
このコンテンツを視聴するには、JoVE 購読が必要です。 サインイン又は無料トライアルを申し込む。
Method Article
小胞体とミトコンドリアの相互作用の研究によって、
要約
ここでは、固定された細胞内小胞体とミトコンドリアの間に内因性の相互作用を視覚化し、高感度で定量化するための手順について説明します。プロトコルは、ミトコンドリア関連膜界面での75 /電位依存性アニオンチャネル/シクロフィリンD複合体イノシトール1,4,5-三リン酸受容体/グルコース調節タンパク質を標的と現場での近接連結アッセイに最適化されています。
要約
Structural interactions between the endoplasmic reticular (ER) and mitochondrial membranes, in domains known as mitochondria-associated membranes (MAM), are crucial hubs for cellular signaling and cell fate. Particularly, these inter-organelle contact sites allow the transfer of calcium from the ER to mitochondria through the voltage-dependent anion channel (VDAC)/glucose-regulated protein 75 (GRP75)/inositol 1,4,5-triphosphate receptor (IP3R) calcium channeling complex. While this subcellular compartment is under intense investigation in both physiological and pathological conditions, no simple and sensitive method exists to quantify the endogenous amount of ER-mitochondria contact in cells. Similarly, MAMs are highly dynamic structures, and there is no suitable approach to follow modifications of ER-mitochondria interactions without protein overexpression. Here, we report an optimized protocol based on the use of an in situ proximity ligation assay to visualize and quantify endogenous ER-mitochondria interactions in fixed cells by using the close proximity between proteins of the outer mitochondrial membrane (VDAC1) and of the ER membrane (IP3R1) at the MAM interface. Similar in situ proximity ligation experiments can also be performed with the GRP75/IP3R1 and cyclophilin D/IP3R1 pairs of antibodies. This assay provides several advantages over other imaging procedures, as it is highly specific, sensitive, and suitable to multiple-condition testing. Therefore, the use of this in situ proximity ligation assay should be helpful to better understand the physiological regulations of ER-mitochondria interactions, as well as their role in pathological contexts.
概要
ミトコンドリアと小胞体(ER)は、細胞内の独立した小器官ではありませんが、ミトコンドリア関連小胞体膜(MAM)のように定義された接触部位での構造的および機能的に相互作用します。実際には、MAMSは、ERおよびミトコンドリアの膜が密接に両側からのタンパク質間の相互作用を可能にする、並置された領域に対応します。それにもかかわらず、これらの細胞小器官の膜は、これらの領域内に融合していないので、彼らは別々のエンティティを維持します。 MAMSは、エネルギー代謝や細胞の生存1-3に影響を与え、カルシウム(Ca 2+)で重要な役割を果たし、ミトコンドリアへのERからのリン脂質の転送。
ERとミトコンドリアの間の関連付けは、第1の電子顕微鏡で1970年代の可視化しました。それ以来、透過型電子顕微鏡4,5、電子線トモグラフィー6,7またはERおよびミトコンドリア特異的蛍光体の免疫局在S /蛍光タンパク質は、8は、古典的ER-ミトコンドリア相互作用を研究するために使用しました。 MAMの分析のための別の有用なツールは、細胞下分画の使用に基づいています。これは、パーコール勾配9に接続された差動超遠心分離によってMAM画分の単離を可能にします。しかし、最終生成物は、濃縮されたMAM画分ではなく、純粋な画分を含んでいます。要するに、これらの戦略は特に敏感および/または定量的ではない、と彼らは大規模なスクリーニングに容易に適していません。あるいは、薬物誘導性蛍光インターオルガネラリンカーを用いて遺伝的アプローチが浮上しているが、それらは、タンパク質10の内因性発現レベルでの細胞小器官の相互作用の分析を可能にしません。
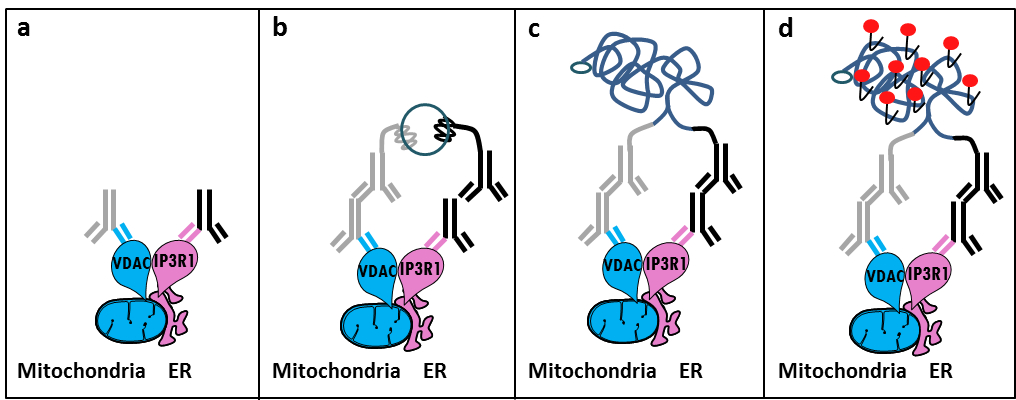
MAM 11でIP3R / GRP75 / VDAC複合体のSzabadkaiの発見に基づいて、我々はER-ミトコンドリアの相互作用を分析するための定量法を開発しました。私たちは、 その場に近接して使用さligatiアッセイに、固定された細胞12内MAM界面でのCa 2+ -channeling複雑に関与する2つの細胞小器官表面タンパク質をVDAC1とIP3R1の間の相互作用を検出し、定量化します。簡単に言えば、我々は、ミトコンドリア外膜(マウス抗VDAC1一次抗体)でVDAC1をプローブし、ER膜(ウサギ抗IP3R1一次抗体)でIP3R1(図1、パネルA)。そして、分析によれば、我々は、相補的なオリゴヌクレオチド拡張にコンジュゲートされた抗マウス及び抗ウサギIgG(マウスおよびウサギ近接ライゲーションアッセイプローブ)の両方を添加しました。 2標的タンパク質が40nm以下の距離にある場合、オリゴヌクレオチドは、環状DNA鋳型(図1、パネルB)の形成を可能にするために、続いて添加コネクタオリゴとハイブリダイズすることができます。この環状DNA分子は、共有結合(図1、パネルC)の近接プローブの1つに取り付けられた一本鎖DNA産物を作成し、連結し、増幅され NG。 MAM界面におけるERおよびミトコンドリアとの間の距離が10nmから25nmの6の範囲にあるので、近接ライゲーションおよび増幅によるテキサスレッド標識オリゴヌクレオチドプローブ(図1、パネルDのハイブリダイゼーション後に検出をもたらす、行うことができます)。各蛍光ドットは、このように、個々の細胞におけるその場での ER-ミトコンドリアの相互作用の定量化を可能にする、VDAC1 / IP3R1間の相互作用を表しています。

図1: その場近接ライゲーションアッセイでによって小胞体、ミトコンドリア相互作用の検出の概略図。 A)VDAC1およびMAMの界面に近接してそのエピトープに結合することができるIP3R1に対するウサギ一次抗体に対するマウス一次抗体、b)は、近接ライゲーションプローブ対の付加マウス及びウサギIgGに対して向け。これらのプローブは、コネクタオリゴのライゲーションのためのテンプレートを形成することができるDNA鎖を添付しています。 c)のライゲーション後に形成された環状DNA鎖を増幅することができると、d)、テキサスレッドで標識されたオリゴヌクレオチドを用いて、蛍光ドットとして顕微鏡によって可視化しました。 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}
その場での近接連結アッセイ実験で同様にCypDがMAMの界面でIP3R / GRP75 / VDAC複合体と相互作用することが示されたことを考慮すると、抗体のGRP75 / IP3R1のペアだけでなく、シクロフィリンD(CypD)/ IP3R1抗体を用いて行うことができます12-14。
Access restricted. Please log in or start a trial to view this content.
プロトコル
ソリューションの調製
- PBSの14.5ミリリットルで37%ホルムアルデヒドの5.5ミリリットルを希釈することにより、PBS中10%ホルムアルデヒド(低塩)を準備します。 PBS 50ml中にグリシンを3.8gを溶解させ、1 Mグリシン、pHが8.0を調製します。 1×PBS中の100mMグリシンを得るために、この溶液を希釈。
- 1×PBS中の0.1%のTriton-X100を準備します。 3.0 M塩化ナトリウムおよび25℃でpH7.0の脱イオン水緩衝液中で調製0.30 Mクエン酸三ナトリウムを用いて20倍生理食塩水クエン酸ナトリウム(SSC)を準備します。 1Xにこのバッファーを希釈し、脱イオン水を使用して、0.01x。
細胞の2.固定
注:我々は、この研究でのHuh7肝癌細胞株を使用したが、この方法は、他の接着細胞の培養にも適用可能です。
- コーティングされていない35 mmのガラスボトムディッシュ15万個の細胞で(10%ウシ胎児血清および0.01%ペニシリン - ストレプトマイシンストックを補充したDMEM 1グラム/ Lのグルコースで培養し、)プレートHuh7細胞。一次電池で作業するとき文化、コラーゲンコートディッシュを使用します。
- 翌日、培地を除去します。 PBSを吸引1mlで細胞を洗浄。ホルムアルデヒド1mlの10%添加して細胞を固定し、攪拌下、室温(RT)で10分間インキュベートします。
- 1 Mグリシンの1ミリリットルで反応を停止し、回転させることにより混合します。停止反応液を除去し、1×PBSを1mlを加えることによって細胞を洗浄します。回転と吸引によって攪拌します。細胞を100mMグリシン、1 mlを加え、攪拌下、室温で15分間、それらをインキュベートし、その後吸引。
注:プロトコルはここで停止することができ、次の手順では、別の日に延期することができます。その場合には、100 mMグリシンの1ミリリットルを追加し、必要に応じて、4℃で維持。
細胞の3透過処理
- 、1×PBS中0.1%のTriton-X100 1 mlを加え、攪拌下、室温で15分間インキュベートし、その後吸引。一次細胞culturで作業するときに20分 - このインキュベーション時間は15にまで増加させることができましたエス( 例えば、一次マウス肝細胞)。 1×1mlのPBSを加えることによって細胞を洗浄します。吸引。
4.ブロッキング
- (キットにより提供される)を各試料にブロッキング溶液の40μlのを追加します。このボリュームは、サンプルをカバーするために増加させることができます。加湿チャンバー内で37℃で30分間皿をインキュベートします。
- 料理のオフブロッキング溶液をタップします。これは再現性に影響を与えるように、スライドごとに同等の残留ボリュームを取得してみてください。サンプルは乾燥させてはいけません!
5.一次抗体
- 1×PBS中で一次抗体を希釈し、皿(VDAC1マウス抗体:1/100、IP3R1ウサギ抗体:1/500)にソリューションを追加します。代替的に、GRP75またはCypD抗体(1/500で使用し、両方のマウス抗体)は、代わりにVDAC1を用いることができます。
- 4℃で湿度チャンバ内で一晩インキュベートします。 0.01%のTween(TBS-T)でトリス緩衝生理食塩水を使用して、スライドを2回洗浄します。
- 近接ライゲーションアッセイプローブは、キットが提供されます。一次抗体の種類に応じたプローブを選択してください。
- 抗体希釈液で5:2近接ライゲーションアッセイプローブ1を準備します。混合物を室温で20分間静置します。希釈された近接連結アッセイプローブ溶液を追加します。 37℃で1時間予備加熱湿度チャンバ内で皿をインキュベートします。 TBS-Tで皿を2回洗浄します。
7.ライゲーション
- in situ検出試薬テキサスレッドキットに使用します。
- 高純度水に5とよく混ぜる:(キットにより提供される)5倍ライゲーション株式1を希釈します。 (キットにより提供される)1×ライゲーション溶液1時40分、ボルテックスでリガーゼを希釈します。すぐにサンプルに添加する前までリガーゼを追加するのを待ちます。
- 各サンプル(35 mmのガラスボトムディッシュのための40μl)にこのソリューションを追加し、予備加熱湿度チャムでスライドをインキュベート37℃で30分間のBER。 TBS-Tでスライドを2回洗浄します。
8.増幅
注:注意して、敏感な試薬を点灯。
- 高純度の水に5:1(キットにより提供される)5倍の増幅ストックを希釈します。凍結ブロック(-20℃)を使用して、冷凍庫からポリメラーゼを削除します。 (キットにより提供)ポリメラーゼの1x増幅溶液と渦で1:80に希釈します。混合物を使用する前にすぐにポリメラーゼを追加します。
- 各サンプル(35 mmのガラスボトムディッシュのための40μl)に、このソリューションを追加します。 37℃で100分間予備加熱湿度チャンバ内のスライドをインキュベートします。スライドのオフ増幅ポリメラーゼソリューションをタップします。
9.最後の洗浄
- 2分間の1×SSC洗浄バッファーでスライドを洗浄します。 2分間0.01x SSC洗浄バッファーでスライドを洗浄します。料理は暗闇の中で室温で乾燥させます。
イメージングのための10の準備
- 空気の泡がカバースリップの下に巻き込まれなかっ取得することを保証し、(水性および固化しない)DAPIを含む封入剤の最小量を使用してスライドをマウントします。マニキュアは、縁部をシールするために使用することができます。 (:594 nmの発光:624 nmで、倍率:63X励起)蛍光または共焦点顕微鏡を用いて分析する前に、約15分間待ちます。
- 撮影後、暗所で-20℃でスライドを保存します。
Access restricted. Please log in or start a trial to view this content.
結果
このプロトコルを使用して、我々の経験に基づいて、我々は安全に固定された細胞におけるER-ミトコンドリア間相互作用の可視化と定量化のため、この方法をお勧めすることができます。代表的な画像はその場での抗体のいくつかのペアを使用したHuH7肝細胞癌細胞株における近接ライゲーションアッセイ可視化ER-ミトコンドリア相互作用は、示されています。...
Access restricted. Please log in or start a trial to view this content.
ディスカッション
Collectively, our studies indicate that the in situ proximity ligation assay is truly a relevant strategy to follow and quantify endogenous ER-mitochondria interactions in fixed cells, without the need for using organelle-specific fluorophores or fluorescent proteins. The specific use of VDAC1/IP3R1 antibodies has been adapted to study ER-mitochondria interactions in HuH7 cells. However, alternative isoforms of VDAC and IP3R may be used, depending on the cell type. In this case, antibodies need to be validated b...
Access restricted. Please log in or start a trial to view this content.
開示事項
The authors declare that they have no competing financial interests.
謝辞
我々は、プロトコルを最適化し、検証するために貢献し、私たちの研究室でのすべての人々に感謝します。この作品は、INSERMと国立研究機関(ANR-09-JCJC-0116とANR-11-BSV1-033-02)によってサポートされていました。 ETは、高等教育と研究のフランスの省から研究フェローシップによって博士号中にサポートされていました。
Access restricted. Please log in or start a trial to view this content.
資料
| Name | Company | Catalog Number | Comments |
| Formaldehyde | Sigma | F-8775 | |
| Glycine | Sigma | G-8898 | |
| Triton | Sigma | T8532 | |
| 35 mm Glass bottom culture dishes | MatTeK corporation | P35G-0-14-C | |
| Blocking solution | Sigma | DUO-92004 or DUO-92002 | provided in the Duolink PLA probes, Sigma |
| VDAC1 antibody | Abcam | ab14734 | |
| IP3R1-H80 antibody | Santa Cruz | sc28614 | |
| CypD antibody | Abcam | ab110324 | |
| Grp75 antibody | Santa Cruz | sc13967 | |
| TBS 10x | euromedex | ET220 | Dilute to obtain 1x |
| Tween 100x | euromedex | 2001-B | dilute in TBS to obtain 0,01% |
| PLA Probes Mouse MINUS | Sigma | DUO-92004 | Duolink, Sigma |
| PLA Probes Rabbit PLUS | Sigma | DUO-92002 | Duolink, Sigma |
| Duolink detection reagents red | Sigma | DUO-92008 | Duolink, Sigma |
| Ligation solution | Sigma | DUO-92008 | Part of the Duolink detection reagents red, Sigma |
| Ligase | Sigma | DUO-92008 | Part of the Duolink detection reagents red, Sigma |
| Amplification solution | Sigma | DUO-92008 | Part of the Duolink detection reagents red, Sigma |
| Polymerase | Sigma | DUO-92008 | Part of the Duolink detection reagents red, Sigma |
| Duolink Mounting Medium | Sigma | DUO80102 | Duolink, Sigma |
| Softwares | |||
| Blob-finder software | BlobFinder is a freely distributed software that can perform calculations on cells from fluorescence microscopy images. This software can be downloaded for free from The Centre for Image Analysis at Uppsala University who have developed the software and the work was supported by the EU FP6 Project ENLIGHT and Olink Bioscience. http://www.cb.uu.se/~amin/BlobFinder/index_files/Page430.htm | ||
| ImageJ software | Can be downloaded for free from: http://rsb.info.nih.gov/ij/download.html | ||
参考文献
- Bravo-Sagua, R., et al. Organelle communication: signaling crossroads between homeostasis and disease. The international journal of biochemistry & cell biology. 50, 55-59 (2014).
- Giorgi, C., et al. Mitochondria-associated membranes: composition, molecular mechanisms, and physiopathological implications. Antioxidants & redox signaling. 22, 995-1019 (2015).
- Phillips, M. J., Voeltz, G. K. Structure and function of ER membrane contact sites with other organelles. Nature reviews. Molecular cell biology. 17, 69-82 (2016).
- Cosson, P., et al. The RTM resistance to potyviruses in Arabidopsis thaliana: natural variation of the RTM genes and evidence for the implication of additional genes. PLoS One. 7, 39169(2012).
- Mannella, C. A. Structure and dynamics of the mitochondrial inner membrane cristae. Biochim Biophys Acta. 1763, 542-548 (2006).
- Csordas, G., et al. Structural and functional features and significance of the physical linkage between ER and mitochondria. The Journal of cell biology. 174, 915-921 (2006).
- Mannella, C. A., Buttle, K., Rath, B. K., Marko, M. Electron microscopic tomography of rat-liver mitochondria and their interaction with the endoplasmic reticulum. Biofactors. 8, 225-228 (1998).
- Rizzuto, R., et al. Close contacts with the endoplasmic reticulum as determinants of mitochondrial Ca2+ responses. Science. 280, 1763-1766 (1998).
- Wieckowski, M. R., Giorgi, C., Lebiedzinska, M., Duszynski, J., Pinton, P. Isolation of mitochondria-associated membranes and mitochondria from animal tissues and cells. Nat Protoc. 4, 1582-1590 (2009).
- Csordas, G., et al. Imaging interorganelle contacts and local calcium dynamics at the ER-mitochondrial interface. Mol Cell. 39, 121-132 (2010).
- Szabadkai, G., et al. Chaperone-mediated coupling of endoplasmic reticulum and mitochondrial Ca2+ channels. J Cell Biol. 175, 901-911 (2006).
- Tubbs, E., et al. Mitochondria-associated endoplasmic reticulum membrane (MAM) integrity is required for insulin signaling and is implicated in hepatic insulin resistance. Diabetes. 63, 3279-3294 (2014).
- Paillard, M., et al. Depressing Mitochondria-Reticulum Interactions Protects Cardiomyocytes From Lethal Hypoxia-Reoxygenation Injury. Circulation. 128, 1555-1565 (2013).
- Rieusset, J., et al. Disruption of calcium transfer from ER to mitochondria links alterations of mitochondria-associated ER membrane integrity to hepatic insulin resistance. Diabetologia. 59, 614-623 (2016).
- Allalou, A., Wahlby, C. BlobFinder, a tool for fluorescence microscopy image cytometry. Computer methods and programs in biomedicine. 94, 58-65 (2009).
- Theurey, P., et al. Mitochondria-associated endoplasmic reticulum membranes allow adaptation of mitochondrial metabolism to glucose availability in the liver. Journal of molecular cell biology. , (2016).
- de Brito, O. M., Scorrano, L. Mitofusin 2 tethers endoplasmic reticulum to mitochondria. Nature. 456, 605-610 (2008).
- Soderberg, O., et al. Direct observation of individual endogenous protein complexes in situ by proximity ligation. Nature methods. 3, 995-1000 (2006).
- De Pinto, V., Messina, A., Lane, D. J., Lawen, A. Voltage-dependent anion-selective channel (VDAC) in the plasma membrane. FEBS letters. 584, 1793-1799 (2010).
- Kaul, S. C., Taira, K., Pereira-Smith, O. M., Wadhwa, R. Mortalin: present and prospective. Experimental gerontology. 37, 1157-1164 (2002).
Access restricted. Please log in or start a trial to view this content.
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved