
Method Article
Göz İzleyici ve Elektroensefalogram Sistemleri ile Birleştirilmiş Görsel-Uzamsal Bir Planlama Görevi
Bu Makalede
Özet
EEG ve göz izleme sistemlerini birleştiren bilişsel planlama çalışması, insanlarda bilişsel kontrol ve hedefe yönelik davranışa aracılık eden nöral mekanizmaları araştırmak için çok modlu bir yaklaşım sağlar. Burada, planlama performansında beyin salınımlarının ve göz hareketlerinin rolünü araştırmak için bir protokol açıklıyoruz.
Özet
Bir hedefe ulaşmak için organize bir plan formüle etme yeteneği ile karakterize edilen planlama süreci, insanın hedefe yönelik davranışı için gereklidir. Bazı nöropsikiyatrik bozukluklarda planlama tehlikeye girdiğinden, planlamayı incelemek için uygun klinik ve deneysel testlerin uygulanması kritik öneme sahiptir. Çeşitli bilişsel alanların katıldığı planlamanın konuşlandırılmasının doğası gereği, planlamanın değerlendirilmesi ve nörogörüntüleme yöntemleriyle birleştirilmiş davranışsal paradigmaların tasarımı, bilişsel sinirbilimdeki güncel zorluklardır. 27 sağlıklı yetişkin katılımcıda bir elektroensefalogram (EEG) sistemi ve göz hareketi kayıtları ile birlikte bir planlama görevi değerlendirildi. Planlama iki aşamaya ayrılabilir: bir dizi adımın dahili olarak temsil edildiği bir zihinsel planlama aşaması ve önceden planlanmış bir hedefe ulaşmak için motor eylemin kullanıldığı bir yürütme aşaması. Protokolümüz bir planlama görevi ve bir kontrol görevi içeriyordu. Planlama görevi, her biri bir hayvanat bahçesi haritasını temsil eden 36 labirent denemesini çözmeyi içeriyordu. Görevin dört dönemi vardı: i) deneklere bir dizi kurala göre dört hayvanın yerlerini ziyaret etmek için bir yol planlamaları talimatının verildiği planlama; ii) deneklerin planlanan yolu çalışma belleklerinde tutmaları gereken bakım; iii) deneklerin, göz takip sistemi tarafından belirtildiği gibi önceden planlanan yolu izlemek için göz hareketlerini kullandığı uygulama; ve iv) deneklerin ziyaret edilen hayvanların sırasını bildirdiği yanıt. Kontrol görevi benzer bir yapıya sahipti, ancak bilişsel planlama bileşeni, görev hedefi değiştirilerek kaldırıldı. EEG'nin uzamsal ve zamansal örüntüleri, planlamanın zaman içinde frontal-orta hat teta aktivitesinde (FMθ) kademeli ve kalıcı bir artışa neden olduğunu ortaya koydu. Bu aktivitenin kaynağı, kaynak analizleri ile prefrontal korteks içinde tespit edildi. Sonuçlarımız, EEG ve göz izleyici sistemlerini birleştiren deneysel paradigmanın bilişsel planlamayı değerlendirmek için en uygun olduğunu göstermiştir.
Giriş
Son 10 yılda, salınımlı nöral dinamiklerin hem biliş hem de davranış üzerindeki rolünü incelemek için kapsamlı araştırmalar yapılmıştır. Bu çalışmalar, özelleşmiş ve yaygın kortikal bölgeler arasındaki frekansa özgü etkileşimlerin biliş ve bilişsel kontrolde çok önemli bir rol oynadığını ortaya koymuştur 1,2,3. Bu yaklaşım, büyük ölçekli kortikal dinamikleri koordine etmeye yardımcı olan ve bilişsel işlemeyi ve hedefe yönelik davranışı destekleyen beyin aktivitesinin ritmik doğasını vurgular 4,5. Beyindeki ritmik salınımların, algı6, attentio 7,8,9, karar verme 10, hafıza yeniden aktivasyonu11, çalışma belleği12 ve bilişsel kontrol13 dahil olmak üzere çeşitli bilişsel süreçlerde yer aldığını gösteren önemli kanıtlar vardır. Bilişsel işlemeiçin bir çerçeve sağlayan geçici büyük ölçekli frekansa özgü ağlarla, hedefe yönelik davranışı yönlendirmek için farklı salınım mekanizmaları önerilmiştir 1,14,15. Örneğin, son bulgular, beyindeki belirli frekans bantlarının, ani yükselme aktivitesini düzenleyen bir geri bildirim mekanizmasını yansıtabileceğini, davranış üretmek için kortikal uyarılabilirliği ve ani yükselme zamanlamasını koordine etmek için zamansal bir referans çerçevesi sağlayabileceğinigöstermektedir 16,17,18. Helfrich ve Knight19 tarafından bir inceleme sağlanmıştır.
Bu kanıtlar, prefrontal korteksin (PFC) planlamayı, görev bağlamlarını ve ilgili davranışsal olarak ilgili kuralları nasıl kodladığı hakkında soruları gündeme getiriyor. PFC'nin, ürettiği nöral aktivitenin salınım kalıpları aracılığıyla bilişsel kontrolü ve hedefe yönelik davranışı desteklediği, uzak beyin bölgelerindeki nöral aktiviteyi seçici olarak önyargılı hale getirdiği ve büyük ölçekli sinir ağlarındaki bilgi akışını kontrol ettiği uzun zamandır düşünülmektedir20. Ek olarak, yerel senkronizasyon sergileyen bölgelerin bölgeler arası aktiviteye katılma olasılığının daha yüksek olduğu önerilmiştir 21,22,23. Özellikle, kafa derisi elektroensefalogramı (EEG) ile ölçülen kortikal teta bandı (4-8 Hz) salınımları, geniş ağlar boyunca yukarıdan aşağıya kontrolü iletmek için potansiyel bir mekanizma olarak önerilmiştir13. Spesifik olarak, insanlarda teta bandı aktivitesi, bellek kodlama ve geri alma, çalışma belleği tutma, yenilik algılama, karar verme ve yukarıdan aşağıya kontrol gibi üst düzey bilişsel süreçleri yansıtır 12,24,25,26.
Bununla ilgili olarak, Cavanagh ve Frank13, kontrol süreçleri için iki ardışık mekanizma önerdi: kontrol ihtiyacının tanınması ve kontrolün somutlaştırılması. Kontrol ihtiyacının tanınması, medial prefrontal korteksten (mPFC) kaynaklanan frontal orta hat teta (FMθ) aktivitesi ile gösterilebilir ve bu, yeni bilgiler 27,28,29, çelişkili uyaran-tepki gereksinimleri 30, hata geri bildirimi 31 gibi çeşitli durumlara yanıt olarak mPFC ile ilgili kontrol süreçlerini yansıtan olayla ilgili potansiyel (ERP) bileşenleri açısından tanımlanmıştır, ve hata tespiti32. Yenilik, çatışma, ceza veya hata varlığında artan bilişsel kontrol ihtiyacını yansıtan bu ERP bileşenleri, frontal orta hat elektrotlarındakaydedilen teta bandında ortak bir spektral imza sergiler 26,27,33,34,35,36,37,38,39,40, 41,42,43,44.
FMθ aktivitesinin EEG yanıtları, teta frekans bandı26'da bir faz sıfırlama ve güç geliştirme modeli gösterir. EEG yönteminin uzamsal çözünürlüğü açısından sınırlamalarına rağmen, FMθ aktivitesinin orta singulat korteks (MCC) tarafından üretildiğini göstermek için çeşitli kanıt kaynakları toplanmıştır13. Bu teta dinamiklerinin, mPFC'nin nöronal süreçlerini düzenleyen zamansal çerçeveler olarak hizmet ettiğine inanılmaktadır ve bunlar daha sonra yüksek kontrol gerektiren olaylara yanıt olarak arttırılmaktadır26. Bu, kaynak analizi 31,33,45,46,47, eşzamanlı EEG ve fonksiyonel manyetik rezonans görüntüleme (fMRI) kayıtları 48,49 ve insanlarda 50 ve maymunlarda51,52,53 invaziv EEG kayıtları yoluyla kurulmuştur.
Bu gözlemlere dayanarak, frontal orta hat tetanının, planlama sırasında olduğu gibi, eylemler ve sonuçlarla ilgili kesinlik eksikliğinin olduğu farklı durumlarda uyarlanabilir kontrolü yürütmek için evrensel bir mekanizma, ortak bir dil olarak hizmet ettiği düşünülmektedir. Bu protokolde önerdiğimiz davranışsal paradigma, bilişsel planlamayı ve onun zamansal ve sinirsel özelliklerini incelemek için kullanılmıştır. Diğer senaryolarda bilişsel kontrol için çeşitli mekanizmalar bildirilmiş olsa da, mevcut protokol planlamanın ve bununla ilişkili nöral ve zamansal özelliklerin yakın zamanda tanımlanmasına izin vermiştir54. Planlamanın bilişsel süreci iki farklı aşamadan oluşur: bir dizi planın içsel bir temsilinin geliştirildiği zihinsel planlama aşaması55 ve daha önce planlanan hedefe ulaşmak için bir dizi motor eylemin yürütüldüğü planlama yürütme aşaması56. Planlamanın, çalışma belleği, dikkat kontrolü ve tepki engelleme dahil olmak üzere yürütücü işlevlerin çeşitli bileşenlerinin entegrasyonunu gerektirdiği bilinmektedir, bu da bu süreçlerin deneysel manipülasyonunu ve izole ölçümünü zorlaştırmaktadır57,58.
Bilişsel planlama üzerine yapılan nörogörüntüleme çalışmaları, Londra Kulesi 59,60,61; Bununla birlikte, kafa karıştırıcı faktörleri kontrol etmek için, bilişsel planlamayı incelemek için kullanılan görevler sınırlı ve yapay hale gelebilir, bu da daha az öngörücü ve ekolojik geçerliliğe yol açabilir 62,63,64,65. Nöropsikoloji alanındaki bu sorunun üstesinden gelmek için, gerçek dünya planlama durumları ekolojik görevler olarak önerilmiştir62,63. Diseksekütif Sendromun Davranışsal Değerlendirmesindeki Hayvanat Bahçesi Haritası Görevi alt testi, planlama ve organizasyon becerilerini daha doğal ve ilgili bir şekilde ölçer64,66. Bu test, bir hayvanat bahçesi haritasındaki 12 konumdan 6'sını ziyaret etmek için bir rota planlamayı içeren bir kalem ve kağıt testidir. Konumlar, fil evi, aslan kafesi, dinlenme alanı, kafe vb. gibi normal bir hayvanat bahçesinde bulunabilecek yaygın yerlerdir. Farklı planlama düzeylerini değerlendiren iki koşul vardır: i) deneklere, seçtikleri sıraya göre, ancak bir dizi kurala göre altı yeri ziyaret etmek için bir rota planlamaları talimatının verildiği formülasyon koşulu; ve ii) deneklere belirli bir sırayla ve bir dizi kurala uyarak altı yeri ziyaret etmeleri talimatının verildiği infaz koşulu. Bu iki koşul, kötü yapılandırılmış (formülasyon) ve iyi yapılandırılmış (uygulama) problemlerde planlama becerileri hakkında bilgi sağlar67. Birincisi, açık bir durumda daha zorlu bir bilişsel görev olarak sunulur, çünkü deneklerin hedefe ulaşmak için mantıklı bir strateji geliştirmesini gerektirir. Bir yolu izlemeden önce, bir operatör dizisi tasarlanmalıdır; aksi takdirde hataların oluşması muhtemeldir. Öte yandan, yürütme koşulu daha düşük bir bilişsel talep gerektirir, çünkü belirli bir empoze edilen stratejiyi takip etmeyi içeren bir görevi çözmek, öznenin yalnızca hedefe ulaşmak için formüle edilmiş planın uygulanmasını izlemesini gerektirir66. Öte yandan, Porteus Labirenti psikoloji alanında, özellikle bilişsel psikoloji ve nöropsikoloji alanlarında iyi bilinen bir görevdir ve problem çözme ve planlama gibi bilişin çeşitli yönlerini değerlendirmek için bir araç olarak yaygın olarak kullanılmaktadır68,69. Porteus Labirenti görevi, basit bir görsel uyaran analiziyle başlayan ve giderek zorlaşan bir kalem ve kağıt görevidir. Denek, kesişen yollardan ve çıkmaz sokaklardan kaçınmak ve mümkün olduğunca çabuk hareket etmek gibi kurallara uyarken, bir başlangıç noktasından bir çıkışa kadar (birkaç seçenek arasından) doğru yolu bulmalı ve izlemelidir68. Yol çizilirken her çatal belirdiğinde, denekler hedefe ulaşmak ve verilen kuralları çiğnemekten kaçınmak için kararlar alırlar69.
Yaygın olarak kullanılan ve ekolojik görevlerin sınırlamalarını ve güçlü yönlerini göz önünde bulundurarak, davranışsal paradigmamızı temel olarak Hayvanat Bahçesi Haritası Görev66 ve Porteus Labirent Görevi68'e dayanarak tasarladık. Davranışsal paradigma, günlük bir yaşam senaryosunda bilişsel planlama sürecini kapsayan dört farklı aşamadan oluşur. Bu aşamalar şu şekildedir: Aşama 1, katılımcıların bir harita üzerinde çeşitli yerleri ziyaret etmek için bir rota oluşturmakla görevlendirildiği, belirlenen kurallara bağlılığın sağlandığı planlama; Aşama 2, katılımcıların planlanan rotayı çalışma hafızalarında tutmaları gereken bakım; Aşama 3, katılımcıların önceden planladıkları rotayı çizerek ve doğruluğunu yakından izleyerek yürüttükleri yürütme; ve Katılımcıların planladıkları rota54'e göre ziyaret edilen hayvanların sırasını bildirdiği Aşama 4, yanıt. Paradigmamız, planlamanın çeşitli bileşenlerini (çalışma belleği, yönetici dikkati ve görsel-uzamsal beceriler gibi) daha gerçekçi bir şekilde yansıtan farklı aşamaları kullanarak planlama yeteneğinin farklı parametrelerinin ölçülmesini sağlar, çünkü rotaların haritalandırılması günlük yaşamda yaygın bir durumdur. Ek olarak, karıştırıcı faktörleri kontrol etmek için, paradigma, planlamada da yer alan yürütücü bilişsel bileşenleri meşgul eden ancak planlama süreci bileşenini dışlayan bir planlama görev yapısı ve eşdeğer uyaranlara sahip bir kontrol görevi içerir. Bu, hem elektrofizyolojik belirteçlerin hem de davranışsal parametrelerin karşılaştırılması için planlama süreci bileşeninin ayrılmasına izin verir54.
Ayrıca, göz izleme, algı, dikkat ve bilişsel işlevlerin altında yatan bilişsel süreçler ve sinirsel mekanizmalar hakkında değerli bilgiler sağlayabilen, göz hareketlerini ölçmek ve analiz etmek için invaziv olmayan bir yöntem sağlayarak bilişsel sinirbilim çalışmalarına önemli katkılarda bulunmuştur. Bir göz izleme sistemi ile farklı göz hareketlerini ölçmek, planlamada yer alan bilişsel süreçler ve sinirsel mekanizmalar hakkında değerli bilgiler sağlayabilir. Örneğin, aşağıdaki hususlar ölçülebilir: görsel bilginin edinildiği sabit bakış dönemleri olan fiksasyonlar70; Bakışları bir yerden başka bir yere kaydırmak için kullanılan hızlı göz hareketleri olan sakkadlar71; gözlerin hareket eden bir nesneyi sorunsuz bir şekilde takip etmesini sağlayan bir tür göz hareketi olan smooth pursuit72; fiksasyonlar sırasında bile ortaya çıkan küçük, hızlı göz hareketleri olan mikrosakkadlar73; ve gözleri yağlı tutmaya ve yabancı cisimlerden korumaya yardımcı olan bir refleks eylemi olan göz kırpma74. Bu göz hareketleri, planlama ve bilişsel kontrol için önemli bileşenler olan görsel arama, dikkat tahsisi70, görsel izleme72, algı73 ve çalışma belleği74 ile ilgili bilişsel süreçler hakkında içgörü sağlayabilir.
Öte yandan, locus coeruleus-norepinefrin (LC-NE) sistemi üzerine yapılan son çalışmalar, bilişsel kontroldeki ilgili rolünü göstermiştir75. Locus coeruleus (LC), serebral korteks, hipokampus, talamus, orta beyin, beyin sapı, beyincik ve omurilik gibi çeşitli beyin bölgelerine projeksiyon yapar 76,77,61. Özellikle yoğun LC-NE innervasyonları, bilişsel kontrol75 ile ilişkili PFC beyin alanlarını alır. Ayrıca, bazı çalışmalar LC sisteminin kronik hiperaktivitesinin dürtüsellik ve uykusuzluk gibi manik-depresif bozukluk semptomlarına katkıda bulunabileceğini göstermektedir. Buna karşılık, LC fonksiyonundaki kronik bir azalma, depresyondan muzdarip hastalar arasında yaygın bir özellik olan duygusal ifadenin azalmasıyla ilişkilendirilmiştir78. Locus coeruleus'un uyaranlara aşırı aktif tepkisi, stres veya anksiyete bozukluğu olan bireylerde aşırı tepkiye yol açabilir79. Bu nedenle, LC-NE sistemindeki değişiklikler bilişsel ve/veya duygusal düzensizlik semptomlarına katkıda bulunabilir. Non-invaziv teknikler, lokus coeruleus aktivitesini incelemek için kullanılabilir, bunlardan biri çoğunlukla locus coeruleus'tan salınan noradrenalin tarafından kontrol edilen pupil çapı değişiklikleridir. Noradrenalin, alfa-adrenoseptörleri uyararak iris dilatör kası ve siliyer gangliona sinyaller gönderen ve postsinaptik alfa-2 adrenoseptörlerinin(66,80,81,82) aktivasyonu yoluyla iris genişlemesini kontrol eden Edinger-Westphal çekirdeği üzerinde etkilidir. Maymunlardan alınan doğrudan LC nöronal kayıtları, LC-NE aktivitesi, göz bebeği çapı ve bilişsel performans arasındaki ilişkiyi doğrulamıştır83. Çeşitli bilişsel görevlerde gelişmiş işleme taleplerine yanıt olarak öğrenci genişlemesi tekrar tekrar gözlenmiştir 71,84,85,86,87.
Göz izleme ve göz bebeği kayıtları ile birleştirilen bilişsel kontrolün elektrofizyolojik belirteçleri, bilişsel kontrol ve planlamanın beyinde nasıl uygulandığına dair önemli soruları çözebilir. EEG ve göz takip sistemlerini birleştiren protokolümüzü kullanmanın önemi iki yönlüdür. Bir yandan, bilişsel kontrol, dağıtılmış beyin aktivitesinin kesin zamansal ilişkilere katılımını gerektiriyor gibi görünmektedir, bu da beyin ağı işlevini incelemek için ideal adayları oluşturmaktadır. Öte yandan, dikkat eksikliği/hiperaktivite bozukluğu 88,89, majör depresif bozukluk90,91, bipolar bozukluk91, şizofreni92, frontotemporal demans93 gibi çeşitli bilişsel ve nöropsikiyatrik bozukluklarda olduğu gibi, bu kapasitelerin herhangi birindeki anormalliklerin normal davranış üzerinde ciddi bir etkisi vardır ve ayrıca frontal lezyonlara bağlı bozukluklar94. Ek olarak, mevcut protokol, göz izleme ve elektroensefalografi kullanarak LC-NE aktivitesini ve salınımlarını karşılaştırmak için pupillometrinin bir parametre olarak kullanılmasına izin verir. Bu, yalnızca insanlarda LC-NE, pupillometri ve nöral belirteçler arasındaki teorik ilişki için kanıt sağlamakla kalmaz, aynı zamanda bilişsel planlama sırasında LC-NE sistemi ile ilgili özelliklerin gelişimsel yörüngesinin izlenmesine de izin verebilir. Bununla birlikte, modelimizde, planlama sırasında potansiyel olarak belirli salınım değişikliklerine neden olabilecek belirli bir sakkad modeli olup olmadığını test etmeye odaklandık95. Ek olarak, davranışsal paradigmamızın yürütme aşamasında bir planın davranışsal uygulamasını incelemenin önemli bir parçası olarak bir göz izleyici sistemi kullandık.
Özetlemek gerekirse, bu protokol hem daha ileri temel araştırmalar hem de nihai klinik ve terapötik uygulamalar için bir platform olarak hizmet edebilecek test edilebilir beyin ağı dinamikleri modelleri üretebilir.
Protokol
Bu protokolde yer alan tüm prosedürler Pontificia Universidad Católica de Chile Tıp Fakültesi biyoetik kurulu tarafından onaylanmış ve tüm katılımcılar çalışma başlamadan önce bilgilendirilmiş onam formu imzalamışlardır (araştırma proje no: 16-251).
1. Katılımcı alımı
- Normal veya düzeltilmiş normal görüşe sahip sağ elini kullanan sağlıklı yetişkinleri (erkekler ve kadınlar) işe alın ve onları dahil etme / dışlama kriterlerine göre tarayın.
NOT: Bu çalışmaya yaşları 19 ile 38 arasında değişen ve akıcı konuşabilen 27 sağlıklı birey dahil edilmiştir. Örneklem büyüklüğü, istenen istatistiksel güç düzeyine bağlı olarak değişebilir ve yaş aralığı, ele alınacak spesifik araştırma sorusuna bağlı olarak değişebilir. Protokolümüzde örneklem büyüklüğünü, Faul ve ark.96'da tarif edildiği gibi istatistiksel Wilcoxon işaretli sıra testi, 0.7 etki büyüklüğü, 0.05 alfa seviyesi ve 0.95 kuvveti dikkate alarak hesapladık. Katılımcıları dahil etme/dışlama kriterleri açısından değerlendirmek için eğitimli bir psikolog tarafından uygulanan MINI-Uluslararası Nöropsikiyatrik Görüşme97'yi kullandık. EEG sinyalindeki değişkenliği azaltmak için sadece sağ elini kullanan denekleri işe alın, çünkü solak bireyler EEG aktivitesinin farklı bir topografik dağılımı sunabilir 98,99,100.
2. Uyaran hazırlama
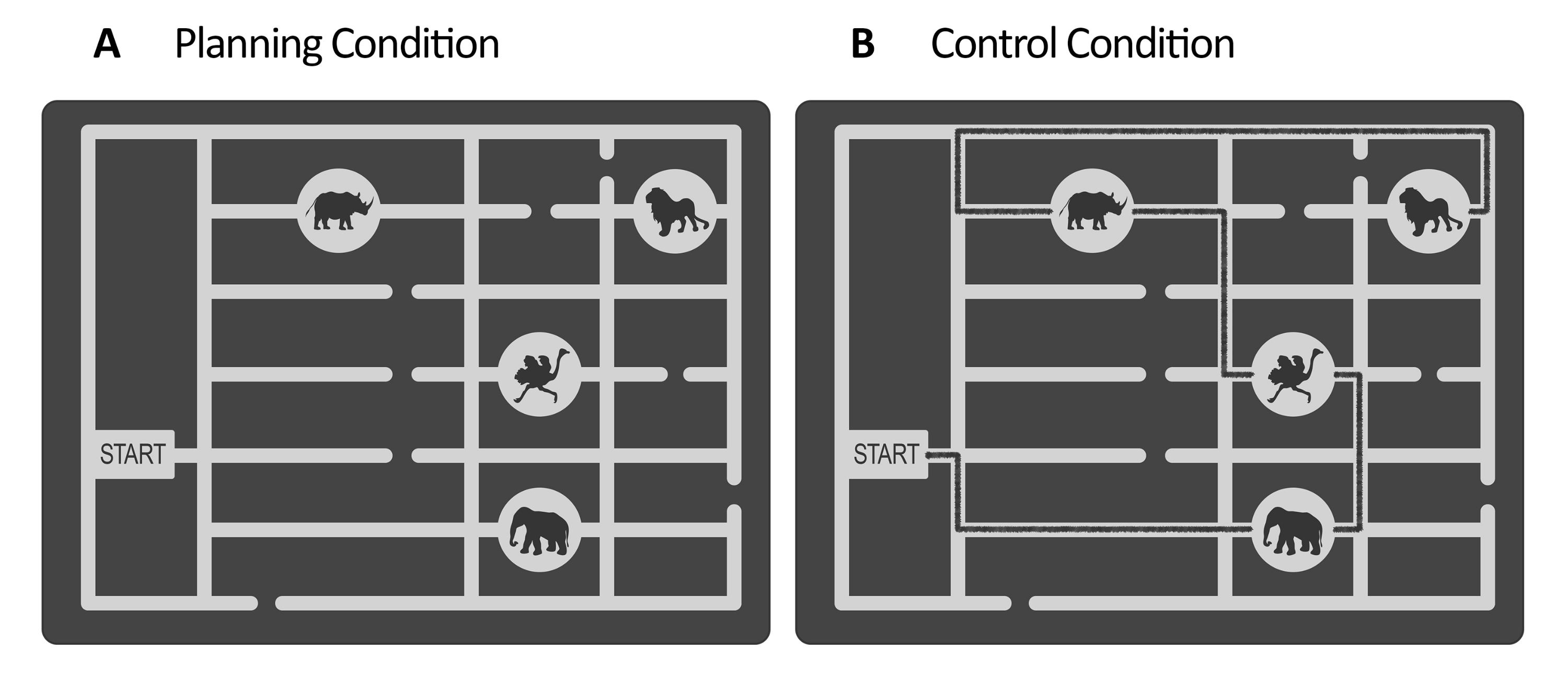
- Planlama görevi için, bir vektör grafik düzenleme yazılımı kullanarak bir dizi uyaran oluşturun (bkz. Malzeme Tablosu). Her uyaran için, bir hayvanat bahçesi haritasını temsil eden gri tonlamalı bir labirent tasarlayın. Labirentin içinde bir geçit ve hayvan konumlarına giden birkaç yol açın (örneğin, bu çalışmada dört hayvan konumu vardı, bkz. Şekil 1).
NOT: Bu çalışmada, her bir uyaranın başlangıç kapısı olan bir hayvanat bahçesi haritası, labirentte bulunan dört hayvan görüntüsü ve birkaç yoldan oluştuğu 36 labirent oluşturduk. Labirent üzerindeki yollar hayvanların bulunduğu yerlere gidebilir veya çıkmayabilir. Azaltılmış kontrastlı gri tonlamalı uyaranların kullanılması, renkli görmeden sorumlu olan retina konilerinin uyarılmasını azalttığı için pupillometri için sıklıkla tercih edilir. Uyaran vurgusundaki bu kayma, uyarılma veya dikkat durumundaki değişiklikleri yansıttığı düşünülen göz bebeğinin boyutundaki değişikliklerin daha hassas bir şekilde ölçülmesine izin verir. Ek olarak, gri tonlamalı uyaranların kullanılması, bireyler arasındaki renk görme farklılıklarından dolayı ölçümdeki değişkenliği azaltır101. - Nihai görevde farklı karmaşıklık seviyeleri elde etmek için, uyaranları belirlenen hedef ve kurallara uygun olarak geçerli çözümlerin sayısına göre bölün (özellikle amaç, hayvan yerlerini ziyaret etmek için bir yol planlamaktır). Geçerli çözümlerin sayısı, kurallara göre planlanabilecek yolların sayısını ifade eder ( Şekil 2 ve adım 5.12.1.'deki kurallara bakın). Beşten fazla olası çözümü olan uyaranları "kolay" ve beş veya daha az olası çözümü olanları "zor" olarak sınıflandırın. Ardından, her kategori için eşit sayıda deneme sürümü oluşturun.
NOT: Alternatif olarak, Domic-Siede ve ark.54 için oluşturulan uyaranları yazarlardan isteyin, çünkü bu uyaranlar bu talimatlara göre oluşturulmuştur. Tüm malzemelerin istek üzerine temin edildiğini, ancak teknik özelliklerin burada ayrıntılı olarak açıklandığını göz önünde bulundurun. Bu çalışmada 18 kolay deneme ve 18 zor deneme oluşturuldu. Davranışsal ve elektrofizyolojik düzeylerdeki zorluk düzeylerindeki farklılıkları değerlendirmek önemlidir, çünkü bilişsel talep/bilişsel çaba/zorluk veya bilişsel planlamanın içsel yönlerini ölçüp ölçmediğinizi belirlemeye yardımcı olacaktır (temsili sonuçlara ve tartışma bölümlerine bakınız). - Kontrol görevi için, planlama göreviyle aynı yapıyı kullanın (değerlendirme periyodu, bakım, yürütme, yanıt, bkz. Şekil 2) ve planlama görevi için oluşturulan aynı uyaranları kullanın, ancak kapıdan başlayarak son konuma kadar hayvan konumlarının sırasını ziyaret etmek için işaretli bir yolu temsil eden çizilmiş bir çizgi ekleyin (bkz. Şekil 1B). İşaretli yolu, labirentin ana yollarından biraz daha koyu bir renk yapın, düşük kontrast bir lüks ölçer kullanılarak değerlendirilir (bkz. adım 2.4).
NOT: Bunun arkasındaki fikir, her iki koşulun (planlama ve kontrol görevleri) psikofiziksel özelliklerini mümkün olduğunca benzer tutmaktır. Labirentlerin izlenen yolları kurallara uyabilir veya uymayabilir (katılımcılara verilen talimatlarla ilgili daha fazla ayrıntı için protokolün 5. bölümüne bakın). Bu çalışmada, uyaranların yarısı kurallara uygun olarak doğru bir ziyaret sırasına sahipken, diğer yarısı hatalar sundu (aynı yolu iki kez kullanmak veya çıkmaz sokakları geçmek gibi, bkz. adım 5.12.1 ve adım 5.12.3 ve Şekil 2). - Deneklerin kullanacağı çene dayanağına yerleştirilmiş bir lüks ölçer kullanarak (bkz. adım 4.5 ve adım 4.6) ve ekrandan aynı mesafede uyaranların aydınlatmasını değerlendirin. Her koşulun her uyaranı bir lux değeri üretir. Daha fazla analiz için her değeri manuel olarak kaydedin.
NOT: Koşullar arasında aydınlatma açısından herhangi bir fark beklenmez (bkz. adım 4.6). Aksi takdirde, uyaranların kontrastını kontrol edin. Bu, göz bebeği çapının102 ölçülmesi durumunda geçerlidir. - Bir vektör grafik editörü (bkz. Malzeme Tablosu) kullanarak doğru geri bildirimi temsil eden bir uyaran (doğru olduğunda beğenme) ve yanlış geri bildirimi temsil eden başka bir uyaran (yanlış olduğunda beğenme) oluşturun (bkz. Malzeme Tablosu) (Şekil 2).

Şekil 1: Deney ve kontrol görevinin uyaranları. (A) planlama ve (B) kontrol görevi uyaranlarının açıklayıcı örnekleri gösterilmiştir. Uyaranlar, bir kapı, farklı yerlerde dört hayvan konumu ve birkaç yoldan oluşan bir hayvanat bahçesi haritasını temsil eder. Her iki durum için de uyaranlar benzerdi; tek fark, kontrol görevi için, (B) uyaranların zaten var olan bir yolu gösteren işaretli bir çizgiye sahip olmasıydı (burada açıklama amaçlı siyah çizgi). Gerçek kontrol uyaranlarındaki bu çizgi biraz daha koyuydu ve düşük kontrast aydınlatma tarafından kontrol ediliyordu (bkz. adım 2.4). Bu rakam Domic-Siede et al.54'ten değiştirilmiştir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

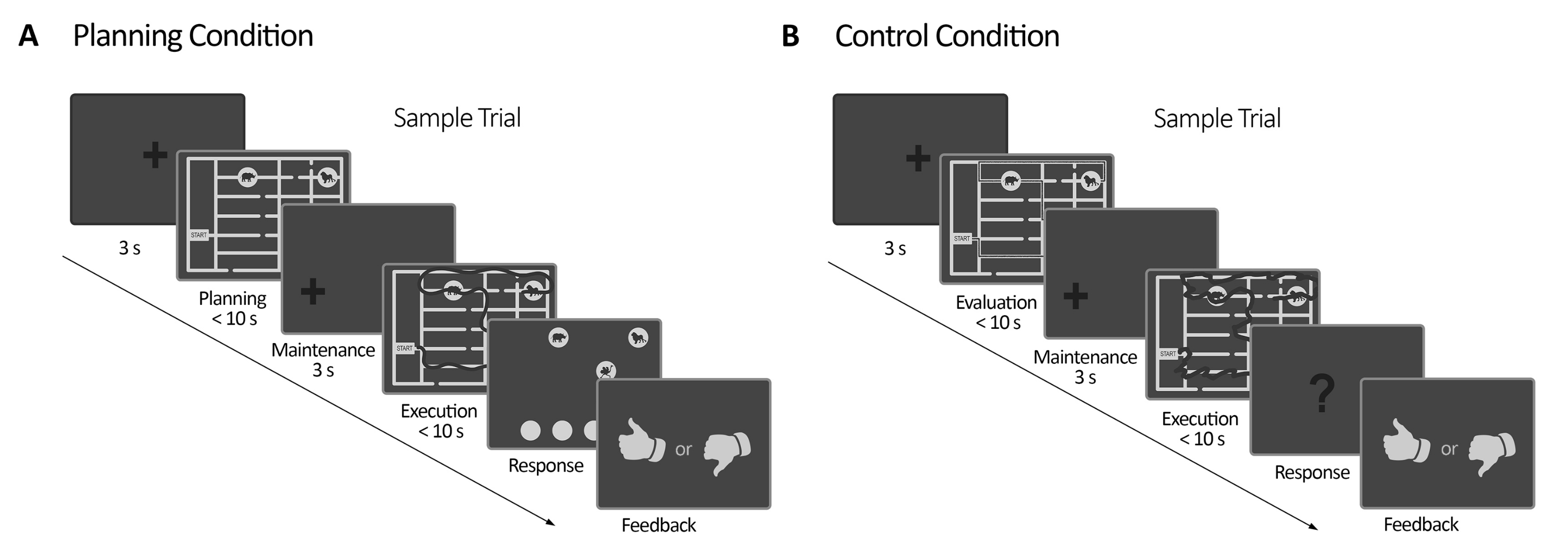
Şekil 2: Deney tasarımı. (A) Görev denemesinin planlanması. Bu durumdaki denemeler 3 sn fiksasyon çaprazı ile başladı. Daha sonra, katılımcılara bir dizi kuralı (en fazla 10 s) izleyerek dört hayvanın bulunduğu yeri ziyaret etmek için bir yol planlamaları talimatı verildi. Daha sonra, kaydırılmış bir sabitleme haçı belirdi (3 s), ardından labirentin tekrar ortaya çıkması izledi. Bu dönemde (uygulama), deneklerin, bakış hareketlerini gerçek zamanlı olarak (koyu çizgi) (en fazla 10 s) tanımlayan çevrimiçi görsel geri bildirim (göz izleyici sistemi tarafından verilen) ile bakışlarını kullanarak önceki planlama döneminde planlanan izi gerçekleştirmeleri gerekiyordu. Daha sonra, yanıt döneminde, denekler, ziyaret edilen hayvanları emrederek infaz sırasında yapılan sırayı raporlamak zorunda kaldılar. Gelen yanıtlara göre geri dönüş sağlandı. (B) Kontrol görevi denemesi. Bu durumdaki denemeler 3 sn fiksasyon çaprazı ile başladı. Daha sonra, katılımcılara izlenen bir yolun (koyu çizgi) kurallara uyup uymadığını değerlendirmeleri talimatı verildi. Daha sonra, kaydırılmış bir sabitleme haçı belirdi (3 s), ardından labirentin tekrar ortaya çıkması izledi. Bu dönemde, denekler, planlama yürütme döneminde olduğu gibi (en fazla 10 saniye) çevrimiçi görsel geri bildirimle önceden izlenen yolu yeniden çizmek zorunda kaldılar. Daha sonra, yanıt süresinde, deneklerin izlenen dizinin daha önce belirtilen kurallara uyup uymadığını (evet veya hayır) yanıtlamaları gerekiyordu Yanıtlarına göre geri bildirim verildi. Bu rakam Domic-Siede et al.54'ten değiştirilmiştir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
3. Planlama ve kontrol görevi programlama
- Bir uyaran sunumu/davranışsal deneyler yazılımı kullanarak Hayvanat Bahçesi Haritası Görev66 ve Porteus Labirenti68'edayalı bir planlama görevi paradigmasını kodlayan bir komut dosyası yazın (bkz. Malzeme Tablosu ve Ek Dosya).
- Görevi, bölüm 2 ve bölüm 4'te açıklanana benzer bir yapıya sahip iki koşulu (bir planlama koşulu ve bir kontrol koşulu) göz önünde bulundurarak kodlayın (bkz. Şekil 2 ve Ek Dosya).
NOT: Görev taleplerini çözme sürecinde yer alan karıştırıcı faktörleri ve algısal bileşenleri kontrol etmek için her iki koşulda da aynı yapıyı kullanmak önemlidir (Şekil 2). Aynı yapıyı kullanmak, bilişsel planlamada yer alan belirli bilişsel sürecin değerlendirilmesini geliştirir. - Görüntü bilgisayarı, EEG bilgisayarı ve ana bilgisayar (göz izleyici bilgisayar) arasındaki iletişimi, ekran bilgisayarından transistör-transistör mantığı (TTL) darbeleri göndererek ethernet ve paralel port iletişimi aracılığıyla senkronize edin (bkz. Şekil 3).
- Planlama ve kontrol görevlerinin başında ve tamamlanan her beş denemeden sonra göz hareketlerini göz izleyici sistemi ile kalibre etmek için bir kod yazın, çünkü ekrandaki bakış konumu yürütme süresi için çok önemlidir (protokolün 3.2.3 ve 3.3.3 adımlarına, tartışma bölümüne ve Ek Dosyadaki koda bakın).
NOT: Bilgisayar iletişiminde gecikmeler olabilir. İki farklı bilgisayarda TTL darbeleri arasındaki gecikmeyi ölçmenin birkaç yöntemi vardır, ancak yaygın bir yaklaşım, dijital osiloskop veya mantık analizörü gibi bir donanım aygıtı kullanmaktır. Başka bir yaklaşım, TTL darbelerini bir ağ bağlantısı üzerinden göndermek ve gecikmeyi ölçmek için ağ analiz araçlarını kullanmak gibi yazılım tabanlı yöntemler kullanmaktır. Başka bir yaklaşım, küresel konumlandırma sistemi (GPS) veya ağ zaman protokolü (NTP) sunucusu kullanarak veya donanım tabanlı bir senkronizasyon çözümü kullanarak iki bilgisayarın saatlerini senkronize etmek, zaman damgası ile her darbe için varış zamanı arasındaki gecikmeyi hesaplamak ve iki bilgisayar arasındaki genel gecikmeyi elde etmek için sonuçların ortalamasını almaktır.
- Görevi, bölüm 2 ve bölüm 4'te açıklanana benzer bir yapıya sahip iki koşulu (bir planlama koşulu ve bir kontrol koşulu) göz önünde bulundurarak kodlayın (bkz. Şekil 2 ve Ek Dosya).
- Planlama görevi için aşağıdaki yapıya sahip bir kod yazın: planlama dönemi, bakım dönemi, planlama yürütme dönemi, yanıt süresi ve geri bildirim (Şekil 2, Ek Dosya).
- Planlama dönemi: Temel olarak 3 sn için sunulan bir sabitleme çaprazını ayarlayarak planlama koşulunu başlatın.
- Labirent kümesini rastgele bir şekilde tek tek sunun (bu çalışmada 36).
NOT: Bu planlama döneminde, katılımcılardan bir dizi kuralı izleyerek en fazla 10 saniye olmak üzere dört hayvan bölgesini ziyaret etmek için bir yol planlamaları istenir (kurallar kendilerine daha önce açıklanmıştır; verilen kuralları görmek için protokolün 5. bölümüne ve Şekil 2'ye bakınız). - Bir etiket kodu kullanarak uyaran sunumunun başlangıcını işaret eden koda bir TTL tetikleyicisi ekleyin ve bu tetikleyiciyi daha dar ve pencereli analizler için EEG bilgisayarına ve göz izleyici ana bilgisayarına gönderin.
- Planlama süresinin, konu planlamayı bitirdiğinde veya maksimum süre aşıldığında joystick/klavyeden bir düğmeye basıldığında sona erdiğini koda yazın. Reaksiyon süresi (RT), daha fazla analiz için günlük dosyasına kaydedilmelidir.
NOT: Bu dönem için 1 rakamını kullanarak bir tetikleme kodu kullandık, ancak sayısal kodlar yerine hiyerarşik olay tanımlayıcıları (HED) etiketlerinin kullanılması önerilir, çünkü HED etiketleri içeriğe anlam ve yapı sağlar, böylece diğer araştırmacıların veya ortak çalışanların verilerin içeriğini anlamasını kolaylaştırır. - Bakım periyodu: Bu periyodu 3 saniye boyunca sunulan kaydırılmış bir fiksasyon çaprazı kullanarak başlatın. Hayvanat bahçesi haritasının başlangıç konumunu (kapısı) tahmin etmek için labirentin kapısının bulunduğu uzamsal konumda kaydırılmış sabitleme haçını bulun (bkz. Şekil 2).
NOT: Bu dönemin amacı üç yönlüdür. İlk olarak, kaydırılmış fiksasyon çaprazı, bir sonraki dönem için planlanan yolu temsil eden izin yürütülmesini kolaylaştırır (bkz. adım 3.2.8). İkincisi, bu süre zarfında katılımcılar, planlama döneminde detaylandırılan planı çalışma hafızalarında tutarlar. Son olarak, bu dönem, planlama döneminin sonunu ve bir sonraki dönemin başlangıcını - planlama yürütme dönemini sınırlamak için duruşmalar arası bir aralık görevi görür. - Planlama yürütme dönemi: Bakım periyodu boyunca 3 saniye boyunca kaydırılmış sabitleme haçı gösterildikten sonra, labirenti tekrar sunun.
- Belirli bir etiket kodu kullanarak bu dönemin başlangıcını belirtmek için EEG'ye ve ana göz izleyici bilgisayarına bir TTL tetikleyicisi gönderin.
- Bu sürenin başlamasından yaklaşık 992 ms sonra öznenin bakış pozisyonunun gerçek zamanlı görsel geri bildirimini (koyu bir çizgi, Şekil 2'deki yürütme süresine bakın) vermek için bir kod yazın.
NOT: Gecikmeli olarak (yaklaşık 1.000 ms) betimlemeye başlamak, deneklere labirentte yönlerini bulmaları için zaman verir ve önceki planladıkları yolu (planlama süresi boyunca) karanlık bir çizgiyle tanımlamalarına olanak tanır. - Denekler tarafından yapılan yolların daha fazla yeniden yapılandırılması için yolların koordinatlarını kaydedin ve performansı çevrimdışı olarak puanlayın (bkz. adım 6.1.1, Şekil 4).
- Planlanan yolu izlemek için maksimum 10 sn olduğundan emin olun ve deneklerin bir düğmeye basarak bu süreyi tamamlamalarına izin verin. Bu şekilde, denekler çizilen yollarını ne zaman bitirdiklerini kontrol edebilirler.
- Daha fazla analiz için RT'yi günlük dosyasına kaydedin.
- Yanıt süresi: Planlama yürütmesinden 10 saniye sonra veya planlama yürütme süresinin sonunda bir düğmeye basıldıktan sonra başlayan, labirentin kaybolduğu ancak hayvanların ve uzamsal konumlarının ekranda kaldığı yanıt süresi için bir kod yazın.
- Yanıt döneminde ekranın alt kısmına yatay olarak dört boş daire yerleştirin.
NOT: Bu sürenin amacı, deneklerin planlama yürütme döneminde ziyaret edilen hayvanların sırasını, hayvanları bir joystick veya klavye kullanarak ziyaret ettikleri sırayla dairelere koyarak belirtmelerini sağlamaktır. - Programı/kodu, deneklerin daha önce sunulan hayvanların her birini (bu çalışmada dört hayvan) seçmek için joystick veya klavyeyi kullanmalarına ve bunları dört dairenin her birine yerleştirmelerine izin verecek şekilde yapılandırın (bkz. Ek Dosya ve Şekil 2).
- Geri besleme: Katılımcılara 3 saniye geri bildirim sağlamak için bir kod yazın. Kurallara uyulması durumunda ziyaret edilen hayvanların geçerli kombinasyonlarına yanıt olarak bir başparmak yukarı resmi gösterilmeli, bildirilen kombinasyon geçersizse bir başparmak aşağı resmi gösterilmelidir.
- Doğru geri bildirim için belirli bir etiket kodu ve yanlış için başka bir etiket kodu kullanarak bir TTL tetikleyicisini EEG ve göz izleyici bilgisayarlara gönderin.
NOT: Geri bildirim vermenin nedeni, görev sırasında performansın izlenmesini kolaylaştırmak ve motivasyonu korumaktır. Bu gerçek zamanlı geri bildirimin sağlanması, ödül etkisini artırır ve uygun görev performansını teşvik eder103.
- Kontrol görevi için planlama koşulu ile aynı yapıya sahip bir kod yazın: bir kontrol periyodu, bir bakım periyodu, bir kontrol yürütme periyodu, bir yanıt periyodu ve geri bildirim (bkz. Ek Dosya, Şekil 2).
- Kontrol periyodu: Kafa karıştırıcı faktörleri azaltmak için kontrol koşulu dönemi için bir kod yazın. Bu periyodun kodu, temel olarak 3 sn için sunulan bir sabitleme çaprazı ile başlamalıdır.
NOT: Planlama görevi ağırlıklı olarak planlamanın uygulanmasını gerektirdiğinden, ancak aynı zamanda görsel-uzamsal işlev, çalışma belleği, dikkat kontrolü, engelleyici kontrol vb. gibi yürütücü işlevin bir parçası olarak diğer bilişsel alanları da işe aldığından.66,88,104,105, kafa karıştırıcı faktörleri azaltmak için bir kontrol görevi çok önemlidir. Bu nedenle, bu görevin temel amacı, bilişsel planlamanın uygulanmasını ortadan kaldırırken, planlama görevini çözmek için gereken tüm bilişsel ve algısal işlevleri talep etmektir54. - Kontrol koşulunun labirentlerini rastgele bir şekilde tek tek sunun (önceden izlenen işaretli bir yola sahip labirentler). Maksimum 10 s'lik bir süre kodlayın.
- Bir etiket kullanarak uyaran sunumunun başlangıcını işaret eden koda bir TTL tetikleyicisi ekleyin ve bu tetikleyiciyi EEG bilgisayarına ve göz izleyici ana bilgisayarına gönderin.
- Bu kontrol süresinin, konu bittiğinde veya maksimum süre aşıldığında joystick/klavyeden bir düğmeye basıldığında doruğa ulaştığını koda yazın.
NOT: Deneklere işaretli yolları değerlendirmeleri talimatı verilir (kurallara uyup uymadıklarına bakılmaksızın, katılımcılara verilen talimatlarla ilgili ayrıntılar için adım 5.12'ye bakın). - Daha fazla analiz için reaksiyon süresini (RT) günlük dosyasına kaydedin.
- Bakım periyodu: Kontrol periyodu sona erdiğinde, 3 saniye boyunca kaydırılmış bir sabitleme haçı sunun.
- Planlama yürütme dönemi olarak, bir sonraki dönem için bakış çizimini kolaylaştırmak için sabitleme haçını kapı girişinin bulunduğu yere yerleştirin.
- Kontrolün yürütme süresi: Labirenti tekrar sunun ve aynı anda EEG'ye bir TTL tetikleyicisi gönderin ve yürütme süresinin başlangıcını işaret eden bir etiketle göz izleyici bilgisayarlara ev sahipliği yapın.
- Bakış konumu hakkında çevrimiçi geri bildirim vermek ve bakışlarını izlenen yolla tanımlamak ve örtüştürmek için planlama yürütme dönemindekiyle aynı kodu tekrarlayın.
- Yolu izlemek için en fazla 10 saniyelik bir süre olduğundan emin olun ve deneklerin bir düğmeye basarak bu süreyi tamamlamalarına izin verin.
- Daha fazla analiz için RT'yi günlük dosyasına kaydedin.
- Kontrol yanıt süresi: Kontrol yürütme süresi sona erdiğinde, yanıt süresini gösteren bir soru işareti gösterin.
- Deneklerin bir joystick veya klavye kullanarak yanıt vermesi için sırasıyla iki düğme programlayın.
NOT: Burada, deneklerden izleme tarafından işaretlenen dizinin doğru olup olmadığını, doğru/EVET için bir düğme ve yanlış/HAYIR için başka bir düğme seçerek yanıtlamaları istenir. - Doğruluğu günlük dosyasına kaydedin.
- Geri besleme: Denekler doğru yanıt verdiğinde 3 sn doğru geri bildirim vermek için bir kod yazın (beğenme görüntüsü) ve denekler yanlış yanıt verdiğinde 3 sn yanlış geri bildirim verin (beğenmeme resmi).
- Planlama koşulunda olduğu gibi, EEG'ye bir TTL tetikleyicisi gönderin ve doğru geri bildirim için bir etiket ve yanlış geri bildirim için başka bir etiket ile göz izleyici bilgisayarları barındırın.
- Kontrol periyodu: Kafa karıştırıcı faktörleri azaltmak için kontrol koşulu dönemi için bir kod yazın. Bu periyodun kodu, temel olarak 3 sn için sunulan bir sabitleme çaprazı ile başlamalıdır.
- Eğitim görevleri: Uyaranlar oluşturun, bir kod yazın ve yukarıda belirtilen planlama ve kontrol görevlerinden önce her koşul için yaklaşık altı deneme/labirentten oluşan kısa bir eğitim oturumu sunun (planlama ve kontrol)
NOT: Buradaki fikir, görev ayarına aşina olmayı sağlamaktır. Devam etmek için kriterler belirlemeniz önerilir. Bu çalışmada, son üç deneme doğruysa ve katılımcılar eğitim oturumunun sonunda hedefi ve prosedürü anladıklarını bildirmişlerse, katılımcılar daha sonra deney oturumuna geçmişlerdir.

Şekil 3: Laboratuvar kurulum örneği. Birbirine bağlı üç bilgisayarı gösteren bir laboratuvar düzeneğinin şematik gösterimi. Ana bilgisayar (göz izleyici bilgisayar), göz hareketi verilerinin izlenmesinden ve saklanmasından sorumludur. EEG bilgisayarı, EEG sinyallerini alır ve saklar. Görüntü bilgisayarı davranışsal deneyi kontrol eder, uyaranları deneklere sunar ve veri toplamayı senkronize etmek için paralel bağlantı noktaları ve LAN bağlantıları aracılığıyla ana bilgisayar ve EEG bilgisayarlarına olay tetikleyicileri gönderir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 4: Göz izleyici sistemi tarafından verilen görsel çevrimiçi geri bildirimden yol rekonstrüksiyonu. Bir planın motor uygulamasından (A, mor renkte, planlama yürütme dönemi) ve bir kontrol yürütme periyodundan (B, yeşil çizgi) ve göz izleyici verileriyle bir yol yeniden yapılandırmasının açıklayıcı örnekleri. Planlama yürütme döneminde yeniden yapılandırılan yol, her planlama görevi denemesinin doğruluğunu değerlendirmek için kullanılır. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
4. Laboratuvar ortamı ve ekipmanları
- EEG elektrotları uluslararası 10-20 sistem106'ya göre yerleştirilmiş olarak katılımcının kafa derisinden EEG aktivitesini kaydetmek için bir EEG alım sistemi kullanın. Çevrimdışı yeniden referans için katılımcının mastoidlerine iki elektrot yerleştirin. Görsel inceleme sırasında dikey, yatay ve göz kırpan göz hareketi sinyallerini tanımlamak için elektrookülogram elektrotlarını kullanın.
- EEG bilgisayarında 2,048 Hz veya 1,024 Hz örnekleme hızı ve 0.1-100 Hz arasında bir bant geçiren filtre ile EEG veri toplama için EEG toplama yazılımını kullanın.
NOT: 1,024 Hz ve 2,048 Hz örnekleme hızları, düşük frekanslı salınımları analiz etmek için yeterli çözünürlük sağlar. Düşük frekanslı salınımları analiz ederken, düşük frekanslı sinyallerin örtüşmemesini sağlamak için düşük frekanslı salınımları analiz ederken, 1.000 Hz'in üzerinde gibi yüksek örnekleme hızına sahip EEG sinyallerini elde etmek önemlidir. - Uyaranları minimum 1.920 piksel x 1.080 piksel çözünürlüğe ve 60 Hz yenileme hızına sahip genişletilmiş bir monitöre yansıtmak için EEG bilgisayarına ve ana göz izleyici bilgisayara paralel portlar ve ethernet aracılığıyla bağlanan ve üzerinde davranışsal deneyler yapmak için bir platforma sahip bir ekran bilgisayarı kullanın. Bu monitörü konudan yaklaşık 82 cm uzağa yerleştirin.
NOT: Katılımcıdan 24 cm uzakta bulunan yenileme hızı 144 Hz olarak ayarlanmış 82 inç monitör kullandık. EEG ve göz hareketlerinin kaydedilmesini içeren bilişsel deneyler için ekran boyutu en az 19 inç olan bir monitör kullanılması önerilir. Ek olarak, genel bir öneri, monitörü, katılımcının görevi rahatça yerine getirmesine ve EEG ve göz hareketlerinin doğru bir şekilde kaydedilmesine izin verirken ekrana sabit bir bakış atmasına olanak tanıyan bir mesafeye yerleştirilmesidir. En iyi sonuçları elde etmek için kurulumu gerektiği gibi test etmeniz ve ayarlamanız önerilir. - Yürütme dönemlerinde katılımcılara göz hareketleri hakkında gerçek zamanlı geri bildirim vermek için bir göz izleme sistemi kullanın ve göz bebeği boyutunu kaydedin. Yeterli zamansal çözünürlük için örnekleme hızını 1.000 Hz'e ayarlayın.
- Baş hareketlerinden kaçının. Gözü video kameranın görüş alanı içinde tutmak için sola ve sağa ve yukarı ve aşağı baş hareketi kısıtlamaları gereklidir. Gözü video kameranın odak aralığında tutmak için ileri ve geri hareket kısıtlaması gereklidir. Hareketleri bu aralıkta tutmak için alın/çene desteklerinin bir kombinasyonunu kullanın.
- Planlama uyaranları ile kontrol uyaranları arasındaki farkları karşılaştırmak için dijital bir lüks ölçer veya benzeri bir şey kullanarak uyaranların parlaklığını değerlendirin.
NOT: İki durumun uyaranları arasındaki farkları değerlendirmek için t-testi veya Wilcoxon gibi istatistiksel bir test kullanılabilir. - En az dört düğmeli bir kontrol joystick'i veya klavyesi kullanın: kontrol koşulundan Evet/Hayır soruları için iki düğme; denemeleri bitirmek için bu iki düğmeden biri; ve hayvanları ekranın altındaki dört dairenin her birine yerleştirmek için ileri veya geri hareket etmek için planlama koşulunun yanıt süresi için iki düğme daha.
5. Elektroensefalografi ve göz izleme kayıt seansları
- Çalışmaya başlamadan önce, katılımcıların yazılı ve imzalı bilgilendirilmiş onam almalarını sağlayın.
- Kayıt oturumundan önce, katılımcılardan makyaj yapmamalarını (maskara ve göz kalemi göz takip sistemi tarafından göz bebeği olarak algılanabilir), uyuşturucu veya kafein107,108 almamalarını veya şiddetli yorgunluk109 hissetmelerini (stres, uyku yoksunluğu vb.) isteyin.
- Katılımcılardan, eğitimli bir psikolog tarafından uygulanan MINI-Uluslararası nöropsikiyatrik görüşme97 aracılığıyla cinsiyetleri, yaşları, el becerileri, ana dilleri ve nöropsikiyatrik geçmişleri hakkında bilgi sağlamak için demografik bir anket doldurmalarını sağlayın.
- Deneğin alnını, kafa derisini, mastoidlerini ve elektrookülogram (EOG) cilt pozisyonunu alkollü bir mendille temizleyin.
- Tüm harici elektrotları katılımcının üzerine yerleştirin. Yatay EOG'yi bipolar olarak her iki gözün dış kantisine ve dikey EOG'yi katılımcının sağ gözünün üstüne ve altına yerleştirin. Daha sonra yeniden referans vermek için sağ ve sol mastoide iki harici elektrot yerleştirin.
- Deneğin kafasını ölçün ve genişletilmiş uluslararası 10-20 sistemine göre doğru boyuttaki EEG kapağını yerleştirin. Bunu yapmak için, aşağıdaki adımları izleyerek Cz elektrodunu bulun ve yerleştirin:
- Saç çizgisini ve burnun üst kısmını görsel olarak inceleyerek kafa derisinin orta çizgisini belirleyin. Orta çizgiyi tanımlamak için bu iki noktayı birleştiren bir çizgi belirleyin.
- Cz'yi bulun. Cz tipik olarak iki preauriküler nokta arasındaki orta nokta (yani, her bir kulağın hemen önünde bulunan noktalar) olarak tanımlanır. Bu noktaları bulun ve ardından Cz'nin yaklaşık konumunu belirlemek için bunları birbirine bağlayan bir çizgi belirleyin.
- Cz'yi ölçün ve işaretleyin. Nasion'dan (yani burnun üst kısmındaki kemikli çıkıntı) Cz'ye olan mesafeyi ölçün. Nasiondan Cz'ye olan mesafe tipik olarak 10-20 sistemindeki toplam baş çevresinin yaklaşık% 53'ü kadardır. Bir kalem veya başka bir işaretleme aracı kullanarak Cz'nin konumunu işaretleyin.
NOT: Hataları en aza indirmek ve EEG verilerinin geçerliliğini sağlamak için elektrot yerleştirme için tutarlı ve standartlaştırılmış bir prosedür izlemek önemlidir. Standart bir yerleştirme prosedürü geliştirilmesi tavsiye edilir. Elektrotları her deneğin kafa derisine yerleştirmek için standart bir prosedür oluşturun ve her denek için aynı prosedürün kullanıldığından emin olun. Kayıtları gerçekleştiren bir ekip veya personel olması durumunda, elektrotları tutarlı ve doğru bir şekilde yerleştirdiklerinden emin olmak için teknisyenleri veya araştırma görevlilerini uygun yerleştirme prosedürü konusunda eğitin. Ayrıca, her denek için elektrot konumlarının sayısallaştırılması, daha sonra kaynak analizi yapmak için arzu edilen bir yaklaşım olacaktır. EEG çalışmalarında, her bir elektrotun deneğin kafasındaki kesin üç boyutlu (3D) konumu, EEG verileri ile karşılık gelen beyin aktivitesi110 arasında bir korelasyon kurmak için sıklıkla bir gerekliliktir. Bu bilgi aynı zamanda EEG verilerinin MRI veya BT görüntüleme tekniklerinden elde edilen anatomik görüntülerle uygun şekilde hizalanması için de kritik öneme sahiptir111,112.
- Körelmiş bir iğneye sahip bir şırınga kullanarak kapağın her bir deliğine iletken jel yerleştirin ve katılımcının saçını uçla uzaklaştırın. Daha sonra, tüm kafa derisi elektrotlarını EEG kapağına yerleştirin.
- EEG kayıt yazılımını kullanarak empedansları kontrol edin ve EEG sistemi tarafından önerilen direnç seviyesinin altında olduklarından emin olun.
- Katılımcıdan deney sırasında mümkün olduğunca hareketsiz kalmasını isteyin. EEG sinyalini inceleyin ve katılımcıdan gözlerini kırpmasını, çene yapmasını ve gözleri kapalıyken birkaç saniye kalmasını isteyerek test edin.
- Katılımcıyı karanlık ve sesi azaltılmış bir odaya oturtun. Başlarını stabilize etmek ve hareketi en aza indirmek için bir çene desteği kullanın ve çene desteği ile uyaran sunum ekranının merkezi arasında yaklaşık 82 cm'lik bir mesafe olduğunu kontrol edin.
- Yanıtlar için katılımcının önüne bir oyun çubuğu veya klavye yerleştirin.
- Talimat -ları: Her duruma başlamadan önce görsel yardımcılar kullanarak sözlü talimatlar verin (planlama ve kontrol). Talimatlarda, uyaranların görsel örneklerini ekleyin ve sırasıyla planlama ve kontrol koşullarında labirentlerin nasıl çözüleceğini açıklayın.
- Planlama görevi için, deneklere, labirentin farklı yerlerinde herhangi bir sırayla ve bir dizi kuralı izleyerek belirli hayvan konumlarına (bu çalışmada dört yer) bir dizi ziyareti tamamlamak için bir yol bulmalarını söyleyin: "(1) Yolu en fazla 10 saniye içinde mümkün olduğunca hızlı planlayın; (2) Geçitten başlayın ve ziyaret edilen dördüncü hayvanda yolu sonlandırın; (3) Aynı yoldan veya köşeden iki kez geçmeyin; (4) Çıkmaz bir sokağı geçmeyin; (5) Bir yolu dik olarak geçmeyin"54.
NOT: Kural 2'den kural 5'e kadar olan bölümler için katılımcıya görsel örnekler göstermenizi öneririz. - Daha sonra, altı denemeden oluşan planlama görevi eğitim oturumunu başlatın.
NOT: Katılımcılara, özellikle yürütme periyodundaki yolu tanımlarken, görevi yerine getirirken herhangi bir sorun olup olmadığını her göz izleyici kalibrasyon periyodundan önce bildirmeleri talimatını verin. Çevrimdışı/son işlemeyi kontrol etmek için bir deneme olup olmadığını not edin (bkz. adım 6.1.1). - Kontrol koşulu için, deneklere labirent üzerinde önceden işaretlenmiş yolun daha önce öğrenilen kurallar göz önünde bulundurularak doğru mu yoksa yanlış mı yapıldığını değerlendirmelerini söyleyin.
NOT: Hata tespit edildiğinde yeni bir yol planlamaya çalışmamak (çizimlerin aynı yolu iki kez kullanması, çıkmaz bir sokağı geçmesi vb.) gibi planlama stratejileri kullanmadan labirentlerin nasıl değerlendirileceğine dair görsel destekli örnekler verin. Bir hata bulunduğunda, yolu düzeltmek yerine yalnızca hatanın algılandığını bildirmeye odaklanılmalıdır. Her denemeden sonra, katılımcılara uyguladıkları stratejiler hakkında sorular sorun. Ardından, çizilen yolları değerlendirdiklerinden ve yeni yollar planlamaktan kaçındıklarından emin olmak için performansları hakkında sözlü geri bildirimde bulunun. Daha sonra, altı denemeden oluşan kontrol görevi eğitim oturumunu başlatın.
- Planlama görevi için, deneklere, labirentin farklı yerlerinde herhangi bir sırayla ve bir dizi kuralı izleyerek belirli hayvan konumlarına (bu çalışmada dört yer) bir dizi ziyareti tamamlamak için bir yol bulmalarını söyleyin: "(1) Yolu en fazla 10 saniye içinde mümkün olduğunca hızlı planlayın; (2) Geçitten başlayın ve ziyaret edilen dördüncü hayvanda yolu sonlandırın; (3) Aynı yoldan veya köşeden iki kez geçmeyin; (4) Çıkmaz bir sokağı geçmeyin; (5) Bir yolu dik olarak geçmeyin"54.
- Tüm kanalların doğru bir şekilde alındığından emin olmak için EEG sinyalini kontrol edin. EEG kaydını başlatın.
- Göz izleyiciyi kalibre edin.
NOT: Katılımcı bakışlarını ekranın çeşitli bölgelerine yönelttiğinde göz izleyicinin bakış konumunu belirleme yeteneğini doğrulayın.- Katılımcıya göz izleyicinin kalibre edileceğini ve ekranın dört köşesine rastgele hareket eden beyaz bir daire (küçük gri noktalı) göreceklerini bildirin (beş noktalı kalibrasyon prosedürü). Bakışlarını daireye sabitlemelerini söyleyin ve daire başka bir yere hareket ettiğinde dairenin konumunu takip etmeleri ve bakışlarını bu yeni pozisyonda tekrar sabitlemeleri gerektiğini bildirin.
- Deneyi çalıştırın, Çıktı/Kaydet'e tıklayarak göz hareketlerini kaydetmeye başlayın ve katılımcıdan daha önce verilen talimatları izlemesini isteyin ve deneyin şimdi başlayacağını bildirin.
- Laboratuvar odasını karanlık bir ortamda muhafaza ediniz. Göz bebeği genişlemesindeki en büyük değişiklikler, parlaklıktaki değişikliklere yanıt olarak meydana gelir113. Deney ortamında tutarlı bir ışık seviyesini koruyun.
6. Veri analizleri
- Davranışsal analizler
- İstatistiksel yazılım kullanarak davranışsal verileri analiz edin (bkz . Malzeme Tablosu). Doğruluğu (doğru yanıtların yüzde oranı) hem planlama hem de kontrol koşullarında nicel bir davranışsal parametre olarak ölçün. Planlama koşulu için, yürütme süresi boyunca alınan yolları çevrimdışı olarak yeniden oluşturmak için göz izleyici verilerini (bakış konumunun x ve y koordinatları) kullanın ve planlanan yolların doğruluğunu gerçek izlenen yollarla karşılaştırmalı olarak belirleyin (Şekil 4). Bunu yapmak için, yanıt döneminde doğru/yanlış yapılan kombinasyonlar ile yapılan izleme arasındaki uyumu manuel olarak kontrol edin.
- Planlama dönemi için labirentleri çözmek için harcanan ortalama süre ve kontrol dönemi için işaretli yolları değerlendirmek için harcanan ortalama süre olan RT'yi hesaplayın.
- Planlama ve kontrol koşulları için yürütme sürelerinin ortalama RT'sini hesaplayın. Özellikle, yalnızca doğru denemelere karşılık gelen RT'yi kullanın.
NOT: Tamamlayıcı olarak, reaksiyon süresini ve doğruluğunu dikkate alan birleşik bir ölçüm sağlayan Domic-Siede ve ark.54'te açıklanan doğrusal entegre hız-doğruluk puanını (LISAS)114,115 kullanmak mümkündür. Planlama yürütme süresi sırasındaki tepki süresi ve planlamanın doğruluğu birbiriyle ilişkili olduğundan, LISAS, yapılan hataların sayısı için düzeltilen tepki süresini hesaba katan bir endeksi hesaplamak için kullanılabilir. Ek olarak, LISAS indeksi, elektrofizyolojik sinyaller ile davranışsal performans arasındaki korelasyonu değerlendirmek için de kullanılabilir. Reaksiyon süresi (RT) ve hata oranının (PE) doğrusal bir kombinasyonu olarak hesaplanır. - Levene Testi116,117 gibi istatistiksel bir test kullanarak homoskedastisiteyi değerlendirin ve karşılaştırmalar için uygun istatistiksel testi (parametrik veya parametrik olmayan) seçmek için D'Agostino ve Pearson omnibus normallik dağılım testi118 veya Shapiro Wilk testi119 kullanarak normalliği test edin.
- Koşullar arasındaki davranışsal parametreleri karşılaştırmak için Wilcoxon işaretli sıra testi120 veya eşleştirilmiş çift t-testi121 kullanarak planlama koşulundaki planlama bileşeninin kontrol koşulundan bilişsel olarak daha talepkar olup olmadığını değerlendirin.
NOT: Bu şekilde, davranışsal paradigmanın bilişsel planlamayı değerlendirmek için en uygun olduğunu doğrulayın. - Planlama koşulundaki denemeleri "kolay" ve "zor" kategorilere ayırın (adım 2.2'ye bakın) ve ardından "kolay" ve "zor" denemeler arasındaki planlama ve yürütme dönemlerindeki doğruluk ve tepki sürelerini karşılaştırmak için eşleştirilmiş bir çift t-testi kullanın.
- EEG ve göz hareketi ön işleme
- Delorme ve Makeig122'de, Dimigen ve ark.123'te ve Mognon ve ark.124'te bir programlama dili yazılımında açıklananlar gibi, kendi kendine yapılan komut dosyalarını ve/veya yerleşik araç kutularını kullanarak aşağıdaki noktalarda açıklanan EEG veri ön işleme hattını gerçekleştirin (bkz.
- Daha iyi görsel inceleme veya daha ileri analizler için fiksasyonları, sakkadları ve göz kırpma olaylarını içe aktarmak için göz hareketi aktivitesini EEG kayıtlarıyla senkronize edin (bkz. adım 3.1.2 ve Ek Dosya).
NOT: Bu çalışmada, göz hareketi olaylarını bir programlama dili yazılımında EEG verilerine aktarmak için Domic-Siede ve ark.54 ve Dimigen ve ark.123'te açıklandığı gibi göz izleme verileri üzerindeki zaman damgalarını ve EEG verileri üzerindeki zaman damgalarını kullandık. - Hesaplama taleplerini azaltmak için 2.028 Hz'de kaydedilmiş olmaları durumunda verileri 1.024 Hz'e kadar alt örnekleyin.
NOT: 1,024-4 Hz'lik ilgilenilen frekans aralığına, beklenen frekans çözünürlüğüne ve analizin hesaplama gereksinimlerine göre 8 Hz'lik bir örnekleme hızı yeterlidir. - EEG sinyalini mastoidler üzerindeki elektrotların ortalamasına yeniden referans verin.
NOT: Başka referanslar da mümkündür. Referans seçimi, EEG analizinin sonuçlarını ve verilerin yorumlanmasını etkileyebilir, bu nedenle farklı referans seçeneklerinin artılarını ve eksilerini dikkatlice değerlendirmek önemlidir. Ortalama mastoid referansı, EEG çalışmaları için popüler bir seçimdir, çünkü hesaplanması kolay kararlı bir referans sağlar ve birçok farklı EEG sinyalini analiz etmek için etkili olduğu gösterilmiştir. EEG verilerini mastoidlerin ortalamasına (ortalama mastoid referansı olarak bilinir) atıfta bulunmak, kafa derisi EEG verilerindeki frontal teta aktivitesini analiz etmek için yaygın bir yaklaşımdır. Mastoid elektrotlar kulağın yakınında bulunur ve EEG sinyalleri için bir referans sağlar. Mastoidlerin ortalamasına atıfta bulunmak, ilgilenilen sinyali iptal etmekten kaçınırken, ilgi çekici olmayan gürültü ve artefaktların etkisini azaltmaya yardımcı olabilir, bu da kullanıcının EEG sinyallerinin daha net bir temsilini elde etmesine yardımcı olur. - Bir programlama dili yazılımı kullanarak genişletilmiş sinyal üzerinden (epoching olmadan) 1 Hz'lik yüksek geçişli kesme frekansı ve 40 Hz'lik düşük geçişli kesme frekansına sahip sıfır fazlı bir sonlu darbe tepkisi (FIR) uygulayın.
NOT: Bu çalışmada Delorme ve Makeig122'de anlatılan araç kutusu kullanılmıştır. - Her koşul için, deneme sayısını göz önünde bulundurarak, verileri sırasıyla planlama ve kontrol dönemlerinin başlangıcına odaklanan dönemlere bölün. Labirent sunumunun başlamasından önceki 1 saniyeyi temel olarak ve planlama veya kontrol döneminden sonraki 4 saniyeyi ilgilenilen bölümler olarak kullanın. Bir programlama dili yazılımı kullanın.
NOT: Bu çalışmada Delorme ve Makeig122ve 36 dönem/denemelerinde anlatılan araç kutusu kullanılmıştır. - Bakım periyodu olarak bitimden 4 sn önce ve 1 sn sonra kullanarak planlama ve kontrol periyotlarının sonuna odaklanan ikinci bir segmentasyon oluşturun.
NOT: Planlama ve kontrol periyotlarının ilk ve son 4 saniyesinin seçilmesinin nedeni (adım 6.2.6 ve adım 6.2.7), her iki koşulda da her dönemin süresinin değişebilmesi ve planlamanın ilk ve son saniyelerinin analiz edilmesinin planlama sürecine ilişkin daha kapsamlı bir görünüm sağlayabilmesidir. Bu nedenle, bu pencere uzunlukları, planlamanın altında yatan salınım dinamiklerini analiz etmek için yeterli ve yeterlidir. - Segmentlere ayrılmış sinyal üzerinde, yapay bileşenleri tanımlamak ve kaldırmak için Logistic Infomax bağımsız bileşenler analizi (ICA) algoritması125'içalıştırın.
- Potansiyel gürültülü bileşenleri otomatik olarak tespit etmek için Plöchl ve ark.126'daönerilen sakkad-fiksasyon varyans oranı kriterini kullanın ve Mognon ve ark.124'te önerilen uzamsal ve zamansal özelliklerin ortak kullanımına dayalı otomatik EEG artefakt dedektörünü kullanın.
NOT: Pion-Tonachini ve ark.127'de önerilen, bağımsız bileşen sınıflandırmalarını yedi kategoride bileşimsel vektörler olarak tahmin eden ve artefaktların tanımlanmasına yardımcı olan bağımsız bileşen sınıflandırıcısının kullanılmasını öneririz. - EMG, elektrot hareketi veya beyinle ilgili olmayan bileşenler gibi diğer potansiyel artefaktal bileşenleri inceleyin. Zaman içinde topografyaları, spektrumları ve aktivasyonları görsel olarak inceleyerek bu bileşenlerin reddini doğrulayın.
- Basıklık kriterini kullanarak otomatik kanal reddi ile gürültülü kanalları enterpolasyon (küresel enterpolasyon) (eşik olarak 5 z-skoru ile).
- Zaman-frekans analizleri
- 250 ms'lik bir pencere uzunluğu ve 5 ms'lik bir zaman adımı kullanarak kısa süreli hızlı Fourier dönüşümü (FFT) (1 Hz - 40 Hz) gerçekleştirin. Bir Hanning penceresi kullanın. Zaman-frekans çizelgelerini taban çizgisine (-1 s ila -0,1 s) normalleştirmek için z-skorunu kullanın.
NOT: Spektrumun görselleştirilmesi, pencere boyutu ile zamansal çözünürlük arasında bir dengeye tabidir. 4 Hz ila 8 Hz teta aralığı da dahil olmak üzere tüm spektrumun kapsamlı bir görünümünü elde etmek için, her deneme ve görev sırasında daha yüksek bir zamansal çözünürlük sağlamak için pencere boyutunun alt sınırı olan 250 ms'yi kullanmanızı öneririz. Ek olarak, bu durumlar için yaygın olarak geleneksel bir seçim olarak kabul edildiğinden, bir Hanning penceresi kullanmanızı öneririz. Zaman ve frekansta daha iyi çözünürlük için sonraki adımlara bakın. - Fz gibi bir frontosantral elektrottan veya ortalama bir frontal elektrot grubundan bir zaman-frekans tablosu seçin.
NOT: Bilişsel kontrol ile frontal orta hat teta 12,128,129 arasındaki ilişkiye ilişkin geniş kanıtları göz önünde bulundurun. - Karşılaştırmaları ilerletmek için Pz ve Oz elektrotları gibi elektrotlardan önden olmayan kontrol zaman-frekans çizelgelerini seçin.
- Frontal ve kontrol elektrotları için, her iki koşuldan zaman-frekans çizelgelerinin grup düzeyinde karşılaştırmaları için p değeri < 0,05 olan eşleştirilmiş numuneler için parametrik olmayan küme tabanlı bir permütasyon testi gerçekleştirin. 1.000 rastgele çekiliş ile Monte Carlo yöntemini kullanın. 130 permütasyon testini gerçekleştirmek için kümenin maksimum istatistik değerini kullanın.
- Sırasıyla planlama ve kontrolün ilk 4 saniyesinden teta frekans bandının (4-8 Hz) ve son 4 saniye segmentinin de ortalamasını alın.
- Eşleştirilmiş çift t-testi veya Wilcoxon işaretli sıra testi kullanarak koşullar arasındaki ortalama teta aktivitesini karşılaştırın.
- Teta aktivitesinin zaman profilini analiz edin. Bunu yapmak için, deneğe göre denemeler boyunca ortalama 4-8 Hz frekans aralığının ortalamasını alın.
- Yanlış keşif oranı (FDR) ile eşleştirilmiş ve düzeltilmiş bir Wilcoxon işaretli sıra testi kullanarak koşullar arasındaki teta aktivite dinamiklerini karşılaştırın.
NOT: Wilcoxon testinde örtüşmeyen pencerelerin 88 ms'lik adımlarını kullandık.
- 250 ms'lik bir pencere uzunluğu ve 5 ms'lik bir zaman adımı kullanarak kısa süreli hızlı Fourier dönüşümü (FFT) (1 Hz - 40 Hz) gerçekleştirin. Bir Hanning penceresi kullanın. Zaman-frekans çizelgelerini taban çizgisine (-1 s ila -0,1 s) normalleştirmek için z-skorunu kullanın.
- Kaynak rekonstrüksiyonu
- Tadel ve ark.131'de açıklanan açık erişimli araç kutusu veya benzer bir araç kutusu gibi kaynak analizi yeniden yapılandırması için bir araç kutusu kullanın.
- Standartlaştırılmış düşük çözünürlüklü beyin elektromanyetik tomografisi (sLORETA)132ve minimum norm görüntüleme yönteminin yanı sıra simetrik sınır eleman yöntemi (simetrik BEM) gibi bir algoritma kullanarak, ters problemi çözmek için Gramfort et al.133'teanlatıldığı gibi bir araç kutusu yardımıyla planlamanın ilk 4 saniyesinden itibaren önceden işlenmiş EEG sinyalinin kaynaklarını hesaplayın.
- Elektrotların 3D sayısallaştırılması olmaması durumunda her katılımcı için varsayılan elektrot konumlarıyla anatomik bir MNI şablonunda (Beyin Fırtınası "Colin27"de MNI şablonunu kullandık) kaynak algoritmayı (bu çalışmada sLORETA algoritması) kullanın (bkz. adım 5.6).
NOT: Varsayılan elektrot konumlarını kullanmanın, beyin aktivitesinin kaynaklarını belirlemek için en verimli yöntem olmadığı unutulmamalıdır. Bununla birlikte, yine de faaliyetin kökeni hakkında genel bir anlayış sağlayabilir. Bu yöntemlerle elde edilen yerelleştirme kaynaklarının kaba yaklaşımlar olduğunu ve sonuçların analizi sırasında dikkatle yorumlanması gerektiğini akılda tutmak önemlidir. - Önceden işlenmiş sinyalin üzerine 4-8 Hz bant geçiren bir filtre uygulayın.
- Temel olarak deneme başlangıcından önceki −1.000 ms ila −10 ms dönemini kullanarak bir z-skoru normalizasyonu uygulayın.
- Deneme başlangıcından sonra 1 saniye ile 4 saniye arasında bir ilgi penceresi kullanarak teta aktivitesinin ortalamasını alın.
- Monte Carlo örneklemesi (1.000 randomizasyon) ile parametrik olmayan bir permütasyon işareti testi kullanarak koşullar arasındaki ortalama uzay kaynaklarını karşılaştırın131.
- İlgi alanlarını (ROI'ler) belirlemek için, korteksi bir beyin atlası kullanarak etiketleyin.
NOT: Tadel veark.131'de açıklanan araç kutusunda uygulanan Destrieux Atlas 134'ü kullandık. - İlgilenilen beyin bölgelerini (ROI'ler) seçin.
NOT: Bilateral superior frontal gyri (SF), bilateral transvers frontopolar gyri ve sulci (FP), bilateral ACC, bilateral MCC ve bilateral dorsolateral prefrontal korteks137,138 gibi prefrontal korteks bölgelerinin bilişsel kontrol işlevlerinde yer aldığını bildiren kanıtları dikkate aldık135,136. - Her ROI için önceki önceden işlenmiş EEG sinyali (1-40 Hz aralığı) üzerinde temel bileşen analizi (PCA) gerçekleştirin ve her ROI için PCA ayrıştırmasının ilk modunu alın.
- Kısa süreli hızlı bir Fourier dönüşümü kullanarak spektral bir analiz yapın ve parametrik olmayan, küme tabanlı bir permütasyon testi130 kullanarak sol ve sağ ilgilenilen bölgeler arasındaki sonuçları karşılaştırın.
- Fark göstermeyen sol ve sağ ROI'leri tek bir ikili zaman serisi olarak ayıklayın ve temsil edin: SF, ACC ve MCC. Ardından, zaman-frekans çizelgelerini çizin ve koşullar arasında karşılaştırma yapın.
- Her bir ROI için planlama görevinin karmaşıklık düzeyine (kolay ve zor denemeler) göre zaman-frekans çizelgelerini karşılaştırın.
- 512 örneğin her biri için sinyalin kenarını yansıtın ve seçilen ROI'ler için 4 Hz ile 8 Hz arasında bir bant geçiren filtre gerçekleştirin.
- Bir programlama dili yazılımından bir sinyal işleme araç kutusu kullanarak anlık genlik139'uelde etmek için bir Hilbert dönüşümü uygulayın (bkz. Malzeme Tablosu).
- Z-skoru normalleştirmesini (temel olarak -1.000 ila -10 ms) kullanarak sinyali düzeltin ve deneğe göre denemelerin ortalamasını alın.
- Wilcoxon işaretli sıra testini (eşleşen çiftler, 1 sn örtüşmeyen pencereler) kullanarak koşullar arasındaki her ROI teta bandı zaman profilini karşılaştırın ve FDR ile düzeltin.
- EEG aktivitesi ile davranışsal performans arasındaki korelasyonlar
- Z skorunu kullanarak ROI'lerin kaynak zaman serisini taban çizgisine göre normalleştirin. Planlama veya kontrol başlangıcından sonra 1 sn ile 4 sn arasında bir pencere seçin (zaman-frekans çizelgelerinde belirgin teta aktivitesinin gözlemlendiği yer).
- Kontrol koşuluna kıyasla planlama koşulundaki teta aktivitesindeki artışı belirlemek için, önce sinyali frekans alanına (1-40 Hz) dönüştürün Chronux araç kutusu140 gibi bir araç kutusu aracılığıyla multitaper yöntemini kullanarak ilgili bölgelerdeki her koşul ve kaynak.
- Teta bandının ortalama frekansını (4-8 Hz) hesaplayın ve teta gücünün iki ölçüsünü hesaplayın: i) planlama periyodu (θ planlama) ve kontrol periyodu (θ kontrolü) sırasındaki teta gücü arasındaki fark, Δ teta olarak gösterilir ve ii) Δ teta (Δ θ) ve kontrol periyodu boyunca teta aktivitesinin oranı (θ kontrolü) olarak ifade edilen teta aktivitesindeki nispi artış, Domic-Siede ve ark.54'te olduğu gibi:
 (1)
(1)

- İki davranışsal parametreyi hesaplayın: iii) Δ LISAS planlaması, LISAS planlamasından LISAS kontrolünü çıkararak ve iv) Δ LISAS planlama yürütmesi, LISAS kontrol yürütmesini LISAS planlama yürütmesinden çıkararak, Domic-Siede ve ark.54'te olduğu gibi:


- Hesaplanan elektrofizyolojik ve davranışsal parametreleri kullanarak Spearman'ın rho korelasyonlarını gerçekleştirin ve ardından FDR ile düzeltin.
- Göz hareketi analizi: Farklı salınım dinamiklerine neden olabilecek her durum için göz hareketlerindeki potansiyel farklılıkları kontrol etmek için aşağıdaki analizi gerçekleştirin:
- Planlama ve kontrol koşulları sırasında tüm denemeden ve 0 s ila 3.75 s arasında sakkad genliğini ve sakkad tepe hızını belirleyin.
- Sonuçları, hangisi uygunsa, Wilcoxon işaretli sıra testi veya eşleşen çift t-testi kullanarak karşılaştırın.
NOT: Dimigen ve ark.123'teaçıklanana benzer bir araç kutusu yardımcı olabilir. - Bir frontal elektrottaki Fourier EEG gücü (örneğin, Fz veya ortalama bir frontal ROI elektrodu) ile Sato ve Yamaguchi141'de açıklandığı gibi sakkad hızı arasındaki tutarlılığı hesaplayın ve değerlendirin.
- İki koşul arasındaki her denemenin ilk 4 saniyesinin tutarlılık güç-sakkad hızı değerlerini karşılaştırmak için Wilcoxon işaretli sıra testini kullanın.
Sonuçlar
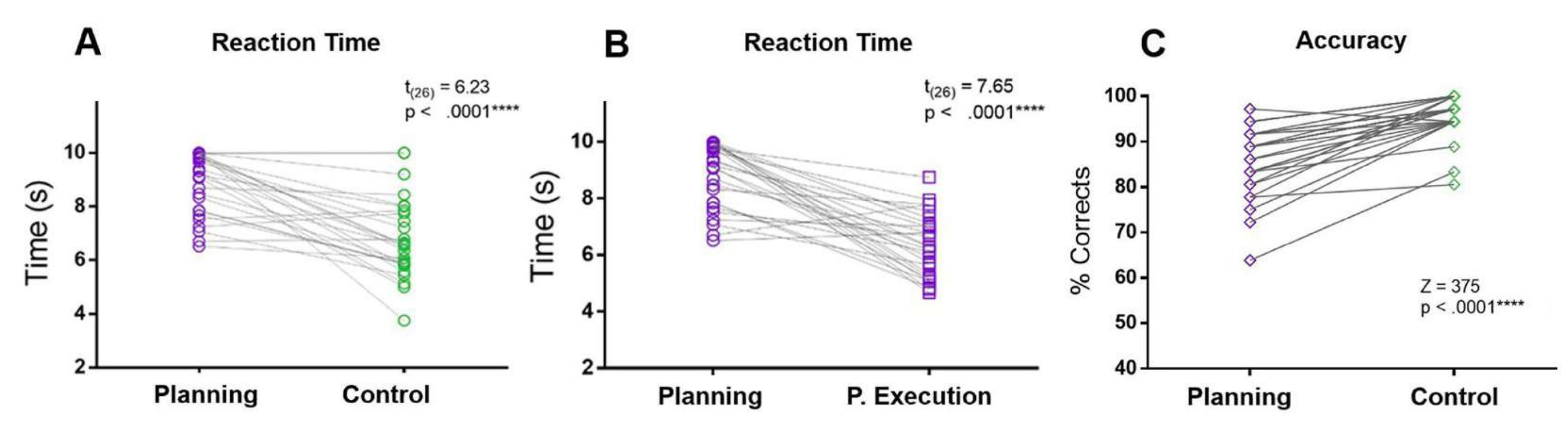
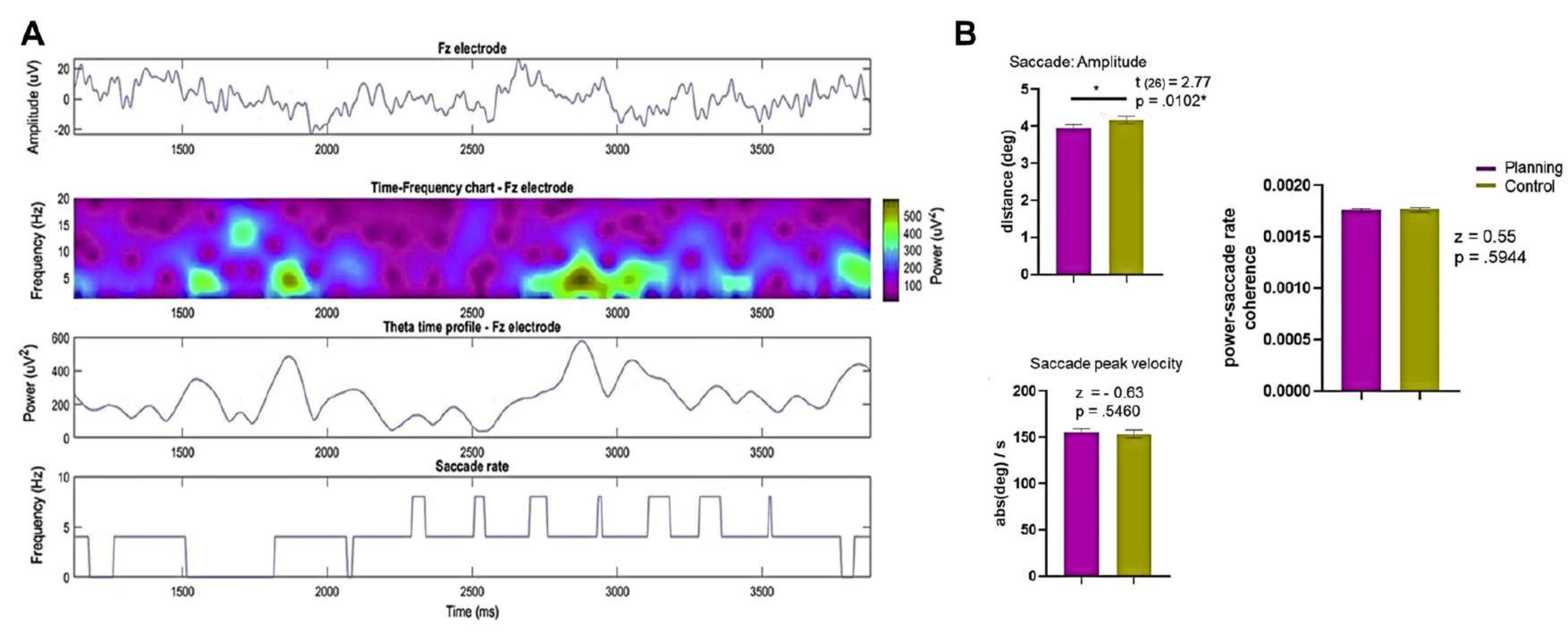
Mevcut protokolde, planlama döneminin RT'si, kontrol döneminin ve planlama yürütme döneminin RT'leri ile karşılaştırılmıştır. Ek olarak, kontrol koşulu ile karşılaştırıldığında, katılımcılar planlama süresi boyunca daha fazla hata yaptılar ve daha düşük doğruluğa sahiptiler (Şekil 5).

Şekil 5: Planlama görevi için tepki süresi ve doğruluğu. Planlama periyodundaki (A) reaksiyon süreleri (mor daireler) ve kontrol periyodu (yeşil daireler) arasında eşleştirilmiş eşleştirilmiş t-testi kullanılarak karşılaştırma. (B) Planlama dönemindeki reaksiyon süreleri (mor daireler) ile planlama yürütme dönemindeki (mor kareler) reaksiyon süreleri arasında eşleştirilmiş-eşleştirilmiş t-testi kullanılarak karşılaştırma. (C) Planlama koşulundaki (mor elmaslar) ve kontrol koşulundaki (yeşil elmaslar) doğruluk oranının bir Wilcoxon işaretli sıra testi kullanılarak karşılaştırılması. Bu rakam Domic-Siede et al.54'ten değiştirilmiştir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
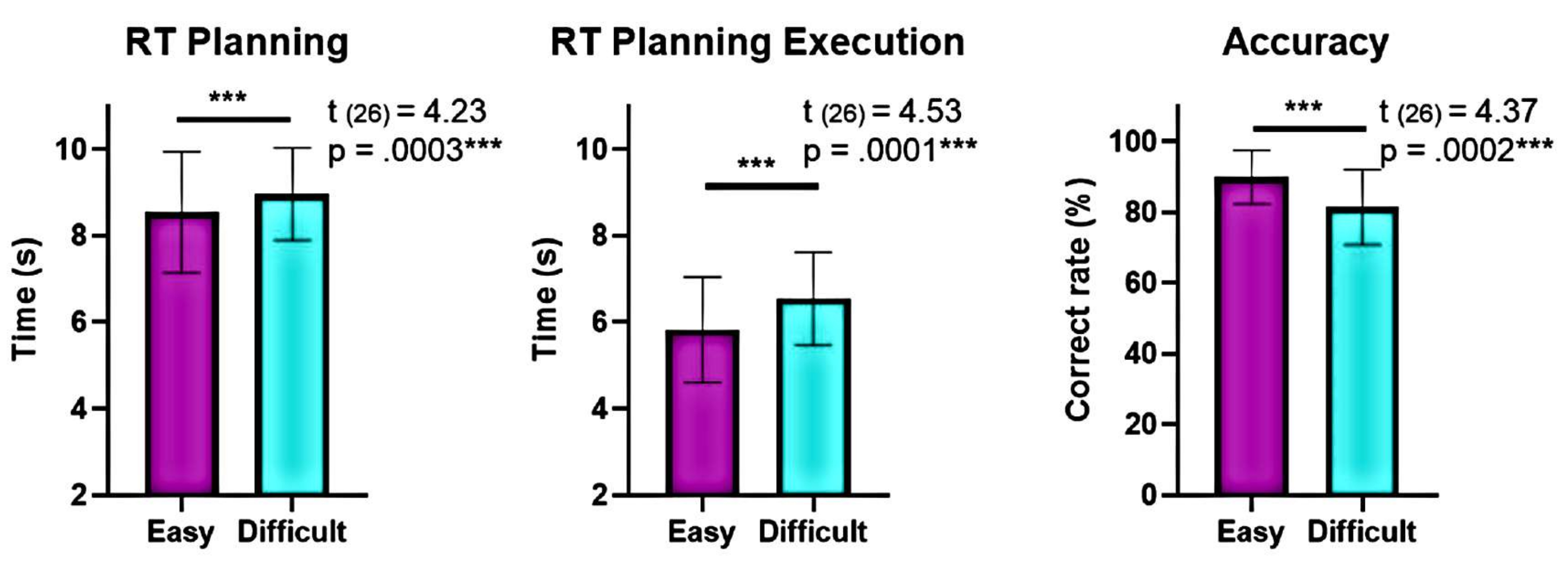
Ayrıca, planlama karmaşıklık seviyelerinin analizi, planlama ve yürütme sırasında "zor" ve "kolay" seviyeler arasında doğruluk ve tepki sürelerinde (RT'ler) önemli farklılıklar göstermiştir (Şekil 6). "Zor" seviye daha uzun RT'lere ve daha düşük doğruluğa sahipti. Bu bulgular, denemelerin geçerli çözümlerin sayısına göre bölünmesinin "kolay" denemeleri "zor" denemelerden ayırt edebileceğini göstermektedir.

Şekil 6: Farklı karmaşıklık seviyelerinde davranışsal performansın karşılaştırılması. "Kolay" ve "zor" karmaşıklık seviyelerinde davranışsal performanstaki anlamlı farklılıklar, eşleştirilmiş çift t-testi kullanılarak tanımlandı. "Kolay" seviye için planlama ve yürütme sırasında "zor" seviyeye kıyasla daha düşük reaksiyon süreleri (RT'ler) görüldü ve "kolay" seviye için doğruluk daha yüksekti. Hata çubukları SEM'i (ortalamanın standart hatası) temsil eder. Bu rakam Domic-Siede et al.54'ten değiştirilmiştir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
Bu sonuçlar, planlama bileşeni kontrol koşulundan başarılı bir şekilde çıkarıldığında (komut manipülasyonu yoluyla ), planlama görevinin bilişsel olarak daha karmaşık, talepkar, zorlayıcı ve zaman alıcı olduğunu göstermiştir. Bu nedenle, görevlerin neden olduğu sinirsel korelasyonlar birbiriyle karşılaştırılabilir.
Planlama sırasında frontal orta hat teta aktivitesini analiz etmek için, Fz elektrodu için planlama sırasındaki ortalama teta frekans bandı, kontrol periyodununkiyle karşılaştırıldı ve planlama sırasında teta bandı frekansında önemli bir artış bulundu (Şekil 7).

Şekil 7: Bilişsel planlama sırasında frontal orta hat teta aktivitesi. (A) Planlama görevi (solda), kontrol görevi (ortada) ve planlama etkisi (sağda) sırasında z-skorlarına normalize edilen tüm konularda teta bandı gücünü temsil eden topografik haritalar. Bilişsel planlama sırasında, denekler frontal orta hat teta aktivitesinde bir artış sergiledi. Renk çubuğu, -0,5 ile 1,5 arasındaki z değerlerini gösterir. (B) Fz (sol), Pz (orta) ve Oz (sağ) elektrotları için kontrol periyoduna (yeşil) kıyasla planlama sırasında denekler arasında teta aktivitesinin minimum, çeyrek, medyan ve maksimum z-skoru değerlerini gösteren bir keman grafiği eşleştirilmiş çift t-testi kullanılarak. Bu rakam Domic-Siede et al.54'ten değiştirilmiştir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
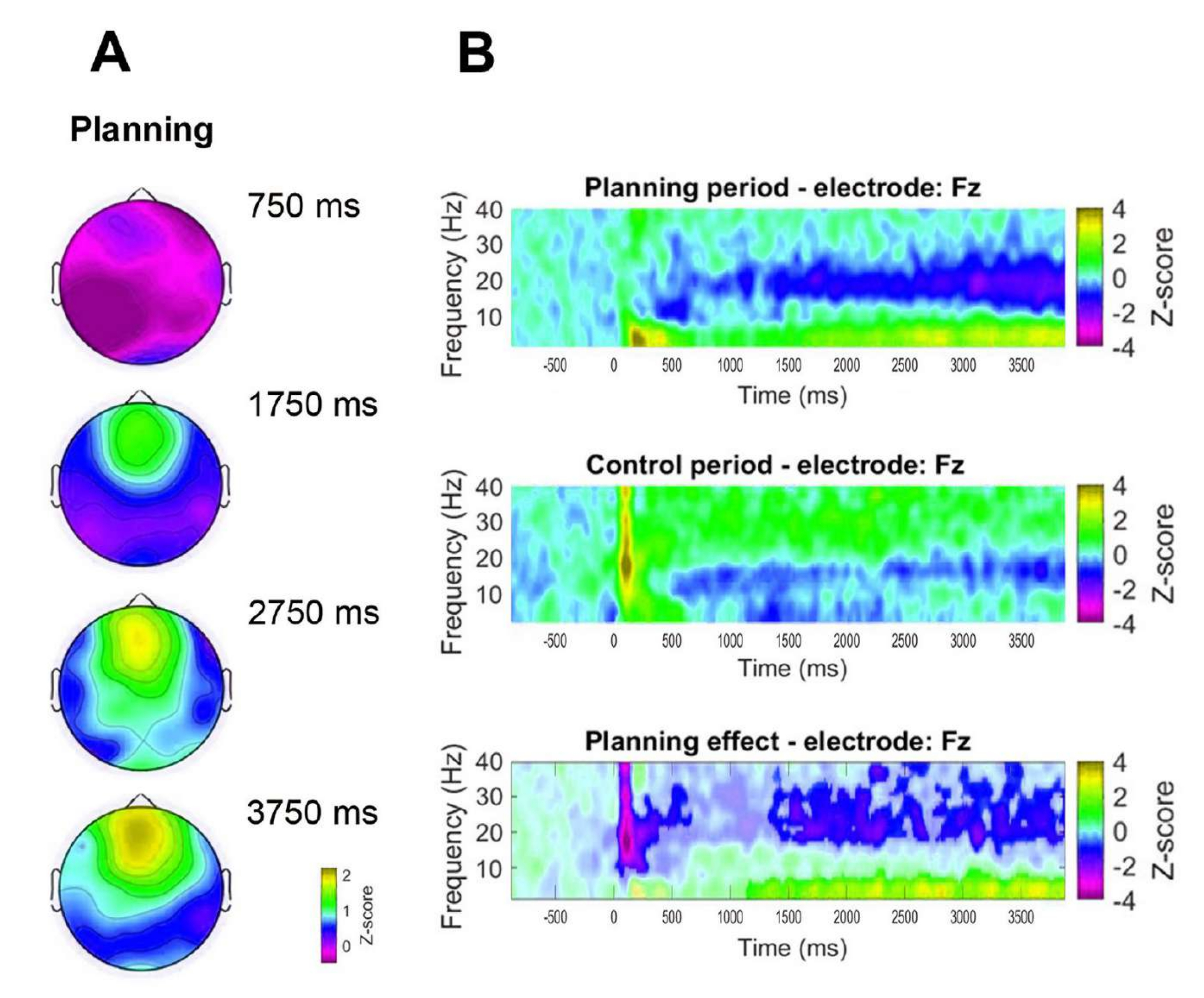
Ek olarak, gözlemlenen frontal teta aktivitesinin zamansal dinamiklerini değerlendirmek için, teta bandı gücünün belirli zaman noktalarına (750 ms, 1,750 ms, 2,750 ms ve 3,750 ms) karşılık gelen topografik haritalar formüle edilmiştir (Şekil 8A). Ayrıca, kontrol periyodu ile karşılaştırıldığında, zaman-frekans analizi, planlama periyodunun başlamasından 1 saniye sonra başlayan teta aktivitesinde önemli, aşamalı ve sürekli bir artış göstermiştir (Şekil 8B).

Şekil 8: Frontal orta hat teta zamansal dinamiği. (A) Teta aktivitesinin topografik zaman dilimleri. Planlama uygulaması (planlama dönemi) sırasında frontal orta hat teta aktivitesinde zaman içinde ilerleyici bir artış gözlenmiştir. Renk çubuğu, z-skoru birimlerini (-0,5 ila 2,2) gösterir. (B) Planlama dönemi (üstte), kontrol döneminde (ortada) ve planlama etkisi için zaman-frekans çizelgeleri, kontrol döneminin planlama döneminden çıkarılmasıyla hesaplanır (altta). Eşleştirilmiş örnekler için parametrik olmayan küme tabanlı bir permütasyon testi kullanılarak belirlendiği üzere anlamlı olmayan pikseller, planlama etkisi grafiğinde daha açık gösterilir. Renk çubuğu, z-skoru birimlerini (-4 ila 4) gösterir. Bu rakam Domic-Siede et al.54'ten değiştirilmiştir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
Teta aktivitesinin kaynak rekonstrüksiyonu için, bir beyin modeli şablonu görselleştirildi ve koşullar arasında karşılaştırıldı ve bu, teta aktivitesinin prefrontal korteks alanlarından (frontal superior korteks, FS; anterior singulat korteks, ACC; ve orta singulat korteks, MCC) kaynaklandığını ve koşullar arasında önemli farklılıklar olduğunu gösterdi (bilateral SF'de, bilateral ACC ve bilateral MCC) (Şekil 9), planlama döneminde daha yüksek teta aktivitesi gözlenmiştir (Şekil 9).

Şekil 9: Kaynak rekonstrüksiyonu. Farklı beyin kaynaklarından teta aktivitesini tahmin etmek için bir sLORETA algoritması kullanıldı. teta aktivitesi 4-8 Hz bant geçirerek filtrelendi, z-skoru normalize edildi, taban çizgisine göre düzeltildi, planlama veya kontrol başlangıcından sonra sırasıyla 1 s veya 4 s arasında ortalama alındı ve koşullar arasında karşılaştırıldı. Bilateral frontal superior bölgede, bilateral anterior singulat kortekste ve bilateral orta singulat kortekste teta aktivitesinde önemli bir artış bulundu. Şekil, permütasyon testinden elde edilen önemli t değerlerini göstermektedir. Kısaltmalar: FS = frontal superior; ACC = anterior singulat korteks; MCC = orta singulat korteks. Bu rakam Domic-Siede et al.54'ten değiştirilmiştir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
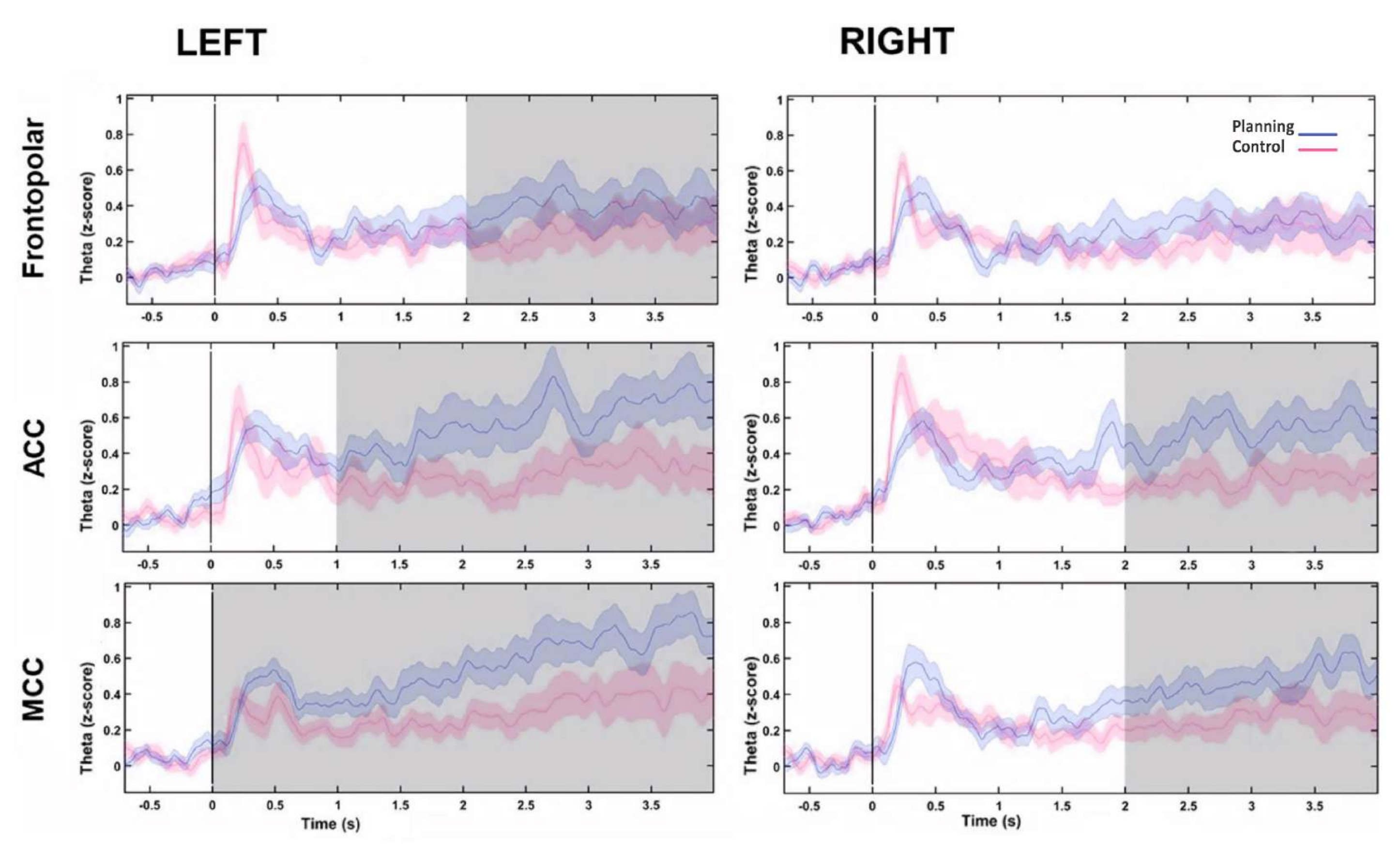
Daha sonra, her bir kaynak için teta değişikliklerinin zaman profili, bir Hilbert dönüşümü hesaplanarak değerlendirildi ve daha sonra koşullar arasındaki teta aktivitesinin anlık genliğini karşılaştırdık. Sol frontopolar, bilateral AKK ve bilateral MCC kaynaklarının planlama dönemi başlangıcından sonra daha yüksek teta aktivitesi gösterdiğini bulduk (Şekil 10). Bu sonuçlar, bilişsel planlama gerektiren deneysel paradigmamızın, PFC bölgelerinden kaynaklanan teta aktivitesini indüklediğini göstermiştir.

Şekil 10: PFC kaynaklarının teta aktivite zaman profili. Hilbert dönüşümü ile hesaplanan anlık genlik, her bir frontal kaynak için PCA ayrışmasının ilk bileşenine uygulandı ve zaman içindeki frontal teta aktivitesini göstermek için hem koşullar hem de taban çizgisi z-skoruna normalleştirildi. Gri gölgeli alanlar, FDR tarafından düzeltilen 1 s'lik adımlarla (Wilcoxon işaretli sıra testi) örtüşmeyen hareketli bir pencere kullanılarak belirlenen önemli farklılıkları gösterir. Gölgeli bölgeler %95 güven aralıklarını temsil eder. Sol FP bölgesi, bilateral AKK ve bilateral MKK, planlama başlangıcından sonra teta aktivitesinde artış gösterdi. Kısaltmalar: ACC = anterior singulat korteks; MCC = orta singulat korteks. Planlama koşulu mor renkle gösterilir. Kontrol koşulu kırmızı ile gösterilmiştir. Bu rakam Domic-Siede et al.54'ten değiştirilmiştir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
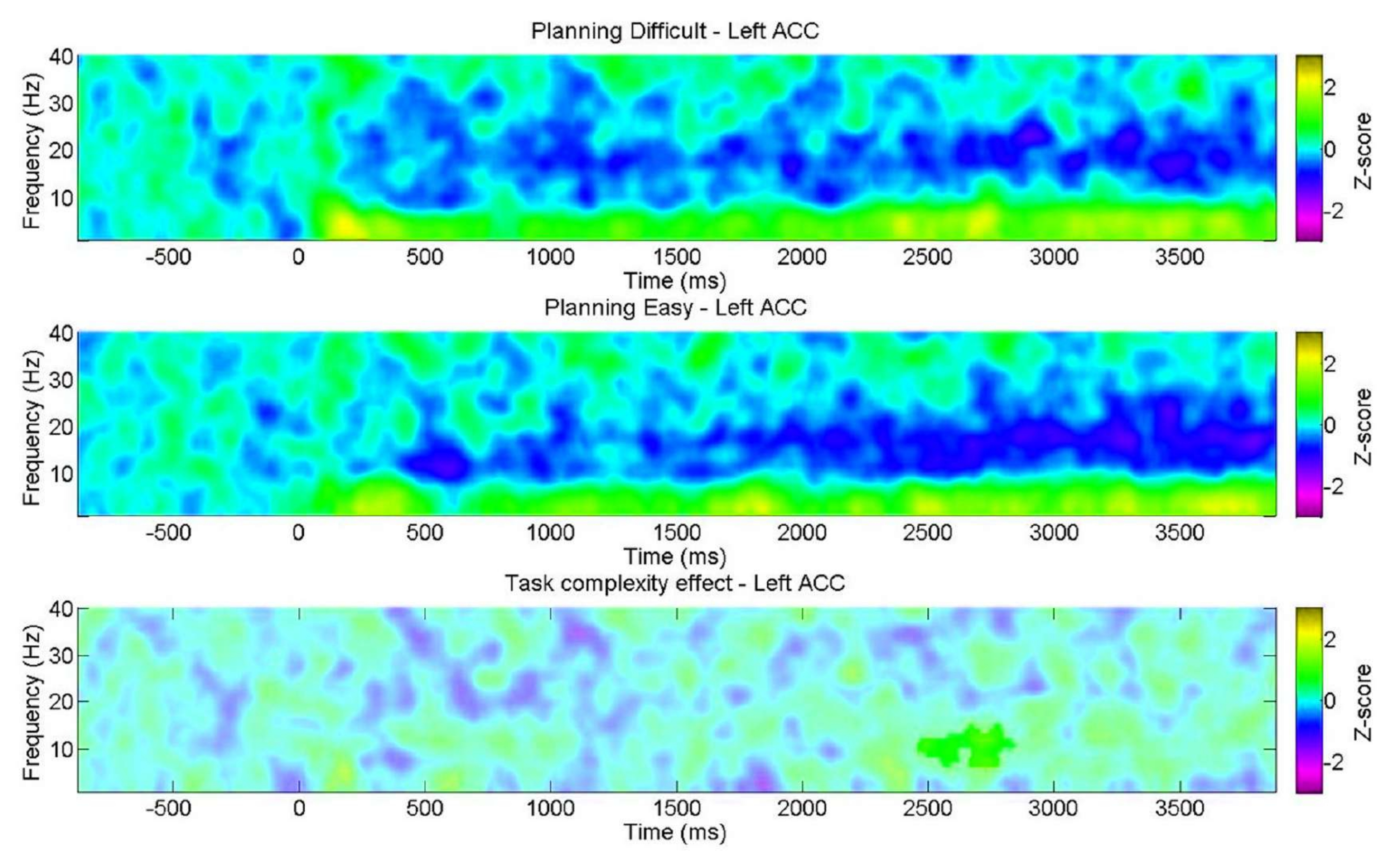
Ayrıca, davranışsal sonuçların gösterdiği gibi, karmaşıklık seviyesine göre planlama sırasında spektral özelliklerdeki değişimi incelemeyi amaçladık. Dikkat çekici bir şekilde, alfa bandı içinde sadece sol ACC'de önemli bir tutarsızlık bulundu. Bu, planlama görevimizin, teta salınımlarındaki değişiklikler yoluyla planlamanın içsel yönlerini, bilişsel kontrol görevlerinde tipik olarak karşılaşılan genel bilişsel taleplerden (çaba) daha büyük ölçüde değerlendirdiği fikrini desteklemektedir (Şekil 11).

Şekil 11: Planlama karmaşıklık seviyelerinde EEG. ROI zaman-frekans çizelgeleri, alfa bandında "zor" seviye için yalnızca sol ön singulat kortekste (ACC) önemli bir pozitif küme gösterdi. Eşleştirilmiş örnekler için parametrik olmayan küme tabanlı bir permütasyon testi kullanılarak belirlendiği üzere anlamlı olmayan pikseller, renk çubuğu -3 ile 3 arasındaki z-skor birimlerini gösterecek şekilde çizim üzerinde daha açık bir gölgede gösterilir. Bu rakam Domic-Siede et al.54'ten değiştirilmiştir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
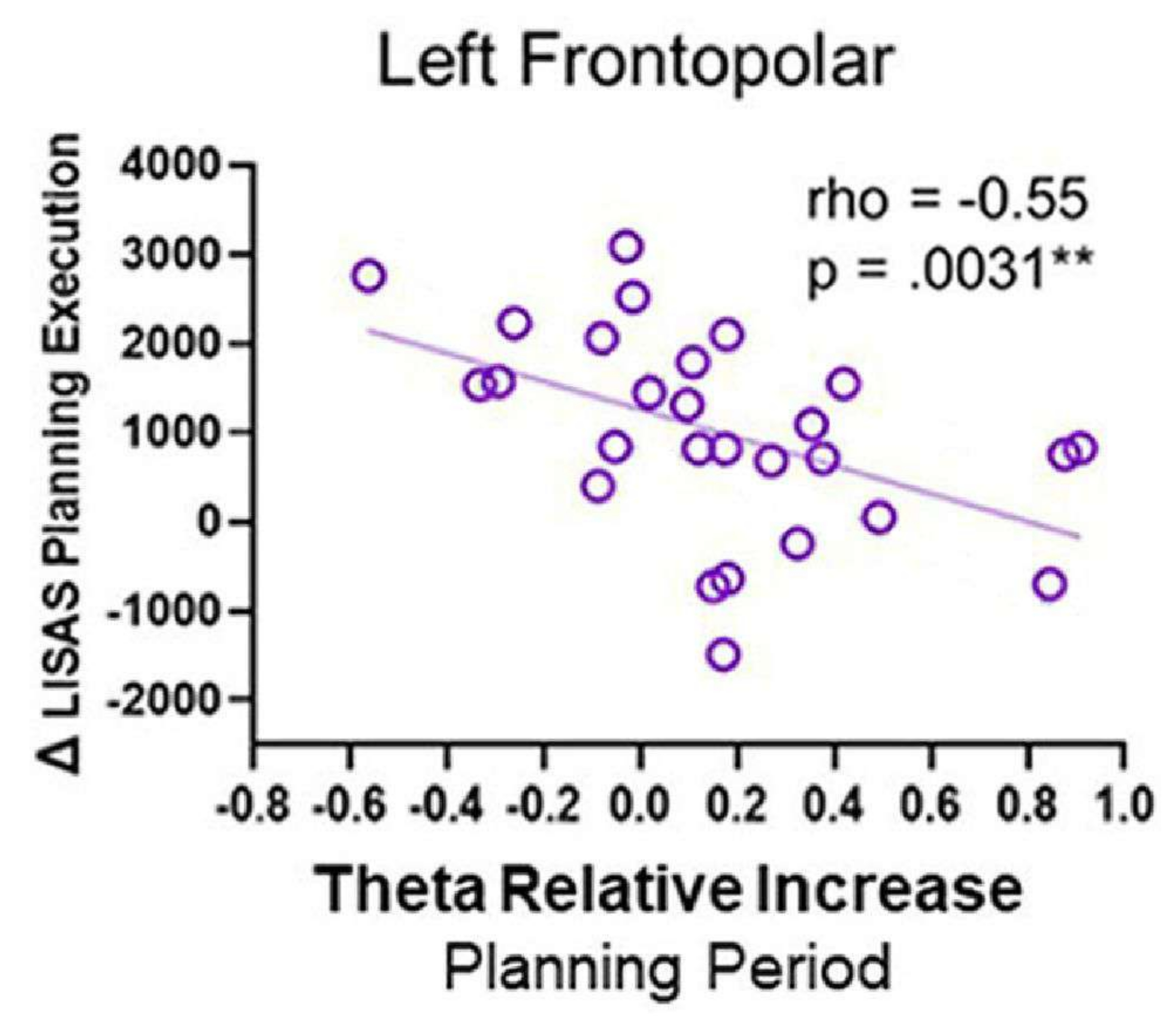
teta aktivitesi ile davranışsal performans arasındaki korelasyonlar gerçekleştirildiğinde, negatif bir korelasyon gözlendi; özellikle, planlama periyodu boyunca sol frontopolar bölgedeki teta aktivitesi arttıkça, LISAS planlama yürütme puanı azalmıştır (Şekil 12). Bu model, daha sonra bir planı başarılı bir şekilde yürütmek için planlama detaylandırması sırasında sol FP bölgesinin gerekli olabileceğini yansıtabilir ve teta aktivitesi için bir rol önerir.

Şekil 12: Teta aktivitesi ve davranışsal performans. Sol frontopolar korteksten gelen teta aktivitesi ile Δ LISAS planlama uygulaması arasındaki Spearman rho korelasyonu anlamlı bir negatif korelasyon gösterdi. Bu rakam Domic-Siede et al.54'ten değiştirilmiştir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
Ek olarak, her bir koşulun neden olduğu değişen bilişsel talepler ve hedefler, planlama ve kontrol koşulları arasında zıt göz hareketlerine neden olmuş ve farklı salınımlı aktivite modellerine yol açmış olabilir95. Yukarıdaki sorunu ele almak için, tek denekli, tek deneme verilerini çeşitli düzeylerde analiz ettik. Özellikle, Fz kanalı zaman serisi ve teta aktivitesi zaman dinamiklerinin zaman içindeki sakkad oranı ile hiçbir bağlantısı olmadığı görülmüştür (Şekil 13A).

Şekil 13: EEG sonuçları ve göz hareketleri kayıtları. (A) Sıralar, EEG'yi (üstte), zaman-frekans tablosunu (ortada üstte), elektrot Fz'den teta zaman profilini (orta altta) ve denek 8 ve deneme 9'un (altta) planlama koşulundaki sakkad oranını sunar. (B) Koşullar arasındaki sakkad genliği, sakkad tepe hızı ve güç-sakkad oranı tutarlılığının Wilcoxon imzalı sıra testi karşılaştırması gösterilir ve planlama ve kontrol koşulları arasındaki sakkad genliğinde önemli istatistiksel farklılıklar ortaya çıkar. SEM, hata çubuklarıyla temsil edilir. Bu rakam Domic-Siede et al.54'ten değiştirilmiştir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
Daha sonra, tüm denemeden ve karşılaştırma için 0 sn ila 3.75 s arasında sakkad genliğini ve tepe hızını elde ettik (Şekil 13B). Kontrol koşulunda sakkad genliğinin daha büyük olduğunu keşfettik. Bununla birlikte, elektrot Fz'deki Fourier teta gücü ile sakkad hızı arasındaki tutarlılık indeksindeki koşullar arasında istatistiksel olarak anlamlı bir fark bulunmamıştır (Şekil 10B), bu da sakkadlar ve teta aktivitesi arasındaki herhangi bir potansiyel ilişkinin koşullar arasında tutarlı olduğunu göstermektedir.
Birlikte ele alındığında, bu sonuçlar, açıklanan deneysel protokolün bilişsel planlamayı bir bilişsel kontrol işlevi olarak incelemek için uygun olduğunu desteklemektedir.
Tartışmalar
Burada açıklanan protokol, salınım ve pupiller dinamikler gibi ilgili ve tamamlayıcı davranışsal ve fizyolojik ölçümlerle bağlantılı olarak yeni ve ekolojik bir planlama görevi sırasında bilişsel planlama ve bilişsel kontrolü değerlendirmek için yenilikçi bir yaklaşım sunmaktadır. Deneylerimiz sırasında, katılımcılar planlama görevini yerine getirirken EEG aktivitesi kaydedildi, bu görevde katılımcılara önce bir plan hazırlamaları ve sonra yürütmeleri talimatı verildi. Hayvanat bahçesi haritasında önceden çizilmiş bir yolun değerlendirilmesini içeren kontrol koşulu, benzer bir ortam ve yapıyı korurken bilişsel planlama yönünü ortadan kaldırmak için oluşturulmuştur. Bu yaklaşım, bilişsel bir kontrol işlevi olarak bilişsel planlamanın, PFC bölgelerinden frontal teta aktivitesinin üretilmesine yol açıp açmadığının ve farklı PFC teta salınım kaynaklarının planlama performansının farklı yönleriyle bağlantılı olup olmadığının değerlendirilmesini sağlar. Bu protokol kullanılarak değerlendirilebilecek bir diğer husus, plan detaylandırma, plan yürütme, plan başarısı ve geri bildirim işleme gibi planlama aşamalarında yer alan çeşitli bilişsel süreçlerin farklılaşmasıdır. Planlamanın, bilişsel kontrol ile ilişkili kanonik bir frontal teta aktivitesine neden olduğunu ve bunun da bir hedefe verimli bir şekilde ulaşılmasına katkıda bulunduğunu bulduk. Bu sonuçlar, bu protokolün ilgisini desteklemektedir.
Bilişsel sinirbilimdeki büyük gelişmelere rağmen, çoğu nörogörüntüleme deneyi, duyu-motordan yoksun ortamlarda yapay görevler ve karışıklıkları kontrol etmek için aşırı basitleştirilmiş uyaranlar kullanarak izole bilişsel işlevleri inceler. Sorunlu bir şekilde, bu deneyler, günlük bir durumda (ekolojik durumlarda) bilişsel bir işlev uygulandığında ortaya çıkan gerçek beyin mekanizmalarını tanımlayamayabilir138,140. Özellikle, hedefleri formüle etme, planları etkili bir şekilde planlama ve yürütme kapasitelerinin, çeşitli bilişsel işlevler (çalışma belleği, engelleyici kontrol, bilişsel esneklik vb.) gerektirdiğinden değerlendirilmesi zordur.104.144. Bu nedenle, bilişsel sinirbilimdeki mevcut eğilimlere dayalı olarak ekolojik davranışsal bir görev tasarlamak teşvik edilmekte ve önerilmektedir 142,143,145,146.
Planlama görevimiz, bir laboratuvar ortamında (uyaranların bir ekranda görüntülendiği bir odanın içinde) gerçekleşmesine rağmen, ekranda etkileşime girebilecekleri anlamlı uyaranlar ve hedefler kullanılarak katılımcılar için ilgi çekici ve etkileşimli hale getirildi. Ek olarak, görev, katılımcıların çeşitli yerleri ziyaret etmek için bir yol planlayan gerçek hayattaki bir duruma katılmalarını gerektirir. Ekolojik bir görev tasarımına sahip olmak için, paradigma, denekleri günlük yaşamda yapmak zorunda kalacaklarına benzer bir şekilde belirli bir davranışı veya bilişsel işlevi yerine getirmeye zorlamalıdır 62,63,147. Ekolojik bir görev tasarımı geliştirmek için, burada kullanılan planlama görevi, birkaç aşamada farklı yerleri ziyaret etmek için bir yol planlamayı içerir56. İlk aşama, katılımcıların bir dizi kurala uymasını sağlarken bir plan oluşturmasını içerir. İkinci aşama, planın çalışma belleğinde tutulmasını içerirken, üçüncü aşama planın yürütülmesini ve kurallara bağlılığının izlenmesini içerir. Bu aşamalar, planlamanın farklı aşamalarını ve bilişsel esneklik, engelleyici kontrol ve çalışma belleği gibi diğer yürütücü işlevlerin orkestrasyonunu temsil eder. Geçerli bir ekolojik bilişsel görev tasarımına sahip olmak için, görevin günlük yaşamlarında bu belirli bilişsel işlevi yerine getirmekte zorluk çeken psikiyatrik veya bilişsel bozukluğu olan hastalarda belirli bilişsel bozuklukları tespit edebilmesi gerekir105. Bu, bu protokolü kullanarak gelecekteki araştırmalarla başarılabilir.
Bu protokolün kullanılmasıyla elde edilen davranışsal sonuçlar, deneysel tahminlerle uyumlu hale getirildi. Planlama bileşeni bir kontrol koşulu oluşturmak için kontrol görevinden çıkarıldığında davranışta önemli bir fark gözlendi, böylece daha fazla karşılaştırma kolaylaştırıldı. Planlama koşulunun, reaksiyon süresi ve doğruluk gibi parametrelerle kanıtlandığı gibi, bilişsel olarak kontrol koşulundan daha zorlu olduğu bulundu. Bu, 23,55,56,57,148,149,150 planlamasının uygulanmasında yüksek bilişsel işlevlerin artan katılımını yansıtabilir.
Daha hızlı tepki süreleri, daha iyi performanslar ve gereken farklı bilişsel süreçler (kuralların değerlendirilmesi) ile kanıtlandığı gibi, kontrol koşulunun daha az karmaşık biliş içerdiği göz önüne alındığında, olası bir alternatif değişiklik, planlama görevinde mevcut olan karmaşıklık seviyelerinden yararlanmak, bunları manipüle etmek ve planlama fonksiyonunu farklı karmaşıklık seviyelerine göre parametrik olarak analiz etmektir (örneğin, deneme sayısının artırılması ve zor, orta ve kolay deneme koşullarının oluşturulması). Bununla birlikte, protokolümüzden elde edilen sonuçlar, davranışsal ölçümlere dayalı olarak "kolay" ve "zor" denemeler arasında ayrım yapmanın mümkün olmasına rağmen, elektrofizyolojik ölçümlerde herhangi bir fark tespit edilmediğini göstermiştir. Bu, protokolümüzdeki sonuçların, dikkat, zihinsel çaba, zorluk düzeyi veya yüksek düzeyde bilişsel talep gibi bilişsel kontrolün daha geniş yönlerinden ziyade, planlama işlevinin içsel özelliklerini daha doğru bir şekilde yansıttığını göstermektedir54. Bununla birlikte, daha fazla araştırma, dört hayvan bölgesini ziyaret eden işaretli bir yolu takip etmek ve aynı zamanda dizi sırasını hatırlamak gibi diğer kontrol koşullarını da dikkate alabilir. Bu şekilde, zorluk seviyesi daha iyi kontrol edilebilir ve planlama, çalışma belleğinden ayırt edilebilir, ancak bunun olası bir dezavantajı yorgunluktur, çünkü denekler iki zorlu görevi yerine getirmek zorunda kalacaklardır.
Birkaç çalışma, farklı göz hareketi parametrelerini belirli bilişsel olaylarla ilişkilendirmiştir. Bir yandan, bazı çalışmalar bilişsel görevler sırasında teta salınımları ile göz bebeği çapı arasında korelasyonlar bulmuş ve bu iki bilişsel işlev ölçüsü arasında bir ilişki olduğunu düşündürmektedir. Örneğin, Lin ve ark.152 , orta frontal teta aktivitesi ile öğrenci boyutundaki değişiklikler arasında, değişen derecelerde öznel çatışmayı yansıtan bir ilişki buldu. Bulguları, bu sinyallerin çatışma işlemeyi, artan dikkati ve esnek davranışsal tepkileri temsil ettiğini göstermektedir. Bu nedenle, midfrontal teta aktivitesi ile öğrenci tepkileri arasındaki ilişki, karar verme süreçlerinde maliyet ve faydaların tartılmasında rol oynuyor gibi görünmektedir. Başka bir çalışmada, Yu ve ark.153 , kontrol inhibitör tepkilerini modüle eden bir çalışma belleği görevi aracılığıyla görev başında zaman katılımının nörofizyolojik olarak bilişsel kontrolü nasıl etkilediğini inceledi. Öğrenci çapı verileri ile frontal teta aktivitesi arasındaki ilişkiyi incelediler ve görev süresi arttıkça performansın düştüğünü ve buna pupil genişleme modülasyonunda ve frontal teta aktivitesinde bir azalma eşlik ettiğini gösterdiler. Görevin başlangıcında, esas olarak yüksek çalışma belleği ve engelleyici kontrol gerektiren zorlu görevler için, öğrenci genişleme modülasyonu ile gösterildiği gibi, görev katılımı, teta aktivitesi ve bilişsel kontrol arasında güçlü bir ilişki buldular. Bununla birlikte, bu ilişki sonlara doğru dağıldı ve harcanan çaba ile görevi yerine getirmek için kullanılan bilişsel kontrol arasında bir kopukluğa işaret etti, bu da prefrontal görev süresi etkilerinin bir işaretidir153. Öte yandan, diğer çalışmalar sakkadları ve salınımları araştırmıştır. Örneğin, Nakatani ve ark.154 , algısal bir görevde, oksipital bölgelerden gelen alfa bandı genliğinin göz kırpmaları ve sakkad etkilerini öngördüğünü ortaya koydu. Ayrıca, Velasques ve ark.155 , prosakkadik bir dikkat görevi sırasında sakkad genliğinin frontal gama değişiklikleri ile ilişkili olduğunu göstermiştir. Ayrıca, Bodala ve ark.156 , frontal orta hat tetasındaki azalmalara, sürekli dikkatin yanı sıra sakkadların genliği ve hızındaki azalmaların eşlik ettiğini bulmuşlardır. Bu bulgular, göz hareketlerinin, özellikle sakkadların, EEG sinyallerindeki arka plan gürültüsüne katkıda bulunmak yerine bilişsel süreçleri yansıtabileceğini düşündürmektedir. Bu çalışmada, sakkadat-fiksasyon varyans oranı kriteri126 ile ICA algoritmasını kullanarak göz hareketleriyle ilişkili artefaktların eliminasyonunu geliştirdik. Bu kriter, ücretsiz görüntüleme görevleri için artefakt kaldırmayı geliştirir157. Çalışmamızda, koşullar arasında sakkad tepe hızı ve teta gücü ile sakkad hızı arasındaki tutarlılıkta herhangi bir fark gözlenmemiştir. Ancak, bu soruları ele almak için daha fazla çalışmaya ihtiyaç vardır.
Bu protokolü kullanmanın kritik bir adımı, deney sırasında göz izleyiciyi sürekli olarak kalibre etmektir, çünkü kameradan gelen bakış verilerinin kaybı, görevi kirletecek ve doğru yanıtlar almayı zorlaştıracak hatalara neden olabilir. Bu nedenle, mümkün olduğunca sık kalibre etmek çok önemlidir. Bununla birlikte, kalibre edilmiş bir göz izleyici ile yapılan denemelerin sayısı ile deneyin uzunluğu arasında bir değiş tokuş vardır. Çalışmamızda, her beş denemede bir kalibrasyon yapmaya karar verdik.
Bu planlama görevi sırasında teta salınımları ile göz bebeği çapı arasındaki ilişkiyi araştıran gelecekteki araştırmalar yapılmalıdır. Planlama, dikkat kaynaklarının tahsisini ve çoklu bilişsel süreçlerin koordinasyonunu gerektiren yönetici kontrolünün kritik bir yönüdür. Planlama görevleri sırasında teta salınımları ile göz bebeği çapı arasındaki ilişkiyi anlamak, yürütücü kontrolün altında yatan nöral mekanizmalar ve bunların zaman içinde nasıl değiştiği hakkında değerli bilgiler sağlayabilir. Ayrıca, bu tür çalışmalar, yorgunluk veya dikkat kaybı gibi bilişsel işlevlerdeki değişikliklerin, planlama görevlerindeki performansı ve kaynakları etkili bir şekilde tahsis etme yeteneğini nasıl etkilediğinin daha derin bir şekilde anlaşılmasına yol açabilir. Bu bilgiler, diğerlerinin yanı sıra bilişsel eğitim programları veya dikkat eksikliği / hiperaktivite bozukluğu (DEHB) gibi durumlar için tedaviler gibi planlama performansını iyileştirmeyi amaçlayan müdahaleler geliştirmek için önemli etkilere sahip olabilir.
Önceki araştırmalar, sonuçlarımızla da onaylandığı gibi, PFC'nin bilişsel planlamada çok önemli bir rol oynadığını göstermiştir. Bu sonuçlar, bilişsel planlamanın PFC'de, özellikle ön singulat kortekste, orta singulat kortekste ve üst frontal bölgelerde FMθ aktivitesini indüklediğini göstermektedir54. Bu bulgular, yürütücü işlevlerle ilgili önceki araştırmalarla uyumludur. FMθ aktivitesinin, zorlu görevler sırasında kontrolü başlatmak ve beyin bölgeleri arasındaki iletişimi kolaylaştırmak için yukarıdan aşağıya bir süreç olarak hareket ettiği fikrini destekleyen önemli kanıtlar vardır13. Bilişsel kontrol ile ilişkili FMθ aktivitesinin zamansal dinamiklerini sadece birkaç çalışma incelemiş olsa da, FMθ'nin zaman profilinin bilişsel kontrolün çeşitli yönleri ve farklı PFC bölgelerinin katılımı hakkında bilgi sağlayabileceği konusunda yaygın bir fikir birliği vardır. Bilişsel planlamayı değerlendirmek için protokolümüzü kullanmak, planlama sırasında FMθ aktivitesinin zaman profilini karakterize etmemizi sağladı. Spesifik olarak, planlama koşulu sırasındaki FMθ aktivitesi aşamalı bir artış gösterdi. Bu protokolün uygulanmasıyla, ilk kez, FMθ'nin, diğer üst düzey bilişsel işlevlerde olduğu gibi, planlama uygulaması sırasında da mevcut olduğu ve zamansal dinamiklerinin bilişsel kontrolün bir göstergesi olarak hizmet edebileceği gösterilmiştir.
Sonuçlarımız ve protokolümüz, sanal nöropsikolojik değerlendirmelerin iyileştirilmesi ve depresyon ve dikkat eksikliği / hiperaktivite bozukluğu gibi ilişkili bilişsel planlama sorunları ile nöropsikiyatrik bozuklukların tedavisi de dahil olmak üzere sinirbilim alanında potansiyel uygulamalara sahiptir. Örneğin, değerlendirmeler, davranışsal performans düzeyinde farklı hata modellerini, elektrofizyolojik düzeyde farklı salınım modellerini ve farklı göz hareketlerini incelemeyi içerebilir. Ek olarak, bu çalışmanın sonuçları, bilişsel planlama yeteneklerini geliştirmeyi amaçlayan beyin-bilgisayar arayüzlerinin ve bilişsel eğitim programlarının geliştirilmesini bilgilendirebilir.
Mevcut protokol, nörotipik ve nöropsikiyatrik popülasyonlarda bilişsel planlamanın anlaşılması zor bilişsel kontrol işlevinin temelini oluşturan sinirsel mekanizmaların anlaşılmasına yeni kanıtlar sağlayabilir. Dahası, davranışsal paradigmamız, elektrofizyolojik, pupillometri ve davranışsal ölçümlerin incelenmesi yoluyla, teta salınımlarındaki değişikliklerde yansıtıldığı gibi, bilişsel kontrol görevlerinde tipik olarak mevcut olan genel bilişsel taleplerden ziyade planlamanın içsel yönlerini inceleyen pratik bir planlama görevi ile bilişsel kontrol ve planlamanın nörobiyolojisi hakkında içgörüler sunabilir.
Açıklamalar
Yazarların ifşa edecek hiçbir şeyi yok.
Teşekkürler
Bu araştırma, ANID 21150295, FONDECYT düzenli hibe 1180932, FONDECYT düzenli hibe 1230383, FONDECYT de Iniciación hibe 11220009, Postdoc hibe Universidad de O'Higgins ve Institut Universitaire de France (IUF) doktora burs programı Becas de Doctorado Nacional año 2015 tarafından finansal olarak desteklenmiştir. Paradigma tasarımı hakkındaki geri bildirimleri için Profesör Pablo Billeke'ye teşekkür etmek istiyoruz. Profesör Eugenio Rodríguez'e zaman-frekans analiz kodlarını paylaşma nezaketini gösterdiği için teşekkür ederiz. Son olarak, geri bildirimleri için Milan Domic, Vicente Medel, Josefina Ihnen, Andrea Sánchez, Gonzalo Boncompte, Catalina Fabar ve Daniela Santander'e teşekkür ederiz.
Malzemeler
| Name | Company | Catalog Number | Comments |
| EEG System | Biosemi | ActiveTwo Base system, 64 channels | |
| Eye-tracker System | Eyelink SR Research | EyeLink 1000 Plus Core Unit, High-speed camera, Host PC/Monitor, | |
| CPU display | Intel | Hard drive 221 GB, Processor Intel Core i7-4790 3.60 Hz, OS Windows 7, 4GB RAM | |
| CPU EEG | Intel | Hard drive 223 GB, Processor Intel Core i7-4790 3.60 Hz, OS Windows 7, 4GB RAM | |
| Monitor | ASUS | ASUS VG248QE 24" LCD monitor | |
| Joytsick | Logitech | Model F310 | |
| Luxmeter | Focket | LCD screen (0-200.000 lux) model Liebe WH LX1330B | |
| Statistics software | GraphPad Prism | GraphPad Prism version 8 for Windows | |
| MATLAB Programming Software | The MathWorks | MATLAB R2014a and R2018b | |
| SVG tool Inkscape | Inkscape Project | vector graphic editor software | |
| Presentation Software | Neurobehavioral Systems | stimulus delivery and experiment control program for neuroscience |
Referanslar
- Siegel, M., Donner, T. H., Engel, A. K. Spectral fingerprints of large-scale neuronal interactions. Nature Reviews Neuroscience. 13 (2), 121-134 (2012).
- Fries, P. A mechanism for cognitive dynamics: Neuronal communication through neuronal coherence. Trends in Cognitive Sciences. 9 (10), 474-480 (2005).
- Fries, P. Rhythms for cognition: Communication through coherence. Neuron. 88 (1), 220-235 (2015).
- Thut, G., Miniussi, C., Gross, J. The functional importance of rhythmic activity in the brain. Current Biology. 22 (16), 658-663 (2012).
- Fröhlich, F., McCormick, D. A. Endogenous electric fields may guide neocortical network activity. Neuron. 67 (1), 129-143 (2010).
- Spaak, E., de Lange, F. P., Jensen, O. Local entrainment of alpha oscillations by visual stimuli causes cyclic modulation of perception. Journal of Neuroscience. 34 (10), 3536-3544 (2014).
- Fiebelkorn, I. C., Saalmann, Y. B., Kastner, S. Rhythmic sampling within and between objects despite sustained attention at a cued location. Current Biology. 23 (24), 2553-2558 (2013).
- Landau, A. N., Fries, P. Attention samples stimuli rhythmically. Current Biology. 22 (11), 1000-1004 (2012).
- Song, K., Meng, M., Lin, C., Zhou, K., Luo, H. Behavioral oscillations in attention: Rhythmic α pulses mediated through θ band. Journal of Neuroscience. 34 (14), 4837-4844 (2014).
- Wyart, V., Nobre, A. C., Summerfield, C. Dissociable prior influences of signal probability and relevance on visual contrast sensitivity. Proceedings of the National Academy of Sciences of the United States of America. 109 (9), 3593-3598 (2012).
- Leszczyński, M., Fell, J., Axmacher, N. Rhythmic working memory activation in the human hippocampus. Cell Reports. 13 (6), 1272-1282 (2015).
- Onton, J., Delorme, A., Makeig, S. Frontal midline EEG dynamics during working memory. NeuroImage. 27 (2), 341-356 (2005).
- Cavanagh, J. F., Frank, M. J. Frontal theta as a mechanism for cognitive control. Trends in Cognitive Sciences. 18 (8), 414-421 (2014).
- Siegel, M., Buschman, T. J., Miller, E. K. Cortical information flow during flexible sensorimotor decisions. Science. 348 (6241), 1352-1355 (2015).
- Weisz, N., et al. Prestimulus oscillatory power and connectivity patterns predispose conscious somatosensory perception. Proceedings of the National Academy of Sciences of the United States of America. 111 (4), 417-425 (2014).
- Buzsáki, G., Draguhn, A. Neuronal oscillations in cortical networks. Science. 304 (5679), 1926-1929 (2004).
- Cohen, M. X. A neural microcircuit for cognitive conflict detection and signaling. Trends in Neurosciences. 37 (9), 480-490 (2014).
- Yuste, R. From the neuron doctrine to neural networks. Nature Reviews Neuroscience. 16 (8), 487-497 (2015).
- Helfrich, R. F., Knight, R. T. Oscillatory dynamics of prefrontal cognitive control. Trends in Cognitive Sciences. 20 (12), 916-930 (2016).
- Miller, E. K., Cohen, J. D. An integrative theory of prefrontal cortex function. Annual Review of Neurosciences. 24 (1), 167-202 (2001).
- von Nicolai, C., et al. Corticostriatal coordination through coherent phase-amplitude coupling. Journal of Neuroscience. 34 (17), 5938-5948 (2014).
- Sweeney-Reed, C. M., et al. Thalamic theta phase alignment predicts human memory formation and anterior thalamic cross-frequency coupling. ELife. 4, 07578(2015).
- Voytek, B., et al. Oscillatory dynamics coordinating human frontal networks in support of goal maintenance. Nature Neuroscience. 18 (9), 1318-1324 (2015).
- Raghavachari, S., et al. Theta oscillations in human cortex during a working-memory task: Evidence for local generators. Journal of Neurophysiology. 95 (3), 1630-1638 (2006).
- Jacobs, J., Hwang, G., Curran, T., Kahana, M. J. EEG oscillations and recognition memory: Theta correlates of memory retrieval and decision making. NeuroImage. 32 (2), 978-987 (2006).
- Itthipuripat, S., Wessel, J. R., Aron, A. R. Frontal theta is a signature of successful working memory manipulation. Experimental Brain Research. 224 (2), 255-262 (2013).
- Cavanagh, J. F., Zambrano-Vazquez, L., Allen, J. J. B. Theta lingua franca: A common mid-frontal substrate for action monitoring processes. Psychophysiology. 49 (2), 220-238 (2012).
- Mas-Herrero, E., Marco-Pallarés, J. Frontal theta oscillatory activity is a common mechanism for the computation of unexpected outcomes and learning rate. Journal of Cognitive Neuroscience. 26 (3), 447-458 (2014).
- Folstein, J. R., Van Petten, C. Influence of cognitive control and mismatch on the N2 component of the ERP: A review. Psychophysiology. 45 (1), 152-170 (2008).
- Cohen, M. X., Donner, T. H. Midfrontal conflict-related theta-band power reflects neural oscillations that predict behavior. Journal of Neurophysiology. 110 (12), 2752-2263 (2013).
- Walsh, M. M., Anderson, J. R. Modulation of the feedback-related negativity by instruction and experience. Proceedings of the National Academy of Sciences of the United States of America. 108 (47), 19048-19053 (2011).
- Luu, P., Tucker, D. M., Makeig, S. Frontal midline theta and the error-related negativity: Neurophysiological mechanisms of action regulation. Clinical Neurophysiology. 115 (8), 1821-1835 (2004).
- Hanslmayr, S., et al. The electrophysiological dynamics of interference during the Stroop task. Journal of Cognitive Neuroscience. 20 (2), 215-225 (2008).
- Cavanagh, J. F., Cohen, M. X., Allen, J. J. B. Prelude to and resolution of an error: EEG phase synchrony reveals cognitive control dynamics during action monitoring. Journal of Neuroscience. 29 (1), 98-105 (2009).
- Cohen, M. X., van Gaal, S., Ridderinkhof, K. R., Lamme, V. A. F. Unconscious errors enhance prefrontal-occipital oscillatory synchrony. Frontiers in Human Neuroscience. 3, 54(2009).
- Cavanagh, J. F., Frank, M. J., Klein, T. J., Allen, J. J. B. Frontal theta links prediction errors to behavioral adaptation in reinforcement learning. NeuroImage. 49 (4), 3198-3209 (2010).
- Cohen, M. X., Cavanagh, J. F. Single-trial regression elucidates the role of prefrontal theta oscillations in response conflict. Frontiers in Psychology. 2, 30(2011).
- Cohen, M. X., Van Gaal, S. Dynamic interactions between large-scale brain networks predict behavioral adaptation after perceptual errors. Cerebral Cortex. 23 (5), 1061-1072 (2013).
- Nigbur, R., Cohen, M. X., Ridderinkhof, K. R., Stürmer, B. Theta dynamics reveal domain-specific control over stimulus and response conflict. Journal of Cognitive Neuroscience. 24 (5), 1264-1274 (2012).
- van Driel, J., Swart, J. C., Egner, T., Ridderinkhof, K. R., Cohen, M. X. No) time for control: Frontal theta dynamics reveal the cost of temporally guided conflict anticipation. Cognitive, Affective, and Behavioral Neuroscience. 15 (4), 787-807 (2015).
- van de Vijver, I., Ridderinkhof, K. R., Cohen, M. X. Frontal oscillatory dynamics predict feedback learning and action adjustment. Journal of Cognitive Neuroscience. 23 (12), 4106-4121 (2011).
- Narayanan, N. S., Cavanagh, J. F., Frank, M. J., Laubach, M. Common medial frontal mechanisms of adaptive control in humans and rodents. Nature Neuroscience. 16 (12), 1888-1895 (2013).
- Anguera, J. A., et al. Video game training enhances cognitive control in older adults. Nature. 501 (7465), 97-101 (2013).
- Smit, A. S., Eling, P. A. T. M., Hopman, M. T., Coenen, A. M. L. Mental and physical effort affect vigilance differently. International Journal of Psychophysiology. 57 (3), 211-217 (2005).
- Cohen, M. X., Ranganath, C. Reinforcement learning signals predict future decisions. Journal of Neuroscience. 27 (2), 371-378 (2007).
- Gehring, W. J., Goss, B., Coles, M. G. H., Meyer, D. E., Donchin, E. A neural system for error detection and compensation. Psychological Science. 4 (6), 385-390 (1993).
- Yeung, N., Botvinick, M. M., Cohen, J. D. The neural basis of error detection: Conflict monitoring and the error-related negativity. Psychological Review. 111 (4), 931-959 (2004).
- Debener, S., et al. Trial-by-trial coupling of concurrent electroencephalogram and functional magnetic resonance imaging identifies the dynamics of performance monitoring. Journal of Neuroscience. 25 (50), 11730-11737 (2005).
- Hauser, T. U., et al. The feedback-related negativity (FRN) revisited: New insights into the localization, meaning, and network organization. NeuroImage. 84, 159-168 (2014).
- Wang, C., Ulbert, I., Schomer, D. L., Marinkovic, K., Halgren, E. Responses of human anterior cingulate cortex microdomains to error detection, conflict monitoring, stimulus-response mapping, familiarity, and orienting. Journal of Neuroscience. 25 (3), 604-613 (2005).
- Tsujimoto, S., Genovesio, A. Firing variability of frontal pole neurons during a cued strategy task. Journal of Cognitive Neuroscience. 29 (1), 25-36 (2017).
- Womelsdorf, T., Johnston, K., Vinck, M., Everling, S. Theta-activity in anterior cingulate cortex predicts task rules and their adjustments following errors. Proceedings of the National Academy of Sciences of the United States of America. 107 (11), 5248-5253 (2010).
- Womelsdorf, T., Vinck, M., Stan Leung, L., Everling, S. Selective theta synchronization of choice-relevant information subserves goal-directed behavior. Frontiers in Human Neuroscience. 4, 2010(2010).
- Domic-Siede, M., Irani, M., Valdés, J., Perrone-Bertolotti, M., Ossandón, T. Theta activity from frontopolar cortex, mid-cingulate cortex and anterior cingulate cortex shows different roles in cognitive planning performance. NeuroImage. 226, 117557(2021).
- Wilensky, R. Planning and Understanding: A Computational Approach to Human Reasoning. , Addison-Wesley. Reading, MA. (1983).
- Grafman, J., Hendler, J. Planning and the brain. Behavioral and Brain Sciences. 14, 563-564 (1991).
- Hayes-Roth, B., Hayes-Roth, F. A cognitive model of planning. Cognitive Science. 3 (4), 275-310 (1979).
- Tremblay, M., et al. Brain activation with a maze test: An EEG coherence analysis study in healthy subjects. Neuroreport. 5 (18), 2449-2453 (1994).
- Shallice, T. Specific impairments of planning. Philosophical Transactions of the Royal Society B: Biological Sciences. 298 (1089), 199-209 (1982).
- Unterrainer, J. M., Owen, A. M. Planning and problem solving: From neuropsychology to functional neuroimaging. Journal of Physiology. 99 (4-6), 308-317 (2006).
- Domic-Siede, M., et al. La planificación cognitiva en el contexto de la evaluación neuropsicológica e investigación en neurociencia cognitiva: Una revisión sistemática. Terapia Psicológica. 40 (3), 367-395 (2022).
- Miotto, E. C., Morris, R. G. Virtual planning in patients with frontal lobe lesions. Cortex. 34 (5), 639-657 (1998).
- Burgess, P. W., Simons, J. S., Coates, L. M. A., Channon, S. The search for specific planning processes. The Cognitive Psychology of Planning. Morris, R., Ward, G. , Psychology Press. Hove, UK. 199-227 (2005).
- Oosterman, J. M., Wijers, M., Kessels, R. P. C. Planning or something else? Examining neuropsychological predictors of zoo map performance. Applied Neuropsychology. 20 (2), 103-109 (2013).
- Campbell, Z., et al. Utilizing virtual reality to improve the ecological validity of clinical neuropsychology: An fMRI case study elucidating the neural basis of planning by comparing the Tower of London with a three-dimensional navigation task. Applied Neuropsychology. 16 (4), 295-306 (2009).
- Wilson, B. A., Alderman, N., Burgess, P. W., Emslie, H., Evans, J. J. Behavioural Assessment of the Dysexecutive Syndrome. , Thames Valley Test Company. St Edmunds, UK. (1996).
- Spector, L., Grafman, J. Planning, neuropsychology, and artificial intelligence: Cross fertilization. Handbook of Neuropsychology. Boller, F., Grafman, J. 9, Elsevier Science Publishers. Amsterdam, the Netherlands. 377-392 (1994).
- Porteus, S. D. The Maze Test and Clinical Psychology. , Pacific Books. Palo Alto, California. (1959).
- Lis, S., Krieger, S., Wilhelm, J., Gallhofer, B. Feedback about previous action improves executive functioning in schizophrenia: An analysis of maze solving behaviour. Schizophrenia Research. 78 (2-3), 243-250 (2005).
- Zelinsky, G. J. A theory of eye movements during target acquisition. Psychological Review. 115 (4), 787-835 (2008).
- Wurtz, R. H., Goldberg, M. E., Robinson, D. L. Brain mechanisms of visual attention. Scientific American. 246 (6), 124-135 (1982).
- Burr, D., Morrone, M. C. Eye movements: Building a stable world from glance to glance. Current Biology. 15 (20), 839-840 (2005).
- Rolfs, M. Microsaccades: Small steps on a long way. Vision Research. 49 (20), 2415-2441 (2009).
- Ortega, J., Plaska, C. R., Gomes, B. A., Ellmore, T. M. Spontaneous eye blink rate during the working memory delay period predicts task accuracy. Frontiers in Psychology. 13, 788231(2022).
- Grueschow, M., Kleim, B., Ruff, C. C. Role of the locus coeruleus arousal system in cognitive control. Journal of Neuroendocrinology. 32 (12), 12890(2020).
- Aston-Jones, G., Shaver, R., Dinan, T. Cortically projecting nucleus basalis neurons in rat are physiologically heterogeneous. Neuroscience Letters. 46 (1), 19-24 (1984).
- Foote, S. L., Bloom, F. E., Aston-Jones, G. Nucleus locus coeruleus: New evidence of anatomical and physiological specificity. Physiological Reviews. 63 (3), 844-914 (1983).
- Siever, L. J., Davis, K. L. Overview: Toward a dysregulation hypothesis of depression. The American Journal of Psychiatry. 142 (9), 1017-1031 (1985).
- Aston-Jones, G., Gold, J. I. How we say no: Norepinephrine, inferior frontal gyrus, and response inhibition. Biological Psychiatry. 65 (7), 548-549 (2009).
- Aston-Jones, G., Cohen, J. D. An integrative theory of locus coeruleus norepinephrine function: adaptive gain and optimal performance. Annual Review of Neurosciences. 28, 403-450 (2005).
- Einhauser, W., Stout, J., Koch, C., Carter, O. Pupil dilation reflects perceptual selection and predicts subsequent stability in perceptual rivalry. Proceedings of the National Academy of Sciences of the United States of America. 105 (5), 1704-1709 (2008).
- Yoshitomi, T., Ito, Y., Inomata, H. Adrenergic excitatory and cholinergic inhibitory innervations in the human iris dilator. Experimental Eye Research. 40 (3), 453-459 (1985).
- Rajkowski, J., Kubiak, P., Aston-Jones, G. Locus coeruleus activity in monkey: Phasic and tonic changes are associated with altered vigilance. Brain Research Bulletin. 35 (5-6), 607-616 (1994).
- Beatty, J. Task-evoked pupillary responses, processing load, and the structure of processing resources. Psychological Bulletin. 91 (2), 276-292 (1982).
- Beatty, J., Kahneman, D. Pupillary changes in two memory tasks. Psychonomic Science. 5, 371-372 (1966).
- Hess, E. H., Polt, J. M. Pupil size in relation to mental activity during simple problem-solving. Science. 143 (3611), 1190-1192 (1964).
- Kuchinke, L., Vo, M. L., Hofmann, M., Jacobs, A. M. Pupillary responses during lexical decisions vary with word frequency but not emotional valence. International Journal of Psychophysiology. 65 (2), 132-140 (2007).
- Barkley, R. A. Adolescents with attention-deficit/hyperactivity disorder: An overview of empirically based treatments. Journal of Psychiatric Practice. 10 (1), 39-56 (2004).
- Gau, S. S. F. em89, Shang, C. Y. Executive functions as endophenotypes in ADHD: Evidence from the Cambridge Neuropsychological Test Battery (CANTAB). Journal of Child Psychology and Psychiatry and Allied Disciplines. 51 (7), 838-849 (2010).
- Bora, E., Harrison, B. J., Yücel, M., Pantelis, C. Cognitive impairment in euthymic major depressive disorder: A meta-analysis. Psychological Medicine. 43 (10), 2017-2026 (2013).
- Rive, M. M., Koeter, M. W. J., Veltman, D. J., Schene, A. H., Ruhé, H. G. Visuospatial planning in unmedicated major depressive disorder and bipolar disorder: Distinct and common neural correlates. Psychological Medicine. 46 (11), 2313-2328 (2016).
- Holt, D. V., Wolf, J., Funke, J., Weisbrod, M., Kaiser, S. Planning impairments in schizophrenia: Specificity, task independence and functional relevance. Schizophrenia Research. 149 (1-13), 174-179 (2013).
- Lima-Silva, T. B., et al. Functional profile of patients with behavioral variant frontotemporal dementia (bvFTD) compared to patients with Alzheimer's disease and normal controls. Dementia and Neuropsychologia. 7 (1), 96-103 (2013).
- Karnath, H. O., Wallesch, C. W., Zimmermann, P. Mental planning and anticipatory processes with acute and chronic frontal lobe lesions: A comparison of maze performance in routine and non-routine situations. Neuropsychologia. 29 (4), 271-290 (1991).
- Staudigl, T., Hartl, E., Noachtar, S., Doeller, C. F., Jensen, O. Saccades are phase-locked to alpha oscillations in the occipital and medial temporal lobe during successful memory encoding. PLoS Biology. 15 (12), 2003404(2017).
- Faul, F., Erdfelder, E., Lang, A. G., Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behavior Research Methods. 39 (2), 175-191 (2007).
- Ferrando, L., Bobes, J., Gibert, J., Soto, M. MINI Entrevista Neuropsiquiátrica Internacional (MINI International Neuropsychiatric Interview, MINI). Instrumentos de Detección y Orientación Diagnóstica. , Retrieved from www.novartis.es (2000).
- McAssey, M., Dowsett, J., Kirsch, V., Brandt, T., Dieterich, M. Different EEG brain activity in right and left-handers during visually induced self-motion perception. Journal of Neurology. 267 (1), 79-90 (2020).
- Kelly, R., Mizelle, J. C., Wheaton, L. A. Distinctive laterality of neural networks supporting action understanding in left- and right-handed individuals: An EEG coherence study. Neuropsychologia. 75, 20-29 (2015).
- O'Hare, A. J., Atchley, R. A., Young, K. M. Central and divided visual field presentation of emotional images to measure hemispheric differences in motivated attention. Journal of Visualized Experiments. (129), 56257(2017).
- Beatty, J. Task-evoked pupillary responses, processing load, and the structure of processing resources. Psychological Bulletin. 91 (2), 276-292 (1982).
- Wainstein, G., et al. Pupil size tracks attentional performance in attention-deficit/hyperactivity disorder. Scientific Reports. 7 (1), 8228(2017).
- Drueke, B., et al. Neural correlates of positive and negative performance feedback in younger and older adults. Behavioral and Brain Functions. 11, 17(2015).
- Lezak, M. D. The problem of assessing executive functions. International Journal of Psychology. 17 (1-4), 281-297 (1982).
- Oosterman, J. M., Wijers, M., Kessels, R. P. C. Planning or something else? Examining neuropsychological predictors of zoo map performance. Applied Neuropsychology. 20 (2), 103-109 (2013).
- Keil, A., et al. Committee report: Publication guidelines and recommendations for studies using electroencephalography and magnetoencephalography. Psychophysiology. 51 (1), 1-21 (2014).
- Abokyi, S., Owusu-Mensah, J., Osei, K. A. Caffeine intake is associated with pupil dilation and enhanced accommodation. Eye. 31 (4), 615-619 (2017).
- Wilhelm, B., Stuiber, G., Lüdtke, H., Wilhelm, H. The effect of caffeine on spontaneous pupillary oscillations. Ophthalmic & Physiological Optics. 34 (1), 73-81 (2014).
- McGinley, M. J., David, S. V., McCormick, D. A. Cortical membrane potential signature of optimal states for sensory signal detection. Neuron. 87 (1), 179-192 (2015).
- Slotnick, S. D. High density event-related potential data acquisition in cognitive neuroscience. Journal of Visualized Experiments. (38), 1945(2010).
- Lamm, C., Windischberger, C., Leodolter, U., Moser, E., Bauer, H. Co-registration of EEG and MRI data using matching of spline interpolated and MRI-segmented reconstructions of the scalp surface. Brain Topography. 14 (2), 93-100 (2001).
- Schwartz, D., et al. Registration of MEG/EEG data with 3D MRI: Methodology and precision issues. Brain Topography. 9, 101-116 (1996).
- Laeng, B., Sirois, S., Gredebäck, G. Pupillometry: A window to the preconscious? Perspectives on psychological science. Journal of the Association for Psychological Science. 7 (1), 18-27 (2012).
- Vandierendonck, A. A comparison of methods to combine speed and accuracy measures of performance: A rejoinder on the binning procedure. Behavior Research Methods. 49 (2), 653-673 (2017).
- Vandierendonck, A. Further tests of the utility of integrated speed-accuracy measures in task switching. Journal of Cognition. 1 (1), 8(2018).
- Levene, H. Robust tests for equality of variances. In Contributions to Probability and Statistics: Essays in Honor of Harold Hotelling. Olkin, I. , Stanford University Press. Redwood City, CA. 278-292 (1960).
- Zimmerman, D. W. A note on preliminary tests of equality of variances. The British Journal of Mathematical and Statistical Psychology. 57, 173-181 (2004).
- D'Agostino, R. B., Belanger, A., D'Agostino, R. B. A suggestion for using powerful and informative tests of normality). The American Statistician. 44 (4), 316(1990).
- Shapiro, S. S., Wilk, M. B. An analysis of variance test for normality (complete samples). Biometrika. 52 (3-4), 591-611 (1965).
- Wilcoxon, F. Individual comparisons by ranking methods. Biometrics Bulletin. 1 (6), 80(1945).
- Fay, M. P., Proschan, M. A. Wilcoxon-Mann-Whitney or t-test? On assumptions for hypothesis tests and multiple interpretations of decision rules. Statistics Surveys. 4, 1-39 (2010).
- Delorme, A., Makeig, S. EEGLAB: An open-source toolbox for analysis of single-trial EEG dynamics including independent component analysis. Journal of Neuroscience Methods. 134 (1), 9-21 (2004).
- Dimigen, O., Sommer, W., Hohlfeld, A., Jacobs, A. M., Kliegl, R. Coregistration of eye movements and EEG in natural reading: Analyses and review. Journal of Experimental Psychology. 140 (4), 552-572 (2011).
- Mognon, A., Jovicich, J., Bruzzone, L., Buiatti, M. ADJUST: An automatic EEG artifact detector based on the joint use of spatial and temporal features. Psychophysiology. 48 (2), 229-240 (2011).
- Bell, A. J., Sejnowski, T. J. A non-linear information maximization algorithm that performs blind separation. Advances in Neural Information Processing Systems 7. Leen, T. K., Tesauro, G., Touretsky, D. S. , MIT Press. Cambridge, MA. edited by (1995).
- Plöchl, M., Ossandón, J. P., König, P. Combining EEG and eye tracking: Identification, characterization, and correction of eye movement artifacts in electroencephalographic data. Frontiers in Human Neuroscience. 6, 278(2012).
- Pion-Tonachini, L., Kreutz-Delgado, K., Makeig, S. ICLabel: An automated electroencephalographic independent component classifier, dataset, and website. NeuroImage. 198, 181-197 (2019).
- Gartner, M., Grimm, S., Bajbouj, M. Frontal midline theta oscillations during mental arithmetic: Effects of stress. Frontiers in Behavioral Neuroscience. 9, 96(2015).
- Wang, W., Viswanathan, S., Lee, T., Grafton, S. T. Coupling between theta oscillations and cognitive control network during cross-modal visual and auditory attention: Supramodal vs modality-specific mechanisms. PLoS One. 11 (7), 0158465(2016).
- Maris, E., Oostenveld, R. Nonparametric statistical testing of EEG- and MEG-data. Journal of Neuroscience Methods. 164 (1), 177-190 (2007).
- Tadel, F., Baillet, S., Mosher, J. C., Pantazis, D., Leahy, R. M. Brainstorm: A user-friendly application for MEG/EEG analysis. Computational Intelligence and Neuroscience. 2011, 879716(2011).
- Pascual-Marqui, R. D. Standardized low-resolution brain electromagnetic tomography (sLORETA): Technical details. Methods and Findings in Experimental and Clinical Pharmacology. 24, 5-12 (2002).
- Gramfort, A., Papadopoulo, T., Olivi, E., Clerc, M. OpenMEEG: Opensource software for quasistatic bioelectromagnetics). Biomedical Engineering Online. 9, 45(2010).
- Destrieux, C., Fischl, B., Dale, A., Halgren, E. Automatic parcellation of human cortical gyri and sulci using standard anatomical nomenclature. Neuroimage. 53 (1), 1-15 (2010).
- Orr, J. M., Weissman, D. H. Anterior cingulate cortex makes 2 contributions to minimizing distraction. Cerebral Cortex. 9 (3), 703-711 (2009).
- Christoff, K., Gabrieli, J. D. E. The frontopolar cortex and human cognition: Evidence for a rostrocaudal hierarchical organization within the human prefrontal cortex. Psychobiology. 28 (2), 168-186 (2000).
- Nitschke, K., Köstering, L., Finkel, L., Weiller, C., Kaller, C. P. A meta-analysis on the neural basis of planning: Activation likelihood estimation of functional brain imaging results in the Tower of London task. Human Brain Mapping. 38 (1), 396-413 (2017).
- Barbey, A. K., Koenigs, M., Grafman, J. Dorsolateral prefrontal contributions to human working memory. Cortex. 49 (5), 1195-1205 (2013).
- Le Van Quyen, M., et al. Comparison of Hilbert transform and wavelet methods for the analysis of neuronal synchrony. Journal of Neuroscience Methods. 111 (2), 83-98 (2001).
- Bokil, H., Andrews, P., Kulkarni, J. E., Mehta, S., Mitra, P. P. Chronux: A platform for analyzing neural signals. Journal of Neuroscience Methods. 192 (1), 146-151 (2010).
- Sato, N., Yamaguchi, Y. EEG theta regulates eye saccade generation during human object-place memory encoding. Advances in Cognitive Neurodynamics ICCN 2007. , Springer. Dordrecht, the Netherlands. 429-434 (2008).
- Zaki, J., Ochsner, K. The need for a cognitive neuroscience of naturalistic social cognition. Annals of the New York Academy of Sciences. 1167, 16-30 (2009).
- Shamay-Tsoory, S. G., Mendelsohn, A. Real-life neuroscience: An ecological approach to brain and behavior research. Perspectives on Psychological Science. 14 (5), 841-859 (2019).
- Diamond, A. Executive functions. Annual Review of Psychology. 64, 135-168 (2013).
- Dudai, Y. Memory from A to Z: Keywords, Concepts and Beyond. , Oxford University Press. New York, NY. (2002).
- Kingstone, A., Smilek, D., Ristic, J., Friesen, C. K., Eastwood, J. D. Attention, researchers! It is time to take a look at the real world. Current Directions in Psychological Science. 12 (5), 176-180 (2003).
- Morris, R. G., Ward, G. Introduction to the psychology of planning. The Cognitive Psychology of Planning. Morris, R., Ward, G. , Psychology Press. Hove, UK. (2005).
- Zwosta, K., Ruge, H., Wolfensteller, U. Neural mechanisms of goal-directed behavior: Outcome-based response selection is associated with increased functional coupling of the angular gyrus. Frontiers in Human Neuroscience. 9, 180(2015).
- Owen, A. M., Doyon, J., Petrides, M., Evans, A. C. Planning and spatial working memory: A positron emission tomography study in humans. European Journal of Neuroscience. 8 (2), 353-364 (1996).
- Ossandon, T., et al. Efficient "pop-out" visual search elicits sustained broadband gamma activity in the dorsal attention network. Journal of Neuroscience. 32 (10), 3414-3421 (2012).
- Lezak, M. D. Neuropsychological Assessment., 3rd edition. , Oxford University Press. New York, NY. (1995).
- Lin, H., Saunders, B., Hutcherson, C. A., Inzlicht, M. Midfrontal theta and pupil dilation parametrically track subjective conflict (but also surprise) during intertemporal choice. NeuroImage. 172, 838-852 (2018).
- Yu, S., Mückschel, M., Beste, C. Superior frontal regions reflect the dynamics of task engagement and theta band-related control processes in time-on task effects. Scientific Reports. 12 (1), 846(2022).
- Nakatani, H., van Leeuwen, C. Antecedent occipital alpha band activity predicts the impact of oculomotor events in perceptual switching. Frontiers in Systems Neuroscience. 7, 19(2013).
- Velasques, B., et al. Changes in saccadic eye movement (SEM) and quantitative EEG parameter in bipolar patients. Journal of Affective Disorders. 145 (3), 378-385 (2013).
- Bodala, I. P., Li, J., Thakor, N. V., Al-Nashash, H. EEG and eye tracking demonstrate vigilance enhancement with challenge integration. Frontiers in Human Neuroscience. 10, 273(2016).
- Dimigen, O. Optimizing the ICA-based removal of ocular EEG artifacts from free viewing experiments. NeuroImage. 207, 116117(2020).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır