
Method Article
塩基対解像度でDNAメチル化の評価のための強化されたの減少表現重亜硫酸塩シークエンシング
* これらの著者は同等に貢献しました
要約
Enhanced Reduced Representation Bisulfite Sequencing is a method for the preparation of sequencing libraries for DNA methylation analysis based on restriction enzyme digestion combined with cytosine bisulfite conversion. This protocol requires 50 ng of starting material and yields base pair resolution data at GC-rich genomic regions.
要約
DNAメチル化パターンのマッピングが重く正常および疾患組織中で検討されている。種々の方法は、細胞中のシトシンメチル化パターンを調べるために確立されている。全ゲノムバイサルファイトシークエンシングの減少表現は、GCリッチなゲノム遺伝子座での定量的な塩基対の分解能シトシンメチル化パターンを検出するために開発された。これは、亜硫酸水素塩変換に続いて、制限酵素の使用を組み合わせることによって達成される。強化縮小表現重亜硫酸塩シークエンシング(ERRBS)が覆われ、生物学的に関連するゲノム遺伝子座を増加し、ヒト、マウス、および他の生物由来のDNA中のシトシンのメチル化をプロファイルするために使用されている。 ERRBSは、ライブラリーの調製において使用するための低分子量フラグメントを生成するために、DNAの制限酵素消化を開始する。これらの断片は、次世代配列決定のための標準ライブラリ構築にかけられる。最終amplificati前の非メチル化シトシンの重亜硫酸塩変換ステップに覆われたゲノム遺伝子座におけるシトシンのメチル化レベルの定量的なベースの解像度を可能にします。プロトコルは、4日以内に完了することができる。指定されたシーケンシング対照レーンを使用した場合、配列決定最初の3つの塩基で低複雑度にもかかわらず、ERRBSライブラリは、高品質のデータを得る。マッピングおよびバイオインフォマティクス解析は、次に実行され、歩留まりのデータを容易にゲノムワイドな様々なプラットフォームと一体化することができる。 ERRBSは、実現可能な研究用途の範囲内で、ヒト臨床サンプルおよび適用を処理すること小さな入力材料量を利用することができる。生成されたビデオは、ERRBSプロトコルの重要なステップを示しています。
概要
シトシン(5-メチル)におけるDNAメチル化インプリンティング、X染色体不活性化、開発、および遺伝子発現1-8の調節を含むがこれらに限定されない生物学的過程、種々の哺乳動物細胞において重要なエピジェネティックなマークである。悪性および他の疾患におけるDNAメチル化パターンの研究は、疾患特異的パターンを決定し、疾患の病因と潜在的なバイオマーカーの発見9-17の理解に貢献してきました。 DNAのメチル化状態のためのエピゲノムを調べる多くのプロトコルがある。これらは、親和性に基づく、制限酵素ベース、および下流マイクロアレイまたは配列決定プラットフォームを利用して亜硫酸水素塩変換ベースのアッセイに分けることができる。さらに、複合重亜硫酸塩制限分析18および減少表現重亜硫酸塩シークエンシング(のCFM.rrb 19)、を含むがこれらに限定されますが、いないこれらの一般的なカテゴリを埋めるいくつかのプロトコルがあります。
のCFM.rrbはもともとマイスナーらによって記載された。19,20。プロトコルは、費用対効果の高い21,22で定量的な塩基対の解像度データが得られた重亜硫酸塩シークエンシング、続くGCリッチなゲノム領域を豊かにする手順を紹介しました。 GCに富む領域はMspIを(C ^ CGG)制限酵素によって標的化される、およびシトシンのメチル化は、ポリメラーゼ連鎖反応(PCR)増幅、続いてシトシンの重亜硫酸塩変換(ウラシルに修飾されていないシトシンの脱アミノ化)によって解決される。のCFM.rrbは、遺伝子のプロモーターおよび全ゲノム配列決定のために必要な画分におけるCpGアイランドの大部分をカバー。しかしのCFM.rrbは、CpGの海岸と生物学的関連性の他の遺伝子間領域の限られた範囲を持っていた。いくつかのグループは、これらのゲノム領域23-25 の方法論と結果の報道に改良元のレポート以降に更新のCFM.rrbプロトコルを公開している。強化されたの減少表現重亜硫酸塩Sequencing(ERRBSは)のCFM.rrbに比べライブラリ準備の修正および代替データ整列アプローチ26を含む。 ERRBSは生成されたデータで表現CpGの高い数をもたらし、26を尋問すべてのゲノム領域のカバレッジを増加させた。この方法は、ヒト患者及び他の動物標本26-30におけるDNAメチル化パターンを解決するために使用されている。
ERRBSプロトコルが完了し、代表的なヒトDNAを使用して生成されたデータに必要なすべてのステップにオファーの詳細を説明した(サンプルは、以前に報告され、非識別患者サンプル31から得られ、正常なヒトのドナーからのCD34 +骨髄サンプルされた)。プロトコルは、サンプルあたりの処理時間が短縮され、ライブラリーサイズの選択において高い精度を可能にする自動化されたサイズ選択プロセスを含む。プロトコルは、確立された分子生物学技術のシリーズを組み合わせた。高分子量DNAはワット消化エンド修理、-テーリング、およびメチル化されたアダプターの連結に続いてメチル化非感受性制限酵素(MspIで)i番目。 GCリッチフラグメントのサイズ選択は、亜硫酸水素塩変換し、配列決定の前にPCR増幅に続いている。重亜硫酸塩変換は、以前に32に記載されているとデータ分析やアプリケーションの詳細なレビューはお薦めや参照が読者の使用のために含まれていますが、この論文の範囲を超えている。プロトコルは、4日間にわたって行われ、小さな入力(50 NG以下)材料量に適していることができます。差動メチル化部位と領域決定のためだけでなく、エピジェネティックな多型検出のためだけでなく、十分なCpG部位当りの高いカバレッジ記載収量データなどのプロトコルは、 ら 33 Landanに記載されているように。
プロトコル
実行されるすべての手順は、医療機関動物実験委員会のインディアナ大学大学院によって承認され、健康ガイドラインの国立研究所に従っている。
1.手術手技
- NIHガイドライン25に従って無菌滅菌手袋を使用して、この手順の間に技術、機器、および無菌手術野を維持します。 (完全なリストに特異的な試薬/機器の表を参照)、それらをオートクレーブで手術を開始する前に、ツールを滅菌する。動作中のツールを殺菌するためにガラスビーズ滅菌器を使用してください。
2.麻酔と準備
- 獣医イソフルラン気化器システムを用いて、0.9リットル/分の酸素と2.5%のイソフルランの混合物で麻酔ボックス内にマウスを麻酔。マウスを箱から取り出す前に、身体の位置の変化に応答しないことを確認してください。
- MOUに眼軟膏を適用します乾燥からそれらを保護するために、それ自体の目。
- ノーズコーンにあるボックスからのガスの流れを切り替えます。コーンの内側の鼻と口を備えた外科用パッドと吸収ベンチ紙で覆われて加熱されたパッドの上に真正面からその左側にマウスを置きます。継続的にマウスの呼吸のリズムと速度を監視し、必要に応じてイソフルランレベルを調整 - 麻酔の適切なレベルを維持するために、(2.5の間で3%イソフルラン)、および総鎮静を確認するために、つま先ピンチ反射を使用しています。
3.外科的アプローチ
- 合わせて、手術野でステレオスコープを集中。それは視野の端に沿って配置されるように、ノーズコーンとテープを上下に調整する。
- マウスは、その左側に横たわっていると、切開が行われる耳の後ろの領域を露出させ、ノーズコーンに右耳の端をテープで固定します。後耳介静脈が耳全体に水平に移動することを確認してください。 Tの正しい配置に注意してください彼は耳の動物とテーピングはすぐに顔面神経を見つけるために重要である。
- にし、70%エタノールで耳の後ろに毛皮を湿らせ、かみそりまたは外科用メスの刃を使用して、手術部位を剃る。毛皮を事前に湿潤この解剖学的位置に簡単に剃ることができます。
- 70%エタノールに続くようなベタジン外科スクラブ(7.5%ポビドンヨード)などのヨウ素溶液、で皮膚をきれいにしてください。徹底的に領域を消毒するために、この清掃をさらに2回繰り返します。
- 、切開する耳の突起にエリア後方に尾側耳から後耳介静脈をトレースする場所を決定する。隆起に3ミリメートルの後部 - 春のはさみを使用して、4ミリメートル切開2を作る。
- 鈍的切開を使用して皮下脂肪および筋膜を通して解剖。 、血管または筋肉組織を簡単に破損する可能性があるため、ハサミで直接切断しないでください。
- 出血が発生した場合は、無菌の綿棒で手術部位に圧力を適用する少なくとも30秒間。重要な流体損失が発生した場合、25または27 G針を使用して0.9%滅菌生理食塩水までの0.5mlを腹腔内にマウスを注入する。
- 顔面神経を見つけるために、いくつかの主要なランドマーク、脊髄副神経、外耳道を使用し、(後述)二腹筋を前方。顔面神経の枝が可視化されるまで、これらのランドマークの周りに解剖。それは明らかにし、筋膜の層が下にある構造に付着されたときに神経が重要な白色固体構造として表示されます。
- 皮下脂肪と筋膜が解剖された後、僧帽筋を支配するために頭蓋骨の尾の部分から移動する脊髄副神経を検索します。顔面神経は、脊髄アクセサリ神経に深い。
- 真珠のような白に見え、顔面神経への吻側見ることができる軟骨外耳道を探す。
- の上部とC上に位置し、前二腹筋の筋腹を探す顔面神経にaudal。
- 顔面神経の主枝を可視化されると、茎乳突孔からその起源を見つけるために背側にそれらをトレースします。細かい使用すると、開いて新しく高度な面積を維持するために背側鉗子を移動した後、開いた手術部位を保持する神経のパス以下の春のはさみのヒントを進めるためにデュモン鉗子#45分の5をひっくり返した。
- この時点で、頬骨と顔面神経の幹、頬、および限界下顎枝を可視化する。
注:一時的な分岐が孔に近い見つかります。近い顎にその上下に限界下顎神経の枝、従って、これらの神経枝は、このレベルでは表示されません。- 神経切断を行う場合は、先端の細い鉗子で軽く神経を安定させ、春のはさみで神経を切った。脳幹から神経をavulsing防ぐために、ピンセットで神経にあまりにも多くのトラクションを適用することは避けてください。押す互いに離れる切り株、または全く再接続が発生しないことを保証するために、遠位神経の部分をカットし、削除する。
- 挫滅損傷を実行した場合は、すべての軸索を切断する一定の圧力を用いて30秒間の神経を圧縮するデュモン#45分の5鉗子を使用し、最初のクラッシュサイトに直交する第2の角度でこのクラッシュを繰り返します。そうでなければ怪我は動物の間一貫性がなくなります、30秒のクラッシュ時の圧力の可変量を適用することは避けてください。
4.閉会と回復
- 下にある構造上の脂肪と筋肉を再配置します。
- 切開のエッジを近似すると7.5ミリメートルの創傷クリップを使用して傷を閉じます。縫合糸または接着剤は、創傷閉鎖のためにも許容される。術後の鎮痛薬は、この時点で提供することができる。
- マウスの耳からテープを外します。イソフルランの流れをオフにして、マウスが1分30秒純酸素を呼吸することができます。 PL麻酔から回復する寝具で空のケージにマウスをエース。
- マウスが回復されると、顔面神経麻痺の確証的兆候がその挙動を調べる。ウィスカーが麻痺して戻って頬に向かって傾斜してなり、鼻がずれてしまう、との目は、空気のパフに応じて点滅しません。
- ハウス動物共同で、彼らが女性である場合は、手術後。彼らはより積極的であり、強制的に感染につながる、彼らのcagemateの創傷クリップを削除する傾向があるため、共同で雄マウスを収納しないでください。必要であれば、この時点では術後の鎮痛薬を提供します。
- 全く感染または他の合併症は術後に発生していないことを確認するために操作した後、数日間一日一回のマウスを監視します。彼らは自分の上で落ちていない場合は、10日手術後 - 7創傷クリップを外します。
- 目のまばたき反射が再いずれかになるまで、毎日の角膜合併症を防ぐために、罹患した眼に眼軟膏を潤滑適用カバーまたは安楽死まで、。
結果
図1は、記載されたプロトコルを通じて説明されているキーの手順を、ハイライト、ERRBSの概要を説明します。 ERRBSライブラリは、50ngのインプットDNAを用いて調製した。
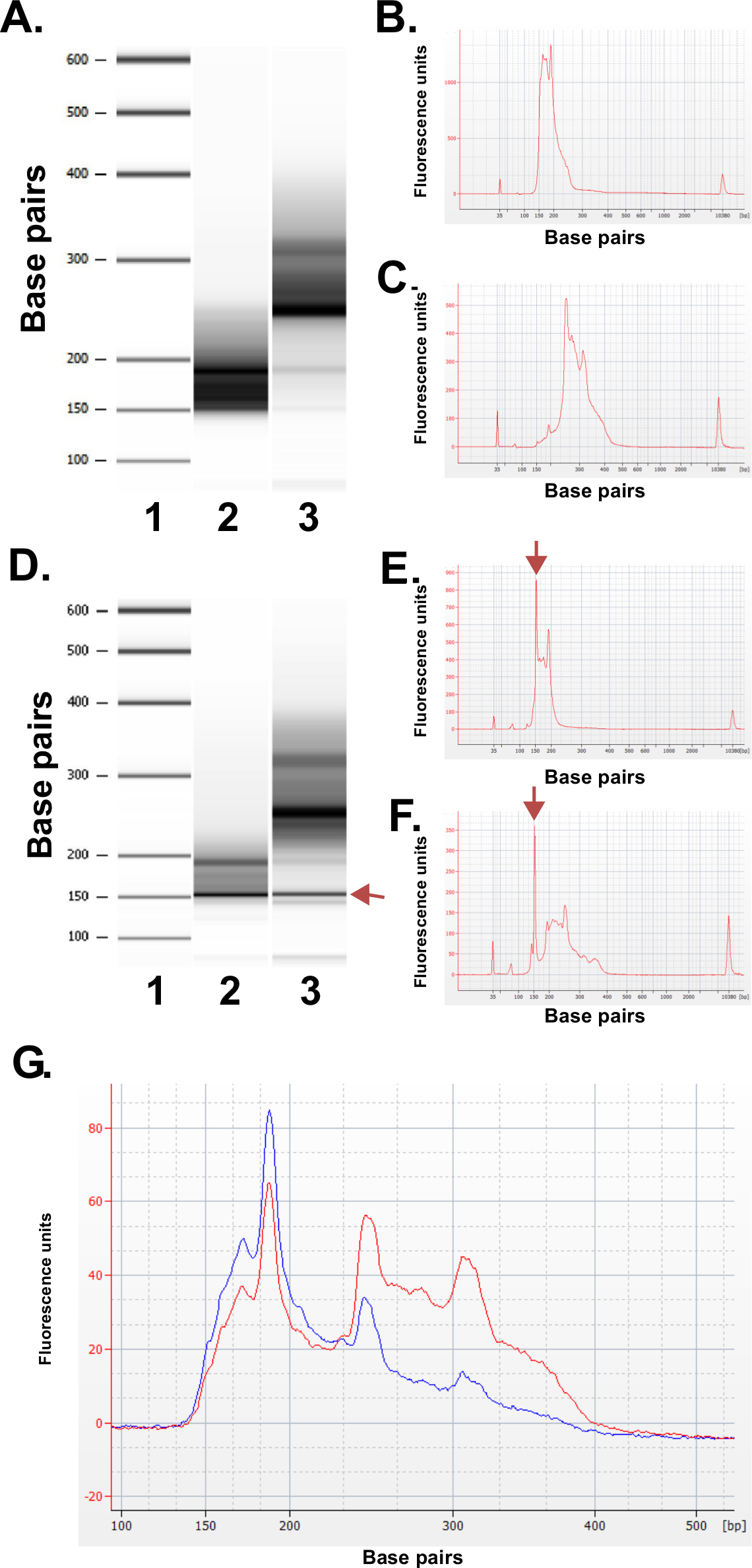
準備ライブラリの品質を評価する。図書館生産は日常的に150〜250 bpおよび250〜400 BP( 図3A-C)の割合の大きさが得られます。サンプル間のライブラリのサイズ分布のわずかな違いが予想される。特定のシーケンスの濃縮を示す、両方のより低いおよびより高いライブラリの分画で非常に強いDNAの大きさがあることに注意してください。反復DNAの家族の濃縮でのMspI消化の結果は190塩基対、250 bpおよびERRBSライブラリ内の310 bpのでヒトゲノム中に存在する配列。これら三つの反復は、( 図3A-Cおよび3Gを参照)ERRBSライブラリ20の特性署名を表す。代表的なライブラリーは、次世代の配列を配列決定したRを用いて、シングルエンドの読み取り。イルミナHiSeq 2500シーケンサー上で推奨されるライブラリ濃度でロードするときは、1mm 2あたり500,000-700,000のクラスタ密度が期待されている。クラスタのこのクラスタリング密度、81.6パーセント±3.14パーセント(N = 81)のフィルタ( 図4A)を渡す。 、ライブラリインサート(MspIで認識部位:C ^ CGG)の複雑度の低い端に独立した対照レーン場合、シーケンス処理中に記録された強度値と品質スコアは、しかし、最初の3つの塩基( 図4B-C)で非常に可変である(; 図4D Q30値)基地の85%が30以上の品質スコアを持つことになります、(説明を参照)に含まれています。
プロトコル収量の塩基対解像度データに記載されているように、データのアライメントおよびシトシンメチル化決定した( 表7)。高出力モードでヒトゲノム、HiSeq 2500の1つのレーンでERRBSライブラリーの51サイクル単読み取るシークエンス実行のレギュラーLY 153194882±12918302合計は品質フィルタリングおよびアダプタが152231183±13189678収量をトリミングした後、解析パイプラインへの入力のために読み取ることを読み取って生成します。 ERRBSライブラリの平均マッピング効率は10倍ののCpGあたりの最小カバレッジと84.94±16.29(N = 100)ののCpGあたりの平均カバレッジ3183594±713547 CpGの表現と5.92パーセント±通常は62.95パーセントである。
ERRBSプロトコルは、(:多重化された配列決定のためのプロトコル適合補足ファイル1を参照)を多重化に適している。代表シングからのデータで動作を図5に要約されたデータは、多重化されたシークエンシングランから(51サイクル単読みシングラン; N =レーンあたり2ライブラリのための128であり、n =レーンあたり3つのライブラリのための11であり、n = 11 4用のライブラリはn = 100)、ならびにsimulat単一のレーンをダウンサンプリング(51サイクル単読み取るシング実行ERRBSライブラリの完全な車線シングと比較した)レーン当たり電子50%、33%、25%は、レーンごとに読み込む(図2、図3、レーン当たり4サンプル多重それぞれであり; n = 3)。サンプルあたり読み込み数は多重化率で減少すると、10倍の最小被覆率で覆われたCpGの数とのCpGあたりのカバー率は( 図5および表8)にも減少する。期待非CpG部位の平均コンバージョン率は0.04%(N = 400)±99.85パーセントである。変換率は99%以上が偽メチル化レベルの高い速度をもたらすことができ、最適な重亜硫酸塩変換未満を示すことができる。
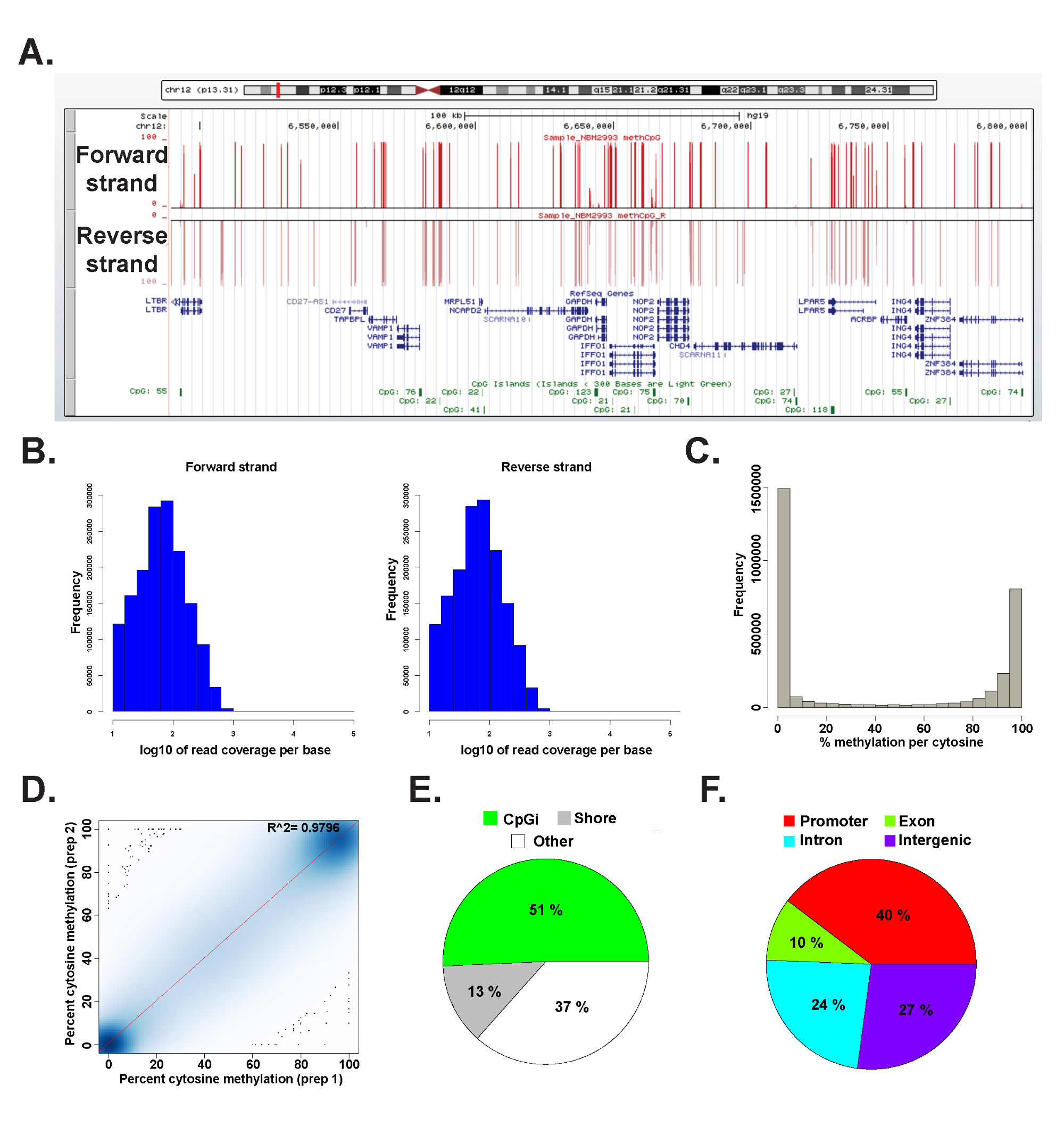
代表的なヒトのゲノムDNAから調製ERRBSライブラリからのデータはmethylKitパッケージ26を (コマンドの詳細については補足コードファイル1を参照)を使用してR 2.15.2 45で分析した。データは、一般的に使用されるゲノムブラウザ( 図6A)で可視化することができる。シトシンメチル化データが均等に両方の鎖( 図6B)に由来し、全体の範囲である潜在的なシトシンメチル化レベルのスペクトル( 図6C)。データ結果との間に代表的なヒトDNAサンプルの利回り高い一致( 図6D)から技術的反復の分析とゲノム遺伝子座の広いスペクトルにCpGをカバーしています( 図6E及びFと、以前に26を説明する)。技術的複製が高いR 2値(97%以上)が得られますが、生物学的レプリカは0.92から0.96 26までの範囲のR 2値が得られると、異なるヒト細胞型を比較した(データは示さず)0.86以上2の値より低いRが得られる。

図1:ERRBSプロトコルステップのフロー· チャート。チャートは伝統的な作業日で完了することができるステップを表します。*潜在的な一時停止ポイントを示す(すぐに従うる連結は、クリーンアップし、サンプルは、プロトコルの期間に進む前に、-20℃で凍結することが可能なサイズ選択、プロトコルステップ5)の前に。

図2:サイズ選択プロトコル。 (A)ERRBSピピン準備プロトコルで使用されている設定のスクリーンショット(プロトコルセクション5.1.2参照- 5.1.6)を:(1)を選択しカセットタイプ。 (2)選択基準が使用される。 (3)各レーンのコレクションモードを選択します。 (4)回収bpの範囲を入力します。 (5)プロトコルを保存するプロトコルセクション5.2で使用される手動のゲル抽出の(B)ステージ:(1)可視化ゲルはしご。 (2)カミソリの刃を使用して、サイズ選択のためのサイズのマーク。切除したサンプルの(3)画像(下分数:150〜250 bpおよび高い割合:250〜400塩基対)。">この図の拡大版をご覧になるにはこちらをクリックしてください。

図3:バイオアナライザー機を使用して、ヒトDNAサンプルから調製した代表的ERRBSライブラリの精度管理の結果。標準ラダー(1)、下側のライブラリの画分(ピピンプレップから135から240 bpの画分)を示す(A)ゲル状の画像。 2)以上のライブラリ画分(ピピンプレップから240から410 bpの画分)。 。期待下ライブラリ分の3)(B)バイオアナライザー電気泳動(C)が予想より高いライブラリ画分のバイオアナライザー電気泳動図D - 。。F)質の悪いライブラリ準備から代表的なデータ。標準はしご(1)のゲル様画像(D)、下位ライブラリの画分(2)以上のライブラリの画分(3)。 arroでマーク150 bpのでバンドwは、アダプタの過剰な量を示している。下の電気泳動(E)と(矢印でマーク)の150bpでの過剰アダプタピークが高いライブラリ画分(F)。シークエンシングのためにプールERRBSライブラリの(G)バイオアナライザー電気泳動図。レッドトレースは、上位と下位画分の同じ表現と高品質プールされたライブラリを表します。ブルートレースは、より高いと低分画の同じ表現の不足のためにシークエンシングに適していないプールされたライブラリを表します。 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}

図4:代表のためのチャートを配列決定は、高出力でHiSeq 2500シーケンサーで51サイクル単読みシングランをERRBSモード。 (A)クラスタ密度(ミリメートル当たり2 =千のクラスタが乗K / mmの、青)とERRBSライブラリと2レーンにフィルター(緑)を通過クラスター密度(B)とのレーンに最初の30サイクルで見られる典型的な強度。 ERRBSライブラリ。のための最初の3サイクルの強度でのMspI消化からCGG署名を注意してください。1 ERRBSレーンにおける各サイクルのために30以上の品質スコアと拠点の(C)の割合(> Q30%)。(D)品質スコア分布1 ERRBSレーン内のすべてのサイクル。青= Q30、グリーンより小さい=以上Q30に等しい大きい。このレーンにおいて、塩基の84.7%は、30以上の品質スコアを持っていた。

図5:出力結果の配列決定。レーンシーケンシングRUあたりの多重化して単一のサンプルから実験データのボックスプロット NS(緑のボックスとして表示される)と3 ERRBSライブラリのシーケンシングの実行からシミュレートされたダウンサンプリングにより得られたデータの(青いボックスとして表示、各シークエンス実行のために5回サンプリング)51サイクル単読みシークエンシングから実行される多重化係数が対応する。 ERRBSライブラリの数は、レーンごとに配列決定した。レーンごとに単一ERRBSライブラリから1 =全部レーンまたは読み取り、表現するデータを100%;レーン2 = 50%、レーンごとに2つのERRBSライブラリからのデータを表す。レーン3 = 33%、レーンあたり3 ERRBSライブラリからのデータを表す。レーンと、4 = 25%レーンあたり4 ERRBSライブラリからのデータを示す。(A)は、読み出し回数、または多重化率ごとに、分析された配列の数(B)のCpGの数は多重化ごとに配列決定データによってカバー要因。(C)多重化因数あたりのCpGあたりの平均カバレッジ。_blank ">この図の拡大版をご覧になるにはこちらをクリックしてください。

図6:ヒトゲノムDNAから調製ERRBSライブラリからの代表的なデータカリフォルニアの(A)大学サンタクルーズ校(UCSC)ゲノムブラウザERRBSシーケンシングレーンからの代表的なデータの43の画像。。 y軸のスケールバーは10倍の最小で覆わ各シトシンで0-100%メチル化を表す。トップカスタムトラックはフォワード鎖を表し、下側のカスタムトラックは、逆鎖を表す。に沿って前方のCpGカバレッジの6,489,523-6,802,422(hg19)のRefSeq遺伝子と、このゲノム領域内のCpGアイランドを含む(B)の分布ヒストグラムと代表的なヒトCD34 +骨髄サンプル中の鎖を逆に(C)分布ヒストグラム:。chr12ですが示さ。ERRBSれる覆わCpGの割合を示す代表的なヒトCD34 +骨髄試料中の両方の鎖に沿って、CpGメチル化のレベルが、ヒトDNA試料の代表的な技術的複製からのCpGメチル化のレベル(D)、相関プロットの(E)円グラフ(F)円グラフは、遺伝子のプロモーターに注釈付きERRBSでカバーCpGの割合を示す。CpGアイランド(ライトグリーン)、ヒトゲノムDNAから調製した代表的なサンプル中のCpGの海岸(グレー)、その他の地域(白)に注釈付き(赤)、エクソン(緑)、イントロン(青)と遺伝子間領域(紫)。 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}
| 試薬 | ボリューム | コメント |
| 10倍のT4 DNAリガーゼ反応バッファー | 10μL | |
| デオキシ三リン酸(dNTPを)溶液ミックス | 4μL | 各ヌクレオチドの10mmのミックス |
| T4 DNAポリメラーゼ | 5μL | 3000単位/ ml |
| DNAポリメラーゼIラージ(クレノウ)フラグメント | 1μL | 5000単位/ ml |
| T4ポリヌクレオチドキナーゼ | 5μL | 万単位/ ml |
| DNアーゼを含まない水 | 45μL |
表1:最終修復反応試薬端修復反応(プロトコールステップ2.1)に使用される試薬名と数量。
| 試薬 | ボリューム | コメント |
| 10倍反応バッファー | 5μL | 例えば、NEバッファー2 |
| 1 mMの2'-デオキシアデノシン5'-三リン酸(dATPを) | 10μL | |
| ノウフラグメント(3 '→5'エキソ) | 3μL | 5000単位/ ml |
表2:A-テーリング反応試薬 A-テーリング反応(プロトコルステップ3.1)で使用される試薬名と数量。。
| 試薬 | ボリューム | コメント |
| DNアーゼを含まない水で15μMにアニールアダプター | 3μL | PEアダプタ1.0とPEアダプター2.0。シーケンスおよび参照については、表4を参照してください。 |
| 10倍のT4 DNAリガーゼ反応バッファー | 581; L | |
| T4 DNAリガーゼ | 1μL | 200万単位/ ml |
| DNアーゼを含まない水 | 31μL |
表3:アダプターライゲーション反応試薬アダプターライゲーション反応(プロトコールステップ4.2)に使用される試薬名と数量。

表4:ライゲーション反応(プロトコルステップ4)、PCR増幅ステップ(プロトコルステップ7)でERRBSプロトコル全体で使用されるオリゴのERRBSプロトコルで使用されるオリゴリスト。
| 試薬 | ボリューム | コメント |
| 10倍のファストスタートハイ·フィデリティ反応緩衝のWi第18 mMの塩化マグネシウム | 20μL | |
| 10のdNTP溶液ミックス | 5μL | |
| 25μMPCR PEプライマー1.0 | 4μL | 表4を参照してください |
| 25μMPCR PEプライマー2.0 | 4μL | 表4を参照してください |
| ファストスタートハイフィデリティ酵素 | 2μL | 5ユニット/μlファストスタートTaq DNAポリメラーゼ |
| DNアーゼを含まない水 | 125μL |
表5:PCR反応試薬 PCR増幅反応(プロトコールステップ7.1)に使用される試薬名と数量。
| プロトコルステップ | 試薬/プロトコルの詳細 | インプットDNA量 | ||
| 5-10 NG | 25 NG | 50 NG | ||
| 1 | MspIを酵素 | 1μL | 2μL | 2μL |
| MspIを、反応体積を消化 | 50 | 100 | 100 | |
| 4 | 連結反応でアダプタ | 1μL | 2μL | 3μL |
| ライゲーション反応容積 | 20μL | 25μL | 50μL | |
| 5 | サイズ選択プロトコル | マニュアルゲルのみ | ピピンプレップまたは手動ゲル | ピピンプレップまたは手動ゲル |
| 7 | PCRプライマー濃度 | 25μM | 25μM | 14サイクルの10μM。 18サイクルの25μM |
| PCRサイクルの数 | 18 | 18 | 14-18 | |
表6:5-50 NGに至るまでの入力物質の量のためのプロトコルのステップの変更プロトコルを通していくつかのステップは、出発物質の様々な量から質の高いライブラリを生成するために使用される試薬量の修正が必要。キー試薬量への変更は、ここに含まれています。それに応じて反応におけるバッファと水のボリュームを調整します。
| CHR | ベース | ストランド | 報道 | freqC | FREQT |
| CHR1 | 10564 | R | 366 | 85.52 | 14.48 |
| CHR1 | 10571 | F | 423 | 91.25 | 8.75 |
| CHR1 | 10542 | F | 432 | 91.2 | 8.8 |
| CHR1 | 10563 | F | 429 | 94.64 | 5.36 |
| CHR1 | 10572 | R | 366 | 96.99 | 3.01 |
| CHR1 | 10590 | R | 370 | 88.11 | 11.89 |
| CHR1 | 10526 | R | 350 | 92 | 8 |
| CHR1 | 10543 | R | 368 | 92.93 | 7.07 |
| CHR1 | 10525 | F | 433 | 91.92 | 8.08 |
| CHR1 | 10497 | F | 435 | 88.74 | 11.26 |
| レーンあたりERRBSのライブラリの数 | 一意に整列の数が読み取る平均 | CpGの数をカバー意味 | のCpGあたりの平均報道 |
| 1 | 152231184±13189678 | 3183594±713547 | 85±16 |
| 2 | 77680837±7657058 | 2674823153494± | 49±9 |
| 3 | 49938156±2436865 | 2552186± - 76624 | 39±2 |
| 4 | 34457208±4441686 | 1814461±144339 | 28±4 |
表8:シングル多重ERRBSライブラリを配列決定から代表的なパラメータ示すのは、51サイクル単読みシークエンスランからのレーンあたりのデータです:一意に整列の平均値と標準偏差を読み取り、単一の配列決定から得られたカバーのCpGとCpG部位ごとの報道の数レーンあたりERRBSライブラリ(N = 100)、レーンあたり2 ERRBSライブラリ(N = 128)、レーン(N = 11)ごとに3つのERRBSライブラリ、およびレーン(N = 11)ごとに4つのERRBSライブラリ。
ディスカッション
生体関連ゲノム領域におけるシトシンのメチル化のプロトコル提示収量の塩基対解像度データ。出発物質の50 ngのために最適化されて書かれたプロトコルは、しかし、それは、入力材料の範囲(5ngのまたは複数)26を処理するように適合させることができる。 表6に見られるように、これは、プロトコルのステップのいくつかの調整が必要となる。ERRBSライブラリはペアエンド配列決定およびさらなるゲノムカバーに適しているシング長い51サイクルより読み込むことによっても達成することができる。多重化された配列決定は、サンプルあたりのコストプロトコルを提供するが、これはデータに表さCpG部位当りの縮小範囲( 図5および表8)になり、かつ当たり高いカバー力を必要とする分析を実行するために、カバレッジの十分な深さが得られないであろうCpG部位(Landan ら 33で説明した例のように)。最後に、このプロトコル(または重亜硫酸塩ベースのprotocol)はメチル-シトシンとヒドロキシメチルシトシン46,47を区別することができません。しかしながら、他のプロトコルと統合することができる生成されたデータは、異なる修飾を線引きする48,49をもたらし、他のシトシン修飾は最近、関心があるべきであり、50を報告した。
図3A-Cに示されており、一回シークエンシングのためにプールのような高品質のライブラリが表示されます両方のライブラリ留分から等しいモルの寄与を表す図3Gに示すようにトレース(赤トレース)が得られる。ライブラリ調製障害は手順中の任意の工程から生じ得る。分解したDNAが処理される場合は、このプロトコルで説明シングパラメータを使用してのMspI断片が濃縮され、したがって、低いのCpGカバレッジされていないライブラリになります。酵素は、非機能的または不注意の反応のいずれかから除外された場合、プロトコルが期待ライブラリをもたらさないであろう。ライゲーションREAた場合ctionは非効率的であり、アダプタは、予想よりも高濃度であり、および/または使用したプライマー濃度は、ライブラリー、障害が発生することができ、最終的な増幅工程のための制限試薬である。 (バイオアナライザ結果の〜150 bpのピークと見られ; 図3D-F)過剰アダプタは、ライブラリ内にも、ライブラリと過剰のアダプターの両方の無差別クラスタリングにシークエンシングを妨害する。そのようなライブラリは、明らかに正常にシーケンスかもしれないが、読み込みのかなりの部分は、単にアダプタのシーケンスになります。過剰のアダプタがライブラリに認められた場合、それは材料が数量比をアダプタために最適な入力素材を使用して利用可能な場合、ライブラリの準備を繰り返すのがベストです。最後に、ライブラリの効率的なPCR増幅を確実にするために、より低いおよびより高いライブラリ画分を亜硫酸水素塩変換し、PCR濃縮ステップを通して別々の試料として維持されている。これを怠ると、Pの間に、増幅の微分効率を生み出すのCR( 図3G青のトレースに見られるように)上位と下位画分の反応および配列決定の際に、各ライブラリの画分に覆われたそれぞれのゲノム遺伝子座の不均等な表現の可能性。ユーザーがすぐに生成されているライブラリーを増幅するために必要な最適なPCRサイクルのさらなる滴定亜硫酸水素塩変換後に定量的PCRステップを含むように選択してもよい。
ERRBSライブラリ調製プロトコルは、特定の試薬が推奨されているいくつかの重要なステップがあります。エンドリペア工程では、四ヌクレオチドdNTPミックスの使用は、MspIで酵素スター活性に起因するものと、元のDNAサンプルに存在する剪断DNA断片として、CGオーバーハングを含有していない製品の最終修復を可能にする。これが結果に改善されたのCpG表現になる。連結工程では、ライゲーションをReactiことを保証するために高濃度リガーゼ(200万単位/ ml)およびメチル化アダプターを使用することが重要である上は効率的であり、亜硫酸水素塩変換が正確なデータの位置合わせのための不可欠なアダプター配列に影響しないこと。 PCR工程では、亜硫酸水素塩処理したGCリッチなDNA断片を増幅することができるポリメラーゼを用いて、高い特異性のために必要である。最後に、過剰のアダプターおよびプライマーの除去を確実にするために、(例:アジェンコートAMPure XP)SPRIビーズ精製は、連結およびPCR産物の単離のために列に基づいたアッセイではなく、推奨されます。
高品質のデータを生成するためには、効率的な亜硫酸水素塩変換を保証することが重要である。提示制御は、ユーザーにシークエンシングに先立って変換効率を決定する能力を提供しています。代替として、そのようなラムダDNAなどの非ヒトDNA、内部対照(スパイクイン)として使用することができる。による種の違いにより、このタイプの制御は、直接下流の配列決定に含めることができる( 例えば、ユーによって使用されるような、 ら 34)。しかし、スパイク-iの場合利用sの、それを一意に増幅され、独立して、ライブラリーの配列決定の前に配列決定されない限り、ライブラリの配列決定に先立って変換効率を決定するために使用することができない。決定された換算率は、非CpG部位でのメチル化状態に基づいている。これは、非CpGコンテキストで高いシトシンメチル化の文脈で使用するのに適切でないかもしれない(例えば、胚性幹細胞)と平行サンプルまたは変換効率を評価する他の手段は、この目的のために利用することができる。
ERRBSライブラリの配列に固有であるアドレスには、いくつかの注意事項があります。配列決定されたライブラリの画分の最初の3つの塩基が原因のMspI認識切断部位にほぼ一様に非ランダムである(C ^ CGG; 図4B、Cを参照のこと)。低い品質に重要なデータの損失の可能性で、この結果は、シークエンシング時に見かけ上高いクラスター密度にもかかわらず、貧しいクラスタのローカリゼーションの結果を読み込みます。この障壁を克服するために、専用の制御車線として独立したレーン(PHIX制御や他のライブラリタイプ)の高い複雑さのライブラリが含まれています。高い複雑さのライブラリーは、配列決定され、最初の4つの塩基は、A、C、TおよびGのバランスの表現を含む端部を有する。適切なコントロールレーンは、RNA-seqの、チップ配列、全ゲノム配列決定、または配列決定機メーカー( 例えば PHIXコントロールV3)によって提供されるコントロールなどのライブラリが含まれています。各配列決定実行の対照レーンとして指定する場合は、クラスタの位置を検出するために配列決定の最初の4つの塩基の間に利用される行列生成のための基礎として役立つことができる。高い品質が5.2によりCpG部位あたりの平均カバレッジを上げる(n = 4)をしますキャプチャさ読み込む。代替的に、この技術的な困難は、以前23に記載のようにダーククエンシングアプローチを用いて克服することができる。その他のシーケンシング基準は製造業者のプロトコルごとの標準操作手順に従ってください。のCpG Cあたりの最後に、報道データ分析のための芳泉は、目的の生物学的質問によってユーザによって部分的に案内される。 10倍のカバレージ閾値は、関心のものであるべきであるが、このしきい値を下げることができ、高いカバレッジ分析アプローチを与える。
ERRBSデータ解析の完全な議論はしかし、差別的メチル化シトシンと地域は、オープンソースツール31,51-53を用いて決定することができる、この記事の範囲を超えています。追加の分析の考慮事項とアプローチは54,55を十分に記載されており、読者は計画された分析に最も適切なツールについての文献を検索することが奨励されている。
他の公開方法と比較して、ERRBSは再現性の記載された歩留まり率が高いように行う4日間のプロトコルを提供しています。それのMassARRAY EpiTYPER 26標準金に比べて検証された、費用対効果の高いカバレッジデータ用であり、各種の入力材料に適応可能であるおよび配列決定アプローチ(臨床サンプル処理及び他の低周波の細胞型に有利な)量。これは、ゲノムワイドな結合転写因子、クロマチンリモデリング、エピジェネティックマークと関心のある他のシトシン修飾プロファイリング他の技術と分析、生物学的に関連する遺伝子座での塩基対の分解能を提供し、統合的に使用することができる。データがこのような研究で使用ERRBSは、総合的な分子的アプローチに貢献し、生物学的なモデルとヒト疾患の研究では、高次元解析を可能にすることができる。
開示事項
著者らは、開示する利害の競合がない。
謝辞
We thank all the authors of the original ERRBS report. We thank Mame Fall for technical assistance. We acknowledge the Weill Cornell Medical College Epigenomics Core for technical services and assistance. The work was supported by a Sass Foundation Judah Folkman Fellowship, an NCI K08CA169055 and ASH-AMFDP12005 to FGB, NIH R01HG006798 and R01NS076465, funding from the Irma T. Hirschl and Monique Weill-Caulier Charitable Trusts and STARR Consortium (I7-A765) to CEM, and an LLS SCORE grant (7006-13) to AMM.
資料
| Name | Company | Catalog Number | Comments |
| MspI | New England Biolabs | R0106M | 100,000 units/ml |
| NEBuffer 2 | New England Biolabs | B7002S | Reaction buffer for MspI enzyme; protocol step 1.2 |
| Phenol solution | Sigma-Aldrich | P4557 | Equilibrated with 10 mM Tris HCl, pH 8.0; see safety and handling instructions at http://www.sigmaaldrich.com/catalog/product/sigma/p4557 |
| Chloroform | Sigma-Aldrich | C2432 | See safety and handling instructions at http://www.sigmaaldrich.com/catalog/product/sial/c2432 |
| Glycogen | Sigma-Aldrich | G1767 | 19-22 mg/ml |
| NaOAc | Sigma-Aldrich | S7899 | 3 M, pH 5.2 |
| Ethanol | Sigma-Aldrich | E7023 | 200 proof, for molecular biology |
| Buffer EB | Qiagen | 19086 | 10 mM Tris-Cl, pH 8.5 |
| tris(hydroxymethyl)aminomethane (Tris) | Sigma-Aldrich | T1503 | prepare a 1 M, pH 8.5 solution |
| Tris- Ethylenediaminetetraacetic acid (TE) | Sigma-Aldrich | T9285 | Dilute to 1x buffer solution per manufacturer's recommendations |
| T4 DNA Ligase Reaction Buffer | New England Biolabs | B0202S | 10x concentration |
| Deoxynucleotide triphosphate (dNTP) Solution Mix | New England Biolabs | N0447L | 10 mM each nucleotide |
| T4 DNA Polymerase | New England Biolabs | M0203L | 3,000 units/ml |
| DNA Polymerase I, Large (Klenow) Fragment | New England Biolabs | M0210L | 5,000 units/ml |
| T4 Polynucleotide Kinase | New England Biolabs | M0201L | 10,000 units/ml |
| QIAquick PCR Purification Kit | Qiagen | 28104 | Used for DNA product purification in protocol step 2.3 |
| 2'-deoxyadenosine 5'-triphosphate (dATP) | Promega | U1201 | 100 mM |
| Klenow Fragment (3'→5' exo-) | New England Biolabs | M0212L | 5,000 units/ml |
| MinElute PCR Purification Kit | Qiagen | 28004 | Used for DNA product purification in protocol step 3.3 |
| T4 DNA Ligase | New England Biolabs | M0202M | 2,000,000 units/ml |
| Methylation Adapter Oligo Kit | Illumina | ME-100-0010 | |
| Agencourt AMPure XP | Beckman Coulter | A63881 | Used in protocol sections that implement magnetic bead purification steps (steps 4.3 and 8.2). Equilibrate to room temperature before use. |
| Pippin Prep Gel Cassettes, 2% Agarose, dye-free | Sage Science | CDF2010 | with internal standards |
| Certified Low Range Ultra Agarose | Bio-Rad | 161-3106 | |
| Tris-Borate-EDTA (TBE) buffer | Sigma-Aldrich | T4415 | |
| Ethidium bromide solution | Sigma-Aldrich | E1510 | 10 mg/ml |
| 50 bp DNA Ladder | NEB | N3236S | |
| 100 bp DNA Ladder | NEB | N3231S | |
| Gel Loading Dye, Orange (6x) | NEB | B7022S | |
| Scalpel Blade No. 11 | Fisher Scientific | 3120030 | |
| QIAquick Gel Extraction Kit | Qiagen | 28704 | |
| EZ DNA Methylation Kit | Zymo Research | D5001 | Used in protocol step 6.2 |
| EZ DNA Methylation-Lightning Kit | Zymo Research | D5030 | Alternative for step 6.2 |
| Universal Methylated Human DNA Standard | Zymo Research | D5011 | Used as bisulfite conversion control |
| FastStart High Fidelity PCR System | Roche | 03553426001 | |
| Qubit dsDNA High Sensitivity Assay Kit | Life Technologies | Q32854 | A fluorescence-based DNA quantitation assay; used in protocol steps 1.1, 9.1 and 10.1 |
| DynaMag-2 Magnet | Life Technologies | 12321D | |
| High Sensitivity DNA Kit | Agilent Technologies | 5067-4626 | |
| 2100 Bioanalyzer | Agilent Technologies | ||
| PhiX Control v3 | Illumina | FC-110-3001 | |
| HiSeq 2500 | Illumina | ||
| Pippin Prep | Sage Science | ||
| Qubit 2.0 Fluorometer | Life Technologies | Q32872 | |
| TruSeq SR Cluster Kit v3-cBot-HS | Illumina | GD-401-3001 | |
| TruSeq SBS Kit v3-HS | Illumina | FC-401-3002 | |
| TruSeq RNA Sample prep | Illumina | RS-122-2001 | Barcoded adapters used for multiplexing libraries; See Supplemental file for multiplexing protocol. |
| Microcentrifuge | |||
| Vortex Mixer | |||
| Dry Block Heater | |||
| Thermal Cycler | |||
| Water Bath | |||
| Gel electrophoresis system | |||
| Electrophoresis power supply | |||
| Gel doc | |||
| UV or blue light transilluminator |
参考文献
- Jones, P. A. Functions of DNA methylation: islands, start sites, gene bodies and beyond. Nat Rev Genet. 13 (7), 484-492 (2012).
- Barlow, D. P. Genomic imprinting: a mammalian epigenetic discovery model. Annual Review Of Genetics. 45, 379-403 (2011).
- Thiagarajan, R. D., Morey, R., Laurent, L. C. The epigenome in pluripotency and differentiation. Epigenomics. 6 (1), 121-137 (2014).
- Reik, W. Stability and flexibility of epigenetic gene regulation in mammalian development. Nature. 447 (7143), 425-432 (2007).
- Hartnett, L., Egan, L. J. Inflammation, DNA methylation and colitis-associated cancer. Carcinogenesis. 33 (4), 723-731 (2012).
- Smith, Z. D., Meissner, A. DNA methylation: roles in mammalian development. Nat Rev Genet. 14 (3), 204-220 (2013).
- Li, E., Bestor, T. H., Jaenisch, R. Targeted mutation of the DNA methyltransferase gene results in embryonic lethality. Cell. 69 (6), 915-926 (1992).
- Okano, M., Bell, D. W., Haber, D. A., Li, E. DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell. 99 (3), 247-257 (1999).
- Feinberg, A. P. Phenotypic plasticity and the epigenetics of human disease. Nature. 447 (7143), 433-440 (2007).
- Bock, C. Epigenetic biomarker development. Epigenomics. 1 (1), 99-110 (2009).
- Laird, P. W. The power and the promise of DNA methylation markers. Nat Rev Cancer. 3 (4), 253-266 (2003).
- How Kit, A., Nielsen, H. M., Tost, J. DNA methylation based biomarkers: practical considerations and applications. Biochimie. 94 (11), 2314-2337 (2012).
- Mikeska, T., Bock, C., Do, H., Dobrovic, A. DNA methylation biomarkers in cancer: progress towards clinical implementation. Expert Review Of Molecular Diagnostics. 12 (5), 473-487 (2012).
- Gyparaki, M. T., Basdra, E. K., Papavassiliou, A. G. DNA methylation biomarkers as diagnostic and prognostic tools in colorectal cancer. Journal of Molecular Medicine. 91 (11), 1249-1256 (2013).
- Figueroa, M. E., et al. DNA methylation signatures identify biologically distinct subtypes in acute myeloid leukemia. Cancer Cell. 17 (1), 13-27 (2010).
- Heyn, H., Mendez-Gonzalez, J., Esteller, M. Epigenetic profiling joins personalized cancer medicine. Expert review of Molecular Diagnostics. 13 (5), 473-479 (2013).
- Kulis, M., Esteller, M. DNA methylation and cancer. Advances in Genetics. 70, 27-56 (2010).
- Xiong, Z., Laird, P. W. COBRA: a sensitive and quantitative DNA methylation assay. Nucleic Acids Res. 25 (12), 2532-2534 (1997).
- Meissner, A., et al. Reduced representation bisulfite sequencing for comparative high-resolution DNA methylation analysis. Nucleic Acids Res. 33 (18), 5868-5877 (2005).
- Gu, H., et al. Preparation of reduced representation bisulfite sequencing libraries for genome-scale DNA methylation profiling. Nat Protoc. 6 (4), 468-481 (2011).
- Bock, C., et al. Quantitative comparison of genome-wide DNA methylation mapping technologies. Nat Biotechnol. 28 (10), 1106-1114 (2010).
- Harris, R. A., et al. Comparison of sequencing-based methods to profile DNA methylation and identification of monoallelic epigenetic modifications. Nat Biotechnol. 28 (10), 1097-1105 (2010).
- Boyle, P., et al. Gel-free multiplexed reduced representation bisulfite sequencing for large-scale DNA methylation profiling. Genome Biol. 13 (10), R92 (2012).
- Chatterjee, A., Rodger, E. J., Stockwell, P. A., Weeks, R. J., Morison, I. M. Technical considerations for reduced representation bisulfite sequencing with multiplexed libraries. Journal of Biomedicine & Biotechnology. 2012, 741542 (2012).
- Lee, Y. K., et al. Improved reduced representation bisulfite sequencing for epigenomic profiling of clinical samples. Biological Procedures Online. 16 (1), 1 (2014).
- Akalin, A., et al. Base-pair resolution DNA methylation sequencing reveals profoundly divergent epigenetic landscapes in acute myeloid leukemia. PLoS Genet. 8 (6), e1002781 (2012).
- Hatzi, K., et al. A Hybrid Mechanism of Action for BCL6 in B Cells Defined by Formation of Functionally Distinct Complexes at Enhancers and Promoters. Cell Reports. 4 (3), 578-588 (2013).
- Will, B., et al. Satb1 regulates the self-renewal of hematopoietic stem cells by promoting quiescence and repressing differentiation commitment. Nature Immunology. 14 (5), 437-445 (2013).
- Lu, C., et al. Induction of sarcomas by mutant IDH2. Genes Dev. 27 (18), 1986-1998 (2013).
- Kumar, R., et al. AID stabilizes stem-cell phenotype by removing epigenetic memory of pluripotency genes. Nature. 500 (7460), 89-92 (2013).
- Li, S., et al. An optimized algorithm for detecting and annotating regional differential methylation. BMC Bioinformatics. 14, S10 (2013).
- Patterson, K., Molloy, L., Qu, W., Clark, S. DNA methylation: bisulphite modification and analysis. Journal of Visualized Experiments. (56), 3170 (2011).
- Landan, G., et al. Epigenetic polymorphism and the stochastic formation of differentially methylated regions in normal and cancerous tissues. Nat Genet. 44 (11), 1207-1214 (2012).
- Yu, M., et al. Tet-assisted bisulfite sequencing of 5-hydroxymethylcytosine. Nat Protoc. 7 (12), 2159-2170 (2012).
- Goecks, J., Nekrutenko, A., Taylor, J., Galaxy, T. Galaxy: a comprehensive approach for supporting accessible, reproducible, and transparent computational research in the life sciences. Genome Biol. 11 (8), R86 (2010).
- Dorff, K. C., et al. GobyWeb: simplified management and analysis of gene expression and DNA methylation sequencing data. PLoS One. 8 (7), e69666 (2013).
- Roehr, J. T., Dodt, M., Ahmed, R., Dieterich, C. Flexbar − flexible barcode and adapter processing for next-generation sequencing platforms. MDPI Biology. 1 (3), 895-905 (2012).
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet.journal, North America. 17 (1), 10-12 (2011).
- Bolger, A. M., Lohse, M., Usadel, B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics. 30 (15), 2114-2120 (2014).
- Needleman, S. B., Wunsch, C. D. A general method applicable to the search for similarities in the amino acid sequence of two proteins. J Mol Biol. 48 (3), 443-453 (1970).
- Krueger, F., Andrews, S. R. Bismark: a flexible aligner and methylation caller for Bisulfite-Seq applications. Bioinformatics. 27 (11), 1571-1572 (2011).
- Li, H., et al. The Sequence Alignment/Map format and SAMtools. Bioinformatics. 25 (16), 2078-2079 (2009).
- Kent, W. J., et al. The human genome browser at UCSC. Genome Res. 12 (6), 996-1006 (2002).
- Thorvaldsdottir, H., Robinson, J. T., Mesirov, J. P. Integrative Genomics Viewer (IGV): high-performance genomics data visualization and exploration. Briefings in Bioinformatics. 14 (2), 178-192 (2013).
- Team, R. C. R. A language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria. ISBN 3-900051-07-0, (2012).
- Nestor, C., Ruzov, A., Meehan, R., Dunican, D. Enzymatic approaches and bisulfite sequencing cannot distinguish between 5-methylcytosine and 5-hydroxymethylcytosine in DNA. BioTechniques. 48 (4), 317-319 (2010).
- Huang, Y., et al. The behaviour of 5-hydroxymethylcytosine in bisulfite sequencing. PLoS One. 5 (1), e8888 (2010).
- Yu, M., et al. Base-resolution analysis of 5-hydroxymethylcytosine in the mammalian genome. Cell. 149 (6), 1368-1380 (2012).
- Song, C. X., et al. Genome-wide profiling of 5-formylcytosine reveals its roles in epigenetic priming. Cell. 153 (3), 678-691 (2013).
- Ito, S., et al. Tet proteins can convert 5-methylcytosine to 5-formylcytosine and 5-carboxylcytosine. Science. 333 (6047), 1300-1303 (2011).
- Akalin, A., et al. methylKit: a comprehensive R package for the analysis of genome-wide DNA methylation profiles. Genome Biol. 13 (10), R87-1186 (2012).
- Stockwell, P. A., Chatterjee, A., Rodger, E. J., Morison, I. M. DMAP: Differential Methylation Analysis Package for RRBS and WGBS data. Bioinformatics. 30 (13), 1814-1822 (2014).
- Sun, D., et al. MOABS: model based analysis of bisulfite sequencing data. Genome Biol. 15 (2), R38 (2014).
- Bock, C. Analysing and interpreting DNA methylation data. Nat Rev Genet. 13 (10), 705-719 (2012).
- Rivera, C. M., Ren, B. Mapping human epigenomes. Cell. 155 (1), 39-55 (2013).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved