
Method Article
Одновременная томографическая визуализация Брайтфилда, флуоресценции и оптической когерентности сократителения сердечных трабекул Ex Vivo
В этой статье
Резюме
Этот протокол представляет собой коллекцию саркомера, кальция и макроскопических геометрических данных из активно сокращающейся сердечной трабекулы ex vivo. Эти одновременные измерения стали возможными благодаря интеграции трех методов визуализации.
Аннотация
В сердечной мышце внутриклеточные транзиторы Ca2+ активируют сократительные миофиламенты, вызывая сокращение, макроскопическое укорочение и геометрическую деформацию. Наше понимание внутренних отношений между этими событиями было ограничено, потому что мы не можем ни «видеть» внутри мышцы, ни точно отслеживать пространственно-временную природу динамики возбуждения-сокращения. Чтобы решить эти проблемы, мы создали устройство, которое сочетает в себе набор модальностей визуализации. В частности, он объединяет микроскоп Яркого поля для измерения локальных изменений длины саркомера и деформации ткани, флуоресцентный микроскоп для визуализации переходного периода Ca2 + и оптический когерентный томограф для захвата геометрических изменений ткани на протяжении всего времени сердечного цикла. Здесь мы представляем инфраструктуру обработки изображений и связанную с ней структуру сбора данных. Данные собираются из изолированных палочкоподобных тканевых структур, известных как trabeculae carneae. В нашем инструменте пара платиновых крючков с позиционным управлением удерживает каждый конец образца мышц ex vivo, в то время как он постоянно наполняется богатым питательными веществами солевым раствором. Крючки находятся под независимым контролем, что позволяет в режиме реального времени контролировать длину и силу мышц. Продольная трансляция позволяет фрагментазировать образец, преодолевая ограничения, связанные с относительным размером окна визуализации микроскопа (540 мкм на 540 мкм) и длиной типичной трабекулы (>2000 мкм). Платиновые электроды на любом конце мышечной камеры стимулируют трабекулу с заданной пользователем скоростью. Мы используем сигнал стимуляции в качестве триггера для синхронизации данных из каждого окна изображения, чтобы восстановить все подергивание образца в стационарных условиях. Применение методов обработки изображений к этим данным визуализации brightfield обеспечивает смещение тканей и карты длины саркомера. Такой набор данных, когда он будет включен в конвейер моделирования экспериментов, обеспечит более глубокое понимание мышечной сократительной однородности и гетерогенности в физиологии и патофизиологии.
Введение
Суперфузия изолированных препаратов сердечной мышечной ткани является стандартным и широко используемым протоколом для изучения сердечной ионной активации и механики1. В частности, выделение трабекул, палочкоподобных структур от желудочковых стенок, позволило оценить явления, включая зависящие от длины активации сокращения2,растягивательно-зависимый ответ сокращения3,4и диастолическую вязкоупругость 5 сердечнойткани. Тер Керс, инициатор этой техники суперфузии изолированных трабекул, первоначально использовал комбинацию флуоресцентной визуализации для измерений Ca2+ и лазерной дифракции для определения длин саркомера2,5. Со времени этих ранних исследований становится все более распространенным извлечение информации о длине саркомера с большим пространственным разрешением с использованиемметодов 6 на основе 2D-быстрого преобразования Фурье (FFT) на изображениях микроскопии яркого поля. Две системы визуализации позволяют частично оценить основную связь между высвобождением Ca2+ и производством силы, зависящей от длины саркомера.
Сердечная мышца поползнена, с видимыми полосами, связанными с нижележащей серией сократительных единиц, состоящих из толстых и толстых нитей. Взаимодействие этих составляющих нитей, составляющих саркомеру, лежит в основе генерации силы, которая начинается следующим образом: деполяризующий электрический сигнал, или потенциал действия, вызывает открытие зависимых от напряжения каналов L-типа Ca2+ в клеточной мембране; последующий клеточный приток Ca2+ индуцирует высвобождение Ca2+ из саркоплазматического стикулума (SR), внутриклеточного хранилища Ca2+, в процессе, известном как Ca2+-индуцированный Ca2+ высвобождение7; это внезапное увеличение внутриклеточной концентрации Ca2+ от наномолярного до микромолярного диапазона позволяет происходить производство силы; Насосы Ca2+ непрерывно выдавливают Ca2+ из цитозола обратно в SR и внеклеточный компартмент; когда внутриклеточная концентрация Ca2+ возвращается в наномолярный диапазон, производство силы прекращается, и мышца расслабляется. Во время производства силы составляющие толстые и тонкие нити скользят друг над другом. Длина саркомера диктует относительную степень перекрытия и, следовательно, потенциал для производства силы мышцы макроскопически.
В этой статье мы расширяем эти методы флуоресцентно-яркой визуализации, включая оптическую когерентную томографию (OCT). OCT использует физический принцип интерференции и способен собирать геометрическую деформацию ткани для понимания сократительной гетерогенности мышц8. Наше устройство(рисунок 1)использует спектрально-доменную ОКТ-систему (SD-OCT). В SD-OCT светоотвод разделяет свет от широкополосного сверхлюминесцентного диода короткой когерентности на эталонный и измерительный диапазоны. Опорный рычаг содержит фиксированное зеркало, а измерительный рычаг содержит 2D-гальванометр для управления светом. Свет, рассеянный от образца, собирается и мешает отражению света в опорном плече, образуя интерференционную картину. Информация о глубине кодируется в частоте спектральной полосы. Для извлечения информации сигнал пропускается через спектрометр и к результату применяется обратный БПФ. Соответствующий 1D сигнал представляет собой структуры на разных глубинах, соответствующие изменениям показателя преломления9 (A-scan). Направляя лазер по одной оси, можно построить поперечное сечение интересующего образца (B-сканирование) и, аналогично, повторяя процесс по пошаговому рисунку на оставшейся оси, может быть получено трехмерное изображение (C-scan). Таким образом, можно собрать серию B-сканов на одном срезе для повторяющегося изменяющегося во времени объекта на основе внешнего триггера и повторить для создания трехмерного сканирования, представляющего изменяющееся во времени планарное изображение10.
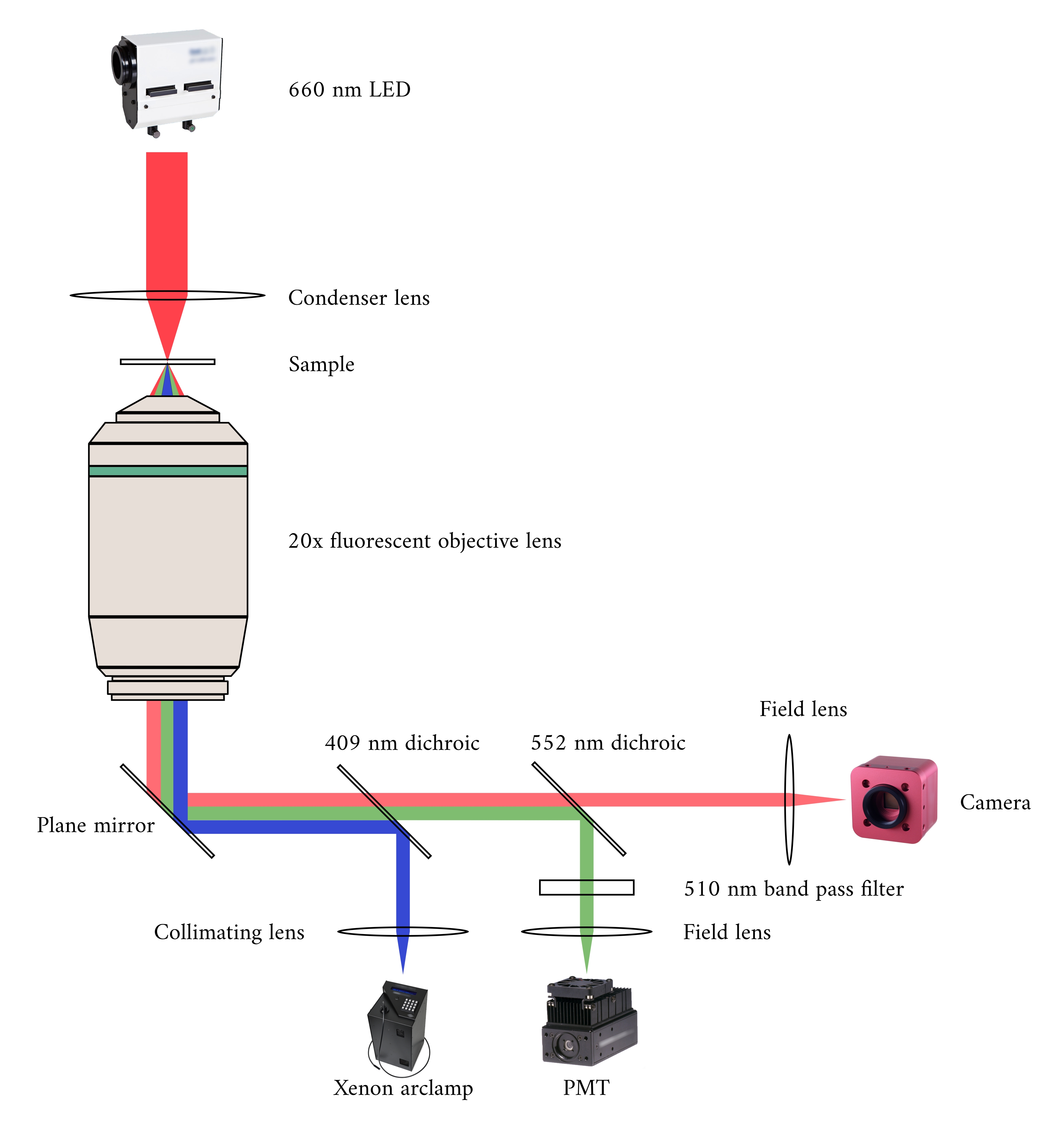
При интеграции трех систем визуализации мы рассмотрели следующие два принципа. Во-первых, датчики изображения не должны обнаруживать свет от альтернативного метода визуализации, а во-вторых, физическая конструкция должна содержать свободное пространство для по крайней мере трех одновременных плоскостей визуализации. Чтобы удовлетворить первое требование, микроскоп Brightfield использует светодиод с длиной волны 660 нм для освещения образца в перевернутой конфигурации. Флуоресцентный микроскоп находится в конфигурации эпифлуоресценции, где одна и та же объективная линза используется как для возбуждения, так и для сбора излучаемого света. Свет возбуждения имеет длину волны от 340 нм до 380 нм, а фотоумножительная трубка (PMT) измеряет излучаемый свет на длине волны 510 нм. Пара дихроичных зеркал позволяет этим двум оптическим путям использовать один и тот же физический след, не мешая противоположному измерению(рисунок 2). Наконец, OCT использует широкополосный (спектральная ширина 100 нм) свет с центральной длиной волны 840 нм, отличный от двух других модальностей. Из-за низкой когерентности света, используемого для OCT, любой рассеянный свет от источников флуоресценции яркого поля не будет способствовать интерференционной картине, которая кодирует информацию о глубине. Для второго требования конструкция корпуса капиллярной трубки имеет доступные оптические пути к передней, нижней и верхней плоскостям образца. Во время экспериментов два платиновых крючка удерживают трабекулу в капиллярной трубке, перфузивированной кислородом раствором Krebs-Henseleit (KH). Головка гальванометра ОКТ ориентирована ортогонально на путь визуализации яркого поля-флуоресценции, чтобы воспользоваться преимуществами третьей ортогональной оптической плоскости(рисунок 3).
В этой статье излагаются конструктивные соображения для создания устройства, способного одновременно визуализировать кальций, длину саркомера и геометрию мышц. Чтобы продемонстрировать эти измерительные возможности, мы описываем процесс выделения желудочковой трабекулы, получение необходимых буферных растворов, а также критические этапы, связанные с обработкой и флуоресцентной нагрузкой ex vivo trabecula. Наконец, в этом документе описываются процессы, необходимые для преобразования набора данных в более полезные визуализации.
протокол
Комитет по этике животных Оклендского университета одобрил обращение с крысами и подготовку образцов тканей.
1. Калибровка изображений
- Калибровка пикселей микроскопа Brightfield
- Наполните измерительную камеру дистиллированной водой.
- Поместите дифракционную решетку с известными линиями на мкм в измерительную камеру.
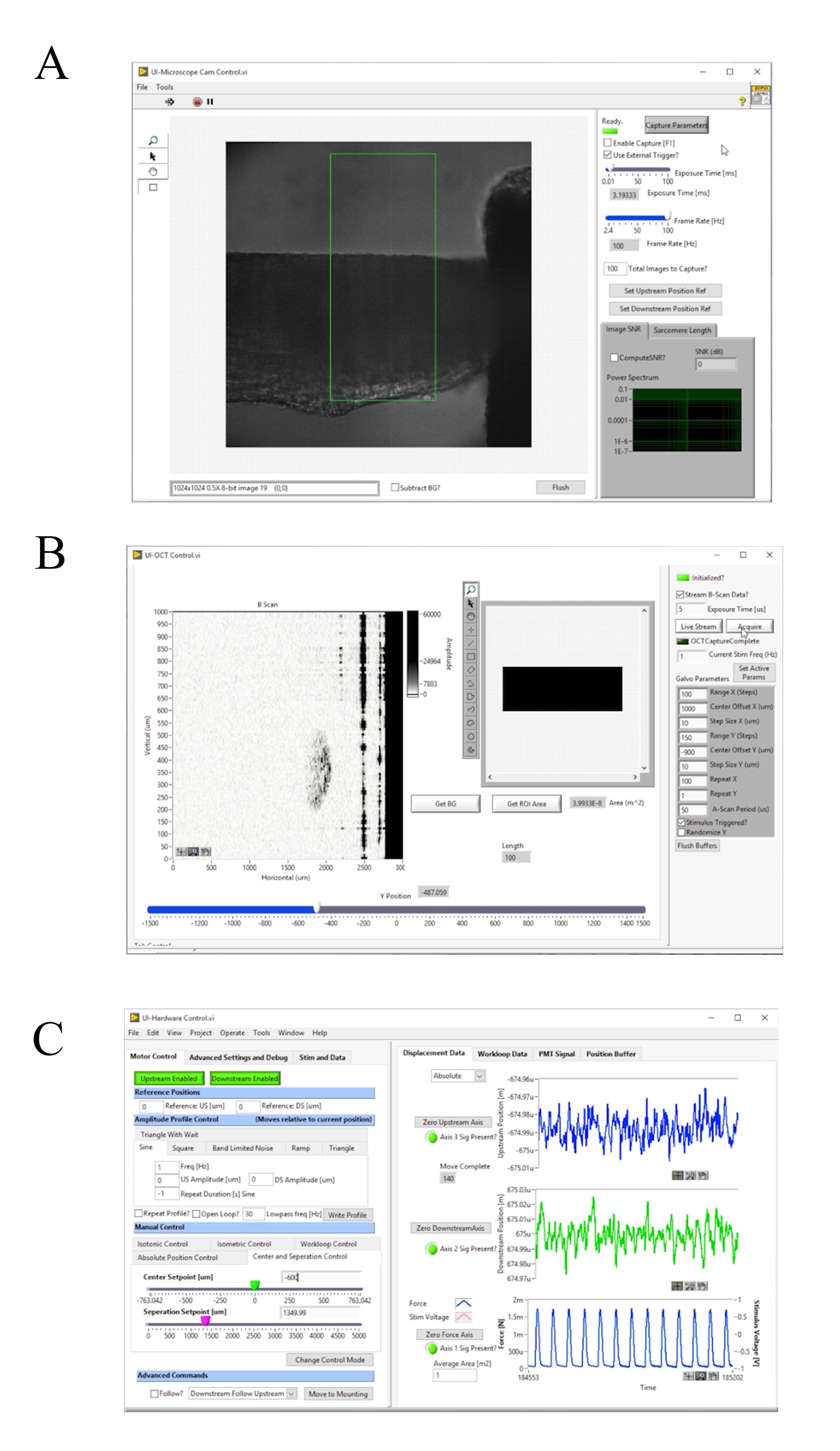
- Нажмите клавишу F1, чтобы включить захват и отрегулировать частоту кадров [Гц], пока дифракционная решетка не будет хорошо видна(рисунок 4A). Убедитесь, что дифракционная решетка проходит параллельно краю рамы. Нажмите клавишу F1 еще раз, чтобы остановить захват.
- Установите для параметра Total Images to Capture? значение один, нажмите клавиши CTRL + SHIFT + S для потоковой передачи данных на диск и нажмите клавишу F1, чтобы захватить изображение дифракционной решетки.
- Откройте ImageJ и импортируйте изображение дифракционной решетки (Файл > Открыть > Выбрать калибровочное изображение). Удерживая сдвиг, провести линию, которая охватывает 20 светлых и темных полос дифракционной решетки.
- Калибровка изображения (Анализ > Задать масштаб). Длина линии из шага 1.1.5 задает значение Расстояние в пикселях. Установите значение «Известное расстояние» в 20 раз больше линий на мкм измерения, а для единицы длины — значение мкм. Обратная величина — это количество микрометров, представленных пикселем.
- Калибровка глубины OCT
- Измерьте толщину стекла микроскопа с помощью суппортов Вернье.
- Включите источник лазера OCT.
- Накройте головку гальванометра и нажмите Get BG, чтобы измерить фоновую интерференционную картину и вычесть ее из измерения(рисунок 4B).
- Зажмите измеряемый стеклянный микроскоп (из шага 1.2.1) в измерительном рычаге OCT.
- Щелкните Live Stream, чтобы просмотреть изображение центра развертывания Office. Отрегулируйте слайд стеклянного микроскопа до тех пор, пока он не будет виден в B-сканировании.
- Чтобы записать изображение B-сканирования, установите для диапазона Y (Шаги) значение один, щелкните Поток данных B-сканирования?и нажмите кнопку Получить.
- Импортируйте изображение B-сканирования в ImageJ (файл > Открыть > Выбрать B-скан). Удерживая сдвиг, проведите линию между границами стекла микроскопа.
- Установите масштаб (Анализ > Задать масштаб). Установите известное расстояние для измерения, собранного на шаге 1.2.1.
- Чтобы рассчитать разрешение глубины в воздухе, скорректируйте показатель преломления слайда микроскопа(nстекла = 1,5175)11, умножив значение измерения на пиксель на nстекол.
ПРИМЕЧАНИЕ: N стекла, которое указано, предназначено для боросиликатного стекла. Слайды микроскопа могут быть изготовлены из различных материалов. Используйте соответствующий показатель преломления для измеряемого слайда. - Чтобы масштабировать разрешение глубины для миокарда, разделите значение с шага 1.2.9. по nмиокарда = 1,38 (значение, о котором сообщалосьранее 12).
2. Подготовка образцов мышц
- Подготовьте установку для вскрытия.
- Вылейте часть раствора для рассечения (описанного в таблице 1)в небольшую металлическую миску и поместите в морозильную камеру примерно за час до иссечения сердца.
- Установите рассеченную установку, гарантирующую, что раствор для рассечения хорошо насыщен кислородом (100% кислород) и промывывается через каждую из линий труб. Заполните камеру рассечения кислородным раствором для рассечения и свободно свяжите 3/0 шва вокруг перфузионного катетера.
- Иссекаем сердце.
- Обезболить крысу Wistar 8-10 недель с использованием газообразного изофлурана (< 5% кислорода). Подтвердите анестезию путем защемления хвоста.
- Поместите анестезированную крысу в положение лежа на спине и подкожно вбрызгиваемую область введения раствором гепарина (1000 МЕ/кг). Поддерживайте анестезию еще пять минут, чтобы позволить гепарину циркулировать.
- Извлеките металлическую чашу, содержащую раствор для рассечения, из морозильной камеры и поместите ее рядом со скамейкой для эвтаназии.
ПРИМЕЧАНИЕ: Избегайте полного замораживания раствора для рассечения, чтобы обеспечить полное погружение рассеченного сердца. - Переместят обезболенную крысу на скамейку эвтаназии и усыпить при вывихе шейки матки.
- Откройте грудь крысы ножницами, сначала разрезав стенку тела вдоль нижней стороны грудной клетки, затем диафрагму, прежде чем продолжить вдоль боковых границ грудной клетки. Поднимите грудь с дороги.
- Возьмите сердце одной рукой, в то время как другая рука использует пару изогнутых ножниц, чтобы разрезать соединительные сосуды (аорта, полая вена и т. Д.).
- Быстро погружает сердце в холодный рассеченный раствор.
- Выделяют трабекулу.
- Определите аорту, пока сердце находится в металлической чаше, затем перенесите сердце в камеру рассечения. Используя два изогнутых щипца, потяните аорту над перфузионной канюлью.
- Удерживайте аорту на месте одним щипцом. Между тем, откройте линию трубки, чтобы раствор для рассечения протекал через перфузионную канюлю.
ПРИМЕЧАНИЕ: Стремитесь завершить перфузию в течение минуты после иссечения сердца. - Как только коронарная сосудистая клетка очищена от крови и сердце полностью перфузировано раствором для рассечения, остановите перфузионный поток и закрепите аорту на месте с помощью шва. Включите поток обратно и перфузируйте канюлированное сердце.
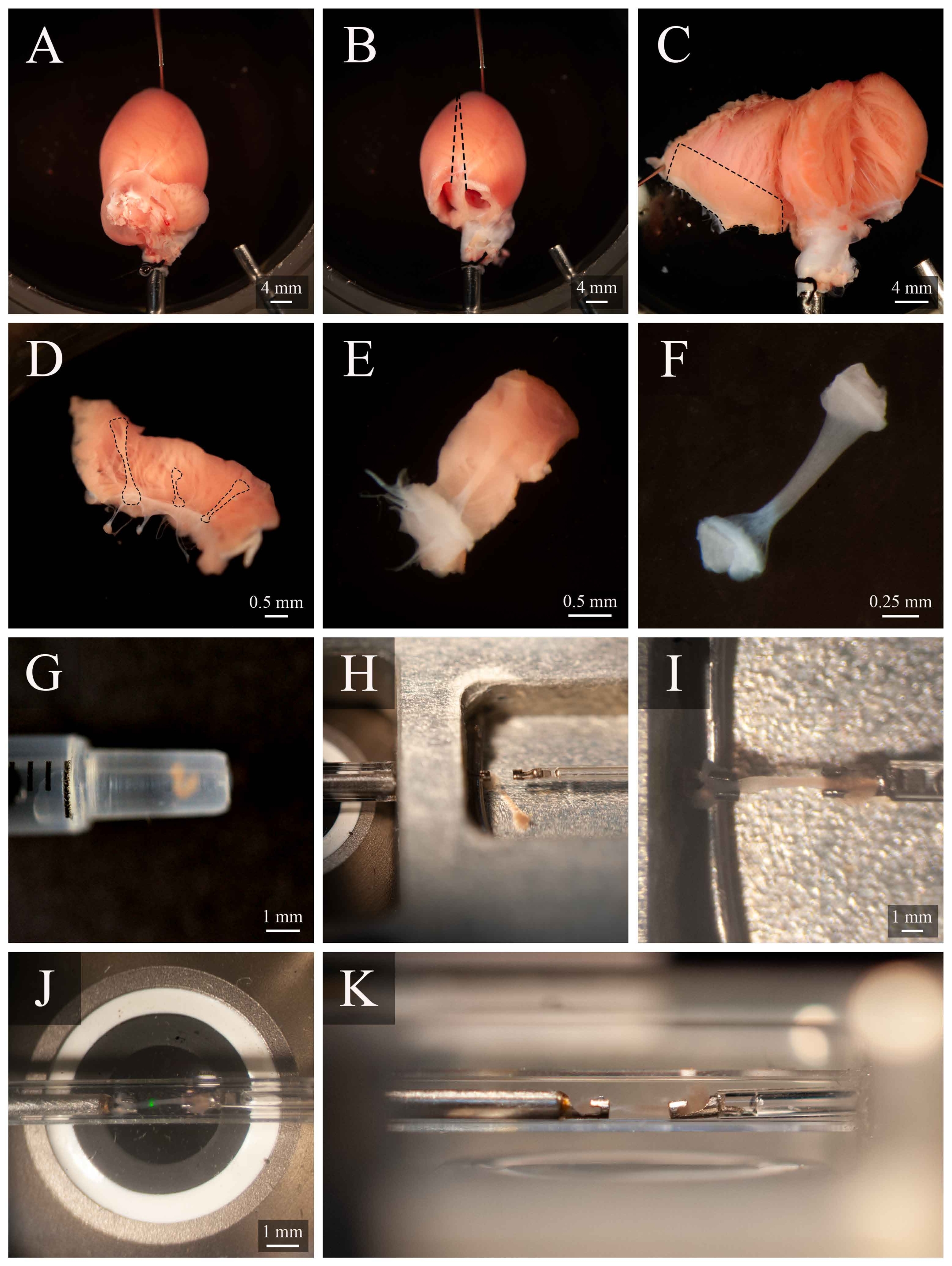
- Поверните канюлю так, чтобы левая коронарная артерия была видна на верхней поверхности. Прикрепить вершину сердца к нижней части рассеченной камеры(рисунок 5А). Отрежьте оба предсердия(рисунок 5В).
- С помощью набора пружинных ножниц разрезают вдоль правой стороны перегородки до вершины сердца (как показано на рисунке 5В). Прикрепить открытый левый желудочек к основанию рассеченной камеры. Затем разрезайте вдоль левой стороны перегородки, откройте правый желудочек и прикрепите его к основанию рассеченной камеры(рисунок 5C).
ПРИМЕЧАНИЕ: Для того, чтобы закрепить желудочки в открытом положении, некоторые сосочковые мышцы придется разрезать. Определите свободно работающий трабекул в правом желудочке(рисунок 5D-E). - С помощью пружинных ножниц и щипцов разрезают ткань стенки, окружающую трабекулу, затем разрезают ткань стенки пополам ортогонально в направлении трабекулы. Обрежьте ткань стены до тех пор, пока ее размер не будет соответствовать используемой конфигурации монтажа. В этом случае примерно половина размера кунжутного семени(рисунок 5F).
ПРИМЕЧАНИЕ: Трабекулы могут быть рассечены из правого и левого желудочков, но те, что слева, как правило, более мутные и менее применимы для измерения саркомера и геометрии. - Оставьте иссеченную трабекулу в рассеченной камере, непрерывно наливая раствором для рассечения.
3. Экспериментальный протокол
ПРИМЕЧАНИЕ: Устройство13, используемое для этого эксперимента, было построено собственной установкой и использует пользовательский код управления. Необходимыми соображениями для проектирования устройства, построенного для воспроизведения этих данных, являются два независимо приводимых в действие монтажных крючка, измерительная камера с тремя оптически четкими осями(рисунок 3)и внешняя триггерная линия, которая синхронизирует камеры Brightfield и OCT со стимулятором. Сигнал напряжения и силы PMT собирался с помощью аналоговых карт DAQ, изображения с OCT и микроскопа Brightfield собирались с помощью карт захвата кадров Camera Link, а сигнал стимула собирался с помощью цифровой карты ввода/вывода. Данные хранились в автономном режиме с использованием набора потребительских циклов производителя для поддержания временного выравнивания.
- Подготовьте кардиомиометр.
- Промывайте горячую (~60 °C) воду, дистиллированную воду (комнатной температуры), а затем раствор суперфусат через измерительную камеру. Непрерывно пузырьки раствора суперфусата с карбогеном.
- Включите источник освещения микроскопа Brightfield и нажмите клавишу F1, чтобы включить захват(рисунок 4A). Вручную отрегулируйте нисходящий крюк, пока он не будет центрирован на изображении яркого поля. Щелкните Нулевая ось нисходящего потока,затем Downstream Отключено, чтобы включить двигатель(рисунок 4C). Перемещайте ползунок DS Setpoint [um] до тех пор, пока конец перехватчика не выровняется с краем интересуемой области по умолчанию.
- Повторно обнулите нисходящую ось, затем переместите ползунок DS Setpoint [um] на 1000. Повторите процесс с помощью восходящего хука, но не перемещайте ползунок US Setpoint [um].
- Щелкните Move to Mounting (рисунок 4C).
- Запустите систему флуоресцентного освещения, переключив переключатель Лампы перед быстрым включением подсистем контроллера, переключив главный переключатель.
ПРИМЕЧАНИЕ: Некоторые источники ультрафиолетового света производят большое количество озона. Если это так, подключите экстрактор озона к выходу источника света и убедитесь, что он работает до включения источника флуоресцентной подсветки. - Переключите режим работы на Turbo-Blanking, нажав кнопку Mode на передней панели, затем 2,затем 1. Нажмите кнопку On-line, чтобы разрешить управляющее коду информировать операцию.
- Смонтировать трабекулу.
- Приостановите поток суперсмешка через измерительную камеру. Заполните монтажную камеру раствором для рассечения.
- Используя шприц объемом 1 мл, транспортировать трабекулу из камеры рассечения в монтажную камеру(рисунок 5G).
- Чтобы перенести трабекулу, поместите шприц вертикально и соприкоснись с поверхностью раствора монтажной камеры. Позвольте трабекуле опуститься в монтажную камеру под действием силы тяжести(рисунок 5H).
- Опустить уровень жидкости в монтажной камере так, чтобы она была на одном уровне со средней частью крючков.
- Отрегулируйте расстояние между крючками, чтобы отразить длину слабины трабекулы, переместив ползунок DS Setpoint [um].
- Используя микроскоп для облегчения визуализации, слегка захватите один из кусков торцевой ткани щипцами и установите его на крючок вверх по течению. Установите другой кусок торцовой ткани на нисходящий крючок(рисунок 5I).
- После надежного монтажа переместите трабекулу обратно в измерительную камеру(рисунок 5J),нажав кнопку Move to Chamber (рисунок 4C). Возобновить поток суперсмешков и экстракцию жидкости.
- Установите частоту стимула [Гц] на 1, длительность стимула [мс] на 10 и напряжение стимула на 10. Начните стимуляцию, нажав Stimulus On?.
- Приготовьте трабекулу.
- Примерно через 1 ч акклиматизации постепенно уменьшают напряжение стимула и продолжительность стимула на 1 В и 1 мс шагов соответственно. Типичный набор значений составляет 3 В и 3 мс.
- Включите систему освещения brightfield. Нажмите клавишу F1 и выберите интересуяшую область, которая заключает в себя полосатую область пользовательского интерфейса. Щелкните Compute SL?, чтобы вычислить среднюю длину саркомера в выделенной области. Увеличьте длину мышц до тех пор, пока средняя длина саркомера не увеличится до 2,32 мкм, увеличив ползунок «Заданное значение разделения»...
ПРИМЕЧАНИЕ: Вычислительная SL? использует 2D FFT, описанный в шаге 4.3. Область интереса, используемая для расчета средней длины саркомера, обычно составляет от 100 мкм до 150 мкм в квадрате. Поэтому, когда мышца приближается к оптимальной длине саркомера, для расчета средней длины саркомера используется от 43 до 65 саркомер. - Перемещайте мышцу, регулируя ползунок «Центральная заданная точка» [um] на вкладке«Управление центром и разделением» (рисунок 4C)так, чтобы край нисходящего крючка был виден только на изображении яркого поля. Соберите информацию о флуоресценции для десяти подергиваний.
- Увеличьте значение Центрального заданного значения [um] на 200 и соберите еще десять подергиваний информации о флуоресценции. Повторяйте этот процесс до тех пор, пока изображение яркого поля не будет содержать восходящий крюк. Соберите информацию о флуоресценции последнего окна.
- Верните трабекулу в центральное положение, установив для параметра Центральное заданное значение [um] значение 0.
- Уменьшить частоту стимула до 0,2 Гц и переключиться с KH суперфузата на нагрузочный раствор Fura-2 (подробно описано в таблице 1).
- Измеряйте флуоресцентный сигнал каждые 10 минут, щелкнув Включить источник флуоресценции на вкладке Stim and Data. Визуализируйте флуоресцентный сигнал на вкладке PMT Signal.
- После того, как сигнал 360 нм увеличился в 10 раз или продолжительность процедуры загрузки превысила 2 ч, верните частоту стимула к 1 Гц и переключитесь обратно на раствор суперфузата KH.
- Проверяйте измерение соотношения каждые 10 минут до тех пор, пока измерение коэффициента не стабилизируется, после чего может начаться сбор данных.
- Собирайте данные ярко-полевого и флуоресцентного изображения.
- Верните мышцу в положение, где край нисходящего крючка только что присутствует в ярком изображении. Начните потоковую передачу аппаратных данных, щелкнув Потоковые данные на диск на вкладке Stim и Данные пользовательского интерфейса аппаратного управления. Захватите информацию о флуоресценции, щелкнув Включить источник флуоресценции.
- В пользовательском интерфейсе brightfield imaging установите режим захвата на внешний триггер, увеличьте частоту кадров до 100 Гц и установите количество изображений для захвата на 100. Нажмите клавиши CTRL+SHIFT+S, а затем клавишу F1, чтобы записать данные изображения яркого поля для этого окна.
- Увеличьте значение центрального заданного значения [um] на 200 и повторите шаг 3.4.2. Продолжайте идти по протоколу сканирования до тех пор, пока данные изображения не будут собраны для последнего окна из шага 3.3.4.
- Верните трабекулу в центральное положение, установив для параметра Центральное заданное значение [um] значение 0.
- Сбор данных изображений центра развертывания Office.
- Включите лазерный источник OCT, повернув главный ключ к | , нажав кнопку питания, а затем кнопку SDS.
- Накройте головку гальванометра и нажмите Get BG, чтобы измерить фоновую интерференционную картину и вычесть ее из измерения(рисунок 4B).
- Установите режим захвата изображения в режим live-view.
- Отрегулируйте положение yдо тех пор, пока изображение B-scan не будет содержать только восходящий хук. Разделите длину мышцы, отображаемую на передней панели управления(рисунок 4B),на две и вычтите текущее положение y. Введите это значение во входные данные "y-offset". Отрегулируйте значение"x-offset" до тех пор, пока поперечное сечение трабекулы не будет центрировано в кадре.
- Центрируя трабекулу, сканируйте вдоль оси y,регулируя положение y,,чтобы найти положения, соответствующие крючкам вверх и вниз по течению. Запишите эти позиции вниз. Установите для диапазона Y (шагов) абсолютную разницу между этими значениями, деленную на десять.
- Установите для режима захвата изображения значение Стимул срабатывает?, Диапазон X (шаги) 100 и нажмите кнопку Задать активные параметры.
- Щелкните Поток данных B-Scan?,затем Получить.
ПРИМЕЧАНИЕ: Закрытый протокол визуализации требует 200 подергиваний для захвата всей геометрии мышц для образца длиной 2 мм, что соответствует времени захвата ~ 3 мин 20 с.
4. Обработка набора данных изображения яркого поля
- Подготовьте изображения к анализу.
- Импорт изображений в ImageJ (Файл > Импорт > Последовательность изображений > Выбрать изображение).
- Увеличьте контрастность изображения (Image > Adjust > Brightness/Contrast > Переместить минимальные и максимальные ползунки для централизации гистограммы изображения).
- Увеличьте резкость изображения (Фильтры обработки > > Нерезкая маска > установите радиус (Sigma) на 1,0 пикселя, а вес маски (0,1-0,9) на 0,6).
- Экспортируйте последовательность изображений (Сохранить как > Последовательность изображений > Установите формат в PNG, начните с 0, а цифры (1-8) - 4).
- Сшивайте изображения, измеряйте локализованное смещение и вычисляйте локальные длины саркомера.
- Откройте "TrabeculaProcessing.m" (доступно по запросу) и установите для переменной FolderPath основную папку, содержащую все данные, а ImagePath - папку, в которой была сохранена последовательность изображений из Шага 4.1.4. Задайте для разделов количество окон изображения, а для фреймов — количество кадров, захваченных в окне.
- Запустите код.
ПРИМЕЧАНИЕ: Выходные данные будут присутствовать в пути к выходной папке, указанном пользователем. (По умолчанию для пути задано значение FolderPath/Output).
- Техника длины саркомера БПФ
- Используйте программное обеспечение для обработки изображений для выполнения БПФ в области изображения, где саркомы хорошо видны.
- Умножьте результаты калибровки пикселей на мкм с шага 1.1.6 на 1,6 мкм и 3,0 мкм перед вычислением обратного, чтобы получить диапазон пространственных частот, представляющих интерес.
- Подогнать экспоненциальную величину к результату БПФ, игнорируя частотную информацию в диапазоне частот, рассчитанном на шаге 4.3.2, и вычтите ее из результата преобразования, чтобы удалить член постоянного тока.
- Подогнать кривую Гаусса к интересуемой полосе частот.
- Вычислите обратную пику кривой Гаусса. Это средняя длина саркомера для интересуемого региона.
ПРИМЕЧАНИЕ: Расчет БПФ и подгонка экспоненциальных и гауссовских уравнений выполнялись с использованием пользовательского кода LabVIEW.
5. Обработка данных флуоресценции
- Вычтите зависящее от окна автофлуоресценцию из соответствующего окна и вычислите коэффициент сигналов, связанных с длинами волн возбуждения 340 нм и 380 нм.
6. Обработка данных визуализации ЦЕНТРА РАЗВЕРТЫВАНИЯ Office
- Подготовьте набор образов центра развертывания Office к сегментации.
- Откройте ImageJ и импортируйте изображения (Файл > Импорт > Последовательность изображений). Откроется окно проводника, найдите изображения, выберите одно из них и нажмите кнопку Открыть.
ПРИМЕЧАНИЕ: Если код элемента управления для центра развертывания Office не хранит изображения в формате, читаемом ImageJ, преобразуйте их в PNG. - Чтобы упростить визуализацию, организуйте последовательность изображений в гиперстек (Image > Hyperstacks > Stack to Hyperstack). В открывшемся диалоговом окне задайте для числа фрагментов число B-сканов на фрагмент, а для X — количество фрагментов по длине трабекулы.
- Нарисуйте прямоугольник, который заключает в себе трабекулу. Убедитесь, что он заключает в себе весь объем во времени, используя ползунки в окне гиперстека. Обрезать изображение до окна (Image > Crop).
- Удалите фрагменты, содержащие образы крючков для монтирования (Стеки > Инструменты > Хранитель фрагментов). Выберите диапазон фрагментов, содержащий только информацию о трабекуле.
- Откройте ImageJ и импортируйте изображения (Файл > Импорт > Последовательность изображений). Откроется окно проводника, найдите изображения, выберите одно из них и нажмите кнопку Открыть.
- Сегментация поездов WEKA.
- Открытая сегментация WEKA(плагины > сегментация > обучаемая сегментация Weka).
- Установите режим выбора в Freehand.
- Нажмите «Настройки» и настройте параметры классификатора и обучения. (Для этой модели использовались следующие особенности обучения: гауссовское размытие, фильтр Собеля, гессенский, разностные гауссовы, мембранные проекции, двусторонние и липшицы. Толщина мембраны была установлена на 1, размер пластыря мембраны на 8, минимальная сигма на 1 и максимальная сигма на 32. Классификатор был установлен в FastRandomForest, а параметры классификатора были установлены в: batchSize 100, maxDepth to 32, numFeatures в 32, numThreads в 0 и numTrees в 200.)
- Вручную сегментируйте изображения до тех пор, пока обучение не привьёт удовлетворительную сегментацию.
- Сохраните классификатор.
- Сегментация обработанных B-сканов
- Запустите сегментацию WEKA в соответствии с шагом 6.2.1.
- Загрузите классификатор из шага 6.2.5.
- Нажмите кнопку Создать результат.
- Преобразование изображений в 8-битные (Image > Type > 8-bit).
- Преобразуйте изображения в двоичные (Обработать > Двоичный > Сделать двоичный > Метод по умолчанию и фон по умолчанию).
- Сохранить как последовательность изображений (PNG).
- Рассчитайте среднее значение CSA в сегментированных изображениях B-сканирования.
- Подсчитайте количество белых пикселей в двоичном изображении B-сканирования.
- Умножьте площадь пикселя на калиброванное разрешение глубины (из шага 1.2) и 10 мкм (расстояние между соседними A-сканами).
- Повторите для всех B-сканов между крючками и усредните измерения.
- Преобразование сегментированного изображения в сетку.
- Откройте "OCTmain.m" (доступно по запросу) и установите imageDirectory в папку, содержащую выходные данные из Шага 6.3.6. При необходимости задайте параметр outputPath.
- Установите для фрагментов значение "Диапазон Y (шаги)" (Шаг 3.5.5) и кадры значение "Повторить X" (Шаг 3.5.6), z_dim разрешение глубины (Шаг 1.2.10) и x_dim и y_dim значение, присвоенное 10.
- Нажмите кнопку Выполнить.
Результаты
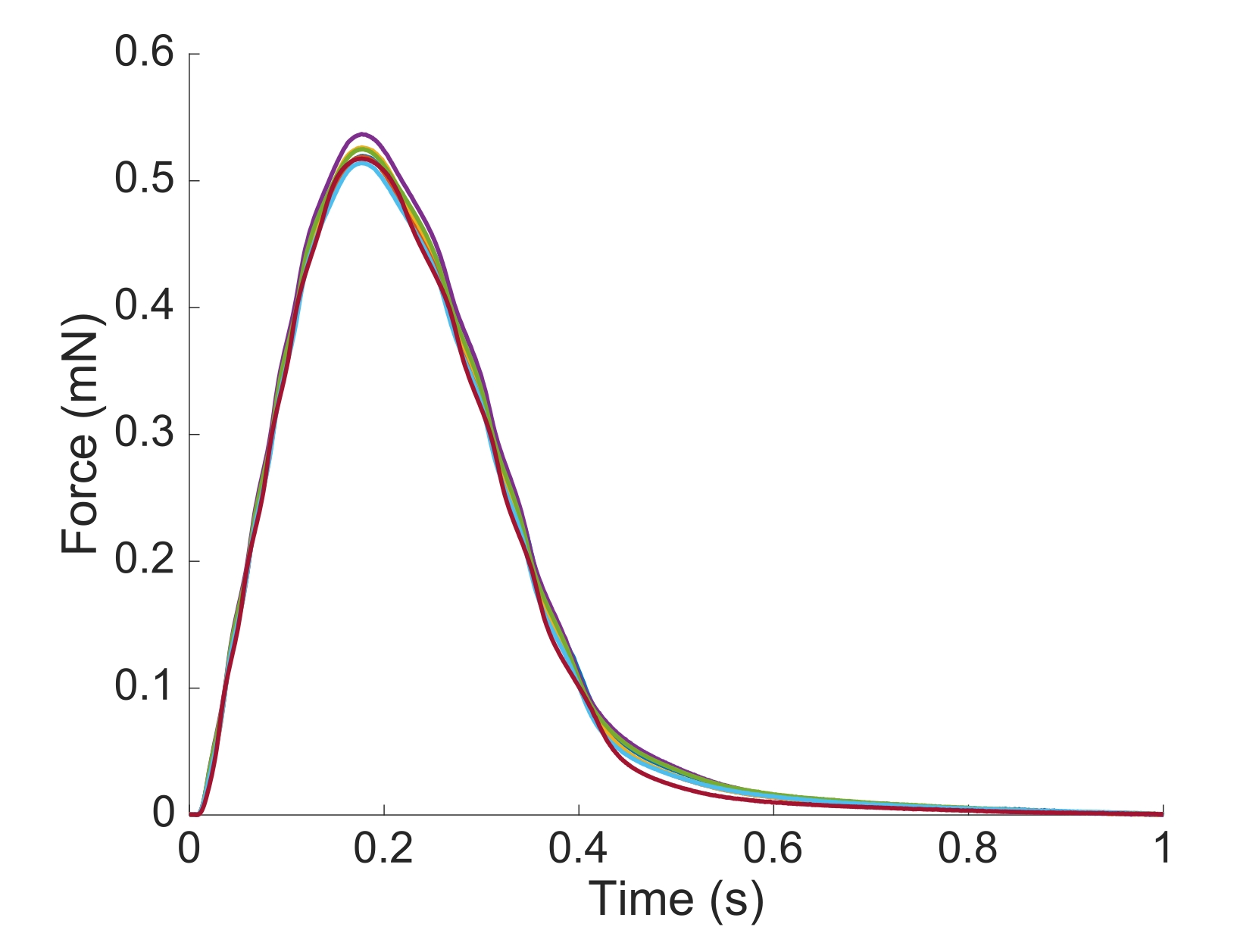
Для того, чтобы захватить региональную информацию Ca2+ и brightfield для всей длины trabecula, представленной здесь, потребовалось семь мышечных позиций. Рисунок 6 предполагает, что сила подергивания не была тронута этим движением, показывая, что не было никакой зависимости положения от производства активной силы.
B-сканы, собранные с помощью оптической когерентной томографии со скоростью 100 Гц, были сегментированы с помощью плагина ImageJ WEKA14 (рисунок 7A). Каждое поперечное сечение выглядит искаженным из-за разницы между боковым (10 мкм) и глубинным (1,73 мкм (в миокарде)) разрешениями. Это искажение было исправлено путем масштабирования оси глубины изображения по соотношению бокового разрешения и глубины разрешения. Рисунок 7B,C демонстрирует, что после масштабирования необработанного C-сканирования трабекулы он приблизительно цилиндрический по геометрии. Отражение стенки измерительной камеры иногда может перекрываться с мышечными данными(рисунок 7A,B),но программное обеспечение сегментации может быть обучено учитывать это(рисунок 7D,E). После сегментировки площадь поперечного сечения по длине мышцы может быть рассчитана на протяжении всего подергивания(рисунок 7F). Отметим, что именно эта трабекула имеет небольшой придаток, ответвляющийся от него. Движение ветви очевидно ~0,75 мм вдоль трабекулы. Наконец, сегментированные изображения могут быть преобразованы в сетки, чтобы помочь в построении геометрических моделей(рисунок 7G).
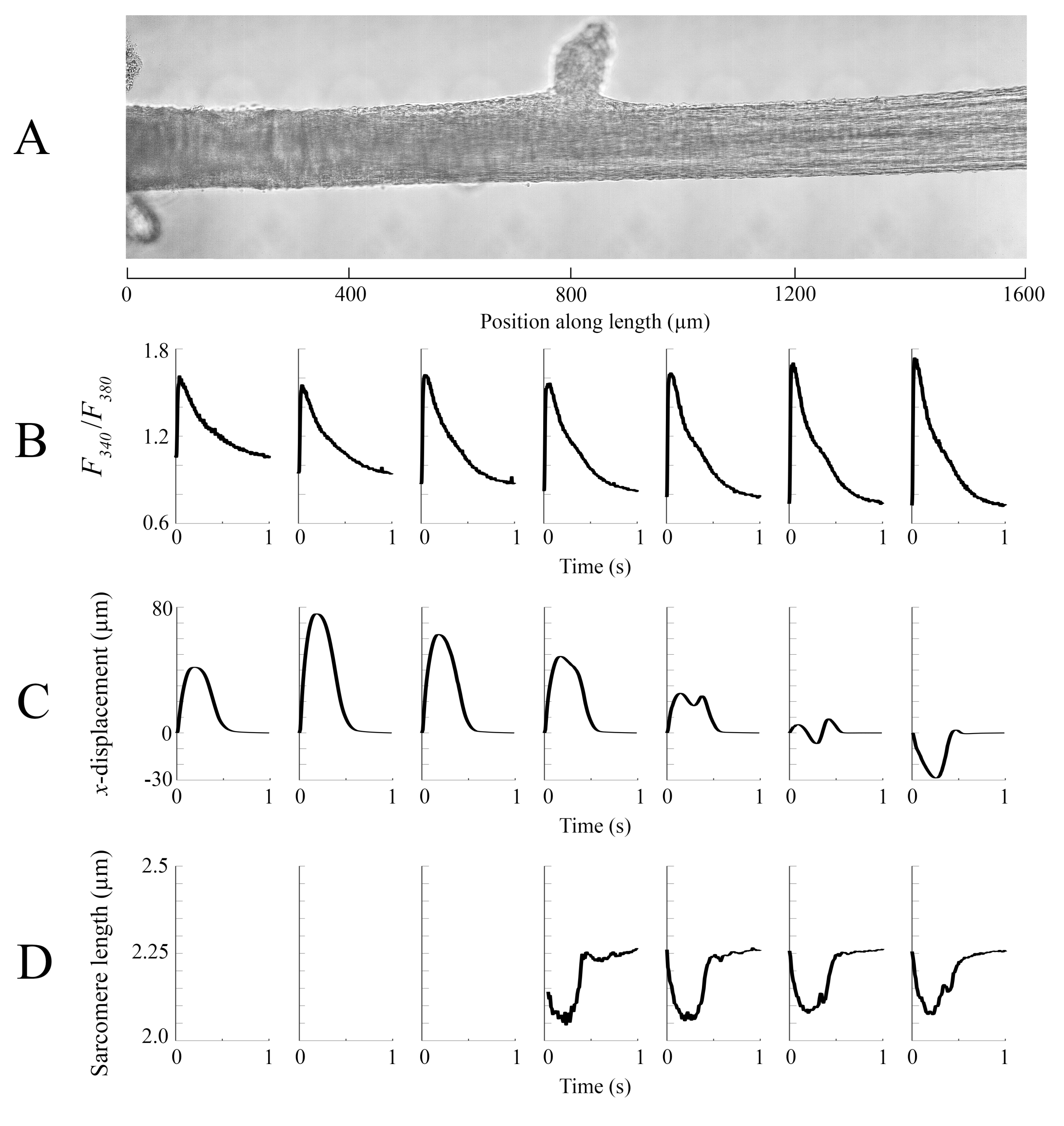
Данные визуализации, полученные в каждом из различных положений трабекулы со скоростью 100 кадров в секунду, были сшиты вместе, чтобы создать единое полное изображение трабекулы(рисунок 8A). Разрешение этих изображений составляет 0,535 мкм/пиксель. Использование функций линейного взвешивания в перекрывающихся областях соседних окон облегчает визуализацию и сводит к минимуму влияние виньетки, присутствующей в изображениях яркого поля. Для измерения флуоресцентного сигнала окно трабекулы размером 540 мкм на 540 мкм циклически освещается светом длин волн 340 нм, 365 нм и 380 нм со скоростью 600 Гц. Отношение излучаемой флуоресценции, связанной со светом возбуждения 340 нм и 380 нм, коррелирует с внутриклеточным кальцием после того, как трабекула была загружена Fura-2. Поскольку это измерение является соотношением, эффективная скорость измерения составляет 200 Гц. Усредненные(n = 10) внутриклеточные переходные ca2+ из каждого окна выравниваются с областью, в которую они были изображены(рисунок 8B). В то время как пик переходных процессов кажется достаточно последовательным, диастолический [Ca2+] ниже в области между 900 мкм и 1800 мкм вдоль трабекулы. Аналогичным образом, результаты расчетов слежения за смещением(рисунок 8С)и длины саркомера(рисунок 8D)также указывают на наличие региональной изменчивости. Используемая техника отслеживания без маркеров способна обрабатывать смещение каждого пикселя, учитывая достаточную контрастность. При отображении распределения длин саркомера в посте для расчета региональной длины саркомера использовалась перекрестная корреляционная область 128 пикселей на 128 пикселей (~67 мкм на 67 мкм). Эта область инкапсулирует примерно 29 саркомер, когда близкий образец близок к оптимальной длине саркомера. Размер шага (как в x-, так и в y-направлениях)между центроидом каждого окна перекрестной корреляции был установлен на 50 пикселей (~26 мкм) для обработки этих данных. Пригодность оценок длины саркомера была проверена на основе ширины и амплитуды гауссовского соответствия сигналу БПФ. Эти условия не были выполнены в мышечной области между 0 мкм и 500 мкм, поэтому там не было вычислено никакой информации о длине саркомера. Учитывая связанные с этим перемещения, вполне вероятно, что саркомы в этом регионе удлинялись во время фазы сокращения. В соответствии с этим предположением, средняя длина саркомера с правой стороны трабекулы сокращается в течение этого периода. Благодаря объединению информации, представленной каждой из панелей, получается, что область с наибольшей площадью поперечного сечения не производит наиболее значительных сил. Исходя из предположения, что региональная вариация переходных процессов Ca2+ имеет приблизительно гладкий градиент, рисунок 8B показывает, что наибольшая амплитуда Ca2+ переходного действия происходит где-то между 1300 мкм и 1600 мкм вдоль трабекулы. Карта смещения показывает, что область, которая подвергается наименьшему движению, хорошо согласуется с пиковым переходным процессом Ca2+. Однако эта область имеет наименьшие области поперечного сечения выборки. Имея в виду эти данные, можно сделать вывод, что этот регион вызвал наибольший стресс.

Рисунок 1:Аннотированное изображение кардиомиометра. Очерчен каждый из основных оптических компонентов. Вставка содержит крупный план, вид сзади, объектива микроскопа in situ,под измерительной камерой. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 2:Оптический путь для одновременной яркой и флуоресцентной микроскопии. Источником освещения для флуоресцентного микроскопа является ксеноновая дуговая лампа, выход которой циклически переключается между 340 нм, 365 нм и 380 нм света. Выходной путь дуговой лампы содержит дихроичное зеркало с длиной волны отсечки 409 нм, которое отражает ультрафиолетовый свет на зеркало, которое направляет свет в объектив флуоресцентного микроскопа. Линза фокусирует свет возбуждения на образце и собирает излучаемый свет, который имеет более длинную длину волны 510 нм. Этот излучаемый свет проходит через первое дихроичное зеркало, но не второе, так как оно имеет длину волны отсечки 552 нм. Затем полевая линза фокусирует отраженный свет на датчике PMT. Между тем, источник освещения (660 нм LED) для микроскопа Brightfield расположен над образцом. Проходящий свет фокусируется на образце конденсаторной линзой, и 20× флуоресцентный объектив захватывает результирующее изображение передачи. Длина волны, используемая для яркого освещения поля, превышает длину волны отсечки каждого дихроичного зеркала, поэтому она проходит через оба из них, прежде чем изображение будет сфокусировано на датчике КМОП-камеры. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 3:Конструкция держателя измерительной камеры. (A)Изометрический вид держателя измерительной камеры с наложителями оптических путей. Яркое освещение происходит с верхней поверхности(z-ось); Флуоресцентное освещение происходит с нижней поверхности (осьz),а сигнал OCT измерительного рычага ортогоналив к другой осиосвещения (ось y). Во время эксперимента два платиновых крючка удерживают трабекулу в стеклянной капиллярной трубке, которая функционирует как измерительная камера. Двигатели с голосовыми катушками управляют каждым крюком, а их положение измеряется с помощью лазерной интерферометрии. Текущее положение сравнивается с заданным пользователем заданным значением и с помощью ПИД-контроллера, закодированного в FPGA, ошибка сводится к минимуму. (B)Измерительная камера in situ с включенной яркой подсветкой. Вид сзади показан на рисунке 1. (C)Схема потока суперфузата через держатель измерительной камеры. Суперсварка поступает в заднюю часть блока и течет в направлении, указанном стрелками. Электроды вверх и вниз по течению устанавливают стимуляцию поля для получения сокращения установленной трабекулы в измерительной камере. Синее затенение указывает на области, где суперсырье течет во время эксперимента. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 4:Передняя панель программного обеспечения для сбора и управления изображениями. (A) Пользовательский интерфейс Brightfield Imaging. (B) Пользовательский интерфейс визуализации OCT. (C) Пользовательский интерфейс аппаратного управления. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 5:Протокол рассечения и монтажа Trabecula. (A)Лангендорф-перфузированное сердце крысы в камере рассечения. (B ) То же самое сердце с удаленными предсердиями. Пунктирные линии указывают траекторию иссечения для открытия желудочков. (C) Открытое сердце для обнажения внутренней части обоих желудочков. Пунктирное поле указывает регион, где обычно встречаются трабекулы. (D)Иссеченная область стенки правого желудочка (такая же, как указано пунктирной графой в С). Пунктирные линии выделяют три трабекулы. (E) Трабекула, выбранная из трех в D. (F) Трабекула из панели E с удаленной тканью стенки. (G) Изолированная трабекула в конце шприца 1 мл. (H) Трабекула в монтажной камере. (I) Трабекула установлена между двумя платиновыми крючками. (J) Трабекула, установленная между крючками, внутри измерительной камеры(рисунок 3B). Зеленое пятно является артефактом из первого дихроичного фильтра. (K)Вторичный угол установленной трабекулы внутри измерительной камеры. Расстояние между трабекулой и объективом микроскопа составляет примерно 1 мм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 6:Позиционная зависимость измерения силы. Сила, создаваемая мышцей из каждого из положений визуализации(n = 7), наложена. Средняя выработка активной силы составила 0,527 мН ± 0,003 мН, время до 50 % сокращения 77,1 мс ± 0,3 мс, а время до 50 % релаксации 328,1 мс ± 0,9 мс (все данные представлены как среднее ± SE). Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 7:Анализ изображений OCT. (A) Пример сегментации WEKA. Сегментарное поперечное сечение мышцы выделено красным цветом, а фон выделен зеленым. (B) Превосходное представление необработанных данных C-сканирования трабекулы. Яркая угловая линия в верхней части изображения является отражением стенки измерительной камеры. (C) Боковой вид необработанных данных C-сканирования трабекулы. (D)Превосходное представление сегментированных данных центра развертывания Office. (E) Боковое представление сегментированных данных центра развертывания Office. (F) Площадь поперечного сечения по длине трабекулы (осьx)вовремени (ось y). Средняя площадь поперечного сечения по длине мышцы составляла 0,0326 мм2 ± 0,0005мм2 (среднее ± S.E.) (G) Сетка трабекулы. Сетка была приблизительно выровнена с участком площади поперечного сечения панели F. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 8:Анализ изображений Brightfield и флуоресцентной визуализации. (A) Сшитое изображение (семь окон визуализации) трабекулы. (B) Ca2+ переходных процессов по длине трабекулы. (C) Среднее x- смещение от каждого окна изображения. Положительное смещение представляет собой движение вправо и отрицательное влево. (D) Средняя длина саркомера от каждого окна изображения, которое имело необходимую контрастность изображения. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Таблица 1: Таблица решений Пожалуйста, нажмите здесь, чтобы загрузить эту таблицу.
Обсуждение
В этом исследовании мы представляем конфигурацию, которая позволяет собирать три оптические системы, сочетающие яркое поле, флуоресценцию и визуализацию OCT, для сбора данных от активно сжимающейся сердечной трабекулы ex vivo (рисунок 1 и рисунок 2). Такое оркестрованное интегрирование возможно благодаря конструкции измерительной камеры(рисунок 3),позволяющей ортогональное расположение ОКТ к оси яркого поля-флуоресценции. Система мышечного крепления играет не менее важную роль в успешности одновременных количественных показателей ключевых показателей в характеристике динамики возбуждения-сокращения сердечной мышцы. Его новизна заключается в том, что он позволяет проводить процедуры сканирования мышц без видимого нарушения механической производительности мышцы(рисунок 6). Благодаря комбинированной конфигурации визуализации и системе моторизованного крючка для измерения силы эта система может оценивать региональную гетерогенность в переходной, смещенной и саркомерной длине Ca2+ вместе с макроскопической геометрической информацией о сжимающейся трабекуле на протяжении всего временного хода подергивания(рисунки 7 и рисунок 8).
Учитывая повсеместное распространенность систем визуализации brightfield-epifluorescence в кардиологических исследовательских лабораториях, воспроизведение этих результатов может быть достигнуто с некоторыми незначительными аппаратными соображениями. Здесь мы представляем инструментарий обработки изображений для объединения ярко-эпифлуоресценции и OCT, что имеет важное значение для анализа лежащей в основе сократительной гетерогенности. Интеграция OCT требует беспрепятственного оптического пути, в то время как закрытая визуализация требует внешней триггерной линии между стимулом и камерой визуализации OCT и Brightfield, а также крючков для крепления мышц, которые способны перемещать образец по всей измерительной камере. Необходимое программное обеспечение и методы постобработки находятся в свободном доступе. В частности, используемое программное обеспечение сегментации, WEKA14,является открытым исходным кодом. Метод безмаркерного отслеживания точек материала8,длины саркомера, закрытой объемной визуализации10и кодов генерации сетки также доступен и может быть предоставлен по запросу соответствующего автора.
Мышечная жизнеспособность, оптимальная нагрузка Fura-2 и фокус изображения — вот три столпа, которые составляют основу успешного эксперимента. Использование раствора для рассечения, содержащего БДМ, для предотвращения контрактуры, транспортировки мышцы в шприце, непрерывной оксигенации раствора и приготовления новых экспериментальных растворов в день эксперимента , все это способствует высокой скорости жизнеспособности мышц. Перед загрузкой трабекулы Fura-2AM необходимо собрать автофлуоресценцию для каждого состояния, которое интересно изучить, поскольку она может оказать значительное влияние на измеренный переходный процесс Ca2+ 15. Оксигенация нагрузочного раствора Фура-2АМ осложняется необходимым включением поверхностно-активного раствора плуроника-Ф127 для облегчения загрузки красителя. Для борьбы с образованием избыточных пузырьков, вызванных этим поверхностно-активным веществом, небольшая капля антипены в нагрузочном растворе позволяет пользователю увеличить скорость оксигенации, тем самым повышая вероятность того, что трабекула сохраняет функциональную жизнеспособность на протяжении всего процесса загрузки. Наконец, фокус изображения должен быть равномерным по длине мышцы, чтобы максимизировать отношение сигнал/шум информации о ярком поле и флуоресценции.
Есть два ограничения, которые следует учитывать при отношении методов, представленных здесь. Во-первых, это пространственное разрешение флуоресцентного микроскопа. В то время как пространственные разрешения OCT и изображения яркого поля высоки, разрешение флуоресцентного микроскопа ограничено интегралом флуоресценции от объема, захваченного в окне визуализации 540 мкм на 540 мкм. Существует возможность увеличения пространственного разрешения флуоресцентного микроскопа за счет использования камеры с высоким коэффициентом усиления зарядовой связи вместо PMT для захвата флуоресцентного сигнала за счет отношения сигнал/шум16. Во-вторых, диаметр трабекулы, который можно изучать с точки зрения измеримой длины саркомера и геометрической глубины. Подход с оконным БПФ для вычисления длины саркомера использует преимущество улучшенного пространственного разрешения, но связан с уменьшением надежности(рисунок 8D). В тех случаях, когда необходимо изучить мутные или большие трабекулы, рассасываемость БПФ будет значительно снижена из-за снижения контраста, связанного с саркомерическим полосированием в более крупных образцах тканей. Аналогичным образом, в пределах ОКТ обратные отражения от глубины изображения более 300 мкм будут слишком слабыми, чтобы их можно было разрешить на этапе сегментации. Следовательно, наша техника ограничена трабекулами диаметром менее 300 мкм. Однако не рекомендуется изучать образцы большого диаметра, так как могут возникнуть проблемы с диффузной оксигенацией мышечного ядра во время высоких показателей стимуляции17.
Наш метод позволяет оценить ионную механическую функцию в сочетании с геометрией мышц в здоровых и больных мышцах, обеспечивая мощный подход к пониманию физиологии сердечной мышцы, патофизиологии и фармакологии. Конвейер обработки изображений, описанный здесь, извлекает данные, которые будут иметь решающее значение для более глубокого понимания сократительной гетерогенности. Одним из способов полной реализации потенциала такого богатого набора данных является построение математических моделей, которые интегрируют и интерпретируют эти данные, и делать прогнозы, которые могут быть проверены экспериментально с помощью нашего устройства.
Раскрытие информации
Авторам нечего раскрывать.
Благодарности
Это исследование финансировалось докторскими стипендиями Оклендского университета (присуждаемыми JD и MC), стипендиями сэра Чарльза Геркуса (20/011 и 21/116) от Совета по исследованиям в области здравоохранения Новой Зеландии (присуждаемыми J-CH KT, соответственно), докторской стипендией, присуждаемой Национальным фондом сердца (присуждаемой AA), грантами Marsden Fast-Start (UOA1504 и UOA1703) от Королевского общества Новой Зеландии (присуждается J-CH и KT, соответственно) и исследовательскую стипендию Джеймса Кука от Королевского общества Новой Зеландии (присуждается AT). Первоначальная разработка этого инструмента финансировалась за счет гранта Марсдена (11-UOA-199) от Королевского общества Новой Зеландии (присужден AT и PN).
Материалы
| Name | Company | Catalog Number | Comments |
| 2,3-Butanedione monoxime | Acros Organics | 150375000 | |
| 20× microscope lens | Nikon | CFI Super Fluor 20× | NA 0.75 |
| 2D Galvanometer | Thorlabs | GVSM002/M | |
| 50-50 beam splitter | Thorlabs | FC850-40-50-APC | |
| 90-10 beam-splitter | Thorlabs | TW850R2A2 | |
| Analogue input module | National Instruments | NI-9205 | Records the PMT signal at 200 kHz |
| Brightfield imaging light source | CoolLED | PE-2 | 660 nm LAM |
| Broadband light source | Superlum | Broadlighter-840 | |
| CaCl2 | Sigma-Aldrich | C4901 | |
| Cameralink card | National Instruments | NI-1429 | Brightfield imaging frame grabber |
| Carbogen 5 | BOC | Gas code: 181 | |
| Condensor lens | Nikon | LWD 0.52 | |
| D(+)-Glucose | Merck | 108337 | |
| DAQ | National Instruments | NI-6259 | Triggers the galvanometer movement |
| Dichroic mirror 1 | Semrock | FF409-Di03 | |
| Dichroic mirror 2 | Semrock | FF552-Di02 | |
| Diffraction grating | Wasatch Photonics | 1200 lines/mm @840 nm | |
| Dimethyl sulfoxide | Sigma-Aldrich | 276855 | |
| Direct-Q 3 UV System | Merck Millipore | ZRQSVR3WW | Distilled water machine |
| Dry bath | Corning | 6875-SB | LSE digital dry bath |
| FIJI | ImageJ | Open-source image processing software | |
| Fura-2AM pentapotassium salt | Thermofisher | F14186 | |
| Hardware FPGA card | National Instruments | NI-7813R | Also controls the triggering of the brightfield capture |
| Heparin | Pfizer | 61024 | |
| HEPES | PanReac AppliChem | A1069 | |
| Inverted microscope | Nikon | TI-DH illumination pillar | |
| Isofluorane | MedSource | VAPDRUGISO250 | |
| KCl | Sigma-Aldrich | P9541 | |
| KH2PO4 | Sigma-Aldrich | P5655 | |
| Line-scan camera | Basler | spL2048-70km | Spectrometer camera |
| Magnetic stirrer | IKA | 3810000 | RCT basic |
| Matlab | Mathworks | Data processing code | |
| MgCl2 | Sigma-Aldrich | M8266 | |
| MgSO4.7H2O | Sigma-Aldrich | M1880 | |
| NaCl | Sigma-Aldrich | 71376 | |
| NaH2PO4.2H2O | Sigma-Aldrich | 71505 | |
| NaHCO3 | Sigma-Aldrich | S6014 | |
| OCT FPGA card | National Instruments | NI-1483R | |
| Oxygen tank | BOC | Gas code: 100D | |
| pH meter | Mettler Toledo | MP220 | |
| Photomultiplier tube | Hamamatsu | H7422-20 | |
| Powerload | Thermofisher | P10020 | |
| Superluminescent diode | Broadlighter | D-840 | |
| Transimpedance amplifier | Custom | ||
| Tris(hydroxymethyl)amino-methane | Sigma-Aldrich | 252859 | |
| Wistar rat | Vernon Jansen Unit | 8 – 10 weeks | |
| Xenon arc lamp | Sutter Instrument | DG-4 | Lambda DG-4 |
Ссылки
- Han, J. -C., et al. Energetics of stress production in isolated cardiac trabeculae from the rat. American Journal of Physiology. Heart and circulatory physiology. 299 (5), 1382-1394 (2010).
- Ter Keurs, H. E. D. J., Rijnsburger, W. H., Van Heuningen, R., Nagelsmit, M. J. Tension development and sarcomere length in rat cardiac trabeculae. Evidence of length-dependent activation. Circulation Research. 46 (5), 703-714 (1980).
- Shen, X., Cannell, M. B., Ward, M. L. Effect of SR load and pH regulatory mechanisms on stretch-dependent Ca2+ entry during the slow force response. Journal of Molecular and Cellular Cardiology. 63, 37-46 (2013).
- Dowrick, J. M., et al. The slow force response to stretch: Controversy and contradictions. Acta Physiologica. 226 (1), 13250(2019).
- Stuyvers, B. D. M. Y., Miura, M., Ter Keurs, H. E. D. J. Diastolic viscoelastic properties of rat cardiac muscle; involvement of Ca2+. Advances in Experimental Medicine and Biology. 430, 13-28 (1997).
- Tang, E. J. L. P., Laven, R. C., Hajirassouliha, A., Nielsen, P. M. F., Taberner, A. J. Measurement of displacement in isolated heart muscle cells using markerless subpixel image registration. Conference Record - IEEE International Instrumentation and Measurement Technology Conference. , (2019).
- Bers, D. M. Cardiac excitation-contraction coupling. Nature. 415 (6868), 198-205 (2002).
- Cheuk, M. L., et al. A method for markerless tracking of the strain distribution of actively contracting cardiac muscle preparations. Experimental Mechanics. 61 (1), 95-106 (2020).
- Lippok, N., Coen, S., Nielsen, P., Vanholsbeeck, F. Dispersion compensation in Fourier domain optical coherence tomography using the fractional Fourier transform. Optics Express. 20 (21), 23398(2012).
- Cheuk, M. L., et al. Four-Dimensional imaging of cardiac trabeculae contracting in vitro using gated OCT. IEEE Transactions on Biomedical Engineering. 64 (1), 218-224 (2017).
- Ritland, H. N. Relation between refractive index and density of a glass at constant temperature. Journal of the American Ceramic Society. 38 (2), 86-88 (1955).
- Tuchina, D. K., Bashkatov, A. N., Genina, E. A., Tuchin, V. V. Quantification of glucose and glycerol diffusion in myocardium. Journal of Innovative Optical Health Sciences. 8 (3), (2015).
- Taberner, A., et al. A dynamometer for nature's engines. IEEE Instrumentation and Measurement Magazine. 22 (2), 10-16 (2019).
- Arganda-Carreras, I., et al. Trainable Weka Segmentation: A machine learning tool for microscopy pixel classification. Bioinformatics. 33 (15), 2424-2426 (2017).
- Jiang, Y., Julian, F. J. Pacing rate, halothane, and BDM affect fura 2 reporting of [Ca2+](i) in intact rat trabeculae. American Journal of Physiology - Cell Physiology. 273 (6), 2046-2056 (1997).
- Miura, M., Boyden, P. A., Ter Keurs, H. E. D. J. Ca2+ waves during triggered propagated contractions in intact trabeculae. American Journal of Physiology - Heart and Circulatory Physiology. 274 (1), (1998).
- Han, J. -C., et al. Radius-dependent decline of performance in isolated cardiac muscle does not reflect inadequacy of diffusive oxygen supply. American Journal of Physiology-Heart and Circulatory Physiology. 300 (4), 1222-1236 (2011).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены