
Method Article
Задача визуально-пространственного планирования в сочетании с системами отслеживания глаз и электроэнцефалограммы
В этой статье
Резюме
Изучение когнитивного планирования, сочетающее системы ЭЭГ и отслеживания взгляда, обеспечивает мультимодальный подход к исследованию нейронных механизмов, которые опосредуют когнитивный контроль и целенаправленное поведение у людей. В этой статье мы опишем протокол исследования роли колебаний мозга и движений глаз в планировании производительности.
Аннотация
Процесс планирования, характеризующийся способностью сформулировать организованный план для достижения цели, имеет важное значение для целенаправленного поведения человека. Поскольку при некоторых нервно-психических расстройствах планирование нарушено, проведение надлежащих клинических и экспериментальных тестов для изучения планирования имеет решающее значение. Из-за характера развертывания планирования, в котором участвуют несколько когнитивных областей, оценка планирования и проектирование поведенческих парадигм в сочетании с методами нейровизуализации являются актуальными задачами когнитивной нейронауки. Задача планирования оценивалась в сочетании с системой электроэнцефалограммы (ЭЭГ) и записями движений глаз у 27 здоровых взрослых участников. Планирование можно разделить на две стадии: стадию ментального планирования, на которой внутренне представлена последовательность шагов, и стадию выполнения, на которой двигательные действия используются для достижения ранее запланированной цели. Наш протокол включал в себя задачу планирования и задачу контроля. Задача планирования включала в себя решение 36 лабиринтов, каждое из которых представляло собой карту зоопарка. Задание состояло из четырех периодов: i) планирование, где испытуемым было поручено спланировать путь к местам обитания четырех животных в соответствии с набором правил; ii) сопровождение, при котором испытуемые должны были сохранить запланированный путь в своей рабочей памяти; iii) выполнение, в котором испытуемые с помощью движений глаз отслеживали ранее запланированный путь, указанный системой отслеживания взгляда; и iv) ответ, в котором испытуемые сообщали о порядке посещенных животных. Контрольная задача имела аналогичную структуру, но компонент когнитивного планирования был удален путем модификации цели задачи. Пространственные и временные модели ЭЭГ показали, что планирование вызывает постепенное и длительное повышение лобно-срединной тета-активности (FMθ) с течением времени. Источник этой активности был идентифицирован в префронтальной коре с помощью анализа источников. Наши результаты показали, что экспериментальная парадигма, сочетающая ЭЭГ и системы айтрекера, является оптимальной для оценки когнитивного планирования.
Введение
В течение последних 10 лет были проведены обширные исследования для изучения роли колебательной нейронной динамики как в познании, так и в поведении. Эти исследования установили, что частотно-специфические взаимодействия между специализированными и широко распространенными областями коры головного мозга играют решающую роль в познании и когнитивном контроле 1,2,3. Этот подход подчеркивает ритмическую природу мозговой активности, которая помогает координировать крупномасштабную динамику коры головного мозга и лежит в основе когнитивной обработки и целенаправленного поведения 4,5. Существуют существенные доказательства того, что ритмические колебания в мозге участвуют в различных когнитивных процессах, включая восприятие6, аттентио 7,8,9, принятие решений10, реактивацию памяти11, рабочую память12 и когнитивный контроль13. Были предложены различные осцилляторные механизмы для управления целенаправленным поведением, при этом переходные крупномасштабные частотно-специфичные сети обеспечивают основу для когнитивной обработки 1,14,15. Например, недавние результаты показывают, что определенные частотные полосы в мозге могут отражать механизм обратной связи, который регулирует активность спайков, обеспечивая временную систему отсчета для координации возбудимости коры головного мозга и времени возникновения спайков для продуцированияповедения. Обзор предоставлен Helfrich and Knight19.
Эта совокупность данных поднимает вопросы о том, как префронтальная кора (ПФК) кодирует контексты задач планирования и связанные с ними поведенчески релевантные правила. Долгое время считалось, что ПФК поддерживает когнитивный контроль и целенаправленное поведение через осцилляторные паттерны нейронной активности, которые она генерирует, избирательно смещая нейронную активность в отдаленных областях мозга и контролируя поток информации в крупномасштабных нейронных сетях. Кроме того, было высказано предположение, что регионы, демонстрирующие локальную синхронность, с большей вероятностью будут участвовать в межрегиональной деятельности 21,22,23. В частности, корковые колебания тета-диапазона (4-8 Гц), измеряемые с помощью электроэнцефалограммы (ЭЭГ) кожи головного мозга, были предложены в качестве потенциального механизма передачи нисходящего контроля по широким сетям13. В частности, активность тета-полос у людей отражает когнитивные процессы высокого уровня, такие как кодирование и извлечение памяти, сохранение рабочей памяти, обнаружение новизны, принятие решений и нисходящий контроль 12,24,25,26.
В связи с этим Кавана и Франкпредложили два последовательных механизма процессов управления: признание необходимости контроля и реализация контроля. На осознание необходимости контроля может указывать активность лобной срединной тета (FMθ), исходящая из медиальной префронтальной коры (mPFC), которая была описана в терминах компонентов, связанных с потенциалом события (ERP), которые отражают связанные с mPFC процессы управления в ответ на различные ситуации, такие как новая информация 27,28,29, конфликтующие требования стимул-реакция 30, обратная связь по ошибкам31. и обнаружение ошибок32. Эти компоненты ERP, которые отражают потребность в усилении когнитивного контроля при наличии новизны, конфликта, наказания или ошибки, демонстрируют общую спектральную сигнатуру в тета-диапазоне, зарегистрированную на фронтальных срединных электродах 26,27,33,34,35,36,37,38,39,40. 41,42,43,44.
Реакции ЭЭГ активности FMθ демонстрируют паттерн сброса фазы и усиления мощности в тета-диапазоне частот26. Несмотря на ограничения метода ЭЭГ с точки зрения его пространственного разрешения, были собраны различные источники доказательств, демонстрирующих, что активность FMθ генерируется средней поясной корой (MCC)13. Считается, что эти тета-динамики служат временными рамками, регулирующими нейронные процессы mPFC, которые впоследствии усиливаются в ответ на события, требующие повышенногоконтроля. Это было установлено с помощью анализа источников 31,33,45,46,47, одновременных записей ЭЭГ и функциональной магнитно-резонансной томографии (фМРТ) 48,49, а также инвазивных записей ЭЭГ у людей 50 и обезьян51,52,53.
На основании этих наблюдений считается, что лобная срединная тета служит универсальным механизмом, общим языком для осуществления адаптивного контроля в различных ситуациях, где отсутствует определенность в отношении действий и результатов, например, во время планирования. Поведенческая парадигма, которую мы предлагаем в этом протоколе, была использована для изучения когнитивного планирования и его временных и нейронных характеристик. Несмотря на то, что в других сценариях сообщалось о различных механизмах когнитивного контроля, текущий протокол позволил недавно описать планирование и связанные с ним нейронные и временные свойства. Когнитивный процесс планирования состоит из двух различных фаз: фазы ментального планирования, в ходекоторой вырабатывается внутреннее представление последовательности планов, и фазы выполнения планирования, в ходе которой выполняется набор двигательных действий для достижения ранее запланированной цели. Известно, что планирование требует интеграции различных компонентов исполнительных функций, включая рабочую память, контроль внимания и торможение реакции, что делает экспериментальное манипулирование и изолированное измерение этих процессов сложнойзадачей.
В нейровизуализационных исследованиях когнитивного планирования широко используются поведенческие парадигмы, такие как Лондонский Тауэр 59,60,61; Тем не менее, для того, чтобы контролировать вмешивающиеся факторы, задачи, используемые для изучения когнитивного планирования, могут стать ограниченными и искусственными, что приведет к снижению прогностической и экологической валидности. Для преодоления этой проблемы в области нейропсихологии в качестве экологических задач были предложены ситуации планирования реального мира62,63. Субтест Zoo Map Task в Поведенческой оценке батареи дисисполнительного синдрома измеряет планирование и организационные навыки более естественным и актуальным образом64,66. Этот тест представляет собой тест с использованием карандаша и бумаги, который включает в себя планирование маршрута для посещения 6 из 12 мест на карте зоопарка. Локации представляют собой обычные места, которые можно найти в обычном зоопарке, такие как домик для слонов, клетка со львами, зона отдыха, кофейня и т.д. Существуют два условия, которые оценивают различные уровни планирования: i) условие формулировки, когда испытуемым предлагается спланировать маршрут для посещения шести мест в порядке их выбора, но в соответствии с набором правил; и ii) условие выполнения, когда субъектам дается указание посетить шесть мест в определенном порядке и в соответствии с набором правил. Эти два условия дают информацию о навыках планирования в плохо структурированных (формулировка) и хорошо структурированных (исполнение) задачах67. Первая представлена как более сложная когнитивная задача в открытой ситуации, поскольку требует от испытуемых выработки логической стратегии для достижения цели. Перед трассировкой пути необходимо разработать последовательность операторов; В противном случае возможны ошибки. С другой стороны, условие выполнения требует более низкого когнитивного спроса, поскольку решение задачи, включающей следование определенной навязанной стратегии, требует от испытуемого лишь контроля за выполнением сформулированного плана для достижения цели66. С другой стороны, лабиринт Портеуса является хорошо известной задачей в области психологии, особенно в области когнитивной психологии и нейропсихологии, и он широко используется в качестве инструмента для оценки различных аспектов познания, таких как решение проблем и планирование68,69. Задача «Лабиринт Портеуса» — это задача, выполненная карандашом и бумагой, которая начинается с простого анализа визуальных стимулов и становится все более сложной. Субъект должен найти и проследить правильный путь от начальной точки до выхода (из нескольких вариантов), следуя при этом правилам, таким как избегание пересекающихся путей и тупиков, и действоватькак можно быстрее. Каждый раз, когда при рисовании пути появляется развилка, испытуемые принимают решения, чтобы достичь цели и не нарушить заданные правила69.
Учитывая ограничения и сильные стороны широко используемых и экологических задач, мы разработали нашу поведенческую парадигму в основном на основе задачи66 на карте зоопарка и задачи68 на лабиринте Портеус. Поведенческая парадигма состоит из четырех отдельных стадий, которые охватывают когнитивный процесс планирования в сценарии повседневной жизни. Эти этапы следующие: Этап 1, планирование, где перед участниками ставится задача создания маршрута для посещения различных локаций на карте, обеспечения соблюдения установленных правил; 2 этап, техническое обслуживание, где участники обязаны сохранить запланированный маршрут в рабочей памяти; Этап 3, исполнение, где участники выполняют свой ранее запланированный маршрут, рисуя и внимательно следя за его точностью; и этап 4, реагирование, где участники сообщают о последовательности посещенных животных в соответствии с запланированным маршрутом54. Наша парадигма позволяет измерять различные параметры способности к планированию с использованием различных этапов, которые отражают различные компоненты планирования (такие как рабочая память, исполнительное внимание и зрительно-пространственные навыки) более реалистичным образом, поскольку составление маршрутов является обычным явлением в повседневной жизни. Кроме того, для контроля искажающих факторов парадигма включает в себя контрольную задачу со структурой задачи планирования и эквивалентными стимулами, которая задействует исполнительные когнитивные компоненты, также участвующие в планировании, но исключает компонент процесса планирования. Это позволяет разделить компонент процесса планирования для сравнения как электрофизиологических маркеров, так и поведенческих параметров54.
Кроме того, отслеживание движения глаз внесло значительный вклад в исследования когнитивной нейробиологии, предоставив неинвазивный метод измерения и анализа движений глаз, который может предоставить ценную информацию о когнитивных процессах и нейронных механизмах, лежащих в основе восприятия, внимания и когнитивных функций. Измерение различных типов движений глаз с помощью системы отслеживания взгляда может предоставить ценную информацию о когнитивных процессах и нейронных механизмах, участвующих в планировании. Например, можно измерить следующие аспекты: фиксации, которые представляют собой периоды устойчивого взгляда, в течение которых усваивается зрительная информация70; саккады, которые представляют собой быстрые движения глаз, используемые для перевода взгляда с одного места на другое71; плавное преследование, которое представляет собой тип движения глаз, позволяющий глазам плавно следовать за движущимся объектом72; микросаккады, представляющие собой небольшие, быстрые движения глаз, происходящие даже во время фиксаций73; и моргание, которое является рефлекторным действием, помогающим сохранить глаза увлажненными и защитить их от посторонних предметов74. Эти движения глаз могут дать представление о когнитивных процессах, участвующих в визуальном поиске, распределении внимания70, визуальном отслеживании72, восприятии73 и рабочей памяти74, которые являются важными компонентами планирования и когнитивного контроля.
С другой стороны, недавние исследования системы locus coeruleus-норадреналин (LC-NE) показали его релевантную роль в когнитивном контроле75. Locus coeruleus (LC) проецируется в несколько областей мозга, таких как кора головного мозга, гиппокамп, таламус, средний мозг, ствол мозга, мозжечок и спинной мозг 76,77,61. Особенно плотные иннервации LC-NE получают области мозга PFC, связанные с когнитивным контролем75. Кроме того, некоторые исследования указывают на то, что хроническая гиперактивность системы ЛК может способствовать появлению симптомов маниакально-депрессивного расстройства, таких как импульсивность и бессонница. Напротив, хроническое снижение функции LC было связано со снижением эмоциональной экспрессии, что является распространенной характеристикой среди пациентов, страдающих депрессией. Сверхактивная реакция locus coeruleus на стимулы может привести к чрезмерной реакции у лиц со стрессом или тревожными расстройствами79. Таким образом, изменения в системе LC-NE могут способствовать появлению симптомов когнитивной и/или эмоциональной дисрегуляции. Неинвазивные методы могут быть использованы для изучения активности locus coeruleus, одним из которых является изменение диаметра зрачка, которое в основном контролируется норадреналином, высвобождаемым из locus coeruleus. Норадреналин действует на мышцу расширения радужной оболочки, стимулируя альфа-адренорецепторы, и на ядро Эдингера-Вестфаля, которое посылает сигналы в цилиарный ганглий и контролирует расширение радужки через активацию постсинаптических альфа-2 адренорецепторов 66,80,81,82. Прямые нейронные записи LC у обезьян подтвердили связь между активностью LC-NE, диаметром зрачка и когнитивными способностями83. Расширение зрачков неоднократно наблюдалось в ответ на повышенные потребности в обработке информации в нескольких когнитивных задачах 71,84,85,86,87.
Электрофизиологические маркеры когнитивного контроля в сочетании с отслеживанием глаз и записью зрачков могут решить важнейшие вопросы о том, как когнитивный контроль и планирование реализуются в мозге. Важность использования нашего протокола, сочетающего в себе системы ЭЭГ и айтрекера, двояка. С одной стороны, когнитивный контроль, по-видимому, требует участия распределенной мозговой активности в точных временных отношениях, которые представляют собой идеальных кандидатов для изучения функционирования сети мозга. С другой стороны, аномалии в любой из этих способностей оказывают серьезное влияние на нормальное поведение, как это может быть в случае различных когнитивных и нервно-психических расстройств, таких как синдром дефицита внимания/гиперактивности88,89, большое депрессивное расстройство90,91, биполярное расстройство 91, шизофрения92, лобно-височная деменция93, а также расстройства, вызванные лобными поражениями94. Кроме того, текущий протокол позволяет использовать пупиллометрию в качестве параметра для сравнения активности и колебаний LC-NE с помощью отслеживания движения глаз и электроэнцефалографии. Это может не только предоставить доказательства теоретической связи между LC-NE, пупиллометрией и нейронными маркерами у людей, но также может позволить отслеживать траекторию развития характеристик, связанных с системой LC-NE во время когнитивного планирования. Тем не менее, в нашей модели мы сосредоточились на проверке того, существует ли определенный паттерн саккад во время планирования, который потенциально может привести к конкретным изменениям колебаний95. Кроме того, мы использовали систему отслеживания взгляда в качестве важной части изучения поведенческого выполнения плана на этапе выполнения нашей поведенческой парадигмы.
Подводя итог, можно сказать, что этот протокол может создать тестируемые модели динамики сети мозга, которые могут служить платформой как для дальнейших фундаментальных исследований, так и для возможных клинических и терапевтических применений.
протокол
Все процедуры, описанные в этом протоколе, были одобрены комитетом по биоэтике медицинского факультета Папского католического университета Чили, и все участники подписали форму информированного согласия до начала исследования (номер исследовательского проекта: 16-251).
1. Набор участников
- Набирайте здоровых взрослых правшей (мужчин и женщин) с нормальным зрением или зрением с коррекцией к норме и проверяйте их по критериям включения/исключения.
Примечание: В этом исследовании приняли участие 27 здоровых людей в возрасте от 19 до 38 лет, свободно говорящих на этих языках. Размер выборки может варьироваться в зависимости от желаемого уровня статистической мощности, а возрастной диапазон может варьироваться в зависимости от конкретного исследовательского вопроса, который будет рассмотрен. В нашем протоколе мы рассчитали размер выборки, принимая во внимание статистический критерий Вилкоксона со знаком ранга, размер эффекта 0,7, альфа-уровень 0,05 и степень 0,95, как описано в Faul et al.96. Мы использовали MINI-Международное нейропсихиатрическое интервью97, проведенное квалифицированным психологом, для оценки участников с точки зрения критериев включения/исключения. Набирайте только правшей, чтобы уменьшить вариабельность сигнала ЭЭГ, потому что у левшей может наблюдаться другое топографическое распределение активности ЭЭГ 98,99,100.
2. Подготовка стимулов
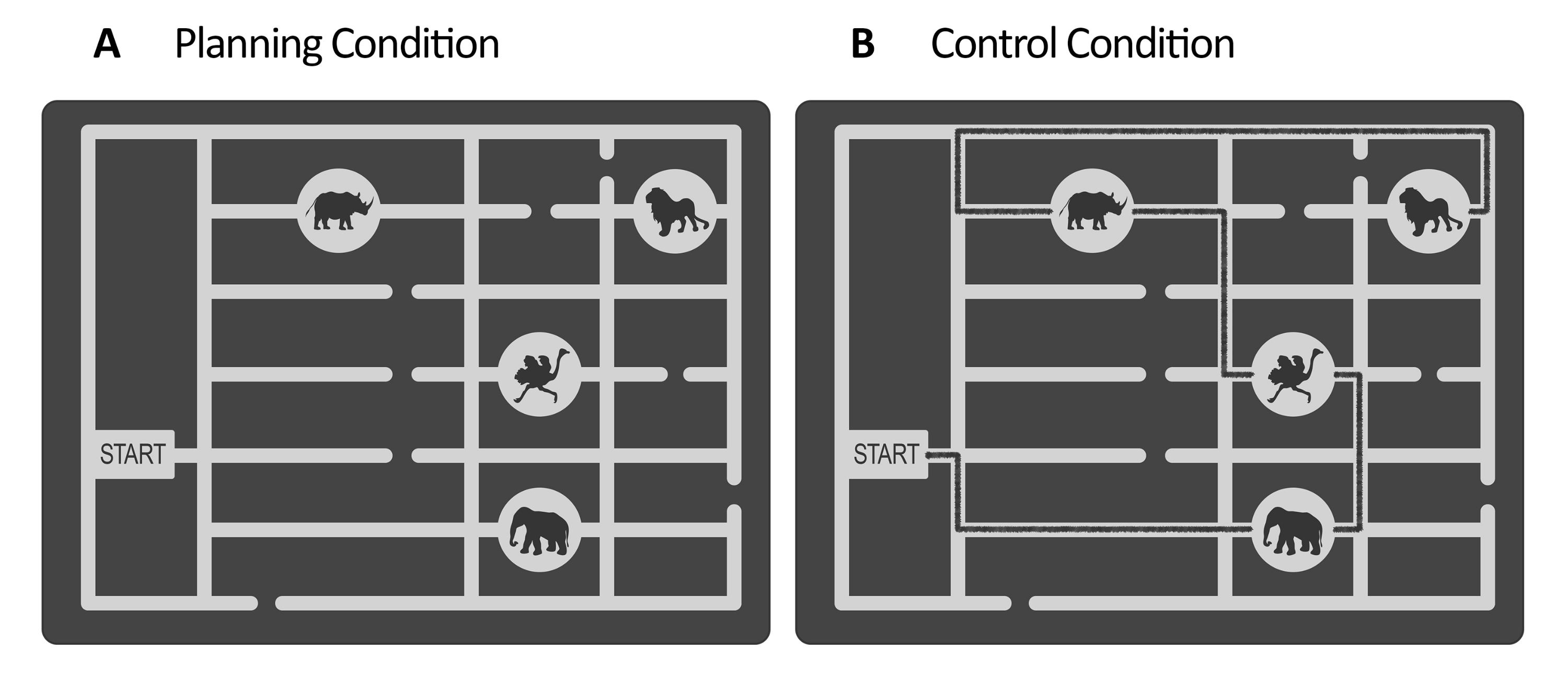
- Для задачи планирования создайте набор стимулов с помощью векторного графического редактора (см. Таблицу материалов). Для каждого стимула создайте лабиринт в оттенках серого, представляющий собой карту зоопарка. Внутри лабиринта сделайте ворота и несколько тропинок, ведущих к местам обитания животных (например, в этом исследовании было четыре места обитания животных, см. рисунок 1).
Примечание: В этом исследовании мы создали 36 лабиринтов, в которых каждый стимул состоял из карты зоопарка со стартовыми воротами, четырех изображений животных, расположенных в лабиринте, и нескольких дорожек. Тропинки в лабиринте могут вести или не вести к местам обитания животных. Использование стимулов в оттенках серого с пониженным контрастом часто предпочтительно для пупиллометрии, потому что это снижает стимуляцию колбочек сетчатки, которые отвечают за цветовое зрение. Этот сдвиг в акценте стимула позволяет более чутко измерять изменения в размере зрачка, которые, как считается, отражают изменения в состоянии возбуждения или внимания. Кроме того, использование стимулов в оттенках серого снижает вариабельность в измерении из-за различий в цветовом зрении между индивидуумами101. - Для того, чтобы получить разные уровни сложности в итоговом задании, разделите стимулы на основе количества валидных решений в соответствии с установленной целью и правилами (в частности, цель состоит в том, чтобы спланировать путь к посещению мест обитания животных). Число допустимых решений относится к числу путей, которые можно спланировать в соответствии с правилами (см. правила на рисунке 2 и шаг 5.12.1). Классифицируйте стимулы с более чем пятью возможными решениями как «легкие», а стимулы с пятью или менее возможными решениями — как «сложные». Затем создайте равное количество пробных версий для каждой категории.
ПРИМЕЧАНИЕ: В качестве альтернативы запросите стимулы, созданные для Domic-Siede et al.54 , у авторов, поскольку эти стимулы были созданы в соответствии с этими инструкциями. Учтите, что все материалы доступны по запросу, но технические характеристики подробно описаны здесь. В этом исследовании мы создали 18 простых испытаний и 18 сложных испытаний. Оценка различий в уровнях сложности на поведенческом и электрофизиологическом уровнях важна, потому что это поможет определить, измеряете ли вы когнитивные потребности/когнитивные усилия/трудности или внутренние аспекты когнитивного планирования (см. репрезентативные результаты и разделы обсуждения). - Для задачи управления используйте ту же структуру, что и в задаче планирования (период оценки, обслуживание, выполнение, реагирование, см. рисунок 2), и используйте те же стимулы, созданные для задачи планирования, но добавьте нарисованную линию, представляющую отмеченный путь для посещения последовательности мест расположения животных, начиная от ворот до последнего места (см. рисунок 1B). Сделайте отмеченный путь немного более темным цветом, чем основные пути лабиринта, с низким контрастом, оцененным с помощью люксметра (см. шаг 2.4).
Примечание: Идея состоит в том, чтобы сохранить психофизические особенности обоих состояний (задач планирования и контроля) как можно более схожими. Проложенные пути лабиринтов могли либо следовать правилам, либо не следовать (см. раздел 5 протокола для получения более подробной информации об инструкциях, данных участникам). В этом исследовании половина стимулов имела правильную последовательность посещения, соответствующую правилам, в то время как другая половина демонстрировала ошибки (например, использование одного и того же пути дважды или пересечение тупиков, см. шаг 5.12.1 и шаг 5.12.3 и рисунок 2). - Оцените освещенность стимулов с помощью люксметра, расположенного в подбородке, которым будут пользоваться испытуемые (см. шаг 4.5 и шаг 4.6) и на одинаковом расстоянии от экрана. Каждый стимул каждого условия производит величину люкса. Запишите каждое значение вручную для дальнейшего анализа.
ПРИМЕЧАНИЕ: Никаких различий в освещенности между условиями не ожидается (см. шаг 4.6). В противном случае проверьте контрастность раздражителей. Это актуально, если диаметр зрачка будет измеряться102. - Создайте один стимул, обозначающий правильную обратную связь (большой палец вверх при правильном) и другой стимул, представляющий неправильную обратную связь (большой палец вниз при неправильной) с помощью векторного графического редактора (см. Таблицу материалов) (рис. 2).

Рисунок 1: Стимулы экспериментальной и контрольной задачи. Приведены наглядные примеры стимулов (А) планирования и (В) контрольной задачи. Стимулы представляют собой карту зоопарка, состоящую из ворот, четырех мест расположения животных в разных местах и нескольких тропинок. Стимулы для обоих состояний были схожими; единственное отличие состояло в том, что для контрольной задачи (В) стимулы имели отмеченную линию, указывающую на уже существующий путь (черная линия здесь для иллюстративных целей). Эта линия в реальных контрольных стимулах была немного темнее, с низким контрастом, контролируемым освещенностью (см. шаг 2.4). Этот рисунок был изменен по Domic-Siede et al.54. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этой цифры.
{kind=link}

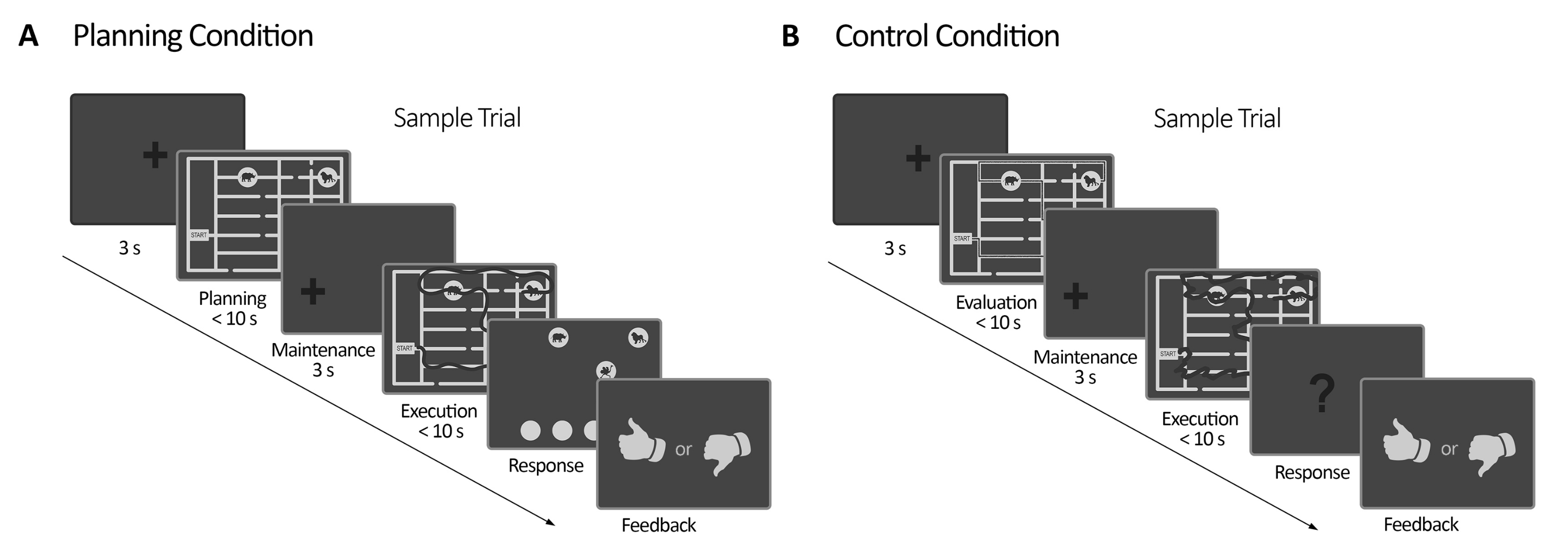
Рисунок 2: План эксперимента. (А) Планирование испытания задачи. Испытания в этом состоянии начинались с 3 с фиксационного креста. Затем участникам было поручено спланировать маршрут для посещения всех четырех мест с животными в соответствии с набором правил (максимум 10 секунд). Далее появился смещенный крест фиксации (3 с), за которым снова последовало появление лабиринта. В этот период (выполнение) испытуемые должны были выполнить трассировку, запланированную в предыдущем периоде планирования, используя свой взгляд с помощью визуальной обратной связи в режиме онлайн (предоставляемой системой отслеживания взгляда), которая обрисовывала движение их взгляда в режиме реального времени (темная линия) (максимум 10 секунд). Впоследствии, в период ответа, испытуемые должны были сообщить о последовательности, установленной во время казни, приказав посещенным животным. Согласно их ответам, была получена обратная связь. (Б) Испытание контрольной задачи. Испытания в этом состоянии начинались с 3 с фиксационного креста. Затем участникам было поручено оценить, соответствует ли правила проложенный путь (темная линия) правилам или нет. Далее появился смещенный крест фиксации (3 с), за которым снова последовало появление лабиринта. В этот период испытуемые должны были перерисовать уже прорисованный путь с визуальной обратной связью в режиме онлайн, как и в период планирования выполнения (максимум 10 с). Впоследствии, в период ответа, испытуемые должны были ответить (да или нет), следовала ли отслеживаемая последовательность ранее установленным правилам. Этот рисунок был изменен по Domic-Siede et al.54. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этой цифры.
{kind=link}
3. Планирование и управление программированием задач
- Напишите сценарий, кодирующий парадигму задачи планирования на основе задачиZoo Map Task 66 и Porteus Maze68, используя программное обеспечение для презентации стимулов/поведенческих экспериментов (см. Таблицу материалов и Дополнительный файл).
- Закодируйте задачу с учетом двух условий (условия планирования и контрольного условия) со структурой, аналогичной той, которая описана в разделах 2 и 4 (см. рисунок 2 и дополнительный файл).
Примечание: Важно использовать одну и ту же структуру в обоих условиях, чтобы контролировать искажающие факторы и воспринимающие компоненты, участвующие в процессе решения требований задачи (рис. 2). Использование одной и той же структуры улучшает оценку конкретного когнитивного процесса, участвующего в когнитивном планировании. - Синхронизируйте обмен данными между компьютером дисплея, компьютером ЭЭГ и главным компьютером (компьютером с отслеживанием глаза) через Ethernet и параллельный порт, отправляя импульсы транзисторно-транзисторной логики (TTL) с компьютера дисплея (см. рис. 3).
- Напишите код для калибровки движений глаз с помощью системы отслеживания взгляда в начале задач планирования и контроля, а также после завершения каждых пяти попыток, поскольку положение взгляда на экране имеет решающее значение для периода выполнения (см. шаги 3.2.3 и 3.3.3 протокола, раздел обсуждения и код в дополнительном файле).
ПРИМЕЧАНИЕ: Возможны задержки в компьютерной связи. Существует несколько методов измерения задержки между импульсами TTL на двух разных компьютерах, но одним из распространенных подходов является использование аппаратного устройства, такого как цифровой осциллограф или логический анализатор. Другой подход заключается в использовании программных методов, таких как отправка импульсов TTL по сетевому соединению и использование инструментов сетевого анализа для измерения задержки. Другой подход заключается в синхронизации часов двух компьютеров либо с помощью глобальной системы позиционирования (GPS) или сервера сетевого протокола времени (NTP), либо с помощью аппаратного решения для синхронизации, вычисления задержки между временной меткой и временем прихода для каждого импульса и усреднения результатов для получения общей задержки между двумя компьютерами.
- Закодируйте задачу с учетом двух условий (условия планирования и контрольного условия) со структурой, аналогичной той, которая описана в разделах 2 и 4 (см. рисунок 2 и дополнительный файл).
- Напишите код для задачи планирования со следующей структурой: период планирования, период обслуживания, период выполнения планирования, период отклика и обратная связь (Рисунок 2, Дополнительный файл).
- Планируемый период: Инициируйте условие планирования, установив фиксирующий крест, представленный в течение 3 секунд в качестве базовой линии.
- Случайным образом представьте набор лабиринтов один за другим (36 в этом исследовании).
ПРИМЕЧАНИЕ: В течение этого периода планирования участников просят спланировать путь для посещения четырех мест для животных, с максимальным временем 10 секунд, следуя набору правил (правила им предварительно объясняются; см. раздел 5 протокола, чтобы ознакомиться с приведенными правилами, а также рисунок 2). - Включите TTL-триггер в код, сигнализирующий о начале предъявления стимула с помощью тег-кода, и отправьте этот триггер на компьютер ЭЭГ и хост-компьютер с айтрекером для дальнейшего более узкого и оконного анализа.
- Пропишите в коде, что период планирования завершается после нажатия кнопки с джойстика/клавиатуры, когда объект заканчивает планирование или если превышено максимальное время. Время реакции (RT) должно быть записано в файле журнала для дальнейшего анализа.
ПРИМЕЧАНИЕ: Для этого периода мы использовали код триггера с цифрой 1, но рекомендуется использовать теги иерархических дескрипторов событий (HED) вместо числовых кодов, потому что теги HED придают смысл и структуру контенту, тем самым облегчая понимание содержимого данных другими исследователями или сотрудниками. - Период технического обслуживания: Инициируйте этот период с помощью смещенного креста фиксации, представленного в течение 3 с. Расположите смещенный крест фиксации в том месте, где расположены ворота лабиринта, чтобы предугадать начальную позицию (ворота) карты зоопарка (см. рис. 2).
ПРИМЕЧАНИЕ: Цель этого периода состоит из трех частей. Во-первых, смещенный крест фиксации облегчает выполнение трассировки, представляющей запланированный путь на следующий период (см. шаг 3.2.8). Во-вторых, в течение этого периода участники держат в своей рабочей памяти план, разработанный в течение планового периода. Наконец, этот период служит межсудебным интервалом для разграничения конца планового периода и начала следующего периода – периода выполнения планирования. - Планируемый период выполнения: После смещения фиксирующего креста показывается в течение 3 с в течение периода обслуживания, снова предъявите лабиринт.
- Отправьте триггер TTL на ЭЭГ и главный компьютер айтрекера, чтобы указать начало этого периода с помощью определенного кода метки.
- Напишите код, обеспечивающий визуальную обратную связь в режиме реального времени (темная линия, см. период выполнения на рисунке 2) положения взгляда субъекта примерно через 992 мс после наступления этого периода.
ПРИМЕЧАНИЕ: Начало очерчивания с задержкой (примерно 1000 мс) дает испытуемым время сориентироваться в лабиринте, что позволяет им очертить свой предыдущий запланированный путь (в течение периода планирования) темной линией. - Запишите координаты путей для дальнейшей реконструкции траекторий, проложенных испытуемыми, и оцените производительность в автономном режиме (см. шаг 6.1.1, рисунок 4).
- Обеспечьте максимальное время в 10 секунд для отслеживания запланированного пути и позвольте испытуемым завершить этот период нажатием кнопки. Таким образом, субъекты могут контролировать, когда они закончат рисовать свой контур.
- Сохраните RT в файле журнала для дальнейшего анализа.
- Срок ответа: Напишите код для периода отклика, который начинается через 10 секунд после выполнения планирования или после нажатия кнопки в конце периода выполнения планирования, в котором лабиринт исчезает, но животные и их пространственное положение остаются на экране.
- Поместите четыре пустых круга горизонтально в нижней части экрана в период отклика.
ПРИМЕЧАНИЕ: Цель этого периода состоит в том, чтобы позволить субъектам указать последовательность посещенных животных в течение периода выполнения планирования, расположив животных в кругах в том же порядке, в котором они посещали их с помощью джойстика или клавиатуры. - Настройте программу/код таким образом, чтобы испытуемые могли использовать джойстик или клавиатуру для выбора каждого из представленных ранее животных (четырех животных в этом исследовании) и вставлять их в каждый из четырех кругов (см. дополнительный файл и рисунок 2).
- Обратная связь: Напишите код, чтобы предоставить участникам 3 секунды обратной связи. Изображение с большим пальцем вверх должно отображаться в ответ на допустимые комбинации посещенных животных, если правила соблюдены, в то время как изображение с большим пальцем вниз должно отображаться, если комбинация недопустима.
- Отправьте триггер TTL, используя определенный код метки для правильной обратной связи и еще один код метки для неправильной версии, на компьютеры ЭЭГ и айтрекера.
ПРИМЕЧАНИЕ: Причина предоставления обратной связи заключается в том, чтобы облегчить контроль за выполнением и поддерживать мотивацию во время выполнения задачи. Такое предоставление обратной связи в режиме реального времени усиливает эффект вознаграждения и стимулирует надлежащее выполнение задачи103.
- Напишите код для контрольной задачи с той же структурой, что и условие планирования: контрольный период, период обслуживания, период выполнения контроля, период отклика и обратная связь (см. дополнительный файл, рисунок 2).
- Контрольный период: Напишите код для периода контрольного условия, чтобы смягчить искажающие факторы. Код для этого периода должен начинаться с фиксирующего креста, представленного за 3 с в качестве исходного уровня.
Примечание: Поскольку задача планирования в основном требует осуществления планирования, но также задействует другие когнитивные области как часть исполнительной функции, такие как зрительно-пространственная функция, рабочая память, контроль внимания, ингибиторный контроль и т.д.66,88,104,105, задача управления имеет решающее значение для смягчения искажающих факторов. Таким образом, основная цель этой задачи состоит в том, чтобы задействовать все когнитивные и перцептивные функции, необходимые для решения задачи планирования, исключив при этом реализацию когнитивного планирования54. - Случайным образом поочередно представляйте лабиринты контрольного условия (лабиринты с уже проложенным отмеченным путем). Время кодирования не более 10 с.
- Включите TTL-триггер в код, сигнализирующий о начале предъявления стимула с помощью метки, и отправьте этот триггер на компьютер ЭЭГ и главный компьютер айтрекера.
- Пропишите в коде, что этот период управления завершается после нажатия кнопки джойстика/клавиатуры всякий раз, когда объект заканчивает работу или при превышении максимального времени.
ПРИМЕЧАНИЕ: Испытуемым предлагается оценить отмеченные пути (следуют ли они правилам или нет, см. шаг 5.12 для получения подробной информации об инструкциях, данных участникам). - Сохраните время реакции (RT) в файле журнала для дальнейшего анализа.
- Период технического обслуживания: По окончании контрольного периода предъявите смещенный фиксирующий крест в течение 3 с.
- В качестве периода планирования поместите фиксирующий крест в том месте, где расположен вход в ворота, чтобы облегчить рисование взгляда на следующий период.
- Период выполнения контроля: Снова представьте лабиринт и одновременно отправьте TTL-триггер на ЭЭГ и хост-компьютеры с отслеживанием глаз с меткой, сигнализирующей о начале периода выполнения.
- Повторите тот же код, что и в период выполнения планирования, чтобы дать онлайн-обратную связь о положении взгляда, а также очертить и перекрыть свой взгляд с трассируемым путем.
- Обеспечьте максимальное время трассировки пути в 10 секунд и позвольте объектам завершить этот период нажатием кнопки.
- Сохраните RT в файле журнала для дальнейшего анализа.
- Период срабатывания контроля: По окончании периода выполнения контроля предъявите вопросительный знак с указанием периода ответа.
- Запрограммируйте соответственно две кнопки, чтобы испытуемые давали ответ с помощью джойстика или клавиатуры.
ПРИМЕЧАНИЕ: Здесь испытуемым предлагается ответить, была ли последовательность, отмеченная трассировкой, правильной или нет, выбрав одну кнопку для правильного/ДА и другую для неверного/НЕТ. - Сохраните точность в файле журнала.
- Обратная связь: Напишите код, который будет выдавать 3 секунды правильной обратной связи, когда испытуемые отвечают правильно (изображение с большим пальцем вверх), и 3 секунды неправильной обратной связи, когда испытуемые отвечают неправильно (изображение с большим пальцем вниз).
- Как и в условии планирования, отправьте триггер TTL на ЭЭГ и разместите компьютеры с айтрекером с тегом для правильной обратной связи и другой меткой для неправильной обратной связи.
- Контрольный период: Напишите код для периода контрольного условия, чтобы смягчить искажающие факторы. Код для этого периода должен начинаться с фиксирующего креста, представленного за 3 с в качестве исходного уровня.
- Учебные задачи: Создание стимулов, написание кода и проведение перед вышеупомянутыми задачами планирования и контроля краткой обучающей сессии, состоящей примерно из шести испытаний/лабиринтов для каждого условия (планирование и контроль)
ПРИМЕЧАНИЕ: Идея состоит в том, чтобы обеспечить ознакомление с постановкой задачи. Рекомендуется установить критерии для дальнейших действий. В этом исследовании, если последние три испытания были правильными, и участники сообщали о понимании цели и процедуры в конце тренировочной сессии, участники затем переходили к экспериментальной сессии.

Рисунок 3: Пример лабораторной установки. Схематическое изображение лабораторной установки с изображением трех взаимосвязанных компьютеров. Главный компьютер (компьютер айтрекера) отвечает за отслеживание и хранение данных о движении глаз. Компьютер ЭЭГ получает и сохраняет сигналы ЭЭГ. Компьютер с дисплеем управляет поведенческим экспериментом, представляет стимулы испытуемым и отправляет триггеры событий на хост и компьютеры ЭЭГ через параллельные порты и соединения LAN для синхронизации сбора данных. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этой цифры.
{kind=link}

Рисунок 4: Реконструкция траектории по визуальной онлайн-обратной связи, полученной от системы отслеживания взгляда. Наглядные примеры реконструкции траектории по моторному выполнению плана (А, фиолетовый, период планирования выполнения) и контрольного периода выполнения (В, линия зеленого цвета) и с данными айтрекера. Путь, восстановленный в период выполнения планирования, используется для оценки точности каждой пробной версии задачи планирования. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этой цифры.
{kind=link}
4. Лабораторные установки и оборудование
- Используйте систему регистрации ЭЭГ для регистрации активности ЭЭГ с кожи головы участника, при этом электроды ЭЭГ размещены в соответствии с международной системой10-20 106. Расположите два электрода на сосцевидном отростке участника для повторного отображения в автономном режиме. Используйте электроды электроокулограммы для определения сигналов вертикальных, горизонтальных и мигающих движений глаз во время визуального осмотра.
- Используйте программное обеспечение для сбора данных ЭЭГ с частотой дискретизации 2 048 Гц или 1 024 Гц и полосовым фильтром в диапазоне от 0,1 до 100 Гц в компьютере ЭЭГ.
ПРИМЕЧАНИЕ: Частоты дискретизации 1,024 Гц и 2,048 Гц обеспечивают достаточное разрешение для анализа низкочастотных колебаний. При анализе низкочастотных колебаний важно регистрировать сигналы ЭЭГ с высокой частотой дискретизации, например, более 1000 Гц, чтобы гарантировать, что низкочастотные сигналы не будут искажены. - Используйте компьютер с дисплеем, который подключен к компьютеру ЭЭГ и компьютеру с отслеживанием глаз через параллельные порты и Ethernet и имеет установленную на нем платформу для проведения поведенческих экспериментов, чтобы проецировать стимулы на расширенный монитор с минимальным разрешением 1 920 пикселей x 1 080 пикселей и частотой обновления 60 Гц. Поместите этот монитор на расстоянии примерно 82 см от объекта съемки.
ПРИМЕЧАНИЕ: Мы использовали монитор с диагональю 24 дюйма и частотой обновления 144 Гц, расположенный на расстоянии 82 см от участника. Для когнитивных экспериментов с записью ЭЭГ и движений глаз рекомендуется использовать монитор с размером экрана не менее 19 дюймов. Кроме того, общая рекомендация заключается в том, чтобы расположить монитор на расстоянии, которое позволяет участнику комфортно выполнять задачу и сохранять устойчивый взгляд на экране, обеспечивая при этом точную запись ЭЭГ и движений глаз. Рекомендуется протестировать и отрегулировать настройку по мере необходимости, чтобы обеспечить наилучшие результаты. - Используйте систему отслеживания взгляда, чтобы дать участникам обратную связь в режиме реального времени о движениях их глаз во время выполнения и записать размер зрачка. Установите частоту дискретизации на частоте 1 000 Гц для получения адекватного временного разрешения.
- Избегайте движений головой. Ограничения движения головы влево и вправо, вверх и вниз необходимы для того, чтобы глаз оставался в поле зрения видеокамеры. Ограничение движения вперед и назад необходимо для того, чтобы глаз оставался в фокусном диапазоне видеокамеры. Используйте комбинацию упоров для лба и подбородка, чтобы движения оставались в этом диапазоне.
- Оцените яркость стимулов с помощью цифрового люксметра или аналогичного устройства, чтобы сравнить различия между планирующими стимулами и контрольными стимулами.
Примечание: Статистический тест, такой как t-критерий или критерий Вилкоксона, может быть использован для оценки различий между стимулами двух состояний. - Используйте джойстик управления или клавиатуру как минимум с четырьмя кнопками: две кнопки для вопросов «Да/Нет» из условия управления; одна из этих двух кнопок для завершения испытаний; и еще две кнопки для периода отклика условия планирования для перемещения вперед или назад, чтобы вставить животных в каждый из четырех кругов в нижней части экрана.
5. Сеансы записи электроэнцефалографии и отслеживания движения глаз
- Перед началом исследования попросите участников заполнить письменное и подписанное информированное согласие.
- Перед началом сеанса записи попросите участников не приходить с макияжем (тушь для ресниц и подводка для глаз могут быть распознаны как зрачок системой айтрекера), если они принимали наркотики или кофеин107,108 или если они чувствуют сильную усталость109 (стресс, недосыпание и т.д.).
- Предложите участникам заполнить демографический опрос, чтобы предоставить информацию об их поле, возрасте, роде рук, родном языке и психоневрологическом анамнезе с помощью нейропсихиатрического интервью MINI-International97 , проводимого квалифицированным психологом.
- Очистите лоб, кожу головы, сосцевидный отросток и положение кожи объекта на электроокулограмме (ЭОГ) спиртовой салфеткой.
- Поместите все внешние электроды на участника. Поместите горизонтальную ЭОГ биполярно на внешнюю поверхность обоих глаз, а вертикальную ЭОГ над и ниже правого глаза участника. Установите по два внешних электрода на правый и левый сосцевидный отросток для последующего повторного использования.
- Измерьте голову испытуемого и установите ЭЭГ-шапочку нужного размера в соответствии с расширенной международной системой 10-20. Для этого найдите и установите электрод Cz, выполнив следующие действия:
- Определите среднюю линию кожи головы, визуально осмотрев линию роста волос и верхнюю часть носа. Определите линию, соединяющую эти две точки, чтобы определить среднюю линию.
- Найдите Cz. Cz обычно определяется как средняя точка между двумя преаурикулярными точками (т.е. точками, расположенными непосредственно перед каждым ухом). Найдите эти точки, а затем определите линию, соединяющую их, чтобы определить приблизительное местоположение буквы Cz.
- Измерьте и отметьте Cz. Измерьте расстояние от nasion (т.е. костного выступа в верхней части носа) до Cz. Расстояние от насиона до Cz обычно составляет около 53% от общей окружности головы в системе 10-20. Отметьте местоположение буквы Cz с помощью ручки или другого инструмента для разметки.
ПРИМЕЧАНИЕ: Важно следовать последовательной и стандартизированной процедуре установки электродов, чтобы свести к минимуму ошибки и обеспечить достоверность данных ЭЭГ. Рекомендуется разработать стандартную процедуру размещения. Разработайте стандартную процедуру размещения электродов на коже головы каждого субъекта и убедитесь, что для каждого субъекта используется одна и та же процедура. В случае, если запись выполняет команда или персонал, обучите техников или научных сотрудников правильной процедуре размещения, чтобы обеспечить последовательное и точное размещение электродов. Кроме того, оцифровка положений электродов для каждого объекта была бы желательным подходом для последующего выполнения анализа источника. В исследованиях ЭЭГ точное трехмерное (3D) расположение каждого электрода на голове испытуемого часто является требованием для установления корреляции между данными ЭЭГ исоответствующей активностью мозга. Эта информация также имеет решающее значение для правильного согласования данных ЭЭГ с анатомическими изображениями, полученными с помощью методов МРТ или КТ111,112.
- Введите проводящий гель в каждое отверстие колпачка с помощью шприца с тупой иглой, отодвигая кончиком волосы участника. После этого наденьте все электроды кожи головы на шапочку ЭЭГ.
- Проверьте импедансы с помощью программного обеспечения для записи ЭЭГ и убедитесь, что они находятся ниже уровня сопротивления, рекомендованного системой ЭЭГ.
- Попросите участника оставаться как можно более неподвижным во время эксперимента. Изучите сигнал ЭЭГ и проверьте его, попросив участника моргнуть, сделать челюсть и остаться на несколько секунд с закрытыми глазами.
- Усадите участника в темную и шумоизолированную комнату. Используйте подбородок, чтобы стабилизировать голову и свести к минимуму движения, и убедитесь, что расстояние между подбородком и центром экрана для предъявления стимулов составляет примерно 82 см.
- Поместите джойстик или клавиатуру перед участником для ответов.
- Резолюция: Давайте устные инструкции с помощью наглядных пособий перед началом каждого состояния (планирование и контроль). В инструкции включите наглядные примеры стимулов, а также объясните, как решать лабиринты в условиях планирования и контроля соответственно.
- В задаче на планирование попросите испытуемых найти путь для завершения последовательности посещений определенных мест обитания животных (четыре места в этом исследовании) в разных местах лабиринта в любом порядке и следуя набору правил: «(1) Спланируйте путь как можно быстрее, максимум за 10 секунд; (2) Начните путь от ворот и завершите путь у четвертого посещенного животного; (3) Не проходите через одну и ту же дорожку или угол дважды; (4) Не переходите в тупик; (5) Не переходите дорогу перпендикулярно»54.
ПРИМЕЧАНИЕ: Для правил 2 - 5 мы рекомендуем показать участнику наглядные примеры. - После этого начните тренировку по планированию из шести испытаний.
ПРИМЕЧАНИЕ: Проинструктируйте участников сообщать перед каждым периодом калибровки айтрекера, если возникли какие-либо проблемы с выполнением задачи, особенно во время определения контура в период выполнения. Обратите внимание на то, проводилась ли пробная версия для проверки в автономном режиме/постобработке (см. шаг 6.1.1). - В качестве контрольного условия попросите испытуемых оценить, правильно или неправильно проложен уже отмеченный путь в лабиринте с учетом ранее изученных правил.
ПРИМЕЧАНИЕ: Приведите примеры с визуальной поддержкой того, как оценить лабиринты без использования стратегий планирования, например, не пытаться спланировать новый путь при обнаружении ошибок (например, рисование с использованием одного и того же пути дважды, пересечение тупика и т. д.). При обнаружении ошибки основное внимание следует уделять исключительно сообщению об обнаружении ошибки, а не исправлению пути. После каждого испытания спрашивайте участников о стратегиях, которые они применили. Затем предоставьте устный отзыв об их работе, чтобы убедиться, что они оценили нарисованные пути и не планировали новые пути. После этого начните тренировочную сессию по контрольной задаче из шести попыток.
- В задаче на планирование попросите испытуемых найти путь для завершения последовательности посещений определенных мест обитания животных (четыре места в этом исследовании) в разных местах лабиринта в любом порядке и следуя набору правил: «(1) Спланируйте путь как можно быстрее, максимум за 10 секунд; (2) Начните путь от ворот и завершите путь у четвертого посещенного животного; (3) Не проходите через одну и ту же дорожку или угол дважды; (4) Не переходите в тупик; (5) Не переходите дорогу перпендикулярно»54.
- Проверьте сигнал ЭЭГ, чтобы убедиться, что все каналы регистрируются правильно. Начните запись ЭЭГ.
- Откалибруйте айтрекер.
ПРИМЕЧАНИЕ: Проверьте способность айтрекера определять положение взгляда, когда участник направляет свой взгляд на различные области экрана.- Сообщите участнику, что айтрекер будет откалиброван и что он увидит белый круг (с маленькой серой точкой), хаотично движущийся к четырем углам экрана (процедура калибровки по пяти точкам). Дайте им указание зафиксировать свой взгляд на круге и сообщите им, что, когда он перемещается в другое место, они должны следовать за положением круга и снова фиксировать свой взгляд в этом новом положении.
- Запустите эксперимент, начните сохранять движения глаз, нажав на кнопку «Вывод/запись», и попросите участника следовать ранее полученным инструкциям, сообщив ему, что эксперимент начнется.
- Держите лабораторное помещение в темном помещении. Наибольшие изменения в расширении зрачков происходят в ответ на изменения яркости113. Поддерживайте постоянный уровень освещенности в экспериментальной среде.
6. Анализ данных
- Поведенческий анализ
- Анализируйте поведенческие данные с помощью статистического программного обеспечения (см. Таблицу материалов). Измеряйте точность (процент точных ответов) в качестве количественного поведенческого параметра как в условиях планирования, так и в условиях контроля. В качестве условия планирования используйте данные айтрекера (координаты x и y положения взгляда), чтобы воссоздать пути, пройденные в течение периода выполнения в автономном режиме, и определить точность запланированных путей по сравнению с фактическими трассированными путями (рис. 4). Для этого вручную проверьте соответствие между правильно / неправильно сделанными комбинациями в период отклика и сделанной трассировкой.
- Вычислите RT, который представляет собой среднее время, затраченное на решение лабиринтов за период планирования, и среднее время, затраченное на оценку отмеченных путей за контрольный период.
- Рассчитайте среднее значение RT периодов выполнения для условий планирования и контроля. В частности, используйте ОТ, соответствующую только правильным испытаниям.
ПРИМЕЧАНИЕ: В дополнение к этому, можно использовать линейную интегрированную оценку скорости-точности (LISAS)114,115, описанную в Domic-Siede et al.54, которая обеспечивает комбинированную меру, учитывающую время реакции и точность. Поскольку время реакции в течение периода выполнения планирования и точность планирования взаимосвязаны, LISAS может быть использован для вычисления индекса, который учитывает время реакции, скорректированное на количество допущенных ошибок. Кроме того, индекс LISAS может быть использован для оценки корреляции между электрофизиологическими сигналами и поведенческими показателями. Он рассчитывается как линейная комбинация времени реакции (RT) и доли ошибок (PE). - Оцените гомоскедастичность с помощью статистического теста, такого как тест Левина116,117, и проверьте нормальность с помощью омнибусного теста распределения нормальности Д'Агостино и Пирсона118 или теста Шапиро Уилка119, чтобы выбрать подходящий статистический тест для сравнения (параметрический или непараметрический).
- Оцените, является ли компонент планирования в условиях планирования более когнитивно требовательным, чем в контрольном условии, используя либо тест Вилкоксона со знакомранга 120 , либо t-критерий121 на основе сопоставленной пары для сравнения поведенческих параметров между условиями.
ПРИМЕЧАНИЕ: Таким образом, подтвердите, что поведенческая парадигма является оптимальной для оценки когнитивного планирования. - Разделите испытания в условиях планирования на категории «легкие» и «сложные» (см. шаг 2.2), а затем используйте t-критерий согласованной пары для сравнения точности и времени реакции в периоды планирования и выполнения между «легкими» и «сложными» испытаниями.
- ЭЭГ и предварительная обработка движений глаз
- Выполните конвейер предварительной обработки данных ЭЭГ, описанный в следующих пунктах, с использованием самодельных сценариев и/или установленных наборов инструментов, таких как описанные в Delorme and Makeig122, в Dimigen et al.123 и в Mognon et al.124, в программном обеспечении на языке программирования (см. Таблицу материалов).
- Синхронизируйте активность движений глаз с записями ЭЭГ, чтобы импортировать фиксации, саккады и события моргания для лучшего визуального контроля или дальнейшего анализа (см. шаг 3.1.2 и дополнительный файл).
ПРИМЕЧАНИЕ: В этом исследовании мы использовали временные метки на данных отслеживания движения глаз и временные метки на данных ЭЭГ, как описано в Domic-Siede et al.54 и в Dimigen et al.123 для импорта событий движения глаз в данные ЭЭГ в программном обеспечении на языке программирования. - Уменьшите частоту дискретизации данных до 1 024 Гц, если они были записаны с частотой 2 028 Гц, чтобы снизить вычислительные требования.
ПРИМЕЧАНИЕ: Частота дискретизации 1 024 Гц является достаточной в соответствии с интересующим частотным диапазоном 4-8 Гц, ожидаемым частотным разрешением и вычислительными требованиями анализа. - Повторно соотнесите сигнал ЭЭГ со средним значением электродов на сосцевидных отростках.
ПРИМЕЧАНИЕ: Возможны и другие ссылки. Выбор референса может повлиять на результаты анализа ЭЭГ и интерпретацию данных, поэтому важно тщательно рассмотреть плюсы и минусы различных вариантов референса. Средний эталон сосцевидного отростка является популярным выбором для исследований ЭЭГ, потому что он обеспечивает стабильный эталон, который легко вычислить, и он показал свою эффективность для анализа множества различных сигналов ЭЭГ. Соотнесение данных ЭЭГ со средним значением сосцевидного отростка (известным как средний эталон сосцевидного отростка) является распространенным подходом к анализу лобной тета-активности в данных ЭЭГ кожи головы. Сосцевидные электроды расположены рядом с ухом и служат эталоном для сигналов ЭЭГ. Обращение к среднему значению сосцевидных отростков может помочь уменьшить влияние шума и артефактов, которые не представляют интереса, избегая при этом подавления интересующего сигнала, что помогает пользователю получить более четкое представление сигналов ЭЭГ. - Примените конечную импульсную характеристику (КИХ) с нулевой фазой с частотой среза верхних частот 1 Гц и частотой среза нижних частот 40 Гц к расширенному сигналу (без эпохации) с помощью программного обеспечения на языке программирования.
ПРИМЕЧАНИЕ: В этом исследовании мы использовали набор инструментов, описанный в Delorme and Makeig122. - Для каждого условия, учитывая количество испытаний, разделите данные на эпохи, сосредоточенные вокруг начала периодов планирования и контроля соответственно. Используйте 1 с до начала представления лабиринта в качестве исходной линии и 4 с после периода планирования или контроля в качестве сегментов интереса. Используйте программное обеспечение на языке программирования.
ПРИМЕЧАНИЕ: В этом исследовании мы использовали набор инструментов, описанный в Delorme and Makeig122и 36 эпох/испытаний. - Создайте вторую сегментацию, сосредоточенную вокруг конца периодов планирования и контроля, используя 4 с до конца и 1 с после в качестве периода обслуживания.
ПРИМЕЧАНИЕ: Причина выбора первых и последних 4 секунд периодов планирования и контроля (шаг 6.2.6 и шаг 6.2.7) заключается в том, что продолжительность каждого периода в обоих условиях может варьироваться, и анализ первой и последней секунд планирования может обеспечить более полное представление о процессе планирования. Таким образом, эти длины окон являются адекватными и достаточными для анализа колебательной динамики, лежащей в основе планирования. - Над сегментированным сигналом запустите алгоритм125анализа независимых компонентов (ICA) Logistic Infomax для идентификации и удаления артефактных компонентов.
- Используйте критерий соотношения дисперсии саккады к фиксации, рекомендованный в Plöchl et al.126, для автоматического обнаружения потенциальных зашумленных компонентов, а также используйте автоматический детектор артефактов ЭЭГ, основанный на совместном использовании пространственных и временных особенностей, рекомендованный в Mognon et al.124.
ПРИМЕЧАНИЕ: Мы рекомендуем использовать независимый классификатор компонентов, предложенный в Pion-Tonachini et al.127, который оценивает независимые классификации компонентов как векторы композиции по семи категориям, помогая идентифицировать артефакты. - Осмотрите другие потенциальные артефактные компоненты, такие как ЭМГ, движение электродов или компоненты, не связанные с мозгом. Проверьте отбраковку этих компонентов путем визуального изучения топографий, спектров и активаций с течением времени.
- Интерполяция (сферическая интерполяция) зашумленных каналов путем автоматического подавления каналов с использованием критерия эксцесса (с z-оценкой 5 в качестве порога).
- Частотно-временной анализ
- Выполните кратковременное быстрое преобразование Фурье (БПФ) (от 1 Гц до 40 Гц) с длиной окна 250 мс и временным шагом 5 мс. Используйте окно Хеннинга. Используйте z-оценку для нормализации диаграмм временной частоты до базового уровня (от −1 с до −0,1 с).
ПРИМЕЧАНИЕ: Визуализация спектра зависит от компромисса между размером окна и временным разрешением. Для получения полного представления обо всем спектре, включая тета-диапазон от 4 Гц до 8 Гц, мы рекомендуем использовать нижний предел размера окна, который составляет 250 мс, чтобы обеспечить более высокое временное разрешение во время каждого испытания и задачи. Кроме того, мы рекомендуем использовать окно Хеннинга, так как оно считается обычным выбором для таких случаев. Для лучшего разрешения по времени и частоте см. дальнейшие действия. - Выберите частотно-временную диаграмму от лобно-центрального электрода, такого как Fz, или усредненной группы фронтальных электродов.
Примечание: Рассмотрите обширные доказательства связи между когнитивным контролем и лобной средней тетой 12,128,129. - Выберите нефронтальные управляющие частотно-временные диаграммы от электродов, таких как электроды Pz и Oz, для дальнейшего сравнения.
- Для фронтальных и контрольных электродов выполните непараметрический кластерный тест перестановок для парных образцов с p-значением < 0,05 для сравнения на уровне группы графиков времени и частот в обоих условиях. Используйте метод Монте-Карло с 1 000 случайных розыгрышей. Используйте максимальное статистическое значение кластера для выполнения теста130 на перестановку.
- Усредняем полосу тета-частот (4-8 Гц) от первых 4 с планирования и управления соответственно, а также от последних 4 с отрезка.
- Сравните усредненную тета-активность между условиями с помощью t-критерия согласованной пары или теста Вилкоксона со знаком ранга.
- Проанализируйте временной профиль тета-активности. Для этого усредните частотный диапазон 4-8 Гц по всем испытаниям по испытуемым.
- Сравните динамику тета-активности между условиями с помощью теста Вилкоксона со знаком ранга, сопоставленного и скорректированного с помощью коэффициента ложных открытий (FDR).
ПРИМЕЧАНИЕ: В тесте Вилкоксона мы использовали 88 мс неперекрывающихся окон.
- Выполните кратковременное быстрое преобразование Фурье (БПФ) (от 1 Гц до 40 Гц) с длиной окна 250 мс и временным шагом 5 мс. Используйте окно Хеннинга. Используйте z-оценку для нормализации диаграмм временной частоты до базового уровня (от −1 с до −0,1 с).
- Реконструкция источника
- Используйте набор инструментов для реконструкции анализа источников, такой как набор инструментов с открытым доступом, описанный в Tadel et al.131 , или другой подобный.
- Рассчитайте источники предварительно обработанного сигнала ЭЭГ в течение первых 4 с планирования с помощью такого алгоритма, как стандартизированная электромагнитная томография мозга с низким разрешением (sLORETA)132и метод визуализации с минимальной нормой, а также метод симметричных граничных элементов (симметричная МЭМ), с помощью набора инструментов, подобного описанному в Gramfort et al.133, для решения обратной задачи.
- Используйте исходный алгоритм (алгоритм sLORETA в этом исследовании) на анатомическом шаблоне MNI (мы использовали шаблон MNI в Brainstorm "Colin27") с расположением электродов по умолчанию для каждого участника в случае, если не происходит 3D-оцифровка электродов (см. шаг 5.6).
ПРИМЕЧАНИЕ: Следует отметить, что использование стандартного расположения электродов не является самым эффективным методом определения источников мозговой активности. Тем не менее, он все же может дать общее представление о происхождении деятельности. Важно иметь в виду, что источники локализации, полученные с помощью этих методов, являются грубыми приближениями и должны интерпретироваться с осторожностью при анализе результатов. - Примените полосовой фильтр 4–8 Гц к предварительно обработанному сигналу.
- Примените нормализацию z-оценки, используя в качестве исходного уровня период от −1000 мс до начала исследования.
- Усредните тета-активность, используя интересующее нас временное окно от 1 до 4 с после начала исследования.
- Сравните средние пространственные источники между условиями с помощью непараметрического теста на знак перестановки с выборкой по методу Монте-Карло (1 000 рандомизаций)131.
- Чтобы определить области интереса (ROI), обозначьте кору головного мозга с помощью атласа мозга.
ПРИМЕЧАНИЕ: Мы использовали Destrieux Atlas134 , реализованный в наборе инструментов, описанном в Tadel et al.131. - Выберите интересующие области мозга (ROI).
ПРИМЕЧАНИЕ: Мы рассмотрели доказательства, свидетельствующие о том, что области префронтальной коры, такие как двусторонняя верхняя лобная извилина (SF), двусторонняя поперечная лобно-полярная извилины и борозды (FP), двусторонняя ACC, двусторонняя MCC и двусторонняя дорсолатеральная префронтальная кора137,138, участвуют в функциях когнитивного контроля135,136. - Выполните анализ главных компонент (PCA) по предыдущему предварительно обработанному сигналу ЭЭГ (диапазон 1-40 Гц) для каждого ROI и выберите первый режим разложения PCA для каждого ROI.
- Выполните спектральный анализ с использованием кратковременного быстрого преобразования Фурье и сравните результаты между левой и правой областями интереса с помощью непараметрического кластерного теста перестановок130.
- Извлеките и представьте левый и правый ROI, не показывающие различий, в виде одного двустороннего временного ряда: SF, ACC и MCC. Затем постройте графики времени и частоты и сравните условия.
- Сравните временные и частотные диаграммы в соответствии с уровнем сложности задачи планирования (простые и сложные пробные версии) для каждого ROI.
- Зеркально отразите фронт сигнала для каждой из 512 выборок и выполните полосовой фильтр в диапазоне от 4 Гц до 8 Гц для выбранных ROI.
- Примените преобразование Гильберта для получения мгновенной амплитуды139с помощью набора инструментов для обработки сигналов из программного обеспечения на языке программирования (см. Таблицу материалов).
- Корректируйте сигнал с помощью нормализации z-оценки (от −1000 до −10 мс в качестве исходного уровня) и усредняйте по испытаниям по испытуемым.
- Сравните временной профиль каждого тета-полосы ROI между условиями с помощью теста Вилкоксона со знаком ранга (совпавшие пары, 1 с неперекрывающихся окон) и скорректируйте с помощью FDR.
- Корреляция между активностью ЭЭГ и поведенческими показателями
- Нормализуйте исходные временные ряды ROI до базового уровня с помощью z-оценки. Выберите окно от 1 с до 4 с после начала планирования или контроля (в котором наблюдается заметная тета-активность на графиках временных частот).
- Чтобы определить увеличение тета-активности в условиях планирования по сравнению с контрольными условиями, сначала преобразуйте сигнал в частотную область (1-40 Гц) с помощью метода мультиконуса с помощью набора инструментов, такого как набор инструментовChronux 140 для каждого условия и источника в интересующих областях.
- Вычислите среднюю частоту тета-диапазона (4-8 Гц) и рассчитайте две меры тета-мощности: i) разницу между тета-мощностью в течение планового периода (θ планирование) и контрольного периода (θ контроль), обозначаемую как Δ тета, и ii) относительное увеличение тета-активности, выраженное в виде отношения Δ тета (Δ θ) и тета-активности в течение контрольного периода (θ контроль), как в Domic-Siede et al.54:
 (1)
(1)

- Рассчитаем два поведенческих параметра: iii) Δ планирование LISAS, вычитая управление LISAS из планирования LISAS, и iv) Δ выполнение планирования LISAS, вычитая выполнение управления LISAS из выполнения планирования LISAS, как в Domic-Siede et al.54:


- Выполните рокорреляцию Спирмена с использованием рассчитанных электрофизиологических и поведенческих параметров, а затем скорректировайте ее с помощью FDR.
- Анализ движений глаз: Чтобы контролировать потенциальные различия в движениях глаз для каждого состояния, которые могут привести к различной динамике колебаний, выполните следующий анализ:
- Определите амплитуду саккады и пиковую скорость саккады на протяжении всего испытания и от 0 с до 3,75 с в условиях планирования и контроля.
- Сравните результаты, используя либо тест Вилкоксона со знаком, либо t-критерий согласованной пары, в зависимости от того, что подходит.
ПРИМЕЧАНИЕ: Набор инструментов, подобный описанному в Dimigen et al.123, может быть полезен. - Рассчитайте и оцените когерентность между мощностью ЭЭГ Фурье на одном фронтальном электроде (например, Fz или усредненном фронтальном электроде ROI) и скоростью саккады, как описано в Sato and Yamaguchi141.
- Используйте тест Вилкоксона со знаком ранга для сравнения значений степени когерентности-скорости саккады первых 4 с каждого испытания между двумя условиями.
Результаты
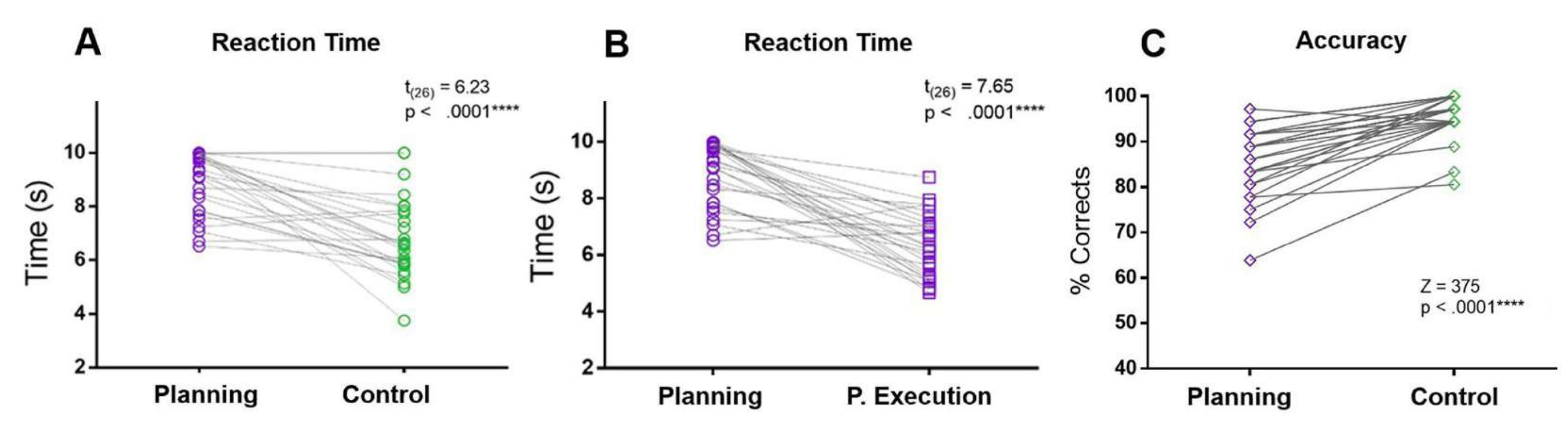
В настоящем протоколе РТ планового периода сравнивался с РТ контрольного периода и планового периода исполнения. Кроме того, по сравнению с контрольным условием, участники допустили больше ошибок и имели более низкую точность в течение периода планирования (рис. 5).

Рисунок 5: Время реакции и точность для задачи планирования. Сравнение времени реакции (А) в период планирования (фиолетовые круги) и контрольного периода (зеленые круги) с использованием t-критерия согласованной пары. (B) Сравнение времени реакции в период планирования (фиолетовые круги) и в период выполнения планирования (фиолетовые квадраты) с использованием t-критерия с подобранной парой. (C) Сравнение коэффициента точности в условиях планирования (фиолетовые ромбы) и контрольных условиях (зеленые ромбы) с использованием критерия Вилкоксона со знаком ранга. Этот рисунок был изменен по Domic-Siede et al.54. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этой цифры.
{kind=link}
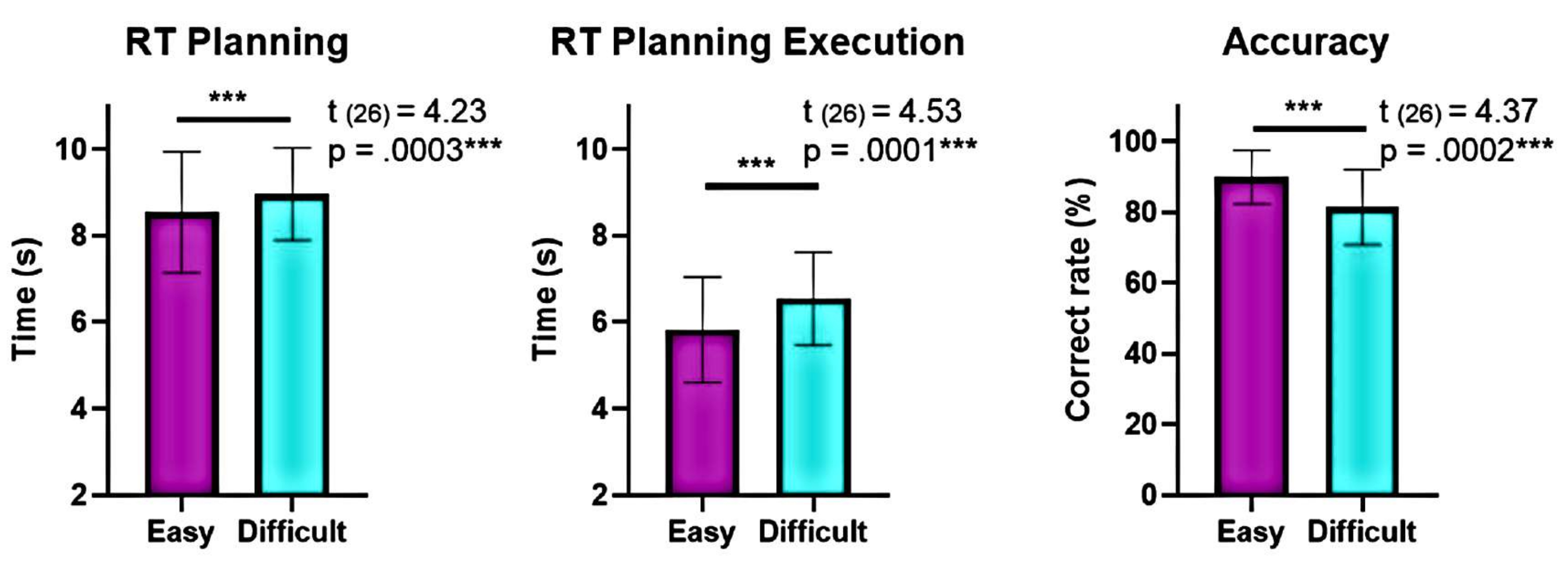
Более того, анализ уровней сложности планирования показал значительные различия в точности и времени реакции (RT) между «сложным» и «легким» уровнями во время планирования и выполнения (рис. 6). «Сложный» уровень имел более длинные RT и меньшую точность. Эти результаты свидетельствуют о том, что разделение испытаний на основе количества действительных решений может отличить «легкие» испытания от «сложных».

Рисунок 6: Сравнение поведенческой производительности на различных уровнях сложности. Значимые различия в поведенческих показателях на «легком» и «сложном» уровнях сложности были выявлены с помощью t-критерия согласованной пары. Более низкое время реакции (RT) наблюдалось во время планирования и выполнения для «легкого» уровня по сравнению с «сложным», а точность была выше для «легкого» уровня. Полосы погрешностей представляют собой SEM (стандартную погрешность среднего значения). Этот рисунок был изменен по Domic-Siede et al.54. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этой цифры.
{kind=link}
Эти результаты показали, что когда компонент планирования был успешно удален из контрольного условия (с помощью манипуляций с инструкциями), задача планирования стала когнитивно более сложной, требовательной, сложной и трудоемкой. Следовательно, нейронные корреляты, индуцированные задачами, могут быть сравнены друг с другом.
Для анализа тета-активности лобной срединной линии во время планирования средняя полоса тета-частот во время планирования для электрода Fz сравнивалась с контрольным периодом, и было обнаружено значительное увеличение частоты тета-полосы во время планирования (рис. 7).

Рисунок 7: Фронтальная срединная тета-активность во время когнитивного планирования. (A) Топографические карты, представляющие мощность тета-полосы у всех испытуемых, нормализованные по z-баллам во время задачи планирования (слева), контрольной задачи (в центре) и эффекта планирования (справа). Во время когнитивного планирования у испытуемых наблюдалось увеличение тета-активности лобной средней линии. Цветная полоса показывает z-значения в диапазоне от −0,5 до 1,5. (B) График скрипки, показывающий минимальные, квартили, медианные и максимальные значения z-оценки тета-активности у испытуемых во время планирования (фиолетовый) по сравнению с контрольным периодом (зеленый) для электродов Fz (слева), Pz (посередине) и Oz (справа) с использованием t-критерия согласованной пары. Этот рисунок был изменен по Domic-Siede et al.54. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этой цифры.
{kind=link}
Кроме того, для оценки временной динамики наблюдаемой фронтальной тета-активности были сформулированы топографические карты, соответствующие конкретным временным точкам мощности тета-диапазона (750 мс, 1750 мс, 2750 мс и 3750 мс) (рис. 8A). Кроме того, по сравнению с контрольным периодом, частотно-временной анализ продемонстрировал значительное, прогрессивное и устойчивое увеличение тета-активности, начиная с 1 с после начала периода планирования (рис. 8B).

Рисунок 8: Лобная срединная тета-временная динамика. (A) Топографические временные срезы тета-активности. Наблюдалось прогрессивное увеличение активности лобной срединной тета-линии с течением времени в течение реализации планирования (период планирования). Цветная полоса указывает единицы z-оценки (от −0,5 до 2,2). (B) Графики частот по времени для планового периода (вверху), контрольного периода (посередине) и эффекта планирования, рассчитанные путем вычитания контрольного периода из планового периода (внизу). Незначащие пиксели, определенные с помощью непараметрического кластерного теста перестановок для парных выборок, отображаются светлее на графике эффекта планирования. Цветная полоса указывает единицы z-оценки (от −4 до 4). Этот рисунок был изменен по Domic-Siede et al.54. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этой цифры.
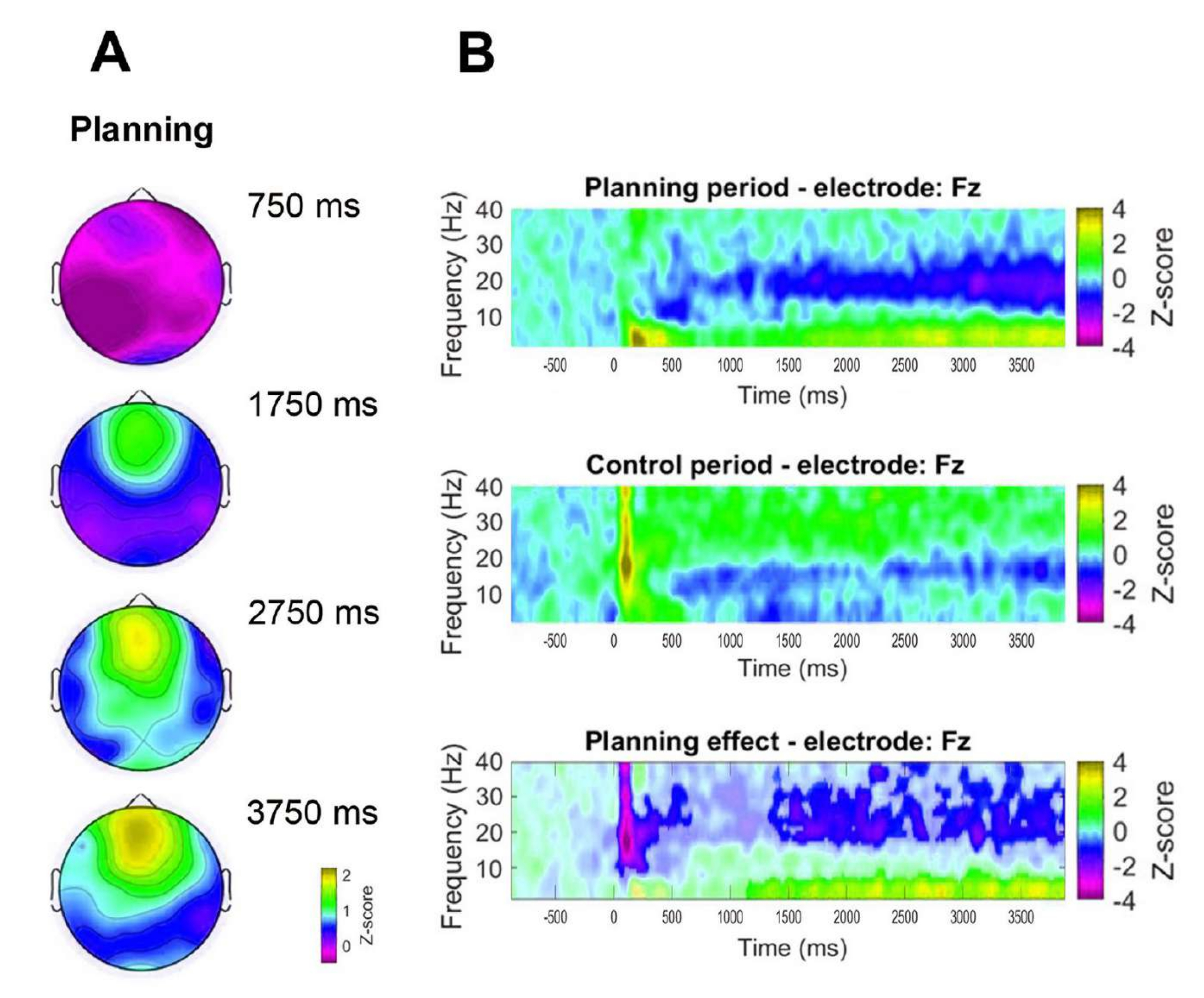{kind=link}
Для реконструкции источника тета-активности был визуализирован шаблон модели мозга и сравнен между условиями, и это указывало на то, что тета-активность возникает в областях префронтальной коры (лобная верхняя кора, FS; передняя поясная кора, ACC; и среднепоясная кора, MCC), а также на наличие существенных различий между условиями (в двусторонней SF, двусторонний ACC и двусторонний MCC) (рисунок 9), при этом более высокая тета-активность наблюдалась в период планирования (рисунок 9).

Рисунок 9: Реконструкция источника. Алгоритм sLORETA был использован для оценки тета-активности из различных источников мозга. Тета-активность была отфильтрована полосой пропускания 4-8 Гц, нормализована z-оценкой, скорректирована по исходному уровню, усреднена между 1 с или 4 с после планирования или начала контроля, соответственно, и сравнивалась между условиями. Значительное увеличение тета-активности было обнаружено в двусторонней передней верхней области, двусторонней передней поясной коре и двусторонней средней поясной коре. На рисунке показаны значимые t-значения из теста перестановки. Сокращения: FS = лобовой вышестоящий ; ACC = передняя поясная кора; MCC = средняя поясная извилина коры. Этот рисунок был изменен по Domic-Siede et al.54. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этой цифры.
{kind=link}
После этого временной профиль тета-изменений с течением времени для каждого источника был оценен путем вычисления преобразования Гильберта, а затем мы сравнили мгновенную амплитуду тета-активности между условиями. Мы обнаружили, что левый лобопольный, двусторонний АКК и двусторонние источники МКК демонстрировали более высокую тета-активность после начала планового периода (рис. 10). Эти результаты позволяют предположить, что наша экспериментальная парадигма, требующая когнитивного планирования, индуцировала тета-активность, возникающую в областях ПФК.

Рисунок 10: Временной профиль тета-активности источников PFC. Мгновенная амплитуда, рассчитанная с помощью преобразования Гильберта, была применена к первому компоненту разложения PCA для каждого фронтального источника, и оба условия и исходная линия нормализованы к z-оценке, чтобы показать фронтальную тета-активность с течением времени. Серыми затененными областями видны значительные различия, определенные с помощью неперекрывающегося движущегося окна с шагом 1 с (критерий Вилкоксона со знаком ранга), скорректированный FDR. Заштрихованные области представляют 95% доверительных интервалов. Левая область FP, двусторонний ACC и двусторонний MCC показали увеличение тета-активности после начала планирования. Сокращения: ACC = передняя поясная кора; MCC = средняя поясная извилина коры. Условия планирования показаны фиолетовым цветом. Условие управления отображается красным цветом. Этот рисунок был изменен по Domic-Siede et al.54. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этой цифры.
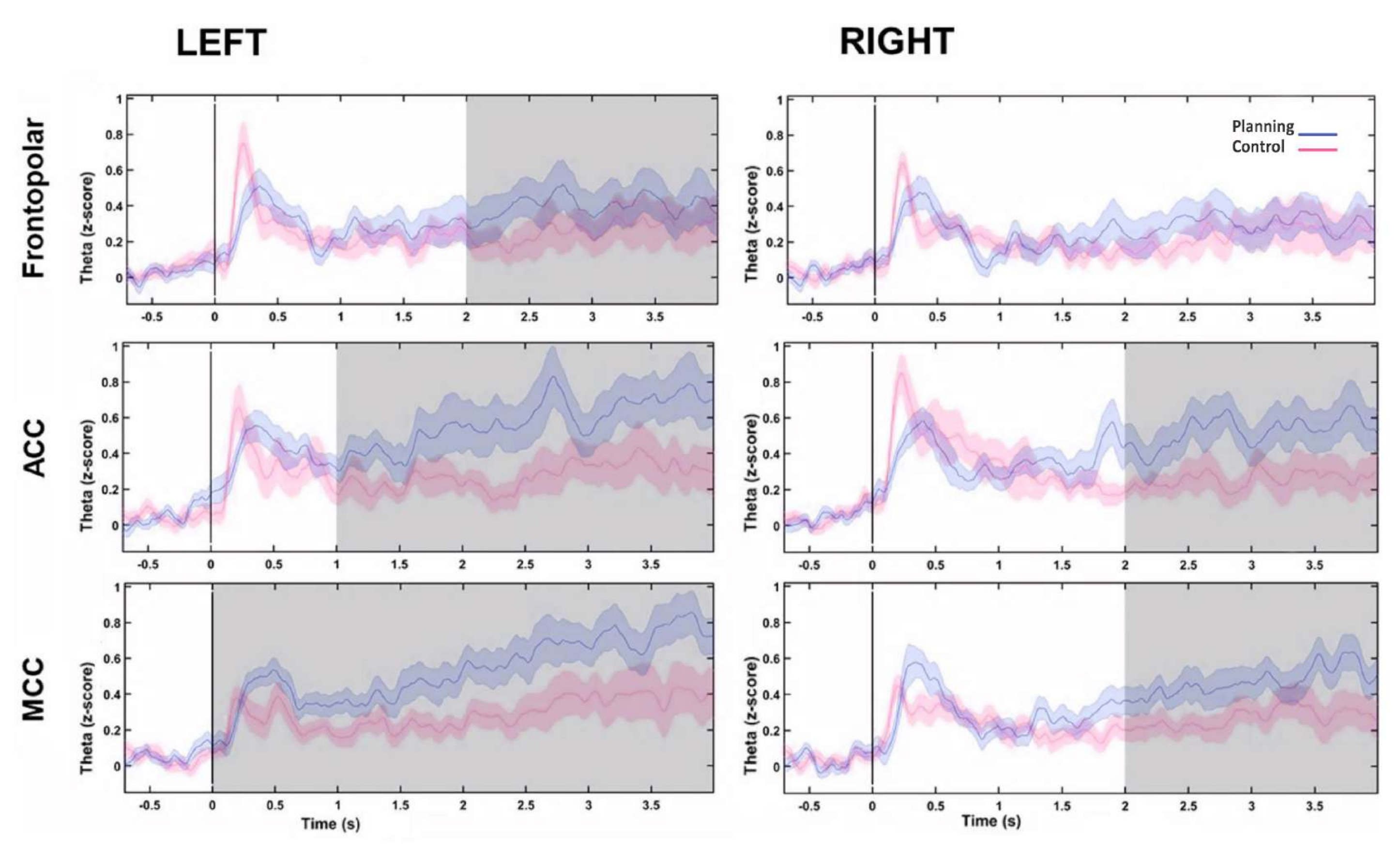{kind=link}
Кроме того, мы стремились изучить вариацию спектральных характеристик во время планирования в зависимости от уровня сложности, на что указывают поведенческие результаты. Следует отметить, что существенное расхождение было обнаружено только в левом ППЦ в альфа-диапазоне. Это подтверждает идею о том, что наша задача планирования оценивает внутренние аспекты планирования через изменения тета-колебаний в большей степени, чем общие когнитивные требования (усилия), обычно встречающиеся в задачах когнитивного контроля (рис. 11).

Рисунок 11: ЭЭГ по уровням сложности планирования. Частотно-временные диаграммы ROI показали значительный положительный кластер в альфа-диапазоне исключительно в левой передней поясной коре (ACC) для «сложного» уровня. Незначащие пиксели, определенные с помощью непараметрического теста перестановок на основе кластеров для парных выборок, отображаются на графике более светлым оттенком, а цветная полоса указывает единицы z-оценки от −3 до 3. Этот рисунок был изменен по Domic-Siede et al.54. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этой цифры.
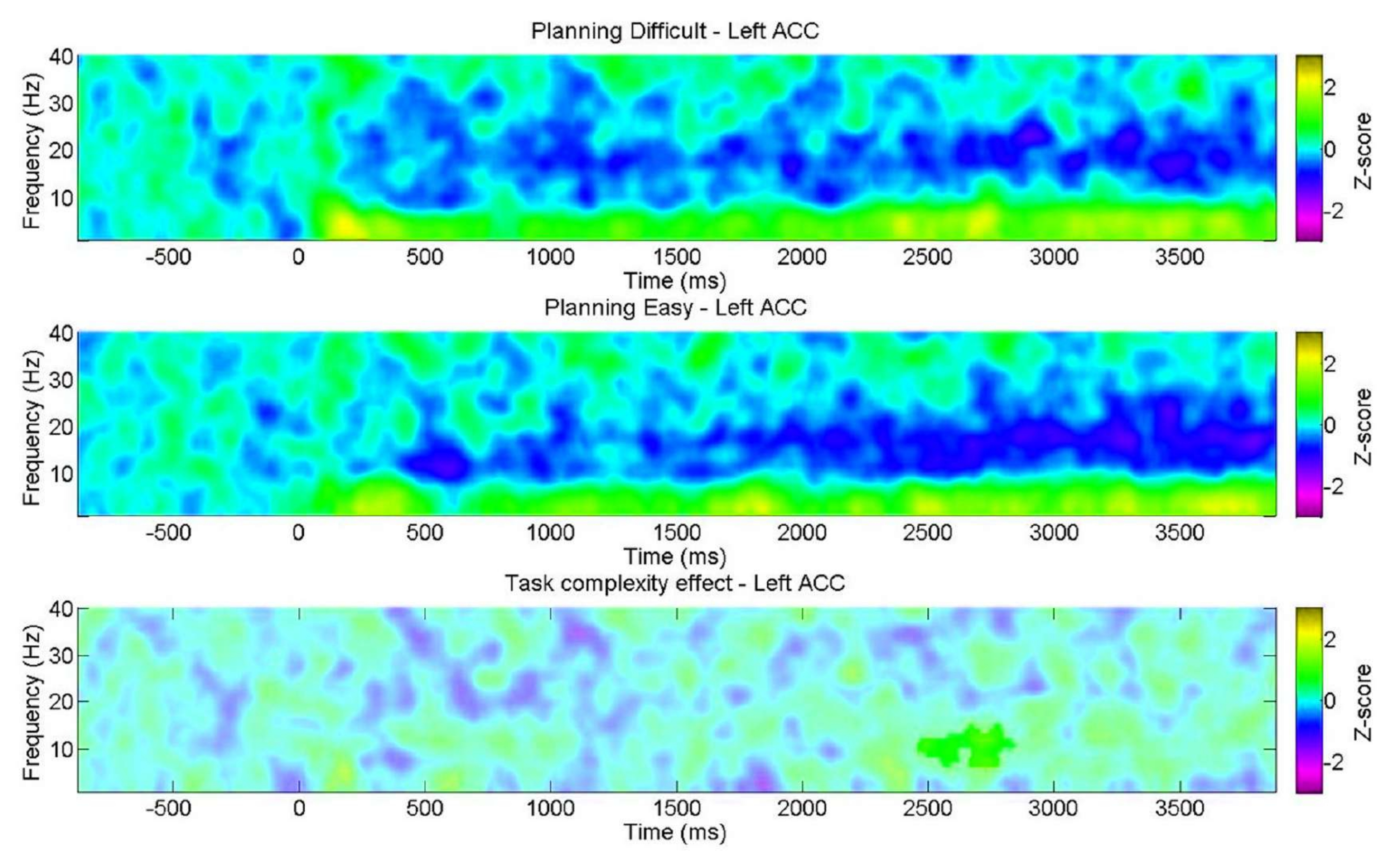{kind=link}
Когда была установлена корреляция между тета-активностью и поведенческими показателями, наблюдалась отрицательная корреляция; в частности, по мере увеличения тета-активности в левой лобно-полярной области в течение периода планирования, оценка выполнения планирования LISAS снижалась (рис. 12). Этот шаблон может отражать то, что левая область FP может быть необходима во время разработки плана для успешного выполнения плана впоследствии, и предполагает роль тета-активности.

Рисунок 12: Тета-активность и поведенческие показатели. Корреляция Спирмена между тета-активностью левой лобнополярной коры и выполнением планирования Δ LISAS показала значимую отрицательную корреляцию. Этот рисунок был изменен по Domic-Siede et al.54. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этой цифры.
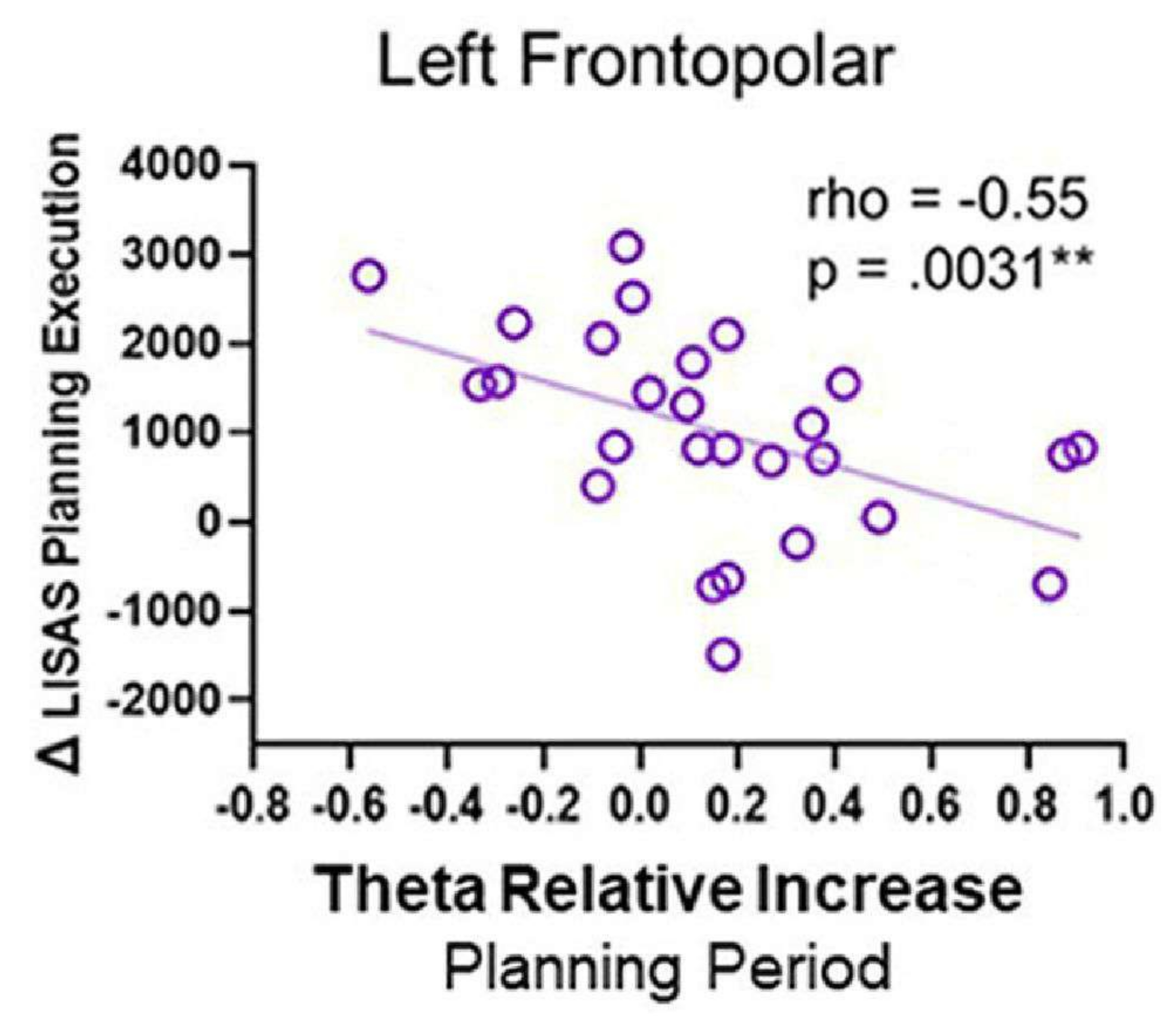{kind=link}
Кроме того, различные когнитивные потребности и цели, вызванные каждым состоянием, могли вызывать контрастные движения глаз между условиями планирования и контроля, что приводило кразличным моделям колебательной активности. Чтобы решить вышеуказанную проблему, мы проанализировали данные по одному субъекту и одному испытанию на различных уровнях. Примечательно, что временные ряды канала Fz и временная динамика тета-активности, по-видимому, не имеют связи с частотой саккад с течением времени (рис. 13A).

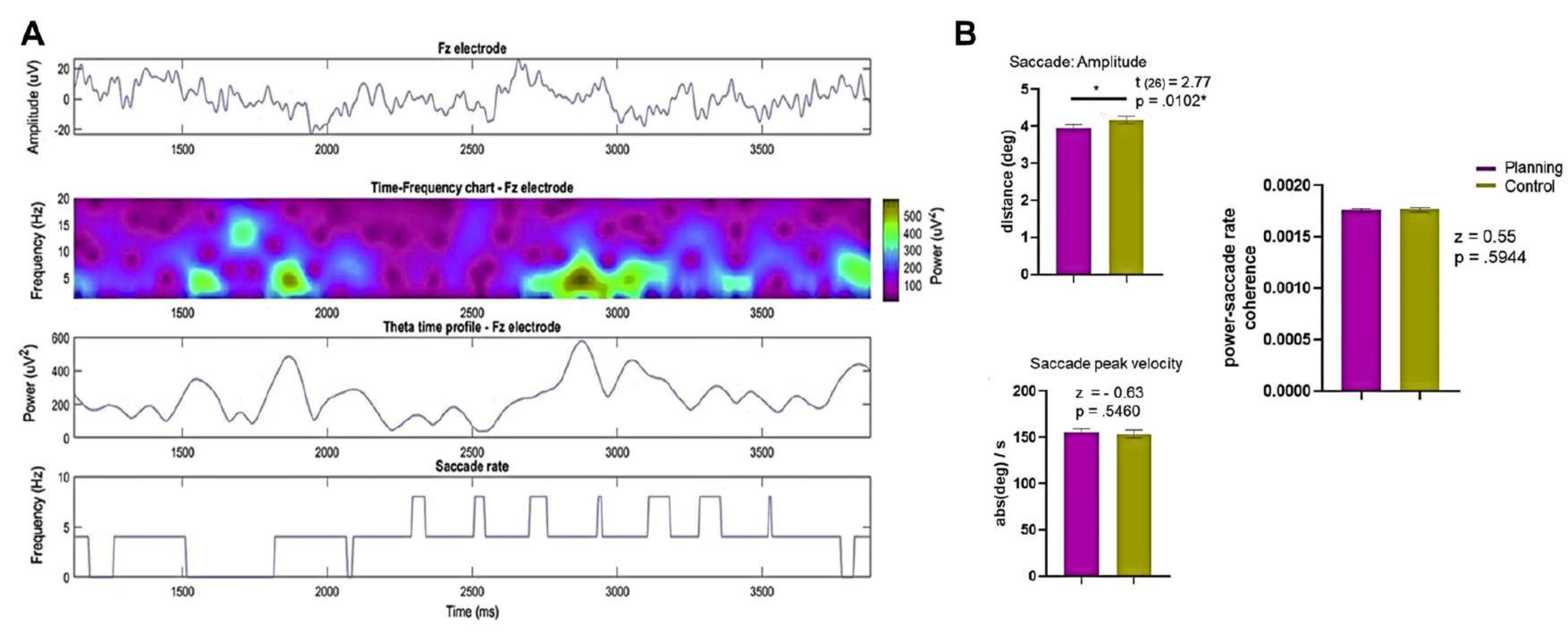
Рисунок 13: Результаты записи ЭЭГ и движений глаз. (A) В строках представлены ЭЭГ (вверху), временная шкала (вверху посередине), профиль времени тета-времени от электрода Fz (внизу) и частота саккады субъекта 8 и испытания 9 (внизу) в условиях планирования. (B) Показано сравнение в критерее Вилкоксона амплитуды саккады, пиковой скорости саккады и когерентности мощности и скорости саккады между условиями, которое выявило значительные статистические различия в амплитуде саккады между условиями планирования и контроля. SEM представлен полосами погрешностей. Этот рисунок был изменен по Domic-Siede et al.54. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этой цифры.
{kind=link}
Затем мы получили амплитуду саккады и пиковую скорость из всего испытания и от 0 с до 3,75 с для сравнения (рис. 13B). Мы обнаружили, что амплитуда саккады была больше в контрольном состоянии. Тем не менее, не было обнаружено статистически значимых различий между условиями в индексе когерентности между тета-мощностью Фурье на электроде Fz и скоростью саккады (рис. 10B), что указывает на то, что любая потенциальная связь между саккадами и тета-активностью была последовательной в разных условиях.
В совокупности эти результаты подтверждают, что описанный экспериментальный протокол подходит для изучения когнитивного планирования как функции когнитивного контроля.
Обсуждение
Описанный здесь протокол предлагает инновационный подход к оценке когнитивного планирования и когнитивного контроля во время задачи нового и экологического планирования в сочетании с соответствующими и дополнительными поведенческими и физиологическими измерениями, такими как колебательная и зрачковая динамика. Во время наших экспериментов активность ЭЭГ записывалась, пока участники выполняли задание на планирование, в котором участникам было поручено сначала разработать, а затем выполнить план. Контрольное условие, которое включало в себя оценку предварительно нарисованного пути на карте зоопарка, было установлено для устранения когнитивного аспекта планирования при сохранении аналогичной обстановки и структуры. Этот подход позволяет оценить, приводит ли когнитивное планирование как функция когнитивного контроля к генерации фронтальной тета-активности из областей ПФК и связаны ли различные источники тета-осцилляций ПФК с различными аспектами эффективности планирования. Еще одним аспектом, который может быть оценен с помощью этого протокола, является дифференциация различных когнитивных процессов, задействованных на этапах планирования, таких как разработка плана, выполнение плана, достижение плана и обработка обратной связи. Мы обнаружили, что планирование индуцировало каноническую фронтальную тета-активность, связанную с когнитивным контролем, что способствовало эффективному достижению цели. Эти результаты подкрепляют интерес к данному протоколу.
Несмотря на обширное развитие когнитивной нейробиологии, большинство экспериментов по нейровизуализации изучают изолированные когнитивные функции с использованием искусственных задач в сенсомоторно-деприструтной среде и чрезмерно упрощенных стимулов для контроля путаницы. Проблема заключается в том, что эти эксперименты могут оказаться не в состоянии выявить реальные механизмы мозга, задействованные при реализации когнитивной функции в повседневной ситуации (во время экологических ситуаций)138,140. В частности, способности к формулировке целей, планированию и эффективному выполнению планов трудно оценить, поскольку они требуют различных когнитивных функций (рабочей памяти, ингибиторного контроля, когнитивной гибкости и т. д.)104,144. Таким образом, разработка экологической поведенческой задачи поощряется и предлагается на основе современных тенденций в когнитивной нейронауке 142,143,145,146.
Наша задача по планированию, несмотря на то, что она выполнялась в лабораторных условиях (в комнате с стимулами, отображаемыми на экране), была сделана увлекательной и интерактивной для участников за счет использования значимых стимулов и целей, с которыми они могли взаимодействовать на экране. Кроме того, задание требует от участников участия в реальной ситуации — планирования пути к посещению различных локаций. Чтобы иметь экологическую задачу, парадигма должна побуждать испытуемых выполнять определенное поведение или когнитивную функцию таким же образом, как они должны были бы делать вповседневной жизни. Для разработки дизайна экологической задачи используемая здесь задача планирования включает в себя планирование маршрута для посещения различных мест в несколько этапов56. На первом этапе участники создают план, следя за тем, чтобы он соответствовал набору правил. Второй этап включает в себя поддержание плана в рабочей памяти, в то время как третий этап включает в себя выполнение плана и контроль за его соблюдением правил. Эти стадии представляют собой различные фазы планирования и координации других исполнительных функций, таких как когнитивная гибкость, ингибиторный контроль и рабочая память. Для того, чтобы иметь обоснованный экологический когнитивный дизайн задачи, задача должна быть способна выявлять специфические когнитивные нарушения у пациентов с психическими или когнитивными расстройствами, которые испытывают трудности с выполнением этой специфической когнитивной функции в своейповседневной жизни. Это может быть достигнуто за счет будущих исследований с использованием этого протокола.
Поведенческие результаты, полученные с помощью этого протокола, были согласованы с экспериментальными прогнозами. Значительная разница в поведении наблюдалась при удалении компонента планирования из задачи управления для формирования контрольного условия, что облегчало дальнейшие сравнения. Было обнаружено, что условия планирования являются когнитивно более требовательными, чем контрольные условия, о чем свидетельствуют такие параметры, как время реакции и точность. Это может отражать повышенное участие высоких когнитивных функций в реализации планирования 23,55,56,57,148,149,150.
Учитывая, что условие контроля включает в себя менее сложное познание, о чем свидетельствует более быстрое время реакции, лучшие результаты и другие необходимые когнитивные процессы (оценка правил), возможной альтернативой модификации является использование уровней сложности, присутствующих в задаче планирования, манипулирование ими и параметрический анализ функции планирования в соответствии с различными уровнями сложности (например, увеличение количества испытаний, а также создание сложных, средних и легких условий испытаний). Тем не менее, результаты нашего протокола показали, что, хотя можно было различить «легкие» и «сложные» испытания на основе поведенческих показателей, не было обнаружено различий в электрофизиологических показателях. Это говорит о том, что результаты в нашем протоколе более точно отражают внутренние характеристики функции планирования, а не более широкие аспекты когнитивного контроля, такие как внимание, умственное усилие, уровень сложности или высокий уровень когнитивныхпотребностей. Тем не менее, дальнейшие исследования могут рассмотреть другие типы условий контроля, такие как следование по отмеченной тропе, которая проходит через четыре места обитания животных, а также запоминание порядка последовательности. Таким образом, уровень сложности можно было бы лучше контролировать, а планирование можно было бы отличить от рабочей памяти, но одним из возможных недостатков этого является усталость, потому что испытуемым пришлось бы выполнять две очень сложные задачи.
Несколько исследований связывают различные параметры движения глаз с конкретными когнитивными событиями. С одной стороны, некоторые исследования обнаружили корреляцию между тета-колебаниями и диаметром зрачка во время выполнения когнитивных задач, что предполагает связь между этими двумя показателями когнитивной функции. Например, Lin et al.152 обнаружили корреляцию между среднелобной тета-активностью и изменениями в размере зрачка, отражающую различную степень субъективного конфликта. Их результаты показывают, что эти сигналы представляют собой обработку конфликта, повышенное внимание и гибкие поведенческие реакции. Следовательно, взаимосвязь между тета-активностью в середине лоба и реакциями зрачков, по-видимому, играет роль в взвешивании затрат и выгод в процессе принятия решений. В другом исследовании Yu et al.153 изучали, как нейрофизиологическое влияние времени на выполнение задачи влияет на когнитивный контроль через задачу на рабочую память, которая модулирует контрольные тормозные реакции. Они изучили взаимосвязь между данными о диаметре зрачка и лобной тета-активностью и показали, что по мере увеличения длительности задачи производительность снижалась, и это сопровождалось снижением модуляции расширения зрачков и лобной тета-активности. В начале задания они обнаружили сильную корреляцию между вовлеченностью в задачу, тета-активностью и когнитивным контролем, на что указывает модуляция расширения зрачка, в основном для сложных задач, требующих высокой рабочей памяти и ингибиторного контроля. Однако к концу эта связь рассеялась, сигнализируя о разрыве связи между затраченными усилиями и когнитивным контролем, используемым для выполнения задачи, что является отличительной чертой префронтальногоэффекта времени на выполнение задачи. С другой стороны, в других исследованиях изучались саккады и осцилляции. Например, Nakatani et al.154 показали, что в задаче восприятия амплитуда альфа-диапазона от затылочных областей предсказывает мигание и саккадные эффекты. Более того, Velasques et al.155 показали, что во время выполнения задачи на просаккадическое внимание амплитуда саккады была связана с изменениями фронтального гамма-излучения. Кроме того, Bodala et al.156 обнаружили, что снижение лобной средней теты сопровождалось снижением устойчивого внимания, а также амплитуды и скорости саккад. Эти результаты свидетельствуют о том, что движения глаз, особенно саккады, могут отражать когнитивные процессы, а не просто способствовать возникновению фонового шума в сигналах ЭЭГ. В данном исследовании мы улучшили устранение артефактов, связанных с движением глаз, с помощью алгоритма ICA с критерием дисперсии саккады и фиксации126. Этот критерий улучшает удаление артефактов для заданий свободного просмотра157. В нашем исследовании не наблюдалось различий в пиковой скорости саккады и когерентности между тета-мощностью и скоростью саккады в разных условиях. Тем не менее, для решения этих вопросов необходимы дополнительные исследования.
Важным шагом в использовании этого протокола является постоянная калибровка айтрекера во время эксперимента, так как потеря данных о взгляде с камеры может привести к ошибкам, которые загрязнят задачу и затруднят получение точных ответов. Поэтому крайне важно проводить калибровку как можно чаще. Тем не менее, существует компромисс между количеством испытаний с откалиброванным айтрекером и продолжительностью эксперимента. В нашем исследовании мы решили проводить калибровку каждые пять испытаний.
В будущем следует провести исследования, изучающие взаимосвязь между тета-колебаниями и диаметром зрачка во время этой задачи планирования. Планирование является важнейшим аспектом исполнительного контроля, который требует распределения ресурсов внимания и координации многочисленных когнитивных процессов. Понимание взаимосвязи между тета-колебаниями и диаметром зрачка во время выполнения задач планирования может дать ценную информацию о нейронных механизмах, лежащих в основе исполнительного контроля, и о том, как они изменяются с течением времени. Кроме того, такие исследования могут привести к более глубокому пониманию того, как изменения в когнитивных функциях, такие как усталость или потеря внимания, влияют на выполнение задач планирования и способность эффективно распределять ресурсы. Эта информация может иметь важное значение для разработки вмешательств, направленных на повышение эффективности планирования, таких как программы когнитивной тренировки или лечения таких состояний, как синдром дефицита внимания и гиперактивности (СДВГ), среди прочих.
Предыдущие исследования показали, что ПФК играет решающую роль в когнитивном планировании, что подтверждают наши результаты. Эти результаты демонстрируют, что когнитивное планирование индуцирует активность FMθ в ПФК, особенно в передней поясной коре, средней поясной коре и верхних лобных областях54. Эти результаты согласуются с предыдущими исследованиями исполнительных функций. Существуют существенные доказательства в поддержку идеи о том, что активность FMθ действует как нисходящий процесс, инициирующий контроль и облегчающий коммуникацию между областями мозга во время выполнениясложных задач. Несмотря на то, что лишь в нескольких исследованиях изучалась временная динамика активности FMθ, связанная с когнитивным контролем, существует широко распространенное мнение о том, что временной профиль FMθ может предоставить информацию о различных аспектах когнитивного контроля и вовлечении отдельных областей ПФК. Использование нашего протокола для оценки когнитивного планирования позволило нам охарактеризовать временной профиль активности FMθ во время планирования. В частности, активность FMθ в условиях планирования показала прогрессирующее увеличение. При реализации данного протокола впервые было продемонстрировано, что FMθ присутствует и при реализации планирования, как и в других когнитивных функциях более высокого порядка, а его временная динамика может служить индикатором когнитивного контроля.
Наши результаты и протокол имеют потенциальное применение в области нейробиологии, в том числе для улучшения виртуальных нейропсихологических оценок и лечения нейропсихиатрических расстройств с сопутствующими проблемами когнитивного планирования, такими как депрессия и синдром дефицита внимания и гиперактивности. Например, оценка может включать в себя изучение различных паттернов ошибок на уровне поведенческих показателей, различных колебательных паттернов на электрофизиологическом уровне и различных движений глаз. Кроме того, результаты этой работы могут послужить основой для разработки интерфейсов мозг-компьютер и программ когнитивной тренировки, направленных на улучшение способностей к когнитивному планированию.
Настоящий протокол может внести новый вклад в понимание нейронных механизмов, лежащих в основе неуловимой функции когнитивного контроля когнитивного планирования в нейротипичных и нейропсихиатрических популяциях. Более того, наша поведенческая парадигма может предложить понимание нейробиологии когнитивного контроля и планирования через изучение электрофизиологических, пупиллометрических и поведенческих измерений с практической задачей планирования, которая изучает внутренние аспекты планирования, а не общие когнитивные требования, обычно присутствующие в задачах когнитивного контроля, что отражается в изменениях тета-осцилляций.
Раскрытие информации
Авторам нечего раскрывать.
Благодарности
Это исследование было финансово поддержано программой докторских стипендий Becas de Doctorado Nacional año 2015 от ANID 21150295, регулярным грантом FONDECYT 1180932, регулярным грантом FONDECYT 1230383, грантом FONDECYT de Iniciación 11220009, грантом постдока Университета О'Хиггинса и Институтом Университета Франции (IUF). Мы хотим поблагодарить профессора Пабло Биллеке за его отзыв о дизайне парадигмы. Мы благодарим профессора Эухенио Родригеса за любезно предоставленные им коды частотно-временного анализа. Наконец, мы благодарим Милана Домича, Висенте Меделя, Жозефину Инен, Андреа Санчеса, Гонсало Бонкомпте, Каталину Фабар и Даниэлу Сантандер за их отзывы.
Материалы
| Name | Company | Catalog Number | Comments |
| EEG System | Biosemi | ActiveTwo Base system, 64 channels | |
| Eye-tracker System | Eyelink SR Research | EyeLink 1000 Plus Core Unit, High-speed camera, Host PC/Monitor, | |
| CPU display | Intel | Hard drive 221 GB, Processor Intel Core i7-4790 3.60 Hz, OS Windows 7, 4GB RAM | |
| CPU EEG | Intel | Hard drive 223 GB, Processor Intel Core i7-4790 3.60 Hz, OS Windows 7, 4GB RAM | |
| Monitor | ASUS | ASUS VG248QE 24" LCD monitor | |
| Joytsick | Logitech | Model F310 | |
| Luxmeter | Focket | LCD screen (0-200.000 lux) model Liebe WH LX1330B | |
| Statistics software | GraphPad Prism | GraphPad Prism version 8 for Windows | |
| MATLAB Programming Software | The MathWorks | MATLAB R2014a and R2018b | |
| SVG tool Inkscape | Inkscape Project | vector graphic editor software | |
| Presentation Software | Neurobehavioral Systems | stimulus delivery and experiment control program for neuroscience |
Ссылки
- Siegel, M., Donner, T. H., Engel, A. K. Spectral fingerprints of large-scale neuronal interactions. Nature Reviews Neuroscience. 13 (2), 121-134 (2012).
- Fries, P. A mechanism for cognitive dynamics: Neuronal communication through neuronal coherence. Trends in Cognitive Sciences. 9 (10), 474-480 (2005).
- Fries, P. Rhythms for cognition: Communication through coherence. Neuron. 88 (1), 220-235 (2015).
- Thut, G., Miniussi, C., Gross, J. The functional importance of rhythmic activity in the brain. Current Biology. 22 (16), 658-663 (2012).
- Fröhlich, F., McCormick, D. A. Endogenous electric fields may guide neocortical network activity. Neuron. 67 (1), 129-143 (2010).
- Spaak, E., de Lange, F. P., Jensen, O. Local entrainment of alpha oscillations by visual stimuli causes cyclic modulation of perception. Journal of Neuroscience. 34 (10), 3536-3544 (2014).
- Fiebelkorn, I. C., Saalmann, Y. B., Kastner, S. Rhythmic sampling within and between objects despite sustained attention at a cued location. Current Biology. 23 (24), 2553-2558 (2013).
- Landau, A. N., Fries, P. Attention samples stimuli rhythmically. Current Biology. 22 (11), 1000-1004 (2012).
- Song, K., Meng, M., Lin, C., Zhou, K., Luo, H. Behavioral oscillations in attention: Rhythmic α pulses mediated through θ band. Journal of Neuroscience. 34 (14), 4837-4844 (2014).
- Wyart, V., Nobre, A. C., Summerfield, C. Dissociable prior influences of signal probability and relevance on visual contrast sensitivity. Proceedings of the National Academy of Sciences of the United States of America. 109 (9), 3593-3598 (2012).
- Leszczyński, M., Fell, J., Axmacher, N. Rhythmic working memory activation in the human hippocampus. Cell Reports. 13 (6), 1272-1282 (2015).
- Onton, J., Delorme, A., Makeig, S. Frontal midline EEG dynamics during working memory. NeuroImage. 27 (2), 341-356 (2005).
- Cavanagh, J. F., Frank, M. J. Frontal theta as a mechanism for cognitive control. Trends in Cognitive Sciences. 18 (8), 414-421 (2014).
- Siegel, M., Buschman, T. J., Miller, E. K. Cortical information flow during flexible sensorimotor decisions. Science. 348 (6241), 1352-1355 (2015).
- Weisz, N., et al. Prestimulus oscillatory power and connectivity patterns predispose conscious somatosensory perception. Proceedings of the National Academy of Sciences of the United States of America. 111 (4), 417-425 (2014).
- Buzsáki, G., Draguhn, A. Neuronal oscillations in cortical networks. Science. 304 (5679), 1926-1929 (2004).
- Cohen, M. X. A neural microcircuit for cognitive conflict detection and signaling. Trends in Neurosciences. 37 (9), 480-490 (2014).
- Yuste, R. From the neuron doctrine to neural networks. Nature Reviews Neuroscience. 16 (8), 487-497 (2015).
- Helfrich, R. F., Knight, R. T. Oscillatory dynamics of prefrontal cognitive control. Trends in Cognitive Sciences. 20 (12), 916-930 (2016).
- Miller, E. K., Cohen, J. D. An integrative theory of prefrontal cortex function. Annual Review of Neurosciences. 24 (1), 167-202 (2001).
- von Nicolai, C., et al. Corticostriatal coordination through coherent phase-amplitude coupling. Journal of Neuroscience. 34 (17), 5938-5948 (2014).
- Sweeney-Reed, C. M., et al. Thalamic theta phase alignment predicts human memory formation and anterior thalamic cross-frequency coupling. ELife. 4, 07578 (2015).
- Voytek, B., et al. Oscillatory dynamics coordinating human frontal networks in support of goal maintenance. Nature Neuroscience. 18 (9), 1318-1324 (2015).
- Raghavachari, S., et al. Theta oscillations in human cortex during a working-memory task: Evidence for local generators. Journal of Neurophysiology. 95 (3), 1630-1638 (2006).
- Jacobs, J., Hwang, G., Curran, T., Kahana, M. J. EEG oscillations and recognition memory: Theta correlates of memory retrieval and decision making. NeuroImage. 32 (2), 978-987 (2006).
- Itthipuripat, S., Wessel, J. R., Aron, A. R. Frontal theta is a signature of successful working memory manipulation. Experimental Brain Research. 224 (2), 255-262 (2013).
- Cavanagh, J. F., Zambrano-Vazquez, L., Allen, J. J. B. Theta lingua franca: A common mid-frontal substrate for action monitoring processes. Psychophysiology. 49 (2), 220-238 (2012).
- Mas-Herrero, E., Marco-Pallarés, J. Frontal theta oscillatory activity is a common mechanism for the computation of unexpected outcomes and learning rate. Journal of Cognitive Neuroscience. 26 (3), 447-458 (2014).
- Folstein, J. R., Van Petten, C. Influence of cognitive control and mismatch on the N2 component of the ERP: A review. Psychophysiology. 45 (1), 152-170 (2008).
- Cohen, M. X., Donner, T. H. Midfrontal conflict-related theta-band power reflects neural oscillations that predict behavior. Journal of Neurophysiology. 110 (12), 2752-2263 (2013).
- Walsh, M. M., Anderson, J. R. Modulation of the feedback-related negativity by instruction and experience. Proceedings of the National Academy of Sciences of the United States of America. 108 (47), 19048-19053 (2011).
- Luu, P., Tucker, D. M., Makeig, S. Frontal midline theta and the error-related negativity: Neurophysiological mechanisms of action regulation. Clinical Neurophysiology. 115 (8), 1821-1835 (2004).
- Hanslmayr, S., et al. The electrophysiological dynamics of interference during the Stroop task. Journal of Cognitive Neuroscience. 20 (2), 215-225 (2008).
- Cavanagh, J. F., Cohen, M. X., Allen, J. J. B. Prelude to and resolution of an error: EEG phase synchrony reveals cognitive control dynamics during action monitoring. Journal of Neuroscience. 29 (1), 98-105 (2009).
- Cohen, M. X., van Gaal, S., Ridderinkhof, K. R., Lamme, V. A. F. Unconscious errors enhance prefrontal-occipital oscillatory synchrony. Frontiers in Human Neuroscience. 3, 54 (2009).
- Cavanagh, J. F., Frank, M. J., Klein, T. J., Allen, J. J. B. Frontal theta links prediction errors to behavioral adaptation in reinforcement learning. NeuroImage. 49 (4), 3198-3209 (2010).
- Cohen, M. X., Cavanagh, J. F. Single-trial regression elucidates the role of prefrontal theta oscillations in response conflict. Frontiers in Psychology. 2, 30 (2011).
- Cohen, M. X., Van Gaal, S. Dynamic interactions between large-scale brain networks predict behavioral adaptation after perceptual errors. Cerebral Cortex. 23 (5), 1061-1072 (2013).
- Nigbur, R., Cohen, M. X., Ridderinkhof, K. R., Stürmer, B. Theta dynamics reveal domain-specific control over stimulus and response conflict. Journal of Cognitive Neuroscience. 24 (5), 1264-1274 (2012).
- van Driel, J., Swart, J. C., Egner, T., Ridderinkhof, K. R., Cohen, M. X. No) time for control: Frontal theta dynamics reveal the cost of temporally guided conflict anticipation. Cognitive, Affective, and Behavioral Neuroscience. 15 (4), 787-807 (2015).
- van de Vijver, I., Ridderinkhof, K. R., Cohen, M. X. Frontal oscillatory dynamics predict feedback learning and action adjustment. Journal of Cognitive Neuroscience. 23 (12), 4106-4121 (2011).
- Narayanan, N. S., Cavanagh, J. F., Frank, M. J., Laubach, M. Common medial frontal mechanisms of adaptive control in humans and rodents. Nature Neuroscience. 16 (12), 1888-1895 (2013).
- Anguera, J. A., et al. Video game training enhances cognitive control in older adults. Nature. 501 (7465), 97-101 (2013).
- Smit, A. S., Eling, P. A. T. M., Hopman, M. T., Coenen, A. M. L. Mental and physical effort affect vigilance differently. International Journal of Psychophysiology. 57 (3), 211-217 (2005).
- Cohen, M. X., Ranganath, C. Reinforcement learning signals predict future decisions. Journal of Neuroscience. 27 (2), 371-378 (2007).
- Gehring, W. J., Goss, B., Coles, M. G. H., Meyer, D. E., Donchin, E. A neural system for error detection and compensation. Psychological Science. 4 (6), 385-390 (1993).
- Yeung, N., Botvinick, M. M., Cohen, J. D. The neural basis of error detection: Conflict monitoring and the error-related negativity. Psychological Review. 111 (4), 931-959 (2004).
- Debener, S., et al. Trial-by-trial coupling of concurrent electroencephalogram and functional magnetic resonance imaging identifies the dynamics of performance monitoring. Journal of Neuroscience. 25 (50), 11730-11737 (2005).
- Hauser, T. U., et al. The feedback-related negativity (FRN) revisited: New insights into the localization, meaning, and network organization. NeuroImage. 84, 159-168 (2014).
- Wang, C., Ulbert, I., Schomer, D. L., Marinkovic, K., Halgren, E. Responses of human anterior cingulate cortex microdomains to error detection, conflict monitoring, stimulus-response mapping, familiarity, and orienting. Journal of Neuroscience. 25 (3), 604-613 (2005).
- Tsujimoto, S., Genovesio, A. Firing variability of frontal pole neurons during a cued strategy task. Journal of Cognitive Neuroscience. 29 (1), 25-36 (2017).
- Womelsdorf, T., Johnston, K., Vinck, M., Everling, S. Theta-activity in anterior cingulate cortex predicts task rules and their adjustments following errors. Proceedings of the National Academy of Sciences of the United States of America. 107 (11), 5248-5253 (2010).
- Womelsdorf, T., Vinck, M., Stan Leung, L., Everling, S. Selective theta synchronization of choice-relevant information subserves goal-directed behavior. Frontiers in Human Neuroscience. 4, 2010 (2010).
- Domic-Siede, M., Irani, M., Valdés, J., Perrone-Bertolotti, M., Ossandón, T. Theta activity from frontopolar cortex, mid-cingulate cortex and anterior cingulate cortex shows different roles in cognitive planning performance. NeuroImage. 226, 117557 (2021).
- Wilensky, R. . Planning and Understanding: A Computational Approach to Human Reasoning. , (1983).
- Grafman, J., Hendler, J. Planning and the brain. Behavioral and Brain Sciences. 14, 563-564 (1991).
- Hayes-Roth, B., Hayes-Roth, F. A cognitive model of planning. Cognitive Science. 3 (4), 275-310 (1979).
- Tremblay, M., et al. Brain activation with a maze test: An EEG coherence analysis study in healthy subjects. Neuroreport. 5 (18), 2449-2453 (1994).
- Shallice, T. Specific impairments of planning. Philosophical Transactions of the Royal Society B: Biological Sciences. 298 (1089), 199-209 (1982).
- Unterrainer, J. M., Owen, A. M. Planning and problem solving: From neuropsychology to functional neuroimaging. Journal of Physiology. 99 (4-6), 308-317 (2006).
- Domic-Siede, M., et al. La planificación cognitiva en el contexto de la evaluación neuropsicológica e investigación en neurociencia cognitiva: Una revisión sistemática. Terapia Psicológica. 40 (3), 367-395 (2022).
- Miotto, E. C., Morris, R. G. Virtual planning in patients with frontal lobe lesions. Cortex. 34 (5), 639-657 (1998).
- Burgess, P. W., Simons, J. S., Coates, L. M. A., Channon, S., Morris, R., Ward, G. The search for specific planning processes. The Cognitive Psychology of Planning. , 199-227 (2005).
- Oosterman, J. M., Wijers, M., Kessels, R. P. C. Planning or something else? Examining neuropsychological predictors of zoo map performance. Applied Neuropsychology. 20 (2), 103-109 (2013).
- Campbell, Z., et al. Utilizing virtual reality to improve the ecological validity of clinical neuropsychology: An fMRI case study elucidating the neural basis of planning by comparing the Tower of London with a three-dimensional navigation task. Applied Neuropsychology. 16 (4), 295-306 (2009).
- Wilson, B. A., Alderman, N., Burgess, P. W., Emslie, H., Evans, J. J. . Behavioural Assessment of the Dysexecutive Syndrome. , (1996).
- Spector, L., Grafman, J., Boller, F., Grafman, J. Planning, neuropsychology, and artificial intelligence: Cross fertilization. Handbook of Neuropsychology. 9, 377-392 (1994).
- Porteus, S. D. . The Maze Test and Clinical Psychology. , (1959).
- Lis, S., Krieger, S., Wilhelm, J., Gallhofer, B. Feedback about previous action improves executive functioning in schizophrenia: An analysis of maze solving behaviour. Schizophrenia Research. 78 (2-3), 243-250 (2005).
- Zelinsky, G. J. A theory of eye movements during target acquisition. Psychological Review. 115 (4), 787-835 (2008).
- Wurtz, R. H., Goldberg, M. E., Robinson, D. L. Brain mechanisms of visual attention. Scientific American. 246 (6), 124-135 (1982).
- Burr, D., Morrone, M. C. Eye movements: Building a stable world from glance to glance. Current Biology. 15 (20), 839-840 (2005).
- Rolfs, M. Microsaccades: Small steps on a long way. Vision Research. 49 (20), 2415-2441 (2009).
- Ortega, J., Plaska, C. R., Gomes, B. A., Ellmore, T. M. Spontaneous eye blink rate during the working memory delay period predicts task accuracy. Frontiers in Psychology. 13, 788231 (2022).
- Grueschow, M., Kleim, B., Ruff, C. C. Role of the locus coeruleus arousal system in cognitive control. Journal of Neuroendocrinology. 32 (12), 12890 (2020).
- Aston-Jones, G., Shaver, R., Dinan, T. Cortically projecting nucleus basalis neurons in rat are physiologically heterogeneous. Neuroscience Letters. 46 (1), 19-24 (1984).
- Foote, S. L., Bloom, F. E., Aston-Jones, G. Nucleus locus coeruleus: New evidence of anatomical and physiological specificity. Physiological Reviews. 63 (3), 844-914 (1983).
- Siever, L. J., Davis, K. L. Overview: Toward a dysregulation hypothesis of depression. The American Journal of Psychiatry. 142 (9), 1017-1031 (1985).
- Aston-Jones, G., Gold, J. I. How we say no: Norepinephrine, inferior frontal gyrus, and response inhibition. Biological Psychiatry. 65 (7), 548-549 (2009).
- Aston-Jones, G., Cohen, J. D. An integrative theory of locus coeruleus norepinephrine function: adaptive gain and optimal performance. Annual Review of Neurosciences. 28, 403-450 (2005).
- Einhauser, W., Stout, J., Koch, C., Carter, O. Pupil dilation reflects perceptual selection and predicts subsequent stability in perceptual rivalry. Proceedings of the National Academy of Sciences of the United States of America. 105 (5), 1704-1709 (2008).
- Yoshitomi, T., Ito, Y., Inomata, H. Adrenergic excitatory and cholinergic inhibitory innervations in the human iris dilator. Experimental Eye Research. 40 (3), 453-459 (1985).
- Rajkowski, J., Kubiak, P., Aston-Jones, G. Locus coeruleus activity in monkey: Phasic and tonic changes are associated with altered vigilance. Brain Research Bulletin. 35 (5-6), 607-616 (1994).
- Beatty, J. Task-evoked pupillary responses, processing load, and the structure of processing resources. Psychological Bulletin. 91 (2), 276-292 (1982).
- Beatty, J., Kahneman, D. Pupillary changes in two memory tasks. Psychonomic Science. 5, 371-372 (1966).
- Hess, E. H., Polt, J. M. Pupil size in relation to mental activity during simple problem-solving. Science. 143 (3611), 1190-1192 (1964).
- Kuchinke, L., Vo, M. L., Hofmann, M., Jacobs, A. M. Pupillary responses during lexical decisions vary with word frequency but not emotional valence. International Journal of Psychophysiology. 65 (2), 132-140 (2007).
- Barkley, R. A. Adolescents with attention-deficit/hyperactivity disorder: An overview of empirically based treatments. Journal of Psychiatric Practice. 10 (1), 39-56 (2004).
- Gau, S. S. F. e. m. 8. 9., Shang, C. Y. Executive functions as endophenotypes in ADHD: Evidence from the Cambridge Neuropsychological Test Battery (CANTAB). Journal of Child Psychology and Psychiatry and Allied Disciplines. 51 (7), 838-849 (2010).
- Bora, E., Harrison, B. J., Yücel, M., Pantelis, C. Cognitive impairment in euthymic major depressive disorder: A meta-analysis. Psychological Medicine. 43 (10), 2017-2026 (2013).
- Rive, M. M., Koeter, M. W. J., Veltman, D. J., Schene, A. H., Ruhé, H. G. Visuospatial planning in unmedicated major depressive disorder and bipolar disorder: Distinct and common neural correlates. Psychological Medicine. 46 (11), 2313-2328 (2016).
- Holt, D. V., Wolf, J., Funke, J., Weisbrod, M., Kaiser, S. Planning impairments in schizophrenia: Specificity, task independence and functional relevance. Schizophrenia Research. 149 (1-13), 174-179 (2013).
- Lima-Silva, T. B., et al. Functional profile of patients with behavioral variant frontotemporal dementia (bvFTD) compared to patients with Alzheimer's disease and normal controls. Dementia and Neuropsychologia. 7 (1), 96-103 (2013).
- Karnath, H. O., Wallesch, C. W., Zimmermann, P. Mental planning and anticipatory processes with acute and chronic frontal lobe lesions: A comparison of maze performance in routine and non-routine situations. Neuropsychologia. 29 (4), 271-290 (1991).
- Staudigl, T., Hartl, E., Noachtar, S., Doeller, C. F., Jensen, O. Saccades are phase-locked to alpha oscillations in the occipital and medial temporal lobe during successful memory encoding. PLoS Biology. 15 (12), 2003404 (2017).
- Faul, F., Erdfelder, E., Lang, A. G., Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behavior Research Methods. 39 (2), 175-191 (2007).
- Ferrando, L., Bobes, J., Gibert, J., Soto, M. MINI Entrevista Neuropsiquiátrica Internacional (MINI International Neuropsychiatric Interview, MINI). Instrumentos de Detección y Orientación Diagnóstica. , (2000).
- McAssey, M., Dowsett, J., Kirsch, V., Brandt, T., Dieterich, M. Different EEG brain activity in right and left-handers during visually induced self-motion perception. Journal of Neurology. 267 (1), 79-90 (2020).
- Kelly, R., Mizelle, J. C., Wheaton, L. A. Distinctive laterality of neural networks supporting action understanding in left- and right-handed individuals: An EEG coherence study. Neuropsychologia. 75, 20-29 (2015).
- O'Hare, A. J., Atchley, R. A., Young, K. M. Central and divided visual field presentation of emotional images to measure hemispheric differences in motivated attention. Journal of Visualized Experiments. (129), 56257 (2017).
- Beatty, J. Task-evoked pupillary responses, processing load, and the structure of processing resources. Psychological Bulletin. 91 (2), 276-292 (1982).
- Wainstein, G., et al. Pupil size tracks attentional performance in attention-deficit/hyperactivity disorder. Scientific Reports. 7 (1), 8228 (2017).
- Drueke, B., et al. Neural correlates of positive and negative performance feedback in younger and older adults. Behavioral and Brain Functions. 11, 17 (2015).
- Lezak, M. D. The problem of assessing executive functions. International Journal of Psychology. 17 (1-4), 281-297 (1982).
- Oosterman, J. M., Wijers, M., Kessels, R. P. C. Planning or something else? Examining neuropsychological predictors of zoo map performance. Applied Neuropsychology. 20 (2), 103-109 (2013).
- Keil, A., et al. Committee report: Publication guidelines and recommendations for studies using electroencephalography and magnetoencephalography. Psychophysiology. 51 (1), 1-21 (2014).
- Abokyi, S., Owusu-Mensah, J., Osei, K. A. Caffeine intake is associated with pupil dilation and enhanced accommodation. Eye. 31 (4), 615-619 (2017).
- Wilhelm, B., Stuiber, G., Lüdtke, H., Wilhelm, H. The effect of caffeine on spontaneous pupillary oscillations. Ophthalmic & Physiological Optics. 34 (1), 73-81 (2014).
- McGinley, M. J., David, S. V., McCormick, D. A. Cortical membrane potential signature of optimal states for sensory signal detection. Neuron. 87 (1), 179-192 (2015).
- Slotnick, S. D. High density event-related potential data acquisition in cognitive neuroscience. Journal of Visualized Experiments. (38), 1945 (2010).
- Lamm, C., Windischberger, C., Leodolter, U., Moser, E., Bauer, H. Co-registration of EEG and MRI data using matching of spline interpolated and MRI-segmented reconstructions of the scalp surface. Brain Topography. 14 (2), 93-100 (2001).
- Schwartz, D., et al. Registration of MEG/EEG data with 3D MRI: Methodology and precision issues. Brain Topography. 9, 101-116 (1996).
- Laeng, B., Sirois, S., Gredebäck, G. Pupillometry: A window to the preconscious? Perspectives on psychological science. Journal of the Association for Psychological Science. 7 (1), 18-27 (2012).
- Vandierendonck, A. A comparison of methods to combine speed and accuracy measures of performance: A rejoinder on the binning procedure. Behavior Research Methods. 49 (2), 653-673 (2017).
- Vandierendonck, A. Further tests of the utility of integrated speed-accuracy measures in task switching. Journal of Cognition. 1 (1), 8 (2018).
- Levene, H., Olkin, I. Robust tests for equality of variances. In Contributions to Probability and Statistics: Essays in Honor of Harold Hotelling. , 278-292 (1960).
- Zimmerman, D. W. A note on preliminary tests of equality of variances. The British Journal of Mathematical and Statistical Psychology. 57, 173-181 (2004).
- D'Agostino, R. B., Belanger, A., D'Agostino, R. B. A suggestion for using powerful and informative tests of normality). The American Statistician. 44 (4), 316 (1990).
- Shapiro, S. S., Wilk, M. B. An analysis of variance test for normality (complete samples). Biometrika. 52 (3-4), 591-611 (1965).
- Wilcoxon, F. Individual comparisons by ranking methods. Biometrics Bulletin. 1 (6), 80 (1945).
- Fay, M. P., Proschan, M. A. Wilcoxon-Mann-Whitney or t-test? On assumptions for hypothesis tests and multiple interpretations of decision rules. Statistics Surveys. 4, 1-39 (2010).
- Delorme, A., Makeig, S. EEGLAB: An open-source toolbox for analysis of single-trial EEG dynamics including independent component analysis. Journal of Neuroscience Methods. 134 (1), 9-21 (2004).
- Dimigen, O., Sommer, W., Hohlfeld, A., Jacobs, A. M., Kliegl, R. Coregistration of eye movements and EEG in natural reading: Analyses and review. Journal of Experimental Psychology. 140 (4), 552-572 (2011).
- Mognon, A., Jovicich, J., Bruzzone, L., Buiatti, M. ADJUST: An automatic EEG artifact detector based on the joint use of spatial and temporal features. Psychophysiology. 48 (2), 229-240 (2011).
- Bell, A. J., Sejnowski, T. J., Leen, T. K., Tesauro, G., Touretsky, D. S. A non-linear information maximization algorithm that performs blind separation. Advances in Neural Information Processing Systems 7. , (1995).
- Plöchl, M., Ossandón, J. P., König, P. Combining EEG and eye tracking: Identification, characterization, and correction of eye movement artifacts in electroencephalographic data. Frontiers in Human Neuroscience. 6, 278 (2012).
- Pion-Tonachini, L., Kreutz-Delgado, K., Makeig, S. ICLabel: An automated electroencephalographic independent component classifier, dataset, and website. NeuroImage. 198, 181-197 (2019).
- Gartner, M., Grimm, S., Bajbouj, M. Frontal midline theta oscillations during mental arithmetic: Effects of stress. Frontiers in Behavioral Neuroscience. 9, 96 (2015).
- Wang, W., Viswanathan, S., Lee, T., Grafton, S. T. Coupling between theta oscillations and cognitive control network during cross-modal visual and auditory attention: Supramodal vs modality-specific mechanisms. PLoS One. 11 (7), 0158465 (2016).
- Maris, E., Oostenveld, R. Nonparametric statistical testing of EEG- and MEG-data. Journal of Neuroscience Methods. 164 (1), 177-190 (2007).
- Tadel, F., Baillet, S., Mosher, J. C., Pantazis, D., Leahy, R. M. Brainstorm: A user-friendly application for MEG/EEG analysis. Computational Intelligence and Neuroscience. 2011, 879716 (2011).
- Pascual-Marqui, R. D. Standardized low-resolution brain electromagnetic tomography (sLORETA): Technical details. Methods and Findings in Experimental and Clinical Pharmacology. 24, 5-12 (2002).
- Gramfort, A., Papadopoulo, T., Olivi, E., Clerc, M. OpenMEEG: Opensource software for quasistatic bioelectromagnetics). Biomedical Engineering Online. 9, 45 (2010).
- Destrieux, C., Fischl, B., Dale, A., Halgren, E. Automatic parcellation of human cortical gyri and sulci using standard anatomical nomenclature. Neuroimage. 53 (1), 1-15 (2010).
- Orr, J. M., Weissman, D. H. Anterior cingulate cortex makes 2 contributions to minimizing distraction. Cerebral Cortex. 9 (3), 703-711 (2009).
- Christoff, K., Gabrieli, J. D. E. The frontopolar cortex and human cognition: Evidence for a rostrocaudal hierarchical organization within the human prefrontal cortex. Psychobiology. 28 (2), 168-186 (2000).
- Nitschke, K., Köstering, L., Finkel, L., Weiller, C., Kaller, C. P. A meta-analysis on the neural basis of planning: Activation likelihood estimation of functional brain imaging results in the Tower of London task. Human Brain Mapping. 38 (1), 396-413 (2017).
- Barbey, A. K., Koenigs, M., Grafman, J. Dorsolateral prefrontal contributions to human working memory. Cortex. 49 (5), 1195-1205 (2013).
- Le Van Quyen, M., et al. Comparison of Hilbert transform and wavelet methods for the analysis of neuronal synchrony. Journal of Neuroscience Methods. 111 (2), 83-98 (2001).
- Bokil, H., Andrews, P., Kulkarni, J. E., Mehta, S., Mitra, P. P. Chronux: A platform for analyzing neural signals. Journal of Neuroscience Methods. 192 (1), 146-151 (2010).
- Sato, N., Yamaguchi, Y. EEG theta regulates eye saccade generation during human object-place memory encoding. Advances in Cognitive Neurodynamics ICCN 2007. , 429-434 (2008).
- Zaki, J., Ochsner, K. The need for a cognitive neuroscience of naturalistic social cognition. Annals of the New York Academy of Sciences. 1167, 16-30 (2009).
- Shamay-Tsoory, S. G., Mendelsohn, A. Real-life neuroscience: An ecological approach to brain and behavior research. Perspectives on Psychological Science. 14 (5), 841-859 (2019).
- Diamond, A. Executive functions. Annual Review of Psychology. 64, 135-168 (2013).
- Dudai, Y. . Memory from A to Z: Keywords, Concepts and Beyond. , (2002).
- Kingstone, A., Smilek, D., Ristic, J., Friesen, C. K., Eastwood, J. D. Attention, researchers! It is time to take a look at the real world. Current Directions in Psychological Science. 12 (5), 176-180 (2003).
- Morris, R. G., Ward, G., Morris, R., Ward, G. Introduction to the psychology of planning. The Cognitive Psychology of Planning. , (2005).
- Zwosta, K., Ruge, H., Wolfensteller, U. Neural mechanisms of goal-directed behavior: Outcome-based response selection is associated with increased functional coupling of the angular gyrus. Frontiers in Human Neuroscience. 9, 180 (2015).
- Owen, A. M., Doyon, J., Petrides, M., Evans, A. C. Planning and spatial working memory: A positron emission tomography study in humans. European Journal of Neuroscience. 8 (2), 353-364 (1996).
- Ossandon, T., et al. Efficient "pop-out" visual search elicits sustained broadband gamma activity in the dorsal attention network. Journal of Neuroscience. 32 (10), 3414-3421 (2012).
- Lezak, M. D. . Neuropsychological Assessment., 3rd edition. , (1995).
- Lin, H., Saunders, B., Hutcherson, C. A., Inzlicht, M. Midfrontal theta and pupil dilation parametrically track subjective conflict (but also surprise) during intertemporal choice. NeuroImage. 172, 838-852 (2018).
- Yu, S., Mückschel, M., Beste, C. Superior frontal regions reflect the dynamics of task engagement and theta band-related control processes in time-on task effects. Scientific Reports. 12 (1), 846 (2022).
- Nakatani, H., van Leeuwen, C. Antecedent occipital alpha band activity predicts the impact of oculomotor events in perceptual switching. Frontiers in Systems Neuroscience. 7, 19 (2013).
- Velasques, B., et al. Changes in saccadic eye movement (SEM) and quantitative EEG parameter in bipolar patients. Journal of Affective Disorders. 145 (3), 378-385 (2013).
- Bodala, I. P., Li, J., Thakor, N. V., Al-Nashash, H. EEG and eye tracking demonstrate vigilance enhancement with challenge integration. Frontiers in Human Neuroscience. 10, 273 (2016).
- Dimigen, O. Optimizing the ICA-based removal of ocular EEG artifacts from free viewing experiments. NeuroImage. 207, 116117 (2020).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены