
Method Article
Uma tarefa de planejamento visuoespacial acoplada a sistemas de rastreador ocular e eletroencefalograma
Neste Artigo
Resumo
O estudo do planejamento cognitivo combinando EEG e sistemas de rastreamento ocular fornece uma abordagem multimodal para investigar os mecanismos neurais que medeiam o controle cognitivo e o comportamento direcionado a objetivos em humanos. Aqui, descrevemos um protocolo para investigar o papel das oscilações cerebrais e dos movimentos oculares no planejamento do desempenho.
Resumo
O processo de planejamento, caracterizado pela capacidade de formular um plano organizado para atingir uma meta, é essencial para o comportamento humano direcionado a objetivos. Como o planejamento está comprometido em vários transtornos neuropsiquiátricos, a implementação de testes clínicos e experimentais adequados para examinar o planejamento é fundamental. Devido à natureza da implantação do planejamento, no qual vários domínios cognitivos participam, a avaliação do planejamento e o design de paradigmas comportamentais juntamente com métodos de neuroimagem são desafios atuais na neurociência cognitiva. Uma tarefa de planejamento foi avaliada em combinação com um sistema de eletroencefalograma (EEG) e registros de movimentos oculares em 27 participantes adultos saudáveis. O planejamento pode ser separado em dois estágios: um estágio de planejamento mental no qual uma sequência de etapas é representada internamente e um estágio de execução no qual a ação motora é usada para atingir um objetivo previamente planejado. Nosso protocolo incluía uma tarefa de planejamento e uma tarefa de controle. A tarefa de planejamento envolveu a resolução de 36 testes de labirinto, cada um representando um mapa do zoológico. A tarefa teve quatro períodos: i) planejamento, onde os sujeitos foram instruídos a planejar um caminho para visitar os locais de quatro animais de acordo com um conjunto de regras; ii) manutenção, onde os sujeitos tiveram que manter o caminho planejado em sua memória de trabalho; iii) execução, onde os sujeitos utilizaram movimentos oculares para traçar o caminho previamente planejado conforme indicado pelo sistema eye-tracker; e iv) resposta, onde os sujeitos relataram a ordem dos animais visitados. A tarefa de controle tinha uma estrutura semelhante, mas o componente de planejamento cognitivo foi removido modificando o objetivo da tarefa. Os padrões espaciais e temporais do EEG revelaram que o planejamento induz um aumento gradual e duradouro na atividade frontal-média (FMθ) ao longo do tempo. A fonte dessa atividade foi identificada no córtex pré-frontal por análises de fontes. Nossos resultados sugeriram que o paradigma experimental que combina sistemas de EEG e rastreador ocular foi ideal para avaliar o planejamento cognitivo.
Introdução
Durante os últimos 10 anos, uma extensa pesquisa foi conduzida para examinar o papel da dinâmica neural oscilatória na cognição e no comportamento. Esses estudos estabeleceram que as interações específicas de frequência entre regiões corticais especializadas e generalizadas desempenham um papel crucial na cognição e no controle cognitivo 1,2,3. Essa abordagem destaca a natureza rítmica da atividade cerebral, que ajuda a coordenar a dinâmica cortical em larga escala e sustenta o processamento cognitivo e o comportamento direcionado a objetivos 4,5. Há evidências substanciais mostrando que as oscilações rítmicas no cérebro estão envolvidas em vários processos cognitivos, incluindo percepção6, atenção 7,8,9, tomada de decisão10, reativação da memória11, memória de trabalho12 e controle cognitivo13. Diferentes mecanismos oscilatórios foram propostos para orientar o comportamento direcionado a objetivos, com redes transitórias específicas de frequência em larga escala fornecendo uma estrutura para o processamento cognitivo 1,14,15. Por exemplo, descobertas recentes sugerem que bandas de frequência específicas no cérebro podem refletir um mecanismo de feedback que regula a atividade de pico, fornecendo um quadro de referência temporal para coordenar a excitabilidade cortical e o tempo de pico para produzir comportamento 16,17,18. Uma revisão é fornecida por Helfrich e Knight19.
Esse corpo de evidências levanta questões sobre como o córtex pré-frontal (PFC) codifica contextos de tarefas de planejamento e regras comportamentais relevantes relacionadas. Há muito se pensa que o PFC apóia o controle cognitivo e o comportamento direcionado a objetivos por meio dos padrões oscilatórios de atividade neural que gera, influenciando seletivamente a atividade neural em regiões cerebrais distantes e controlando o fluxo de informações em redes neurais de grande escala20. Além disso, foi proposto que as regiões que exibem sincronia local são mais propensas a participar de atividades inter-regionais 21,22,23. Em particular, as oscilações corticais da banda (4-8 Hz), medidas pelo eletroencefalograma (EEG) do couro cabeludo, foram propostas como um mecanismo potencial para transmitir o controle de cima para baixo em redes amplas13. Especificamente, a atividade da banda em humanos reflete processos cognitivos de alto nível, como codificação e recuperação de memória, retenção de memória de trabalho, detecção de novidades, tomada de decisão e controle de cima para baixo 12,24,25,26.
Relacionado a isso, Cavanagh e Frank13 propuseram dois mecanismos sequenciais de processos de controle: o reconhecimento da necessidade de controle e a instanciação do controle. O reconhecimento da necessidade de controle pode ser indicado pela atividade da linha média frontal (FMθ) originada do córtex pré-frontal medial (mPFC), que tem sido descrita em termos de componentes de potencial relacionado a eventos (ERP) que refletem os processos de controle relacionados ao mPFC em resposta a várias situações, como novas informações 27,28,29, requisitos conflitantes de estímulo-resposta 30, feedback de erro31, e detecção de erros32. Esses componentes do ERP, que refletem a necessidade de maior controle cognitivo na presença de novidade, conflito, punição ou erro, exibem uma assinatura espectral comum na banda registrada nos eletrodos da linha média frontal 26,27,33,34,35,36,37,38,39,40, 41,42,43,44.
As respostas de EEG da atividade de FMθ exibem um padrão de redefinição de fase e aumento de potência na banda de frequência26. Apesar das limitações do método EEG em termos de sua resolução espacial, várias fontes de evidência foram coletadas para demonstrar que a atividade da FMθ é gerada pelo córtex cingulado médio (MCC)13. Acredita-se que essa dinâmica sirva como estruturas temporais que regulam os processos neuronais do mPFC, que são subsequentemente aumentados em resposta a eventos que requerem maior controle26. Isso foi estabelecido por meio de análise de fonte 31,33,45,46,47, registros simultâneos de EEG e ressonância magnética funcional (fMRI) 48,49 e registros invasivos de EEG em humanos50 e macacos 51,52,53.
Com base nessas observações, considera-se que o da linha média frontal serve como um mecanismo universal, uma linguagem comum, para executar o controle adaptativo em diferentes situações em que há falta de certeza sobre as ações e resultados, como durante o planejamento. O paradigma comportamental que propomos neste protocolo tem sido utilizado para estudar o planejamento cognitivo e suas características temporais e neurais. Embora vários mecanismos de controle cognitivo tenham sido relatados em outros cenários, o protocolo atual permitiu a descrição recente do planejamento e suas propriedades neurais e temporais associadas54. O processo cognitivo de planejamento compreende duas fases distintas: a fase de planejamento mental, durante a qual uma representação interna de uma sequência de planos é desenvolvida55, e a fase de execução do planejamento, na qual um conjunto de ações motoras é executado para atingir a meta previamente planejada56. Sabe-se que o planejamento requer a integração de vários componentes das funções executivas, incluindo memória de trabalho, controle atencional e inibição de resposta, tornando a manipulação experimental e a medição isolada desses processos desafiadoras57,58.
Estudos de neuroimagem sobre planejamento cognitivo têm comumente usado paradigmas comportamentais como a Torre de Londres 59,60,61; No entanto, para controlar os fatores de confusão, as tarefas utilizadas para estudar o planejamento cognitivo podem se tornar limitadas e artificiais, levando a uma menor validade preditiva e ecológica 62,63,64,65. Para superar esse problema no campo da neuropsicologia, situações de planejamento do mundo real têm sido propostas como tarefas ecológicas62,63. O subteste Zoo Map Task na bateria de Avaliação Comportamental da Síndrome Disexecutiva mede as habilidades de planejamento e organização de maneira mais natural e relevante64,66. Este teste é um teste de lápis e papel que envolve o planejamento de uma rota para visitar 6 dos 12 locais em um mapa do zoológico. Os locais são locais comuns que podem ser encontrados em um zoológico comum, como uma casa de elefantes, gaiola de leão, área de descanso, cafeteria, etc. Existem duas condições que avaliam diferentes níveis de planejamento: i) a condição de formulação, onde os sujeitos são instruídos a planejar uma rota para visitar seis lugares na ordem de sua escolha, mas de acordo com um conjunto de regras; e ii) a condição de execução, onde os sujeitos são instruídos a visitar seis locais em uma ordem específica e seguindo um conjunto de regras também. Essas duas condições fornecem informações sobre habilidades de planejamento em problemas mal estruturados (formulação) e bem estruturados (execução)67. O primeiro é apresentado como uma tarefa cognitiva mais exigente em uma situação aberta, pois exige que os sujeitos desenvolvam uma estratégia lógica para atingir o objetivo. Antes de traçar um caminho, uma sequência de operadores deve ser concebida; caso contrário, é provável que ocorram erros. Por outro lado, a condição de execução requer uma menor demanda cognitiva, pois a resolução de uma tarefa que envolve o cumprimento de uma estratégia específica imposta requer apenas que o sujeito monitore a implementação do plano formulado para atingir a meta66. Por outro lado, o Labirinto de Porteus é uma tarefa bem conhecida no campo da psicologia, particularmente nas áreas da psicologia cognitiva e da neuropsicologia, e tem sido amplamente utilizado como ferramenta para avaliar vários aspectos da cognição, como resolução de problemas e planejamento68,69. A tarefa do Labirinto de Porteus é uma tarefa de lápis e papel que começa com uma simples análise de estímulos visuais e se torna cada vez mais difícil. O sujeito deve encontrar e traçar o caminho correto de um ponto de partida para uma saída (entre várias opções) enquanto segue regras, como evitar caminhos que se cruzam e becos sem saída, e agir o mais rápido possível68. Cada vez que uma bifurcação aparece ao traçar o caminho, os sujeitos tomam decisões para atingir a meta e evitar quebrar as regras dadas69.
Considerando as limitações e pontos fortes das tarefas comumente usadas e ecológicas, projetamos nosso paradigma comportamental baseado principalmente na Tarefa66 do Mapa do Zoológico e na Tarefa68 do Labirinto de Porteus. O paradigma comportamental consiste em quatro estágios distintos que englobam o processo cognitivo de planejamento em um cenário de vida diária. Essas etapas são as seguintes: Etapa 1, planejamento, onde os participantes têm a tarefa de criar uma rota para visitar vários locais em um mapa, garantindo o cumprimento das regras estabelecidas; Estágio 2, manutenção, onde os participantes são obrigados a manter a rota planejada em sua memória de trabalho; Etapa 3, execução, onde os participantes executam sua rota previamente planejada desenhando e monitorando de perto sua precisão; e Etapa 4, resposta, onde os participantes relatam a sequência de animais visitados de acordo com a rota planejada54. Nosso paradigma permite a medição de diferentes parâmetros de capacidade de planejamento usando diferentes estágios, que refletem os vários componentes do planejamento (como memória de trabalho, atenção executiva e habilidades visuoespaciais) de maneira mais realista, uma vez que mapear rotas é uma ocorrência comum na vida diária. Além disso, para controlar fatores de confusão, o paradigma inclui uma tarefa de controle com uma estrutura de tarefa de planejamento e estímulos equivalentes, que envolve os componentes cognitivos executivos também envolvidos no planejamento, mas exclui o componente do processo de planejamento. Isso permite a separação do componente do processo de planejamento para a comparação de marcadores eletrofisiológicos e parâmetros comportamentais54.
Além disso, o rastreamento ocular fez contribuições significativas para os estudos de neurociência cognitiva, fornecendo um método não invasivo para medir e analisar os movimentos oculares, que pode fornecer informações valiosas sobre os processos cognitivos e mecanismos neurais subjacentes à percepção, atenção e funções cognitivas. Medir diferentes tipos de movimentos oculares com um sistema de rastreamento ocular pode fornecer informações valiosas sobre os processos cognitivos e mecanismos neurais envolvidos no planejamento. Por exemplo, os seguintes aspectos podem ser medidos: fixações, que são os períodos de olhar estável durante os quais a informação visual é adquirida70; sacadas, que são os movimentos rápidos dos olhos usados para mudar o olhar de um local para outro71; perseguição suave, que é um tipo de movimento ocular que permite que os olhos sigam um objeto em movimento suavemente72; microssacadas, que são movimentos oculares pequenos e rápidos que ocorrem mesmo durante as fixações73; e piscar, que são uma ação reflexa que ajuda a manter os olhos lubrificados e protegê-los de objetos estranhos74. Esses movimentos oculares podem fornecer informações sobre os processos cognitivos envolvidos na busca visual, alocação de atenção70, rastreamento visual72, percepção73 e memória de trabalho74, que são componentes importantes para o planejamento e controle cognitivo.
Por outro lado, estudos recentes sobre o sistema locus coeruleus-norepinefrina (LC-NE) têm mostrado seu papel relevante no controle cognitivo75. O locus coeruleus (LC) se projeta para várias regiões do cérebro, como córtex cerebral, hipocampo, tálamo, mesencéfalo, tronco encefálico, cerebelo e medula espinhal 76,77,61. Inervações LC-NE particularmente densas recebem áreas cerebrais PFC associadas ao controle cognitivo75. Além disso, alguns estudos indicam que a hiperatividade crônica do sistema LC pode contribuir para sintomas de transtorno maníaco-depressivo, como impulsividade e insônia. Em contraste, uma diminuição crônica na função LC tem sido associada à redução da expressão emocional, uma característica prevalente entre pacientes que sofrem de depressão78. Uma resposta hiperativa do locus coeruleus a estímulos pode levar a uma resposta excessiva em indivíduos com transtornos de estresse ou ansiedade79. Portanto, alterações no sistema LC-NE podem contribuir para os sintomas de desregulação cognitiva e/ou emocional. Técnicas não invasivas podem ser usadas para examinar a atividade do locus coeruleus, uma das quais são as alterações do diâmetro da pupila, que são controladas principalmente pela noradrenalina liberada do locus coeruleus. A noradrenalina atua no músculo dilatador da íris estimulando os receptores alfa-adrenérgicos e no núcleo de Edinger-Westphal, que envia sinais para o gânglio ciliar e controla a dilatação da íris por meio da ativação de adrenoceptores alfa-2 pós-sinápticos 66,80,81,82. Gravações neuronais diretas de LC de macacos confirmaram a relação entre a atividade de LC-NE, o diâmetro da pupila e o desempenho cognitivo83. A dilatação da pupila tem sido repetidamente observada em resposta a demandas de processamento aprimoradas em várias tarefas cognitivas 71,84,85,86,87.
Marcadores eletrofisiológicos de controle cognitivo combinados com rastreamento ocular e registros pupilares podem desvendar questões cruciais sobre como o controle cognitivo e o planejamento são implementados no cérebro. A importância de usar nosso protocolo combinando sistemas de EEG e rastreador ocular é dupla. Por um lado, o controle cognitivo parece exigir a participação da atividade cerebral distribuída em relações temporais precisas, que constituem candidatos ideais para estudar a função da rede cerebral. Por outro lado, anormalidades em qualquer uma dessas capacidades têm um impacto severo no comportamento normal, como pode ser no caso de uma variedade de distúrbios cognitivos e neuropsiquiátricos, como transtorno de déficit de atenção/hiperatividade88,89, transtorno depressivo maior90,91, transtorno bipolar91, esquizofrenia92, demência frontotemporal93, bem como distúrbios devido a lesões frontais94. Além disso, o protocolo atual permite o uso da pupilometria como parâmetro para comparar a atividade e as oscilações de LC-NE usando rastreamento ocular e eletroencefalografia. Isso pode não apenas fornecer evidências para a relação teórica entre LC-NE, pupilometria e marcadores neurais em humanos, mas também pode permitir o rastreamento da trajetória de desenvolvimento de características relacionadas ao sistema LC-NE durante o planejamento cognitivo. No entanto, em nosso modelo, nos concentramos em testar se havia um padrão específico de sacadas durante o planejamento que poderia resultar em mudanças específicas de oscilação95. Além disso, usamos um sistema de rastreamento ocular como uma parte importante do exame da execução comportamental de um plano na fase de execução de nosso paradigma comportamental.
Resumindo, este protocolo pode produzir modelos testáveis de dinâmica de rede cerebral que podem servir como uma plataforma para pesquisas básicas adicionais e eventuais aplicações clínicas e terapêuticas.
Protocolo
Todos os procedimentos deste protocolo foram aprovados pelo comitê de bioética da Faculdade de Medicina da Pontifícia Universidade Católica do Chile, e todos os participantes assinaram um termo de consentimento livre e esclarecido antes do início do estudo (projeto de pesquisa número: 16-251).
1. Recrutamento de participantes
- Recrute adultos saudáveis destros (homens e mulheres) com visão normal ou corrigida para normal e faça a triagem deles de acordo com os critérios de inclusão/exclusão.
NOTA: Neste estudo, foram recrutados 27 indivíduos saudáveis, com idade entre 19 e 38 anos e falantes fluentes. O tamanho da amostra pode variar de acordo com o nível desejado de poder estatístico, e a faixa etária pode variar dependendo da questão de pesquisa específica a ser abordada. Em nosso protocolo, calculamos o tamanho da amostra levando em consideração o teste estatístico dos postos sinalizados de Wilcoxon, um tamanho de efeito de 0,7, um nível alfa de 0,05 e um poder de 0,95, conforme descrito em Faul et al.96. Utilizou-se o MINI-International Neuropsychiatric Interview97, aplicado por um psicólogo treinado, para avaliar os participantes quanto aos critérios de inclusão/exclusão. Recrute apenas indivíduos destros para reduzir a variabilidade no sinal de EEG, pois indivíduos canhotos podem apresentar uma distribuição topográfica diferente da atividade de EEG 98,99,100.
2. Preparação de estímulos
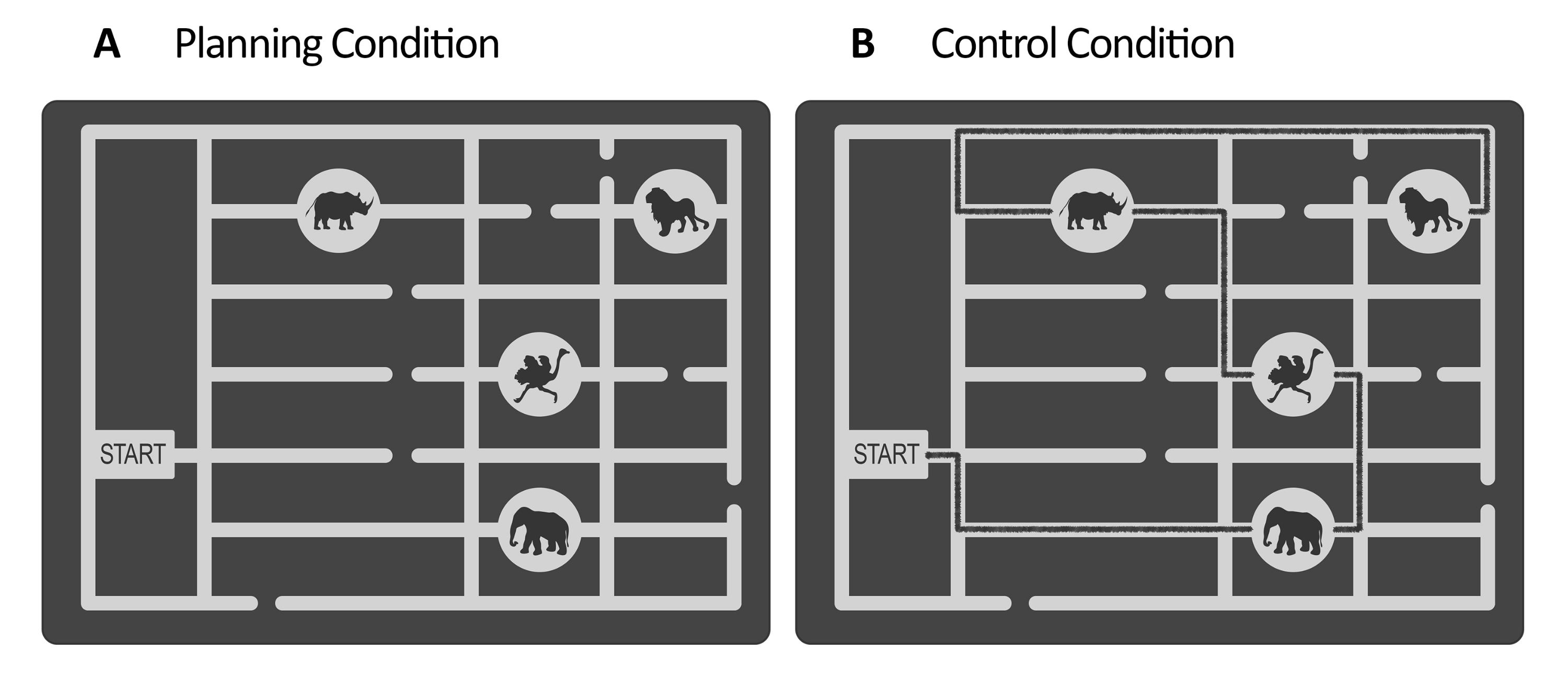
- Para a tarefa de planejamento, crie um conjunto de estímulos usando um software editor de gráficos vetoriais (consulte Tabela de Materiais). Para cada estímulo, projete um labirinto em tons de cinza que represente um mapa do zoológico. Dentro do labirinto, faça um portal e vários caminhos que levam aos locais dos animais (por exemplo, neste estudo, havia quatro locais de animais, veja a Figura 1).
NOTA: Neste estudo, criamos 36 labirintos em que cada estímulo consistia em um mapa do zoológico com um portão de partida, quatro imagens de animais localizados no labirinto e vários caminhos. Os caminhos no labirinto podem ou não levar à localização dos animais. O uso de estímulos em tons de cinza com contraste reduzido é frequentemente preferido para a pupilometria porque reduz a estimulação dos cones da retina, responsáveis pela visão de cores. Essa mudança na ênfase do estímulo permite uma medição mais sensível das mudanças no tamanho da pupila, que refletem mudanças no estado de excitação ou atenção. Além disso, o uso de estímulos em tons de cinza reduz a variabilidade na medida devido às diferenças na visão de cores entre os indivíduos101. - Para obter diferentes níveis de complexidade na tarefa final, divida os estímulos com base no número de soluções válidas de acordo com o objetivo e as regras estabelecidas (particularmente, o objetivo é planejar um caminho para visitar locais de animais). O número de soluções válidas refere-se ao número de caminhos possíveis de planejar seguindo as regras (consulte as regras na Figura 2 e na etapa 5.12.1.). Classifique os estímulos com mais de cinco soluções possíveis como "fáceis" e aqueles com cinco ou menos soluções possíveis como "difíceis". Em seguida, crie um número igual de testes para cada categoria.
NOTA: Alternativamente, solicite os estímulos criados para Domic-Siede et al.54 aos autores, pois esses estímulos foram criados seguindo estas instruções. Considere que todos os materiais estão disponíveis mediante solicitação, mas as especificações são detalhadas aqui. Neste estudo, criamos 18 tentativas fáceis e 18 tentativas difíceis. Avaliar as diferenças nos níveis de dificuldade nos níveis comportamental e eletrofisiológico é importante porque ajudará a determinar se você está medindo a demanda cognitiva / esforço cognitivo / dificuldade ou aspectos intrínsecos do planejamento cognitivo (consulte os resultados representativos e as seções de discussão). - Para a tarefa de controle, use a mesma estrutura da tarefa de planejamento (período de avaliação, manutenção, execução, resposta, consulte a Figura 2) e use os mesmos estímulos criados para a tarefa de planejamento, mas adicione uma linha desenhada representando um caminho marcado para visitar a sequência de locais de animais desde o portão até o último local (consulte a Figura 1B). Faça o caminho marcado com uma cor ligeiramente mais escura do que os caminhos principais do labirinto, com baixo contraste avaliado usando um luxímetro (consulte a etapa 2.4).
NOTA: A ideia por trás disso é manter as características psicofísicas de ambas as condições (tarefas de planejamento e controle) o mais semelhantes possível. Os caminhos traçados dos labirintos podem seguir as regras ou não (consulte a seção 5 do protocolo para obter mais detalhes sobre as instruções dadas aos participantes). Neste estudo, metade dos estímulos teve uma sequência de visitas correta seguindo as regras, enquanto a outra metade apresentou erros (como usar o mesmo caminho duas vezes ou cruzar becos sem saída, ver passo 5.12.1 e passo 5.12.3 e Figura 2). - Avalie a iluminância dos estímulos usando um luxímetro posicionado no apoio de queixo que os sujeitos usarão (consulte a etapa 4.5 e a etapa 4.6) e à mesma distância da tela. Cada estímulo de cada condição produz um valor lux. Registre cada valor manualmente para análises posteriores.
NOTA: Não são esperadas diferenças na iluminância entre as condições (consulte a etapa 4.6). Caso contrário, verifique o contraste dos estímulos. Isso é relevante se o diâmetro da pupila for medido102. - Crie um estímulo representando o feedback correto (polegar para cima quando correto) e outro estímulo representando feedback incorreto (polegar para baixo quando incorreto) usando um editor gráfico vetorial (consulte a Tabela de Materiais) também (Figura 2).

Figura 1: Estímulos da tarefa experimental e controle. Exemplos ilustrativos dos estímulos (A) planejamento e (B) tarefa de controle são mostrados. Os estímulos representam um mapa do zoológico que consiste em um portão, quatro localizações de animais em lugares diferentes e vários caminhos. Os estímulos para ambas as condições foram semelhantes; a única diferença era que, para a tarefa de controle, (B) os estímulos tinham uma linha marcada indicando um caminho já existente (linha preta aqui para fins ilustrativos). Essa linha nos estímulos de controle reais era ligeiramente mais escura, com baixo contraste controlado pela iluminância (consulte a etapa 2.4). Essa figura foi modificada de Domic-Siede et al.54. Clique aqui para ver uma versão maior desta figura.
{kind=link}

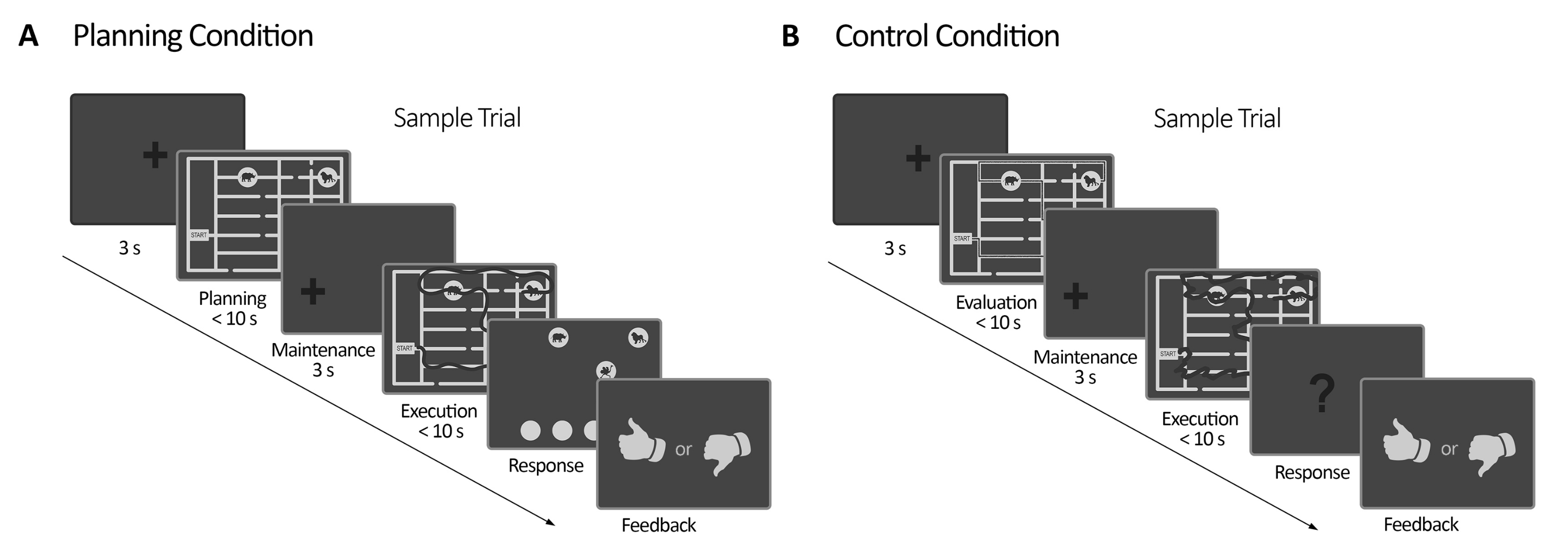
Figura 2: Desenho experimental. (A) Teste de tarefa de planejamento. As tentativas nesta condição começaram com uma cruz de fixação de 3 s. Em seguida, os participantes foram instruídos a planejar um caminho para visitar todos os quatro locais dos animais seguindo um conjunto de regras (máximo de 10 s). Em seguida, uma cruz de fixação deslocada apareceu (3 s), seguida pelo aparecimento do labirinto novamente. Nesse período (execução), os sujeitos tiveram que executar o traçado planejado no período de planejamento anterior usando seu olhar com feedback visual on-line (dado pelo sistema de rastreamento ocular), que delineou o movimento do olhar em tempo real (linha escura) (máximo de 10 s). Posteriormente, no período de resposta, os sujeitos tiveram que relatar a sequência feita durante a execução, ordenando que os animais fossem visitados. De acordo com suas respostas, o feedback foi entregue. (B) Teste de tarefa de controle. As tentativas nesta condição começaram com uma cruz de fixação de 3 s. Em seguida, os participantes foram instruídos a avaliar se um caminho traçado (linha escura) seguia as regras ou não. Em seguida, uma cruz de fixação deslocada apareceu (3 s), seguida pelo aparecimento do labirinto novamente. Nesse período, os sujeitos tiveram que redesenhar o caminho já traçado com feedback visual online, como no período de execução do planejamento (máximo de 10 s). Posteriormente, no período de resposta, os sujeitos tiveram que responder (sim ou não) se a sequência traçada seguia as regras previamente declaradas. Essa figura foi modificada de Domic-Siede et al.54. Clique aqui para ver uma versão maior desta figura.
{kind=link}
3. Programação de tarefas de planejamento e controle
- Escreva um script codificando um paradigma de tarefa de planejamento baseado na Tarefa66 do Mapa do Zoológico e no Labirinto68de Porteus usando um software de apresentação de estímulos/experimentos comportamentais (consulte a Tabela de Materiais e o Arquivo Suplementar).
- Codifique a tarefa considerando duas condições (uma condição de planejamento e uma condição de controle) com uma estrutura semelhante à explicada na seção 2 e na seção 4 (consulte a Figura 2 e o Arquivo Suplementar).
NOTA: É importante utilizar a mesma estrutura em ambas as condições para controlar os fatores de confusão e os componentes perceptivos envolvidos no processo de resolução das demandas da tarefa (Figura 2). O uso da mesma estrutura melhora a avaliação do processo cognitivo específico envolvido no planejamento cognitivo. - Sincronize a comunicação entre o computador de exibição, o computador de EEG e o computador host (computador rastreador ocular) via ethernet e comunicação de porta paralela enviando pulsos de lógica transistor-transistor (TTL) do computador de exibição (consulte a Figura 3).
- Escreva um código para calibrar os movimentos oculares com o sistema de rastreamento ocular no início das tarefas de planejamento e controle e após cada cinco tentativas concluídas, pois a posição do olhar na tela é crucial para o período de execução (consulte a etapa 3.2.3 e a etapa 3.3.3 do protocolo, a seção de discussão e o código no Arquivo Suplementar).
NOTA: Pode haver atrasos na comunicação com o computador. Existem vários métodos para medir o atraso entre pulsos TTL em dois computadores diferentes, mas uma abordagem comum é usar um dispositivo de hardware, como um osciloscópio digital ou um analisador lógico. Outra abordagem é usar métodos baseados em software, como enviar os pulsos TTL por uma conexão de rede e usar ferramentas de análise de rede para medir o atraso. Outra abordagem é sincronizar os relógios dos dois computadores, usando um sistema de posicionamento global (GPS) ou servidor NTP (protocolo de tempo de rede) ou usando uma solução de sincronização baseada em hardware, calcular o atraso entre o carimbo de data/hora e o tempo de chegada para cada pulso e calcular a média dos resultados para obter o atraso geral entre os dois computadores.
- Codifique a tarefa considerando duas condições (uma condição de planejamento e uma condição de controle) com uma estrutura semelhante à explicada na seção 2 e na seção 4 (consulte a Figura 2 e o Arquivo Suplementar).
- Escreva um código para a tarefa de planejamento com a seguinte estrutura: o período de planejamento, o período de manutenção, o período de execução do planejamento, o período de resposta e comentários (Figura 2, Arquivo Suplementar).
- O período de planejamento: Inicie a condição de planejamento definindo uma cruz de fixação apresentada por 3 s como linha de base.
- Apresente aleatoriamente o conjunto de labirintos um a um (36 neste estudo).
NOTA: Neste período de planejamento, os participantes são solicitados a planejar um caminho para visitar os quatro locais de animais, com um tempo máximo de 10 s, seguindo um conjunto de regras (as regras são previamente explicadas a eles; consulte a seção 5 do protocolo para ver as regras fornecidas, bem como a Figura 2). - Inclua um gatilho TTL no código sinalizando o início da apresentação do estímulo usando um código de etiqueta e envie esse gatilho para o computador EEG e o computador host do rastreador ocular para análises mais estreitas e em janelas.
- Escreva no código que o período de planejamento culmina quando um botão de um joystick/teclado é pressionado sempre que o sujeito termina o planejamento ou se o tempo máximo é excedido. O tempo de reação (RT) deve ser registado no ficheiro de registo para análises posteriores.
NOTA: Para este período, usamos um código de disparo usando o número 1, mas o uso de tags de descritores hierárquicos de eventos (HED) em vez de códigos numéricos é recomendado, pois as tags HED fornecem significado e estrutura ao conteúdo, facilitando assim a compreensão do conteúdo dos dados por outros pesquisadores ou colaboradores. - O período de manutenção: Inicie este período usando uma cruz de fixação deslocada apresentada por 3 s. Localize a cruz de fixação deslocada na posição espacial onde o portão do labirinto está localizado para antecipar a posição inicial (portão) do mapa do zoológico (veja a Figura 2).
NOTA: O objetivo deste período é triplo. Primeiro, a cruz de fixação deslocada facilita a execução do traço que representa o caminho planejado para o próximo período (consulte a etapa 3.2.8). Em segundo lugar, durante esse período, os participantes mantêm o plano elaborado durante o período de planejamento em sua memória de trabalho. Finalmente, esse período serve como um intervalo entre tentativas para delimitar o final do período de planejamento e o início do próximo período - o período de execução do planejamento. - O período de execução do planejamento: Depois que a cruz de fixação deslocada for mostrada por 3 s durante o período de manutenção, apresente o labirinto novamente.
- Envie um gatilho TTL para o EEG e o computador rastreador ocular host para indicar o início desse período usando um código de tag específico.
- Escreva um código para fornecer feedback visual em tempo real (uma linha escura, veja o período de execução na Figura 2) da posição do olhar do sujeito aproximadamente 992 ms após o início desse período.
NOTA: Começar a delinear com um atraso (1.000 ms aproximadamente) dá aos sujeitos tempo para se orientarem no labirinto, permitindo-lhes delinear seu caminho planejado anterior (durante o período de planejamento) com uma linha escura. - Registre as coordenadas dos caminhos para posterior reconstrução dos caminhos feitos pelos sujeitos e pontue o desempenho offline (consulte a etapa 6.1.1, Figura 4).
- Garanta um tempo máximo de 10 s para traçar o caminho planejado e permita que os sujeitos finalizem esse período pressionando um botão. Desta forma, os sujeitos podem controlar quando terminaram o caminho traçado.
- Salve o RT no arquivo de log para análises adicionais.
- O período de resposta: Escreva um código para o período de resposta, que começa após 10 s de execução do planejamento ou ao pressionar um botão no final do período de execução do planejamento, no qual o labirinto desaparece, mas os animais e suas posições espaciais permanecem na tela.
- Coloque quatro círculos vazios horizontalmente na parte inferior da tela no período de resposta.
NOTA: O objetivo deste período é permitir que os sujeitos indiquem a sequência de animais visitados durante o período de execução do planejamento, colocando os animais nos círculos na mesma ordem em que os visitaram usando um joystick ou teclado. - Configure o programa/código para permitir que os sujeitos usem o joystick ou teclado para selecionar cada um dos animais (quatro animais neste estudo) apresentados anteriormente e inseri-los em cada um dos quatro círculos (ver Arquivo Suplementar e Figura 2).
- Realimentação: Escreva um código para entregar 3 s de feedback aos participantes. Uma imagem de polegar para cima deve ser exibida em resposta a combinações válidas de animais visitados se as regras forem seguidas, enquanto uma imagem de polegar para baixo deve ser exibida se a combinação relatada for inválida.
- Envie um gatilho TTL, usando um código de tag específico para feedback correto e outro código de tag para incorreto, para os computadores de EEG e rastreador ocular.
NOTA: O motivo para dar feedback é facilitar o monitoramento do desempenho e manter a motivação durante a tarefa. Essa provisão de feedback em tempo real aumenta o efeito de recompensa e incentiva o desempenho adequado da tarefa103.
- Escreva um código para a tarefa de controle com a mesma estrutura da condição de planejamento: um período de controle, um período de manutenção, um período de execução de controle, um período de resposta e feedback (consulte Arquivo Suplementar, Figura 2).
- O período de controle: Escreva um código para o período da condição de controle para mitigar fatores de confusão. O código para este período deve começar com uma cruz de fixação apresentada por 3 s como linha de base.
NOTA: Uma vez que a tarefa de planejamento exige predominantemente a implementação do planejamento, mas também recruta outros domínios cognitivos como parte da função executiva, como função visuoespacial, memória de trabalho, controle atencional, controle inibitório, etc.66,88,104,105, uma tarefa de controle é crucial para mitigar fatores de confusão. Assim, o principal objetivo dessa tarefa é exigir todas as funções cognitivas e perceptivas necessárias para resolver a tarefa de planejamento, ao mesmo tempo em que remove a implementação do planejamento cognitivo54. - Apresente aleatoriamente os labirintos da condição de controle um por um (labirintos com um caminho marcado já traçado). Codifique um tempo máximo de 10 s.
- Inclua um gatilho TTL no código que sinaliza o início da apresentação do estímulo usando uma etiqueta e envie esse gatilho para o computador EEG e o computador host do rastreador ocular.
- Escreva no código que este período de controle culmina quando um botão de um joystick/teclado é pressionado sempre que o assunto termina ou se o tempo máximo é excedido.
NOTA: Os sujeitos são instruídos a avaliar os caminhos marcados (estejam eles seguindo as regras ou não, consulte a etapa 5.12 para obter detalhes sobre as instruções dadas aos participantes). - Salve o tempo de reação (RT) no arquivo de log para análises adicionais.
- O período de manutenção: Uma vez terminado o período de controle, apresente uma cruz de fixação deslocada por 3 s.
- Como período de execução do planejamento, coloque a cruz de fixação onde está localizada a entrada do portão para facilitar o desenho do olhar para o próximo período.
- O período de execução do controle: Apresente o labirinto novamente e, simultaneamente, envie um gatilho TTL para o EEG e hospede computadores rastreadores oculares com uma etiqueta sinalizando o início do período de execução.
- Repita o mesmo código do período de execução do planejamento para fornecer comentários on-line sobre a posição do olhar e delinear e sobrepor seu olhar com o caminho rastreado.
- Garanta um tempo máximo de 10 s para traçar o caminho e permita que os sujeitos finalizem esse período pressionando um botão.
- Salve o RT no arquivo de log para análises adicionais.
- O período de resposta de controle: Uma vez terminado o período de execução do controle, apresente um ponto de interrogação indicando o período de resposta.
- Programe dois botões, respectivamente, para que os sujeitos dêem uma resposta usando um joystick ou um teclado.
NOTA: Aqui, os sujeitos são solicitados a responder se a sequência marcada pelo traço estava correta ou não, selecionando um botão para correto/SIM e outro para incorreto/NÃO. - Salve a precisão no arquivo de log.
- Realimentação: Escreva um código para fornecer 3 s de feedback correto sempre que os sujeitos responderem corretamente (uma imagem de polegar para cima) e forneça 3 s de feedback incorreto quando os sujeitos responderem incorretamente (uma imagem de polegar para baixo).
- Como na condição de planejamento, envie um gatilho TTL para o EEG e hospede computadores rastreadores oculares com uma tag para feedback correto e outra tag para feedback incorreto.
- O período de controle: Escreva um código para o período da condição de controle para mitigar fatores de confusão. O código para este período deve começar com uma cruz de fixação apresentada por 3 s como linha de base.
- Tarefas de treinamento: Crie estímulos, escreva um código e apresente antes das tarefas de planejamento e controle mencionadas uma breve sessão de treinamento de aproximadamente seis tentativas/labirintos para cada condição (planejamento e controle)
NOTA: A ideia é garantir a familiarização com a configuração da tarefa. Recomenda-se definir critérios para prosseguir. Neste estudo, se as três últimas tentativas estivessem corretas e os participantes relatassem entender o objetivo e o procedimento ao final da sessão de treinamento, os participantes prosseguiam para a sessão experimental.

Figura 3: Exemplo de uma configuração de laboratório. Representação esquemática de uma configuração de laboratório mostrando três computadores interconectados. O computador host (computador rastreador ocular) é responsável por rastrear e armazenar os dados do movimento ocular. O computador de EEG adquire e armazena os sinais de EEG. O computador de exibição controla o experimento comportamental, apresenta os estímulos aos sujeitos e envia gatilhos de eventos para o host e os computadores de EEG por meio de portas paralelas e conexões LAN para sincronizar a coleta de dados. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 4: Reconstrução do caminho a partir do feedback visual on-line fornecido pelo sistema de rastreamento ocular. Exemplos ilustrativos de uma reconstrução de caminho a partir da execução motora de um plano (A, em roxo, período de execução do planejamento) e um período de execução de controle (B, linha em verde) e com dados de rastreador ocular. O caminho reconstruído no período de execução do planejamento é usado para avaliar a precisão de cada teste de tarefa de planejamento. Clique aqui para ver uma versão maior desta figura.
{kind=link}
4. Ambiente e equipamento de laboratório
- Use um sistema de aquisição de EEG para registrar a atividade de EEG do couro cabeludo do participante, com os eletrodos de EEG colocados de acordo com o sistema internacional 10-20106. Posicione dois eletrodos nas mastóides do participante para re-referenciamento offline. Use eletrodos de eletrooculograma para identificar sinais de movimento ocular verticais, horizontais e piscantes durante a inspeção visual.
- Use o software de aquisição de EEG para a aquisição de dados de EEG com uma taxa de amostragem de 2.048 Hz ou 1.024 Hz e um filtro passa-banda entre 0,1-100 Hz no computador de EEG.
NOTA: Taxas de amostragem de 1.024 Hz e 2.048 Hz fornecem resolução suficiente para analisar oscilações de baixa frequência. É importante adquirir sinais de EEG com alta taxa de amostragem, como acima de 1.000 Hz, ao analisar oscilações de baixa frequência, a fim de garantir que os sinais de baixa frequência não sejam confundidos. - Use um computador de exibição, que esteja conectado ao computador EEG e ao computador rastreador ocular host por meio de portas paralelas e ethernet e tenha uma plataforma para executar experimentos comportamentais instalada nele, para projetar os estímulos em um monitor estendido com resolução mínima de 1.920 pixels x 1.080 pixels e taxa de atualização de 60 Hz. Coloque este monitor a aproximadamente 82 cm de distância do assunto.
NOTA: Usamos um monitor de 24 polegadas com taxa de atualização definida em 144 Hz localizado a 82 cm de distância do participante. Recomenda-se o uso de um monitor com tamanho de tela de pelo menos 19 polegadas para experimentos cognitivos envolvendo o registro de EEG e movimentos oculares. Além disso, uma recomendação geral é colocar o monitor a uma distância que permita ao participante realizar a tarefa confortavelmente e manter um olhar estável na tela, permitindo registros precisos do EEG e dos movimentos oculares. É aconselhável testar e ajustar a configuração conforme necessário para garantir os melhores resultados. - Use um sistema de rastreamento ocular para dar aos participantes feedback em tempo real sobre seus movimentos oculares durante os períodos de execução e registre o tamanho da pupila. Defina a taxa de amostragem em 1.000 Hz para uma resolução temporal adequada.
- Evite movimentos da cabeça. Restrições de movimento da cabeça para a esquerda e para a direita e para cima e para baixo são necessárias para manter o olho dentro do campo de visão da câmera de vídeo. A restrição de movimento para frente e para trás é necessária para manter o olho no alcance focal da câmera de vídeo. Use uma combinação de apoios de testa/queixo para manter os movimentos dentro dessa faixa.
- Avalie a luminância dos estímulos usando um luxímetro digital ou similar para comparar as diferenças entre os estímulos de planejamento e os estímulos de controle.
NOTA: Um teste estatístico como o teste t ou Wilcoxon pode ser usado para avaliar as diferenças entre os estímulos das duas condições. - Use um joystick de controle ou teclado com pelo menos quatro botões: dois botões para perguntas Sim/Não da condição de controle; um desses dois botões para terminar as tentativas; e outros dois botões para o período de resposta da condição de planejamento para avançar ou retroceder para inserir os animais em cada um dos quatro círculos na parte inferior da tela.
5. Sessões de gravação de eletroencefalografia e rastreamento ocular
- Antes de iniciar o estudo, peça aos participantes que preencham o consentimento informado por escrito e assinado.
- Antes da sessão de gravação, peça aos participantes que não compareçam usando maquiagem (rímel e delineador podem ser detectados como uma pupila pelo sistema de rastreamento ocular), tendo tomado drogas ou cafeína107,108, ou se sentirem fadiga severa109 (estresse, privação de sono, etc.).
- Peça aos participantes que preencham uma pesquisa demográfica para fornecer informações sobre sexo, idade, lateralidade, língua nativa e histórico neuropsiquiátrico por meio da entrevista neuropsiquiátrica MINI-International97 aplicada por um psicólogo treinado.
- Limpe a testa, o couro cabeludo, as mastóides e a posição da pele do eletrooculograma (EOG) do sujeito com um lenço umedecido com álcool.
- Coloque todos os eletrodos externos no participante. Coloque o EOG horizontal bipolarmente no canto externo de ambos os olhos e o EOG vertical acima e abaixo do olho direito do participante. Coloque dois eletrodos externos na mastóide direita e esquerda para posterior rereferência.
- Meça a cabeça do sujeito e coloque a touca de EEG de tamanho correto de acordo com o sistema internacional estendido 10-20. Para fazer isso, localize e coloque o eletrodo Cz seguindo estas etapas:
- Identifique a linha média do couro cabeludo inspecionando visualmente a linha do cabelo e a parte superior do nariz. Identifique uma linha conectando esses dois pontos para definir a linha média.
- Localize o Cz. O Cz é normalmente definido como o ponto médio entre os dois pontos pré-auriculares (ou seja, os pontos localizados logo na frente de cada orelha). Localize esses pontos e, em seguida, identifique uma linha conectando-os para identificar a localização aproximada do Cz.
- Meça e marque o Cz. Meça a distância do násio (ou seja, a protuberância óssea na parte superior do nariz) até o Cz. A distância do násio ao Cz é tipicamente em torno de 53% da circunferência total da cabeça no sistema 10-20. Marque a localização do Cz usando uma caneta ou outra ferramenta de marcação.
NOTA: É importante seguir um procedimento consistente e padronizado para a colocação dos eletrodos, a fim de minimizar erros e garantir a validade dos dados do EEG. Recomenda-se desenvolver um procedimento de colocação padrão. Crie um procedimento padrão para colocar os eletrodos no couro cabeludo de cada sujeito e certifique-se de que o mesmo procedimento seja usado para cada indivíduo. Caso haja uma equipe ou equipe realizando as gravações, treine os técnicos ou assistentes de pesquisa sobre o procedimento de colocação adequado para garantir que eles coloquem os eletrodos de forma consistente e precisa. Além disso, digitalizar as posições dos eletrodos para cada sujeito seria uma abordagem desejável para realizar a análise da fonte posteriormente. Em estudos de EEG, a localização tridimensional (3D) precisa de cada eletrodo na cabeça do sujeito é frequentemente um requisito para estabelecer uma correlação entre os dados do EEG e a atividade cerebral correspondente110. Essas informações também são críticas para o alinhamento adequado dos dados do EEG com imagens anatômicas derivadas de técnicas de ressonância magnética ou tomografia computadorizada111,112.
- Insira o gel condutor em cada orifício da tampa usando uma seringa com uma agulha cega, afastando o cabelo do participante com a ponta. Depois, coloque todos os eletrodos do couro cabeludo na touca de EEG.
- Verifique as impedâncias usando o software de gravação de EEG e certifique-se de que estejam abaixo do nível de resistência recomendado pelo sistema de EEG.
- Peça ao participante para ficar o mais imóvel possível durante o experimento. Inspecione o sinal de EEG e teste-o pedindo ao participante que pisque, faça uma mandíbula e permaneça alguns segundos com os olhos fechados.
- Sente o participante em uma sala escura e com atenuação de som. Use um apoio de queixo para estabilizar a cabeça e minimizar o movimento, e verifique se há uma distância de aproximadamente 82 cm entre o apoio de queixo e o centro da tela de apresentação do estímulo.
- Coloque um joystick ou teclado na frente do participante para as respostas.
- Instruções: Dê instruções orais usando recursos visuais antes de iniciar cada condição (planejamento e controle). Nas instruções, inclua exemplos visuais dos estímulos e explique como resolver os labirintos nas condições de planejamento e controle, respectivamente.
- Para a tarefa de planejamento, instrua os sujeitos a encontrar um caminho para completar uma sequência de visitas a determinados locais de animais (quatro locais neste estudo) em diferentes locais do labirinto em qualquer ordem e seguindo um conjunto de regras: "(1) Planeje o caminho o mais rápido possível em no máximo 10 s; (2) Comece no portão e conclua o caminho no quarto animal visitado; (3) Não passe pelo mesmo caminho ou curva duas vezes; (4) Não atravesse um beco sem saída; (5) Não cruze um caminho perpendicularmente"54.
NOTA: Para a regra 2 à regra 5, recomendamos mostrar exemplos visuais ao participante. - Depois, inicie a sessão de treinamento de tarefas de planejamento de seis tentativas.
NOTA: Instrua os participantes a relatar antes de cada período de calibração do rastreador ocular se houve algum problema na execução da tarefa, especialmente durante o delineamento do caminho no período de execução. Anote se houve um teste para verificar offline/pós-processamento (consulte a etapa 6.1.1). - Para a condição de controle, instrua os sujeitos a avaliar se o caminho já marcado no labirinto foi feito corretamente ou incorretamente considerando as regras aprendidas anteriormente.
NOTA: Dê exemplos com suporte visual de como avaliar os labirintos sem usar estratégias de planejamento, como não tentar planejar um novo caminho quando erros forem detectados (como desenhos usando o mesmo caminho duas vezes, cruzando um beco sem saída, etc.). Quando um erro é encontrado, o foco deve ser apenas relatar a detecção do erro, em vez de corrigir o caminho. Após cada tentativa, pergunte aos participantes sobre as estratégias que implementaram. Em seguida, forneça feedback oral sobre seu desempenho para garantir que eles avaliaram os caminhos traçados e evitaram planejar novos caminhos. Em seguida, inicie a sessão de treinamento da tarefa de controle de seis tentativas.
- Para a tarefa de planejamento, instrua os sujeitos a encontrar um caminho para completar uma sequência de visitas a determinados locais de animais (quatro locais neste estudo) em diferentes locais do labirinto em qualquer ordem e seguindo um conjunto de regras: "(1) Planeje o caminho o mais rápido possível em no máximo 10 s; (2) Comece no portão e conclua o caminho no quarto animal visitado; (3) Não passe pelo mesmo caminho ou curva duas vezes; (4) Não atravesse um beco sem saída; (5) Não cruze um caminho perpendicularmente"54.
- Verifique o sinal de EEG para certificar-se de que todos os canais estão sendo adquiridos corretamente. Inicie a gravação do EEG.
- Calibre o rastreador ocular.
NOTA: Verifique a capacidade do rastreador ocular de determinar a posição do olhar quando o participante direciona o olhar para várias regiões da tela.- Informe ao participante que o rastreador ocular será calibrado e que ele verá um círculo branco (com um pequeno ponto cinza) movendo-se aleatoriamente para os quatro cantos da tela (procedimento de calibração de cinco pontos). Instrua-os a fixar o olhar no círculo e informe-os de que, quando ele se mover para outro local, eles devem seguir a posição do círculo e fixar o olhar novamente nessa nova posição.
- Execute o experimento, comece a salvar os movimentos dos olhos clicando em Saída/Registro e peça ao participante que siga as instruções dadas anteriormente, informando que o experimento começará agora.
- Mantenha a sala do laboratório em um ambiente escuro. As maiores mudanças na dilatação da pupila ocorrem em resposta a mudanças na luminância113. Mantenha um nível de luz consistente no ambiente experimental.
6. Análise dos dados
- Análises comportamentais
- Analise os dados comportamentais usando software estatístico (consulte Tabela de Materiais). Meça a precisão (taxa percentual de respostas precisas) como um parâmetro comportamental quantitativo nas condições de planejamento e controle. Para a condição de planejamento, use dados do rastreador ocular (coordenadas x e y da posição do olhar) para recriar os caminhos percorridos durante o período de execução offline e determinar a precisão dos caminhos planejados em comparação com os caminhos rastreados reais (Figura 4). Para isso, verifique manualmente a congruência entre as combinações feitas corretamente/incorretamente no período de resposta e o rastreamento feito.
- Calcule o RT, que é o tempo médio gasto resolvendo os labirintos para o período de planejamento e o tempo médio gasto avaliando os caminhos marcados para o período de controle.
- Calcular o RT médio dos períodos de execução para as condições de planeamento e controlo. Especificamente, use o RT correspondente apenas aos testes corretos.
NOTA: Complementarmente, é possível utilizar o escore linear integrado de velocidade-precisão (LISAS)114,115 descrito em Domic-Siede et al.54, que fornece uma medida combinada que considera o tempo de reação e a precisão. Como o tempo de reação durante o período de execução do planejamento e a precisão do planejamento estão inter-relacionados, o LISAS pode ser utilizado para calcular um índice que contabiliza o tempo de reação corrigido pelo número de erros cometidos. Além disso, o índice LISAS pode ser usado para avaliar a correlação entre sinais eletrofisiológicos e desempenho comportamental também. É calculado como uma combinação linear de tempo de reação (RT) e proporção de erro (PE). - Avalie a homocedasticidade usando um teste estatístico como o Teste de Levene116,117 e teste a normalidade usando o teste de distribuição de normalidade omnibus de D'Agostino e Pearson118 ou o teste de Shapiro Wilk119 para escolher o teste estatístico adequado para comparações (paramétricas ou não paramétricas).
- Avalie se o componente de planejamento na condição de planejamento é mais exigente cognitivamente do que a condição de controle usando o teste de postos sinalizados de Wilcoxon120 ou o teste t de pares pareados121 para comparar os parâmetros comportamentais entre as condições.
NOTA: Desta forma, valide se o paradigma comportamental é ideal para avaliar o planejamento cognitivo. - Separe os ensaios na condição de planejamento em categorias "fáceis" e "difíceis" (consulte a etapa 2.2) e, em seguida, use um teste t de pares pareados para comparar a precisão e os tempos de reação nos períodos de planejamento e execução entre os ensaios "fáceis" e "difíceis".
- Pré-processamento de EEG e movimento ocular
- Execute o pipeline de pré-processamento de dados de EEG explicado nos pontos a seguir usando scripts de fabricação própria e/ou caixas de ferramentas estabelecidas, como as descritas em Delorme e Makeig122, em Dimigen et al.123 e em Mognon et al.124, em um software de linguagem de programação (ver Tabela de Materiais).
- Sincronize a atividade do movimento ocular com as gravações de EEG para importar as fixações, sacadas e eventos de piscar para melhor inspeção visual ou análises adicionais (consulte a etapa 3.1.2 e o Arquivo Suplementar).
NOTA: Neste estudo, usamos os carimbos de data/hora nos dados de rastreamento ocular e os carimbos de data/hora nos dados de EEG, conforme descrito em Domic-Siede et al.54 e em Dimigen et al.123 para importar os eventos de movimento ocular para os dados de EEG em um software de linguagem de programação. - Reduza a amostra dos dados para 1.024 Hz, caso tenham sido gravados a 2.028 Hz, para reduzir as demandas computacionais.
NOTA: Uma taxa de amostragem de 1,024 Hz é suficiente de acordo com a faixa de frequência de interesse de 4-8 Hz, a resolução de frequência esperada e os requisitos computacionais da análise. - Refaça a referência do sinal de EEG à média dos eletrodos nas mastóides.
NOTA: Outras referências são possíveis. A escolha da referência pode afetar os resultados da análise do EEG e a interpretação dos dados, por isso é importante considerar cuidadosamente os prós e contras das diferentes opções de referência. A referência mastóide média é uma escolha popular para estudos de EEG porque fornece uma referência estável que é fácil de calcular e tem se mostrado eficaz para analisar muitos sinais diferentes de EEG. Referenciar os dados de EEG à média das mastóides (conhecida como referência média da mastóide) é uma abordagem comum para analisar a atividade frontal nos dados de EEG do couro cabeludo. Os eletrodos mastóideos estão localizados próximos à orelha e fornecem uma referência para os sinais de EEG. A referência à média das mastóides pode ajudar a reduzir a influência de ruídos e artefatos que não são de interesse, evitando o cancelamento do sinal de interesse, o que ajuda o usuário a obter uma representação mais clara dos sinais de EEG. - Aplique uma resposta de impulso finito de fase zero (FIR) com uma frequência de corte passa-alta de 1 Hz e uma frequência de corte passa-baixa de 40 Hz sobre o sinal estendido (sem epoching) usando um software de linguagem de programação.
NOTA: Neste estudo, usamos a caixa de ferramentas descrita em Delorme e Makeig122. - Para cada condição, considerando o número de tentativas, divida os dados em épocas centradas no início dos períodos de planejamento e controle, respectivamente. Use 1 s antes do início da apresentação do labirinto como linha de base e 4 s após o período de planejamento ou controle como os segmentos de interesse. Use um software de linguagem de programação.
NOTA: Neste estudo, usamos a caixa de ferramentas descrita em Delorme e Makeig122e 36 épocas/ensaios. - Crie uma segunda segmentação centrada no final dos períodos de planejamento e controle usando 4 s antes do final e 1 s depois como o período de manutenção.
NOTA: A razão para selecionar o primeiro e os últimos 4 s dos períodos de planejamento e controle (etapa 6.2.6 e etapa 6.2.7) é que a duração de cada período em ambas as condições pode variar, e a análise do primeiro e do último segundos do planejamento pode fornecer uma visão mais abrangente do processo de planejamento. Assim, esses comprimentos de janela são adequados e suficientes para analisar a dinâmica oscilatória subjacente ao planejamento. - Sobre o sinal segmentado, execute o algoritmo de análise de componentes independentes (ICA) da Logistic Infomax125para identificar e remover componentes artefactuais.
- Use o critério de razão de variância sacadémico-fixação recomendado em Plöchl et al.126para detectar automaticamente potenciais componentes ruidosos e use o detector automático de artefatos de EEG com base no uso conjunto de características espaciais e temporais recomendadas em Mognon et al.124.
NOTA: Recomendamos o uso do classificador de componentes independentes proposto em Pion-Tonachini et al.127, que estima classificações de componentes independentes como vetores composicionais em sete categorias, ajudando a identificar artefatos. - Inspecione outros componentes artefactuais potenciais, como EMG, movimento do eletrodo ou componentes não relacionados ao cérebro. Valide a rejeição desses componentes inspecionando visualmente as topografias, espectros e ativações ao longo do tempo.
- Interpolar (interpolação esférica) canais ruidosos por rejeição automática de canais usando o critério de curtose (com um z-score de 5 como limite).
- Análises de tempo-frequência
- Execute uma FFT (transformada rápida de Fourier) de curta duração (1 Hz a 40 Hz) usando um comprimento de janela de 250 ms e um intervalo de tempo de 5 ms. Use uma janela de Hanning. Use o escore z para normalizar os gráficos de frequência de tempo para a linha de base (−1 s a −0,1 s).
NOTA: A visualização do espectro está sujeita a uma compensação entre o tamanho da janela e a resolução temporal. Para obter uma visão abrangente de todo o espectro, incluindo a faixa de 4 Hz a 8 Hz, recomendamos usar o limite inferior do tamanho da janela, que é de 250 ms, para garantir uma resolução temporal mais alta durante cada teste e tarefa. Além disso, recomendamos o uso de uma janela Hanning, pois ela é amplamente considerada uma escolha convencional para esses casos. Para obter uma melhor resolução no tempo e na frequência, consulte as etapas a seguir. - Selecione um gráfico de frequência de tempo de um eletrodo frontocentral, como o Fz, ou um grupo médio de eletrodos frontais.
NOTA: Considere as amplas evidências sobre a associação entre controle cognitivo e da linha média frontal 12,128,129. - Selecione gráficos de tempo-frequência de controle não frontal de eletrodos como os eletrodos Pz e Oz para promover as comparações.
- Para os eletrodos frontal e de controle, realizar um teste de permutação baseado em cluster não paramétrico para amostras pareadas, com um valor de p < 0,05 para as comparações em nível de grupo dos gráficos de tempo-frequência de ambas as condições. Use o método de Monte Carlo com 1.000 sorteios aleatórios. Use o valor estatístico máximo do cluster para realizar o teste de permutação130.
- Calcule a média da banda de frequência (4-8 Hz) dos primeiros 4 s de planejamento e controle, respectivamente, e também do último segmento de 4 s.
- Compare a atividade média entre as condições usando um teste t de pares pareados ou teste de postos sinalizados de Wilcoxon.
- Analise o perfil de tempo da atividade. Para fazer isso, calcule a média da faixa de frequência de 4-8 Hz nas tentativas por sujeito.
- Compare a dinâmica da atividade entre as condições usando um teste de classificação sinalizada de Wilcoxon emparelhado e corrigido com a taxa de descoberta falsa (FDR).
NOTA: Usamos passos de 88 ms de janelas não sobrepostas no teste de Wilcoxon.
- Execute uma FFT (transformada rápida de Fourier) de curta duração (1 Hz a 40 Hz) usando um comprimento de janela de 250 ms e um intervalo de tempo de 5 ms. Use uma janela de Hanning. Use o escore z para normalizar os gráficos de frequência de tempo para a linha de base (−1 s a −0,1 s).
- Reconstrução da fonte
- Use uma caixa de ferramentas para reconstrução de análise de fontes, como a caixa de ferramentas de acesso aberto descrita em Tadel et al.131 ou outra semelhante.
- Calcule as fontes do sinal de EEG pré-processado dos primeiros 4 s de planejamento usando um algoritmo como a tomografia eletromagnética cerebral de baixa resolução padronizada (sLORETA)132e o método de imagem de norma mínima, bem como o método dos elementos de contorno simétrico (BEM simétrico), com a ajuda de uma caixa de ferramentas como a descrita em Gramfort et al.133para resolver o problema inverso.
- Use o algoritmo de origem (algoritmo sLORETA neste estudo) em um modelo MNI anatômico (usamos o modelo MNI no Brainstorm "Colin27") com os locais padrão dos eletrodos para cada participante, caso não haja digitalização 3D dos eletrodos (consulte a etapa 5.6).
NOTA: Deve-se notar que usar os locais padrão dos eletrodos não é o método mais eficiente para determinar as fontes de atividade cerebral. No entanto, ainda pode fornecer uma compreensão geral da origem da atividade. É importante ter em mente que as fontes de localização obtidas por meio desses métodos são aproximações grosseiras e devem ser interpretadas com cautela durante a análise dos resultados. - Aplique um filtro passa-banda de 4-8 Hz sobre o sinal pré-processado.
- Aplique uma normalização de escore z usando o período de −1.000 ms a −10 ms antes do início do estudo como linha de base.
- Calcule a média da atividade usando uma janela de tempo de interesse entre 1 s e 4 s após o início do teste.
- Compare as fontes de espaço médio entre as condições usando um teste de sinal de permutação não paramétrico com amostragem de Monte Carlo (1.000 randomizações)131.
- Para determinar as regiões de interesse (ROIs), rotule o córtex usando um atlas cerebral.
NOTA: Utilizou-se o Destrieux Atlas134 implementado na caixa de ferramentas descrita em Tadel et al.131. - Selecione as regiões cerebrais de interesse (ROIs).
NOTA: Consideramos as evidências relatando que as regiões do córtex pré-frontal, como os giros frontais superiores bilaterais (SF), giros e sulcos frontopolares transversais bilaterais (FP), ACC bilateral, CCM bilateral e córtex pré-frontal dorsolateral bilateral137,138, estão envolvidas nas funções de controle cognitivo135,136. - Realize a análise de componentes principais (PCA) sobre o sinal de EEG pré-processado anterior (faixa de 1-40 Hz) para cada ROI e use o primeiro modo de decomposição do PCA para cada ROI.
- Realize uma análise espectral usando uma transformada de Fourier rápida de curto prazo e compare os resultados entre as regiões esquerda e direita de interesse usando um teste de permutação não paramétrico baseado em cluster130.
- Extraia e represente as ROIs esquerda e direita que não mostram diferenças como uma série temporal bilateral: SF, ACC e MCC. Em seguida, plote gráficos de tempo-frequência e compare entre as condições.
- Compare os gráficos de tempo-frequência de acordo com o nível de complexidade da tarefa de planejamento (tentativas fáceis versus difíceis) para cada ROI.
- Espelhe a borda do sinal para cada uma das 512 amostras e execute um filtro passa-banda entre 4 Hz e 8 Hz para as ROIs selecionadas.
- Aplique uma transformada de Hilbert para obter a amplitude instantânea139usando uma caixa de ferramentas de processamento de sinal de um software de linguagem de programação (consulte Tabela de Materiais).
- Corrija o sinal usando a normalização do escore z (−1.000 a −10 ms como linha de base) e calcule a média entre os ensaios por sujeito.
- Compare cada perfil de tempo de banda ROI entre as condições usando o teste de postos sinalizados de Wilcoxon (pares correspondentes, 1 s de janelas não sobrepostas) e corrija com o FDR.
- Correlações entre a atividade do EEG e o desempenho comportamental
- Normalize a série temporal de origem dos ROIs para a linha de base usando o escore z. Selecione uma janela de 1 s a 4 s após o início do planejamento ou controle (onde é observada atividade proeminente nos gráficos de frequência de tempo).
- Para determinar o aumento da atividade na condição de planejamento em comparação com a condição de controle, primeiro transforme o sinal no domínio da frequência (1-40 Hz) usando o método multitaper por meio de uma caixa de ferramentas como a caixa de ferramentas Chronux140 para cada condição e fonte nas regiões de interesse.
- Calcule a frequência média da banda (4-8 Hz) e calcule duas medidas da potência: i) a diferença entre a potência durante o período de planejamento (θ planejamento) e o período de controle (θ controle), denotada como Δ, e ii) o aumento relativo na atividade, expressa como a razão entre Δ (Δ θ) e a atividade durante o período de controle (θ controle), como em Domic-Siede et al.54:
 (1)
(1)

- Calcule dois parâmetros comportamentais: iii) Δ planejamento LISAS, subtraindo o controle LISAS do planejamento LISAS, e iv) Δ execução do planejamento LISAS, subtraindo a execução do controle LISAS da execução do planejamento LISAS, como em Domic-Siede et al.54:


- Realizar as correlações rho de Spearman usando os parâmetros eletrofisiológicos e comportamentais calculados e, em seguida, corrigidos pelo FDR.
- Análise do movimento ocular: Para controlar as diferenças potenciais nos movimentos oculares para cada condição que pode resultar em diferentes dinâmicas oscilatórias, execute a seguinte análise:
- Determinar a amplitude sacádica e a velocidade de pico sacádica de todo o ensaio e de 0 s a 3,75 s durante as condições de planeamento e controlo.
- Compare os resultados usando o teste de postos sinalizados de Wilcoxon ou o teste t de pares pareados, o que for apropriado.
NOTA: Uma caixa de ferramentas como a descrita em Dimigen et al.123pode ser útil. - Calcule e avalie a coerência entre a potência do EEG de Fourier em um eletrodo frontal (por exemplo, o Fz ou um eletrodo ROI frontal médio) e a taxa de sacádica, conforme descrito em Sato e Yamaguchi141.
- Use o teste de postos sinalizados de Wilcoxon para comparar os valores da taxa de coerência poder-sacada dos primeiros 4 s de cada tentativa entre as duas condições.
Resultados
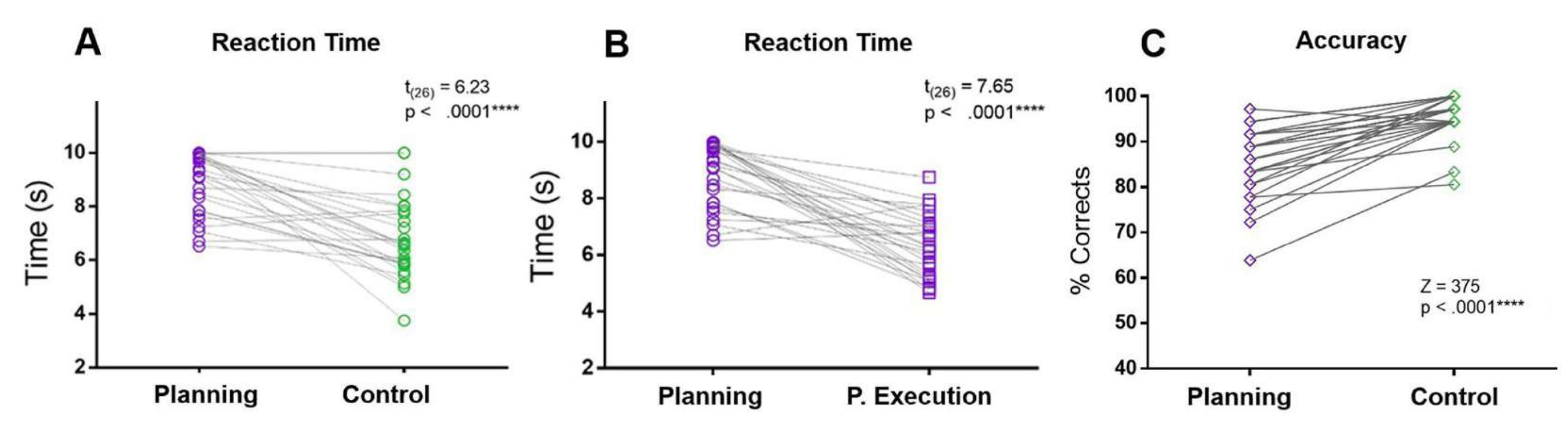
No presente protocolo, o TR do período de planejamento foi comparado com os TRs do período de controle e do período de execução do planejamento. O TR de planejamento foi maior do que os RTs de controle e período de execução de planejamento. Além disso, em comparação com a condição de controle, os participantes cometeram mais erros e tiveram menor precisão durante o período de planejamento (Figura 5).

Figura 5: Tempo de reação e precisão para a tarefa de planejamento. Comparação entre os tempos de reação (A) no período de planejamento (círculos roxos) e no período de controle (círculos verdes) usando um teste t pareado correspondente. (B) Comparação entre os tempos de reação no período de planejamento (círculos roxos) e no período de execução do planejamento (quadrados roxos) usando um teste t pareado pareado. (C) Comparação da taxa de precisão na condição de planejamento (diamantes roxos) e na condição de controle (diamantes verdes) usando um teste de classificação sinalizada de Wilcoxon. Essa figura foi modificada de Domic-Siede et al.54. Clique aqui para ver uma versão maior desta figura.
{kind=link}
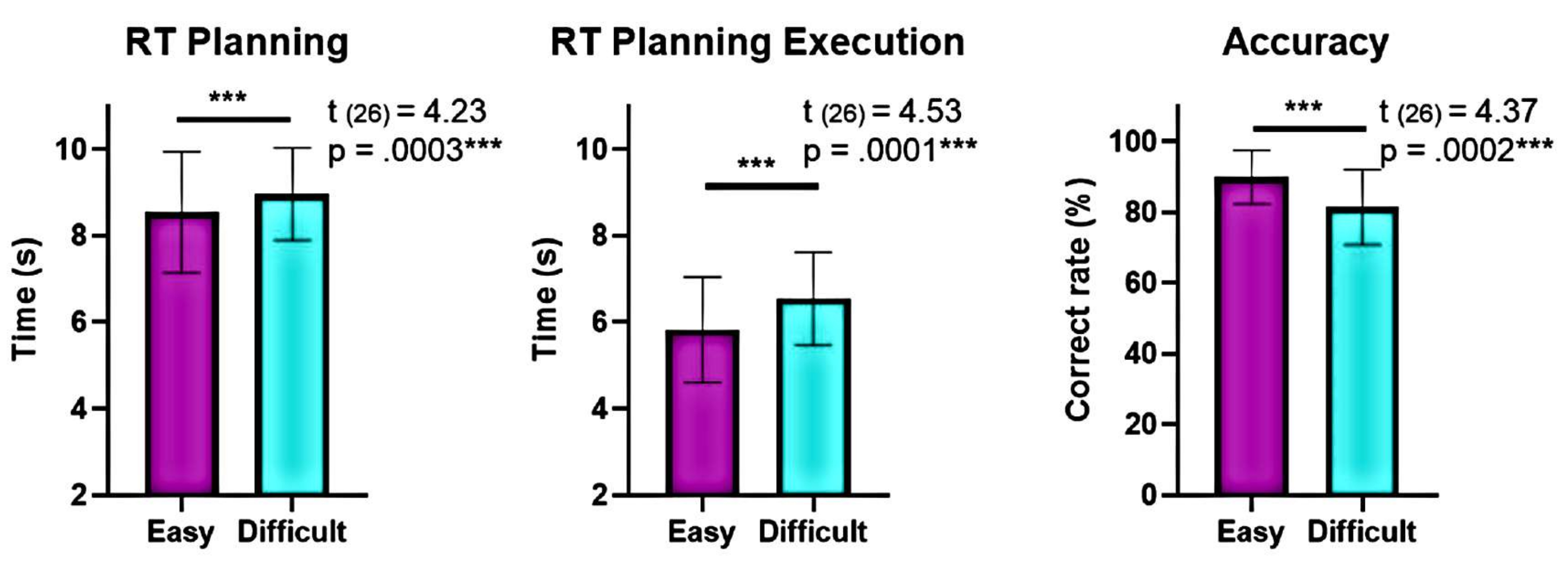
Além disso, a análise dos níveis de complexidade do planejamento mostrou diferenças significativas na precisão e nos tempos de reação (RTs) entre os níveis "difícil" e "fácil" durante o planejamento e a execução (Figura 6). O nível "difícil" teve RTs mais longos e menor precisão. Esses achados sugerem que dividir os ensaios com base no número de soluções válidas pode distinguir ensaios "fáceis" de "difíceis".

Figura 6: Comparação do desempenho comportamental nos diferentes níveis de complexidade. Diferenças significativas no desempenho comportamental nos níveis de complexidade "fácil" e "difícil" foram identificadas usando um teste t de pares pareados. Tempos de reação (RTs) mais baixos foram observados durante o planejamento e execução para o nível "fácil" em comparação com o "difícil", e a precisão foi maior para o nível "fácil". As barras de erro representam o SEM (erro padrão da média). Essa figura foi modificada de Domic-Siede et al.54. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Esses resultados indicaram que, quando o componente de planejamento foi removido com sucesso da condição de controle (por meio da manipulação de instruções), a tarefa de planejamento foi cognitivamente mais complexa, exigente, desafiadora e demorada. Assim, os correlatos neurais induzidos pelas tarefas podem ser comparados entre si.
Para analisar a atividade da linha média frontal durante o planejamento, a banda de frequência média durante o planejamento do eletrodo Fz foi comparada com a do período de controle, e um aumento significativo na frequência da banda foi encontrado durante o planejamento (Figura 7).

Figura 7: Atividade da linha média frontal durante o planejamento cognitivo. (A) Mapas topográficos representando a potência da banda em todos os sujeitos normalizados para os escores z durante a tarefa de planejamento (esquerda), a tarefa de controle (meio) e o efeito de planejamento (direita). Durante o planejamento cognitivo, os sujeitos exibiram um aumento na atividade da linha média frontal. A barra de cores mostra os valores z entre −0,5 a 1,5. (B) Um gráfico de violino mostrando os valores mínimos, quartis, mediana e máximo de pontuação z da atividade entre os indivíduos durante o planejamento (roxo) em comparação com o período de controle (verde) para os eletrodos Fz (esquerda), Pz (meio) e Oz (direita) usando um teste t de pares pareados. Essa figura foi modificada de Domic-Siede et al.54. Clique aqui para ver uma versão maior desta figura.
{kind=link}
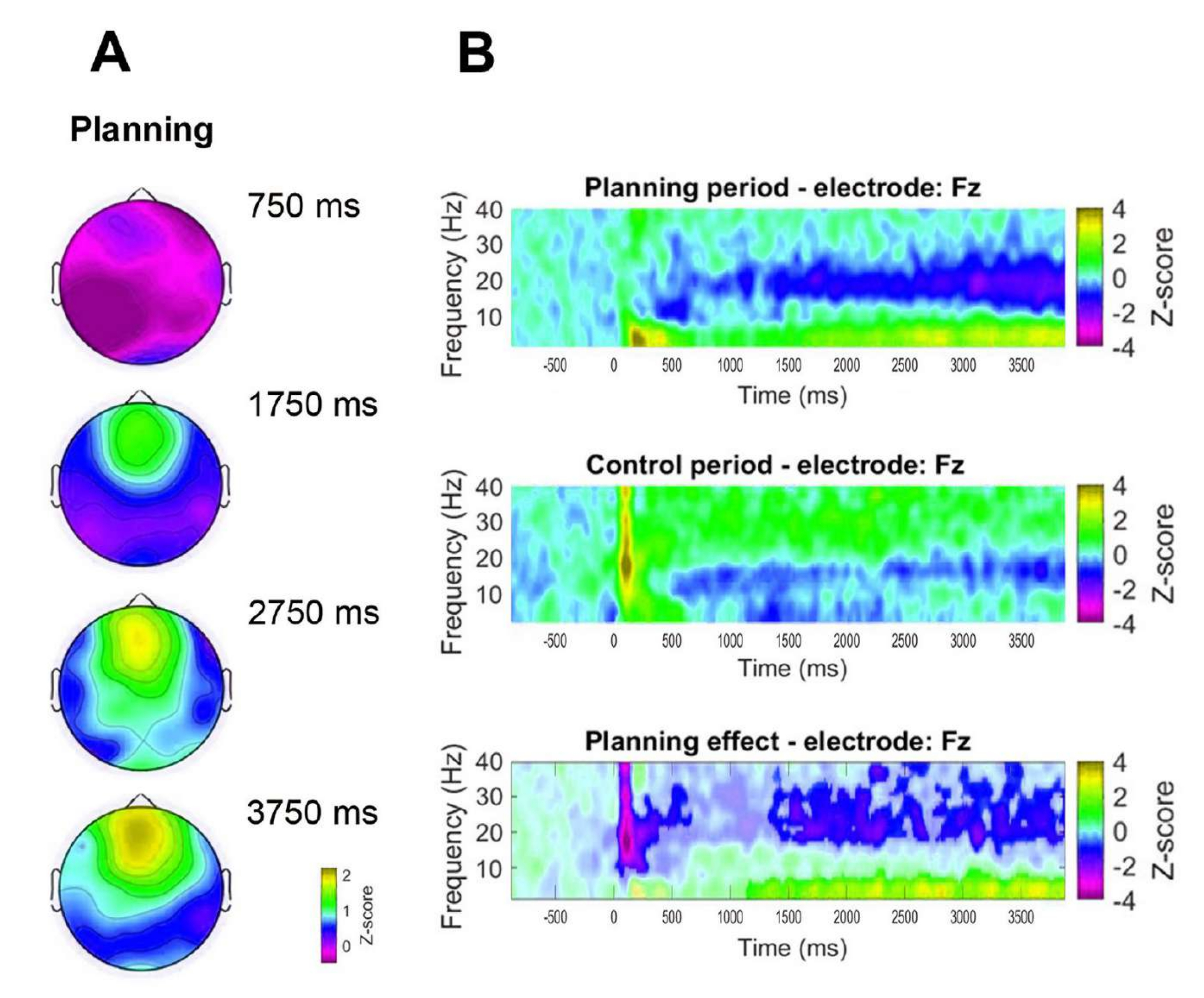
Além disso, para avaliar a dinâmica temporal da atividade frontal observada, foram formulados mapas topográficos correspondentes a pontos de tempo específicos da potência da banda (750 ms, 1.750 ms, 2.750 ms e 3.750 ms) (Figura 8A). Além disso, em comparação com o período de controle, a análise de tempo-frequência demonstrou um aumento significativo, progressivo e sustentado na atividade começando 1 s após o início do período de planejamento (Figura 8B).

Figura 8: Dinâmica temporal da linha média frontal. (A) Fatias de tempo topográficas da atividade. Foi observado um aumento progressivo na atividade da linha média frontal ao longo do tempo durante a implementação do planejamento (período de planejamento). A barra de cores indica as unidades de pontuação z (−0,5 a 2,2). (B) Gráficos de frequência de tempo para o período de planejamento (superior), o período de controle (médio) e o efeito de planejamento, calculados subtraindo o período de controle do período de planejamento (inferior). Pixels não significativos, conforme determinado usando um teste de permutação baseado em cluster não paramétrico para amostras emparelhadas, são mostrados mais claros no gráfico de efeito de planejamento. A barra de cores indica as unidades de pontuação z (−4 a 4). Essa figura foi modificada de Domic-Siede et al.54. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Para a reconstrução da fonte da atividade, um modelo de cérebro foi visualizado e comparado entre as condições, e isso indicou que a atividade se originou nas áreas do córtex pré-frontal (córtex frontal superior, FS; córtex cingulado anterior, ACC; e córtex cingulado médio, MCC), bem como que houve diferenças significativas entre as condições (no SF bilateral, o ACC bilateral e o MCC bilateral) (Figura 9), com maior atividade observada no período de planejamento (Figura 9).

Figura 9: Reconstrução da fonte. Um algoritmo sLORETA foi usado para estimar a atividade de diferentes fontes cerebrais. A atividade foi filtrada por passagem de banda de 4-8 Hz, escore z normalizado, corrigido pela linha de base, média entre 1 s ou 4 s após o início do planejamento ou controle, respectivamente, e comparada entre as condições. Um aumento significativo na atividade foi encontrado na área frontal superior bilateral, no córtex cingulado anterior bilateral e no córtex cingulado médio bilateral. A figura mostra valores t significativos do teste de permutação. Abreviaturas: FS = frontal superior; ACC = córtex cingulado anterior; MCC = córtex cingulado médio. Essa figura foi modificada de Domic-Siede et al.54. Clique aqui para ver uma versão maior desta figura.
{kind=link}
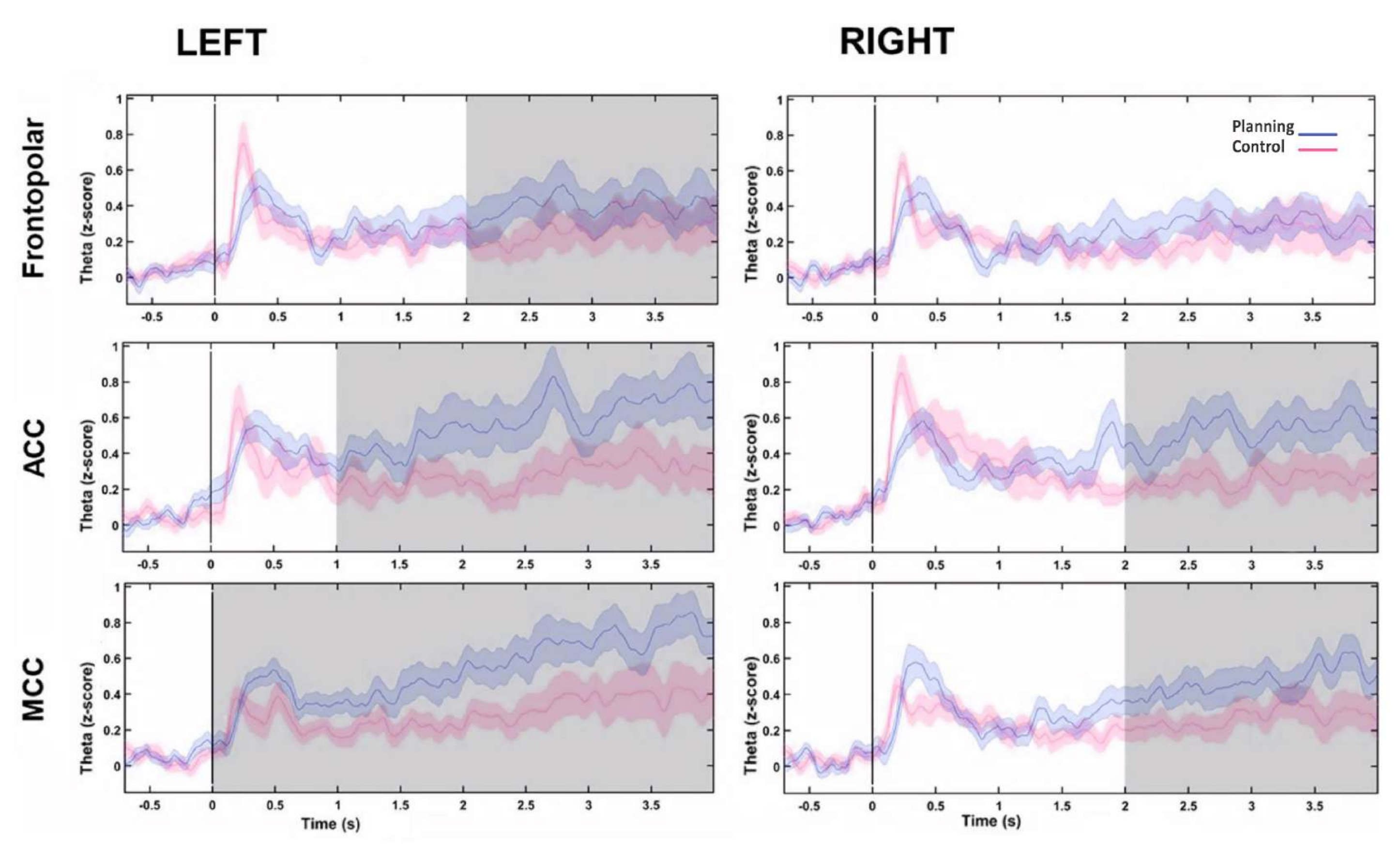
Posteriormente, o perfil temporal das mudanças ao longo do tempo para cada fonte foi avaliado calculando uma transformada de Hilbert e, em seguida, comparamos a amplitude instantânea da atividade entre as condições. Descobrimos que as fontes frontopolares esquerdas, ACC bilateral e CCM bilateral apresentaram maior atividade após o início do período de planejamento (Figura 10). Esses resultados sugeriram que nosso paradigma experimental exigindo planejamento cognitivo induziu a atividade originada nas regiões do PFC.

Figura 10: Perfil de tempo de atividade das fontes PFC. A amplitude instantânea calculada com a transformada de Hilbert foi aplicada ao primeiro componente da decomposição do PCA para cada fonte frontal e ambas as condições e linha de base normalizaram para o escore z para mostrar a atividade frontal ao longo do tempo. As áreas sombreadas em cinza mostram diferenças significativas determinadas por meio de uma janela móvel não sobreposta com passos de 1 s (teste dos postos sinalizados de Wilcoxon) corrigidos pelo FDR. As regiões sombreadas representam intervalos de confiança de 95%. A região PF esquerda, o ACC bilateral e o MCC bilateral mostraram aumentos na atividade após o início do planejamento. Abreviaturas: ACC = córtex cingulado anterior; MCC = córtex cingulado médio. A condição de planejamento é mostrada em roxo. A condição de controle é mostrada em vermelho. Essa figura foi modificada de Domic-Siede et al.54. Clique aqui para ver uma versão maior desta figura.
{kind=link}
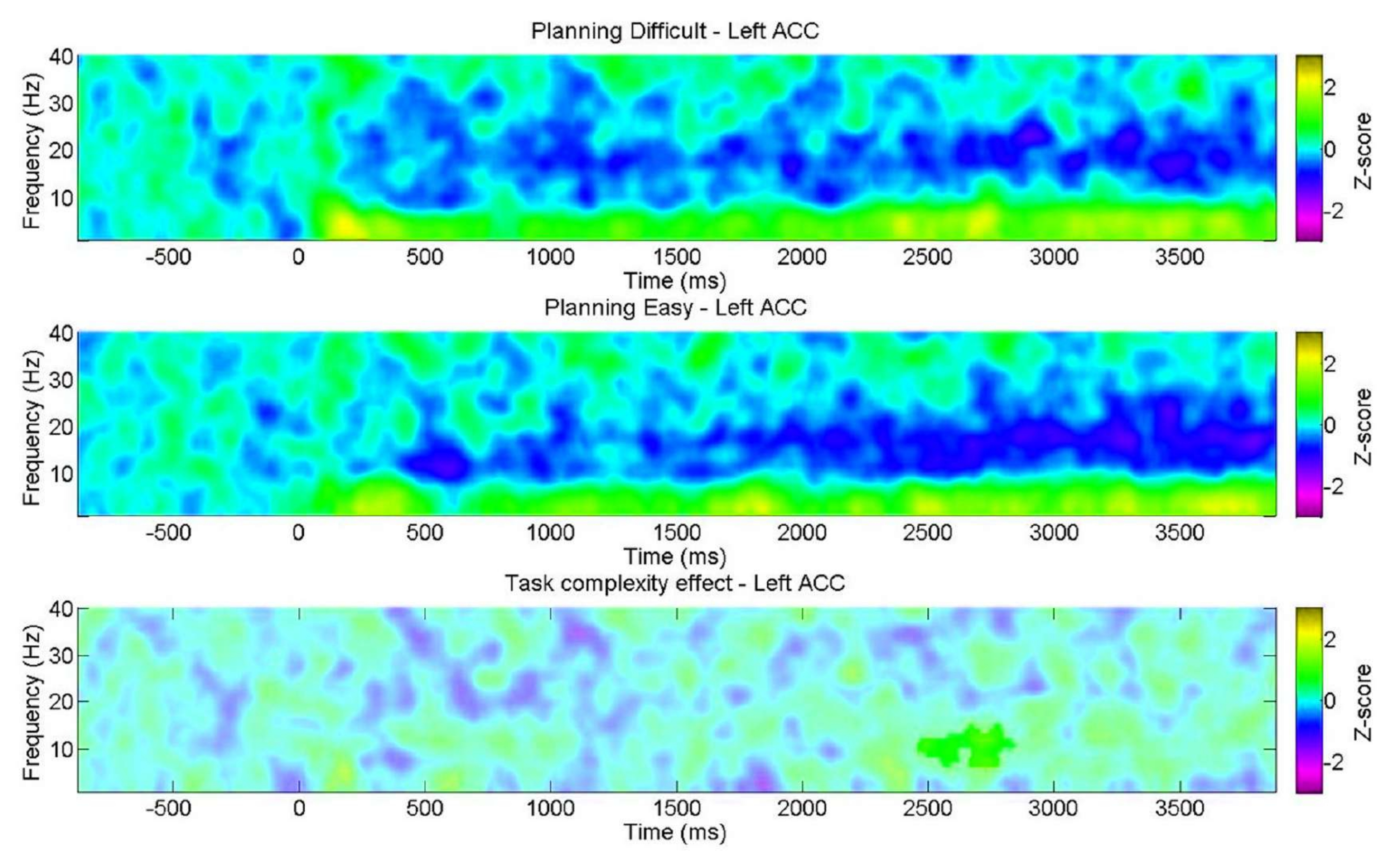
Além disso, pretendemos examinar a variação nas características espectrais durante o planejamento em relação ao nível de complexidade, conforme indicado pelos resultados comportamentais. É importante notar que uma discrepância significativa foi encontrada apenas no ACC esquerdo dentro da banda alfa. Isso apóia a noção de que nossa tarefa de planejamento avalia as facetas intrínsecas do planejamento por meio de mudanças nas oscilações em maior extensão do que as demandas cognitivas gerais (esforço) normalmente encontradas em tarefas de controle cognitivo (Figura 11).

Figura 11: EEG nos níveis de complexidade do planejamento. Os gráficos de tempo-frequência ROI mostraram um cluster positivo significativo na banda alfa exclusivamente no córtex cingulado anterior esquerdo (ACC) para o nível "difícil". Pixels não significativos, conforme determinado usando um teste de permutação baseado em cluster não paramétrico para amostras emparelhadas, são mostrados em um tom mais claro no gráfico, com a barra de cores indicando as unidades de pontuação z de −3 a 3. Essa figura foi modificada de Domic-Siede et al.54. Clique aqui para ver uma versão maior desta figura.
{kind=link}
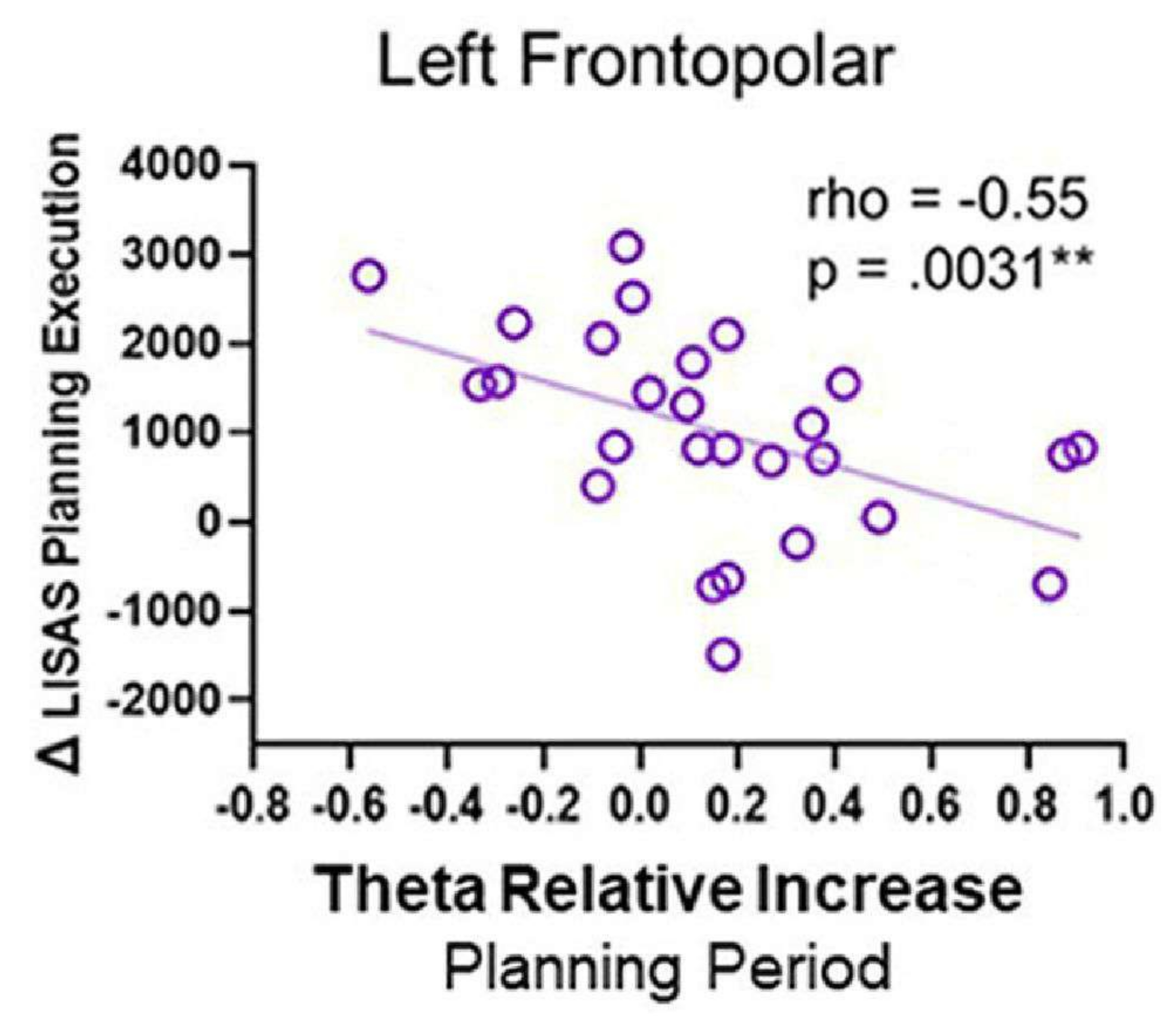
Quando foram realizadas correlações entre a atividade e o desempenho comportamental, observou-se correlação negativa; especificamente, à medida que a atividade na região frontopolar esquerda durante o período de planejamento aumentou, a pontuação de execução de planejamento do LISAS diminuiu (Figura 12). Esse padrão pode refletir que a região FP esquerda pode ser necessária durante a elaboração do planejamento para executar um plano com sucesso posteriormente e sugere um papel para a atividade.

Figura 12: Atividade e desempenho comportamental. A correlação rho de Spearman entre a atividade do córtex frontopolar esquerdo e a execução do planejamento Δ LISAS mostrou uma correlação negativa significativa. Essa figura foi modificada de Domic-Siede et al.54. Clique aqui para ver uma versão maior desta figura.
{kind=link}
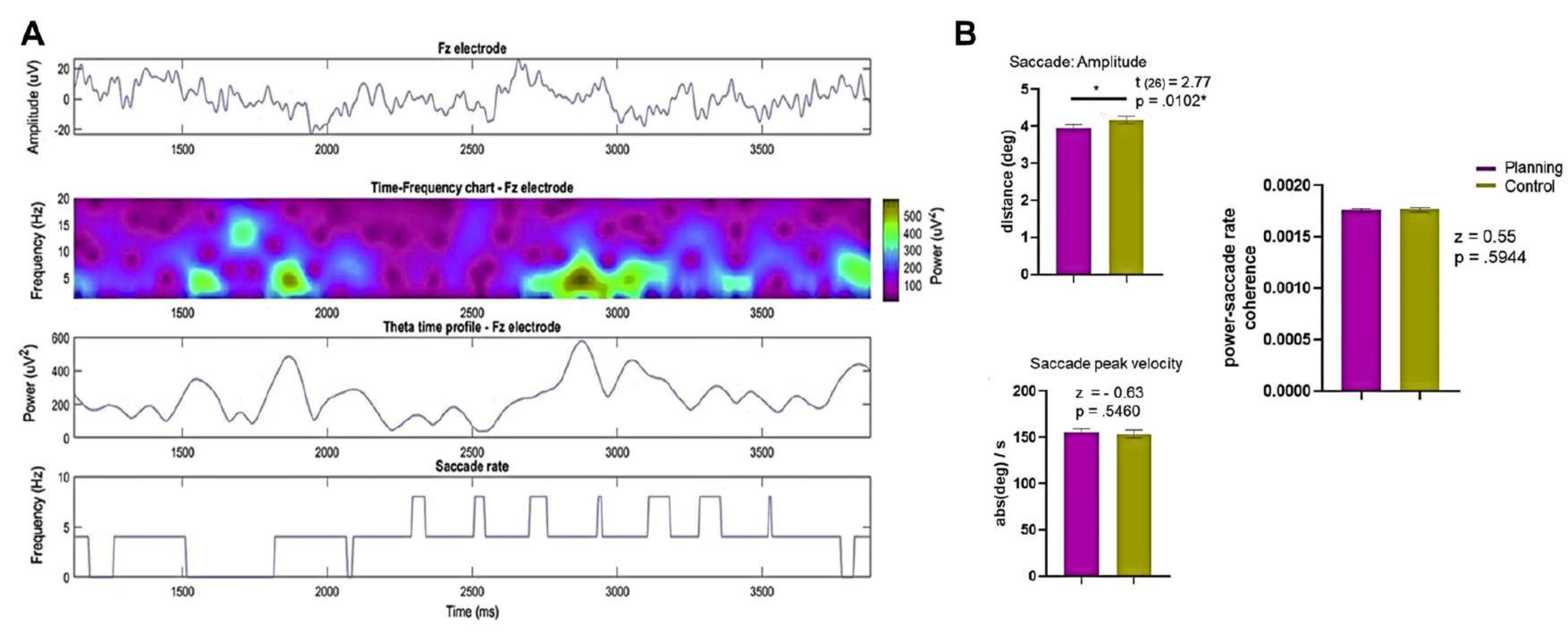
Além disso, as diferentes demandas e objetivos cognitivos induzidos por cada condição podem ter causado movimentos oculares contrastantes entre as condições de planejamento e controle, levando a diferentes padrões de atividade oscilatória95. Para resolver o problema acima, analisamos os dados de um único sujeito e de um único ensaio em vários níveis. Notavelmente, a série temporal do canal Fz e a dinâmica do tempo de atividade pareciam não ter conexão com a taxa de sacadas ao longo do tempo (Figura 13A).

Figura 13: Resultados dos registros de EEG e movimento ocular. (A) As linhas apresentam o EEG (superior), o gráfico de frequência de tempo (topo médio), o perfil de tempo do eletrodo Fz (fundo médio) e a taxa sacádica do sujeito 8 e do ensaio 9 (inferior) na condição de planejamento. (B) Uma comparação do teste de postos sinalizados de Wilcoxon da amplitude sacádica, velocidade de pico sacádica e coerência da taxa de potência-sacada entre as condições é mostrada, revelando diferenças estatísticas significativas na amplitude sacádica entre as condições de planejamento e controle. O SEM é representado pelas barras de erro. Essa figura foi modificada de Domic-Siede et al.54. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Em seguida, obtivemos a amplitude sacádica e a velocidade de pico de todo o ensaio e de 0 s a 3,75 s para comparação (Figura 13B). Descobrimos que a amplitude da sacádica foi maior na condição controle. No entanto, não foram encontradas diferenças estatisticamente significativas entre as condições no índice de coerência entre a potência de Fourier no eletrodo Fz e a taxa sacádica (Figura 10B), indicando que qualquer relação potencial entre as sacadas e a atividade foi consistente entre as condições.
Em conjunto, esses resultados apóiam que o protocolo experimental descrito é adequado para estudar o planejamento cognitivo como uma função de controle cognitivo.
Discussão
O protocolo aqui descrito oferece uma abordagem inovadora para avaliar o planejamento cognitivo e o controle cognitivo durante uma tarefa de planejamento novo e ecológico em associação com medidas comportamentais e fisiológicas pertinentes e complementares, como dinâmica oscilatória e pupilar. Durante nossos experimentos, a atividade do EEG foi registrada enquanto os participantes realizavam a tarefa de planejamento, na qual os participantes foram instruídos a primeiro elaborar e depois executar um plano. A condição de controle, que envolvia a avaliação de um caminho pré-desenhado no mapa do zoológico, foi estabelecida para eliminar o aspecto do planejamento cognitivo, mantendo uma configuração e estrutura semelhantes. Essa abordagem permite avaliar se o planejamento cognitivo, como uma função de controle cognitivo, leva à geração de atividade frontal a partir de regiões PFC e se diferentes fontes de oscilação PFC estão ligadas a diferentes aspectos do desempenho do planejamento. Outro aspecto que pode ser avaliado usando este protocolo é a diferenciação dos vários processos cognitivos envolvidos durante as fases de planejamento, como elaboração do plano, execução do plano, realização do plano e processamento de feedback. Descobrimos que o planejamento induziu uma atividade frontal canônica associada ao controle cognitivo, o que contribuiu para atingir um objetivo de forma eficiente. Estes resultados reforçam o interesse deste protocolo.
Apesar dos vastos desenvolvimentos na neurociência cognitiva, a maioria dos experimentos de neuroimagem examina funções cognitivas isoladas usando tarefas artificiais em ambientes sensório-motores e estímulos simplificados para controlar fatores de confusão. Problematicamente, esses experimentos podem não ser capazes de identificar os mecanismos cerebrais reais envolvidos quando uma função cognitiva é implementada em uma situação cotidiana (durante situações ecológicas) 138 , 140 . Em particular, as capacidades para formular metas, planejar e executar planos de forma eficaz são difíceis de avaliar, pois requerem várias funções cognitivas (memória de trabalho, controle inibitório, flexibilidade cognitiva, etc.)104.144. Assim, o desenho de uma tarefa comportamental ecológica é incentivado e sugerido com base nas tendências atuais da neurociência cognitiva 142,143,145,146.
Nossa tarefa de planejamento, apesar de ocorrer em um ambiente de laboratório (dentro de uma sala com os estímulos exibidos em uma tela), tornou-se envolvente e interativa para os participantes por meio do uso de estímulos e objetivos significativos com os quais eles poderiam interagir na tela. Além disso, a tarefa exige que os participantes se envolvam em uma situação da vida real - planejando um caminho para visitar vários locais. Para ter um desenho de tarefa ecológico, o paradigma deve desafiar os sujeitos a realizar um comportamento específico ou função cognitiva de maneira semelhante ao que eles teriam que fazer na vida cotidiana 62,63,147. Para desenvolver um projeto de tarefa ecológica, a tarefa de planejamento usada aqui envolve o planejamento de um caminho para visitar lugares diferentes em vários estágios56. A primeira etapa envolve os participantes criando um plano, garantindo que ele siga um conjunto de regras. A segunda etapa envolve manter o plano na memória de trabalho, enquanto a terceira etapa envolve a execução do plano e o monitoramento de sua adesão às regras. Esses estágios representam diferentes fases de planejamento e orquestração de outras funções executivas, como flexibilidade cognitiva, controle inibitório e memória de trabalho. Para ter um desenho de tarefa cognitiva ecológica válido, a tarefa deve ser capaz de detectar deficiências cognitivas específicas em pacientes com transtornos psiquiátricos ou cognitivos que têm dificuldade em realizar essa função cognitiva específica em sua vida diária105. Isso pode ser alcançado por meio de pesquisas futuras usando este protocolo.
Os resultados comportamentais obtidos com o uso desse protocolo foram alinhados com as predições experimentais. Uma diferença significativa no comportamento foi observada quando o componente de planejamento foi removido da tarefa de controle para formar uma condição de controle, facilitando assim novas comparações. A condição de planejamento foi considerada cognitivamente mais exigente do que a condição de controle, como evidenciado por parâmetros como o tempo de reação e a precisão. Isso pode refletir o aumento do envolvimento de altas funções cognitivas na implementação do planejamento 23,55,56,57,148,149,150.
Considerando que a condição de controle envolvia cognição menos complexa, como evidenciado pelos tempos de reação mais rápidos, melhores desempenhos e diferentes processos cognitivos necessários (a avaliação de regras), uma possível modificação alternativa é explorar os níveis de complexidade presentes na tarefa de planejamento, manipulá-los e analisar a função de planejamento parametricamente de acordo com diferentes níveis de complexidade (por exemplo, aumentando o número de tentativas e criando condições difíceis, médias e fáceis). No entanto, os resultados do nosso protocolo mostraram que, embora fosse possível distinguir entre ensaios "fáceis" e "difíceis" com base em medidas comportamentais, não foram detectadas diferenças nas medidas eletrofisiológicas. Isso sugere que os resultados em nosso protocolo refletem com mais precisão as características intrínsecas da função de planejamento do que aspectos mais amplos do controle cognitivo, como atenção, esforço mental, nível de dificuldade ou alto nível de demanda cognitiva54. No entanto, pesquisas adicionais podem considerar outros tipos de condições de controle, como seguir um caminho marcado que visita os quatro locais dos animais, mas também lembrar a ordem da sequência. Dessa forma, o nível de dificuldade poderia ser melhor controlado e o planejamento poderia ser distinguido da memória de trabalho, mas uma possível desvantagem disso é a fadiga, pois os sujeitos teriam que realizar duas tarefas altamente exigentes.
Vários estudos associaram diferentes parâmetros do movimento ocular a eventos cognitivos específicos. Por um lado, alguns estudos encontraram correlações entre as oscilações e o diâmetro da pupila durante tarefas cognitivas, sugerindo uma relação entre essas duas medidas da função cognitiva. Por exemplo, Lin et al.152 encontraram uma correlação entre a atividade médio-frontal e mudanças no tamanho da pupila, refletindo vários graus de conflito subjetivo. Suas descobertas sugerem que esses sinais representam processamento de conflitos, maior atenção e respostas comportamentais flexíveis. Portanto, a relação entre a atividade frontal médio e as respostas dos alunos parece desempenhar um papel na ponderação de custos e benefícios durante os processos de tomada de decisão. Em outro estudo, Yu et al.153 examinaram como o envolvimento do tempo na tarefa afeta neurofisiologicamente o controle cognitivo por meio de uma tarefa de memória de trabalho que modula as respostas inibitórias de controle. Eles estudaram a relação entre os dados do diâmetro da pupila e a atividade frontal e mostraram que, à medida que a duração da tarefa aumentava, o desempenho diminuía, e isso era acompanhado por uma diminuição na modulação da dilatação da pupila e na atividade frontal. No início da tarefa, eles encontraram uma forte correlação entre o engajamento na tarefa, a atividade e o controle cognitivo, conforme indicado pela modulação da dilatação da pupila, principalmente para tarefas exigentes que exigiam alta memória de trabalho e controle inibitório. No entanto, essa relação se dissipou no final, sinalizando uma desconexão entre o esforço investido e o controle cognitivo usado para realizar a tarefa, que é uma marca registrada dos efeitos pré-frontais do tempo na tarefa153. Por outro lado, outros estudos investigaram sacadas e oscilações. Por exemplo, Nakatani et al.154 revelaram que, em uma tarefa perceptiva, a amplitude da banda alfa das regiões occipitais previa piscadas e efeitos sacádicos. Além disso, Velasques et al.155 mostraram que, durante uma tarefa de atenção prosacádica, a amplitude sacádica foi associada a alterações gama frontais. Além disso, Bodala et al.156 descobriram que diminuições na da linha média frontal foram acompanhadas por diminuições na atenção sustentada, bem como na amplitude e velocidade das sacadas. Essas descobertas sugerem que os movimentos oculares, especialmente as sacadas, podem refletir processos cognitivos, em vez de apenas contribuir para o ruído de fundo nos sinais de EEG. No presente estudo, aprimoramos a eliminação de artefatos relacionados ao movimento ocular usando o algoritmo ICA com o critério de razão de variância sacada-fixação126. Este critério aprimora a remoção de artefatos para tarefas de visualização livre157. Em nosso estudo, não foram observadas diferenças na velocidade de pico sacádico e na coerência entre a potência e a taxa sacádica entre as condições. No entanto, mais estudos são necessários para abordar essas questões.
Uma etapa crítica no uso desse protocolo é calibrar constantemente o rastreador ocular durante o experimento, pois a perda de dados do olhar da câmera pode resultar em erros que contaminariam a tarefa e dificultariam a obtenção de respostas precisas. Portanto, é crucial calibrar o mais rápido possível. No entanto, há uma compensação entre o número de tentativas com um rastreador ocular calibrado e a duração do experimento. Em nosso estudo, decidimos calibrar a cada cinco tentativas.
Pesquisas futuras explorando a relação entre as oscilações e o diâmetro da pupila durante esta tarefa de planejamento devem ser conduzidas. O planejamento é um aspecto crítico do controle executivo que requer a alocação de recursos atencionais e a coordenação de múltiplos processos cognitivos. Compreender a relação entre as oscilações e o diâmetro da pupila durante as tarefas de planejamento pode fornecer informações valiosas sobre os mecanismos neurais subjacentes do controle executivo e como eles mudam com o tempo. Além disso, esses estudos podem levar a uma compreensão mais profunda de como as mudanças na função cognitiva, como fadiga ou lapsos de atenção, afetam o desempenho em tarefas de planejamento e a capacidade de alocar recursos de forma eficaz. Essas informações podem ter implicações importantes para o desenvolvimento de intervenções destinadas a melhorar o desempenho do planejamento, como programas de treinamento cognitivo ou tratamentos para condições como transtorno de déficit de atenção/hiperatividade (TDAH), entre outras.
Pesquisas anteriores indicaram que o PFC desempenha um papel crucial no planejamento cognitivo, conforme confirmado por nossos resultados. Esses resultados demonstram que o planejamento cognitivo induz a atividade da FMθ no PFC, particularmente no córtex cingulado anterior, córtex cingulado médio e regiões frontais superiores54. Essas descobertas se alinham com pesquisas anteriores sobre funções executivas. Há evidências substanciais para apoiar a ideia de que a atividade da FMθ atua como um processo de cima para baixo para iniciar o controle e facilitar a comunicação entre as regiões do cérebro durante tarefas exigentes13. Embora apenas alguns estudos tenham examinado a dinâmica temporal da atividade da FMθ associada ao controle cognitivo, há um consenso generalizado de que o perfil temporal da FMθ pode fornecer informações sobre várias facetas do controle cognitivo e o envolvimento de regiões distintas do PFC. O uso de nosso protocolo para avaliar o planejamento cognitivo nos permitiu caracterizar o perfil temporal da atividade da FMθ durante o planejamento. Especificamente, a atividade de FMθ durante a condição de planejamento mostrou um aumento progressivo. Com a implementação deste protocolo, pela primeira vez, foi demonstrado que o FMθ também está presente durante a implementação do planejamento, como em outras funções cognitivas de ordem superior, e sua dinâmica temporal pode servir como um indicador de controle cognitivo.
Nossos resultados e protocolo têm aplicações potenciais no campo da neurociência, inclusive para melhorar as avaliações neuropsicológicas virtuais e o tratamento de distúrbios neuropsiquiátricos com problemas de planejamento cognitivo associados, como depressão e transtorno de déficit de atenção/hiperatividade. Por exemplo, as avaliações podem envolver o exame de diferentes padrões de erros no nível de desempenho comportamental, diferentes padrões oscilatórios no nível eletrofisiológico e diferentes movimentos oculares. Além disso, os resultados deste trabalho podem informar o desenvolvimento de interfaces cérebro-computador e programas de treinamento cognitivo que visam aprimorar as habilidades de planejamento cognitivo.
O presente protocolo pode contribuir com novas evidências para a compreensão dos mecanismos neurais que sustentam a função de controle cognitivo indescritível do planejamento cognitivo em populações neurotípicas e neuropsiquiátricas. Além disso, nosso paradigma comportamental pode oferecer insights sobre a neurobiologia do controle cognitivo e planejamento por meio do exame de medidas eletrofisiológicas, pupilométricas e comportamentais com uma tarefa prática de planejamento que examina aspectos intrínsecos do planejamento, em vez das demandas cognitivas gerais tipicamente presentes em tarefas de controle cognitivo, conforme refletido nas mudanças nas oscilações.
Divulgações
Os autores não têm nada a divulgar.
Agradecimentos
Esta pesquisa foi financiada pelo programa de bolsas de doutorado Becas de Doctorado Nacional año 2015 da ANID 21150295, FONDECYT regular grant 1180932, FONDECYT regular grant 1230383, FONDECYT de Iniciación grant 11220009, Postdoc grant Universidad de O'Higgins e o Institut Universitaire de France (IUF). Queremos agradecer ao professor Pablo Billeke por seu feedback sobre o design do paradigma. Agradecemos ao professor Eugenio Rodríguez por gentilmente compartilhar seus códigos de análise de frequência de tempo. Por fim, agradecemos a Milan Domic, Vicente Medel, Josefina Ihnen, Andrea Sánchez, Gonzalo Boncompte, Catalina Fabar e Daniela Santander pelo feedback.
Materiais
| Name | Company | Catalog Number | Comments |
| EEG System | Biosemi | ActiveTwo Base system, 64 channels | |
| Eye-tracker System | Eyelink SR Research | EyeLink 1000 Plus Core Unit, High-speed camera, Host PC/Monitor, | |
| CPU display | Intel | Hard drive 221 GB, Processor Intel Core i7-4790 3.60 Hz, OS Windows 7, 4GB RAM | |
| CPU EEG | Intel | Hard drive 223 GB, Processor Intel Core i7-4790 3.60 Hz, OS Windows 7, 4GB RAM | |
| Monitor | ASUS | ASUS VG248QE 24" LCD monitor | |
| Joytsick | Logitech | Model F310 | |
| Luxmeter | Focket | LCD screen (0-200.000 lux) model Liebe WH LX1330B | |
| Statistics software | GraphPad Prism | GraphPad Prism version 8 for Windows | |
| MATLAB Programming Software | The MathWorks | MATLAB R2014a and R2018b | |
| SVG tool Inkscape | Inkscape Project | vector graphic editor software | |
| Presentation Software | Neurobehavioral Systems | stimulus delivery and experiment control program for neuroscience |
Referências
- Siegel, M., Donner, T. H., Engel, A. K. Spectral fingerprints of large-scale neuronal interactions. Nature Reviews Neuroscience. 13 (2), 121-134 (2012).
- Fries, P. A mechanism for cognitive dynamics: Neuronal communication through neuronal coherence. Trends in Cognitive Sciences. 9 (10), 474-480 (2005).
- Fries, P. Rhythms for cognition: Communication through coherence. Neuron. 88 (1), 220-235 (2015).
- Thut, G., Miniussi, C., Gross, J. The functional importance of rhythmic activity in the brain. Current Biology. 22 (16), 658-663 (2012).
- Fröhlich, F., McCormick, D. A. Endogenous electric fields may guide neocortical network activity. Neuron. 67 (1), 129-143 (2010).
- Spaak, E., de Lange, F. P., Jensen, O. Local entrainment of alpha oscillations by visual stimuli causes cyclic modulation of perception. Journal of Neuroscience. 34 (10), 3536-3544 (2014).
- Fiebelkorn, I. C., Saalmann, Y. B., Kastner, S. Rhythmic sampling within and between objects despite sustained attention at a cued location. Current Biology. 23 (24), 2553-2558 (2013).
- Landau, A. N., Fries, P. Attention samples stimuli rhythmically. Current Biology. 22 (11), 1000-1004 (2012).
- Song, K., Meng, M., Lin, C., Zhou, K., Luo, H. Behavioral oscillations in attention: Rhythmic α pulses mediated through θ band. Journal of Neuroscience. 34 (14), 4837-4844 (2014).
- Wyart, V., Nobre, A. C., Summerfield, C. Dissociable prior influences of signal probability and relevance on visual contrast sensitivity. Proceedings of the National Academy of Sciences of the United States of America. 109 (9), 3593-3598 (2012).
- Leszczyński, M., Fell, J., Axmacher, N. Rhythmic working memory activation in the human hippocampus. Cell Reports. 13 (6), 1272-1282 (2015).
- Onton, J., Delorme, A., Makeig, S. Frontal midline EEG dynamics during working memory. NeuroImage. 27 (2), 341-356 (2005).
- Cavanagh, J. F., Frank, M. J. Frontal theta as a mechanism for cognitive control. Trends in Cognitive Sciences. 18 (8), 414-421 (2014).
- Siegel, M., Buschman, T. J., Miller, E. K. Cortical information flow during flexible sensorimotor decisions. Science. 348 (6241), 1352-1355 (2015).
- Weisz, N., et al. Prestimulus oscillatory power and connectivity patterns predispose conscious somatosensory perception. Proceedings of the National Academy of Sciences of the United States of America. 111 (4), 417-425 (2014).
- Buzsáki, G., Draguhn, A. Neuronal oscillations in cortical networks. Science. 304 (5679), 1926-1929 (2004).
- Cohen, M. X. A neural microcircuit for cognitive conflict detection and signaling. Trends in Neurosciences. 37 (9), 480-490 (2014).
- Yuste, R. From the neuron doctrine to neural networks. Nature Reviews Neuroscience. 16 (8), 487-497 (2015).
- Helfrich, R. F., Knight, R. T. Oscillatory dynamics of prefrontal cognitive control. Trends in Cognitive Sciences. 20 (12), 916-930 (2016).
- Miller, E. K., Cohen, J. D. An integrative theory of prefrontal cortex function. Annual Review of Neurosciences. 24 (1), 167-202 (2001).
- von Nicolai, C., et al. Corticostriatal coordination through coherent phase-amplitude coupling. Journal of Neuroscience. 34 (17), 5938-5948 (2014).
- Sweeney-Reed, C. M., et al. Thalamic theta phase alignment predicts human memory formation and anterior thalamic cross-frequency coupling. ELife. 4, 07578 (2015).
- Voytek, B., et al. Oscillatory dynamics coordinating human frontal networks in support of goal maintenance. Nature Neuroscience. 18 (9), 1318-1324 (2015).
- Raghavachari, S., et al. Theta oscillations in human cortex during a working-memory task: Evidence for local generators. Journal of Neurophysiology. 95 (3), 1630-1638 (2006).
- Jacobs, J., Hwang, G., Curran, T., Kahana, M. J. EEG oscillations and recognition memory: Theta correlates of memory retrieval and decision making. NeuroImage. 32 (2), 978-987 (2006).
- Itthipuripat, S., Wessel, J. R., Aron, A. R. Frontal theta is a signature of successful working memory manipulation. Experimental Brain Research. 224 (2), 255-262 (2013).
- Cavanagh, J. F., Zambrano-Vazquez, L., Allen, J. J. B. Theta lingua franca: A common mid-frontal substrate for action monitoring processes. Psychophysiology. 49 (2), 220-238 (2012).
- Mas-Herrero, E., Marco-Pallarés, J. Frontal theta oscillatory activity is a common mechanism for the computation of unexpected outcomes and learning rate. Journal of Cognitive Neuroscience. 26 (3), 447-458 (2014).
- Folstein, J. R., Van Petten, C. Influence of cognitive control and mismatch on the N2 component of the ERP: A review. Psychophysiology. 45 (1), 152-170 (2008).
- Cohen, M. X., Donner, T. H. Midfrontal conflict-related theta-band power reflects neural oscillations that predict behavior. Journal of Neurophysiology. 110 (12), 2752-2263 (2013).
- Walsh, M. M., Anderson, J. R. Modulation of the feedback-related negativity by instruction and experience. Proceedings of the National Academy of Sciences of the United States of America. 108 (47), 19048-19053 (2011).
- Luu, P., Tucker, D. M., Makeig, S. Frontal midline theta and the error-related negativity: Neurophysiological mechanisms of action regulation. Clinical Neurophysiology. 115 (8), 1821-1835 (2004).
- Hanslmayr, S., et al. The electrophysiological dynamics of interference during the Stroop task. Journal of Cognitive Neuroscience. 20 (2), 215-225 (2008).
- Cavanagh, J. F., Cohen, M. X., Allen, J. J. B. Prelude to and resolution of an error: EEG phase synchrony reveals cognitive control dynamics during action monitoring. Journal of Neuroscience. 29 (1), 98-105 (2009).
- Cohen, M. X., van Gaal, S., Ridderinkhof, K. R., Lamme, V. A. F. Unconscious errors enhance prefrontal-occipital oscillatory synchrony. Frontiers in Human Neuroscience. 3, 54 (2009).
- Cavanagh, J. F., Frank, M. J., Klein, T. J., Allen, J. J. B. Frontal theta links prediction errors to behavioral adaptation in reinforcement learning. NeuroImage. 49 (4), 3198-3209 (2010).
- Cohen, M. X., Cavanagh, J. F. Single-trial regression elucidates the role of prefrontal theta oscillations in response conflict. Frontiers in Psychology. 2, 30 (2011).
- Cohen, M. X., Van Gaal, S. Dynamic interactions between large-scale brain networks predict behavioral adaptation after perceptual errors. Cerebral Cortex. 23 (5), 1061-1072 (2013).
- Nigbur, R., Cohen, M. X., Ridderinkhof, K. R., Stürmer, B. Theta dynamics reveal domain-specific control over stimulus and response conflict. Journal of Cognitive Neuroscience. 24 (5), 1264-1274 (2012).
- van Driel, J., Swart, J. C., Egner, T., Ridderinkhof, K. R., Cohen, M. X. No) time for control: Frontal theta dynamics reveal the cost of temporally guided conflict anticipation. Cognitive, Affective, and Behavioral Neuroscience. 15 (4), 787-807 (2015).
- van de Vijver, I., Ridderinkhof, K. R., Cohen, M. X. Frontal oscillatory dynamics predict feedback learning and action adjustment. Journal of Cognitive Neuroscience. 23 (12), 4106-4121 (2011).
- Narayanan, N. S., Cavanagh, J. F., Frank, M. J., Laubach, M. Common medial frontal mechanisms of adaptive control in humans and rodents. Nature Neuroscience. 16 (12), 1888-1895 (2013).
- Anguera, J. A., et al. Video game training enhances cognitive control in older adults. Nature. 501 (7465), 97-101 (2013).
- Smit, A. S., Eling, P. A. T. M., Hopman, M. T., Coenen, A. M. L. Mental and physical effort affect vigilance differently. International Journal of Psychophysiology. 57 (3), 211-217 (2005).
- Cohen, M. X., Ranganath, C. Reinforcement learning signals predict future decisions. Journal of Neuroscience. 27 (2), 371-378 (2007).
- Gehring, W. J., Goss, B., Coles, M. G. H., Meyer, D. E., Donchin, E. A neural system for error detection and compensation. Psychological Science. 4 (6), 385-390 (1993).
- Yeung, N., Botvinick, M. M., Cohen, J. D. The neural basis of error detection: Conflict monitoring and the error-related negativity. Psychological Review. 111 (4), 931-959 (2004).
- Debener, S., et al. Trial-by-trial coupling of concurrent electroencephalogram and functional magnetic resonance imaging identifies the dynamics of performance monitoring. Journal of Neuroscience. 25 (50), 11730-11737 (2005).
- Hauser, T. U., et al. The feedback-related negativity (FRN) revisited: New insights into the localization, meaning, and network organization. NeuroImage. 84, 159-168 (2014).
- Wang, C., Ulbert, I., Schomer, D. L., Marinkovic, K., Halgren, E. Responses of human anterior cingulate cortex microdomains to error detection, conflict monitoring, stimulus-response mapping, familiarity, and orienting. Journal of Neuroscience. 25 (3), 604-613 (2005).
- Tsujimoto, S., Genovesio, A. Firing variability of frontal pole neurons during a cued strategy task. Journal of Cognitive Neuroscience. 29 (1), 25-36 (2017).
- Womelsdorf, T., Johnston, K., Vinck, M., Everling, S. Theta-activity in anterior cingulate cortex predicts task rules and their adjustments following errors. Proceedings of the National Academy of Sciences of the United States of America. 107 (11), 5248-5253 (2010).
- Womelsdorf, T., Vinck, M., Stan Leung, L., Everling, S. Selective theta synchronization of choice-relevant information subserves goal-directed behavior. Frontiers in Human Neuroscience. 4, 2010 (2010).
- Domic-Siede, M., Irani, M., Valdés, J., Perrone-Bertolotti, M., Ossandón, T. Theta activity from frontopolar cortex, mid-cingulate cortex and anterior cingulate cortex shows different roles in cognitive planning performance. NeuroImage. 226, 117557 (2021).
- Wilensky, R. . Planning and Understanding: A Computational Approach to Human Reasoning. , (1983).
- Grafman, J., Hendler, J. Planning and the brain. Behavioral and Brain Sciences. 14, 563-564 (1991).
- Hayes-Roth, B., Hayes-Roth, F. A cognitive model of planning. Cognitive Science. 3 (4), 275-310 (1979).
- Tremblay, M., et al. Brain activation with a maze test: An EEG coherence analysis study in healthy subjects. Neuroreport. 5 (18), 2449-2453 (1994).
- Shallice, T. Specific impairments of planning. Philosophical Transactions of the Royal Society B: Biological Sciences. 298 (1089), 199-209 (1982).
- Unterrainer, J. M., Owen, A. M. Planning and problem solving: From neuropsychology to functional neuroimaging. Journal of Physiology. 99 (4-6), 308-317 (2006).
- Domic-Siede, M., et al. La planificación cognitiva en el contexto de la evaluación neuropsicológica e investigación en neurociencia cognitiva: Una revisión sistemática. Terapia Psicológica. 40 (3), 367-395 (2022).
- Miotto, E. C., Morris, R. G. Virtual planning in patients with frontal lobe lesions. Cortex. 34 (5), 639-657 (1998).
- Burgess, P. W., Simons, J. S., Coates, L. M. A., Channon, S., Morris, R., Ward, G. The search for specific planning processes. The Cognitive Psychology of Planning. , 199-227 (2005).
- Oosterman, J. M., Wijers, M., Kessels, R. P. C. Planning or something else? Examining neuropsychological predictors of zoo map performance. Applied Neuropsychology. 20 (2), 103-109 (2013).
- Campbell, Z., et al. Utilizing virtual reality to improve the ecological validity of clinical neuropsychology: An fMRI case study elucidating the neural basis of planning by comparing the Tower of London with a three-dimensional navigation task. Applied Neuropsychology. 16 (4), 295-306 (2009).
- Wilson, B. A., Alderman, N., Burgess, P. W., Emslie, H., Evans, J. J. . Behavioural Assessment of the Dysexecutive Syndrome. , (1996).
- Spector, L., Grafman, J., Boller, F., Grafman, J. Planning, neuropsychology, and artificial intelligence: Cross fertilization. Handbook of Neuropsychology. 9, 377-392 (1994).
- Porteus, S. D. . The Maze Test and Clinical Psychology. , (1959).
- Lis, S., Krieger, S., Wilhelm, J., Gallhofer, B. Feedback about previous action improves executive functioning in schizophrenia: An analysis of maze solving behaviour. Schizophrenia Research. 78 (2-3), 243-250 (2005).
- Zelinsky, G. J. A theory of eye movements during target acquisition. Psychological Review. 115 (4), 787-835 (2008).
- Wurtz, R. H., Goldberg, M. E., Robinson, D. L. Brain mechanisms of visual attention. Scientific American. 246 (6), 124-135 (1982).
- Burr, D., Morrone, M. C. Eye movements: Building a stable world from glance to glance. Current Biology. 15 (20), 839-840 (2005).
- Rolfs, M. Microsaccades: Small steps on a long way. Vision Research. 49 (20), 2415-2441 (2009).
- Ortega, J., Plaska, C. R., Gomes, B. A., Ellmore, T. M. Spontaneous eye blink rate during the working memory delay period predicts task accuracy. Frontiers in Psychology. 13, 788231 (2022).
- Grueschow, M., Kleim, B., Ruff, C. C. Role of the locus coeruleus arousal system in cognitive control. Journal of Neuroendocrinology. 32 (12), 12890 (2020).
- Aston-Jones, G., Shaver, R., Dinan, T. Cortically projecting nucleus basalis neurons in rat are physiologically heterogeneous. Neuroscience Letters. 46 (1), 19-24 (1984).
- Foote, S. L., Bloom, F. E., Aston-Jones, G. Nucleus locus coeruleus: New evidence of anatomical and physiological specificity. Physiological Reviews. 63 (3), 844-914 (1983).
- Siever, L. J., Davis, K. L. Overview: Toward a dysregulation hypothesis of depression. The American Journal of Psychiatry. 142 (9), 1017-1031 (1985).
- Aston-Jones, G., Gold, J. I. How we say no: Norepinephrine, inferior frontal gyrus, and response inhibition. Biological Psychiatry. 65 (7), 548-549 (2009).
- Aston-Jones, G., Cohen, J. D. An integrative theory of locus coeruleus norepinephrine function: adaptive gain and optimal performance. Annual Review of Neurosciences. 28, 403-450 (2005).
- Einhauser, W., Stout, J., Koch, C., Carter, O. Pupil dilation reflects perceptual selection and predicts subsequent stability in perceptual rivalry. Proceedings of the National Academy of Sciences of the United States of America. 105 (5), 1704-1709 (2008).
- Yoshitomi, T., Ito, Y., Inomata, H. Adrenergic excitatory and cholinergic inhibitory innervations in the human iris dilator. Experimental Eye Research. 40 (3), 453-459 (1985).
- Rajkowski, J., Kubiak, P., Aston-Jones, G. Locus coeruleus activity in monkey: Phasic and tonic changes are associated with altered vigilance. Brain Research Bulletin. 35 (5-6), 607-616 (1994).
- Beatty, J. Task-evoked pupillary responses, processing load, and the structure of processing resources. Psychological Bulletin. 91 (2), 276-292 (1982).
- Beatty, J., Kahneman, D. Pupillary changes in two memory tasks. Psychonomic Science. 5, 371-372 (1966).
- Hess, E. H., Polt, J. M. Pupil size in relation to mental activity during simple problem-solving. Science. 143 (3611), 1190-1192 (1964).
- Kuchinke, L., Vo, M. L., Hofmann, M., Jacobs, A. M. Pupillary responses during lexical decisions vary with word frequency but not emotional valence. International Journal of Psychophysiology. 65 (2), 132-140 (2007).
- Barkley, R. A. Adolescents with attention-deficit/hyperactivity disorder: An overview of empirically based treatments. Journal of Psychiatric Practice. 10 (1), 39-56 (2004).
- Gau, S. S. F. e. m. 8. 9., Shang, C. Y. Executive functions as endophenotypes in ADHD: Evidence from the Cambridge Neuropsychological Test Battery (CANTAB). Journal of Child Psychology and Psychiatry and Allied Disciplines. 51 (7), 838-849 (2010).
- Bora, E., Harrison, B. J., Yücel, M., Pantelis, C. Cognitive impairment in euthymic major depressive disorder: A meta-analysis. Psychological Medicine. 43 (10), 2017-2026 (2013).
- Rive, M. M., Koeter, M. W. J., Veltman, D. J., Schene, A. H., Ruhé, H. G. Visuospatial planning in unmedicated major depressive disorder and bipolar disorder: Distinct and common neural correlates. Psychological Medicine. 46 (11), 2313-2328 (2016).
- Holt, D. V., Wolf, J., Funke, J., Weisbrod, M., Kaiser, S. Planning impairments in schizophrenia: Specificity, task independence and functional relevance. Schizophrenia Research. 149 (1-13), 174-179 (2013).
- Lima-Silva, T. B., et al. Functional profile of patients with behavioral variant frontotemporal dementia (bvFTD) compared to patients with Alzheimer's disease and normal controls. Dementia and Neuropsychologia. 7 (1), 96-103 (2013).
- Karnath, H. O., Wallesch, C. W., Zimmermann, P. Mental planning and anticipatory processes with acute and chronic frontal lobe lesions: A comparison of maze performance in routine and non-routine situations. Neuropsychologia. 29 (4), 271-290 (1991).
- Staudigl, T., Hartl, E., Noachtar, S., Doeller, C. F., Jensen, O. Saccades are phase-locked to alpha oscillations in the occipital and medial temporal lobe during successful memory encoding. PLoS Biology. 15 (12), 2003404 (2017).
- Faul, F., Erdfelder, E., Lang, A. G., Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behavior Research Methods. 39 (2), 175-191 (2007).
- Ferrando, L., Bobes, J., Gibert, J., Soto, M. MINI Entrevista Neuropsiquiátrica Internacional (MINI International Neuropsychiatric Interview, MINI). Instrumentos de Detección y Orientación Diagnóstica. , (2000).
- McAssey, M., Dowsett, J., Kirsch, V., Brandt, T., Dieterich, M. Different EEG brain activity in right and left-handers during visually induced self-motion perception. Journal of Neurology. 267 (1), 79-90 (2020).
- Kelly, R., Mizelle, J. C., Wheaton, L. A. Distinctive laterality of neural networks supporting action understanding in left- and right-handed individuals: An EEG coherence study. Neuropsychologia. 75, 20-29 (2015).
- O'Hare, A. J., Atchley, R. A., Young, K. M. Central and divided visual field presentation of emotional images to measure hemispheric differences in motivated attention. Journal of Visualized Experiments. (129), 56257 (2017).
- Beatty, J. Task-evoked pupillary responses, processing load, and the structure of processing resources. Psychological Bulletin. 91 (2), 276-292 (1982).
- Wainstein, G., et al. Pupil size tracks attentional performance in attention-deficit/hyperactivity disorder. Scientific Reports. 7 (1), 8228 (2017).
- Drueke, B., et al. Neural correlates of positive and negative performance feedback in younger and older adults. Behavioral and Brain Functions. 11, 17 (2015).
- Lezak, M. D. The problem of assessing executive functions. International Journal of Psychology. 17 (1-4), 281-297 (1982).
- Oosterman, J. M., Wijers, M., Kessels, R. P. C. Planning or something else? Examining neuropsychological predictors of zoo map performance. Applied Neuropsychology. 20 (2), 103-109 (2013).
- Keil, A., et al. Committee report: Publication guidelines and recommendations for studies using electroencephalography and magnetoencephalography. Psychophysiology. 51 (1), 1-21 (2014).
- Abokyi, S., Owusu-Mensah, J., Osei, K. A. Caffeine intake is associated with pupil dilation and enhanced accommodation. Eye. 31 (4), 615-619 (2017).
- Wilhelm, B., Stuiber, G., Lüdtke, H., Wilhelm, H. The effect of caffeine on spontaneous pupillary oscillations. Ophthalmic & Physiological Optics. 34 (1), 73-81 (2014).
- McGinley, M. J., David, S. V., McCormick, D. A. Cortical membrane potential signature of optimal states for sensory signal detection. Neuron. 87 (1), 179-192 (2015).
- Slotnick, S. D. High density event-related potential data acquisition in cognitive neuroscience. Journal of Visualized Experiments. (38), 1945 (2010).
- Lamm, C., Windischberger, C., Leodolter, U., Moser, E., Bauer, H. Co-registration of EEG and MRI data using matching of spline interpolated and MRI-segmented reconstructions of the scalp surface. Brain Topography. 14 (2), 93-100 (2001).
- Schwartz, D., et al. Registration of MEG/EEG data with 3D MRI: Methodology and precision issues. Brain Topography. 9, 101-116 (1996).
- Laeng, B., Sirois, S., Gredebäck, G. Pupillometry: A window to the preconscious? Perspectives on psychological science. Journal of the Association for Psychological Science. 7 (1), 18-27 (2012).
- Vandierendonck, A. A comparison of methods to combine speed and accuracy measures of performance: A rejoinder on the binning procedure. Behavior Research Methods. 49 (2), 653-673 (2017).
- Vandierendonck, A. Further tests of the utility of integrated speed-accuracy measures in task switching. Journal of Cognition. 1 (1), 8 (2018).
- Levene, H., Olkin, I. Robust tests for equality of variances. In Contributions to Probability and Statistics: Essays in Honor of Harold Hotelling. , 278-292 (1960).
- Zimmerman, D. W. A note on preliminary tests of equality of variances. The British Journal of Mathematical and Statistical Psychology. 57, 173-181 (2004).
- D'Agostino, R. B., Belanger, A., D'Agostino, R. B. A suggestion for using powerful and informative tests of normality). The American Statistician. 44 (4), 316 (1990).
- Shapiro, S. S., Wilk, M. B. An analysis of variance test for normality (complete samples). Biometrika. 52 (3-4), 591-611 (1965).
- Wilcoxon, F. Individual comparisons by ranking methods. Biometrics Bulletin. 1 (6), 80 (1945).
- Fay, M. P., Proschan, M. A. Wilcoxon-Mann-Whitney or t-test? On assumptions for hypothesis tests and multiple interpretations of decision rules. Statistics Surveys. 4, 1-39 (2010).
- Delorme, A., Makeig, S. EEGLAB: An open-source toolbox for analysis of single-trial EEG dynamics including independent component analysis. Journal of Neuroscience Methods. 134 (1), 9-21 (2004).
- Dimigen, O., Sommer, W., Hohlfeld, A., Jacobs, A. M., Kliegl, R. Coregistration of eye movements and EEG in natural reading: Analyses and review. Journal of Experimental Psychology. 140 (4), 552-572 (2011).
- Mognon, A., Jovicich, J., Bruzzone, L., Buiatti, M. ADJUST: An automatic EEG artifact detector based on the joint use of spatial and temporal features. Psychophysiology. 48 (2), 229-240 (2011).
- Bell, A. J., Sejnowski, T. J., Leen, T. K., Tesauro, G., Touretsky, D. S. A non-linear information maximization algorithm that performs blind separation. Advances in Neural Information Processing Systems 7. , (1995).
- Plöchl, M., Ossandón, J. P., König, P. Combining EEG and eye tracking: Identification, characterization, and correction of eye movement artifacts in electroencephalographic data. Frontiers in Human Neuroscience. 6, 278 (2012).
- Pion-Tonachini, L., Kreutz-Delgado, K., Makeig, S. ICLabel: An automated electroencephalographic independent component classifier, dataset, and website. NeuroImage. 198, 181-197 (2019).
- Gartner, M., Grimm, S., Bajbouj, M. Frontal midline theta oscillations during mental arithmetic: Effects of stress. Frontiers in Behavioral Neuroscience. 9, 96 (2015).
- Wang, W., Viswanathan, S., Lee, T., Grafton, S. T. Coupling between theta oscillations and cognitive control network during cross-modal visual and auditory attention: Supramodal vs modality-specific mechanisms. PLoS One. 11 (7), 0158465 (2016).
- Maris, E., Oostenveld, R. Nonparametric statistical testing of EEG- and MEG-data. Journal of Neuroscience Methods. 164 (1), 177-190 (2007).
- Tadel, F., Baillet, S., Mosher, J. C., Pantazis, D., Leahy, R. M. Brainstorm: A user-friendly application for MEG/EEG analysis. Computational Intelligence and Neuroscience. 2011, 879716 (2011).
- Pascual-Marqui, R. D. Standardized low-resolution brain electromagnetic tomography (sLORETA): Technical details. Methods and Findings in Experimental and Clinical Pharmacology. 24, 5-12 (2002).
- Gramfort, A., Papadopoulo, T., Olivi, E., Clerc, M. OpenMEEG: Opensource software for quasistatic bioelectromagnetics). Biomedical Engineering Online. 9, 45 (2010).
- Destrieux, C., Fischl, B., Dale, A., Halgren, E. Automatic parcellation of human cortical gyri and sulci using standard anatomical nomenclature. Neuroimage. 53 (1), 1-15 (2010).
- Orr, J. M., Weissman, D. H. Anterior cingulate cortex makes 2 contributions to minimizing distraction. Cerebral Cortex. 9 (3), 703-711 (2009).
- Christoff, K., Gabrieli, J. D. E. The frontopolar cortex and human cognition: Evidence for a rostrocaudal hierarchical organization within the human prefrontal cortex. Psychobiology. 28 (2), 168-186 (2000).
- Nitschke, K., Köstering, L., Finkel, L., Weiller, C., Kaller, C. P. A meta-analysis on the neural basis of planning: Activation likelihood estimation of functional brain imaging results in the Tower of London task. Human Brain Mapping. 38 (1), 396-413 (2017).
- Barbey, A. K., Koenigs, M., Grafman, J. Dorsolateral prefrontal contributions to human working memory. Cortex. 49 (5), 1195-1205 (2013).
- Le Van Quyen, M., et al. Comparison of Hilbert transform and wavelet methods for the analysis of neuronal synchrony. Journal of Neuroscience Methods. 111 (2), 83-98 (2001).
- Bokil, H., Andrews, P., Kulkarni, J. E., Mehta, S., Mitra, P. P. Chronux: A platform for analyzing neural signals. Journal of Neuroscience Methods. 192 (1), 146-151 (2010).
- Sato, N., Yamaguchi, Y. EEG theta regulates eye saccade generation during human object-place memory encoding. Advances in Cognitive Neurodynamics ICCN 2007. , 429-434 (2008).
- Zaki, J., Ochsner, K. The need for a cognitive neuroscience of naturalistic social cognition. Annals of the New York Academy of Sciences. 1167, 16-30 (2009).
- Shamay-Tsoory, S. G., Mendelsohn, A. Real-life neuroscience: An ecological approach to brain and behavior research. Perspectives on Psychological Science. 14 (5), 841-859 (2019).
- Diamond, A. Executive functions. Annual Review of Psychology. 64, 135-168 (2013).
- Dudai, Y. . Memory from A to Z: Keywords, Concepts and Beyond. , (2002).
- Kingstone, A., Smilek, D., Ristic, J., Friesen, C. K., Eastwood, J. D. Attention, researchers! It is time to take a look at the real world. Current Directions in Psychological Science. 12 (5), 176-180 (2003).
- Morris, R. G., Ward, G., Morris, R., Ward, G. Introduction to the psychology of planning. The Cognitive Psychology of Planning. , (2005).
- Zwosta, K., Ruge, H., Wolfensteller, U. Neural mechanisms of goal-directed behavior: Outcome-based response selection is associated with increased functional coupling of the angular gyrus. Frontiers in Human Neuroscience. 9, 180 (2015).
- Owen, A. M., Doyon, J., Petrides, M., Evans, A. C. Planning and spatial working memory: A positron emission tomography study in humans. European Journal of Neuroscience. 8 (2), 353-364 (1996).
- Ossandon, T., et al. Efficient "pop-out" visual search elicits sustained broadband gamma activity in the dorsal attention network. Journal of Neuroscience. 32 (10), 3414-3421 (2012).
- Lezak, M. D. . Neuropsychological Assessment., 3rd edition. , (1995).
- Lin, H., Saunders, B., Hutcherson, C. A., Inzlicht, M. Midfrontal theta and pupil dilation parametrically track subjective conflict (but also surprise) during intertemporal choice. NeuroImage. 172, 838-852 (2018).
- Yu, S., Mückschel, M., Beste, C. Superior frontal regions reflect the dynamics of task engagement and theta band-related control processes in time-on task effects. Scientific Reports. 12 (1), 846 (2022).
- Nakatani, H., van Leeuwen, C. Antecedent occipital alpha band activity predicts the impact of oculomotor events in perceptual switching. Frontiers in Systems Neuroscience. 7, 19 (2013).
- Velasques, B., et al. Changes in saccadic eye movement (SEM) and quantitative EEG parameter in bipolar patients. Journal of Affective Disorders. 145 (3), 378-385 (2013).
- Bodala, I. P., Li, J., Thakor, N. V., Al-Nashash, H. EEG and eye tracking demonstrate vigilance enhancement with challenge integration. Frontiers in Human Neuroscience. 10, 273 (2016).
- Dimigen, O. Optimizing the ICA-based removal of ocular EEG artifacts from free viewing experiments. NeuroImage. 207, 116117 (2020).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoExplore Mais Artigos
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados