטר האישון לפעילות התטא הקדמית והראו שככל שמשך המטלה גדל, הביצועים פחתו, וזה לווה בירידה באפנון הרחבת האישונים ובפעילות התטא הקדמית. בתחילת המשימה, הם מצאו מתאם חזק בין מעורבות במשימה, פעילות תטא ושליטה קוגניטיבית, כפי שמצוין על ידי אפנון הרחבת האישונים, בעיקר עבור משימות תובעניות שדרשו זיכרון עבודה גבוה ושליטה מעכבת. עם זאת, קשר זה התפוגג לקראת הסוף, וסימן ניתוק בין המאמץ המושקע לבין השליטה הקוגניטיבית המשמשת לביצוע המשימה, שהיא סימן היכר של השפעות זמן על משימה קדם-מצחית153. מצד שני, מחקרים אחרים חקרו סקאדות ותנודות. לדוגמה, Nakatani et al.154 גילו כי במשימה תפיסתית, משרעת פס האלפא מאזורי העורף ניבאה מצמוץ והשפעות סקאדה. יתר על כן, Velasques et al.155 הראו כי במהלך משימת קשב פרוסקדית, משרעת הסקדה הייתה קשורה לשינויי גמא פרונטליים. יתר על כן, Bodala et al.156 מצאו כי ירידות בתטה בקו האמצע הקדמי לוו בירידה בקשב מתמשך, כמו גם במשרעת ובמהירות של סקאדות. ממצאים אלה מצביעים על כך שתנועות עיניים, במיוחד סקאדות, עשויות לשקף תהליכים קוגניטיביים ולא רק לתרום לרעשי הרקע באותות EEG. במחקר הנוכחי, שיפרנו את ביטול החפצים הקשורים לתנועת העיניים באמצעות אלגוריתם ICA עם קריטריון יחס השונות של סקאד לקיבוע126. קריטריון זה משפר את הסרת החפצים למשימות צפייה בחינם157. במחקר שלנו, לא נצפו הבדלים במהירות השיא של הסקדה ובקוהרנטיות בין כוח תטא לקצב הסקדה בין תנאים. עם זאת, נדרשים מחקרים נוספים כדי לענות על שאלות אלה.
שלב קריטי בשימוש בפרוטוקול זה הוא כיול מתמיד של עוקב העיניים במהלך הניסוי, שכן אובדן נתוני המבט מהמצלמה עלול לגרום לשגיאות שיזהמו את המשימה ויקשו על קבלת תגובות מדויקות. לכן, חיוני לכייל בתדירות גבוהה ככל האפשר. עם זאת, יש פשרה בין מספר הניסויים עם עוקב עיניים מכויל לבין אורך הניסוי. במחקר שלנו, החלטנו לכייל כל חמישה ניסויים.
יש לערוך מחקר עתידי הבוחן את הקשר בין תנודות תטא וקוטר האישון במהלך משימת תכנון זו. תכנון הוא היבט קריטי של בקרה ניהולית הדורש הקצאת משאבי קשב ותיאום של תהליכים קוגניטיביים מרובים. הבנת הקשר בין תנודות תטא וקוטר האישון במהלך משימות תכנון יכולה לספק תובנות חשובות לגבי המנגנונים העצביים הבסיסיים של שליטה ניהולית וכיצד הם משתנים עם הזמן. יתר על כן, מחקרים כאלה יכולים להוביל להבנה מעמיקה יותר של האופן שבו שינויים בתפקוד הקוגניטיבי, כגון עייפות או הפרעות קשב, משפיעים על הביצועים במשימות תכנון ועל היכולת להקצות משאבים ביעילות. למידע זה עשויות להיות השלכות חשובות על פיתוח התערבויות שמטרתן לשפר את ביצועי התכנון, כגון תוכניות אימון קוגניטיביות או טיפולים במצבים כמו הפרעת קשב וריכוז (ADHD), בין היתר.
מחקרים קודמים הצביעו על כך שקליפת המוח הקדם-מצחית ממלאת תפקיד מכריע בתכנון קוגניטיבי, כפי שאושר על ידי התוצאות שלנו. תוצאות אלה מדגימות כי תכנון קוגניטיבי משרה פעילות FMθ בקליפת המוח הקדם-מצחית, במיוחד בקליפת המוח החגורה הקדמית, בקליפת המוח האמצעית ובאזורים הקדמיים העליונים54. ממצאים אלה עולים בקנה אחד עם מחקרים קודמים על תפקודים ניהוליים. ישנן ראיות משמעותיות התומכות ברעיון שפעילות FMθ פועלת כתהליך מלמעלה למטה ליזום שליטה ולהקל על תקשורת בין אזורי מוח במהלך משימות תובעניות13. בעוד שרק מחקרים מעטים בחנו את הדינמיקה הטמפורלית של פעילות FMθ הקשורה לשליטה קוגניטיבית, קיימת הסכמה רחבה שפרופיל הזמן של FMθ יכול לספק מידע על היבטים שונים של שליטה קוגניטיבית ומעורבות של אזורי PFC מובחנים. השימוש בפרוטוקול שלנו להערכת תכנון קוגניטיבי אפשר לנו לאפיין את פרופיל הזמן של פעילות FMθ במהלך התכנון. באופן ספציפי, פעילות FMθ במהלך תנאי התכנון הראתה עלייה הדרגתית. על ידי יישום פרוטוקול זה, בפעם הראשונה, הוכח כי FMθ, נוכח גם במהלך יישום התכנון, כמו בתפקודים קוגניטיביים אחרים מסדר גבוה יותר, והדינמיקה הטמפורלית שלו עשויה לשמש כאינדיקטור לשליטה קוגניטיבית.
לתוצאות ולפרוטוקול שלנו יש יישומים פוטנציאליים בתחום מדעי המוח, כולל לשיפור הערכות נוירופסיכולוגיות וירטואליות וטיפול בהפרעות נוירו-פסיכיאטריות עם בעיות תכנון קוגניטיביות נלוות, כגון דיכאון והפרעת קשב וריכוז. לדוגמה, הערכות יכולות לכלול בחינת דפוסים שונים של טעויות ברמת הביצועים ההתנהגותיים, דפוסי תנודה שונים ברמה האלקטרופיזיולוגית ותנועות עיניים שונות. בנוסף, תוצאות עבודה זו עשויות לתרום לפיתוח ממשקי מוח-מחשב ותוכניות אימון קוגניטיביות שמטרתן לשפר את יכולות התכנון הקוגניטיבי.
הפרוטוקול הנוכחי עשוי לתרום ראיות חדשות להבנת המנגנונים העצביים העומדים בבסיס פונקציית השליטה הקוגניטיבית החמקמקה של תכנון קוגניטיבי באוכלוסיות נוירוטיפיקליות ונוירו-פסיכיאטריות. יתר על כן, הפרדיגמה ההתנהגותית שלנו עשויה להציע תובנות לגבי הנוירוביולוגיה של שליטה ותכנון קוגניטיביים באמצעות בחינת מדידות אלקטרופיזיולוגיות, אישון והתנהגות עם משימת תכנון מעשית הבוחנת היבטים מהותיים של תכנון במקום הדרישות הקוגניטיביות הכלליות הקיימות בדרך כלל במשימות בקרה קוגניטיביות, כפי שבא לידי ביטוי בשינויים בתנודות התטא.
Disclosures
למחברים אין מה לחשוף.
Acknowledgements
מחקר זה נתמך כספית על ידי תוכנית המלגות לדוקטורט Becas de Doctorado Nacional año 2015 של ANID 21150295, 1180932 המענקים הרגילים של FONDECYT, 1230383 המענקים הרגילים של FONDECYT, מענק FONDECYT de Iniciación 11220009, מענק פוסט-דוקטורט Universidad de O'Higgins והמכון האוניברסיטאי של צרפת (IUF). אנו רוצים להודות לפרופסור פבלו בילקה על המשוב שלו על עיצוב הפרדיגמה. אנו מודים לפרופסור אאוג'ניו רודריגז על שיתוף הקודים שלו לניתוח תדר הזמן. לבסוף, אנו מודים למילאן דומיץ', ויסנטה מדל, ג'וזפינה איהנן, אנדראה סאנצ'ס, גונזלו בונקומפטה, קטלינה פאבר ודניאלה סנטנדר על המשוב שלהם.
Materials
| Name | Company | Catalog Number | Comments |
| EEG System | Biosemi | ActiveTwo Base system, 64 channels | |
| Eye-tracker System | Eyelink SR Research | EyeLink 1000 Plus Core Unit, High-speed camera, Host PC/Monitor, | |
| CPU display | Intel | Hard drive 221 GB, Processor Intel Core i7-4790 3.60 Hz, OS Windows 7, 4GB RAM | |
| CPU EEG | Intel | Hard drive 223 GB, Processor Intel Core i7-4790 3.60 Hz, OS Windows 7, 4GB RAM | |
| Monitor | ASUS | ASUS VG248QE 24" LCD monitor | |
| Joytsick | Logitech | Model F310 | |
| Luxmeter | Focket | LCD screen (0-200.000 lux) model Liebe WH LX1330B | |
| Statistics software | GraphPad Prism | GraphPad Prism version 8 for Windows | |
| MATLAB Programming Software | The MathWorks | MATLAB R2014a and R2018b | |
| SVG tool Inkscape | Inkscape Project | vector graphic editor software | |
| Presentation Software | Neurobehavioral Systems | stimulus delivery and experiment control program for neuroscience |
References
- Siegel, M., Donner, T. H., Engel, A. K. Spectral fingerprints of large-scale neuronal interactions. Nature Reviews Neuroscience. 13 (2), 121-134 (2012).
- Fries, P. A mechanism for cognitive dynamics: Neuronal communication through neuronal coherence. Trends in Cognitive Sciences. 9 (10), 474-480 (2005).
- Fries, P. Rhythms for cognition: Communication through coherence. Neuron. 88 (1), 220-235 (2015).
- Thut, G., Miniussi, C., Gross, J. The functional importance of rhythmic activity in the brain. Current Biology. 22 (16), 658-663 (2012).
- Fröhlich, F., McCormick, D. A. Endogenous electric fields may guide neocortical network activity. Neuron. 67 (1), 129-143 (2010).
- Spaak, E., de Lange, F. P., Jensen, O. Local entrainment of alpha oscillations by visual stimuli causes cyclic modulation of perception. Journal of Neuroscience. 34 (10), 3536-3544 (2014).
- Fiebelkorn, I. C., Saalmann, Y. B., Kastner, S. Rhythmic sampling within and between objects despite sustained attention at a cued location. Current Biology. 23 (24), 2553-2558 (2013).
- Landau, A. N., Fries, P. Attention samples stimuli rhythmically. Current Biology. 22 (11), 1000-1004 (2012).
- Song, K., Meng, M., Lin, C., Zhou, K., Luo, H. Behavioral oscillations in attention: Rhythmic α pulses mediated through θ band. Journal of Neuroscience. 34 (14), 4837-4844 (2014).
- Wyart, V., Nobre, A. C., Summerfield, C. Dissociable prior influences of signal probability and relevance on visual contrast sensitivity. Proceedings of the National Academy of Sciences of the United States of America. 109 (9), 3593-3598 (2012).
- Leszczyński, M., Fell, J., Axmacher, N. Rhythmic working memory activation in the human hippocampus. Cell Reports. 13 (6), 1272-1282 (2015).
- Onton, J., Delorme, A., Makeig, S. Frontal midline EEG dynamics during working memory. NeuroImage. 27 (2), 341-356 (2005).
- Cavanagh, J. F., Frank, M. J. Frontal theta as a mechanism for cognitive control. Trends in Cognitive Sciences. 18 (8), 414-421 (2014).
- Siegel, M., Buschman, T. J., Miller, E. K. Cortical information flow during flexible sensorimotor decisions. Science. 348 (6241), 1352-1355 (2015).
- Weisz, N., et al. Prestimulus oscillatory power and connectivity patterns predispose conscious somatosensory perception. Proceedings of the National Academy of Sciences of the United States of America. 111 (4), 417-425 (2014).
- Buzsáki, G., Draguhn, A. Neuronal oscillations in cortical networks. Science. 304 (5679), 1926-1929 (2004).
- Cohen, M. X. A neural microcircuit for cognitive conflict detection and signaling. Trends in Neurosciences. 37 (9), 480-490 (2014).
- Yuste, R. From the neuron doctrine to neural networks. Nature Reviews Neuroscience. 16 (8), 487-497 (2015).
- Helfrich, R. F., Knight, R. T. Oscillatory dynamics of prefrontal cognitive control. Trends in Cognitive Sciences. 20 (12), 916-930 (2016).
- Miller, E. K., Cohen, J. D. An integrative theory of prefrontal cortex function. Annual Review of Neurosciences. 24 (1), 167-202 (2001).
- von Nicolai, C., et al. Corticostriatal coordination through coherent phase-amplitude coupling. Journal of Neuroscience. 34 (17), 5938-5948 (2014).
- Sweeney-Reed, C. M., et al. Thalamic theta phase alignment predicts human memory formation and anterior thalamic cross-frequency coupling. ELife. 4, 07578 (2015).
- Voytek, B., et al. Oscillatory dynamics coordinating human frontal networks in support of goal maintenance. Nature Neuroscience. 18 (9), 1318-1324 (2015).
- Raghavachari, S., et al. Theta oscillations in human cortex during a working-memory task: Evidence for local generators. Journal of Neurophysiology. 95 (3), 1630-1638 (2006).
- Jacobs, J., Hwang, G., Curran, T., Kahana, M. J. EEG oscillations and recognition memory: Theta correlates of memory retrieval and decision making. NeuroImage. 32 (2), 978-987 (2006).
- Itthipuripat, S., Wessel, J. R., Aron, A. R. Frontal theta is a signature of successful working memory manipulation. Experimental Brain Research. 224 (2), 255-262 (2013).
- Cavanagh, J. F., Zambrano-Vazquez, L., Allen, J. J. B. Theta lingua franca: A common mid-frontal substrate for action monitoring processes. Psychophysiology. 49 (2), 220-238 (2012).
- Mas-Herrero, E., Marco-Pallarés, J. Frontal theta oscillatory activity is a common mechanism for the computation of unexpected outcomes and learning rate. Journal of Cognitive Neuroscience. 26 (3), 447-458 (2014).
- Folstein, J. R., Van Petten, C. Influence of cognitive control and mismatch on the N2 component of the ERP: A review. Psychophysiology. 45 (1), 152-170 (2008).
- Cohen, M. X., Donner, T. H. Midfrontal conflict-related theta-band power reflects neural oscillations that predict behavior. Journal of Neurophysiology. 110 (12), 2752-2263 (2013).
- Walsh, M. M., Anderson, J. R. Modulation of the feedback-related negativity by instruction and experience. Proceedings of the National Academy of Sciences of the United States of America. 108 (47), 19048-19053 (2011).
- Luu, P., Tucker, D. M., Makeig, S. Frontal midline theta and the error-related negativity: Neurophysiological mechanisms of action regulation. Clinical Neurophysiology. 115 (8), 1821-1835 (2004).
- Hanslmayr, S., et al. The electrophysiological dynamics of interference during the Stroop task. Journal of Cognitive Neuroscience. 20 (2), 215-225 (2008).
- Cavanagh, J. F., Cohen, M. X., Allen, J. J. B. Prelude to and resolution of an error: EEG phase synchrony reveals cognitive control dynamics during action monitoring. Journal of Neuroscience. 29 (1), 98-105 (2009).
- Cohen, M. X., van Gaal, S., Ridderinkhof, K. R., Lamme, V. A. F. Unconscious errors enhance prefrontal-occipital oscillatory synchrony. Frontiers in Human Neuroscience. 3, 54 (2009).
- Cavanagh, J. F., Frank, M. J., Klein, T. J., Allen, J. J. B. Frontal theta links prediction errors to behavioral adaptation in reinforcement learning. NeuroImage. 49 (4), 3198-3209 (2010).
- Cohen, M. X., Cavanagh, J. F. Single-trial regression elucidates the role of prefrontal theta oscillations in response conflict. Frontiers in Psychology. 2, 30 (2011).
- Cohen, M. X., Van Gaal, S. Dynamic interactions between large-scale brain networks predict behavioral adaptation after perceptual errors. Cerebral Cortex. 23 (5), 1061-1072 (2013).
- Nigbur, R., Cohen, M. X., Ridderinkhof, K. R., Stürmer, B. Theta dynamics reveal domain-specific control over stimulus and response conflict. Journal of Cognitive Neuroscience. 24 (5), 1264-1274 (2012).
- van Driel, J., Swart, J. C., Egner, T., Ridderinkhof, K. R., Cohen, M. X. No) time for control: Frontal theta dynamics reveal the cost of temporally guided conflict anticipation. Cognitive, Affective, and Behavioral Neuroscience. 15 (4), 787-807 (2015).
- van de Vijver, I., Ridderinkhof, K. R., Cohen, M. X. Frontal oscillatory dynamics predict feedback learning and action adjustment. Journal of Cognitive Neuroscience. 23 (12), 4106-4121 (2011).
- Narayanan, N. S., Cavanagh, J. F., Frank, M. J., Laubach, M. Common medial frontal mechanisms of adaptive control in humans and rodents. Nature Neuroscience. 16 (12), 1888-1895 (2013).
- Anguera, J. A., et al. Video game training enhances cognitive control in older adults. Nature. 501 (7465), 97-101 (2013).
- Smit, A. S., Eling, P. A. T. M., Hopman, M. T., Coenen, A. M. L. Mental and physical effort affect vigilance differently. International Journal of Psychophysiology. 57 (3), 211-217 (2005).
- Cohen, M. X., Ranganath, C. Reinforcement learning signals predict future decisions. Journal of Neuroscience. 27 (2), 371-378 (2007).
- Gehring, W. J., Goss, B., Coles, M. G. H., Meyer, D. E., Donchin, E. A neural system for error detection and compensation. Psychological Science. 4 (6), 385-390 (1993).
- Yeung, N., Botvinick, M. M., Cohen, J. D. The neural basis of error detection: Conflict monitoring and the error-related negativity. Psychological Review. 111 (4), 931-959 (2004).
- Debener, S., et al. Trial-by-trial coupling of concurrent electroencephalogram and functional magnetic resonance imaging identifies the dynamics of performance monitoring. Journal of Neuroscience. 25 (50), 11730-11737 (2005).
- Hauser, T. U., et al. The feedback-related negativity (FRN) revisited: New insights into the localization, meaning, and network organization. NeuroImage. 84, 159-168 (2014).
- Wang, C., Ulbert, I., Schomer, D. L., Marinkovic, K., Halgren, E. Responses of human anterior cingulate cortex microdomains to error detection, conflict monitoring, stimulus-response mapping, familiarity, and orienting. Journal of Neuroscience. 25 (3), 604-613 (2005).
- Tsujimoto, S., Genovesio, A. Firing variability of frontal pole neurons during a cued strategy task. Journal of Cognitive Neuroscience. 29 (1), 25-36 (2017).
- Womelsdorf, T., Johnston, K., Vinck, M., Everling, S. Theta-activity in anterior cingulate cortex predicts task rules and their adjustments following errors. Proceedings of the National Academy of Sciences of the United States of America. 107 (11), 5248-5253 (2010).
- Womelsdorf, T., Vinck, M., Stan Leung, L., Everling, S. Selective theta synchronization of choice-relevant information subserves goal-directed behavior. Frontiers in Human Neuroscience. 4, 2010 (2010).
- Domic-Siede, M., Irani, M., Valdés, J., Perrone-Bertolotti, M., Ossandón, T. Theta activity from frontopolar cortex, mid-cingulate cortex and anterior cingulate cortex shows different roles in cognitive planning performance. NeuroImage. 226, 117557 (2021).
- Wilensky, R. . Planning and Understanding: A Computational Approach to Human Reasoning. , (1983).
- Grafman, J., Hendler, J. Planning and the brain. Behavioral and Brain Sciences. 14, 563-564 (1991).
- Hayes-Roth, B., Hayes-Roth, F. A cognitive model of planning. Cognitive Science. 3 (4), 275-310 (1979).
- Tremblay, M., et al. Brain activation with a maze test: An EEG coherence analysis study in healthy subjects. Neuroreport. 5 (18), 2449-2453 (1994).
- Shallice, T. Specific impairments of planning. Philosophical Transactions of the Royal Society B: Biological Sciences. 298 (1089), 199-209 (1982).
- Unterrainer, J. M., Owen, A. M. Planning and problem solving: From neuropsychology to functional neuroimaging. Journal of Physiology. 99 (4-6), 308-317 (2006).
- Domic-Siede, M., et al. La planificación cognitiva en el contexto de la evaluación neuropsicológica e investigación en neurociencia cognitiva: Una revisión sistemática. Terapia Psicológica. 40 (3), 367-395 (2022).
- Miotto, E. C., Morris, R. G. Virtual planning in patients with frontal lobe lesions. Cortex. 34 (5), 639-657 (1998).
- Burgess, P. W., Simons, J. S., Coates, L. M. A., Channon, S., Morris, R., Ward, G. The search for specific planning processes. The Cognitive Psychology of Planning. , 199-227 (2005).
- Oosterman, J. M., Wijers, M., Kessels, R. P. C. Planning or something else? Examining neuropsychological predictors of zoo map performance. Applied Neuropsychology. 20 (2), 103-109 (2013).
- Campbell, Z., et al. Utilizing virtual reality to improve the ecological validity of clinical neuropsychology: An fMRI case study elucidating the neural basis of planning by comparing the Tower of London with a three-dimensional navigation task. Applied Neuropsychology. 16 (4), 295-306 (2009).
- Wilson, B. A., Alderman, N., Burgess, P. W., Emslie, H., Evans, J. J. . Behavioural Assessment of the Dysexecutive Syndrome. , (1996).
- Spector, L., Grafman, J., Boller, F., Grafman, J. Planning, neuropsychology, and artificial intelligence: Cross fertilization. Handbook of Neuropsychology. 9, 377-392 (1994).
- Porteus, S. D. . The Maze Test and Clinical Psychology. , (1959).
- Lis, S., Krieger, S., Wilhelm, J., Gallhofer, B. Feedback about previous action improves executive functioning in schizophrenia: An analysis of maze solving behaviour. Schizophrenia Research. 78 (2-3), 243-250 (2005).
- Zelinsky, G. J. A theory of eye movements during target acquisition. Psychological Review. 115 (4), 787-835 (2008).
- Wurtz, R. H., Goldberg, M. E., Robinson, D. L. Brain mechanisms of visual attention. Scientific American. 246 (6), 124-135 (1982).
- Burr, D., Morrone, M. C. Eye movements: Building a stable world from glance to glance. Current Biology. 15 (20), 839-840 (2005).
- Rolfs, M. Microsaccades: Small steps on a long way. Vision Research. 49 (20), 2415-2441 (2009).
- Ortega, J., Plaska, C. R., Gomes, B. A., Ellmore, T. M. Spontaneous eye blink rate during the working memory delay period predicts task accuracy. Frontiers in Psychology. 13, 788231 (2022).
- Grueschow, M., Kleim, B., Ruff, C. C. Role of the locus coeruleus arousal system in cognitive control. Journal of Neuroendocrinology. 32 (12), 12890 (2020).
- Aston-Jones, G., Shaver, R., Dinan, T. Cortically projecting nucleus basalis neurons in rat are physiologically heterogeneous. Neuroscience Letters. 46 (1), 19-24 (1984).
- Foote, S. L., Bloom, F. E., Aston-Jones, G. Nucleus locus coeruleus: New evidence of anatomical and physiological specificity. Physiological Reviews. 63 (3), 844-914 (1983).
- Siever, L. J., Davis, K. L. Overview: Toward a dysregulation hypothesis of depression. The American Journal of Psychiatry. 142 (9), 1017-1031 (1985).
- Aston-Jones, G., Gold, J. I. How we say no: Norepinephrine, inferior frontal gyrus, and response inhibition. Biological Psychiatry. 65 (7), 548-549 (2009).
- Aston-Jones, G., Cohen, J. D. An integrative theory of locus coeruleus norepinephrine function: adaptive gain and optimal performance. Annual Review of Neurosciences. 28, 403-450 (2005).
- Einhauser, W., Stout, J., Koch, C., Carter, O. Pupil dilation reflects perceptual selection and predicts subsequent stability in perceptual rivalry. Proceedings of the National Academy of Sciences of the United States of America. 105 (5), 1704-1709 (2008).
- Yoshitomi, T., Ito, Y., Inomata, H. Adrenergic excitatory and cholinergic inhibitory innervations in the human iris dilator. Experimental Eye Research. 40 (3), 453-459 (1985).
- Rajkowski, J., Kubiak, P., Aston-Jones, G. Locus coeruleus activity in monkey: Phasic and tonic changes are associated with altered vigilance. Brain Research Bulletin. 35 (5-6), 607-616 (1994).
- Beatty, J. Task-evoked pupillary responses, processing load, and the structure of processing resources. Psychological Bulletin. 91 (2), 276-292 (1982).
- Beatty, J., Kahneman, D. Pupillary changes in two memory tasks. Psychonomic Science. 5, 371-372 (1966).
- Hess, E. H., Polt, J. M. Pupil size in relation to mental activity during simple problem-solving. Science. 143 (3611), 1190-1192 (1964).
- Kuchinke, L., Vo, M. L., Hofmann, M., Jacobs, A. M. Pupillary responses during lexical decisions vary with word frequency but not emotional valence. International Journal of Psychophysiology. 65 (2), 132-140 (2007).
- Barkley, R. A. Adolescents with attention-deficit/hyperactivity disorder: An overview of empirically based treatments. Journal of Psychiatric Practice. 10 (1), 39-56 (2004).
- Gau, S. S. F. e. m. 8. 9., Shang, C. Y. Executive functions as endophenotypes in ADHD: Evidence from the Cambridge Neuropsychological Test Battery (CANTAB). Journal of Child Psychology and Psychiatry and Allied Disciplines. 51 (7), 838-849 (2010).
- Bora, E., Harrison, B. J., Yücel, M., Pantelis, C. Cognitive impairment in euthymic major depressive disorder: A meta-analysis. Psychological Medicine. 43 (10), 2017-2026 (2013).
- Rive, M. M., Koeter, M. W. J., Veltman, D. J., Schene, A. H., Ruhé, H. G. Visuospatial planning in unmedicated major depressive disorder and bipolar disorder: Distinct and common neural correlates. Psychological Medicine. 46 (11), 2313-2328 (2016).
- Holt, D. V., Wolf, J., Funke, J., Weisbrod, M., Kaiser, S. Planning impairments in schizophrenia: Specificity, task independence and functional relevance. Schizophrenia Research. 149 (1-13), 174-179 (2013).
- Lima-Silva, T. B., et al. Functional profile of patients with behavioral variant frontotemporal dementia (bvFTD) compared to patients with Alzheimer's disease and normal controls. Dementia and Neuropsychologia. 7 (1), 96-103 (2013).
- Karnath, H. O., Wallesch, C. W., Zimmermann, P. Mental planning and anticipatory processes with acute and chronic frontal lobe lesions: A comparison of maze performance in routine and non-routine situations. Neuropsychologia. 29 (4), 271-290 (1991).
- Staudigl, T., Hartl, E., Noachtar, S., Doeller, C. F., Jensen, O. Saccades are phase-locked to alpha oscillations in the occipital and medial temporal lobe during successful memory encoding. PLoS Biology. 15 (12), 2003404 (2017).
- Faul, F., Erdfelder, E., Lang, A. G., Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behavior Research Methods. 39 (2), 175-191 (2007).
- Ferrando, L., Bobes, J., Gibert, J., Soto, M. MINI Entrevista Neuropsiquiátrica Internacional (MINI International Neuropsychiatric Interview, MINI). Instrumentos de Detección y Orientación Diagnóstica. , (2000).
- McAssey, M., Dowsett, J., Kirsch, V., Brandt, T., Dieterich, M. Different EEG brain activity in right and left-handers during visually induced self-motion perception. Journal of Neurology. 267 (1), 79-90 (2020).
- Kelly, R., Mizelle, J. C., Wheaton, L. A. Distinctive laterality of neural networks supporting action understanding in left- and right-handed individuals: An EEG coherence study. Neuropsychologia. 75, 20-29 (2015).
- O'Hare, A. J., Atchley, R. A., Young, K. M. Central and divided visual field presentation of emotional images to measure hemispheric differences in motivated attention. Journal of Visualized Experiments. (129), 56257 (2017).
- Beatty, J. Task-evoked pupillary responses, processing load, and the structure of processing resources. Psychological Bulletin. 91 (2), 276-292 (1982).
- Wainstein, G., et al. Pupil size tracks attentional performance in attention-deficit/hyperactivity disorder. Scientific Reports. 7 (1), 8228 (2017).
- Drueke, B., et al. Neural correlates of positive and negative performance feedback in younger and older adults. Behavioral and Brain Functions. 11, 17 (2015).
- Lezak, M. D. The problem of assessing executive functions. International Journal of Psychology. 17 (1-4), 281-297 (1982).
- Oosterman, J. M., Wijers, M., Kessels, R. P. C. Planning or something else? Examining neuropsychological predictors of zoo map performance. Applied Neuropsychology. 20 (2), 103-109 (2013).
- Keil, A., et al. Committee report: Publication guidelines and recommendations for studies using electroencephalography and magnetoencephalography. Psychophysiology. 51 (1), 1-21 (2014).
- Abokyi, S., Owusu-Mensah, J., Osei, K. A. Caffeine intake is associated with pupil dilation and enhanced accommodation. Eye. 31 (4), 615-619 (2017).
- Wilhelm, B., Stuiber, G., Lüdtke, H., Wilhelm, H. The effect of caffeine on spontaneous pupillary oscillations. Ophthalmic & Physiological Optics. 34 (1), 73-81 (2014).
- McGinley, M. J., David, S. V., McCormick, D. A. Cortical membrane potential signature of optimal states for sensory signal detection. Neuron. 87 (1), 179-192 (2015).
- Slotnick, S. D. High density event-related potential data acquisition in cognitive neuroscience. Journal of Visualized Experiments. (38), 1945 (2010).
- Lamm, C., Windischberger, C., Leodolter, U., Moser, E., Bauer, H. Co-registration of EEG and MRI data using matching of spline interpolated and MRI-segmented reconstructions of the scalp surface. Brain Topography. 14 (2), 93-100 (2001).
- Schwartz, D., et al. Registration of MEG/EEG data with 3D MRI: Methodology and precision issues. Brain Topography. 9, 101-116 (1996).
- Laeng, B., Sirois, S., Gredebäck, G. Pupillometry: A window to the preconscious? Perspectives on psychological science. Journal of the Association for Psychological Science. 7 (1), 18-27 (2012).
- Vandierendonck, A. A comparison of methods to combine speed and accuracy measures of performance: A rejoinder on the binning procedure. Behavior Research Methods. 49 (2), 653-673 (2017).
- Vandierendonck, A. Further tests of the utility of integrated speed-accuracy measures in task switching. Journal of Cognition. 1 (1), 8 (2018).
- Levene, H., Olkin, I. Robust tests for equality of variances. In Contributions to Probability and Statistics: Essays in Honor of Harold Hotelling. , 278-292 (1960).
- Zimmerman, D. W. A note on preliminary tests of equality of variances. The British Journal of Mathematical and Statistical Psychology. 57, 173-181 (2004).
- D'Agostino, R. B., Belanger, A., D'Agostino, R. B. A suggestion for using powerful and informative tests of normality). The American Statistician. 44 (4), 316 (1990).
- Shapiro, S. S., Wilk, M. B. An analysis of variance test for normality (complete samples). Biometrika. 52 (3-4), 591-611 (1965).
- Wilcoxon, F. Individual comparisons by ranking methods. Biometrics Bulletin. 1 (6), 80 (1945).
- Fay, M. P., Proschan, M. A. Wilcoxon-Mann-Whitney or t-test? On assumptions for hypothesis tests and multiple interpretations of decision rules. Statistics Surveys. 4, 1-39 (2010).
- Delorme, A., Makeig, S. EEGLAB: An open-source toolbox for analysis of single-trial EEG dynamics including independent component analysis. Journal of Neuroscience Methods. 134 (1), 9-21 (2004).
- Dimigen, O., Sommer, W., Hohlfeld, A., Jacobs, A. M., Kliegl, R. Coregistration of eye movements and EEG in natural reading: Analyses and review. Journal of Experimental Psychology. 140 (4), 552-572 (2011).
- Mognon, A., Jovicich, J., Bruzzone, L., Buiatti, M. ADJUST: An automatic EEG artifact detector based on the joint use of spatial and temporal features. Psychophysiology. 48 (2), 229-240 (2011).
- Bell, A. J., Sejnowski, T. J., Leen, T. K., Tesauro, G., Touretsky, D. S. A non-linear information maximization algorithm that performs blind separation. Advances in Neural Information Processing Systems 7. , (1995).
- Plöchl, M., Ossandón, J. P., König, P. Combining EEG and eye tracking: Identification, characterization, and correction of eye movement artifacts in electroencephalographic data. Frontiers in Human Neuroscience. 6, 278 (2012).
- Pion-Tonachini, L., Kreutz-Delgado, K., Makeig, S. ICLabel: An automated electroencephalographic independent component classifier, dataset, and website. NeuroImage. 198, 181-197 (2019).
- Gartner, M., Grimm, S., Bajbouj, M. Frontal midline theta oscillations during mental arithmetic: Effects of stress. Frontiers in Behavioral Neuroscience. 9, 96 (2015).
- Wang, W., Viswanathan, S., Lee, T., Grafton, S. T. Coupling between theta oscillations and cognitive control network during cross-modal visual and auditory attention: Supramodal vs modality-specific mechanisms. PLoS One. 11 (7), 0158465 (2016).
- Maris, E., Oostenveld, R. Nonparametric statistical testing of EEG- and MEG-data. Journal of Neuroscience Methods. 164 (1), 177-190 (2007).
- Tadel, F., Baillet, S., Mosher, J. C., Pantazis, D., Leahy, R. M. Brainstorm: A user-friendly application for MEG/EEG analysis. Computational Intelligence and Neuroscience. 2011, 879716 (2011).
- Pascual-Marqui, R. D. Standardized low-resolution brain electromagnetic tomography (sLORETA): Technical details. Methods and Findings in Experimental and Clinical Pharmacology. 24, 5-12 (2002).
- Gramfort, A., Papadopoulo, T., Olivi, E., Clerc, M. OpenMEEG: Opensource software for quasistatic bioelectromagnetics). Biomedical Engineering Online. 9, 45 (2010).
- Destrieux, C., Fischl, B., Dale, A., Halgren, E. Automatic parcellation of human cortical gyri and sulci using standard anatomical nomenclature. Neuroimage. 53 (1), 1-15 (2010).
- Orr, J. M., Weissman, D. H. Anterior cingulate cortex makes 2 contributions to minimizing distraction. Cerebral Cortex. 9 (3), 703-711 (2009).
- Christoff, K., Gabrieli, J. D. E. The frontopolar cortex and human cognition: Evidence for a rostrocaudal hierarchical organization within the human prefrontal cortex. Psychobiology. 28 (2), 168-186 (2000).
- Nitschke, K., Köstering, L., Finkel, L., Weiller, C., Kaller, C. P. A meta-analysis on the neural basis of planning: Activation likelihood estimation of functional brain imaging results in the Tower of London task. Human Brain Mapping. 38 (1), 396-413 (2017).
- Barbey, A. K., Koenigs, M., Grafman, J. Dorsolateral prefrontal contributions to human working memory. Cortex. 49 (5), 1195-1205 (2013).
- Le Van Quyen, M., et al. Comparison of Hilbert transform and wavelet methods for the analysis of neuronal synchrony. Journal of Neuroscience Methods. 111 (2), 83-98 (2001).
- Bokil, H., Andrews, P., Kulkarni, J. E., Mehta, S., Mitra, P. P. Chronux: A platform for analyzing neural signals. Journal of Neuroscience Methods. 192 (1), 146-151 (2010).
- Sato, N., Yamaguchi, Y. EEG theta regulates eye saccade generation during human object-place memory encoding. Advances in Cognitive Neurodynamics ICCN 2007. , 429-434 (2008).
- Zaki, J., Ochsner, K. The need for a cognitive neuroscience of naturalistic social cognition. Annals of the New York Academy of Sciences. 1167, 16-30 (2009).
- Shamay-Tsoory, S. G., Mendelsohn, A. Real-life neuroscience: An ecological approach to brain and behavior research. Perspectives on Psychological Science. 14 (5), 841-859 (2019).
- Diamond, A. Executive functions. Annual Review of Psychology. 64, 135-168 (2013).
- Dudai, Y. . Memory from A to Z: Keywords, Concepts and Beyond. , (2002).
- Kingstone, A., Smilek, D., Ristic, J., Friesen, C. K., Eastwood, J. D. Attention, researchers! It is time to take a look at the real world. Current Directions in Psychological Science. 12 (5), 176-180 (2003).
- Morris, R. G., Ward, G., Morris, R., Ward, G. Introduction to the psychology of planning. The Cognitive Psychology of Planning. , (2005).
- Zwosta, K., Ruge, H., Wolfensteller, U. Neural mechanisms of goal-directed behavior: Outcome-based response selection is associated with increased functional coupling of the angular gyrus. Frontiers in Human Neuroscience. 9, 180 (2015).
- Owen, A. M., Doyon, J., Petrides, M., Evans, A. C. Planning and spatial working memory: A positron emission tomography study in humans. European Journal of Neuroscience. 8 (2), 353-364 (1996).
- Ossandon, T., et al. Efficient "pop-out" visual search elicits sustained broadband gamma activity in the dorsal attention network. Journal of Neuroscience. 32 (10), 3414-3421 (2012).
- Lezak, M. D. . Neuropsychological Assessment., 3rd edition. , (1995).
- Lin, H., Saunders, B., Hutcherson, C. A., Inzlicht, M. Midfrontal theta and pupil dilation parametrically track subjective conflict (but also surprise) during intertemporal choice. NeuroImage. 172, 838-852 (2018).
- Yu, S., Mückschel, M., Beste, C. Superior frontal regions reflect the dynamics of task engagement and theta band-related control processes in time-on task effects. Scientific Reports. 12 (1), 846 (2022).
- Nakatani, H., van Leeuwen, C. Antecedent occipital alpha band activity predicts the impact of oculomotor events in perceptual switching. Frontiers in Systems Neuroscience. 7, 19 (2013).
- Velasques, B., et al. Changes in saccadic eye movement (SEM) and quantitative EEG parameter in bipolar patients. Journal of Affective Disorders. 145 (3), 378-385 (2013).
- Bodala, I. P., Li, J., Thakor, N. V., Al-Nashash, H. EEG and eye tracking demonstrate vigilance enhancement with challenge integration. Frontiers in Human Neuroscience. 10, 273 (2016).
- Dimigen, O. Optimizing the ICA-based removal of ocular EEG artifacts from free viewing experiments. NeuroImage. 207, 116117 (2020).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved