
Method Article
Окрашивание и визуализация высокого разрешения трехмерных органоидных и сфероидных моделей
* Эти авторы внесли равный вклад
В этой статье
Резюме
Здесь мы предоставляем подробные, надежные и взаимодополняющие протоколы для выполнения окрашивания и визуализации субклеточного разрешения фиксированных трехмерных моделей клеточных культур в диапазоне от 100 мкм до нескольких миллиметров, что позволяет визуализировать их морфологию, состав клеточного типа и взаимодействия.
Аннотация
Трехмерные (3D) модели клеточных культур in vitro, такие как органоиды и сфероиды, являются ценными инструментами для многих применений, включая разработку и моделирование заболеваний, открытие лекарств и регенеративную медицину. Чтобы в полной мере использовать эти модели, крайне важно изучить их на клеточном и субклеточном уровнях. Однако характеристика таких моделей 3D-культур клеток in vitro может быть технически сложной и требует специальных знаний для выполнения эффективного анализа. Здесь в этой статье представлены подробные, надежные и взаимодополняющие протоколы для выполнения окрашивания и визуализации субклеточного разрешения фиксированных моделей 3D-культур клеток in vitro в диапазоне от 100 мкм до нескольких миллиметров. Эти протоколы применимы к широкому спектру органоидов и сфероидов, которые различаются по своим клеткам происхождения, морфологии и условиям культивирования. От сбора 3D-структуры до анализа изображений, эти протоколы могут быть завершены в течение 4-5 дней. Вкратце, 3D-структуры собираются, фиксируются и затем могут быть обработаны либо путем встраивания парафина и гистологического / иммуногистохимического окрашивания, либо непосредственно иммуномаркированы и подготовлены к оптической очистке и 3D-реконструкции (глубина 200 мкм) с помощью конфокальной микроскопии.
Введение
За последние десятилетия достижения в биологии стволовых клеток и технологиях 3D-культур in vitro ознаменовали революцию в биологии и медицине. Клеточные модели более высокой сложности в 3D стали очень популярными, поскольку они позволяют клеткам расти и взаимодействовать с окружающей внеклеточной структурой, тесно переименовывая аспекты живых тканей, включая их архитектуру, организацию и взаимодействия клеток или даже диффузионные характеристики. Таким образом, 3D-модели клеточных культур могут дать уникальную информацию о поведении клеток в развивающихся или больных тканях in vitro. Органоиды и сфероиды являются многоклеточными 3D-структурами, от нескольких микрометров до миллиметров, и являются наиболее заметными 3D-структурами in vitro. Оба могут культивироваться в поддерживающем каркасе, включая (i) гидрогели, полученные от животных (экстракт базальной мембраны, коллаген), растений (альгинат/агароза) или синтезированные из химических веществ, или (ii) инертные матрицы, содержащие поры для содействия пролиферации и росту клеток.
Органоиды и сфероиды также могут развиваться без присутствия поддерживающего каркаса, полагаясь на клетки для самособирания в кластеры. Это опирается на различные методы, такие как использование неадгезивных материалов для ингибирования прикрепления клеток, поверхностного натяжения и гравитационной силы (например, методы подвешивания капель) или постоянное круговое вращение сосудов (например, культура спиннера). Во всех случаях эти методы облегчают взаимодействие клетка-клетка и клетка-матрица для преодоления ограничений традиционной однослойной клеточной культуры1. Термины «органоиды» и «сфероиды» использовались взаимозаменяемо в прошлом, но между этими двумя моделями 3D-культур клеток есть ключевые различия. Органоиды представляют собой in vitro 3D клеточные кластеры, полученные из плюрипотентных стволовых клеток или тканеспецифических стволовых клеток, в которых клетки спонтанно самоорганизуются в предшественников и дифференцированные типы клеток и которые повторяют, по меньшей мере, некоторые функции интересующего органа2. Сфероиды содержат более широкий спектр многоклеточных 3D-структур, сформированных в условиях несоблюдения, и могут возникать из большого разнообразия типов клеток, таких как увековеченные клеточные линии или первичные клетки3. Следовательно, присущие их внутреннему происхождению стволовых клеток, органоиды имеют более высокую склонность к самосборке, жизнеспособности и стабильности, чем сфероиды.
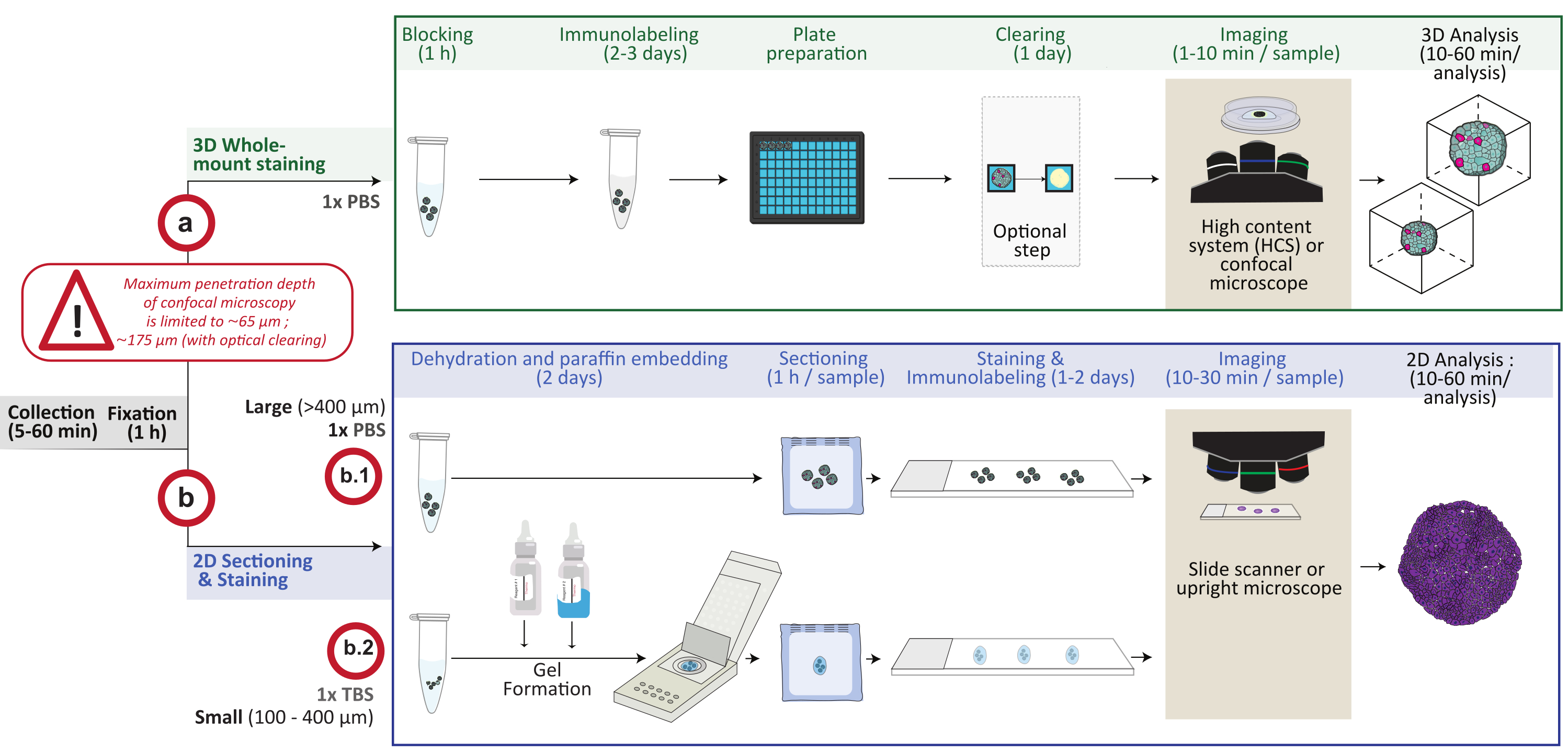
Тем не менее, по сути, эти две модели являются 3D-структурами, состоящими из нескольких клеток, и методы, разработанные для их изучения, таким образом, очень похожи. Например, мощные подходы к визуализации на уровне одноклеточного разрешения необходимы для исследования клеточной сложности как органоидов, так и сфероидов. Здесь, обобщая опыт этой группы и лидеров в области органоидов4, в этой статье описываются подробные процедуры для выполнения двумерного (2D) и 3D-окрашивания, визуализации и анализа клеточного и субклеточного состава и пространственной организации органоидов и сфероидов в диапазоне от 100 мкм до нескольких миллиметров. Действительно, эта процедура представляет два различных и взаимодополняющих типа окрашивания и получения изображений для анализа большого разнообразия размеров и типов моделей 3D-культур клеток in vitro. Использование одного (3D-анализ всего монтажа) или другого (2D-анализ секций) будет зависеть от изученной модели и искомых ответов. 3D-анализ с помощью конфокальной микроскопии может, например, применяться для визуализации клеток в 3D-культуре глубиной до 200 мкм, независимо от общего размера 3D-структуры, тогда как анализ 2D-секций дает представление о образцах любого размера, хотя и на 2D-уровне. Эта процедура была успешно применена к различным органоидам 4,5 и сфероидам, полученным из клеток человека и мышей, происходящих из различных эмбриональных зародышевых слоев. Обзор процедуры показан на рисунке 1. Указаны основные этапы, взаимосвязи между ними, решительные шаги, ожидаемые сроки.

Рисунок 1: Схематический обзор процедуры. Модели 3D-культур клеток in vitro собираются и фиксируются, затем либо подготавливаются для 3D-окрашивания всего крепления (вариант a), либо встраиваются в парафин для 2D-секционирования и окрашивания (вариант b). Для экспериментов по окрашиванию цельного 3D-крепления фиксированные 3D-структуры иммуномепонируются после этапа фиксации. Дополнительный этап оптической очистки может быть выполнен для улучшения качества изображения и глубины оптической микроскопии за счет уменьшения рассеяния света во время обработки изображения. Изображения захватываются на инвертированный конфокальный микроскоп или конфокальную систему высокого содержания и анализируются с помощью соответствующего программного обеспечения. Для встраивания парафина 3D-структуры непосредственно обрабатывают (вариант b.1 для крупных структур ≥ 400 мкм) или включают в гель (b.2; небольшие структуры ≤ 400 мкм) для обезвоживания и встраивания парафина. Затем парафиновые блоки разрезают и окрашивают (гистологическое или иммунохимическое окрашивание). Изображения 2D-участков получаются на цифровом слайд-сканере или вертикальном микроскопе и анализируются на платформе анализа изображений с использованием быстрого цифрового количественного анализа. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
протокол
ПРИМЕЧАНИЕ: Следует ожидать потери ≤25% от первоначального количества 3D-структур на этапах, связанных с изменением реагентов и промывкой в следующей процедуре. Планируйте использовать окончательное количество не менее десяти 3D-структур размером от 100 до 500 мкм на каждое тестируемое условие для выполнения качественного и количественного анализа изображений. При необходимости для более крупных конструкций обрежьте концы наконечников пипетки по 1 мл, чтобы избежать разрушения конструкций. На всех этапах, если осаждение 3D-структуры слишком длинное, клетки можно осторожно раскручивать при 50 × г в течение 5 мин при комнатной температуре (RT). В зависимости от исследуемого вопроса следует учитывать преимущества/недостатки такого шага вращения, так как центрифугирование может поставить под угрозу форму 3D-структур. Избегайте вращения при >100 × г.
1. Сбор и фиксация 3D моделей клеточных культур
ПРИМЕЧАНИЕ: Будьте осторожны, чтобы не аспирировать 3D-структуры, которые будут только слабо прикреплены к стенке трубы.
- Сбор 3D-моделей клеточных культур, встроенных в матрицу
ПРИМЕЧАНИЕ: В этом разделе описывается восстановление 3D-структур, выращенных в каплях экстракта базальной мембраны из мышиной саркомы Энгельбрета-Холма-Роя (BME), но может быть адаптировано к другим матрицам. Смотрите обсуждение ключевых моментов, касающихся ECM.- Удалите культуральную среду из скважин, не нарушая 3D-матрицу. Предварительно обмазите внутреннюю и внешнюю часть наконечника пипетки объемом 1 мл белком ( далее называемым предварительно покрытым наконечником 1 мл ), опустив всю длину наконечника в 0,1% бычьего сывороточного альбумина (BSA) в фосфатно-буферном физиологическом растворе (PBS) (далее называемый PBS-BSA 0,1% раствором ) и пипетирование 1 мл этого раствора вверх и вниз дважды.
ПРИМЕЧАНИЕ: Это предварительное покрытие предотвратит прилипание клеток к наконечнику и минимизирует любые потери. - Предварительно обложите внутреннюю часть центрифужной (15 мл) пробирки белком (далее именуемой предварительно покрытой центрифужной трубкой ) путем многократного заполнения раствором PBS-BSA 0,1% и опорожнения трубки.
ПРИМЕЧАНИЕ: Это предотвратит прилипание клеток к трубке и минимизирует любые потери. - Используя предварительно покрытый наконечник 1 мл, тщательно повторно суспендируйте 3D-структуры скважины, используя 1 мл ледяного холода 1x PBS, и аккуратно перенесите суспензию, содержащую 3D-структуры, в предварительно покрытую центрифужную трубку.
- Осторожно добавьте 13 мл ледяного 1x PBS и позвольте 3D-структурам откладываться на льду в течение не менее 10 минут.
ПРИМЕЧАНИЕ: При необходимости вращайте в течение 5 мин при 50 × г при 4 °C. Избегайте вращения >100 × g, так как это поставит под угрозу форму 3D-структур. - Удалите супернатант. Используя предварительно покрытый наконечник 1 мл, аккуратно повторно суспендируйте 3D-структуры в 1 мл ледяного 1x PBS. Повторите этапы 1.1.4-1.1.5 для получения однородной гранулы без остатка 3D-матрицы.
ПРИМЕЧАНИЕ: Эффективное удаление матрицы зависит от типа матрицы, количества и размера 3D-структур и требует оптимизации для различных условий культуры. Для 3D-структур, выращенных в BME, восстановление после удаления матрицы обычно занимает 45-60 минут. - Используя предварительно покрытый наконечник объемом 1 мл, перенесите суспензию PBS объемом 1 мл 1x PBS, содержащую 3D-структуры, в предварительно покрытую центрифужную трубку объемом 1,5 мл и перейдите к разделу 1.3.
- Удалите культуральную среду из скважин, не нарушая 3D-матрицу. Предварительно обмазите внутреннюю и внешнюю часть наконечника пипетки объемом 1 мл белком ( далее называемым предварительно покрытым наконечником 1 мл ), опустив всю длину наконечника в 0,1% бычьего сывороточного альбумина (BSA) в фосфатно-буферном физиологическом растворе (PBS) (далее называемый PBS-BSA 0,1% раствором ) и пипетирование 1 мл этого раствора вверх и вниз дважды.
- Сбор плавающих 3D-моделей клеточных культур
- Используя предварительно покрытый наконечник 1 мл, тщательно соберите и перенесите 3D-структуры в предварительно покрытую 1,5 мл центрифужную трубку. Позвольте 3D-структурам отстояться или вращаться в течение 5 мин при 50 × г при RT.
- Удалите супернатант. Используя предварительно покрытый наконечник 1 мл, повторно суспендируйте 3D-структуры в 1 мл 1x PBS. Перейдите к разделу 1.3.
- Фиксация 3D моделей клеточных культур
- Позвольте 3D-структурам откладываться. Осторожно удалите супернатант; под вытяжным капюшоном аккуратно повторно суспендировать 3D-структуры в 1 мл формалина с помощью предварительно покрытого наконечника 1 мл.
ПРИМЕЧАНИЕ: Формалин содержит формальдегид, который опасен. Манипулируйте химическим веществом в химической вытяжке. Носите резиновые перчатки и защитные очки для глаз. - Инкубируйте 3D-структуры в течение 30 минут на RT.
ПРИМЕЧАНИЕ: 30-минутный этап фиксации формалином необходим для иммуноокрашивания широкого спектра 3D-структур (различных по размеру, форме и происхождению). Однако, в целом, более длительное время фиксации (>3 ч) лучше подходит для сохранения флуоресценции репортерных белков. - Позвольте 3D-структурам отстояться или вращаться в течение 5 мин при 50 × г при RT. Аккуратно удалите формалин и замените его 1 мл 1x PBS. Повторите этот шаг стирки в 1x PBS дважды. Храните образцы при температуре 4 °C и переходите к разделу 2 или разделу 3.
ПРИМЕЧАНИЕ: Здесь протокол может быть приостановлен, и ячейки могут поддерживаться при 4 ° C для длительного хранения (>1 год).
- Позвольте 3D-структурам откладываться. Осторожно удалите супернатант; под вытяжным капюшоном аккуратно повторно суспендировать 3D-структуры в 1 мл формалина с помощью предварительно покрытого наконечника 1 мл.
2.3D окрашивание, визуализация и анализ 3D-моделей клеточных культур
ПРИМЕЧАНИЕ: Поскольку органоиды слабо прикреплены к стенке трубки, обращайтесь с ними осторожно, так как все последующие изменения реагента могут привести к потере образца. Перед началом работы убедитесь в наличии правильных элементов управления для окрашивания. Положительным и отрицательным контролем могут быть клетки, в которых интересующий белок, как известно, либо сверхэкспрессирован, либо отсутствует, соответственно. Инкубировать образцы без первичного антитела, чтобы определить, обусловлен ли наблюдаемый сигнал неспецифическим связыванием вторичного антитела. Поскольку некоторые клетки, как правило, демонстрируют высокие уровни автофлуоресценции, используйте элементы управления, лишенные вторичных антител, чтобы определить, происходит ли наблюдаемая флуоресценция от фоновой автофлуоресценции. Иммуномаркировка и флуоресцентная репортерная визуализация могут быть объединены.
- Окрашивание всего 3D крепления
- Готовят раствор, блокирующий пермеабилизацию (ПБ), добавляя 1x PBS 0,1%-1% неионного поверхностно-активного вещества (см. Таблицу материалов), 1% диметилсульфоксида, 1% BSA и 1% ослиной сыворотки (или от животного, у которого были подняты вторичные антитела).
ПРИМЕЧАНИЕ: Тщательно оптимизируйте концентрацию неионного поверхностно-активного вещества в зависимости от локализации мишени: мембраны (0-0,5%), цитоплазмы (0,5-1%) и ядра (1%). Этот раствор можно хранить при 4°C до 1 месяца. BSA обычно хорошо работает на этапе блокировки, но в случае высокого фонового шума выполните эмпирический тест, чтобы получить наилучшие возможные результаты для данной комбинации антител. - Перенесите органоиды из центрифужной трубки объемом 1,5 мл в трубку объемом 0,5 мл с помощью предварительно покрытого наконечника объемом 1 мл. Дайте органоидам отстояться, аккуратно удалите 1x PBS и замените его 0,5 мл раствора PB. Инкубируют органоиды с мягким горизонтальным перемешиванием (30-50 об/мин) в течение 1 ч при РТ.
- Дайте органоидам отстояться, аккуратно удалите раствор ПБ и дважды промыть по 1 мл PBS-BSA 0,1% в течение 3 мин.
ПРИМЕЧАНИЕ: Ожидание в течение 3 мин позволяет структурам оседать на дне трубы. - Осторожно удаляют PBS-BSA 0,1% и добавляют 250 мкл первичного антитела, разведенного в соответствующей концентрации в растворе PB:1x PBS (1:10). Для приготовления 10 мл раствора PB:1x PBS (1:10) разводят 1 мл раствора PB в 9 мл 1x PBS. Инкубировать в течение 2-3 дней с мягким горизонтальным перемешиванием (30-50 об/мин) при 4 °C.
ПРИМЕЧАНИЕ: Соответствующее время инкубации антител имеет решающее значение для подходящего проникновения антител, поскольку 3D-структуры иногда могут достигать больших размеров. - Дайте органоидам отстояться, и осторожно удалите раствор первичного антитела. Промывайте 5x в PBS-BSA 0,1% в течение 3 мин на стирку, а затем 2x в 1 мл PBS-BSA 0,1% в течение 15 мин на стирку с мягким горизонтальным перемешиванием.
- Добавьте 250 мкл вторичного антитела, разведенного при 1:250 в растворе PB:1x PBS (1:10). Инкубировать в течение 24 ч при 4°C с мягким горизонтальным перемешиванием (30-50 об/мин). На этом этапе защитите образцы от света.
- Добавьте 250 мкл Hoechst 33342 (20 мкМ раствора), разбавленного при 1:1000 в растворе PB:1x PBS (1:10), и инкубируйте еще 2 ч при 4 °C с мягким горизонтальным перемешиванием (30-50 об/мин).
- Дайте органоидам отстояться и аккуратно удалите раствор, содержащий вторичное антитело + Hoechst 33342. Промывайте органоиды 5x в 1 мл 1x PBS в течение 3 мин на стирку, а затем 2x в 1 мл 1x PBS в течение 15 мин на стирку с мягким горизонтальным перемешиванием (30-50 об/мин).
ПРИМЕЧАНИЕ: Крайне важно тщательно промыть образцы, чтобы избежать фонового шума или потери сигнала. - Храните образцы в PBS при температуре 4 °C до получения изображения. Перейдите к разделу 2.2.
ПРИМЕЧАНИЕ: Здесь протокол может быть приостановлен, и образцы могут храниться при 4 °C в течение нескольких месяцев, защищенные от света.
- Готовят раствор, блокирующий пермеабилизацию (ПБ), добавляя 1x PBS 0,1%-1% неионного поверхностно-активного вещества (см. Таблицу материалов), 1% диметилсульфоксида, 1% BSA и 1% ослиной сыворотки (или от животного, у которого были подняты вторичные антитела).
- Пробоподготовка для конфокальной визуализации
- Используя предварительно покрытый наконечник 1 мл, осторожно перенесите органоиды в 50 мкл из 1x PBS на скважину в 96-луночной черной полистирольной микропластине. Перейдите к этапу 2.2.3 или разделу 2.3.
ПРИМЕЧАНИЕ: На этом этапе образец может быть защищен от света и храниться при температуре 4 °C в течение многих недель. - Очистка
ПРИМЕЧАНИЕ: Стадия очистки является необязательной и может быть использована либо для иммуномаркировки органоидов, либо для обнаружения эндогенной флуоресценции. Очистка может вызвать усадку 3D-структуры, но не изменяет общую морфологию, за исключением сферических однослойных органоидов с большими просветами4. Для этих кистозных органоидов пропустите этап очистки и выполните глубокую визуализацию тканей6.- Готовят 2,5 М раствора для очистки глицерин-фруктозы, содержащего 50% v/v глицерина, 11% v/v дистиллированной воды и 45% мас./v фруктозы, перемешивая на магнитной мешалке, по крайней мере, в течение ночи до полного солюбилизации и однородности раствора. Хранить при температуре 4 °C в темноте до 1 месяца.
- Удалите как можно больше 1x PBS, не прикасаясь к органоидам. Добавьте 200 мкл очищающего раствора, используя наконечник пипетки объемом 1 мл после удаления конца, и осторожно повторно суспендируйте, чтобы предотвратить образование пузырьков. Инкубируйте в RT в течение не менее 12 ч и переходите к разделу 3.
ПРИМЕЧАНИЕ: Поскольку очищающий раствор является вязким, небольшие объемы трудно обрабатывать. Чтобы облегчить обработку, убедитесь, что решение находится на RT, а пипетка медленно. Для оптимальной очистки позвольте образцу отстояться в растворе для очистки в течение не менее 24 ч перед визуализацией. Если 3D-структуры плавают во время захвата, выполните дополнительное вращение в течение 10 минут при <100 × г при RT или дайте больше времени (от одного до нескольких дней), чтобы дать им осадку. Протокол может быть приостановлен на этом этапе, прежде чем приступать к визуализации, если он защищен от света и хранится при 4 ° C (в течение недель) или -20 ° C (в течение нескольких месяцев).
- Используя предварительно покрытый наконечник 1 мл, осторожно перенесите органоиды в 50 мкл из 1x PBS на скважину в 96-луночной черной полистирольной микропластине. Перейдите к этапу 2.2.3 или разделу 2.3.
- Получение и анализ изображений
ПРИМЕЧАНИЕ: Технология секционирования изображений потребуется для изображения 3D-структур.- Используйте конфокальные микроскопы и предпочитайте погружные цели с более высокой числовой апертурой (NA) по сравнению с воздухом. Выбирайте цели увеличения (10x, 20x, 40x) в соответствии с размером 3D-структур, реконструкцией изображения (сшиванием) и решениями, используемыми для анализа.
- При выборе режима сбора учитывайте глубину фокусировки цели, используемой для определения шага для Укладки Z; обеспечивают оптимальный 3D-рендеринг.
ПРИМЕЧАНИЕ: Решения для анализа изображений различаются, и анализ необходимо будет адаптировать к используемому программному обеспечению. Например, этот протокол анализа был создан на программном обеспечении для анализа с высоким содержанием (см. Таблицу материалов и дополнительный рисунок 1 для деталей) и предоставляет данные о сегментации объекта, расчете свойств и выборе клеточной популяции в 3D-реконструированном объекте.
3. 2D-секционирование, окрашивание, визуализация и анализ 3D-моделей клеточных культур
ПРИМЕЧАНИЕ: 3D-модели клеточных культур различаются по размеру. Перейдите к разделу 3.1 или 3.2 для эффективного встраивания парафина (рисунок 2). Дайте достаточно времени для осаждения 3D-структуры перед любыми промывками и изменениями реагентов. Будьте осторожны, чтобы не аспирировать органоиды, которые будут плавать на дне трубки. Для встраивания парафина обратитесь к рисунку 2 для руководства.
- Парафиновое встраивание больших (Ø ≥ 400 мкм) 3D моделей клеточных культур
- За день до встраивания предваряют две колбы по 150 мл, наполненные парафином (парафиновые ванны), небольшую металлическую форму для встраивания на образец и тонкие щипцы до 65 °C.
- Используя предварительно покрытый наконечник 1 мл, осторожно перенесите органоиды в 1x PBS в стеклянную трубку с плоским дном с крышкой бутылки с политетрафторэтиленовой подкладкой. Дайте органоидам отстояться, аккуратно удалите 1x PBS и замените его 70% этанолом. Инкубировать не менее 30 мин.
- Дайте органоидам отстояться и осторожно удалите 70% этанола. Замените его 1 мл готового к употреблению раствора эозина Y. Проведите тюбиком и окрашивайте в течение не менее 30 минут. Осторожно удалите раствор эозина и обезвоживайте органоиды в трех последовательных промывках 1 мл 100% этанола в течение ~ 30 мин каждая.
ПРИМЕЧАНИЕ: Этанол, легковоспламеняющаяся и летучая жидкость, вызывает сильное раздражение глаз и дыхательных путей. Манипулируйте им в вытяжном капюшоне и носите защитные очки для глаз. - Осторожно удалите 100% этанол и под химическим капотом очистите органоиды в 3 последовательных промывках с 1 мл ксилола в течение ~ 30 мин каждая.
ПРИМЕЧАНИЕ: Ксилол является токсичным, жидким легковоспламеняющимся, пары которого могут вызывать раздражение. Манипулируйте им в вытяжном капюшоне. Избегайте прямого контакта с кожей и носите резиновые перчатки и защитные очки для глаз. - Под химическим капотом подготовьте белую микровижневую тканевую кассету, поместив кусок биопсийной прокладки (предварительно пропитанный ксилолом) внутрь одного из отсеков кассеты. Аккуратно перенесите 3D-структуры с помощью предварительно покрытой 2 мл пластиковой пипетки Пастера на биопсийную прокладку. Накройте их другой биопсийной прокладкой, пропитанной ксилолом, чтобы предотвратить движение органоидов, и закройте кассету.
- Если обработано несколько образцов, поместите кассету в ксилоловую ванну, чтобы дождаться дальнейшей обработки. После того, как все образцы будут перенесены в кассеты, поместите кассеты в предварительно приготовленную парафиновую ванну на 30 минут при 65 °C. Переложите кассеты в свежую предварительно приготовленную парафиновую ванну на ночь.
- После пропитки парафином возьмите предварительно застывшую форму для встраивания, и добавьте в нее нагретый парафин. Поместите биопсийную прокладку, содержащую 3D-структуры, в форму и осторожно перемешивайте ее, пока все органоиды не упадут на дно формы. Очень аккуратно разместите 3D-конструкции в центре формы, используя предварительно выровненные тонкие щипцы. Перейдите к разделу 3.3.
ПРИМЕЧАНИЕ: Будьте осторожны, чтобы не нарушить 3D-структуры с помощью щипцов; толкать, но не щипать их.
- Парафиновое встраивание малых (Ø ≤ 400 мкм) 3D моделей клеточных культур
- За день до встраивания предваряют две колбы по 150 мл, наполненные парафином (парафиновые ванны), небольшую металлическую форму для встраивания на образец и тонкие щипцы до 65 °C.
- Осторожно удалите 1x PBS из органоидной суспензии. Аккуратно выполните 3 промывки в 1 мл 1x Tris-буферного физиологического раствора (TBS). Удалите как можно больше 1x TBS, не прикасаясь к органоидам.
ПРИМЕЧАНИЕ: Будьте осторожны, чтобы не аспирировать образец. При необходимости выполните 5-минутное вращение при 50 х г на RT. Оставшиеся следы фосфата будут мешать следующим этапам, в частности, предотвращая полимеризацию геля. Поэтому не используйте решения PBS на любом этапе обработки. Для этого этапа коммерческий набор, содержащий кассеты, реагент No 1 (прозрачная жидкость) и реагент No 2 (цветная жидкость), использовался для облегчения процедуры, встраиваемой парафином, без потенциальной потери крошечных фрагментов (см. Таблицу материалов). Следуйте инструкциям по набору. Кассеты предварительно собраны с подкладочными бумагами и картонными вставками уже на месте. - Добавьте в тюбик 2 капли реагента No2 и аккуратно перемешайте, постукивая по тюбику. Добавьте 2 капли реагента No1 и снова перемешайте постукиванием, чтобы гель затвердел. Используя тонкие щипцы, извлеките гель из тюбика и поместите его в лунку кассеты.
- Под вытяжным шкафом обезвоживать образец, помещая кассету в последовательные ванны следующим образом (используйте колбы по 150 мл и используйте свежий этанол или ксилол для каждой ванны): этанол 70%, 30 мин; этанол 96%, 30 мин; этанол 100%, три промывки, 30 мин каждая; ксилол, три промывки, 30 мин каждая.
- Поместите кассеты в предварительно расплавленную парафиновую ванну на 30 мин при 65 °C и переложите их в свежую предварительно приготовленную парафиновую ванну на ночь. После пропитки парафином возьмите предварительно встраиваемую форму и добавьте в нее нагретый парафин. Откройте кассету, аккуратно вытесните гель мелкими щипцами и поместите гель, содержащий 3D-структуры, в центр встраиваемой формы. Перейдите к разделу 3.3.
- Общие шаги для встраивания парафина
- Аккуратно перенесите форму в холодную область, чтобы парафин затвердел тонким слоем, который будет поддерживать 3D-структуры в соответствующем положении. Добавьте тканевую кассету поверх формы и добавьте горячий парафин, чтобы покрыть эту пластиковую кассету. Удалите форму, как только она полностью затвердеет, и перейдите к разделу 3.4.
ПРИМЕЧАНИЕ: Парафиновые блоки могут храниться при комнатной температуре в течение многих лет.
- Аккуратно перенесите форму в холодную область, чтобы парафин затвердел тонким слоем, который будет поддерживать 3D-структуры в соответствующем положении. Добавьте тканевую кассету поверх формы и добавьте горячий парафин, чтобы покрыть эту пластиковую кассету. Удалите форму, как только она полностью затвердеет, и перейдите к разделу 3.4.

Рисунок 2: Обзор процедуры парафинового встраивания больших и малых моделей 3D-культур клеток in vitro.
(A) Стандартная процедура встраивания парафина. После фиксации и обезвоживания 3D-структуры окрашивают эозином для облегчения их визуализации (сверху и снизу слева). 3D-структуры аккуратно помещаются на биопсийную прокладку (синяя) в кассете с помощью пипетки Пастера объемом 2 мл (посередине). После пропитки парафином 3D-структуры осторожно опускают в жидкий парафин с помощью щипцов и осторожно перемешивают в биопсийной прокладке. Небольшие 3D-структуры теряются на этом этапе, так как они не могут быть освобождены от площадки (внизу справа: неудачное встраивание). Будут встроены только большие 3D-структуры (вверху справа: успешное встраивание). Наконечники стрел указывают на 3D-культуры. (B) Альтернатива стандартному протоколу встраивания парафина. После закрепления небольших 3D-структур используется коммерческий набор для поддержания клеток в геле и облегчения их переноса в форму после пропитки парафином (справа: успешное встраивание). Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
- Секционирование и окрашивание блоков
- Вырежьте 4 мкм сечения с использованием стандартного микротома и выполните стандартные гистологические и иммуногистохимические методы. Перейдите к разделу 3.5.
ПРИМЕЧАНИЕ: Для лучшего сцепления секций использовались специальные слайды (см. Таблицу материалов). Слайды могут храниться при комнатной температуре или при 4 °C в течение многих лет.
- Вырежьте 4 мкм сечения с использованием стандартного микротома и выполните стандартные гистологические и иммуногистохимические методы. Перейдите к разделу 3.5.
- Получение и анализ изображений
- Выполняйте визуализацию с помощью цифрового слайд-сканера или вертикального микроскопа и анализируйте данные с помощью платформы для быстрого цифрового количественного анализа, которая сообщает морфологические и мультиплексированные данные экспрессии на клеточной основе по всем разделам 3D-структуры (см. Дополнительный рисунок 2 для деталей).
ПРИМЕЧАНИЕ: Цель 20x регулярно используется этой группой.
- Выполняйте визуализацию с помощью цифрового слайд-сканера или вертикального микроскопа и анализируйте данные с помощью платформы для быстрого цифрового количественного анализа, которая сообщает морфологические и мультиплексированные данные экспрессии на клеточной основе по всем разделам 3D-структуры (см. Дополнительный рисунок 2 для деталей).
Результаты
Этот протокол предоставляет обзор критических этапов для 2D и 3D окрашивания цельным креплением, а также визуализацию и количественный анализ 3D-моделей клеточных культур (рисунок 3 и рисунок 4). Он применим к широкому спектру моделей 3D-клеточных культур - от сфероидов до органоидов из разных видов или тканей-хозяев - и позволяет получать точную и количественную информацию об архитектуре, организации клеток и взаимодействиях на клеточном и субклеточном уровнях (рисунок 3 и рисунок 4). Лабораториям может потребоваться оптимизировать 2D гистологические и иммуногистохимические методы и концентрации антител в соответствии с их собственными потребностями.
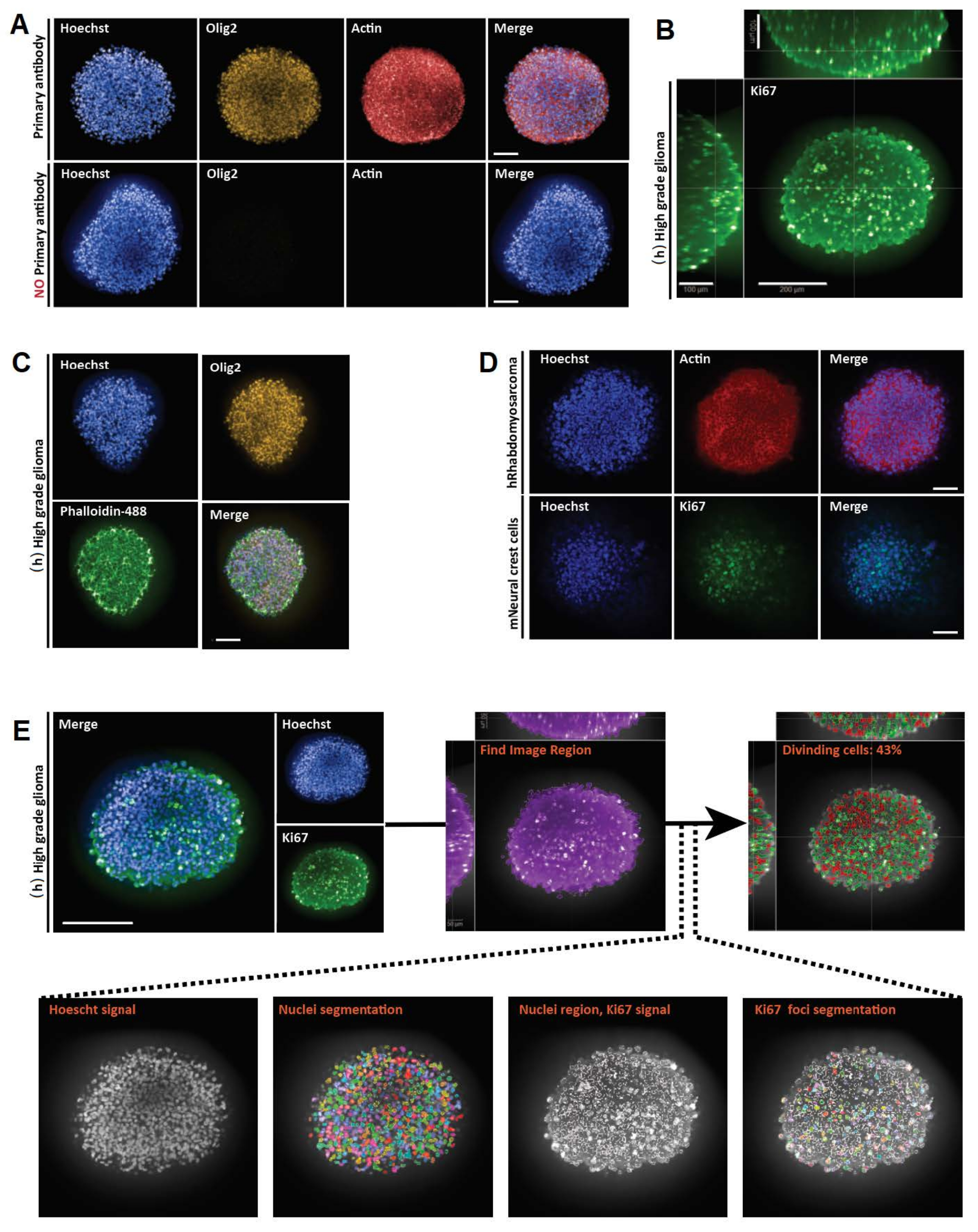
Оба метода дают ценную биологическую информацию. 3D-окрашивание цельного крепления и конфокальная микроскопия предоставляют визуальную информацию о клеточном составе и пространственном положении с полем глубиной до 200 мкм (рисунок 3B). Тем не менее, 2D-секционирование удобно для более крупных 3D-структур, чтобы выявить подробные клеточные морфологические признаки во всем разделе 3D-структур, которые в противном случае может быть сложно наблюдать на месте из-за рассеяния света, которое ставит под угрозу разрешение в более крупных образцах. Более того, оба метода могут предоставить количественные данные. Действительно, полученное разрешение позволяет применять алгоритмы клеточной и субклеточной сегментации для количественной оценки количества клеток и обнаружения наличия различных клеточных маркеров в разных клеточных подтипах (рис. 3F и фиг.4). Таким образом, методы визуализации, описанные здесь, являются воспроизводимыми, простыми и взаимодополняющими и представляют собой ценные инструменты для изучения клеточной гетерогенности.

Рисунок 3: Репрезентативные результаты для 3D-крепления, визуализации и анализа 3D и 2D оптических секций. (A) Конфокальные изображения человеческого (h) полноценного сфероида глиомы, культивируемого в течение недели и помеченного Hoechst (синий), Olig2 (желтый) и Actin (красный) (20x водный объектив). Для всех полученных изображений настройки микроскопа устанавливались с помощью положительного контроля (сверху), а затем отрицательный контроль визуализировали с использованием идентичных настроек для контроля отсутствия флуоресценции при отсутствии первичного антитела (снизу). (B) Ортогональное 3D-изображение окрашивания Ki67, выполненное в (h) полноценном сфероиде глиомы, культивируемом в течение недели (очистка от глицерин-фруктозы; 20x водный объектив, конфокальный). (C) Конфокальные изображения (h) полноценного сфероида глиомы, культивируемого в течение недели и помеченного Hoechst (синий), Olig2 (желтый) и Phalloidine-488 (зеленый) (очистка глицерин-фруктозы; 20x водный объектив). (D) Конфокальные изображения сфероидов человеческих (h) рабдомиосарком (вверху) и мышиных (m) клеток нервного гребня (снизу), культивируемых в течение недели и помеченных Hoechst (синий), Actin (красный) и Ki67 (зеленый), соответственно (очистка глицерин-фруктозы; 20x сухой объектив). (E) Конфокальные изображения (h) полноценного сфероида глиомы, культивируемого в течение недели и помеченного Hoechst (синий) и Ki67 (зеленый) (очистка глицерин-фруктозы; 40x водный объектив) (вверху слева). Сегментированные изображения на канале Hoechst и Ki67-положительных (+) ядерных областях на зеленом канале были сгенерированы с использованием программного обеспечения для анализа высокого содержания (см. Дополнительный рисунок 1 и Таблицу материалов) (внизу). На выходе указан процент ядер Ki67+ на сегментированную 3D-структуру (вверху справа). Шкала = 100 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

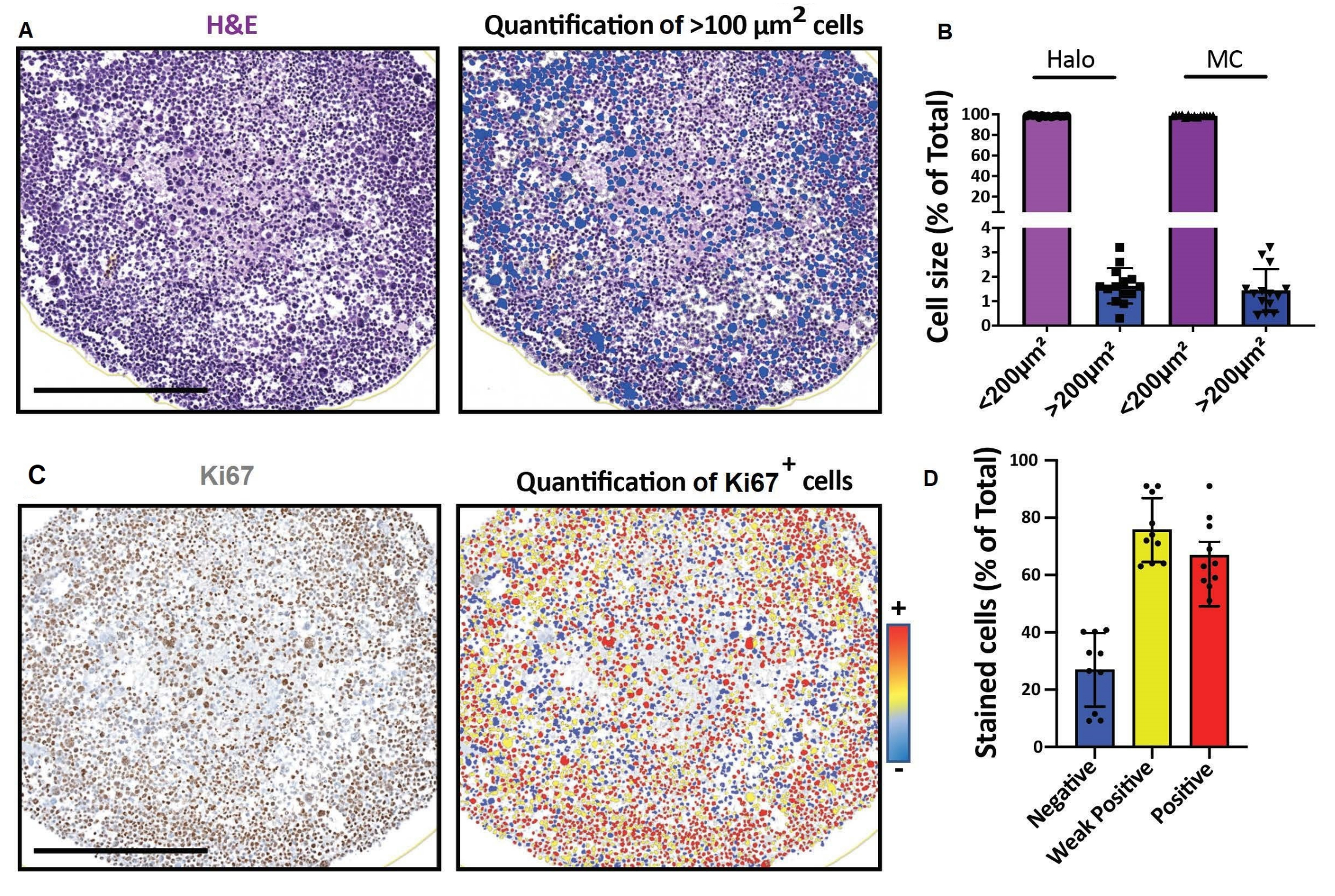
Рисунок 4: Репрезентативные результаты для визуализации и анализа 2D оптических сечений. (A, D) Изображения 2D-секций 3D-модели клеток (сфероиды рабдомиосаркомы человека, культивируемые в течение месяца), полученные с помощью цифрового слайд-сканера и проанализированные на платформе для быстрого цифрового количественного анализа. (A) Окрашивание H&E и обнаружение клеток в соответствии с их размером. Шкала = 500 мкм. (B) Гистограмма показывает процент клеток < 100мкм2 и > 100мкм2 , обнаруженных с помощью программного обеспечения для быстрого цифрового количественного анализа (слева: Halo) или ручного подсчета (справа: MC). (C) Окрашивание Ki67 и обнаружение клеток в соответствии с интенсивностью их сигнала 3,3'-диаминобензидина (DAB). Отрицательный (синий), слабоположительный (желтый), положительный (красный). Шкала = 500 мкм. (D) Гистограмма показывает процент Ki67-отрицательных, слабоположительных и положительных клеток. Сокращения: H&E = гематоксилин и эозин; MC = ручной подсчет. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Дополнительный рисунок 1: Обзор шагов в программном обеспечении для анализа изображений. Анализ основан на ассоциации строительных блоков. Каждый строительный блок, соответствующий функции - сегментации, вычисления, ассоциации, определения вывода - и предлагает несколько алгоритмов и выбор переменных для соответствия визуализируемому биологическому образцу. Программное обеспечение предоставляет несколько протоколов анализа RMS (Ready Made Solution), которые можно легко использовать и изменять. Интегрированные протоколы анализа изображений могут сохраняться, применяться к различным наборам данных и совместно использоваться пользователями. Вкратце, протокол анализа подразумевает последовательную сегментацию объектов: сфероид, ядра и, наконец, карманы Ki67 (A488). Затем подсчитывается средняя интенсивность карманов Ki67 для дальнейшей дискриминации положительных событий. Наконец, положительно отбираются ядра, охватывающие положительные карманы Ki67. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Дополнительный рисунок 2: Обзор этапов процедуры программного обеспечения для количественного анализа. Шаг 1. Загрузите файлы с помощью вкладки Исследования. Файлы будут открыты в разделе Действия с изображениями. Шаг 2. Откройте вкладку «Аннотации», затем нажмите «Действия слоя», чтобы создать новый слой вокруг структуры с помощью инструмента «Круг» на панели инструментов. Для некруглых структур вместо этого можно использовать инструмент пера. Шаг 3. Панель инструментов можно использовать для разработки аннотаций и визуализации количественной оценки с помощью  инструмента. Шаг 4. Откройте вкладку Анализ и выберите наилучшие условия для анализа образца (здесь может потребоваться несколько испытаний). Шаг 4.1. Используйте раздел Выбор пятна для настройки состояния окрашивания. В случае наличия нескольких пятен они могут быть добавлены и переименованы, а виртуальный цвет может быть изменен. Локализация детектирования может быть заданной – окрашивание ядерным или цитоплазмой. Шаг 4.2. Раздел Обнаружение ячеек используется для настройки обнаружения ячеек. Этот раздел будет наиболее важным для анализа. Раздел «Порог ядерного контраста» позволит обнаруживать все ядра. Внимание должно быть обращено на то, что если есть несколько размеров популяции, программное обеспечение может обнаружить несколько клеток вместо уникальной большой. Разделы «Ядерный размер» и «Агрессивность ядерной сегментации» могут использоваться для количественной оценки диапазонов популяции размеров клеток. Шаг 5. Описание выполнения анализа образцов. Выполните действия, показанные на рисунке. Раздел Слой аннотаций будет запускать настройку только на этом слайде. Количественную оценку можно визуализировать с помощью инструмента. Повторяйте шаги 4.1-5 до тех пор, пока не будет достигнута подходящая количественная оценка. Шаги 6-6.1. Эти шаги позволяют нарисовать фигуру с помощью программного обеспечения. Шаг 7. Количественная графика, полученная с помощью программного обеспечения, может быть сохранена. Шаг 8. Данные можно экспортировать. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
инструмента. Шаг 4. Откройте вкладку Анализ и выберите наилучшие условия для анализа образца (здесь может потребоваться несколько испытаний). Шаг 4.1. Используйте раздел Выбор пятна для настройки состояния окрашивания. В случае наличия нескольких пятен они могут быть добавлены и переименованы, а виртуальный цвет может быть изменен. Локализация детектирования может быть заданной – окрашивание ядерным или цитоплазмой. Шаг 4.2. Раздел Обнаружение ячеек используется для настройки обнаружения ячеек. Этот раздел будет наиболее важным для анализа. Раздел «Порог ядерного контраста» позволит обнаруживать все ядра. Внимание должно быть обращено на то, что если есть несколько размеров популяции, программное обеспечение может обнаружить несколько клеток вместо уникальной большой. Разделы «Ядерный размер» и «Агрессивность ядерной сегментации» могут использоваться для количественной оценки диапазонов популяции размеров клеток. Шаг 5. Описание выполнения анализа образцов. Выполните действия, показанные на рисунке. Раздел Слой аннотаций будет запускать настройку только на этом слайде. Количественную оценку можно визуализировать с помощью инструмента. Повторяйте шаги 4.1-5 до тех пор, пока не будет достигнута подходящая количественная оценка. Шаги 6-6.1. Эти шаги позволяют нарисовать фигуру с помощью программного обеспечения. Шаг 7. Количественная графика, полученная с помощью программного обеспечения, может быть сохранена. Шаг 8. Данные можно экспортировать. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Обсуждение
Клеточная культура является незаменимым инструментом для выявления фундаментальных биологических механизмов, участвующих в развитии тканей и органов, функции, регенерации и разрушении, а также заболеваниях. Хотя монослойная 2D-клеточная культура преобладала, недавние исследования сместились в сторону культур, генерирующих 3D-структуры, более отражающие клеточные реакции in vivo, в частности, благодаря дополнительной пространственной организации и контактам между клетками, которые влияют на экспрессию генов и клеточное поведение и, таким образом, могут обеспечить более прогностические данные7. Тем не менее, многие проблемы остаются, в том числе потребность в удобных для пользователя методах окрашивания и визуализации для детальной микроскопической визуализации и оценки сложных 3D-структур на клеточном и субклеточном уровнях. В этом контексте были предоставлены подробные, надежные и взаимодополняющие протоколы для выполнения окрашивания и визуализации клеточного и субклеточного разрешения фиксированных моделей 3D-культур клеток in vitro размером от 100 мкм до нескольких миллиметров.
Эта процедура представляет две различные стратегии для работы с большим разнообразием размеров и типов моделей 3D-культур клеток in vitro. Выбор одного (3D-анализ всего монтажа) или другого (2D-анализ секционирования) будет зависеть от используемой модели и исследуемой проблемы. 3D-анализ цельного крепления методом конфокальной микроскопии позволяет визуализировать ячейки с полем глубиной до 200 мкм, независимо от общего размера 3D-структуры, тогда как 2D-секционирование применимо к образцам любого размера, но визуализация остается 2D-размерной. Ниже приведены некоторые рекомендации по устранению неполадок и технические соображения.
Потеря 3D-структур во время рабочего процесса является наиболее распространенным недостатком. Они могут оставаться прикрепленными к наконечникам и трубкам, поэтому предварительное покрытие наконечников и трубок с 0,1% раствором PBS-BSA является ключевым. Кроме того, крайне важно, чтобы 3D-структуры оседаляли между изменениями реагентов и выполняли все пипетки очень осторожно. Как упоминалось в процедуре, на всех этапах, если осаждение 3D-структуры слишком длинное, клетки могут быть аккуратно раскручены при 50 × g в течение 5 мин при RT. В зависимости от цели исследования, преимущества / недостатки такой стадии вращения следует рассматривать, поскольку центрифугирование может нарушить форму 3D-структур. Кроме того, следует позаботиться о сохранении этой морфологии на этапе фиксации, потому что кистозные органоиды имеют тенденцию к коллапсу. Крепление конструкций размером до 400 мкм должно предотвращать структурные изменения.
Для оптимальной иммуномаркировки восстановление органоидов из их 3D-матриц является решающим шагом. 3D-матрица может препятствовать адекватному проникновению антител или приводить к высокому фоновому окрашиванию из-за неспецифического связывания с матрицей. Удаление ECM может изменить морфологию внешних сегментов органоидов (особенно в случае небольших клеточных выступов, простирающихся от изученных 3D-структур) и частично затруднить анализ. Для таких 3D-структур матрица может сохраняться на протяжении всей процедуры; однако условия культивирования должны быть тщательно адаптированы для выращивания клеток в минимальном количестве матрицы, чтобы предотвратить недостаточное проникновение растворов и антител и избежать последовательных этапов промывки, направленных на снижение чрезмерного фонового шума 6,8.
Стадия оптической очистки, описанная в этом протоколе в 3D-секции окрашивания всего крепления, уместна для визуализации 3D-структур глубиной до 150-200 мкм вместо 50-80 мкм без очистки. По сравнению с другими методами очистки, которые часто требуют нескольких недель и с использованием токсичных очищающих агентов, в этом протоколе 4,9 использовался ранее опубликованный быстрый и безопасный этап очистки. Кроме того, этот этап очистки является обратимым, и новые антитела могут быть добавлены к первоначальному окрашиванию без потери разрешения или яркости4. Тем не менее, в зависимости от изученной модели 3D-культуры клеток, глубина 150-200 мкм может быть недостаточной для информативного изображения 3D-структуры, и этот протокол очистки может вызвать изменения в общей морфологии сферических, монослойных органоидов с большими люменами4. Пользователи должны тщательно спроектировать свой эксперимент и, при необходимости, оптимизировать время стадии пермеабилизации /блокировки (чтобы обеспечить проникновение антител и раствора), стадии очистки (для проникновения глубже 200 мкм образцы должны быть полностью очищены) и получения изображения. Двумя наиболее распространенными технологиями, доступными в основных помещениях, будут световой лист и конфокальная микроскопия. Пользователям нужно будет тщательно выбрать технологию, основанную на размере их 3D-структур и их биологическом вопросе10. Однако, по сравнению с конфокальной микроскопией, разрешение световой листовой микроскопии, полученное для таких глубоких структур, остается неоптимальным для получения субклеточного разрешения.
Здесь сообщается о подробном и надежном процессе, посвященном встраиванию парафина отдельных образцов. Интересно, что Gabriel et al. недавно разработали протокол встраивания 3D-клеточных культур в парафин с повышенной пропускной способностью. Они использовали форму полидиметилсилоксана (PDMS) для ограничения 96 3D-структур в микрочипе в одном блоке, обеспечивая новые перспективы для исследований 3D-моделей опухолей, охватывающих больше групп, временных точек, условий лечения и реплицирует11. Тем не менее, этот метод требует обширных навыков и оборудования, особенно для изготовления предварительного отлива, используемого для создания пресс-форм PDMS.
Таким образом, в этой статье описываются два различных, взаимодополняющих и адаптируемых подхода, позволяющих получать точную и количественную информацию об архитектурном и клеточном составе 3D-клеточных моделей. Оба параметра имеют решающее значение для изучения биологических процессов, таких как внутриопухолевая клеточная гетерогенность и ее роль в устойчивости к лечению.
Раскрытие информации
Авторам нечего раскрывать.
Благодарности
Эта работа была поддержана премией St Baldrick's Robert J. Arceci Innovation Award #604303.
Материалы
| Name | Company | Catalog Number | Comments |
| Equipment | |||
| Biopsy pad Q Path blue | VWR | 720-2254 | |
| Cassettes macrostar III Blc couv. Char. x1500 | VWR | 720-2233 | |
| Cassette microtwin white | VWR | 720-2183 | |
| Chemical hood | Erlab | FI82 5585-06 | |
| Filter tips 1000 µL | Star lab Tip-One | S1122-1730 | |
| Fine forceps | Pyramid innovation | R35002-E | |
| Flat-bottom glass tubes with PTFE lined 2 mL | Fisher Scientific | 11784259 | Excellent for environmental samples, pharmaceuticals and diagnostic reagents. PTFE is designed for the ultimate in product safety. PTFE provides totally inert inner seal and surface facing the sample or product. |
| Glass bottom dish plate 35 mm | Ibedi | 2018003 | |
| Horizontal agitation | N-BIOTEK | NB-205 | |
| Incubator prewarmed to 65 °C | Memmert Incubator | LAB129 | |
| Inox molds 15x15 | VWR | 720-1918 | |
| Microscope Slides Matsunami TOMO-11/90 | Roche diagnostics | 8082286001 | these slides are used for a better adhesion of sections |
| Microtome | Microm Microtech France | HM340E | |
| Panoramic scan II | 3dhistech | 2397612 | |
| Paraffin embedding equipment | Leica | EG1150C | |
| Plastic pipette Pasteur 2 mL | VWR | 612-1681 | |
| Q Path flacon 150mL cape blanc x250 | VWR | 216-1308 | Good for environmental samples, pharmaceuticals and diagnostic reagents. Polypropylene (PP) are rigid, solid, provide excellent stress crack and impact resistance and have a good oil and alcohol barrier and chemical resistance. PE-lined cap is stress crack resistant and offers excellent sealing characteristics. |
| Set of micropipettors (p200, p1000) | Thermo Scientific | 11877351 (20-200) 11887351(p1000) | |
| OPERA PHENIX | PerkinElmer | HH14000000 | |
| SP5 inverted confocal microscope | Leica | LSM780 | |
| Tissue cassette | VWR | 720-0228 | |
| Zeiss Axiomager microscope | Leica | SIP 60549 | |
| Reagent | |||
| Bovine Serum Albumin (BSA) | Sigma-Aldrich | A7030-100G | |
| Cytoblock (kit) | Thermofisher Scientific | 10066588 | |
| Dimethyl sulfoxide (DMSO) | Sigma-Aldrich | 57648266 | CAUTION: toxic and flammable. Vapors may cause irritation. Manipulate in a fume hood. Avoid direct contact with skin. Wear rubber gloves, protective eye goggles. |
| Eosin aqueous 1% | Sigma-Aldrich | HT110316 | |
| Ethanol 96% | VWR | 83804.360 | CAUTION: Causes severe eye irritation. Flammable liquid and vapor. Causes respiratory tract irritation. Manipulate in a fume hood. Wear protective eye goggles. |
| Ethanol 100% | VWR | 20821.365 | CAUTION: Causes severe eye irritation. Flammable liquid and vapor. Causes respiratory tract irritation. Manipulate in a fume hood. Wear protective eye goggles. |
| Formalin 4% | Microm Microtech France | F/40877-36 | CAUTION: Formalin contains formaldehyde which is hazardous. Manipulate in a fume hood. Avoid direct contact with skin. Wear rubber gloves and protective eye goggles. |
| Fructose | Sigma-Aldrich | F0127 | |
| Gill hematoxylin type II | Microm Microtech France | F/CP813 | |
| Glycerol | Sigma-Aldrich | G5516 | 500 mL |
| Hoechst 33342 | Life Technologies | H3570 | CAUTION: Suspected of causing genetic defects. Avoid direct contact with skin. Wear rubber gloves and protective eye goggles. |
| Normal donkey serum | Sigma-Aldrich | D9663 | 10 mL |
| Paraffin Wax tek III | Sakura | 4511 | |
| Phosphate Buffer Saline (PBS) 1 X | Gibco | 14190-094 | |
| Tris-Buffered Saline (TBS) 10X | Microm Microtech France | F/00801 | 100 mL |
| Triton X-100 | Sigma-Aldrich | T8532 | CAUTION: Triton X100 is hazardous. Avoid contact with skin and eyes. |
| Xylene | Sigma-Aldrich | 534056 | CAUTION: Xylene is toxic and flammable. Vapors may cause irritation. Manipulate in a fume hood. Avoid direct contact with skin. Wear rubber gloves, protective eye goggles. |
| Solutions | |||
| Clearing solution | Glycerol-Fructose clearing solution is 60% (vol/w) glycerol and 2.5 M fructose. To prepare 10 mL of this solution, mix 6 mL of glycerol and 4.5 g of fructose. Complete to 10 mL with dH2O. Use a magnetic stirrer overnight. Refractive index = 1.4688 at room temperature (RT: 19–23 °C). Store at 4 °C in dark for up to 1 month. | ||
| PBS-BSA 0,1% solution | To prepare 0,1% (vol/wt) PBS-BSA 0,1% solution, dissolve 500 mg of BSA in 50 mL of PBS-1X (store at 4°C for up to 2 weeks). And dilute 1mL of this solution into 9mL of PBS-1X. This solution can be used to precoat the tip and centrifugation tube. | ||
| Permeabilisation-blocking solution (PB solution) | The PBSDT blocking solution is PBS-1X supplemented with 0.1% – 1% Tritonx-100 (depending on the protein localization membrane/nucleus), 1% DMSO, 1% BSA and 1% donkey serum (or from the animal in which the secondary antibodies were raised). This solution can be stored at 4°C for up to 1 month. | ||
| PB:PBS-1X (1:10) solution | PB:PBS-1X (1:10) solution is a 10 time diluted PB solution. To prepare 10 mL of this solution dilute 1 mL of PB solution in 9 mL of PBS-1X. | ||
| Software | |||
| Halo software | Indicalabs | NM 87114 | |
| Harmony software | PerkinElmer | HH17000010 |
Ссылки
- Ryu, N. E., Lee, S. H., Park, H. Spheroid culture system methods and applications for mesenchymal stem cells. Cells. 8 (12), 1-13 (2019).
- Bartfeld, S., Clevers, H. Stem cell-derived organoids and their application for medical research and patient treatment. Journal of Molecular Medicine. 95 (7), 729-738 (2017).
- Cui, X., Hartanto, Y., Zhang, H. Advances in multicellular spheroids formation. Journal of the Royal Society, Interface. 14 (127), (2017).
- Dekkers, J. F., et al. High-resolution 3D imaging of fixed and cleared organoids. Nature Protocols. 14, 1756-1771 (2019).
- Broutier, L., et al. Culture and establishment of self-renewing human and mouse adult liver and pancreas 3D organoids and their genetic manipulation. Nature Protocols. 11 (9), 1724-1743 (2016).
- Rezanejad, H., Lock, J. H., Sullivan, B. A., Bonner-Weir, S. Generation of pancreatic ductal organoids and whole-mount immunostaining of intact organoids. Current Protocols in Cell Biology. 83 (1), 82 (2019).
- Edmondson, R., Broglie, J. J., Adcock, A. F., Yang, L. Three-dimensional cell culture systems and their applications in drug discovery and cell-based biosensors. Assay and Drug Development Technologies. 12 (4), 207-218 (2014).
- McCray, T., Richards, Z., Marsili, J., Prins, G. S., Nonn, L. Handling and assessment of human primary prostate organoid culture. Journal of Visualized Experiments: JoVE. (143), e59051 (2019).
- Ueda, H. R., et al. Tissue clearing and its applications in neuroscience. Nature Reviews: Neuroscience. 21 (2), 61-79 (2020).
- Lazzari, G., et al. Light sheet fluorescence microscopy versus confocal microscopy: in quest of a suitable tool to assess drug and nanomedicine penetration into multicellular tumor spheroids. European Journal of Pharmaceutics and Biopharmaceutics. 142, 195-203 (2019).
- Gabriel, J., Brennan, D., Elisseeff, J. H., Beachley, V. Microarray embedding/sectioning for parallel analysis of 3D cell spheroids. Scientific Reports. 9, 16287 (2019).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеThis article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены