
Method Article
אפיון פיזיולוגי של הולוביונט האלמוגים באמצעות כלי מיקרו-רספירומטריה חדש
In This Article
Summary
פרוטוקול זה מתאר את ההתקנה וההרצה של מערכת מיקרו-רספירומטריה שניתן להשתמש בה כדי לחקור את התכונות הפיזיולוגיות של הולוביונט האלמוגים.
Abstract
פעילות מטבולית, המוגדרת כסכום התהליכים האורגניזמים המערבים אנרגיה, היא בעלת חשיבות מכרעת בהבנת תפקודם והתפתחותם של החיים על פני כדור הארץ. מדידת קצב חילוף החומרים של אורגניזמים עומדת אפוא במרכז הסבר המצבים הפיזיולוגיים של אורגניזמים, תפקידיהם האקולוגיים והשפעת השינויים הסביבתיים על מינים במערכות אקולוגיות יבשתיות ומיימיות. בשוניות אלמוגים נעשה שימוש במדדי חילוף חומרים כדי לכמת את תפקוד הסימביוזה בין אלמוגים לבין סימביוטות האצות המחייבות שלהם (Symbiodiniaceae), וכן להעריך כיצד גורמי עקה סביבתיים, כולל שינויי אקלים, ישפיעו על בריאות האלמוגים. למרות משמעות זו, חסרות שיטות, ולכן נתונים, הנוגעים למדידות קצב חילוף החומרים בצאצאי אלמוגים, ככל הנראה בשל גודלם הקטן. כדי להתמודד עם פער זה, מחקר זה נועד לפתח מערך מותאם אישית למדידת הנשימה של אקולוגיות קטנות (טווח גודל מילימטרי) של בעלי חיים ימיים. עלות נמוכה זו והתקנה קלה זו אמורה לאפשר מדידה משופרת של קצב חילוף החומרים. זה יהיה חיוני למחקר אקולוגי יישומי המשתמש בייצור מיני של אלמוגים לשיקום שוניות.
Introduction
נשימה היא מדידה ביולוגית קריטית המסמנת את הפעילות המטבולית הכוללת של אורגניזם, אך כמו תכונות קריטיות אחרות (גדילה), קשה למדוד אותה באורגניזמים קטנים1. נשימה יכולה להיות מוגדרת כחמצון של מולקולות אורגניות באמצעות שימוש בחמצן. תהליך זה מייצר את האנרגיה הכימית הדרושה לתפקוד התא (כלומר, חילוף חומרים), החיונית להישרדותם של אורגניזמים. לחלופין, חילוף חומרים אנאירובי גורם לחוב חמצן2. ניתן לקבוע את קצב הנשימה באמצעות אופטודות המודדות את השימוש (ולכן את הירידה) בריכוז החמצן לאורך זמן בתא סגור, נוהג הידוע בדרך כלל בשם רספירומטריה3. בהתחשב בכך שרוב האורגניזמים אינם מאחסנים חמצן, ניתן להסיק את קצב חילוף החומרים באמצעות הקשר הישיר בין נשימה לשימוש בפחמן. מסיבה זו, ניתן להמיר את קצב הנשימה לשימוש יומיומי בפחמן, אשר מודיע על תפקודים מטבוליים קריטיים כמו גדילה, רבייה והיכולת לשמור על הומאוסטזיס מטבולי בתקופות של לחץ סביבתי 4,5, כולל תנאי גלי חום שבדרך כלל מובילים לעקה או להלבנה באלמוגים.
שוניות האלמוגים יורדות בעולם בקצב מואץ. חיית האלמוגים מאכלסת קונסורציום של שותפים (כולל dinoflagellate Symbiodiniaceae, פטריות, חיידקים ווירוסים), המכונים ביחד "holobiont"6. ככל שטמפרטורת האוקיינוס עולה, אלמוגים, ולכן שוניות אלמוגים, נמצאים תחת לחץ גובר לשרוד, שכן טמפרטורות גבוהות מובילות לאובדן של ה-dinoflagellate Symbiodiniaceae (להלן סימביוטים), תופעה הידועה בשם הלבנה7. חומרי מזון רבים אינם זמינים לאלמוגים במים טרופיים אוליגוטרופיים, כולל חנקן אנאורגני וזרחן8. כדי להתמודד, אלמוגים יוצרים סימביוזה תזונתית מחייבת עם הסימביונטים של דינופלגלאט (Symbiodiniaceae), המספקים את רוב חומרי המזון הדרושים למארח האלמוגים כדי לשרוד ולהפקיד את שלדי הסידן הפחמתי שלהם9. סימביוזה מתפקדת עשויה להתאפיין ברמות גבוהות של חלוקת פחמן בין בני זוג10,11, והסדרת הסימביוזה כרוכה בהומאוסטזיס דינמי12.
במהלך עקת חום, הוויסות הדינמי והתקשורת משתבשים, וכתוצאה מכך דיסביוזה והלבנה (נסקר בהתייחסות13). מדידות מטבוליות, כגון פוטוסינתזה ונשימה, טומנות בחובן פוטנציאל להבהיר הן את המצב הדיסביוטי הבריא והן את המצב הדיסביוטי הבלתי מווסת של אלמוגים, ומדידה מדויקת של תהליכים אלה על פני אונטוגניות היא קריטית להבנת תפקוד האורגניזם. זה חשוב במיוחד ככל שהתדירות והעוצמה של אירועי הלבנה המוניים גדלים, עם פוטנציאל להשפיע על שינויים בשיתוף חומרי מזון מהסימביוטים, שם נמצא כי העברת הפחמן יורדת ככל שהטמפרטורות עולות14. זה יכול לנבוע ממנגנונים מכוונים על ידי חומרי המזון הסימביוטיים או מפשרות פיזיולוגיות קשות (סבילות תרמית מוגברת אך ירידה בהישרדות הפונדקאי 15,16,17). שיבושים בסימביוזה עשויים לנבוע הן מהסימביונט והן מהפונדקאי, אם כי גורם מוביל הוא ככל הנראה התקלה התאית של סימביונט18. עם זאת, מתח הנגרם על ידי עלייה בטמפרטורות מי הים מערער את הסימביוזה הזו; חלוקת הפחמן מסימביונט לפונדקאי מצטמצמתב-19,20, ורעב של האלמוגים עלול להיווצר. זה עשוי לבוא לידי ביטוי בירידה במאגרי שומנים ופחמימות באלמוגים עקב שימוש מוגבר בפונדקאי ("קטבוליזם מוגבר של פחמן קבוע"), ככל הנראה עקב שיתוף מופחת על ידי סימביוטים11. לצד תרומת הפוטוסינתזה והנשימה של הסימביונטים של האלמוגים, נשימת חיית האלמוגים היא מדד חשוב להבנת בריאות האלמוגים, השפעות ההלבנה וחילופי החומרים המזינים בין בני זוג אלה, וצמיחת ההולוביונט, פנוטיפ הרלוונטי להישרדות שינוי סביבתי 8,21,22. לבסוף, בהתחשב בכך שאלמוגים רבים הם סימביוטיים, השימוש ברספירומטריה כדי לאפיין פוטוסינתזה בנוסף לנשימה שימושי במיוחד להקשר יחסי P:R ולהבנה אם הסימביוזה יציבה או לא (למשל, הפניה23).
שינויים סביבתיים, אם כן, גורמים לשינויים בתקציבי האנרגיה של האלמוג והסימביוטים שלו, מה שמוביל להבדלים בצמיחה14. כדי להתמודד, פונדקאי האלמוגים עשוי להגביר את הנשימה ואת השימוש בשומנים כדי לענות על הדרישות המטבוליות שלו; לחץ חום יכול להפחית את התפוקה נטו ב -60% עקב נשימה מוגברת זו14, כפי שנמדד על ידי שינוי בחמצן מומס. Symbiodiniaceae עשוי גם להגביר את הטמעת החנקן ואת שימור הפחמן14,24, ולאחר מכן להשתמש במאגרים אלה כדי להעביר אנרגיה לעבר מנגנוני התיקון וההגנה שלהם25,26. האיזון של N ו-C חשוב לוויסות הצמיחה, ו-P בפרט27, מה שעשוי להתבטא בוויסות דינמי של שפע סימביונטי. ואכן, עדויות שנאספו מאלמוגים על פני מרחבי שוניות גדולות (>1,000 ק"מ) מצביעות על כך שלפונדקאים יש את היכולת להגביל את הצמיחה הסימביונטית באמצעות ויסות זרחן, אם כי זה משתנה לפי מיני אלמוגים27.
יחד, מחקרים אלה מצביעים על עלייה של סובלנות לחום עם ירידה במקביל בייצור או טרנסלוקציה של חומרים מזינים (כלומר, הנטייה לסימביוזה) עקב שינויים סביבתיים. לפיכך, יש להשתמש בשיטות חד-נעורים רבות-עוצמה, כגון כימות השימוש בחמצן באמצעות מיקרו-רספירומטריה, כדי להבין את המנגנונים הבסיסיים הקשורים לחילוף חומרים, ולאחר מכן ליישם אותן בשאלות שימור כגון הבנת רכישת סבילות לחום. זה מוצג כאן ככלי מיקרו-רספירומטריה למדדים פיזיולוגיים, שנועד לחקור את הקשר התזונתי בין אלמוגים צעירים לסימביונטות האצות שלהם, אך מתאים לאורגניזמים ימיים קטנים אחרים.
ניתן למדוד את השימוש או הייצור של חמצן על ידי אורגניזמים על ידי הצבתו בתאי ספירומטריה בודדים אטומים הרמטית או "רספירומטרים" (להלן תאים), שבהם שינוי החמצן נמדד באמצעות אופטודות3. אופטודות הן גשושיות המודדות את ריכוז החמצן באמצעות פולסי אור, ורישום מדידות לאורך זמן מאפשר חישוב קצב נשימה ו/או פוטוסינתזה. בפועל, מדידת נשימה דומה למדידת פוטוסינתזה באלמוגים, אלא שהאלמוגים מודגרים בחושך מוחלט. הפחתת סך הנשימה היומית של האלמוגים והסימביוטים מסך הפוטוסינתזה היומית מביאה להפרש חמצן (דלתא חמצן)2,3. באופן כללי, אורגניזמים משתמשים ביותר חמצן ממה שהם מייצרים, וכתוצאה מכך נוצר גירעון. זה יכול להיות מומר שווה ערך פחמן מאז חמצן ופחמן נצרכים ביחס קבוע2. עודפי הפחמן יכולים לשמש את האלמוגים לצמיחה, סינתזת ריר ורבייה, וצרכים מטבוליים חיוניים אחרים12.
פרוטוקול זה מתאר שיטת מיקרו-נשימה (איור 1) ששימשה למדידת קצבי נשימה (R) עבור אלמוגים צעירים בודדים באמצעות עיצוב תא זכוכית מותאם אישית של 1.5 מ"ל (בקבוקון עם חוט GL25 וגובה 20 מ"מ, עם בליטה/רכס, שפת קרקע שטוחה ומכסה בורג עם חור; ראו טבלת חומרים) מלא במי ים מסוננים של 0.5 מיקרומטר. אופטודות סיבים אופטיים (ראו טבלת חומרים) הוכנסו לכל תא דרך חור בצד המכסה. כל אלמוג בודד היה מחובר מעל רשת קשה, פלטפורמת צלחת ערבוב זורם מעל מוט ערבוב מגנטי כדי להבטיח ערבוב נאות של מים בתוך התא. בדוגמה המייצגת כאן, שני פקדים או "ריקים" (תאים שהיו זהים למעט נוכחות הדגימה) נמדדו במקביל לשלושת תאי הדגימה המשוכפלים, מכיוון שהיו לנו מספר בקרים שפעלו בו זמנית. עם זאת, דוגמת ההגדרה (איור 2) מציגה רק את השימוש בארבעה ערוצים; ניתן להגדיל זאת באמצעות בקרים מרובים ועמדות זרימה מרובות. ניתן לשלוט בטמפרטורה גם במערכת זו על ידי טבילת כל תא באמבט מים מותאם אישית עם טמפרטורות מים קבועות מראש (27 ° C לשליטה או 31 ° C ללחץ הטמפרטורה הגבוה בנתונים לדוגמה כאן) באמצעות מערכת זרימה מחזורית (זרימה רציפה ועדינה שנקבעה על 75 L / h). פלטפורמת צלחת הבוחש וצלחת הבוחש עם גלגלי השיניים יכולות להיות בכל גודל וניתן להכין אותן גדולות או קטנות ככל שנדרש כדי להכיל את מספר תאי הזכוכית. בדוגמה זו, המשטח והצלחת היו בערך 34 ס"מ x 26 ס"מ x 3 ס"מ (טבלה של חומרים). כיול האופטודות בוצע לפני כל ריצה באמצעות שתי תמיסות סטנדרטיות המייצגות ריווי חמצן של 0% ו-100% בטמפרטורת המים והמליחות המתאימה לסביבת ניסוי זו.
Protocol
1. התקנת ציוד ואלמוגים בתוך תאי ההנשמה
הערה: אלמוגים מוכנים לרבייה (כלומר, אלה שהיו להם ביציות/צרורות זרע ורדרדים עם פיגמנטציה הנראים מענפים מקוטעים ממושבות המין Acropora tenuis ) נעקרו מהשונית באי המגנטי (19° 6.249'S; 146° 51.728'E) ביום ירח מלא באוקטובר 2018 (מספר היתר: G12/35236.1), נאספו, והובאו למעבדה להשרצה של אלמוגים, שם גידלו וגדלו אלמוגים צעירים.
- ביום המדידות, חברו את שתי פלטות אמבט המים באמצעות פוליצינור כחול ומחברים (ראו טבלת חומרים; איור 1 [5], איור 2A,B). אלה ישמשו כאינקובטורים לאחר חיבור הפוליצינור הכחול לדוד המים/צ'ילר. ודא שניתן לראות בבירור את לוחית המנוע דרך לוחות אמבט המים השקופים כאשר תאי הספירומטריה אינם במקומם.
- חברו את שני הצינורות לדוד/צ'ילר המים (ראו טבלת חומרים). הפעל את דוד השמש / צ'ילר ולאחר מכן הגדר את טמפרטורת הניסוי הרצויה (27 ° C או 31 ° C).
- חברו את בסיס אמבט המים (שלב 1.1) ללוח המנוע הבסיסי באמצעות גלגלי השיניים המגנטיים (איור 1 [6, 7] ואיור 3A), ולאחר מכן חברו את המכלול הזה למקור כוח (איור 3B). הפעל את מקור הכוח כדי להפעיל את גלגלי השיניים, שיפעילו את מוטות הבוחש בתאים.
- לווסת את זרימת המים כנדרש (לדוגמה, זרימה רציפה ועדינה המוגדרת על 75 ליטר/שעה עם ערבוב איטי ב-30 סל"ד) באמצעות ידיות מחבר השסתום (איור 3C).
- כדי להרכיב את תא הרספירומטריה, הוסיפו את החרוז המגנטי (איור 1 [1.5]) לתא הזכוכית (איור 1 [1.6]), ולאחר מכן את בסיס המעמד הפלסטי האטום (איור 1 [1.4]) לתוך תא הזכוכית (איור 4A). גודל החדר והחרוז המגנטי יהיה תלוי באורגניזם ובמערכת המחקר של עניין.
הערה: ישנם חורים בבסיס הפלסטיק המאפשרים זרימת מים וסירקולציה מתנועת החרוז המגנטי בתחתית. - הדביקו את האלמוגים באמצעות דבק אקווריום (ראו טבלת חומרים) לאזיקונית השחורה שמונחת בבסיס הפלסטיק (איור 4B-D). כדי לעשות זאת, ראשית, להדביק את האלמוג הצעיר חתיכת פלסטיק שחור, ולאחר מכן להדביק את החתיכה לבסיס הפלסטיק. ברגע שהאלמוג מודבק היטב (ריפוי הדבק הוא כמעט מיידי), הבריגו את המכסה עם טבעת O (איור 4A) על תא הזכוכית. בצע פעולות אלה מתחת למים באגן נפרד כדי לוודא שאין אוויר בתוך תא הספירומטריה.
הערה: נפח המים ברספירומטר (כלומר, נפח אפקטיבי = 1.5 מ"ל) נקבע באמצעות טבילה מלאה של התא מתחת למים. ההנחה היא שיחסית לנפח המים, התזוזה מהמסה/צפיפות של האלמוגים הקטנים מאוד היא זניחה. צינור המיקרוצנטריפוגה בנפח 1.5 מ"ל מוצג כאן בקנה מידה (איור 4A). - מקמו היטב את התאים בתוך אמבטיות המים (איור 5A). ודא כי תאי הזכוכית נמצאים במגע עם המים מבוקרי הטמפרטורה עבור הניסוי.
- חבר את כבלי הסיב האופטי O2 (ראה טבלת חומרים) כך שיהיו במגע עם נקודות חיישן החמצן (להלן כתמים; ראה טבלת חומרים) על ידי הכנסתם לחור שנקדח בצד תאי המכסה. כתמים קטנים אלה רגישים לחמצן ויזהו ויעבירו את האות מתוך התא דרך כבל הסיב האופטי.
- הוסף סרט אינסטלציה (סרט לבן דק לאיטום עצמי) כדי להתאים את הכבל בצורה נוחה ולאפשר לו להישאר יציב בתוך תא המים. ודאו שניתן לראות את האלמוגים הבודדים (כפי שניתן לראות באיור 5B), עם זרועות ציד חומות הפונות כלפי מעלה, בתוך התא (איור 5B, וידאו 1).
הערה: אומדני עלות של הרכיבים במכשיר מוצגים בטבלה 1.
- הוסף סרט אינסטלציה (סרט לבן דק לאיטום עצמי) כדי להתאים את הכבל בצורה נוחה ולאפשר לו להישאר יציב בתוך תא המים. ודאו שניתן לראות את האלמוגים הבודדים (כפי שניתן לראות באיור 5B), עם זרועות ציד חומות הפונות כלפי מעלה, בתוך התא (איור 5B, וידאו 1).
2. נוהל הפעלה סטנדרטי למדידת נשימה באמצעות מערכת O2
- פתח את התוכנה למדידת חמצן (ראה טבלת חומרים).
- למדוד את הטמפרטורה של החדר שבו הכיול יבוצע. זה יהיה צורך מאוחר יותר לשלב הכיול (שלב 2.8).
- הרכיבו את החיישנים והמכסים האופטיים. לשם כך, חבר את כל הסיב האופטי ליציאה התואמת במודול O2 . הקפד להתאים את כובע 1 לחיישן 1, מכסה 2 לחיישן 2 וכן הלאה.
- כדי להגדיר את התאים לכיול, תחילה יש להרטיב פיסת ספוג נקי עם מעט מי אוסמוזה הפוכה (RO) ולהכניס אותה לכל תא.
הערה: הספוג לא צריך לטפטף, רק להיות לח. הוא עלול לטפטף על נקודת הסיב האופטי. ודא שהכתם אינו רטוב לפני שתמשיך לשלב הבא. - הניחו את התאים הפוכים בעזרת הסיב האופטי התואם (איור 6). זה יאפשר לפתוח את הברגת התא מבלי לגעת בסיב האופטי ולהוסיף את הנתרן הגופריתי לכיול 0%.
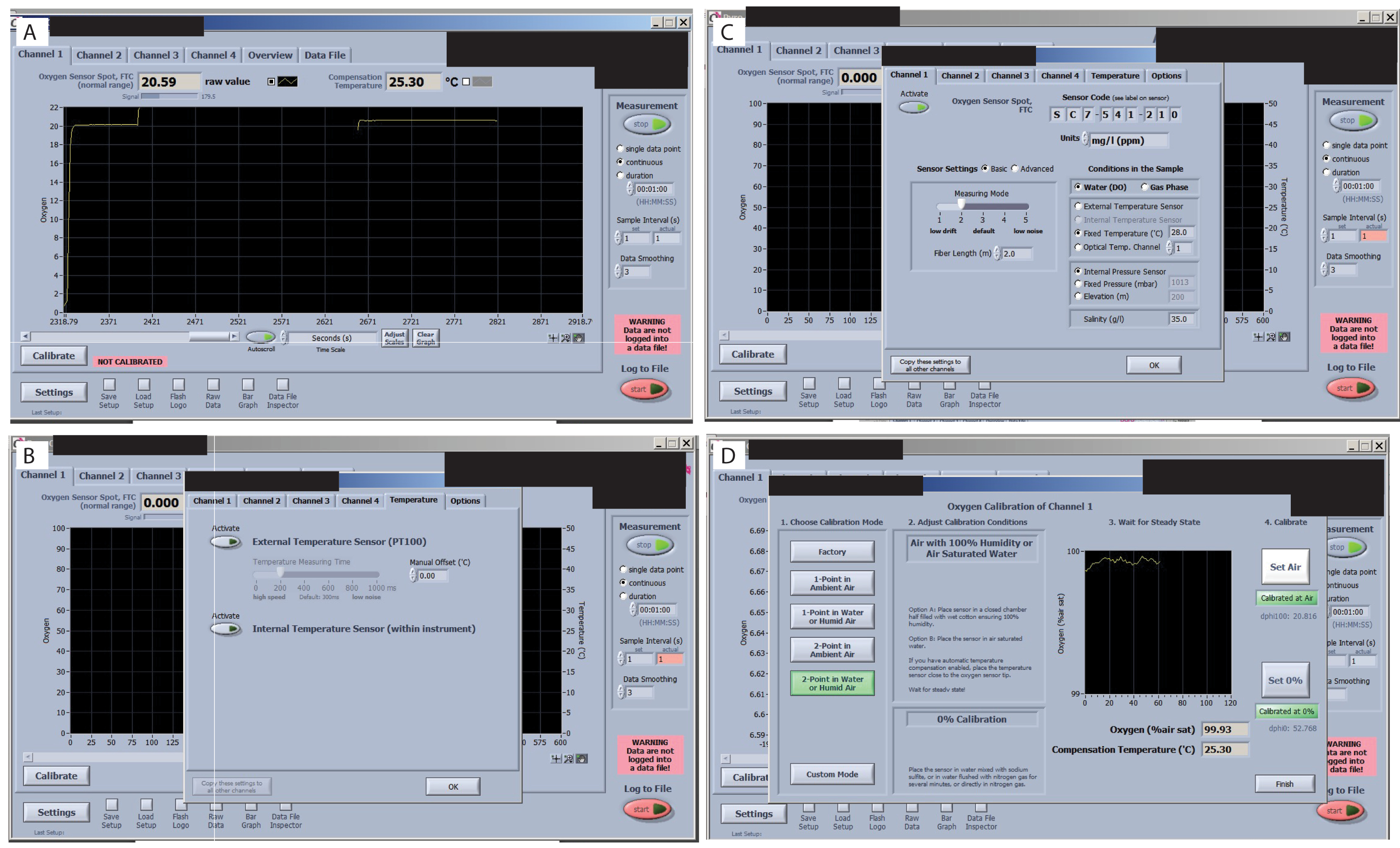
- בדוק את האות של כל חיישן לפני תחילת הכיול. עבור על כל הכרטיסיות בממשק התוכנה ובדוק את ערך האות (בפינה השמאלית העליונה) (איור 7A), כדי לוודא שהן אינן משתנות באופן משמעותי. בדוק את מדריך O2 עבור הערכים המקובלים עבור מערך הניסוי (בהתאם לאורגניזם ולתנאי העניין). עבור הגדרה ספציפית זו, בטמפרטורת חדר של 25.3 מעלות צלזיוס, FTC (זרימת נקודת חיישן חמצן דרך טווח תקין של התא) של 20.59 עם האות ב 179.5 מקובל.
- פתח את תוכנית ובדוק את ההגדרות בתוכנה כדי לוודא שהן נכונות כמתואר להלן. אם החלון המוקפץ אינו מופיע מיד לאחר פתיחת התוכנית, ניתן לעשות זאת על ידי לחיצה על הלחצן הגדרות בפינה השמאלית התחתונה של ממשק התוכנה.
- בדקו שחיישן הטמפרטורה החיצוני מופעל (איור 7B). שנה את ההגדרה לטמפרטורה קבועה (איור 7C). לאחר מכן, הוסף את ערך טמפרטורת החדר ולחץ על העתק הגדרה לכל הערוצים האחרים.
- שנה את ההגדרה להפחתת סחף אותות (איור 7C). לאחר מכן, בחר הגדרות חיישן ובחר רמה 2. אחרת, אם משתמשים בנפחים קטנים, הסחף יהיה כה גבוה שיהיה קשה לכייל אותו.
- בדוק את ההגדרות הכלליות של הערוצים. לחץ על אישור אם מתאים. לחץ על העתק הגדרות לכל הערוצים ולאחר מכן לחץ על אישור.
- כייל את החיישנים. לכיול ערוץ 1, עברו לכרטיסייה ערוץ 1 ולחצו על כפתור הכיול . בחר 2 נקודות במים או אוויר לח.
- לכיול "אוויר", טבלו חתיכת קצף במים, הניחו אותה בתוך התא והמתינו עד שהאות יתייצב (ראו תמונות כיול אוויר לפני ואחרי). כאשר יציב, לחץ על "הגדר אוויר". לחץ על air > כיול > הגדרת אוויר.
- הגדר את הכיול 0% ו- 100% (איור 7D). הסר את התא מהמכסה והנח אותו במכסה הבא כך שאות כיול האוויר יהיה מוכן עם סיום כיול ה- 0% בחיישן הראשון. השתמשו בפיפטת העברה ומלאו את הפקק ב-2% נתרן סולפיט. המתן עד שהאות יתייצב.
הערה: בדרך כלל לוקח לאות זמן רב יותר להתייצב בהשוואה לכיול האוויר. אם מופיעה הודעת אזהרה המציינת כי "הערכים נמצאים מחוץ לטווח האופייני", ודא שהנתרן הגופריתי טרי. חזור על אותו תהליך כיול עבור כל הערוצים. הכינו את נתרן סולפיט על ידי הוספת 2 גרם ב 100 מ"ל של מים RO. - לאחר השלמת הכיול, שטפו היטב את תאי הנשימה וייבשו את התאים והפקקים. ודא שאין מים בחור הסיב האופטי.
- מקמו את האורגניזם (אלמוגים בודדים צעירים, בדוגמה זו) בתוך תאי הנשימה וסגרו עם עפעפיים. בעת הנחת המכסים, הקפד לעשות זאת כאשר החדרים שקועים לחלוטין ואין שום אוויר בפנים.
- מניחים את התאים היטב בצלחת הערבוב ומחברים את הסיב האופטי.
- הפעל את ספק הכוח. ודאו שהמים בתוך התאים מתערבבים לחלוטין. כוונו את הטמפרטורה בצ'ילר/מחמם לטמפרטורות הניסוי שנבחרו.
- הפעל את המשאבה ואת תנור החימום (שלבים 1.2 ו-1.4).
- לחץ על היכנס לממשק תוכנת המדידה O2 כדי להתחיל בהקלטה.
תוצאות
עיבוד וניתוח נתונים
אמנם ישנן שיטות רבות לעבד את הנתונים הגולמיים מניסויי רספירומטריה, אך מומלץ להשתמש בחבילת R respR28. בהתאם לשיתוף הפרוטוקולים לעיל, הדוגלים במדע פתוח וביכולת שחזור, חבילה זו מאפשרת שיתוף וניתוח נתונים בצורה קלה לשכפול ותוכננה תוך התחשבות בכך. זוהי פלטפורמת קוד פתוח חינמית ואגנוסטית למערכת הגששות, וניתנת להתקנה בקלות מ- CRAN או GitHub. קוד מלא ודוגמאות עבור respR נשמרים וניתן למצוא אותם בכתובת https://github.com/januarharianto/respR.
חבילת respR כוללת פונקציות לייבוא, הדמיה וביצוע בקרת איכות על נתוני ספירומטריה, ולחישוב קצבי נשימה באופן אוטומטי או מאזורים שנבחרו ידנית. זה יכול גם להתאים את שיעורי נשימת הרקע ושיעורי ההמרה ליחידות פלט נפוצות. השלבים לעיבוד הנתונים ממערכת המיקרו-רספירומטריה מפורטים להלן. במחקר זה, הנתונים ממערכת הספירומטריה שימשו כדוגמה, אך החבילה מקבלת גם קלט מרוב מערכות בדיקת החמצן הזמינות מסחרית, כמו גם אובייקטי נתונים גנריים מסוג R. פרטים נוספים על החבילה, כולל תיעוד מלא ומדריכים, ניתן למצוא באתר החבילה בכתובת https://januarharianto.github.io/respR/index.html.
ייבוא נתונים גולמיים
קובץ הפלט הגולמי (.txt) מיובא. respR מזהה את התבנית ומנתח אותה למסגרת נתונים כללית R שניתן להשתמש בה בפונקציות הבאות. עם זאת, חשוב לציין כי זה אופציונלי; קבצים אלה וכמעט כל נתוני סדרת זמן חמצן ניתנים לייבוא באמצעות פונקציות בסיס (המפורטות להלן) על ידי כל מי שיש לו ידע בסיסי ב- R.
#load respR
ספריה(respR)
#Import ---
נתונים <- import_file("file.txt")
קובץ #Firesting-Pryo זוהה
בדיקה והצגה חזותית של הנתונים
חלק חיוני בכל משימת ניתוח נתונים הוא לתכנן ולבדוק את הנתונים כדי לחפש חריגות או דפוסים ברורים, או אפילו רק לעזור להבין אותם. פונקציית הבדיקה משמשת כאן (איור 8A), אשר בודקת בעיות נפוצות בנתוני ספירומטריה, כגון ערכים לא מספריים או חסרים.
#inspect עמודת חמצן אחת
Insp <-inspect(נתונים, זמן = 3, חמצן = 8, רוחב = 0.2)
פונקציה זו גם משרטטת את סדרות הזמן של החמצן ומחשבת קצב גלגול (לוח תחתון) כדי לעזור להבהיר כיצד קצב זה עשוי להשתנות במהלך הניסוי. חלקות קצב מתגלגלות אלה עוזרות להודיע אילו אזורים של עקומות הריבית הללו יש לחלץ. במקרה של קצב חילוף חומרים סטנדרטי או שגרתי, האזורים הרצויים הם אלה שבהם הקצב מראה יציבות (למשל, לאחר סביב נקודת זמן 3,000; איור 8B).
כאן, ירידה בחמצן ניתנת לזיהוי רק לאחר כשורה 200 בלוח הזמנים המלא. דפוסים כאלה נפוצים מאוד בנתוני רספירומטריה; לעתים קרובות יש תקופה ממושכת של חוסר יציבות בתחילת הניסוי כאשר המערכת מתייצבת והדגימה מתאקלמת לתנאי הניסוי. מומלץ לחלץ את התעריפים רק מסדרת הזמנים לאחר חוסר יציבות ראשוני זה, מה שמדגיש גם את חשיבות ההדמיות.
שיעורי מיצוי
ל-respR יש שני תפקידים לחילוץ קצבי נשימה. הראשונה היא הפונקציה calc_rate(), המאפשרת חילוץ קצב באופן ידני על ידי ציון אזור של זמן, שורה או רמת חמצן. שיטה זו נפוצה מאוד בניתוחי רספירומטריה, ושיטה מקובלת לחלוטין לקביעת שיעור כל עוד קריטריונים לבחירה נקבעים ומיושמים באופן עקבי28.
דרך חזקה ואובייקטיבית יותר היא להשתמש בפונקציה auto_rate(), המזהה אזורים ליניאריים של הנתונים. אזורים אלה הם אלה של שיעורי נשימה מתמשכים באופן עקבי, המוקצים באופן אוטומטי באמצעות למידת מכונה. פונקציה זו שימושית גם לזיהוי אותות נמוכים (כמו בדגימות המשמשות במחקר הנוכחי, בשל הביומסה הנמוכה בגיל זה). פונקציה זו מאפשרת זיהוי של השיעורים הלינאריים, המינימליים והמקסימליים ביותר באמצעות שיטות עצמאיות, אובייקטיביות וחזקות סטטיסטית28. הדוגמה כאן מזהה אזור ליניארי המתרחש בסביבות נקודות זמן 3,000 עד 5,000. יש לציין כי ניתן לזהות אזורים ליניאריים מרובים, אך קטע זה הוא האזור המדורג הגבוה ביותר, או הליניארי ביותר (איור 8C).
#Determine הקצב הליניארי ביותר (כלומר, עקבי)
דרג <-auto_rate(INSP)
התאמת רקע
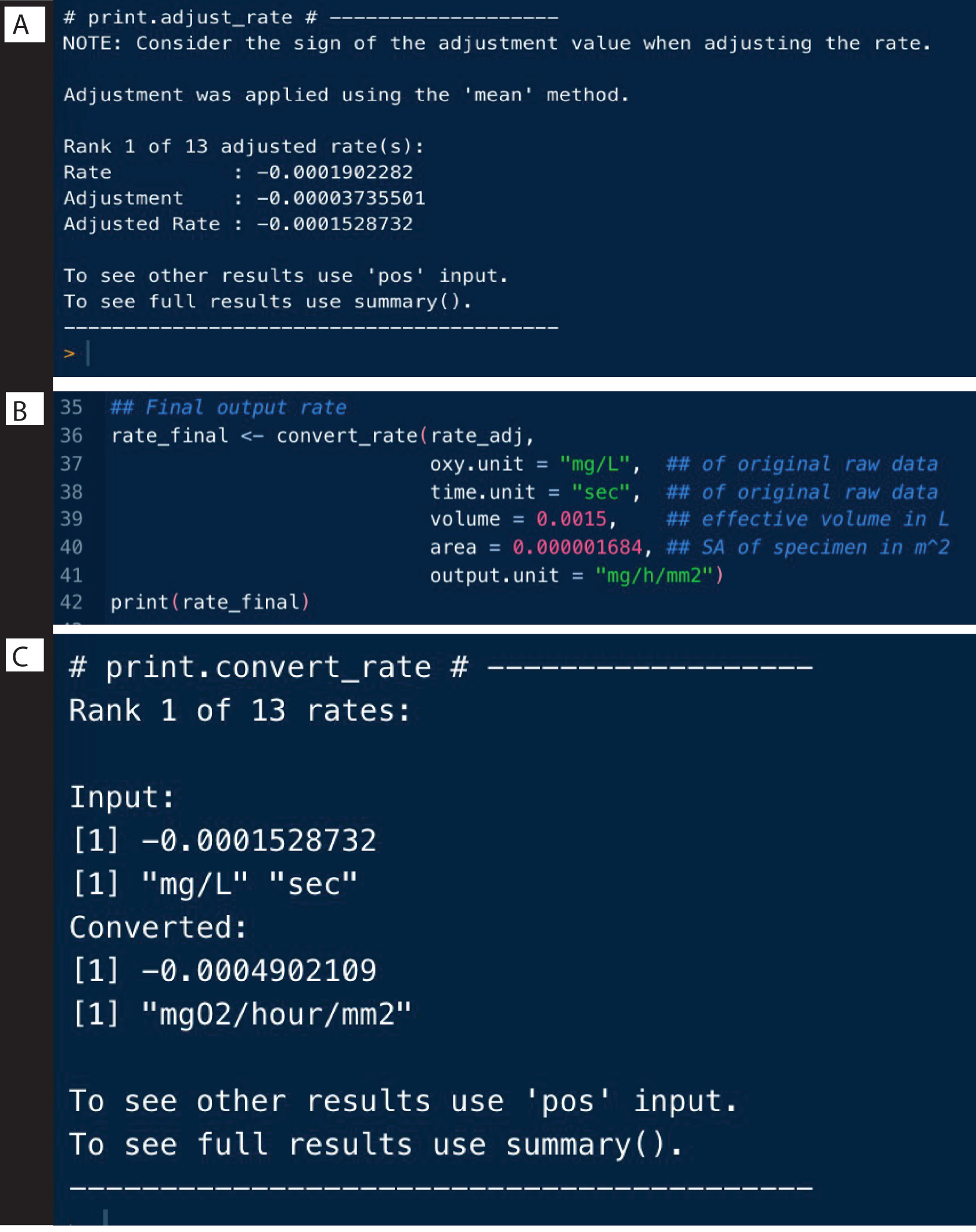
ניתן לקבוע את קצבי הרקע מניסויי בקרה באופן דומה לדוגמה לעיל, וניתן להשתמש בהם כדי להתאים את קצבי הדגימה באמצעות הפונקציה adjust_rate() (איור 9A; שימו לב שהניתוח המלא אינו מוצג כאן, אלא רק ההתאמה). דוגמאות מלאות מפורטות באתר respR .
תעריף #Adjust לרקע
rate_adj <-adjust_rate(rate, by = bg) #saved אובייקט bg
הדפס(rate_adj)
המרת תעריפים
השלב האחרון הוא להמיר את הקצב ליחידות הפלט הרצויות, תוך שימוש ביחידות המקוריות של הנתונים הגולמיים, נפח אפקטיבי של הרספירומטר ונתונים ניסיוניים אחרים, כולל נורמליזציה למדידות ריקות (איור 9B). התפוקה יכולה להיות קצב נשימה מוחלט, כלומר של הדגימה כולה, או קצב מסה או שטח פנים ספציפי. הקצב הספציפי לשטח הפנים היה הפלט ששימש כאן, שהוא באופן ספציפי הקצב האבסולוטי חלקי שטח הפנים של הדגימה (איור 9C).
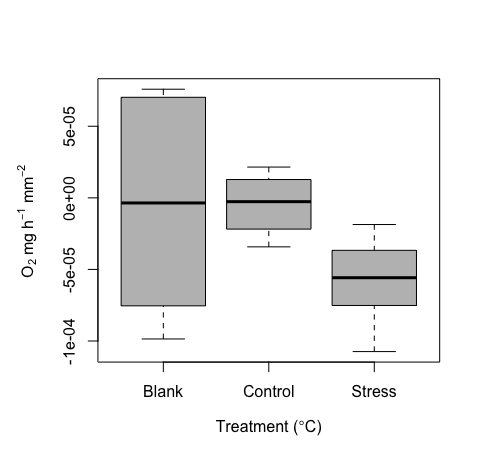
כפי שפורט לעיל, מערכת זו פותחה כדי למדוד דגימות קטנות מאוד. לכן, ציפינו לערכים נמוכים וחפיפה פוטנציאלית עם מדידות ריקות. רמה מסוימת של אות צפויה בתוך החסר, וכאשר בוחנים אותם, ערכים אלה נמצאים בטווח הצפוי של רעש ניסויי כללי, אולי בגלל סחף הגשושיות, שינויי טמפרטורה קלים או בועות על הגשושיות. על פי התכנון ובשל גודל הדגימה הקטן, ולכן נפח אפקטיבי קטן בשימוש, השימוש בחסר חשוב במיוחד כאן, במיוחד עבור כל ריצה. ערכים מייצגים נכללו כאן כדוגמה (איור 10). בהתחשב בגודל הדגימה הקטן, אנו ממליצים להשתמש בחסר בכל ריצה כדי לתקנן מדידות לכל ריצה.
ערכים ריקים אלה משמשים לאחר מכן לסטנדרטיזציה של ערכי מדידות הטיפול. בהינתן שאלמוגים נושמים בנוסף לייצור חמצן, קצב חילוף החומרים יכול לנוע בין ערכים שליליים לחיוביים. דוגמה לתוצאות מייצגות של טווח ערכי הנשימה שזוהו מכלי המיקרו-נשימה ניתנת כאן. התוצאות האלה נקבעו מניסוי מוצלח על אלמוגים צעירים בודדים (איור 10). בסך הכל, הנשימה הייתה צפויה להיות קשה לזיהוי במערך נתונים לדוגמה זה (על ידי תכנון), בהתחשב בגודל הקטן של הדגימות; זה מדגיש את הערך של שיטה זו בלכידת סף אות נמוך זה. תוצאות מייצגות אלה מראות נשימה בחושך בגדלי הדגימות הקטנים ביותר שנבדקו, ומדגישות את סף הזיהוי המינימלי של מערכת זו. מדדנו גם בשני תנאים (בקרה ולחץ טמפרטורה גבוהה). לאחר סטנדרטיזציה לחסר שנמדד בכל ריצה, הערכים נעו בין קרוב לאפס (שליטה) לחציון של ~-5e-5 לטיפול במאמץ. כצפוי, הנשימה הייתה נמוכה. תוצאות אלה מראות בבירור ערכים מייצגים עבור ערכים ריקים, כמו גם בקרת לעומת השוואת טמפרטורה גבוהה עבור דגימות קטנות מאוד אלה.

איור 1: ייצוג סכמטי של כלי המיקרו-רספירומטריה החדש לאפיון פיזיולוגי של הולוביונט אלמוגים (חיית אלמוגים + סימביוטים) או כל אורגניזם קטן (<1 מ"מ). יוצרו תאי ספירומטריה מותאמים אישית (מספר 1; 1.1-1.6). אלה כוללים מכסים (1.1) עם נקודות חיישן חמצן (1.2), והנער הבודד (1.3) מונח על מעמד זורם (1.4) המוצב על גבי מערבל מגנטי (1.5), שכולם מתאימים לתא הזכוכית (1.6). הבקר (2) מחובר למקום באמצעות כבל סיב אופטי שנכנס למכסה (1) ומחובר למחשב (3). המחמם/צ'ילר (4) מתחבר ללוחית הרספירומטריה (5) עם המים הזורמים (מסומנים על ידי חיצי שברון לכיוון), אשר יושב על גבי לוח המערבל (6) עם גלגלי שיניים (7), מופעל על ידי המנוע (8) וספק הכוח (9). אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}

איור 2: הגדרת מיקרו-רספירומטריה. קיימות אפשרויות רבות, כולל (A) לוחית רספירומטריה אחת, או (B) המחוברת למספר לוחות. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}

איור 3: לוח מערבל מגנטי שנבנה בהתאמה אישית על גבי לוח רספירומטריה. לכל תא יש (A) גלגל שיניים בוחש מגנטי מתחתיו, (B) מופעל על ידי מנוע, כאשר (C) לוח הספירומטריה מחובר באמצעות צינורות לתנור החימום/צ'ילר. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}

איור 4: הגדרת תא ספירומטריה מותאם אישית. (A) רכיבים (משמאל לימין: מכסה, בקבוקון זכוכית, מעמד, צינור 1.5 מ"ל לקנה מידה ומוט בוחש). (B) מעמד זרימה בודד שהדגימה יושבת בתוכו. (C) מבט מלמעלה למטה על מעמד הזרימה. (D) וכשהמעמד מונח בתוך בקבוקון הזכוכית כשהמכסה מוברג. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}

איור 5: בקבוקוני זכוכית המונחים בתוך צלחת הבוחש. (A) צלחת מערבל מותאמת אישית עם (B) תקריב של בקבוקון הזכוכית המלא עם הגדרת המכסה. ניתן לראות את האלמוג הצעיר דרך המכסה כאן (נקודה חומה), מעל האזיקון, כאשר הסיב האופטי ממוקם בפתח המכסה. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}

איור 6: תאים מונחים הפוך, מוכנים לכיול. לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}

איור 7: שלבים עיקריים בתוכנה למדידת חמצן. (A) בדוק את האות של כל חיישן. אות אופטימלי למחקר זה ולחיישנים מוצג בנקודת חיישן החמצן FTC (טווח נורמלי). (B) בדוק את סחף האות. (C) הגדר ובדוק את טמפרטורות הטיפול. (ד) הגדר ובדוק את כיולים 0% ו- 100%. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}

איור 8: שלבי פלט ניתוח respR I. (A) בדוק את הפקודה והפלט. (ב) בדוק את יציבות התעריף. (ג) לקבוע את הקצב הליניארי ביותר. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}

איור 9: שלבי פלט ניתוח respR II. (א) התאם את התעריף לרקע, (ב) המר ו-(ג) בדוק את התעריפים. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}

איור 10: תוצאות מייצגות שהופקו מכלי המיקרו-רספירומטריה. נשימה חציונית (O2 ± שגיאת תקן) של שכפול אלמוגים בודדים צעירים, כולל ערכים ריקים וכן נשימה של פרטים תחת בקרה ותנאי עקה בטמפרטורה גבוהה. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
{kind=link}
וידאו 1: מבט מלמעלה למטה על תא הספירומטריה עם האלמוג הצעיר בתוכו במהלך סשן מדידה. אנא לחץ כאן כדי להוריד סרטון זה.
טבלה 1: אומדני עלות של רכיבי מנגנון הרספירומטריה. אנא לחץ כאן כדי להוריד טבלה זו.
Discussion
עבודה זו מתארת את הבנייה של מערך מיקרו-רספירומטריה מותאם אישית שניתן להשתמש בו כדי לכמת את כמות החמצן הנצרכת ומיוצרת על ידי אורגניזמים ימיים קטנים sessile. המרכיבים הקריטיים של פרוטוקול זה כוללים את הגדרת החדרים, כולל הכתמים, וכיול האות הנמוך באמצעות חבילת respR , שבה אות נמוך יכול להיות מוגדר כשיעורים המאופיינים במדרונות רדודים או רועשים. התא המותאם אישית וההתקנה שלו מאפשרים זיהוי של אותות נמוכים אפילו, בעוד השימוש בחבילת R מסייע בהגנה מפני בעיות שבהן התרחשות של מדרונות רדודים או רועשים עלולה להוביל לפרשנות שגויה של התוצאות (למשל, תוצאות חיוביות שגויות).
שינויים פוטנציאליים שיידרשו עבור משתמשים אחרים כוללים את אבטחת האורגניזם של עניין בתוך החדר בהתאמה אישית. במקרה זה, נעשה שימוש באזיקון קטן ונוקשה ובדבק אקווריום כדי לאבטח את הנער הבודד לבסיס הפלסטיק, שהודבק לאחר מכן לעניבה. יש לציין כי לצורך ניסוי זה, צעירי אלמוגים התיישבו על יריעות פלסטיק שחורות. פלסטיק זה איפשר הסרה קלה של צעירי האלמוגים, שהחליקו למעשה מהפלסטיק, כדי לא לפגוע בהם פיזית במהלך ההסרה. צעירי האלמוגים נצמדים למצע עליו הם מתיישבים, ולכן מומלץ ליישב אותם על חומר פלסטי דומה, באמצעות פפטיד מלאכותי16 כדי להקל על הסרתם לצורך תהליך ההדבקה. כדי למזער עוד יותר את הטיפול בעקה ואת ההשפעה על תגובת הנשימה, מומלץ לאפשר לאלמוגים המורכבים על אזיקוני רוכסן להתאקלם במשך שבוע-שבועיים, כפי שמקובל בניסויי עקה רבים של אלמוגים בוגרים. ייתכן שיהיה צורך בשינויים אחרים כדי לאבטח את האורגניזם מעל המקום במכסה ולאפשר זרימת מים. שלב מפתח נוסף לפתרון בעיות כרוך בזיהוי אותות, במיוחד בשיפוע של סדרת הזמן של החמצן שבו יש לקבוע שיעורים. בסופו של דבר, זה מסתכם בשילוב של הפעלת שיקול דעת כדי לא לכלול נתונים בלתי יציבים בעליל, ואת הפונקציות בתוך respR כדי לאפשר שיעורי חילוץ מאזורים שנבחרו באופן עקבי או באופן אוטומטי על ידי זיהוי אזורים ליניאריים של הנתונים. דוגמאות נוספות כיצד לעשות זאת זמינות באתר האינטרנט של respR .
שיטה זו פותחה כדי להרחיב את המדידות של הגבול התחתון של הנשימה לחסרי חוליות ימיים קטנים מאוד. המגבלה הברורה היא שפרוטוקול זה עשוי להיות מועד יותר לתוצאות חיוביות שגויות בהשוואה לפרוטוקולים המיועדים לביומסה גדולה יותר. עם זאת, בהתחשב בכך שזו הייתה מטרת התכנון - למדוד את הגבולות התחתונים הללו - זה נלקח בחשבון בעיצוב, וניתן להשתמש בהליך עם חבילת respR כדי להגן טוב יותר מפני תוצאות חיוביות שגויות. חשוב גם להכיר בכך שקיימות מערכות אחרות למדידת נשימה30, ולמדידת אורגניזמים קטנים, כולל רספירומטריה על קופפודים בודדים31 בנפחים קטנים יותר מזה (~0.5-1 מ"ל), אך הן יקרות או חסרות רכיבים ספציפיים (יכולת ערבוב). עם זאת, מערכת זו היא קוד פתוח ובעלות נמוכה יחסית בהשוואה למערכות מסחריות (למשל, מערכת Core Microplate). מערכת זו משלבת גם שיקולים מתודולוגיים מרכזיים כמו ערבוב, שמערכות אחרות עשויות להיות חסרות. תכונת מוט הערבוב הפנימי חיונית כדי לשכפל את ערבוב המים הטבעי של אורגניזמים ימיים רבים (למשל, קופפודים באמצעות שחייה), שלעתים קרובות אינו אפשרי ועלול להפוך את הנתונים לבלתי שמישים במידה רבה. לעומת זאת, שיטות ערבוב אחרות הזמינות כוללות הנחת כל הרספירומטר על ספסל נדנדה ענק, הדורש ציוד נוסף ויש לו הצלחה מוגבלת בערבוב, או ערבוב באמצעות רטט, אשר יכול לגרום הפרעה לאורגניזם. מסיבה זו, זוהי המערכת היחידה שיכולה לבצע רספירומטריה על אלמוגים צעירים או אורגניזמים קטנים מאוד אחרים. לשם השוואה, טווח הגודל של הדגימות שנכללו כאן נע בין 2.1 ל -3.6 פוליפים (המקבילים לגיל חודשים ספורים בלבד), עם שטח ממוצע מינימלי עד מקסימלי של 1.3 עד 4.5 מ"מ2.
רספירומטריה היא מדד בסיסי במחקרים אקולוגיים, וקיימות שיטות רבות למטרה זו. עם זאת, רוב השיטות הקיימות הללו מכוונות לדגימות ביומסה גבוהות, כולל דגים שלמים, שברי אלמוגים או עשבי ים 32,33,34. שיטה זו היא הראשונה שהשתמשה בצעירי אלמוגים בודדים. בנוסף, ישנם יישומים פוטנציאליים רבים עבור שיטה זו, שכן הוא מספק מידע פיזיולוגי מרכזי על תפקוד האורגניזם. זה יכול להיות חשוב עבור מחקרים המעוניינים לאפיין הערכות בריאות בסיסיות35, להבין את התפקיד של לחץ אקוטי או ארוך טווח במהלך אונטוגניות אלמוגים כגון עקת חום36, או לספק ערכי סף שמנהלים יכולים להגדיר כדי לעזור להגן ולשפר את הבריאות של שוניות אלמוגים37. בהתחשב בכך שהאלמוג הוא הולוביונט וקהילת הסימביונט גמישה יחסית בשלב זה ולאורך השנה הראשונה לחיים38, יהיה מעניין לשדך נתוני ספירומטריה עם שינויים בקהילות לאורך זמן, כדי להקשר באופן מלא את תפקוד האורגניזם בכללותו. חשוב לציין, שיטה זו תורמת לטכניקות "מדע פתוח" המסייעות לספק מתווה ליצירת מערכי ניסוי מותאמים אישית שניתן לשתף, לשפר ולתקנן באופן פתוח.
Disclosures
למחברים אין מה לחשוף.
Acknowledgements
המחברים רוצים להודות לסם נונן על עזרתו ועצתו, לסוון אוטיק על השימוש בתאי הרספירומטריה הראשוניים, לבן שלאב על האיור ההנדסי שלו, ולסדנת המכון האוסטרלי למדעי הים לעיבוד שבבי בהתאמה אישית של מתאמים ומחזיקי תאי ספירומטריה. האלמוגים נאספו תחת היתר הפארק הימי שונית המחסום הגדולה הבא ל- AIMS G12/ 35236.1. אלמוגים אינם זקוקים להיתרי אתיקה.
Materials
| Name | Company | Catalog Number | Comments |
| Cost | |||
| (1.1 – 1.6) Custom respirometry chambers | LabGlass Party Ldt. | 1.5 ml | $407.26 |
| 1.1 lids | AIMS workshop | Vial GL25 thread | ~$10 |
| 1.2 fiber-optics spots (FireStingO2 II fiberoptic optodes) | PyroScience | Oxygen sensor spots, 125 µm PET foil, Ø5 mm, with optical isolation, SN: 183801947 | $41.25 AUD each |
| 1.3 individual organism | NA | NA | NA |
| 1.4 flow-through stand | AIMS workshop | Custom | included in points 5 and 6 price (the workshop gave me an estimate of the lids, stand with gears, motor, incubation flow through |
| 1.5 magnetic stirrer | Any manufactuer is suitable | NA | ~$2? |
| 1.6 glass chamber (vial GL25 thread x 20 mm high, with bump/ridge, flat-ground rim, screw cap with hole, Labglass Pty Ltd, Stafford QLD) | Labglass Pty Ltd, Stafford QLD | Vial GL25 thread x 20 mm high, with bump/ridge, flat-ground rim, screw cap with hole | $50.9 AUD |
| 2 FireSting controller (2) | PyroSciences | NA | 4 sensors is 4000 Euros. 8 sensors used here. |
| 3 computer | NA | NA | NA |
| 4 heater/chiller | VWR International | NA | Small models around $4,000 AUD |
| 5 respirometry plate platform | AIMS workshop | 34 cm x 26 cm x 3 cm (although any dimensions are adequate to fit desired number of chambers) | $1250 AUD |
| 6 stirrer plate with gears (7) | AIMS workshop | 34 cm x 26 cm x 3 cm | $1250 AUD |
| 8 powered by the motor | AIMS workshop | Custom | $700 AUD |
| 9 power supply | Non-specific | NA | ~$300 AUD |
| Aquarium glue | Seachem reef glue | 20g | $14 |
| Oxygen Logger Software | PyroScience | NA | NA |
| Polypipe and connectors | John Guest | NA | $20 |
| Sodium Sulfite | Sigma | S0505-250G (CAS number 7757-83-7) | $54 |
References
- Quigley, K. M. A fast, precise, in-vivo method for micron-level 3D models of corals using dental scanners. Methods in Ecology and Evolution. 13 (10), 2159-2166 (2022).
- Svendsen, M. B. S., Bushnell, P. G., Steffensen, J. F. Design and setup of intermittent-flow respirometry system for aquatic organisms. Journal of Fish Biology. 88 (1), 26-50 (2016).
- Lighton, J. R. B. Measuring Metabolic Rates: a Manual for Scientists. , Oxford University Press. (2018).
- Carey, N., Harianto, J., Byrne, M. Sea urchins in a high-CO2 world: partitioned effects of body size, ocean warming and acidification on metabolic rate. The Journal of Experimental Biology. 219 (Pt 8), 1178-1186 (2016).
- Clark, T. D., Sandblom, E., Jutfelt, F. Aerobic scope measurements of fishes in an era of climate change: respirometry, relevance and recommendations. The Journal of Experimental Biology. 216 (Pt 15), 2771-2782 (2013).
- Voolstra, C. R., et al. Extending the natural adaptive capacity of coral holobionts. Nature Reviews Earth & Environment. 2 (11), 747-762 (2021).
- Hoegh-Guldberg, O. Climate change, coral bleaching and the future of the world's coral reefs. Marine and Freshwater Research. 50 (8), 839-866 (1999).
- Morris, L. A., Voolstra, C. R., Quigley, K. M., Bourne, D. G., Bay, L. K. Nutrient availability and metabolism affect the stability of coral-symbiodiniaceae symbioses. Trends in Microbiology. 27 (8), 678-689 (2019).
- Yellowlees, D., Rees, T. A. V., Leggat, W. Metabolic interactions between algal symbionts and invertebrate hosts. Plant, Cell & Environment. 31 (5), 679-694 (2008).
- Rädecker, N., et al. Using Aiptasia as a model to study metabolic interactions in cnidarian-Symbiodinium symbioses. Frontiers in Physiology. 9, 214(2018).
- Matthews, J. L., et al. Optimal nutrient exchange and immune responses operate in partner specificity in the cnidarian-dinoflagellate symbiosis. Proceedings of the National Academy of Sciences. 114 (50), 13194-13199 (2017).
- Davy, S. K., Allemand, D., Weis, V. M. Cell biology of cnidarian-dinoflagellate symbiosis. Microbiology and Molecular Biology Reviews. 76 (2), 229-261 (2012).
- Weis, V. M. Cellular mechanisms of Cnidarian bleaching: stress causes the collapse of symbiosis. The Journal of Experimental Biology. 211 (Pt 19), 3059-3066 (2008).
- Baker, D. M., Freeman, C. J., Wong, J. C. Y., Fogel, M. L., Knowlton, N. Climate change promotes parasitism in a coral symbiosis. The ISME Journal. 12 (3), 921-930 (2018).
- Chakravarti, L. J., van Oppen, M. J. H. Experimental evolution in coral photosymbionts as a tool to increase thermal tolerance. Frontiers in Marine Science. 5, 227(2018).
- Quigley, K. M., Alvarez Roa, C., Beltran, V. H., Leggat, B., Willis, B. L. Experimental evolution of the coral algal endosymbiont, Cladocopium goreaui: lessons learnt across a decade of stress experiments to enhance coral heat tolerance. Restoration Ecology. 29 (3), e13342(2021).
- Buerger, P., et al. Heat-evolved microalgal symbionts increase coral bleaching tolerance. Science Advances. 6 (20), eaba2498(2020).
- Bieri, T., Onishi, M., Xiang, T., Grossman, A. R., Pringle, J. R. Relative contributions of various cellular mechanisms to loss of algae during cnidarian bleaching. PLoS One. 11 (4), e0152693(2016).
- Tremblay, P., Gori, A., Maguer, J. F., Hoogenboom, M., Ferrier-Pagès, C. Heterotrophy promotes the re-establishment of photosynthate translocation in a symbiotic coral after heat stress. Scientific Reports. 6, 38112(2016).
- Tremblay, P., Grover, R., Maguer, J. F., Hoogenboom, M., Ferrier-Pagès, C. Carbon translocation from symbiont to host depends on irradiance and food availability in the tropical coral Stylophora pistillata. Coral Reefs. 33 (1), 1-13 (2014).
- Wooldridge, S. A. Is the coral-algae symbiosis really 'mutually beneficial' for the partners. Bioessays. 32 (7), 615-625 (2010).
- Wooldridge, S. A. Breakdown of the coral-algae symbiosis: towards formalising a linkage between warm-water bleaching thresholds and the growth rate of the intracellular zooxanthellae. Biogeosciences. 10 (3), 1647-1658 (2013).
- Coles, S. L., Jokiel, P. L. Effects of temperature on photosynthesis and respiration in hermatypic corals. Marine Biology. 43, 209-216 (1977).
- Marubini, F., Davies, P. S. Nitrate increases zooxanthellae population density and reduces skeletogenesis in corals. Marine Biology. 127, 319-328 (1996).
- Iglesias-Prieto, R., Matta, J. L., Robins, W. A., Trench, R. K. Photosynthetic response to elevated temperature in the symbiotic dinoflagellate Symbiodinium microadriaticum in culture. Proceedings of the National Academy of Sciences. 89 (21), 10302-10305 (1992).
- Karako-Lampert, S., Katcoff, D. J., Achituv, Y., Dubinsky, Z., Stambler, N. Responses of Symbiodinium microadriaticum clade B to different environmental conditions. Journal of Experimental Marine Biology and Ecology. 318 (1), 11-20 (2005).
- Blanckaert, A. C. A., Reef, R., Pandolfi, J. M., Lovelock, C. E. Variation in the elemental stoichiometry of the coral-zooxanthellae symbiosis. Coral Reefs. 39, 1071-1079 (2020).
- Harianto, J., Carey, N., Byrne, M. respR-An R package for the manipulation and analysis of respirometry data. Methods in Ecology and Evolution. 10 (6), 912-920 (2019).
- Gamble, S., Carton, A. G., Pirozzi, I. Open-top static respirometry is a reliable method to determine the routine metabolic rate of barramundi. Lates calcarifer. Marine and Freshwater Behaviour and Physiology. 47 (1), 19-28 (2014).
- Burford, B. P., et al. Rapid range expansion of a marine ectotherm reveals the demographic and ecological consequences of short-term variability in seawater temperature and dissolved oxygen. The American Naturalist. 199 (4), 523-550 (2022).
- Morozov, S., McCairns, R. J. S., Merilä, J. FishResp: R package and GUI application for analysis of aquatic respirometry data. Conservation Physiology. 7 (1), coz003(2019).
- Leclercq, N., Gattuso, J. -P., Jaubert, J. Primary production, respiration, and calcification of a coral reef mesocosm under increased CO2 partial pressure. Limnology and Oceanography. 47 (2), 558-564 (2002).
- Anthony, K. R. N., Hoegh-Guldberg, O. Variation in coral photosynthesis, respiration and growth characteristics in contrasting light microhabitats: an analogue to plants in forest gaps and understoreys. Functional Ecology. 17, 246-259 (2003).
- Moulin, L., et al. Long-term mesocosms study of the effects of ocean acidification on growth and physiology of the sea urchin Echinometra mathaei. Marine Environmental Research. 103, 103-114 (2015).
- Quigley, K. M., Bay, L. K., van Oppen, M. J. H. Genome-wide SNP analysis reveals an increase in adaptive genetic variation through selective breeding of coral. Molecular Ecology. 29 (12), 2176-2188 (2020).
- Brunner, C. A., Ricardo, G. F., Uthicke, S., Negri, A. P., Hoogenboom, M. O. Effects of climate change and light limitation on coral recruits. Marine Ecology Progress Series. 690, 65-82 (2022).
- Quigley, K. M., Alvarez Roa, C., Torda, G., Bourne, D., Willis, B. L. Co-dynamics of Symbiodiniaceae and bacterial populations during the first year of symbiosis with Acropora tenuis juveniles. MicrobiologyOpen. 9 (2), e959(2020).
- Quigley, K. M., Bay, L. K., Torda, G., Willis, B. L. Leveraging new knowledge of Symbiodinium community regulation in corals for conservation and reef restoration. Marine Ecology Progress Series, 600. , 245-253 (2018).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved