
Method Article
시공간 계획 작업(Eye-Tracker 및 Electroencephalogram Systems와 결합)
요약
EEG와 시선 추적 시스템을 결합한 인지 계획 연구는 인간의 인지 제어 및 목표 지향적 행동을 중재하는 신경 메커니즘을 조사하기 위한 다중 모드 접근 방식을 제공합니다. 여기에서는 계획 수행에서 뇌 진동과 안구 운동의 역할을 조사하기 위한 프로토콜에 대해 설명합니다.
초록
목표에 도달하기 위해 조직적인 계획을 수립할 수 있는 능력을 특징으로 하는 계획 프로세스는 인간의 목표 지향적 행동에 필수적입니다. 여러 신경 정신 질환에서 계획이 손상되기 때문에 계획을 검토하기 위한 적절한 임상 및 실험 테스트를 구현하는 것이 중요합니다. 여러 인지 영역이 참여하는 계획 배치의 특성으로 인해 계획 평가 및 신경 영상 방법과 결합된 행동 패러다임 설계는 인지 신경 과학의 현재 과제입니다. 계획 과제는 27명의 건강한 성인 참가자를 대상으로 뇌파(EEG) 시스템 및 안구 운동 기록과 함께 평가되었습니다. 계획은 두 단계로 나눌 수 있습니다 : 일련의 단계가 내부적으로 표현되는 정신적 계획 단계와 이전에 계획된 목표를 달성하기 위해 운동 행동을 사용하는 실행 단계. 우리의 프로토콜에는 계획 작업과 제어 작업이 포함되었습니다. 계획 작업에는 각각 동물원 지도를 나타내는 36개의 미로 시험을 해결하는 것이 포함되었습니다. 과제는 4개의 기간으로 나뉩니다: i) 계획, 피험자들은 일련의 규칙에 따라 4마리의 동물의 위치를 방문할 경로를 계획하도록 지시받았습니다. ii) 피험자가 작업 기억에 계획된 경로를 유지해야 하는 유지 관리; iii) 피험자가 안구 추적기 시스템에 표시된 대로 이전에 계획된 경로를 추적하기 위해 안구 움직임을 사용하는 실행; iv) 피험자가 방문한 동물의 순서를 보고한 응답. 통제 과제는 비슷한 구조를 가지고 있었지만 인지 계획 구성 요소는 과제 목표를 수정하여 제거되었습니다. EEG의 공간적, 시간적 패턴은 계획이 시간이 지남에 따라 전두엽-정중선 세타 활성(FMθ)의 점진적이고 지속적인 증가를 유도한다는 것을 보여주었습니다. 이 활동의 원인은 출처 분석에 의해 전전두엽 피질 내에서 확인되었습니다. 우리의 결과는 EEG와 시선 추적기 시스템을 결합한 실험 패러다임이 인지 계획을 평가하는 데 최적임을 시사했습니다.
서문
지난 10년 동안 인지와 행동 모두에 대한 진동 신경 역학의 역할을 조사하기 위해 광범위한 연구가 수행되었습니다. 이러한 연구는 특화된 피질 영역과 광범위한 피질 영역 간의 빈도별 상호 작용이 인지 및 인지 통제에 중요한 역할을 한다는 것을 입증했습니다 1,2,3. 이 접근법은 뇌 활동의 리듬적 특성을 강조하여 대규모 대뇌 피질 역학을 조정하는 데 도움이 되고 인지 처리 및 목표 지향적 행동을 뒷받침합니다 4,5. 뇌의 리듬 진동이 지각6, 주의력 7,8,9, 의사 결정10, 기억 재활성화11, 작업 기억12, 인지 통제13 등 다양한 인지 과정에 관여한다는 것을 보여주는 상당한 증거가 있습니다. 목표 지향적 행동을 안내하기 위해 다양한 진동 메커니즘이 제안되었으며, 일시적인 대규모 주파수 특정 네트워크는 인지 처리를 위한 프레임워크를 제공합니다 1,14,15. 예를 들어, 최근의 연구 결과에 따르면 뇌의 특정 주파수 대역은 스파이크 활동을 조절하는 피드백 메커니즘을 반영하여 피질 흥분성과 스파이크 타이밍을 조정하기 위한 시간적 참조 프레임을 제공할 수 있습니다 16,17,18. 검토는 Helfrich와 Knight19에 의해 제공되었습니다.
이 증거는 전전두엽 피질(PFC)이 계획 작업 컨텍스트 및 관련 행동 관련 규칙을 인코딩하는 방법에 대한 질문을 제기합니다. PFC는 그것이 생성하는 신경 활동의 진동 패턴을 통해 인지 제어 및 목표 지향적 행동을 지원하고, 먼 뇌 영역의 신경 활동을 선택적으로 편향시키며, 대규모 신경망20에서 정보의 흐름을 제어하는 것으로 오랫동안 생각되어 왔습니다. 또한, 지역적 동조를 보이는 지역이 지역간 활동에 참여할 가능성이 더 높다는 것이 제안되었다 21,22,23. 특히, 두피 뇌전도(EEG)로 측정한 피질 세타밴드(4-8Hz) 진동은 광범위한 네트워크를 통해 하향식 제어를 전달하기 위한 잠재적 메커니즘으로 제안되었습니다13. 구체적으로 말하자면, 인간의 세타밴드 활동은 기억 인코딩 및 검색, 작업 기억 유지, 참신함 감지, 의사 결정, 하향식 제어 12,24,25,26과 같은 높은 수준의 인지 과정을 반영합니다.
이와 관련하여, Cavanagh와 Frank13은 통제 과정의 두 가지 순차적 메커니즘을 제안했다: 통제의 필요성에 대한 인식과 통제의 인스턴스화. 통제의 필요성에 대한 인식은 내측 전전두엽 피질(mPFC)에서 유래하는 전두엽 정중선 세타(FMθ) 활성에 의해 지시될 수 있으며, 이는 새로운 정보(27,28,29), 충돌하는 자극-반응 요건(30), 오류 피드백(31), 및 오류 감지32. 새로움, 갈등, 처벌 또는 오류가 있는 경우 향상된 인지 제어의 필요성을 반영하는 이러한 ERP 구성 요소는 전두 엽 정중선 전극 26,27,33,34,35,36,37,38,39,40에 기록된 세타 대역에서 공통 스펙트럼 서명을 나타냅니다. 41,42,43,44.
FMθ 활동의 EEG 응답은 세타 주파수 대역(26)에서 위상 재설정 및 전력 향상의 패턴을 나타낸다. 공간 해상도 측면에서 EEG 방법의 한계에도 불구하고 FMθ 활성이 MCC(mid-cingulate cortex)에 의해 생성된다는 것을 입증하기 위해 다양한 증거 출처가 수집되었습니다13. 이러한 세타 역학은 mPFC의 뉴런 과정을 조절하는 시간적 틀 역할을 하는 것으로 여겨지며, 이는 이후에 고차원의 통제를 필요로 하는 사건에 대한 응답으로 증강됩니다26. 이는 근원 분석 31,33,45,46,47, 동시 EEG 및 기능적 자기공명영상(fMRI) 기록 48,49, 인간 50 및 원숭이51,52,53의 침습적 EEG 기록을 통해 확립되었습니다.
이러한 관찰을 바탕으로 전두엽 정중선 세타는 계획 중과 같이 행동과 결과에 대한 확신이 부족한 다양한 상황에서 적응 제어를 실행하기 위한 보편적인 메커니즘, 공통 언어로 작동하는 것으로 간주됩니다. 이 프로토콜에서 제안하는 행동 패러다임은 인지 계획과 그 시간적 및 신경적 특성을 연구하는 데 사용되었습니다. 다른 시나리오에서도 인지 통제를 위한 다양한 메커니즘이 보고되었지만, 현재의 프로토콜은 계획 및 이와 관련된 신경 및 시간적 속성에 대한 최근의 설명을 허용했다54. 계획의 인지적 과정은 두 가지 뚜렷한 단계로 나뉜다: 일련의 계획들에 대한 내적 표현이 개발되는 정신적 계획 단계(mental planning phase)55와 이전에 계획된 목표를 달성하기 위해 일련의 운동 행동이 실행되는 계획 실행 단계(planning execution phase)56. 계획은 작업 기억, 주의력 조절 및 반응 억제를 포함한 실행 기능의 다양한 구성 요소의 통합을 필요로 하는 것으로 알려져 있으며, 이러한 과정에 대한 실험적 조작 및 격리 측정은57,58 .
인지 계획에 대한 신경 영상 연구는 일반적으로 다음과 같은 행동 패러다임을 사용했습니다 : 런던 타워59 , 60 , 61; 그러나 교란 요인을 통제하기 위해 인지 계획을 연구하는 데 사용되는 작업은 제한적이고 인위적일 수 있으며, 이로 인해 예측 및 생태학적 타당성이 떨어질 수 있습니다 62,63,64,65. 신경심리학 분야에서 이러한 문제를 극복하기 위해 현실 세계의 계획 상황이 생태학적 과제로 제안되어 왔다62,63. Dysexecutive Syndrome 배터리의 행동 평가에서 Zoo Map Task 하위 테스트는 보다 자연스럽고 적절한 방식으로 계획 및 조직 기술을 측정합니다64,66. 이 테스트는 동물원 지도의 12개 위치 중 6개를 방문하는 경로를 계획하는 연필과 종이 테스트입니다. 위치는 코끼리 집, 사자 우리, 휴게소, 커피숍 등과 같이 일반 동물원에서 볼 수 있는 일반적인 장소입니다. 서로 다른 수준의 계획을 평가하는 두 가지 조건이 있습니다 : i) 공식화 조건, 피험자가 선택한 순서대로 그러나 일련의 규칙에 따라 6 곳을 방문하는 경로를 계획하도록 지시받습니다. ii) 피험자가 특정 순서로 6 개의 장소를 방문하고 일련의 규칙에 따라 방문하도록 지시받는 실행 조건. 이 두 가지 조건은 잘못 구조화된 문제(공식화)와 잘 구조화된 문제(실행)에서 계획 기술에 대한 정보를 제공한다67. 첫 번째는 열린 상황에서 더 까다로운 인지 과제로 제시되는데, 그 이유는 피험자가 목표를 달성하기 위해 논리적인 전략을 개발해야 하기 때문입니다. 경로를 추적하기 전에 일련의 연산자를 고안해야 합니다. 그렇지 않으면 오류가 발생할 수 있습니다. 다른 한편으로는, 실행 조건은 더 낮은 인지적 요구를 요구하는데, 그 이유는 부과된 특정 전략을 따르는 것과 관련된 작업을 해결하는 것은 피험자가 목표를 달성하기 위해 공식화된 계획의 실행을 모니터링하기만 하면 되기 때문이다66. 한편, Porteus Maze는 심리학 분야, 특히 인지 심리학 및 신경 심리학 분야에서 잘 알려진 작업이며, 문제 해결 및 계획과 같은인지의 다양한 측면을 평가하는 도구로 널리 사용되어 왔습니다68,69. Porteus Maze 작업은 간단한 시각적 자극 분석으로 시작하여 점점 더 어려워지는 연필과 종이 작업입니다. 피험자는 시작점에서 출구(여러 선택지 중에서)까지 올바른 경로를 찾고 추적해야 하며, 교차하는 경로와 막다른 골목을 피하고 가능한 한 빨리 행동하는 등의 규칙을 따라야 한다68. 경로를 그리는 동안 갈림길이 나타날 때마다, 피험자는 목표에 도달하고 주어진 규칙을 어기지 않기 위해 결정을 내린다69.
일반적으로 사용되는 생태학적 과제의 한계와 강점을 고려하여 동물원 지도 과제66 과 포르테우스 미로 과제68을 중심으로 행동 패러다임을 설계했습니다. 행동 패러다임은 일상 생활 시나리오에서 계획의 인지 과정을 포함하는 4개의 뚜렷한 단계로 구성됩니다. 이 단계는 다음과 같습니다 : 1 단계, 계획, 참가자가지도의 다양한 위치를 방문하기위한 경로를 만들고 설정된 규칙을 준수하는지 확인하는 임무를 맡습니다. 2단계, 유지 보수, 참가자는 계획된 경로를 작업 메모리에 보관해야 합니다. 3단계, 실행: 참가자가 사전에 계획한 경로를 그리고 정확성을 면밀히 모니터링하여 실행합니다. 4단계, 응답, 참가자들은 계획된경로 54에 따라 방문한 동물의 순서를 보고합니다. 우리의 패러다임은 경로를 매핑하는 것이 일상 생활에서 흔히 발생하기 때문에 계획의 다양한 구성 요소(예: 작업 기억, 실행 주의력 및 시공간 기술)를 보다 현실적인 방식으로 반영하는 여러 단계를 사용하여 계획 능력의 다양한 매개변수를 측정할 수 있습니다. 또한, 교란 요인을 통제하기 위해 패러다임에는 계획 작업 구조와 이에 상응하는 자극이 있는 제어 작업이 포함되며, 이는 계획에 관련된 실행 인지 구성 요소를 참여시키지만 계획 프로세스 구성 요소는 제외합니다. 이를 통해 전기생리학적 마커와 행동 매개변수의 비교를 위해 계획 프로세스 구성 요소를 분리할 수 있습니다54.
또한 시선 추적은 안구 움직임을 측정하고 분석하기 위한 비침습적 방법을 제공함으로써 인지 신경 과학 연구에 상당한 기여를 했으며, 이는 지각, 주의력 및 인지 기능의 기저에 있는 인지 과정과 신경 메커니즘에 대한 귀중한 통찰력을 제공할 수 있습니다. 시선 추적 시스템으로 다양한 유형의 안구 움직임을 측정하면 계획과 관련된 인지 과정과 신경 메커니즘에 대한 귀중한 정보를 제공할 수 있습니다. 예를 들어, 다음과 같은 측면들을 측정할 수 있다: 고정(fixations)은 시각 정보가 획득되는 안정된 시선의 기간이다70; 사카데스(saccades)는 한 위치에서 다른 위치로 시선을 옮기는 데 사용되는 빠른 안구 운동이다71; 부드러운 추적(smooth pursuit)은 눈이 움직이는 물체를 부드럽게 따라갈 수 있도록 하는 일종의 안구 운동이다72; 고정 중에도 발생하는 작고 빠른 안구 운동인 마이크로사케이드73; 눈을 깜박이는 것은 눈을 윤활시키고 이물질로부터 눈을 보호하는 데 도움이 되는 반사 작용입니다74. 이러한 안구 운동은 계획 및 인지 통제에 중요한 구성 요소인 시각적 검색, 주의 할당70, 시각적 추적72, 지각73 및 작업 기억(74)과 관련된 인지 과정에 대한 통찰력을 제공할 수 있습니다.
한편, 유전자좌(locus coeruleus-norepinephrine, LC-NE) 시스템에 대한 최근 연구는 인지 통제에서 적절한 역할을 하는 것으로 나타났습니다75. 대뇌 좌위(locus coeruleus, LC)는 대뇌 피질(cerebral cortex), 해마(hippocampus), 시상(thalamus), 중뇌(midbrain), 뇌간(brainstem), 소뇌(cerebellum) 및 척수(spinal cord)와 같은 여러 뇌 영역에 돌출되어 있습니다 76,77,61. 특히 조밀한 LC-NE 신경 분포는 인지 조절과 관련된 PFC 뇌 영역을 받습니다75. 게다가, 일부 연구에서는 LC 시스템의 만성 과잉 행동이 충동성 및 불면증과 같은 조울 장애 증상에 기여할 수 있음을 나타냅니다. 이와는 대조적으로, LC 기능의 만성적인 감소는 우울증을 앓고 있는 환자들 사이에서 널리 퍼져 있는 특징인 감정 표현 감소와 관련이 있다78. 자극에 대한 유전자좌(locus coeruleus)의 과잉 반응은 스트레스 또는 불안 장애가 있는 개인에게 과도한 반응으로 이어질 수 있다79. 따라서 LC-NE 시스템의 변화는 인지 및/또는 정서적 조절 장애의 증상에 기여할 수 있습니다. 비침습적 기법을 사용하여 유전자좌 코에룰루스 활성을 검사할 수 있으며, 그 중 하나는 동공 직경 변화이며, 이는 대부분 코에룰루스 자리에서 방출되는 노르아드레날린에 의해 제어됩니다. 노르아드레날린은 알파-아드레날린 수용체를 자극하여 홍채 확장근과 모양체 신경절에 신호를 보내고 시냅스후 알파-2 아드레날린 수용체 66,80,81,82의 활성화를 통해 홍채 확장을 제어하는 에딩거-웨스트팔 핵에 작용합니다. 원숭이의 직접 LC 뉴런 기록은 LC-NE 활동, 동공 지름 및 인지 능력 사이의 관계를 확인했다83. 동공 확장은 여러 인지 과제에서 향상된 처리 요구에 대한 응답으로 반복적으로 관찰되었습니다 71,84,85,86,87.
시선 추적 및 동공 기록과 결합된 인지 제어의 전기생리학적 마커는 인지 제어 및 계획이 뇌에서 어떻게 구현되는지에 대한 중요한 질문을 풀 수 있습니다. EEG와 시선 추적기 시스템을 결합한 프로토콜 사용의 중요성은 두 가지입니다. 한편으로는, 인지 통제는 정확한 시간적 관계에 분산된 뇌 활동의 참여를 필요로 하는 것으로 보이며, 이는 뇌 네트워크 기능을 연구하기 위한 이상적인 후보를 구성합니다. 다른 한편으로는, 주의력 결핍/과잉 행동 장애(attention deficit/hyperactivity disorder)88,89, 주요우울장애(major depressive disorder) 90,91, 양극성 장애(bipolar disorder)91, 정신분열증(schizophrenia92), 전두측두엽 치매(frontotemporal dementia)93 93 와 같은 다양한 인지 및 신경정신 장애와 같은 인지 및 신경 정신 장애의 경우와 같이 이러한 능력의 이상은 정상적인 행동에 심각한 영향을 미친다. 또한 현재 프로토콜은 시선 추적 및 뇌파를 사용하여 LC-NE 활성과 진동을 비교하기 위한 매개변수로 pupillometry를 사용할 수 있도록 합니다. 이는 인간의 LC-NE, 동공 측정 및 신경 마커 간의 이론적 관계에 대한 증거를 제공할 수 있을 뿐만 아니라 인지 계획 중에 LC-NE 시스템과 관련된 특성의 발달 궤적을 추적할 수 있습니다. 그러나 이 모델에서는 계획 중에 잠재적으로 특정 진동 변화를 초래할 수 있는 특정 패턴의 단속이 있는지 여부를 테스트하는 데 중점을 두었습니다95. 또한, 우리는 행동 패러다임의 실행 단계에서 계획의 행동 실행을 검사하는 중요한 부분으로 시선 추적기 시스템을 사용했습니다.
요약하자면, 이 프로토콜은 추가 기초 연구와 궁극적인 임상 및 치료 응용 모두를 위한 플랫폼 역할을 할 수 있는 뇌 네트워크 역학의 테스트 가능한 모델을 생성할 수 있습니다.
프로토콜
이 프로토콜의 모든 절차는 Pontificia Universidad Católica de Chile 의학부 생명윤리위원회의 승인을 받았으며 모든 참가자는 연구 시작 전에 정보에 입각한 동의서에 서명했습니다(연구 프로젝트 번호: 16-251).
1. 참가자 모집
- 정상 또는 정상에서 정상으로 교정된 시력을 가진 오른손잡이의 건강한 성인(남성 및 여성)을 모집하고 포함/제외 기준에 따라 선별합니다.
참고: 이 연구에서는 19세에서 38세 사이의 유창한 화자인 건강한 개인 27명을 모집했습니다. 표본 크기는 원하는 통계적 검정력 수준에 따라 달라질 수 있으며, 연령대는 다루어야 할 특정 연구 질문에 따라 달라질 수 있습니다. 프로토콜에서는 Faul et al.96에 설명된 대로 통계적 Wilcoxon 부호 순위 테스트, 효과 크기 0.7, 알파 수준 0.05 및 거듭제곱 0.95를 고려하여 표본 크기를 계산했습니다. 우리는 훈련된 심리학자가 신청한 MINI-International Neuropsychiatric Interview97을 사용하여 포함/제외 기준에 따라 참가자를 평가했습니다. 왼손잡이 개인은 EEG 활동의 다른 지형 분포를 나타낼 수 있으므로 EEG 신호의 변동성을 줄이기 위해 오른손잡이 피험자만 모집합니다 98,99,100.
2. 자극 준비
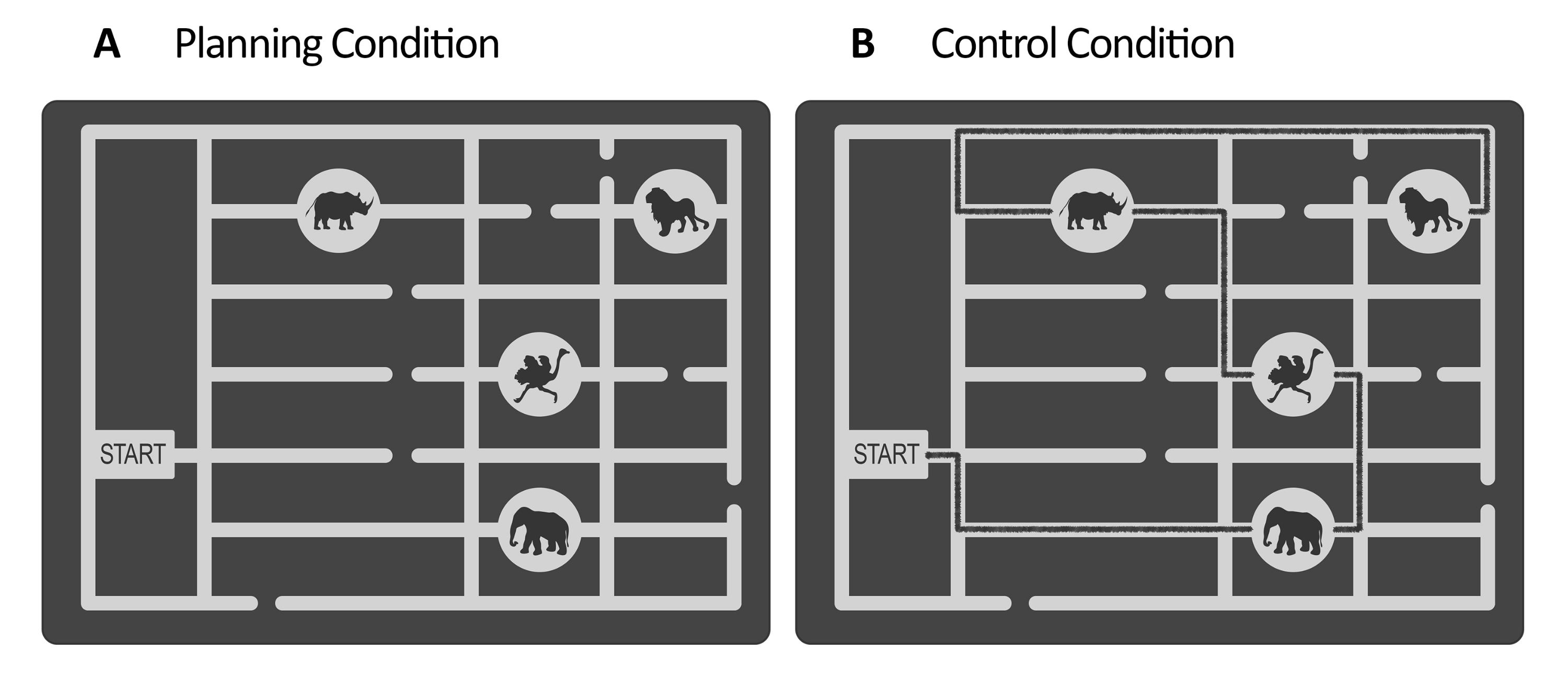
- 계획 작업의 경우 벡터 그래픽 편집기 소프트웨어를 사용하여 일련의 자극을 만듭니다( 재료 표 참조). 각 자극에 대해 동물원 지도를 나타내는 회색조 미로를 설계합니다. 미로 내부에는 동물의 위치로 이어지는 관문과 여러 경로를 만듭니다(예: 이 연구에서는 4개의 동물 위치가 있었습니다. 그림 1 참조).
참고: 이 연구에서는 36개의 미로를 만들었으며, 각 자극은 시작 게이트가 있는 동물원 지도, 미로에 위치한 4개의 동물 이미지 및 여러 경로로 구성되었습니다. 미로의 길은 동물의 위치로 이어질 수도 있고 그렇지 않을 수도 있습니다. 대비가 감소된 그레이스케일 자극을 사용하는 것은 색각을 담당하는 망막 원추세포의 자극을 줄이기 때문에 동공 측정에 선호되는 경우가 많습니다. 이러한 자극 강조점의 변화는 동공 크기의 변화를 보다 민감하게 측정할 수 있게 해주며, 이는 각성 또는 주의력 상태의 변화를 반영하는 것으로 생각됩니다. 또한, 그레이스케일 자극을 사용하면 개인 간의 색각 차이로 인한 측정의 변동성을 줄일 수 있습니다101. - 최종 작업에서 다양한 수준의 복잡성을 얻으려면 설정된 목표와 규칙에 따라 유효한 솔루션의 수에 따라 자극을 나눕니다(특히 목표는 동물 위치를 방문하는 경로를 계획하는 것입니다). 유효한 솔루션의 수는 규칙에 따라 계획할 수 있는 경로의 수를 나타냅니다( 그림 2 및 5.12.1단계의 규칙 참조). 5개 이상의 가능한 해가 있는 자극은 "쉬움"으로, 5개 이하의 해가 있는 자극은 "어려움"으로 분류합니다. 그런 다음 각 범주에 대해 동일한 수의 평가판을 만듭니다.
참고: 또는 Domic-Siede et al.54 에 대해 생성된 자극은 이러한 지침에 따라 생성되었으므로 저자에게 요청하십시오. 모든 재료는 요청 시 사용할 수 있지만 사양은 여기에 자세히 설명되어 있습니다. 본 연구에서는 18건의 쉬운 임상시험과 18건의 어려운 임상시험을 만들었다. 행동 및 전기 생리학적 수준에서 난이도의 차이를 평가하는 것은 인지 요구/인지 노력/난이도 또는 인지 계획의 본질적인 측면을 측정하고 있는지 결정하는 데 도움이 되기 때문에 중요합니다(대표 결과 및 논의 섹션 참조). - 통제 작업의 경우 계획 작업과 동일한 구조(평가 기간, 유지 관리, 실행, 응답, 그림 2 참조)를 사용하고 계획 작업을 위해 생성된 것과 동일한 자극을 사용하되 게이트에서 시작하여 마지막 위치까지 동물 위치 순서를 방문하기 위한 표시된 경로를 나타내는 그려진 선을 추가합니다( 그림 1B 참조)). 표시된 경로를 미로의 주요 경로보다 약간 더 어두운 색상으로 만들고 조도계를 사용하여 낮은 대비를 평가합니다(2.4단계 참조).
참고: 이것의 이면에 있는 아이디어는 두 조건(계획 및 통제 작업)의 정신 물리학적 특징을 가능한 한 유사하게 유지하는 것입니다. 미로의 추적된 경로는 규칙을 따를 수도 있고 따르지 않을 수도 있다(참가자에게 주어진 지시에 대한 자세한 내용은 프로토콜의 섹션 5 참조). 이 연구에서 자극의 절반은 규칙에 따라 올바른 방문 순서를 보인 반면 나머지 절반은 오류를 보였습니다(예: 동일한 경로를 두 번 사용하거나 막다른 골목을 건너는 경우, 5.12.1단계 및 5.12.3단계 및 그림 2 참조). - 피험자가 사용할 턱받이에 위치한 조도계를 사용하여(4.5단계 및 4.6단계 참조) 화면에서 동일한 거리에서 자극의 조도를 평가합니다. 각 조건의 각 자극은 럭스 값을 생성합니다. 추가 분석을 위해 각 값을 수동으로 기록합니다.
참고: 조건 간에 조도의 차이는 예상되지 않습니다(4.6단계 참조). 그렇지 않으면 자극의 대비를 확인하십시오. 이는 동공 직경이102로 측정되는 경우와 관련이 있습니다. - 벡터 그래픽 편집기( Table of Materials)를 사용하여 올바른 피드백(정확할 때 엄지 손가락 위로)을 나타내는 하나의 자극과 잘못된 피드백(정확하지 않은 경우 엄지 손가락 아래로)을 나타내는 다른 자극을 만듭니다(그림 2).

그림 1: 실험 및 통제 과제의 자극. (A) 계획 및 (B) 통제 작업 자극의 예시적인 예가 표시됩니다. 자극은 게이트, 서로 다른 위치에 있는 4개의 동물 위치 및 여러 경로로 구성된 동물원 지도를 나타냅니다. 두 질환에 대한 자극은 비슷했다. 유일한 차이점은 제어 작업의 경우 (B) 자극에 이미 존재하는 경로를 나타내는 표시된 선이 있다는 것입니다(여기서는 설명을 위해 검은색 선). 실제 제어 자극의 이 선은 조도에 의해 제어되는 낮은 대비와 함께 약간 더 어두웠습니다(2.4단계 참조). 이 그림은 Domic-Siede et al.54에서 수정되었습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

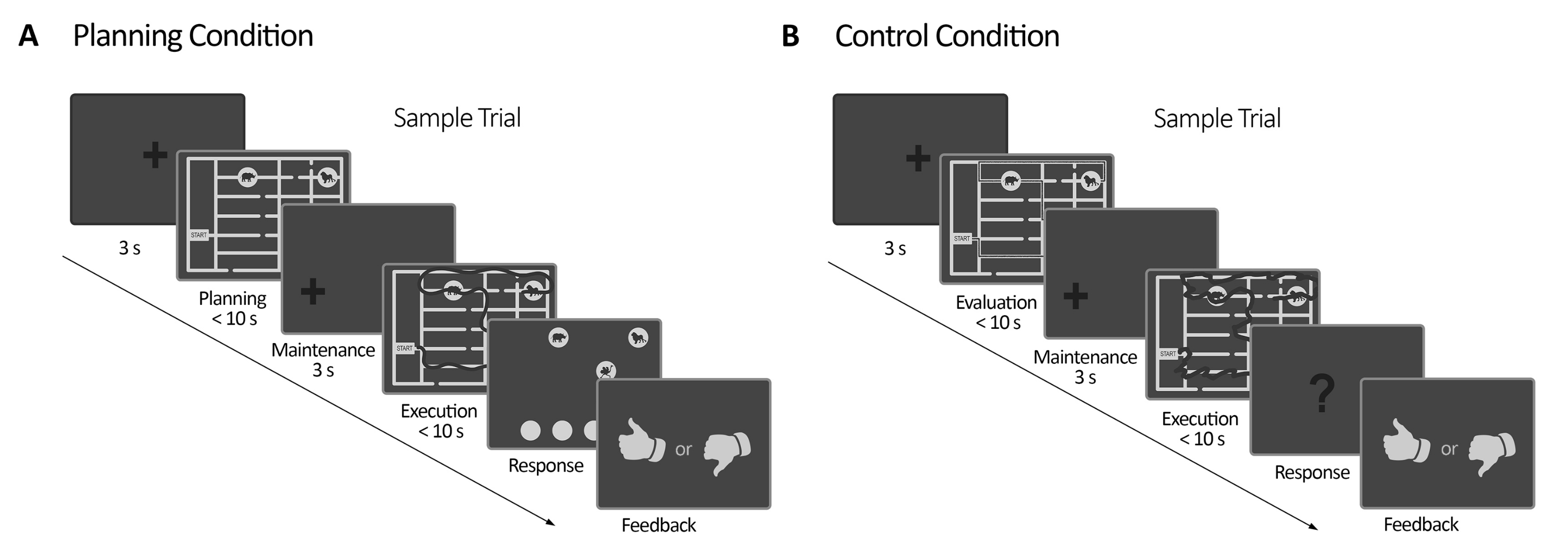
그림 2: 실험 설계. (A) 계획 과제 시험. 이 상태에서의 시험은 3초 고정 십자가로 시작되었습니다. 그런 다음 참가자들은 일련의 규칙(최대 10초)에 따라 4개의 동물 위치를 모두 방문할 수 있는 경로를 계획하도록 지시받았습니다. 다음으로, 이동 된 고정 십자가가 나타 났고 (3 초) 미로가 다시 나타났습니다. 이 기간(실행)에서 피험자는 시선 이동을 실시간(어두운 선)(최대 10초)으로 묘사하는 온라인 시각적 피드백(시선 추적기 시스템에서 제공)과 함께 시선을 사용하여 이전 계획 기간에 계획한 추적을 실행해야 했습니다. 그 후, 응답 시간에, 피험자들은 동물들을 방문하도록 명령함으로써 처형 중에 만들어진 순서를 보고해야 했다. 그들의 응답에 따라 피드백이 전달되었습니다. (B) 제어 작업 시험. 이 상태에서의 시험은 3초 고정 십자가로 시작되었습니다. 그런 다음 참가자들은 추적된 경로(어두운 선)가 규칙을 따랐는지 여부를 평가하도록 지시받았습니다. 다음으로, 이동 된 고정 십자가가 나타 났고 (3 초) 미로가 다시 나타났습니다. 이 기간 동안 피험자들은 계획 실행 기간(최대 10초)과 같이 온라인 시각적 피드백을 통해 이미 추적된 경로를 다시 그려야 했습니다. 그 후, 응답 기간 동안 피험자는 추적된 순서가 이전에 명시된 규칙을 따랐는지 여부에 대해 답변(예 또는 아니오)해야 했습니다. 이 그림은 Domic-Siede et al.54에서 수정되었습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
3. 계획 및 제어 작업 프로그래밍
- 자극 프레젠테이션/행동 실험 소프트웨어를 사용하여 Zoo Map Task66 및 Porteus Maze68을 기반으로 계획 작업 패러다임을 코딩하는 스크립트를 작성합니다( 재료 표 및 보충 파일 참조).
- 두 가지 조건(계획 조건 및 제어 조건)을 고려하여 섹션 2 및 섹션 4에서 설명한 것과 유사한 구조로 작업을 코딩합니다( 그림 2 및 보충 파일 참조).
참고: 작업 요구 사항을 해결하는 과정과 관련된 교란 요인과 지각 구성 요소를 제어하기 위해 두 조건에서 동일한 구조를 사용하는 것이 중요합니다(그림 2). 동일한 구조를 사용하면 인지 계획과 관련된 특정 인지 과정에 대한 평가가 향상됩니다. - 디스플레이 컴퓨터에서 TTL(Transistor-Transistor Logic) 펄스를 전송하는 이더넷 및 병렬 포트 통신을 통해 디스플레이 컴퓨터, EEG 컴퓨터 및 호스트 컴퓨터(아이트래커 컴퓨터) 간의 통신을 동기화합니다( 그림 3 참조).
- 실행 기간 동안 화면상의 시선 위치가 중요하기 때문에 계획 및 제어 작업을 시작할 때와 5번의 시도가 완료될 때마다 시선 추적기 시스템을 사용하여 안구 움직임을 보정하는 코드를 작성합니다(프로토콜의 3.2.3단계 및 3.3.3단계, 논의 섹션 및 보충 파일의 코드 참조).
참고: 컴퓨터 통신이 지연될 수 있습니다. 서로 다른 두 컴퓨터에서 TTL 펄스 간의 지연을 측정하는 방법에는 여러 가지가 있지만 일반적인 접근 방식 중 하나는 디지털 오실로스코프 또는 로직 분석기와 같은 하드웨어 디바이스를 사용하는 것입니다. 또 다른 접근 방식은 네트워크 연결을 통해 TTL 펄스를 전송하고 네트워크 분석 도구를 사용하여 지연을 측정하는 것과 같은 소프트웨어 기반 방법을 사용하는 것입니다. 또 다른 방법은 GPS(Global Positioning System) 또는 NTP(Network Time Protocol) 서버를 사용하거나 하드웨어 기반 동기화 솔루션을 사용하여 두 컴퓨터의 시계를 동기화하고, 각 펄스에 대한 타임스탬프와 도착 시간 사이의 지연을 계산하고, 결과를 평균화하여 두 컴퓨터 간의 전체 지연을 구하는 것입니다.
- 두 가지 조건(계획 조건 및 제어 조건)을 고려하여 섹션 2 및 섹션 4에서 설명한 것과 유사한 구조로 작업을 코딩합니다( 그림 2 및 보충 파일 참조).
- 계획 기간에 대한 코드를 계획 기간, 유지 관리 기간, 계획 실행 기간, 응답 기간 및 피드백(그림 2, 보충 파일).
- 계획 기간: 3초 동안 제시된 고정 교차를 기준선으로 설정하여 계획 조건을 시작합니다.
- 미로 세트를 하나씩 무작위로 제시합니다(이 연구에서는 36개).
참고: 이 계획 기간 동안 참가자들은 일련의 규칙에 따라 최대 10초 동안 4개의 동물 위치를 방문하는 경로를 계획해야 합니다(규칙은 사전에 설명되었습니다. 주어진 규칙을 보려면 프로토콜의 섹션 5와 그림 2를 참조하십시오). - 태그 코드를 사용하여 자극 프레젠테이션의 시작을 알리는 TTL 트리거를 코드에 포함하고, 이 트리거를 EEG 컴퓨터 및 시선 추적기 호스트 컴퓨터로 전송하여 더 좁고 창으로 구성된 분석을 수행합니다.
- 조이스틱/키보드의 버튼을 누르면 피험자가 계획을 완료할 때마다 또는 최대 시간이 초과되는 경우 계획 기간이 절정에 달한다는 코드를 작성합니다. 추가 분석을 위해 반응 시간(RT)을 로그 파일에 기록해야 합니다.
참고: 이 기간에는 숫자 1을 사용하는 트리거 코드를 사용했지만, HED 태그는 콘텐츠에 의미와 구조를 제공하여 다른 연구자나 공동 작업자가 데이터의 내용을 더 쉽게 이해할 수 있도록 하기 때문에 숫자 코드보다 HED(Hierarchical Event Descriptors) 태그를 사용하는 것이 좋습니다. - 유지 보수 기간: 3초 동안 제시된 이동된 고정 십자가를 사용하여 이 기간을 시작합니다. 동물원 지도의 시작 위치(게이트)를 예측하기 위해 미로의 문이 있는 공간적 위치에서 이동된 고정 십자가를 찾습니다( 그림 2 참조).
참고: 이 기간의 목적은 세 가지입니다. 첫째, 이동된 고정 교차는 다음 기간에 대해 계획된 경로를 나타내는 추적의 실행을 용이하게 합니다(3.2.8단계 참조). 둘째, 이 기간 동안 참가자들은 계획 기간 동안 정교하게 다듬어진 계획을 작업 기억에 간직합니다. 마지막으로, 이 기간은 계획 기간의 끝과 다음 기간(계획 실행 기간)의 시작을 구분하는 재판 간 간격 역할을 합니다. - 계획 실행 기간: 유지 보수 기간 동안 3초 동안 이동된 고정 십자가가 표시된 후 미로를 다시 제시합니다.
- TTL 트리거를 EEG 및 호스트 아이 트래커 컴퓨터에 전송하여 특정 태그 코드를 사용하여 이 기간의 시작을 표시합니다.
- 이 기간이 시작된 후 약 992ms 후에 피험자의 시선 위치에 대한 실시간 시각적 피드백(어두운 선, 그림 2의 실행 기간 참조)을 제공하는 코드를 작성합니다.
참고: 지연(약 1,000ms)을 가지고 묘사를 시작하면 피험자가 미로에서 방향을 잡을 시간을 갖게 되며, 이전에 계획된 경로(계획 기간 동안)를 어두운 선으로 묘사할 수 있습니다. - 피험자가 만든 경로를 추가로 재구성하기 위해 경로의 좌표를 기록하고 오프라인에서 성능을 점수화합니다( 그림 4의 6.1.1단계 참조).
- 계획된 경로를 추적하는 데 최대 10초의 시간을 확보하고 피험자가 버튼을 눌러 이 기간을 완료할 수 있도록 합니다. 이러한 방식으로 피사체는 그려진 경로를 언제 완료했는지 제어할 수 있습니다.
- 추가 분석을 위해 RT를 로그 파일에 저장합니다.
- 응답 기간: 계획 실행 10초 후 또는 계획 실행 기간이 끝날 때 버튼을 누르면 시작되는 응답 기간에 대한 코드를 작성하여 미로는 사라지지만 동물과 동물의 공간적 위치는 화면에 남아 있습니다.
- 응답 기간의 화면 아래쪽에 4개의 빈 원을 가로로 배치합니다.
주석: 이 기간의 목적은 피험자가 조이스틱이나 키보드를 사용하여 동물을 방문한 것과 동일한 순서로 동물을 원 안에 넣음으로써 계획 실행 기간 동안 방문한 동물의 순서를 나타낼 수 있도록 하는 것입니다. - 피험자가 조이스틱 또는 키보드를 사용하여 이전에 제시된 각 동물(이 연구에서는 4마리의 동물)을 선택하고 4개의 원 각각에 삽입할 수 있도록 프로그램/코드를 구성합니다( 보충 파일 및 그림 2 참조).
- 피드백: 참가자에게 3초 분량의 피드백을 전달하는 코드를 작성하십시오. 규칙을 따랐을 경우 방문한 동물의 유효한 조합에 대한 응답으로 엄지손가락을 치켜세운 이미지가 표시되어야 하며, 보고된 조합이 유효하지 않은 경우 엄지손가락을 내린 이미지가 표시되어야 합니다.
- 올바른 피드백을 위해 특정 태그 코드를 사용하고 잘못된 피드백을 위해 다른 태그 코드를 사용하여 EEG 및 시선 추적기 컴퓨터에 TTL 트리거를 보냅니다.
참고: 피드백을 제공하는 이유는 성과 모니터링을 용이하게 하고 작업 중에 동기를 유지하기 위함입니다. 이러한 실시간 피드백의 제공은 보상 효과를 높이고 적절한 작업 수행을 장려합니다103.
- 계획 조건과 동일한 구조(제어 기간, 유지 관리 기간, 제어 실행 기간, 응답 기간 및 피드백)로 제어 작업에 대한 코드를 작성합니다( 보충 파일, 그림 2 참조).
- 통제 기간: 교란 요인을 완화하기 위해 제어 조건 기간에 대한 코드를 작성합니다. 이 기간의 코드는 기준선으로 3초 동안 제시된 고정 십자가로 시작해야 합니다.
참고: 계획 과제는 주로 계획의 실행을 요구하지만 시공간 기능, 작업 기억, 주의 통제, 억제 통제 등과 같은 실행 기능의 일부로 다른 인지 영역도 동원하기 때문에66,88,104,105 통제 과제는 교란 요인을 완화하는 데 매우 중요합니다. 따라서, 이 과제의 주요 목표는 인지적 계획(cognitive planning)54의 실행을 제거하면서 계획 과제를 해결하는 데 필요한 모든 인지적 및 지각 기능을 요구하는 것이다. - 제어 조건의 미로를 하나씩 무작위로 표시합니다(표시된 경로가 이미 추적된 미로). 최대 10초의 시간을 코딩합니다.
- 태그를 사용하여 자극 프레젠테이션의 시작을 알리는 TTL 트리거를 코드에 포함시키고 이 트리거를 EEG 컴퓨터와 시선 추적기 호스트 컴퓨터로 보냅니다.
- 이 제어 기간은 조이스틱/키보드의 버튼을 누르면 주제가 끝날 때마다 또는 최대 시간이 초과되는 경우 절정에 달한다고 코드를 작성합니다.
참고: 피험자는 표시된 경로를 평가하도록 지시받습니다(규칙을 따르고 있는지 여부와 관계없이 참가자에게 제공된 지침에 대한 자세한 내용은 5.12단계를 참조하십시오). - 추가 분석을 위해 로그 파일에 반응 시간(RT)을 저장합니다.
- 유지 보수 기간: 제어 기간이 끝나면 3초 동안 이동된 고정 십자가를 제시합니다.
- 계획 실행 기간으로 게이트 입구가 위치한 곳에 고정 십자가를 배치하여 다음 기간 동안 시선을 쉽게 그릴 수 있습니다.
- 제어 실행 기간: 미로를 다시 제시하는 동시에 실행 기간의 시작을 알리는 태그와 함께 EEG 및 호스트 시선 추적기 컴퓨터에 TTL 트리거를 보냅니다.
- 계획 실행 기간과 동일한 코드를 반복하여 응시 위치에 대한 온라인 피드백을 제공하고 추적된 경로와 응시를 묘사하고 겹칩니다.
- 경로를 추적하는 데 최대 10초의 시간을 확보하고 피험자가 버튼을 눌러 이 기간을 완료할 수 있도록 합니다.
- 추가 분석을 위해 RT를 로그 파일에 저장합니다.
- 제어 응답 기간: 제어 실행 기간이 종료되면 응답 기간을 나타내는 물음표를 제시합니다.
- 피험자가 조이스틱이나 키보드를 사용하여 대답하도록 각각 두 개의 버튼을 프로그래밍합니다.
참고: 여기서, 피험자들은 추적에 의해 표시된 순서가 정확한지 아닌지에 대해 하나의 버튼을 선택하고 잘못된/아니오에 대해 다른 버튼을 선택하여 대답하도록 요청받습니다. - 로그 파일에 정확도를 저장합니다.
- 피드백: 피험자가 올바르게 응답할 때마다 3초 동안 올바른 피드백을 전달하고(엄지 손가락 위로 이미지) 피험자가 잘못 응답할 때마다 3초 동안 잘못된 피드백을 제공하도록 코드를 작성합니다(엄지 손가락 아래 이미지).
- 계획 조건에서와 같이 TTL 트리거를 EEG로 보내고 올바른 피드백을 위한 태그와 잘못된 피드백을 위한 태그를 사용하여 시선 추적기 컴퓨터를 호스트합니다.
- 통제 기간: 교란 요인을 완화하기 위해 제어 조건 기간에 대한 코드를 작성합니다. 이 기간의 코드는 기준선으로 3초 동안 제시된 고정 십자가로 시작해야 합니다.
- 교육 과제: 자극을 생성하고, 코드를 작성하고, 앞서 언급한 계획 및 제어 작업 전에 각 조건(계획 및 제어)에 대해 약 6개의 시도/미로로 구성된 간단한 교육 세션을 제시합니다.
참고: 아이디어는 작업 설정에 익숙해지는 것입니다. 진행 기준을 설정하는 것이 좋습니다. 본 연구에서는 마지막 3건의 임상시험이 옳았고 참가자들이 훈련 세션이 끝날 때 목표와 절차를 이해했다고 보고한 경우, 참가자들은 실험 세션을 진행했습니다.

그림 3: 실험실 설정의 예. 상호 연결된 세 대의 컴퓨터를 보여주는 실험실 설정의 개략도. 호스트 컴퓨터(아이트래커 컴퓨터)는 안구 운동 데이터를 추적하고 저장하는 역할을 합니다. EEG 컴퓨터는 EEG 신호를 획득하고 저장합니다. 디스플레이 컴퓨터는 행동 실험을 제어하고, 피험자에게 자극을 제시하고, 병렬 포트와 LAN 연결을 통해 호스트 및 EEG 컴퓨터에 이벤트 트리거를 전송하여 데이터 수집을 동기화합니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 4: 시선 추적기 시스템이 제공하는 시각적 온라인 피드백에서 경로 재구성. 계획의 모터 실행(A, 자주색, 계획 실행 기간) 및 제어 실행 기간(B, 녹색 선) 및 시선 추적기 데이터를 사용하여 경로를 재구성하는 예시. 계획 실행 기간에 재구성된 경로는 각 계획 작업 시도의 정확성을 평가하는 데 사용됩니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
4. 실험실 설정 및 장비
- EEG 수집 시스템을 사용하여 참가자의 두피에서 EEG 활동을 기록하고, EEG 전극은 국제 10-20 시스템106에 따라 배치됩니다. 오프라인 재참조를 위해 참가자의 유양돌기에 두 개의 전극을 배치합니다. 근전도 전극을 사용하여 육안 검사 중에 수직, 수평 및 깜박이는 안구 운동 신호를 식별합니다.
- EEG 컴퓨터에서 2,048Hz 또는 1,024Hz의 샘플링 속도와 0.1-100Hz 사이의 대역 통과 필터로 EEG 데이터 수집을 위해 EEG 수집 소프트웨어를 사용하십시오.
참고: 1,024Hz 및 2,048Hz의 샘플링 속도는 저주파 진동을 분석하기에 충분한 해상도를 제공합니다. 저주파 진동을 분석할 때 저주파 신호가 앨리어싱되지 않도록 하기 위해 1,000Hz 이상과 같은 높은 샘플링 속도로 EEG 신호를 획득하는 것이 중요합니다. - 병렬 포트와 이더넷 을 통해 EEG 컴퓨터 및 호스트 시선 추적기 컴퓨터와 연결되고 행동 실험을 실행할 수 있는 플랫폼이 설치된 디스플레이 컴퓨터를 사용하여 최소 해상도 1,920 x 1,080 픽셀, 주사율 60Hz의 확장 모니터에 자극을 투사합니다. 이 모니터를 피사체에서 약 82cm 떨어진 곳에 놓습니다.
참고: 참가자로부터 82cm 떨어진 곳에 있는 144Hz로 재생 빈도가 설정된 24인치 모니터를 사용했습니다. EEG 및 안구 움직임을 기록하는 것과 관련된 인지 실험을 위해 화면 크기가 19인치 이상인 모니터를 사용하는 것이 좋습니다. 또한 일반적으로 참가자가 작업을 편안하게 수행하고 화면을 안정적으로 응시하면서 EEG 및 안구 움직임을 정확하게 기록할 수 있는 거리에 모니터를 배치하는 것이 좋습니다. 최상의 결과를 보장하기 위해 필요에 따라 설정을 테스트하고 조정하는 것이 좋습니다. - 시선 추적 시스템을 사용하여 참가자에게 실행 기간 동안 안구 움직임에 대한 실시간 피드백을 제공하고 동공 크기를 기록합니다. 적절한 시간 해상도를 위해 샘플링 속도를 1,000Hz로 설정합니다.
- 머리의 움직임을 피하십시오. 비디오 카메라의 시야 내에 시선을 유지하기 위해 왼쪽과 오른쪽, 위아래 머리 움직임 제한이 필요합니다. 비디오 카메라의 초점 범위 내에 시선을 유지하기 위해 앞뒤로 움직이는 제한이 필요합니다. 이마/턱 받침대를 조합하여 이 범위 내에서 움직임을 유지하십시오.
- 계획 자극과 제어 자극 간의 차이를 비교하기 위해 디지털 조도계 또는 이와 유사한 것을 사용하여 자극의 휘도를 평가합니다.
참고: t-test 또는 Wilcoxon과 같은 통계 테스트를 사용하여 두 조건의 자극 간의 차이를 평가할 수 있습니다. - 최소 4개의 버튼이 있는 제어 조이스틱 또는 키보드를 사용하십시오: 제어 조건의 예/아니오 질문을 위한 두 개의 버튼; 시험을 완료하기 위한 이 두 버튼 중 하나; 그리고 계획 조건의 응답 기간에 대한 또 다른 두 개의 버튼은 앞뒤로 이동하여 동물을 화면 하단의 4 개의 원 각각에 삽입합니다.
5. 뇌파 검사 및 시선 추적 기록 세션
- 연구를 시작하기 전에 참가자가 서면 및 서명된 정보에 입각한 동의서를 작성하도록 합니다.
- 녹음 세션 전에, 참가자들에게 화장을 하거나(마스카라와 아이라이너는 시선추적기 시스템에 의해 동공으로 감지될 수 있음), 약물이나 카페인을 복용한 경우(107,108), 또는 극심한 피로109(스트레스, 수면 부족 등)를 느끼지 않도록 요청한다.
- 참가자들이 훈련된 심리학자가 신청한 MINI-International 신경정신과 인터뷰97을 통해 성별, 연령, 손놀림, 모국어 및 신경정신과 병력에 대한 정보를 제공하기 위해 인구통계학적 설문조사를 완료하도록 합니다.
- 알코올 물티슈로 피험자의 이마, 두피, 유양돌기 및 근전도(EOG) 피부 위치를 청소합니다.
- 참가자에게 모든 외부 전극을 놓습니다. 수평 EOG를 양쪽 눈의 바깥쪽 안도에 양극으로 놓고 참가자의 오른쪽 눈 위와 아래에 수직 EOG를 놓습니다. 나중에 다시 참조할 수 있도록 오른쪽과 왼쪽 유양돌기에 두 개의 외부 전극을 놓습니다.
- 피험자의 머리를 측정하고, 확장된 국제 10-20 시스템에 따라 올바른 크기의 EEG 캡을 놓습니다. 이렇게 하려면 다음 단계에 따라 Cz 전극을 찾아 배치합니다.
- 헤어라인과 코 윗부분을 육안으로 검사하여 두피의 정중선을 확인합니다. 이 두 점을 연결하는 선을 식별하여 중간선을 정의합니다.
- Cz를 찾습니다. Cz는 일반적으로 두 귀 앞 지점(즉, 각 귀 바로 앞에 위치한 점) 사이의 중간점으로 정의됩니다. 이러한 점을 찾은 다음 Cz의 대략적인 위치를 식별하기 위해 이를 연결하는 선을 식별합니다.
- Cz를 측정하고 표시합니다. 나시온(즉, 코 꼭대기의 뼈 돌기)에서 Cz까지의 거리를 측정합니다. 나시온에서 Cz까지의 거리는 일반적으로 10-20 시스템에서 전체 머리 둘레의 약 53%입니다. 펜이나 다른 마킹 도구를 사용하여 Cz의 위치를 표시합니다.
알림: 오류를 최소화하고 EEG 데이터의 유효성을 보장하기 위해 전극 배치에 대한 일관되고 표준화된 절차를 따르는 것이 중요합니다. 표준 배치 절차를 개발하는 것이 좋습니다. 각 피험자의 두피에 전극을 삽입하기 위한 표준 절차를 만들고 모든 피험자에게 동일한 절차가 사용되는지 확인합니다. 녹음을 수행하는 팀이나 직원이 있는 경우 기술자 또는 연구 보조원에게 적절한 배치 절차에 대해 교육하여 전극을 일관되고 정확하게 배치할 수 있도록 합니다. 또한 각 피험자에 대한 전극 위치를 디지털화하는 것은 나중에 소스 분석을 수행하기 위해 바람직한 접근 방식이 될 것입니다. EEG 연구에서, 피험자의 머리에 있는 각 전극의 정확한 3차원(3D) 위치는 종종 EEG 데이터와 해당 뇌 활동(110) 사이의 상관관계를 확립하기 위한 요구 사항입니다. 이 정보는 또한 MRI 또는 CT 영상 기법으로부터 유래된 해부학적 이미지와 EEG 데이터의 적절한 정렬에 중요하다111,112.
- 뭉툭한 바늘이 있는 주사기를 사용하여 캡의 각 구멍에 전도성 젤을 삽입하고 끝으로 참가자의 머리카락을 제거합니다. 그런 다음 모든 두피 전극을 EEG 캡에 끼웁니다.
- EEG 기록 소프트웨어를 사용하여 임피던스를 확인하고 EEG 시스템에서 권장하는 저항 수준 미만인지 확인하십시오.
- 참가자에게 실험이 진행되는 동안 가능한 한 가만히 있으라고 요청합니다. EEG 신호를 검사하고 참가자에게 눈을 깜박이고 턱을 만들고 눈을 감고 몇 초 동안 머물도록 요청하여 테스트합니다.
- 참가자를 어둡고 소리가 감쇠된 방에 앉힙니다. 턱받침대를 사용하여 머리를 안정시키고 움직임을 최소화하고, 턱받침대와 자극 제시 화면의 중앙 사이에 약 82cm의 거리가 있는지 확인합니다.
- 응답을 위해 참가자 앞에 조이스틱 또는 키보드를 놓습니다.
- 지시: 각 상태(계획 및 통제)를 시작하기 전에 시각 자료를 사용하여 구두 지침을 제공합니다. 지침에는 자극의 시각적 예가 포함되고 계획 및 제어 조건에서 각각 미로를 해결하는 방법을 설명합니다.
- 계획 과제를 위해 피험자에게 미로의 다른 위치에 있는 특정 동물 위치(이 연구에서는 4개 위치)를 순서에 관계없이 일련의 규칙에 따라 방문 시퀀스를 완료할 수 있는 경로를 찾도록 지시합니다. (2) 관문에서 시작하여 네 번째 방문한 동물에서 길을 마칩니다. (3) 같은 경로나 모퉁이를 두 번 통과하지 마십시오. (4) 막다른 골목을 건너지 마십시오. (5) 길을 수직으로 건너지 마십시오"54.
참고: 규칙 2에서 규칙 5의 경우, 참가자에게 시각적 예시를 보여주는 것이 좋습니다. - 그 후, 6개의 시도로 구성된 계획 작업 교육 세션을 시작합니다.
참고: 참가자들에게 특히 실행 기간 동안 경로를 설명하는 동안 작업을 수행하는 데 문제가 있는 경우 각 시선 추적기 보정 기간 전에 보고하도록 지시합니다. 오프라인/사후 처리를 확인할 수 있는 평가판이 있는지 여부를 기록해 둡니다(6.1.1단계 참조). - 통제 조건의 경우, 피험자들에게 미로에 이미 표시된 경로가 이전에 학습한 규칙을 고려하여 올바르게 만들어졌는지 또는 잘못 만들어졌는지 평가하도록 지시합니다.
참고: 오류가 감지되었을 때 새 경로를 계획하지 않는 것과 같은 계획 전략을 사용하지 않고 미로를 평가하는 방법에 대한 시각적 지원이 포함된 예를 제공합니다(예: 동일한 경로를 두 번 사용하는 그림, 막다른 골목 통과 등). 오류가 발견되면 경로를 수정하는 것보다 오류 감지를 보고하는 데만 집중해야 합니다. 각 실험이 끝나면 참가자들에게 그들이 구현한 전략에 대해 질문합니다. 그런 다음 성과에 대한 구두 피드백을 제공하여 그려진 경로를 평가하고 새로운 경로를 계획하지 않았는지 확인합니다. 그 후, 6번의 시도로 구성된 제어 작업 교육 세션을 시작합니다.
- 계획 과제를 위해 피험자에게 미로의 다른 위치에 있는 특정 동물 위치(이 연구에서는 4개 위치)를 순서에 관계없이 일련의 규칙에 따라 방문 시퀀스를 완료할 수 있는 경로를 찾도록 지시합니다. (2) 관문에서 시작하여 네 번째 방문한 동물에서 길을 마칩니다. (3) 같은 경로나 모퉁이를 두 번 통과하지 마십시오. (4) 막다른 골목을 건너지 마십시오. (5) 길을 수직으로 건너지 마십시오"54.
- EEG 신호를 확인하여 모든 채널이 올바르게 획득되고 있는지 확인합니다. EEG 기록을 시작합니다.
- 아이 트래커를 보정합니다.
참고: 참가자가 화면의 여러 영역으로 시선을 돌릴 때 시선 위치를 결정하기 위해 시선 추적기의 기능을 확인하십시오.- 참가자에게 아이트래커가 보정될 것이며 흰색 원(작은 회색 점이 있음)이 화면의 네 모서리로 무작위로 이동하는 것을 보게 될 것이라고 알립니다(5점 보정 절차). 학생들에게 원에 시선을 고정시키라고 지시하고, 원이 다른 위치로 이동할 때 원의 위치를 따라가다가 다시 그 새로운 위치에 시선을 고정해야 한다고 알려줍니다.
- 실험을 실행하고, 출력/기록을 클릭하여 안구 움직임을 저장하기 시작하고, 참가자에게 이전에 제공된 지침을 따르도록 요청하여 이제 실험이 시작될 것임을 알립니다.
- 실험실을 어두운 환경에 두십시오. 동공 확장의 가장 큰 변화는 휘도113의 변화에 반응하여 발생합니다. 실험 환경에서 일정한 조도를 유지합니다.
6. 데이터 분석
- 행동 분석
- 통계 소프트웨어를 사용하여 행동 데이터를 분석합니다( 재료 표 참조). 정확도(정확한 응답의 백분율 비율)를 계획 조건과 통제 조건 모두에서 정량적 행동 매개변수로 측정합니다. 계획 조건의 경우 시선 추적기 데이터(시선 위치의 x 및 y 좌표)를 사용하여 실행 기간 동안 취한 경로를 오프라인으로 다시 만들고 실제 추적된 경로와 비교하여 계획된 경로의 정확도를 확인합니다(그림 4). 이렇게 하려면 응답 기간에 수행된 조합과 수행된 추적 간의 일치성을 수동으로 확인하십시오.
- 계획 기간 동안 미로를 해결하는 데 소요된 평균 시간과 제어 기간 동안 표시된 경로를 평가하는 데 소요된 평균 시간인 RT를 계산합니다.
- 계획 및 제어 조건에 대한 실행 기간의 평균 RT를 계산합니다. 구체적으로 말하자면, 올바른 시행에만 해당하는 RT를 사용하십시오.
참고: 보완적으로, Domic-Siede et al.54에 기술된 선형 통합 속도-정확도 점수(LISAS)114,115를 사용할 수 있으며, 이는 반응 시간과 정확도를 고려하는 결합된 측정을 제공합니다. 계획 실행 기간 동안의 반응 시간과 계획의 정확성은 상호 관련되어 있기 때문에 LISAS는 오류 수에 대해 수정된 반응 시간을 고려한 인덱스를 계산하는 데 활용할 수 있습니다. 또한 LISAS 지수는 전기생리학적 신호와 행동 수행 간의 상관관계를 평가하는 데 사용할 수 있습니다. 반응 시간(RT)과 오차 비율(PE)의 선형 조합으로 계산됩니다. - Levene 검정116,117과 같은 통계 검정을 사용하여 동질성을 평가하고, D'Agostino 및 Pearson 옴니버스 정규성 분포 검정118 또는 Shapiro Wilk 검정119를 사용하여 정규성을 검정하여 비교를 위한 적절한 통계 검정(모수 또는 비모수)을 선택합니다.
- Wilcoxon 부호 순위 검정120 또는 일치하는 쌍 t-검정121 을 사용하여 계획 조건의 계획 구성 요소가 제어 조건보다 인지적으로 더 까다로운지 여부를 평가하여 조건 간의 행동 매개변수를 비교합니다.
참고: 이러한 방식으로 행동 패러다임이 인지 계획을 평가하는 데 최적인지 검증합니다. - 계획 조건의 시행을 "쉬움"과 "어려움" 범주로 구분한 다음(2.2단계 참조) 매칭 페어 t-검정을 사용하여 "쉬움" 시행과 "어려움" 시행 사이의 계획 및 실행 기간의 정확도와 반응 시간을 비교합니다.
- EEG 및 안구 운동 전처리
- Delorme 및 Makeig122, Dimigen et al.123 및 Mognon et al.124에 설명된 것과 같은 자체 제작 스크립트 및/또는 확립된 도구 상자를 사용하여 프로그래밍 언어 소프트웨어에서 다음 사항에 설명된 EEG 데이터 전처리 파이프라인을 수행합니다( Table of Materials 참조).
- 안구 운동 활동을 EEG 기록과 동기화하여 더 나은 육안 검사 또는 추가 분석을 위해 고정, 단속 및 깜박임 이벤트를 가져옵니다(3.1.2단계 및 추가 파일 참조).
참고: 이 연구에서는 Domic-Siede et al.54 및 Dimigen et al.123 에 설명된 대로 시선 추적 데이터의 타임스탬프와 EEG 데이터의 타임스탬프를 사용하여 프로그래밍 언어 소프트웨어의 EEG 데이터로 안구 운동 이벤트를 가져왔습니다. - 계산 요구를 줄이기 위해 2,028Hz로 기록된 경우 데이터를 1,024Hz로 다운 샘플링합니다.
참고: 1,024Hz의 샘플링 속도는 4-8Hz의 관심 주파수 범위, 예상 주파수 분해능 및 분석의 계산 요구 사항에 따라 충분합니다. - EEG 신호를 유양돌기에 있는 전극의 평균으로 다시 참조합니다.
참고: 다른 참조가 가능합니다. 참조 선택은 EEG 분석 결과와 데이터 해석에 영향을 미칠 수 있으므로 다양한 참조 옵션의 장단점을 신중하게 고려하는 것이 중요합니다. 평균 유양돌기 기준은 계산하기 쉽고 안정적인 기준을 제공하기 때문에 EEG 연구에서 널리 사용되는 선택이며, 다양한 EEG 신호를 분석하는 데 효과적인 것으로 나타났습니다. EEG 데이터를 유양돌기의 평균(평균 유양돌기 기준이라고 함)으로 참조하는 것은 두피 EEG 데이터에서 전두엽 세타 활성을 분석하기 위한 일반적인 접근 방식입니다. 유양돌기 전극은 귀 근처에 위치하며 EEG 신호에 대한 기준을 제공합니다. 유양돌기의 평균을 참조하면 관심 있는 신호를 취소하는 것을 피하면서 관심 없는 노이즈 및 아티팩트의 영향을 줄이는 데 도움이 될 수 있으며, 이는 사용자가 EEG 신호를 보다 명확하게 표현하는 데 도움이 됩니다. - 프로그래밍 언어 소프트웨어를 사용하여 확장 신호(에포크 없음)에 대해 1Hz의 고역 통과 차단 주파수와 40Hz의 저역 통과 차단 주파수를 갖는 0위상 유한 임펄스 응답(FIR)을 적용합니다.
참고: 이 연구에서는 Delorme 및 Makeig122에 설명된 도구 상자를 사용했습니다. - 각 조건에 대해 시행 횟수를 고려하여 데이터를 각각 계획 기간과 통제 기간의 시작을 중심으로 한 에포크로 나눕니다. 미로 프레젠테이션이 시작되기 전 1초를 기준선으로 사용하고 계획 또는 통제 기간 후 4초를 관심 세그먼트로 사용합니다. 프로그래밍 언어 소프트웨어를 사용합니다.
참고: 이 연구에서는 Delorme and Makeig122및 36 epochs/trials에 설명된 도구 상자를 사용했습니다. - 종료 전 4초, 유지 보수 기간 후 1초를 사용하여 계획 및 제어 기간의 끝을 중심으로 하는 두 번째 세분화를 만듭니다.
참고: 계획 및 통제 기간(단계 6.2.6 및 단계 6.2.7)의 첫 번째와 마지막 4초를 선택하는 이유는 두 조건 모두에서 각 기간의 기간이 다를 수 있고 계획의 첫 번째와 마지막 초를 분석하면 계획 프로세스에 대한 보다 포괄적인 관점을 제공할 수 있기 때문입니다. 따라서 이러한 창 길이는 계획의 기초가 되는 진동 역학을 분석하기에 적절하고 충분합니다. - 분할된 신호에 대해 로지스틱 Infomax 독립 성분 분석(ICA) 알고리즘125를 실행하여 아티팩트 성분을 식별하고 제거합니다.
- Plöchl et al.126에서 권장하는 saccade-to-fixation variance ratio criterion을 사용하여 잠재적인 잡음이 있는 구성 요소를 자동으로 감지하고, Mognon et al.124에서 권장하는 공간 및 시간 특징의 공동 사용을 기반으로 하는 자동 EEG 아티팩트 검출기를 사용합니다.
참고: Pion-Tonachini et al.127에서 제안된 독립 성분 분류기를 사용하는 것이 좋으며, 이는 독립적인 성분 분류를 7개 범주에 걸친 조성 벡터로 추정하여 아티팩트를 식별하는 데 도움이 됩니다. - EMG, 전극 움직임 또는 뇌와 관련되지 않은 구성 요소와 같은 다른 잠재적인 인공물 구성 요소를 검사합니다. 시간 경과에 따른 지형, 스펙트럼 및 활성화를 시각적으로 검사하여 이러한 구성 요소의 거부를 검증합니다.
- 첨도 기준(chartosis criterion)을 사용하여 자동 채널 거부를 통해 잡음이 있는 채널을 보간(구형 보간)합니다(z-score를 임계값으로 5).
- 시간-주파수 분석
- 250ms의 윈도우 길이와 5ms의 시간 스텝을 사용하여 단시간 고속 푸리에 변환(FFT)(1Hz-40Hz)을 수행합니다. Hanning 창을 사용합니다. z-score를 사용하여 시간-빈도 차트를 기준선(-1초에서 -0.1초)으로 정규화합니다.
참고: 스펙트럼의 시각화는 창 크기와 시간 해상도 간의 절충에 따라 달라집니다. 4Hz에서 8Hz의 세타 범위를 포함한 전체 스펙트럼을 종합적으로 보려면 윈도우 크기의 하한인 250ms를 사용하여 각 시도 및 작업 중에 더 높은 시간 해상도를 보장하는 것이 좋습니다. 또한 이러한 경우에 일반적으로 선택되는 것으로 널리 알려진 Hanning 창을 사용하는 것이 좋습니다. 시간 및 빈도의 해상도를 개선하려면 추가 단계를 참조하십시오. - Fz와 같은 전두중심 전극 또는 전두엽 전극의 평균 그룹에서 시간-주파수 차트를 선택합니다.
참고: 인지 조절과 전두엽 정중선 세타 12,128,129 사이의 연관성에 관한 광범위한 증거를 고려하십시오. - Pz 및 Oz 전극과 같은 전극에서 비정면 제어 시간-주파수 차트를 선택하여 추가 비교를 수행합니다.
- 정면 전극과 제어 전극의 경우, 두 조건에서 시간-주파수 차트의 그룹 수준 비교를 위해 p-값이 0.05< 쌍을 이룬 샘플에 대해 비모수 클러스터 기반 순열 테스트를 수행합니다. 몬테카를로 방법을 사용하여 1,000번의 무작위 추첨을 합니다. 클러스터의 최대 통계값을 이용하여 치환 테스트130을 수행한다.
- 계획 및 제어의 처음 4초부터 각각 세타 주파수 대역(4-8Hz)과 마지막 4초 세그먼트에서도 평균을 구합니다.
- matched-pair t-test 또는 Wilcoxon signed-rank test를 사용하여 조건 간의 평균 theta activity를 비교합니다.
- 세타 활동의 시간 프로파일을 분석합니다. 이렇게 하려면 피험자별 시행 전반에 걸쳐 4-8Hz의 주파수 범위를 평균화합니다.
- Wilcoxon 부호 순위 테스트를 사용하여 거짓 발견 비율(FDR)과 짝을 이루고 보정한 조건 간의 세타 활동 역학을 비교합니다.
참고: Wilcoxon 테스트에서는 88ms 단계의 겹치지 않는 창을 사용했습니다.
- 250ms의 윈도우 길이와 5ms의 시간 스텝을 사용하여 단시간 고속 푸리에 변환(FFT)(1Hz-40Hz)을 수행합니다. Hanning 창을 사용합니다. z-score를 사용하여 시간-빈도 차트를 기준선(-1초에서 -0.1초)으로 정규화합니다.
- 소스 재구성
- 소스 분석 재구성을 위해 Tadel et al.131 에 설명된 오픈 액세스 툴박스 또는 다른 유사한 툴박스와 같은 툴박스를 사용합니다.
- Gramfort et al.133에 설명된 것과 같은 도구 상자를 사용하여 표준화된 저해상도 뇌 전자기 단층 촬영(sLORETA)132및 최소 노름 이미징 방법, 대칭 경계 요소 방법(대칭 BEM)과 같은 알고리즘을 사용하여 계획의 처음 4초 동안 전처리된 EEG 신호의 소스를 계산합니다.
- 전극의 3D 디지털화가 없는 경우 각 참가자의 기본 전극 위치와 함께 해부학적 MNI 템플릿(Brainstorm "Colin27"에서 MNI 템플릿 사용)에서 소스 알고리즘(이 연구에서는 sLORETA 알고리즘)을 사용합니다(5.6단계 참조).
참고: 기본 전극 위치를 사용하는 것이 뇌 활동의 원인을 결정하는 가장 효율적인 방법은 아니라는 점에 유의해야 합니다. 그러나 여전히 활동의 출처에 대한 일반적인 이해를 제공할 수 있습니다. 이러한 방법을 통해 얻은 국소화 소스는 대략적인 근사치이며 결과를 분석하는 동안 신중하게 해석해야 한다는 점을 명심하는 것이 중요합니다. - 전처리된 신호에 4-8Hz 대역통과 필터를 적용합니다.
- 시험이 시작되기 전 -1,000ms에서 -10ms까지의 기간을 기준으로 z-score 정규화를 적용합니다.
- 시험 시작 후 1초에서 4초 사이의 관심 시간 창을 사용하여 세타 활동의 평균을 구합니다.
- 몬테카를로 표본추출(1,000개 무작위화)131과 함께 비모수적 순열 부호 검정을 사용하여 조건 간의 평균 공간 소스를 비교합니다.
- 관심 영역(ROI)을 결정하려면 뇌 지도(brain atlas)를 사용하여 피질에 레이블을 지정합니다.
참고: Tadel et al.131에 설명된 도구 상자에 구현된 Destrieux Atlas134를 사용했습니다. - 뇌의 관심 영역(ROI)을 선택합니다.
참고: 양측 상전두회(bilateral superior frontal gyri, SF), 양측 횡전두엽 회(bilateral transverse frontopolar gyri) 및 설시(sulci, FP), 양측 ACC, 양측 MCC, 양측 배외측 전전두피질(bilateral dorsolateral prefrontal cortex) 137,138과 같은 전전두엽 피질 영역이 인지 조절 기능135,136에 관여한다는 근거를 고려했다. - 각 ROI에 대해 이전의 전처리된 EEG 신호(1-40Hz 범위)에 대해 주성분 분석(PCA)을 수행하고 각 ROI에 대해 PCA 분해의 첫 번째 모드를 취합니다.
- 단시간 고속 푸리에 변환을 사용하여 스펙트럼 분석을 수행하고, 비-파라메트릭, 클러스터 기반 치환 테스트(130)를 사용하여 관심 영역의 좌측과 우측 사이의 결과를 비교한다.
- 차이가 없는 왼쪽 및 오른쪽 ROI를 추출하여 하나의 양자 시계열(SF, ACC 및 MCC)로 표시합니다. 그런 다음, 시간-주파수 차트를 플로팅하고 조건 간에 비교합니다.
- 각 ROI에 대한 계획 작업의 복잡성 수준(쉬운 시도와 어려운 시도)에 따라 시간-빈도 차트를 비교합니다.
- 512개 샘플 각각에 대한 신호의 에지를 미러링하고 선택한 ROI에 대해 4Hz와 8Hz 사이의 대역통과 필터를 수행합니다.
- 힐베르트 변환을 적용하여 프로그래밍 언어 소프트웨어의 신호 처리 툴박스를 사용하여 순시 진폭139를 구합니다( Table of Materials 참조).
- z-점수 정규화(-1,000에서 -10ms를 기준선으로)를 사용하여 신호를 수정하고 피험자별로 시행 전체의 평균을 구합니다.
- Wilcoxon 부호 순위 테스트(일치하는 쌍, 1초의 겹치지 않는 창)를 사용하여 조건 간의 각 ROI 세타 대역 시간 프로파일을 비교하고 FDR로 수정합니다.
- EEG 활동과 행동 수행 간의 상관 관계
- z-score를 사용하여 ROI의 소스 시계열을 기준선으로 정규화합니다. 계획 또는 제어 시작 후 1초에서 4초 사이의 창을 선택합니다(시간-빈도 차트에서 두드러진 세타 활동이 관찰되는 위치).
- 제어 조건과 비교하여 계획 상태에서의 세타 활동의 증가를 확인하려면 먼저 관심 영역의 각 조건 및 소스에 대해 Chronux 툴박스(140 )와 같은 툴박스를 통해 멀티테이퍼 방법을 사용하여 신호를 주파수 영역(1-40Hz)으로 변환합니다.
- 세타 대역의 평균 주파수(4-8Hz)를 계산하고 세타 파워의 두 가지 측정값을 계산합니다: i) Δ 세타로 표시되는 계획 기간(θ 계획)과 제어 기간(θ 제어) 동안의 세타 파워의 차이, ii) Δ 세타(Δ θ)와 제어 기간(θ 제어) 동안의 세타 활동의 비율로 표시되는 세타 활동의 상대적 증가, Domic-Siede et al.54에서와 같이:
 (1)
(1)

- 두 가지 행동 매개변수를 계산한다: iii) Δ LISAS 계획은 LISAS 계획에서 LISAS 통제를 빼고, iv) Δ LISAS 계획 실행은 LISAS 계획 실행에서 LISAS 통제 실행을 뺀다. Domic-Siede et al.54:


- 계산된 전기생리학적 및 행동적 매개변수를 사용하여 Spearman의 rho 상관관계를 수행한 다음 FDR로 수정합니다.
- 안구 운동 분석: 서로 다른 진동 역학을 초래할 수 있는 각 조건에 대한 안구 운동의 전위차를 제어하려면 다음 분석을 수행합니다.
- 전체 시도에서 그리고 계획 및 제어 조건 동안 0초에서 3.75초까지 단속 진폭과 단속 최고 속도를 결정합니다.
- Wilcoxon 부호 순위 검정 또는 일치 쌍 t-검정 중 적절한 방법을 사용하여 결과를 비교합니다.
참고: Dimigen et al.123에 설명된 것과 같은 도구 상자가 도움이 될 수 있습니다. - Sato 및 Yamaguchi141에 설명된 대로 하나의 정면 전극(예: Fz 또는 평균 정면 ROI 전극)에서 푸리에 EEG 전력과 단속 속도 간의 일관성을 계산하고 평가합니다.
- Wilcoxon 부호순위 검정을 사용하여 두 조건 간 각 시행의 처음 4초에 대한 일관성 검정력 단속 비율 값을 비교합니다.
결과
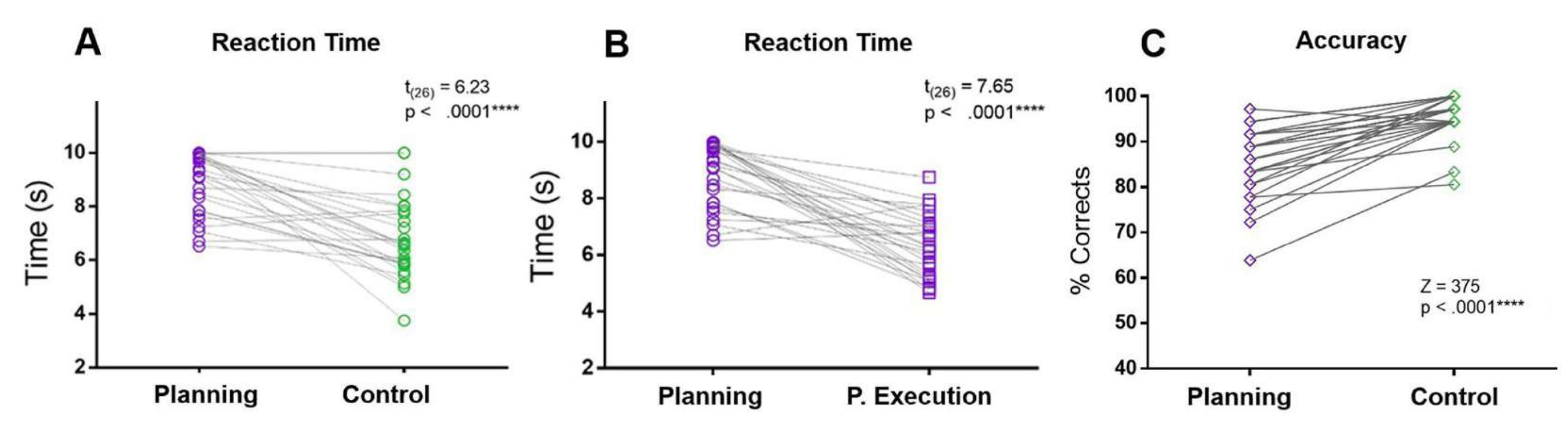
본 프로토콜에서는 계획 기간의 RT를 통제 기간 및 계획 실행 기간의 RT와 비교했습니다. 계획 RT는 통제 및 계획 실행 기간 RT보다 컸습니다. 또한 통제 조건과 비교하여 참가자는 계획 기간 동안 더 많은 실수를 저질렀고 정확도가 낮았습니다(그림 5).

그림 5: 계획 작업에 대한 반응 시간 및 정확도. 일치하는 쌍 t-검정을 사용하여 계획 기간(보라색 원)과 통제 기간(녹색 원)의 (A) 반응 시간을 비교합니다. (B) 일치하는 쌍 t-검정을 사용하여 계획 기간(보라색 원)과 계획 실행 기간(보라색 사각형)의 반응 시간을 비교합니다. (C) Wilcoxon 부호 순위 검정을 사용한 계획 조건(보라색 다이아몬드)과 통제 조건(녹색 다이아몬드)의 정확도 비교. 이 그림은 Domic-Siede et al.54에서 수정되었습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
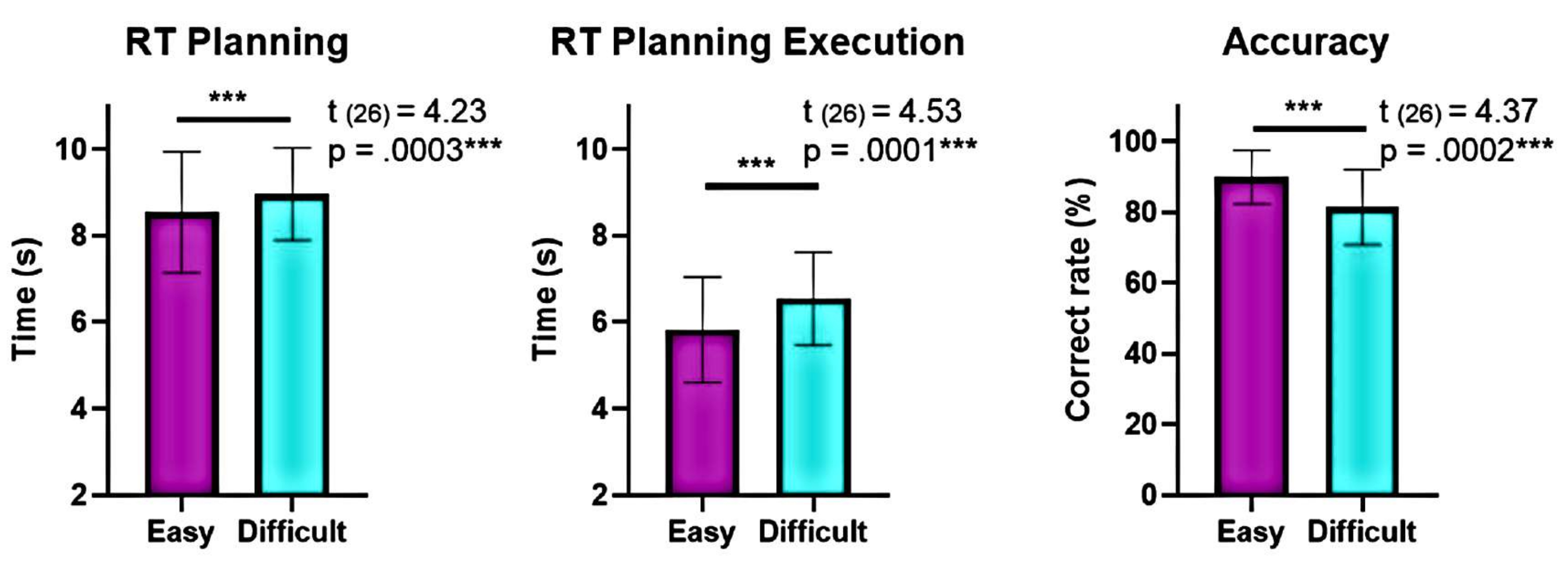
또한 계획 복잡성 수준에 대한 분석은 계획 및 실행 중 "어려움" 수준과 "쉬움" 수준 간에 정확도와 반응 시간(RT)에서 상당한 차이를 보여주었습니다(그림 6). "어려움" 레벨은 RT가 길고 정확도가 낮았습니다. 이러한 결과는 유효한 해결책의 수에 따라 임상시험을 나누는 것이 "쉬운" 임상시험과 "어려운" 임상시험을 구별할 수 있음을 시사합니다.

그림 6: 다양한 복잡성 수준에서의 행동 성능 비교. 복잡성의 "쉬운" 수준과 "어려움" 수준에서 행동 수행의 유의미한 차이는 일치하는 쌍 t-검정을 사용하여 확인되었습니다. 계획 및 실행 중에 "쉬움" 수준이 "어려움"에 비해 더 낮은 반응 시간(RT)이 나타났고 "쉬움" 수준에서 정확도가 더 높았습니다. 오차 막대는 SEM(평균의 표준 오차)을 나타냅니다. 이 그림은 Domic-Siede et al.54에서 수정되었습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
이러한 결과는 계획 구성 요소가 (명령 조작을 통해 ) 제어 조건에서 성공적으로 제거되었을 때 계획 작업이 인지적으로 더 복잡하고 까다롭고 도전적이며 시간이 많이 걸린다는 것을 나타냅니다. 따라서 작업에 의해 유도된 신경 상관관계는 서로 비교할 수 있습니다.
계획 중 정면 정중선 세타 활동을 분석하기 위해 Fz 전극을 계획하는 동안 평균 세타 주파수 대역을 제어 기간의 평균 세타 주파수 대역과 비교했으며, 계획 중 세타 대역 주파수가 크게 증가한 것으로 나타났습니다(그림 7).

그림 7: 인지 계획 중 정면 정중선 세타 활동. (A) 계획 작업(왼쪽), 제어 작업(가운데) 및 계획 효과(오른쪽) 동안 z-점수로 정규화된 모든 피험자에 대한 세타 밴드 파워를 나타내는 지형도. 인지 계획 동안 피험자들은 전두엽 정중선 세타 활동의 증가를 보였습니다. 색상 막대는 -0.5에서 1.5 사이의 z 값을 표시합니다. (B) 정합쌍 t-검정을 사용하여 전극 Fz(왼쪽), Pz(가운데) 및 Oz(오른쪽)에 대한 제어 기간(녹색)과 비교하여 계획 중 피험자(보라색)에 대한 세타 활동의 최소, 사분위수, 중앙값 및 최대 z-점수 값을 보여주는 바이올린 플롯. 이 그림은 Domic-Siede et al.54에서 수정되었습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
또한 관찰된 전두엽 세타 활동의 시간적 역학을 평가하기 위해 세타 밴드 전력의 특정 시점(750ms, 1,750ms, 2,750ms 및 3,750ms)에 해당하는 지형도를 공식화했습니다(그림 8A). 또한, 통제 기간과 비교했을 때, 시간-빈도 분석은 계획 기간이 시작된 후 1초 후부터 세타 활성이 유의미하고 점진적이며 지속적으로 증가하는 것으로 나타났습니다(그림 8B).

그림 8: 정면 정중선 세타 시간 역학. (A) 세타 활동의 지형학적 시간 조각. 계획 실행(계획 기간) 동안 시간에 걸쳐 전두엽 정중선 세타 활동의 점진적인 증가가 관찰되었습니다. 색상 막대는 z-점수 단위(−0.5 - 2.2)를 나타냅니다. (B) 계획 기간(위), 통제 기간(중간) 및 계획 효과에 대한 시간-빈도 차트로, 계획 기간(아래)에서 통제 기간을 빼서 계산합니다. 쌍을 이루는 표본에 대한 비모수 클러스터 기반 순열 테스트를 사용하여 결정된 유의하지 않은 픽셀은 계획 효과 플롯에서 더 밝게 표시됩니다. 색상 막대는 z-점수 단위(-4에서 4)를 나타냅니다. 이 그림은 Domic-Siede et al.54에서 수정되었습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
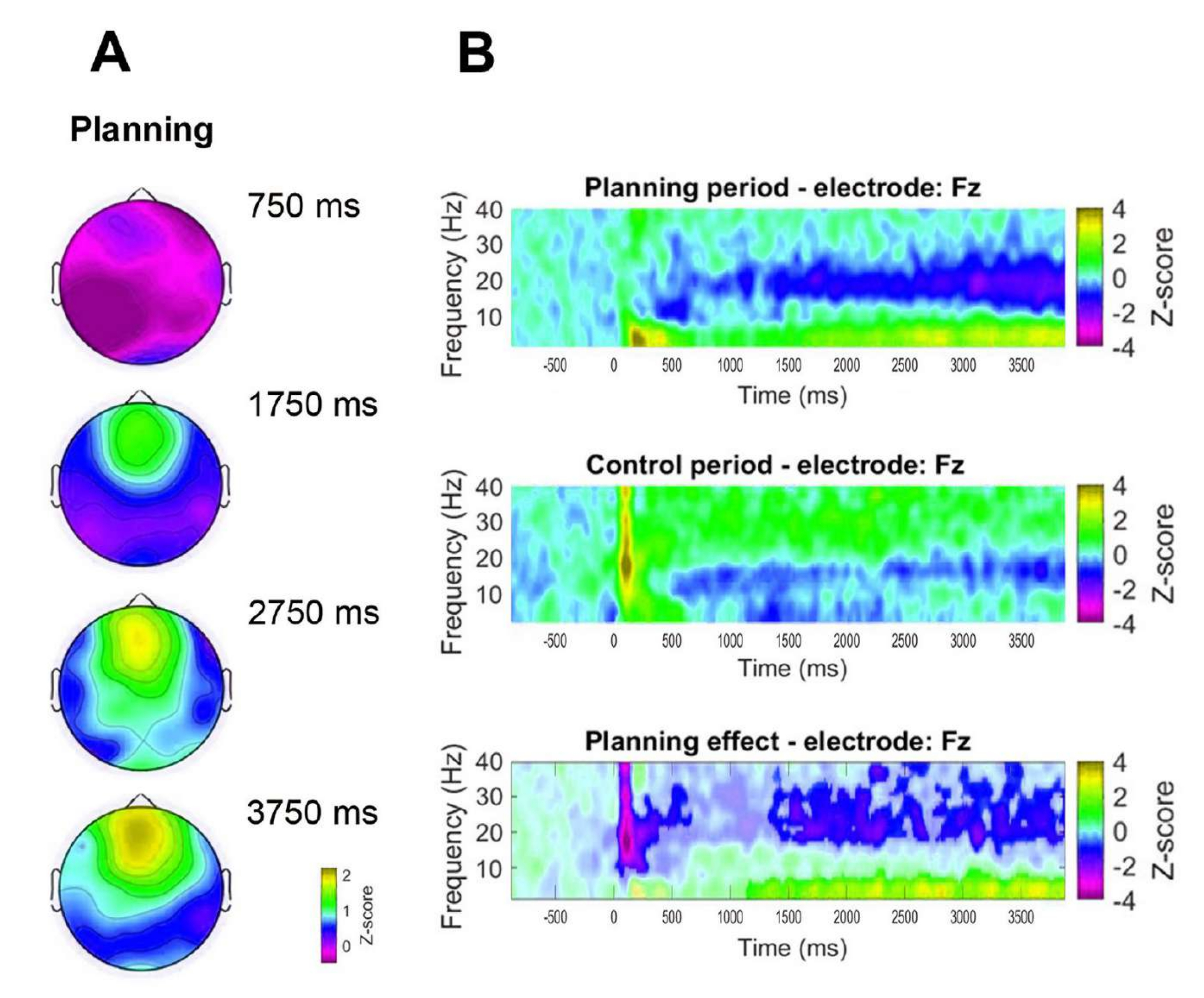{kind=link}
세타 활동의 원천 재구성을 위해 뇌 모델 템플릿을 시각화하고 조건 간에 비교했으며, 이는 세타 활동이 전전두엽 피질 영역(전두엽 상피질, FS, 전방 대상피질, ACC; 및 중대상피질, MCC) 내에서 발생했으며 조건 간에 상당한 차이가 있음을 나타냅니다(양측 SF, 양측 ACC 및 양측 MCC)(그림 9), 계획 기간 동안 더 높은 세타 활성이 관찰되었습니다(그림 9).

그림 9: 소스 재구성. sLORETA 알고리즘을 사용하여 다양한 뇌 소스의 세타 활동을 추정했습니다. 세타 활성은 4-8Hz 대역통과 필터링, z-점수 정규화, 기준선에 의해 수정, 계획 또는 대조 시작 후 각각 1초 또는 4초 사이의 평균을 측정하고 조건 간에 비교했습니다. 세타 활성의 현저한 증가는 양측 전두엽 상영역(bilateral frontal superior area), 양측 전방 대상피질(anterior cingulate cortex) 및 양측 중대상피질(bilateral mid-cingulate cortex)에서 발견되었습니다. 이 그림은 치환 테스트에서 유의한 t-값을 보여줍니다. 약어: FS = frontal superior; ACC = 전대상피질(anterior cingulate cortex); MCC = 중간 대상 피질. 이 그림은 Domic-Siede et al.54에서 수정되었습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
그 후, 힐베르트 변환을 계산하여 각 소스에 대한 시간 경과에 따른 세타 변화의 시간 프로파일을 평가한 다음 조건 간 세타 활성의 순간 진폭을 비교했습니다. 우리는 좌측 전두극, 양측 ACC 및 양측 MCC 소스가 계획 기간 시작 후 더 높은 세타 활성을 나타내는 것을 발견했습니다(그림 10). 이러한 결과는 인지적 계획을 요구하는 우리의 실험 패러다임이 PFC 영역 내에서 발생하는 세타 활동을 유도했음을 시사합니다.

그림 10: PFC 소스의 세타 활동 시간 프로필. 힐베르트 변환으로 계산된 순간 진폭은 각 전두엽 소스에 대한 PCA 분해의 첫 번째 구성 요소에 적용되었으며 시간 경과에 따른 전두엽 세타 활성을 보여주기 위해 조건과 기준선 모두 z-점수로 정규화되었습니다. 회색 음영 영역은 FDR에 의해 수정된 1초(Wilcoxon 부호 순위 테스트) 단계가 있는 겹치지 않는 이동 창을 사용하여 결정된 상당한 차이를 보여줍니다. 음영 처리된 영역은 95% 신뢰 구간을 나타냅니다. 좌측 FP 영역, 양측 ACC 및 양측 MCC는 계획 시작 후 세타 활성이 증가한 것으로 나타났습니다. 약어: ACC = anterior cingulate cortex; MCC = 중간 대상 피질. 계획 조건은 자주색으로 표시됩니다. 제어 조건은 빨간색으로 표시됩니다. 이 그림은 Domic-Siede et al.54에서 수정되었습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
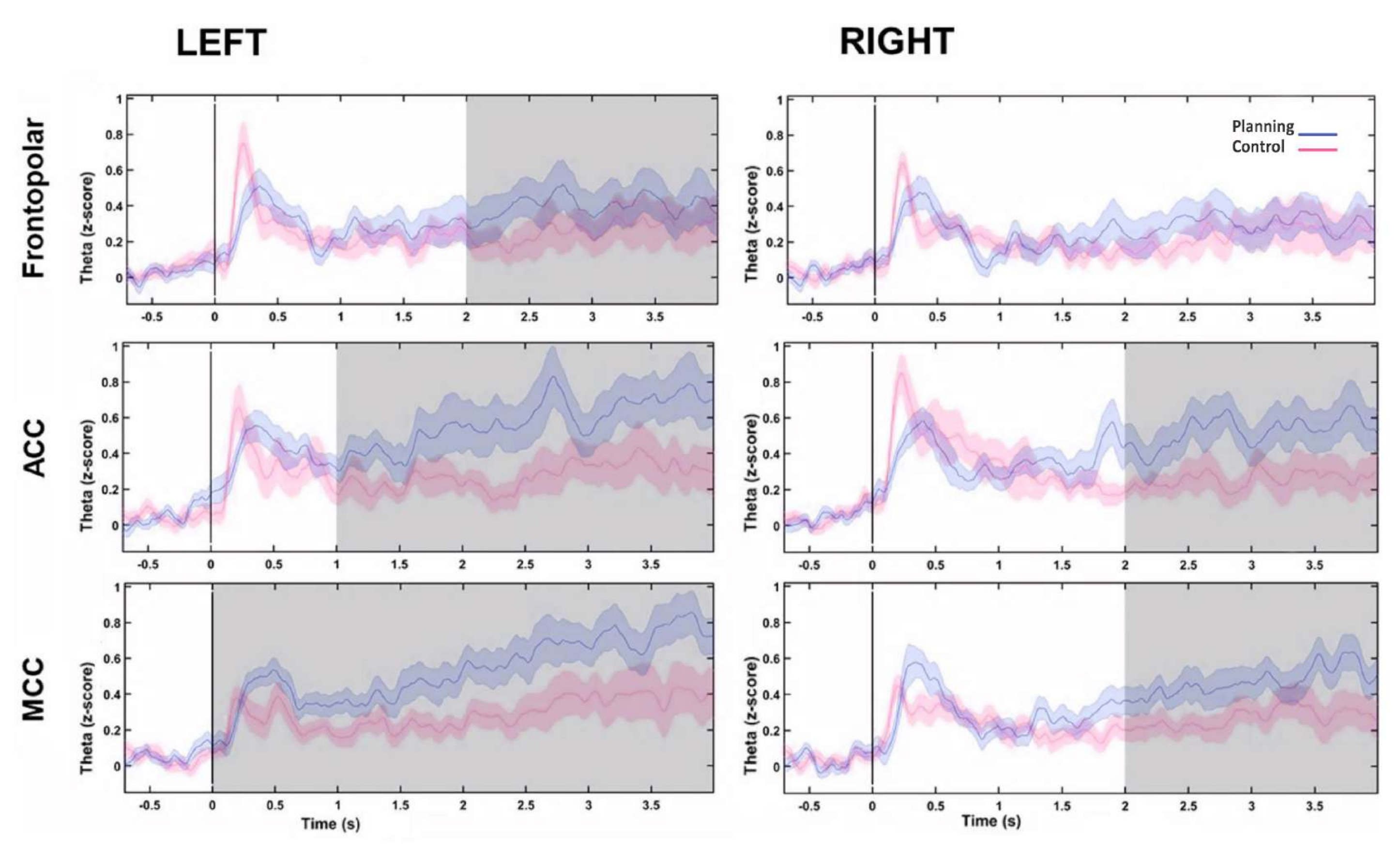{kind=link}
또한, 우리는 행동 결과에서 알 수 있듯이 복잡성 수준과 관련하여 계획 중 스펙트럼 기능의 변화를 조사하는 것을 목표로 했습니다. 주목할 점은 알파 대역 내의 왼쪽 ACC에서만 상당한 불일치가 발견되었다는 것입니다. 이는 우리의 계획 작업이 세타 진동의 변화를 통해 계획의 본질적인 측면을 인지 제어 작업에서 일반적으로 발생하는 일반적인 인지적 요구(노력)보다 더 크게 평가한다는 개념을 뒷받침합니다(그림 11).

그림 11: 계획 복잡성 수준에 따른 EEG. ROI 시간-빈도 차트는 "어려움" 수준에 대해 좌측 전대상피질(ACC)에서만 독점적으로 알파 대역에서 유의한 양의 클러스터를 보여주었습니다. 쌍을 이루는 샘플에 대한 비모수 클러스터 기반 순열 테스트를 사용하여 결정된 유의하지 않은 픽셀은 플롯에서 더 밝은 음영으로 표시되며, 색상 막대는 -3에서 3까지의 z-점수 단위를 나타냅니다. 이 그림은 Domic-Siede et al.54에서 수정되었습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
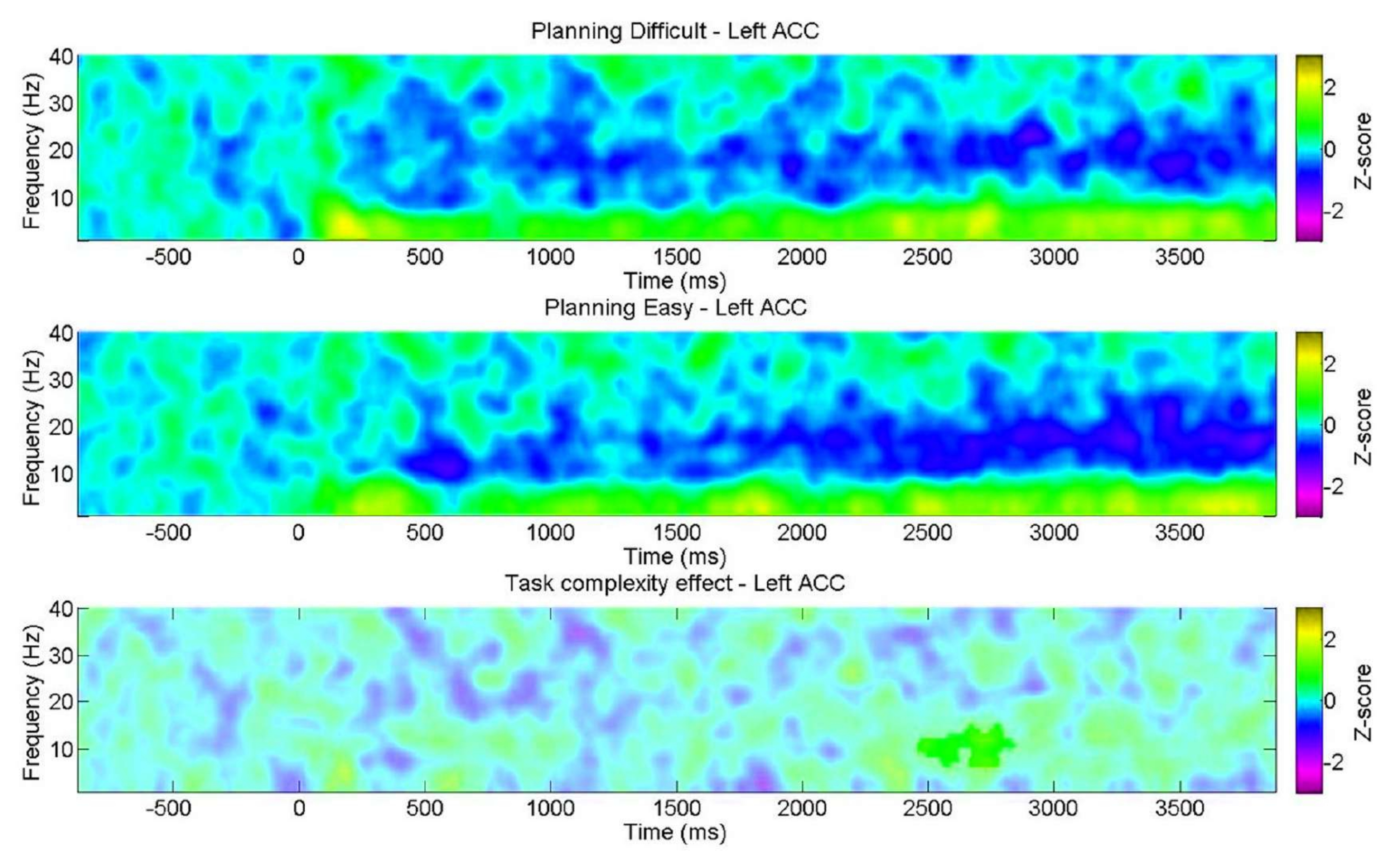{kind=link}
세타 활동과 행동 수행 사이의 상관 관계를 수행했을 때 음의 상관 관계가 관찰되었습니다. 구체적으로, 계획 기간 동안 좌측 전두극 영역에서의 세타 활동이 증가함에 따라 LISAS 계획 실행 점수가 감소했습니다(그림 12). 이 패턴은 계획을 수립하는 동안 계획을 성공적으로 실행하기 위해 왼쪽 FP 영역이 필요할 수 있음을 반영할 수 있으며 세타 활동에 대한 역할을 제안합니다.

그림 12: 세타 활동 및 행동 성과. 좌측 전두극 피질의 세타 활동과 Δ LISAS 계획 실행 사이의 Spearman's rho 상관관계는 유의한 음의 상관관계를 보여주었습니다. 이 그림은 Domic-Siede et al.54에서 수정되었습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
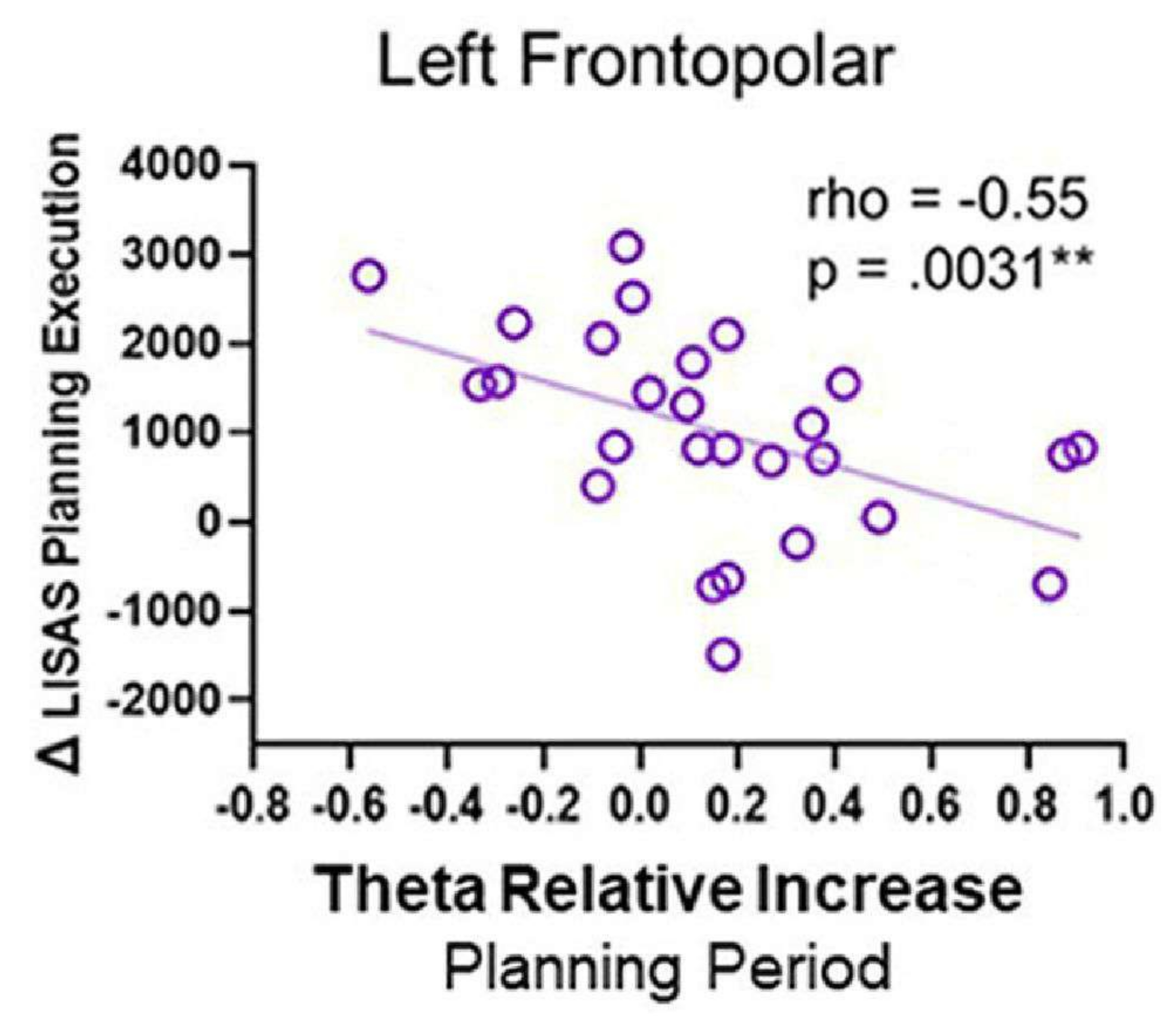{kind=link}
또한, 각 조건에 의해 유발되는 다양한 인지적 요구와 목표는 계획 조건과 통제 조건 사이에 대조적인 안구 움직임을 유발하여 다른 진동 활동 패턴을 유발할 수 있습니다95. 위의 문제를 해결하기 위해 다양한 수준에서 단일 피험자, 단일 시험 데이터를 분석했습니다. 특히, Fz 채널 시계열 및 세타 활동 시간 역학은 시간 경과에 따른 단속 속도와 관련이 없는 것으로 나타났습니다(그림 13A).

그림 13: EEG 및 안구 운동 기록 결과. (A) 행은 EEG(위), 시간-빈도 차트(가운데 위), 전극 Fz의 세타 시간 프로파일(가운데 아래), 계획 조건에서 피험자 8 및 시험 9(아래)의 단속 속도를 나타냅니다. (B) 조건 간의 단속 진폭, 단속 피크 속도 및 전력 단속 속도 일관성에 대한 Wilcoxon 부호 순위 테스트 비교가 표시되며, 이는 계획 조건과 제어 조건 간의 단속 진폭에서 상당한 통계적 차이를 나타냅니다. SEM은 오차 막대로 표시됩니다. 이 그림은 Domic-Siede et al.54에서 수정되었습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
다음으로, 전체 시험에서 비교를 위해 0초에서 3.75초까지의 단속 진폭과 피크 속도를 얻었습니다(그림 13B). 우리는 saccade 진폭이 제어 조건에서 더 크다는 것을 발견했습니다. 그러나 전극 Fz에서 푸리에 세타 전력과 단속 속도 사이의 일관성 지수 조건 간에 통계적으로 유의미한 차이는 발견되지 않았으며(그림 10B), 이는 단속과 세타 활성 간의 잠재적 관계가 조건 전반에 걸쳐 일관되었음을 나타냅니다.
종합하면, 이러한 결과는 설명된 실험 프로토콜이 인지 제어 기능으로서의 인지 계획을 연구하는 데 적합하다는 것을 뒷받침합니다.
토론
여기에 설명된 프로토콜은 진동 및 동공 역학과 같은 적절하고 보완적인 행동 및 생리학적 측정과 관련하여 새롭고 생태학적 계획 작업 중에 인지 계획 및 인지 제어를 평가하기 위한 혁신적인 접근 방식을 제공합니다. 실험 중에는 참가자들이 계획 작업을 수행하는 동안 EEG 활동이 기록되었으며, 참가자들은 먼저 계획을 정교화한 다음 실행하도록 지시받았습니다. 동물원 지도에 미리 그려진 경로를 평가하는 것과 관련된 제어 조건은 유사한 설정과 구조를 유지하면서 인지 계획 측면을 제거하기 위해 설정되었습니다. 이 접근 방식을 사용하면 인지 제어 기능으로서의 인지 계획이 PFC 영역에서 전두엽 세타 활동의 생성으로 이어지는지 여부와 다른 PFC 세타 진동 소스가 계획 수행의 다른 측면과 연결되어 있는지 여부를 평가할 수 있습니다. 이 프로토콜을 사용하여 평가할 수 있는 또 다른 측면은 계획 정교화, 계획 실행, 계획 달성 및 피드백 처리와 같은 계획 단계 중에 관련된 다양한 인지 프로세스의 차별화입니다. 우리는 계획이 인지 통제와 관련된 표준 전두엽 세타 활동을 유도하여 목표를 효율적으로 달성하는 데 기여한다는 것을 발견했습니다. 이러한 결과는 이 프로토콜의 관심을 뒷받침합니다.
인지 신경 과학의 광대한 발전에도 불구하고 대부분의 신경 영상 실험은 감각 운동이 결핍된 환경에서 인공 작업을 사용하여 고립된 인지 기능을 조사하고 혼란을 제어하기 위해 과도하게 단순화된 자극을 조사합니다. 문제적으로, 이러한 실험은 인지 기능이 일상적인 상황(생태학적 상황에서)에서 구현될 때 관련된 실제 뇌 메커니즘을 식별하지 못할 수 있습니다.138,140). 특히, 목표를 수립하고, 계획을 세우고, 계획을 효과적으로 실행하는 능력은 다양한 인지 기능(작업 기억, 억제 제어, 인지 유연성 등)을 필요로 하기 때문에 평가하기 어렵습니다.104,144명. 따라서 생태 행동 과제를 설계하는 것은 인지 신경 과학의 현재 경향에 따라 권장되고 제안됩니다 142,143,145,146.
우리의 계획 작업은 실험실 환경(화면에 자극이 표시되는 방 안)에서 이루어졌음에도 불구하고 참가자들이 화면에서 상호 작용할 수 있는 의미 있는 자극과 목표를 사용하여 참가자를 위해 매력적이고 상호 작용적으로 만들어졌습니다. 또한 이 작업을 수행하려면 참가자가 다양한 위치를 방문할 수 있는 경로를 계획하는 실제 상황에 참여해야 합니다. 생태학적 과제 설계를 갖기 위해서는, 패러다임은 피험자가 일상 생활에서 해야 하는 것과 유사한 방식으로 특정 행동이나 인지 기능을 수행하도록 도전해야 한다 62,63,147. 생태학적 과제 설계를 개발하기 위해, 여기에서 사용되는 계획 과제는 여러 단계에서 다른 장소를 방문하는 경로를 계획하는 것을 포함한다56. 첫 번째 단계는 참가자가 일련의 규칙을 따르도록 하면서 계획을 세우는 것입니다. 두 번째 단계는 작업 메모리에서 계획을 유지 관리하는 것이고, 세 번째 단계는 계획을 실행하고 규칙 준수 여부를 모니터링하는 것입니다. 이 단계는 계획의 여러 단계와 인지적 유연성, 억제 제어 및 작업 기억과 같은 다른 실행 기능의 오케스트레이션을 나타냅니다. 타당한 생태학적 인지 과제 설계를 갖기 위해서는, 과제는 일상 생활에서 특정 인지 기능을 수행하는 데 어려움을 겪는 정신 장애 또는 인지 장애 환자의 특정 인지 장애를 감지할 수 있어야 한다105. 이는 이 프로토콜을 사용하는 향후 연구를 통해 달성할 수 있습니다.
이 프로토콜의 사용을 통해 얻은 행동 결과는 실험적 예측과 일치했습니다. 통제 조건을 형성하기 위해 계획 구성 요소를 제어 작업에서 제거했을 때 행동의 상당한 차이가 관찰되어 추가 비교를 용이하게 했습니다. 계획 조건은 반응 시간 및 정확도와 같은 매개변수에 의해 입증된 바와 같이 제어 조건보다 인지적으로 더 까다롭다는 것이 밝혀졌습니다. 이것은 계획 23,55,56,57,148,149,150의 실행에 높은 인지 기능의 참여가 증가했음을 반영 할 수 있습니다.
더 빠른 반응 시간, 더 나은 성능 및 필요한 다양한 인지 프로세스(규칙 평가)에 의해 입증된 바와 같이 제어 조건이 덜 복잡한 인지를 포함한다는 점을 고려할 때, 가능한 대안 수정은 계획 작업에 존재하는 복잡성 수준을 활용하고, 이를 조작하고, 다양한 복잡성 수준(예: 시행 횟수를 늘리고 어려움, 중간, 쉬움 시행 조건을 만듭니다). 그러나, 본 임상시험계획서의 결과는 행동적 측정에 기초하여 "쉬운" 임상시험과 "어려운" 임상시험을 구별하는 것이 가능했지만, 전기생리학적 측정에서는 차이가 발견되지 않았다는 것을 보여주었다. 이는 본 프로토콜의 결과가 주의력, 정신적 노력, 난이도 또는 높은 수준의 인지적 요구와 같은 인지적 통제의 광범위한 측면보다는 계획 기능의 내재적 특성을 더 정확하게 반영한다는 것을 시사한다54. 그럼에도 불구하고, 추가 연구에서는 4개의 동물 위치를 방문하는 표시된 경로를 따라가는 것과 같은 다른 유형의 제어 조건을 고려할 수 있지만 순서도 기억할 수 있습니다. 이런 식으로, 난이도를 더 잘 제어할 수 있고, 계획을 작업 기억과 구별할 수 있지만, 이것의 한 가지 가능한 단점은 피험자가 두 가지 매우 까다로운 작업을 수행해야 하기 때문에 피로입니다.
여러 연구에서 다양한 안구 운동 매개변수를 특정 인지 사건과 연결했습니다. 한편으로, 특정 연구에서는 인지 작업 중 세타 진동과 동공 직경 사이의 상관 관계를 발견하여 인지 기능의 두 가지 측정 사이의 관계를 시사합니다. 예를 들어, Lin et al.152 은 주관적 갈등의 다양한 정도를 반영하여 중전두엽 세타 활동과 동공 크기의 변화 사이의 상관관계를 발견했습니다. 그들의 연구 결과는 이러한 신호가 갈등 처리, 주의력 증가 및 유연한 행동 반응을 나타낸다는 것을 시사합니다. 따라서 중전두엽 세타 활동과 동공 반응 사이의 관계는 의사 결정 과정에서 비용과 이점을 평가하는 데 중요한 역할을 하는 것으로 보입니다. 또 다른 연구에서, Yu 등[153 ]은 제어 억제 반응을 조절하는 작업 기억 작업을 통해 작업 시간 참여가 신경 생리학적으로 인지 제어에 어떻게 영향을 미치는지 조사했습니다. 연구진은 동공 직경 데이터와 전두엽 세타 활동 사이의 관계를 연구한 결과, 과제 시간이 길어질수록 성능이 감소하며, 이는 동공 확장 조절과 전두엽 세타 활동의 감소를 동반한다는 것을 보여주었습니다. 과제를 시작할 때 그들은 작업 참여, 세타 활동 및 인지 제어 사이에 강력한 상관관계가 있음을 발견했으며, 이는 주로 높은 작업 기억과 억제 제어가 필요한 까다로운 작업에 대한 동공 확장 조절로 나타났습니다. 그러나 이 관계는 말기로 갈수록 소멸되었고, 이는 투자한 노력과 작업을 수행하는 데 사용되는 인지 제어 사이의 단절을 의미하며, 이는 전전두엽 작업 시간 효과의 특징이다153. 반면에, 다른 연구에서는 단속과 진동을 조사했습니다. 예를 들어, Nakatani 등[154 ]은 지각 과제에서 후두 영역의 알파 밴드 진폭이 깜박임과 단속 효과를 예측한다는 것을 밝혔습니다. 더욱이, Velasques 등[155 ]은 프로사카딕 어텐션 과제 동안, 단속 진폭이 전두엽 감마 변화와 연관되어 있음을 보여주었다. 또한, Bodala 등[156 ]은 전두엽 정중선 세타의 감소가 지속적인 주의력의 감소뿐만 아니라 단속의 진폭과 속도의 감소를 동반한다는 것을 발견했습니다. 이러한 결과는 안구 운동, 특히 단속이 EEG 신호의 배경 소음에 기여하는 것이 아니라 인지 과정을 반영할 수 있음을 시사합니다. 본 연구에서는 사카데-고정 분산 비율 기준126과 함께 ICA 알고리즘을 사용하여 안구 운동 관련 아티팩트의 제거를 향상시켰습니다. 이 기준은 무료 보기 작업에 대한 아티팩트 제거를 향상시킵니다(157). 본 연구에서는 조건 전반에 걸쳐 saccade peak 속도와 theta power와 saccade rate 간의 일관성에서 차이가 관찰되지 않았습니다. 그러나 이러한 질문을 해결하기 위해서는 더 많은 연구가 필요합니다.
이 프로토콜을 사용하는 데 있어 중요한 단계는 카메라의 시선 데이터가 손실되면 작업을 오염시키고 정확한 응답을 얻기 어렵게 만드는 오류가 발생할 수 있으므로 실험 중에 시선 추적기를 지속적으로 보정하는 것입니다. 따라서 가능한 한 자주 교정하는 것이 중요합니다. 그러나 보정된 시선추적기를 사용한 임상시험 횟수와 실험 기간 사이에는 상충 관계가 있습니다. 본 연구에서는 5건의 임상시험마다 보정하기로 결정했다.
이 계획 작업 동안 세타 진동과 동공 직경 사이의 관계를 탐구하는 향후 연구가 수행되어야 합니다. 계획은 주의 자원의 할당과 여러 인지 과정의 조정을 필요로 하는 집행 통제의 중요한 측면입니다. 계획 작업 중 세타 진동과 동공 직경 사이의 관계를 이해하면 실행 제어의 기본 신경 메커니즘과 시간이 지남에 따라 어떻게 변화하는지에 대한 귀중한 통찰력을 얻을 수 있습니다. 또한 이러한 연구는 피로 또는 주의력 상실과 같은 인지 기능의 변화가 계획 작업의 성과와 자원을 효과적으로 할당하는 능력에 어떤 영향을 미치는지에 대한 더 깊은 이해로 이어질 수 있습니다. 이 정보는 인지 훈련 프로그램이나 주의력 결핍/과잉 행동 장애(ADHD)와 같은 상태에 대한 치료와 같은 계획 성과를 개선하기 위한 개입을 개발하는 데 중요한 의미를 가질 수 있습니다.
이전 연구에서는 PFC가 인지 계획에서 중요한 역할을 한다는 것을 보여주었으며, 이는 본 연구의 결과에서 확인되었습니다. 이러한 결과는 인지계획이 PFC, 특히 전대상피질(anterior cingulate cortex), 중대상피질(mid-cingulate cortex) 및 상전두엽(superior frontal regions)에서 FMθ 활성을 유도한다는 것을 보여준다54. 이러한 결과는 집행 기능에 대한 이전 연구와 일치합니다. FMθ 활동이 까다로운 작업 중에 뇌 영역 간의 제어를 시작하고 의사소통을 촉진하기 위한 하향식 과정으로 작용한다는 생각을 뒷받침하는 상당한 증거가 있습니다13. 인지 조절과 관련된 FMθ 활동의 시간적 역학을 조사한 연구는 소수에 불과하지만, FMθ의 시간 프로파일이 인지 조절의 다양한 측면과 뚜렷한 PFC 영역의 관여에 대한 정보를 제공할 수 있다는 데 널리 동의하고 있습니다. 인지 계획을 평가하기 위한 프로토콜을 사용하여 계획 중 FMθ 활동의 시간 프로필을 특성화할 수 있었습니다. 구체적으로, 계획 조건 동안의 FMθ 활동은 점진적인 증가를 보였다. 이 프로토콜을 구현함으로써 처음으로 FMθ가 다른 고차 인지 기능과 마찬가지로 계획 구현 중에도 존재하며 시간적 역학이 인지 제어의 지표 역할을 할 수 있음을 입증했습니다.
우리의 결과와 프로토콜은 가상 신경 심리학적 평가를 개선하고 우울증 및 주의력 결핍/과잉 행동 장애와 같은 관련 인지 계획 문제가 있는 신경 정신 장애의 치료를 포함하여 신경 과학 분야에서 잠재적으로 응용될 수 있습니다. 예를 들어, 평가에는 행동 수행 수준에서의 다양한 오류 패턴, 전기 생리학적 수준에서의 다양한 진동 패턴 및 다양한 안구 움직임을 조사하는 것이 포함될 수 있습니다. 또한 이 작업의 결과는 인지 계획 능력을 향상시키는 것을 목표로 하는 뇌-컴퓨터 인터페이스 및 인지 훈련 프로그램의 개발에 영향을 미칠 수 있습니다.
본 프로토콜은 신경 전형 및 신경 정신 질환 인구에서 인지 계획의 애매한 인지 제어 기능을 뒷받침하는 신경 메커니즘을 이해하는 데 새로운 증거에 기여할 수 있습니다. 더욱이, 우리의 행동 패러다임은 세타 진동의 변화에 반영된 인지 통제 과제에 일반적으로 존재하는 일반적인 인지적 요구가 아닌 계획의 본질적인 측면을 조사하는 실용적인 계획 과제와 함께 전기생리학적, pupillometry 및 행동 측정의 검토를 통해 인지 제어 및 계획의 신경생물학에 대한 통찰력을 제공할 수 있습니다.
공개
저자는 공개할 내용이 없습니다.
감사의 말
이 연구는 ANID 21150295의 박사 장학금 프로그램 Becas de Doctorado Nacional año 2015, FONDECYT 정기 보조금 1180932, FONDECYT 정기 보조금 1230383, FONDECYT de Iniciación 보조금 11220009, Postdoc 보조금 Universidad de O'Higgins 및 Institut Universitaire de France(IUF)의 재정 지원을 받았습니다. 패러다임 디자인에 대한 피드백을 주신 Pablo Billeke 교수님께 감사드립니다. 시간-주파수 분석 코드를 공유해 주신 Eugenio Rodríguez 교수님께 감사드립니다. 마지막으로, Milan Domic, Vicente Medel, Josefina Ihnen, Andrea Sánchez, Gonzalo Boncompte, Catalina Fabar, Daniela Santander에게 피드백을 주신 데 감사드립니다.
자료
| Name | Company | Catalog Number | Comments |
| EEG System | Biosemi | ActiveTwo Base system, 64 channels | |
| Eye-tracker System | Eyelink SR Research | EyeLink 1000 Plus Core Unit, High-speed camera, Host PC/Monitor, | |
| CPU display | Intel | Hard drive 221 GB, Processor Intel Core i7-4790 3.60 Hz, OS Windows 7, 4GB RAM | |
| CPU EEG | Intel | Hard drive 223 GB, Processor Intel Core i7-4790 3.60 Hz, OS Windows 7, 4GB RAM | |
| Monitor | ASUS | ASUS VG248QE 24" LCD monitor | |
| Joytsick | Logitech | Model F310 | |
| Luxmeter | Focket | LCD screen (0-200.000 lux) model Liebe WH LX1330B | |
| Statistics software | GraphPad Prism | GraphPad Prism version 8 for Windows | |
| MATLAB Programming Software | The MathWorks | MATLAB R2014a and R2018b | |
| SVG tool Inkscape | Inkscape Project | vector graphic editor software | |
| Presentation Software | Neurobehavioral Systems | stimulus delivery and experiment control program for neuroscience |
참고문헌
- Siegel, M., Donner, T. H., Engel, A. K. Spectral fingerprints of large-scale neuronal interactions. Nature Reviews Neuroscience. 13 (2), 121-134 (2012).
- Fries, P. A mechanism for cognitive dynamics: Neuronal communication through neuronal coherence. Trends in Cognitive Sciences. 9 (10), 474-480 (2005).
- Fries, P. Rhythms for cognition: Communication through coherence. Neuron. 88 (1), 220-235 (2015).
- Thut, G., Miniussi, C., Gross, J. The functional importance of rhythmic activity in the brain. Current Biology. 22 (16), 658-663 (2012).
- Fröhlich, F., McCormick, D. A. Endogenous electric fields may guide neocortical network activity. Neuron. 67 (1), 129-143 (2010).
- Spaak, E., de Lange, F. P., Jensen, O. Local entrainment of alpha oscillations by visual stimuli causes cyclic modulation of perception. Journal of Neuroscience. 34 (10), 3536-3544 (2014).
- Fiebelkorn, I. C., Saalmann, Y. B., Kastner, S. Rhythmic sampling within and between objects despite sustained attention at a cued location. Current Biology. 23 (24), 2553-2558 (2013).
- Landau, A. N., Fries, P. Attention samples stimuli rhythmically. Current Biology. 22 (11), 1000-1004 (2012).
- Song, K., Meng, M., Lin, C., Zhou, K., Luo, H. Behavioral oscillations in attention: Rhythmic α pulses mediated through θ band. Journal of Neuroscience. 34 (14), 4837-4844 (2014).
- Wyart, V., Nobre, A. C., Summerfield, C. Dissociable prior influences of signal probability and relevance on visual contrast sensitivity. Proceedings of the National Academy of Sciences of the United States of America. 109 (9), 3593-3598 (2012).
- Leszczyński, M., Fell, J., Axmacher, N. Rhythmic working memory activation in the human hippocampus. Cell Reports. 13 (6), 1272-1282 (2015).
- Onton, J., Delorme, A., Makeig, S. Frontal midline EEG dynamics during working memory. NeuroImage. 27 (2), 341-356 (2005).
- Cavanagh, J. F., Frank, M. J. Frontal theta as a mechanism for cognitive control. Trends in Cognitive Sciences. 18 (8), 414-421 (2014).
- Siegel, M., Buschman, T. J., Miller, E. K. Cortical information flow during flexible sensorimotor decisions. Science. 348 (6241), 1352-1355 (2015).
- Weisz, N., et al. Prestimulus oscillatory power and connectivity patterns predispose conscious somatosensory perception. Proceedings of the National Academy of Sciences of the United States of America. 111 (4), 417-425 (2014).
- Buzsáki, G., Draguhn, A. Neuronal oscillations in cortical networks. Science. 304 (5679), 1926-1929 (2004).
- Cohen, M. X. A neural microcircuit for cognitive conflict detection and signaling. Trends in Neurosciences. 37 (9), 480-490 (2014).
- Yuste, R. From the neuron doctrine to neural networks. Nature Reviews Neuroscience. 16 (8), 487-497 (2015).
- Helfrich, R. F., Knight, R. T. Oscillatory dynamics of prefrontal cognitive control. Trends in Cognitive Sciences. 20 (12), 916-930 (2016).
- Miller, E. K., Cohen, J. D. An integrative theory of prefrontal cortex function. Annual Review of Neurosciences. 24 (1), 167-202 (2001).
- von Nicolai, C., et al. Corticostriatal coordination through coherent phase-amplitude coupling. Journal of Neuroscience. 34 (17), 5938-5948 (2014).
- Sweeney-Reed, C. M., et al. Thalamic theta phase alignment predicts human memory formation and anterior thalamic cross-frequency coupling. ELife. 4, 07578(2015).
- Voytek, B., et al. Oscillatory dynamics coordinating human frontal networks in support of goal maintenance. Nature Neuroscience. 18 (9), 1318-1324 (2015).
- Raghavachari, S., et al. Theta oscillations in human cortex during a working-memory task: Evidence for local generators. Journal of Neurophysiology. 95 (3), 1630-1638 (2006).
- Jacobs, J., Hwang, G., Curran, T., Kahana, M. J. EEG oscillations and recognition memory: Theta correlates of memory retrieval and decision making. NeuroImage. 32 (2), 978-987 (2006).
- Itthipuripat, S., Wessel, J. R., Aron, A. R. Frontal theta is a signature of successful working memory manipulation. Experimental Brain Research. 224 (2), 255-262 (2013).
- Cavanagh, J. F., Zambrano-Vazquez, L., Allen, J. J. B. Theta lingua franca: A common mid-frontal substrate for action monitoring processes. Psychophysiology. 49 (2), 220-238 (2012).
- Mas-Herrero, E., Marco-Pallarés, J. Frontal theta oscillatory activity is a common mechanism for the computation of unexpected outcomes and learning rate. Journal of Cognitive Neuroscience. 26 (3), 447-458 (2014).
- Folstein, J. R., Van Petten, C. Influence of cognitive control and mismatch on the N2 component of the ERP: A review. Psychophysiology. 45 (1), 152-170 (2008).
- Cohen, M. X., Donner, T. H. Midfrontal conflict-related theta-band power reflects neural oscillations that predict behavior. Journal of Neurophysiology. 110 (12), 2752-2263 (2013).
- Walsh, M. M., Anderson, J. R. Modulation of the feedback-related negativity by instruction and experience. Proceedings of the National Academy of Sciences of the United States of America. 108 (47), 19048-19053 (2011).
- Luu, P., Tucker, D. M., Makeig, S. Frontal midline theta and the error-related negativity: Neurophysiological mechanisms of action regulation. Clinical Neurophysiology. 115 (8), 1821-1835 (2004).
- Hanslmayr, S., et al. The electrophysiological dynamics of interference during the Stroop task. Journal of Cognitive Neuroscience. 20 (2), 215-225 (2008).
- Cavanagh, J. F., Cohen, M. X., Allen, J. J. B. Prelude to and resolution of an error: EEG phase synchrony reveals cognitive control dynamics during action monitoring. Journal of Neuroscience. 29 (1), 98-105 (2009).
- Cohen, M. X., van Gaal, S., Ridderinkhof, K. R., Lamme, V. A. F. Unconscious errors enhance prefrontal-occipital oscillatory synchrony. Frontiers in Human Neuroscience. 3, 54(2009).
- Cavanagh, J. F., Frank, M. J., Klein, T. J., Allen, J. J. B. Frontal theta links prediction errors to behavioral adaptation in reinforcement learning. NeuroImage. 49 (4), 3198-3209 (2010).
- Cohen, M. X., Cavanagh, J. F. Single-trial regression elucidates the role of prefrontal theta oscillations in response conflict. Frontiers in Psychology. 2, 30(2011).
- Cohen, M. X., Van Gaal, S. Dynamic interactions between large-scale brain networks predict behavioral adaptation after perceptual errors. Cerebral Cortex. 23 (5), 1061-1072 (2013).
- Nigbur, R., Cohen, M. X., Ridderinkhof, K. R., Stürmer, B. Theta dynamics reveal domain-specific control over stimulus and response conflict. Journal of Cognitive Neuroscience. 24 (5), 1264-1274 (2012).
- van Driel, J., Swart, J. C., Egner, T., Ridderinkhof, K. R., Cohen, M. X. No) time for control: Frontal theta dynamics reveal the cost of temporally guided conflict anticipation. Cognitive, Affective, and Behavioral Neuroscience. 15 (4), 787-807 (2015).
- van de Vijver, I., Ridderinkhof, K. R., Cohen, M. X. Frontal oscillatory dynamics predict feedback learning and action adjustment. Journal of Cognitive Neuroscience. 23 (12), 4106-4121 (2011).
- Narayanan, N. S., Cavanagh, J. F., Frank, M. J., Laubach, M. Common medial frontal mechanisms of adaptive control in humans and rodents. Nature Neuroscience. 16 (12), 1888-1895 (2013).
- Anguera, J. A., et al. Video game training enhances cognitive control in older adults. Nature. 501 (7465), 97-101 (2013).
- Smit, A. S., Eling, P. A. T. M., Hopman, M. T., Coenen, A. M. L. Mental and physical effort affect vigilance differently. International Journal of Psychophysiology. 57 (3), 211-217 (2005).
- Cohen, M. X., Ranganath, C. Reinforcement learning signals predict future decisions. Journal of Neuroscience. 27 (2), 371-378 (2007).
- Gehring, W. J., Goss, B., Coles, M. G. H., Meyer, D. E., Donchin, E. A neural system for error detection and compensation. Psychological Science. 4 (6), 385-390 (1993).
- Yeung, N., Botvinick, M. M., Cohen, J. D. The neural basis of error detection: Conflict monitoring and the error-related negativity. Psychological Review. 111 (4), 931-959 (2004).
- Debener, S., et al. Trial-by-trial coupling of concurrent electroencephalogram and functional magnetic resonance imaging identifies the dynamics of performance monitoring. Journal of Neuroscience. 25 (50), 11730-11737 (2005).
- Hauser, T. U., et al. The feedback-related negativity (FRN) revisited: New insights into the localization, meaning, and network organization. NeuroImage. 84, 159-168 (2014).
- Wang, C., Ulbert, I., Schomer, D. L., Marinkovic, K., Halgren, E. Responses of human anterior cingulate cortex microdomains to error detection, conflict monitoring, stimulus-response mapping, familiarity, and orienting. Journal of Neuroscience. 25 (3), 604-613 (2005).
- Tsujimoto, S., Genovesio, A. Firing variability of frontal pole neurons during a cued strategy task. Journal of Cognitive Neuroscience. 29 (1), 25-36 (2017).
- Womelsdorf, T., Johnston, K., Vinck, M., Everling, S. Theta-activity in anterior cingulate cortex predicts task rules and their adjustments following errors. Proceedings of the National Academy of Sciences of the United States of America. 107 (11), 5248-5253 (2010).
- Womelsdorf, T., Vinck, M., Stan Leung, L., Everling, S. Selective theta synchronization of choice-relevant information subserves goal-directed behavior. Frontiers in Human Neuroscience. 4, 2010(2010).
- Domic-Siede, M., Irani, M., Valdés, J., Perrone-Bertolotti, M., Ossandón, T. Theta activity from frontopolar cortex, mid-cingulate cortex and anterior cingulate cortex shows different roles in cognitive planning performance. NeuroImage. 226, 117557(2021).
- Wilensky, R. Planning and Understanding: A Computational Approach to Human Reasoning. , Addison-Wesley. Reading, MA. (1983).
- Grafman, J., Hendler, J. Planning and the brain. Behavioral and Brain Sciences. 14, 563-564 (1991).
- Hayes-Roth, B., Hayes-Roth, F. A cognitive model of planning. Cognitive Science. 3 (4), 275-310 (1979).
- Tremblay, M., et al. Brain activation with a maze test: An EEG coherence analysis study in healthy subjects. Neuroreport. 5 (18), 2449-2453 (1994).
- Shallice, T. Specific impairments of planning. Philosophical Transactions of the Royal Society B: Biological Sciences. 298 (1089), 199-209 (1982).
- Unterrainer, J. M., Owen, A. M. Planning and problem solving: From neuropsychology to functional neuroimaging. Journal of Physiology. 99 (4-6), 308-317 (2006).
- Domic-Siede, M., et al. La planificación cognitiva en el contexto de la evaluación neuropsicológica e investigación en neurociencia cognitiva: Una revisión sistemática. Terapia Psicológica. 40 (3), 367-395 (2022).
- Miotto, E. C., Morris, R. G. Virtual planning in patients with frontal lobe lesions. Cortex. 34 (5), 639-657 (1998).
- Burgess, P. W., Simons, J. S., Coates, L. M. A., Channon, S. The search for specific planning processes. The Cognitive Psychology of Planning. Morris, R., Ward, G. , Psychology Press. Hove, UK. 199-227 (2005).
- Oosterman, J. M., Wijers, M., Kessels, R. P. C. Planning or something else? Examining neuropsychological predictors of zoo map performance. Applied Neuropsychology. 20 (2), 103-109 (2013).
- Campbell, Z., et al. Utilizing virtual reality to improve the ecological validity of clinical neuropsychology: An fMRI case study elucidating the neural basis of planning by comparing the Tower of London with a three-dimensional navigation task. Applied Neuropsychology. 16 (4), 295-306 (2009).
- Wilson, B. A., Alderman, N., Burgess, P. W., Emslie, H., Evans, J. J. Behavioural Assessment of the Dysexecutive Syndrome. , Thames Valley Test Company. St Edmunds, UK. (1996).
- Spector, L., Grafman, J. Planning, neuropsychology, and artificial intelligence: Cross fertilization. Handbook of Neuropsychology. Boller, F., Grafman, J. 9, Elsevier Science Publishers. Amsterdam, the Netherlands. 377-392 (1994).
- Porteus, S. D. The Maze Test and Clinical Psychology. , Pacific Books. Palo Alto, California. (1959).
- Lis, S., Krieger, S., Wilhelm, J., Gallhofer, B. Feedback about previous action improves executive functioning in schizophrenia: An analysis of maze solving behaviour. Schizophrenia Research. 78 (2-3), 243-250 (2005).
- Zelinsky, G. J. A theory of eye movements during target acquisition. Psychological Review. 115 (4), 787-835 (2008).
- Wurtz, R. H., Goldberg, M. E., Robinson, D. L. Brain mechanisms of visual attention. Scientific American. 246 (6), 124-135 (1982).
- Burr, D., Morrone, M. C. Eye movements: Building a stable world from glance to glance. Current Biology. 15 (20), 839-840 (2005).
- Rolfs, M. Microsaccades: Small steps on a long way. Vision Research. 49 (20), 2415-2441 (2009).
- Ortega, J., Plaska, C. R., Gomes, B. A., Ellmore, T. M. Spontaneous eye blink rate during the working memory delay period predicts task accuracy. Frontiers in Psychology. 13, 788231(2022).
- Grueschow, M., Kleim, B., Ruff, C. C. Role of the locus coeruleus arousal system in cognitive control. Journal of Neuroendocrinology. 32 (12), 12890(2020).
- Aston-Jones, G., Shaver, R., Dinan, T. Cortically projecting nucleus basalis neurons in rat are physiologically heterogeneous. Neuroscience Letters. 46 (1), 19-24 (1984).
- Foote, S. L., Bloom, F. E., Aston-Jones, G. Nucleus locus coeruleus: New evidence of anatomical and physiological specificity. Physiological Reviews. 63 (3), 844-914 (1983).
- Siever, L. J., Davis, K. L. Overview: Toward a dysregulation hypothesis of depression. The American Journal of Psychiatry. 142 (9), 1017-1031 (1985).
- Aston-Jones, G., Gold, J. I. How we say no: Norepinephrine, inferior frontal gyrus, and response inhibition. Biological Psychiatry. 65 (7), 548-549 (2009).
- Aston-Jones, G., Cohen, J. D. An integrative theory of locus coeruleus norepinephrine function: adaptive gain and optimal performance. Annual Review of Neurosciences. 28, 403-450 (2005).
- Einhauser, W., Stout, J., Koch, C., Carter, O. Pupil dilation reflects perceptual selection and predicts subsequent stability in perceptual rivalry. Proceedings of the National Academy of Sciences of the United States of America. 105 (5), 1704-1709 (2008).
- Yoshitomi, T., Ito, Y., Inomata, H. Adrenergic excitatory and cholinergic inhibitory innervations in the human iris dilator. Experimental Eye Research. 40 (3), 453-459 (1985).
- Rajkowski, J., Kubiak, P., Aston-Jones, G. Locus coeruleus activity in monkey: Phasic and tonic changes are associated with altered vigilance. Brain Research Bulletin. 35 (5-6), 607-616 (1994).
- Beatty, J. Task-evoked pupillary responses, processing load, and the structure of processing resources. Psychological Bulletin. 91 (2), 276-292 (1982).
- Beatty, J., Kahneman, D. Pupillary changes in two memory tasks. Psychonomic Science. 5, 371-372 (1966).
- Hess, E. H., Polt, J. M. Pupil size in relation to mental activity during simple problem-solving. Science. 143 (3611), 1190-1192 (1964).
- Kuchinke, L., Vo, M. L., Hofmann, M., Jacobs, A. M. Pupillary responses during lexical decisions vary with word frequency but not emotional valence. International Journal of Psychophysiology. 65 (2), 132-140 (2007).
- Barkley, R. A. Adolescents with attention-deficit/hyperactivity disorder: An overview of empirically based treatments. Journal of Psychiatric Practice. 10 (1), 39-56 (2004).
- Gau, S. S. F. em89, Shang, C. Y. Executive functions as endophenotypes in ADHD: Evidence from the Cambridge Neuropsychological Test Battery (CANTAB). Journal of Child Psychology and Psychiatry and Allied Disciplines. 51 (7), 838-849 (2010).
- Bora, E., Harrison, B. J., Yücel, M., Pantelis, C. Cognitive impairment in euthymic major depressive disorder: A meta-analysis. Psychological Medicine. 43 (10), 2017-2026 (2013).
- Rive, M. M., Koeter, M. W. J., Veltman, D. J., Schene, A. H., Ruhé, H. G. Visuospatial planning in unmedicated major depressive disorder and bipolar disorder: Distinct and common neural correlates. Psychological Medicine. 46 (11), 2313-2328 (2016).
- Holt, D. V., Wolf, J., Funke, J., Weisbrod, M., Kaiser, S. Planning impairments in schizophrenia: Specificity, task independence and functional relevance. Schizophrenia Research. 149 (1-13), 174-179 (2013).
- Lima-Silva, T. B., et al. Functional profile of patients with behavioral variant frontotemporal dementia (bvFTD) compared to patients with Alzheimer's disease and normal controls. Dementia and Neuropsychologia. 7 (1), 96-103 (2013).
- Karnath, H. O., Wallesch, C. W., Zimmermann, P. Mental planning and anticipatory processes with acute and chronic frontal lobe lesions: A comparison of maze performance in routine and non-routine situations. Neuropsychologia. 29 (4), 271-290 (1991).
- Staudigl, T., Hartl, E., Noachtar, S., Doeller, C. F., Jensen, O. Saccades are phase-locked to alpha oscillations in the occipital and medial temporal lobe during successful memory encoding. PLoS Biology. 15 (12), 2003404(2017).
- Faul, F., Erdfelder, E., Lang, A. G., Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behavior Research Methods. 39 (2), 175-191 (2007).
- Ferrando, L., Bobes, J., Gibert, J., Soto, M. MINI Entrevista Neuropsiquiátrica Internacional (MINI International Neuropsychiatric Interview, MINI). Instrumentos de Detección y Orientación Diagnóstica. , Retrieved from www.novartis.es (2000).
- McAssey, M., Dowsett, J., Kirsch, V., Brandt, T., Dieterich, M. Different EEG brain activity in right and left-handers during visually induced self-motion perception. Journal of Neurology. 267 (1), 79-90 (2020).
- Kelly, R., Mizelle, J. C., Wheaton, L. A. Distinctive laterality of neural networks supporting action understanding in left- and right-handed individuals: An EEG coherence study. Neuropsychologia. 75, 20-29 (2015).
- O'Hare, A. J., Atchley, R. A., Young, K. M. Central and divided visual field presentation of emotional images to measure hemispheric differences in motivated attention. Journal of Visualized Experiments. (129), 56257(2017).
- Beatty, J. Task-evoked pupillary responses, processing load, and the structure of processing resources. Psychological Bulletin. 91 (2), 276-292 (1982).
- Wainstein, G., et al. Pupil size tracks attentional performance in attention-deficit/hyperactivity disorder. Scientific Reports. 7 (1), 8228(2017).
- Drueke, B., et al. Neural correlates of positive and negative performance feedback in younger and older adults. Behavioral and Brain Functions. 11, 17(2015).
- Lezak, M. D. The problem of assessing executive functions. International Journal of Psychology. 17 (1-4), 281-297 (1982).
- Oosterman, J. M., Wijers, M., Kessels, R. P. C. Planning or something else? Examining neuropsychological predictors of zoo map performance. Applied Neuropsychology. 20 (2), 103-109 (2013).
- Keil, A., et al. Committee report: Publication guidelines and recommendations for studies using electroencephalography and magnetoencephalography. Psychophysiology. 51 (1), 1-21 (2014).
- Abokyi, S., Owusu-Mensah, J., Osei, K. A. Caffeine intake is associated with pupil dilation and enhanced accommodation. Eye. 31 (4), 615-619 (2017).
- Wilhelm, B., Stuiber, G., Lüdtke, H., Wilhelm, H. The effect of caffeine on spontaneous pupillary oscillations. Ophthalmic & Physiological Optics. 34 (1), 73-81 (2014).
- McGinley, M. J., David, S. V., McCormick, D. A. Cortical membrane potential signature of optimal states for sensory signal detection. Neuron. 87 (1), 179-192 (2015).
- Slotnick, S. D. High density event-related potential data acquisition in cognitive neuroscience. Journal of Visualized Experiments. (38), 1945(2010).
- Lamm, C., Windischberger, C., Leodolter, U., Moser, E., Bauer, H. Co-registration of EEG and MRI data using matching of spline interpolated and MRI-segmented reconstructions of the scalp surface. Brain Topography. 14 (2), 93-100 (2001).
- Schwartz, D., et al. Registration of MEG/EEG data with 3D MRI: Methodology and precision issues. Brain Topography. 9, 101-116 (1996).
- Laeng, B., Sirois, S., Gredebäck, G. Pupillometry: A window to the preconscious? Perspectives on psychological science. Journal of the Association for Psychological Science. 7 (1), 18-27 (2012).
- Vandierendonck, A. A comparison of methods to combine speed and accuracy measures of performance: A rejoinder on the binning procedure. Behavior Research Methods. 49 (2), 653-673 (2017).
- Vandierendonck, A. Further tests of the utility of integrated speed-accuracy measures in task switching. Journal of Cognition. 1 (1), 8(2018).
- Levene, H. Robust tests for equality of variances. In Contributions to Probability and Statistics: Essays in Honor of Harold Hotelling. Olkin, I. , Stanford University Press. Redwood City, CA. 278-292 (1960).
- Zimmerman, D. W. A note on preliminary tests of equality of variances. The British Journal of Mathematical and Statistical Psychology. 57, 173-181 (2004).
- D'Agostino, R. B., Belanger, A., D'Agostino, R. B. A suggestion for using powerful and informative tests of normality). The American Statistician. 44 (4), 316(1990).
- Shapiro, S. S., Wilk, M. B. An analysis of variance test for normality (complete samples). Biometrika. 52 (3-4), 591-611 (1965).
- Wilcoxon, F. Individual comparisons by ranking methods. Biometrics Bulletin. 1 (6), 80(1945).
- Fay, M. P., Proschan, M. A. Wilcoxon-Mann-Whitney or t-test? On assumptions for hypothesis tests and multiple interpretations of decision rules. Statistics Surveys. 4, 1-39 (2010).
- Delorme, A., Makeig, S. EEGLAB: An open-source toolbox for analysis of single-trial EEG dynamics including independent component analysis. Journal of Neuroscience Methods. 134 (1), 9-21 (2004).
- Dimigen, O., Sommer, W., Hohlfeld, A., Jacobs, A. M., Kliegl, R. Coregistration of eye movements and EEG in natural reading: Analyses and review. Journal of Experimental Psychology. 140 (4), 552-572 (2011).
- Mognon, A., Jovicich, J., Bruzzone, L., Buiatti, M. ADJUST: An automatic EEG artifact detector based on the joint use of spatial and temporal features. Psychophysiology. 48 (2), 229-240 (2011).
- Bell, A. J., Sejnowski, T. J. A non-linear information maximization algorithm that performs blind separation. Advances in Neural Information Processing Systems 7. Leen, T. K., Tesauro, G., Touretsky, D. S. , MIT Press. Cambridge, MA. edited by (1995).
- Plöchl, M., Ossandón, J. P., König, P. Combining EEG and eye tracking: Identification, characterization, and correction of eye movement artifacts in electroencephalographic data. Frontiers in Human Neuroscience. 6, 278(2012).
- Pion-Tonachini, L., Kreutz-Delgado, K., Makeig, S. ICLabel: An automated electroencephalographic independent component classifier, dataset, and website. NeuroImage. 198, 181-197 (2019).
- Gartner, M., Grimm, S., Bajbouj, M. Frontal midline theta oscillations during mental arithmetic: Effects of stress. Frontiers in Behavioral Neuroscience. 9, 96(2015).
- Wang, W., Viswanathan, S., Lee, T., Grafton, S. T. Coupling between theta oscillations and cognitive control network during cross-modal visual and auditory attention: Supramodal vs modality-specific mechanisms. PLoS One. 11 (7), 0158465(2016).
- Maris, E., Oostenveld, R. Nonparametric statistical testing of EEG- and MEG-data. Journal of Neuroscience Methods. 164 (1), 177-190 (2007).
- Tadel, F., Baillet, S., Mosher, J. C., Pantazis, D., Leahy, R. M. Brainstorm: A user-friendly application for MEG/EEG analysis. Computational Intelligence and Neuroscience. 2011, 879716(2011).
- Pascual-Marqui, R. D. Standardized low-resolution brain electromagnetic tomography (sLORETA): Technical details. Methods and Findings in Experimental and Clinical Pharmacology. 24, 5-12 (2002).
- Gramfort, A., Papadopoulo, T., Olivi, E., Clerc, M. OpenMEEG: Opensource software for quasistatic bioelectromagnetics). Biomedical Engineering Online. 9, 45(2010).
- Destrieux, C., Fischl, B., Dale, A., Halgren, E. Automatic parcellation of human cortical gyri and sulci using standard anatomical nomenclature. Neuroimage. 53 (1), 1-15 (2010).
- Orr, J. M., Weissman, D. H. Anterior cingulate cortex makes 2 contributions to minimizing distraction. Cerebral Cortex. 9 (3), 703-711 (2009).
- Christoff, K., Gabrieli, J. D. E. The frontopolar cortex and human cognition: Evidence for a rostrocaudal hierarchical organization within the human prefrontal cortex. Psychobiology. 28 (2), 168-186 (2000).
- Nitschke, K., Köstering, L., Finkel, L., Weiller, C., Kaller, C. P. A meta-analysis on the neural basis of planning: Activation likelihood estimation of functional brain imaging results in the Tower of London task. Human Brain Mapping. 38 (1), 396-413 (2017).
- Barbey, A. K., Koenigs, M., Grafman, J. Dorsolateral prefrontal contributions to human working memory. Cortex. 49 (5), 1195-1205 (2013).
- Le Van Quyen, M., et al. Comparison of Hilbert transform and wavelet methods for the analysis of neuronal synchrony. Journal of Neuroscience Methods. 111 (2), 83-98 (2001).
- Bokil, H., Andrews, P., Kulkarni, J. E., Mehta, S., Mitra, P. P. Chronux: A platform for analyzing neural signals. Journal of Neuroscience Methods. 192 (1), 146-151 (2010).
- Sato, N., Yamaguchi, Y. EEG theta regulates eye saccade generation during human object-place memory encoding. Advances in Cognitive Neurodynamics ICCN 2007. , Springer. Dordrecht, the Netherlands. 429-434 (2008).
- Zaki, J., Ochsner, K. The need for a cognitive neuroscience of naturalistic social cognition. Annals of the New York Academy of Sciences. 1167, 16-30 (2009).
- Shamay-Tsoory, S. G., Mendelsohn, A. Real-life neuroscience: An ecological approach to brain and behavior research. Perspectives on Psychological Science. 14 (5), 841-859 (2019).
- Diamond, A. Executive functions. Annual Review of Psychology. 64, 135-168 (2013).
- Dudai, Y. Memory from A to Z: Keywords, Concepts and Beyond. , Oxford University Press. New York, NY. (2002).
- Kingstone, A., Smilek, D., Ristic, J., Friesen, C. K., Eastwood, J. D. Attention, researchers! It is time to take a look at the real world. Current Directions in Psychological Science. 12 (5), 176-180 (2003).
- Morris, R. G., Ward, G. Introduction to the psychology of planning. The Cognitive Psychology of Planning. Morris, R., Ward, G. , Psychology Press. Hove, UK. (2005).
- Zwosta, K., Ruge, H., Wolfensteller, U. Neural mechanisms of goal-directed behavior: Outcome-based response selection is associated with increased functional coupling of the angular gyrus. Frontiers in Human Neuroscience. 9, 180(2015).
- Owen, A. M., Doyon, J., Petrides, M., Evans, A. C. Planning and spatial working memory: A positron emission tomography study in humans. European Journal of Neuroscience. 8 (2), 353-364 (1996).
- Ossandon, T., et al. Efficient "pop-out" visual search elicits sustained broadband gamma activity in the dorsal attention network. Journal of Neuroscience. 32 (10), 3414-3421 (2012).
- Lezak, M. D. Neuropsychological Assessment., 3rd edition. , Oxford University Press. New York, NY. (1995).
- Lin, H., Saunders, B., Hutcherson, C. A., Inzlicht, M. Midfrontal theta and pupil dilation parametrically track subjective conflict (but also surprise) during intertemporal choice. NeuroImage. 172, 838-852 (2018).
- Yu, S., Mückschel, M., Beste, C. Superior frontal regions reflect the dynamics of task engagement and theta band-related control processes in time-on task effects. Scientific Reports. 12 (1), 846(2022).
- Nakatani, H., van Leeuwen, C. Antecedent occipital alpha band activity predicts the impact of oculomotor events in perceptual switching. Frontiers in Systems Neuroscience. 7, 19(2013).
- Velasques, B., et al. Changes in saccadic eye movement (SEM) and quantitative EEG parameter in bipolar patients. Journal of Affective Disorders. 145 (3), 378-385 (2013).
- Bodala, I. P., Li, J., Thakor, N. V., Al-Nashash, H. EEG and eye tracking demonstrate vigilance enhancement with challenge integration. Frontiers in Human Neuroscience. 10, 273(2016).
- Dimigen, O. Optimizing the ICA-based removal of ocular EEG artifacts from free viewing experiments. NeuroImage. 207, 116117(2020).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기더 많은 기사 탐색
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유