
Для просмотра этого контента требуется подписка на Jove Войдите в систему или начните бесплатную пробную версию.
Method Article
Непрерывный флуоресцентный анализ метилирования ДНК на основе эндонуклеазы для скрининга ингибиторов ДНК-метилтрансферазы
В этой статье
Резюме
ДНК-метилтрансферазы являются потенциальными мишенями лекарств от рака. Здесь представлен протокол для оценки малых молекул ингибирования ДНК-метилтрансферазы. Этот анализ использует эндонуклеазу для соединения метилирования ДНК с генерацией флуоресценции и позволяет контролировать активность ферментов в режиме реального времени.
Аннотация
Метилирование ДНК, форма эпигенетической регуляции генов, важно для нормальной клеточной функции. В клетках белки, называемые ДНК-метилтрансферазами (DDNMT), устанавливают и поддерживают паттерн метилирования ДНК. Изменения в нормальном паттерне метилирования ДНК связаны с развитием и прогрессированием рака, что делает DDNMT потенциальными мишенями для лекарств от рака. Таким образом, выявление и характеристика новых низкомолекулярных ингибиторов этих ферментов имеет большое значение. В этой статье представлен протокол, который может быть использован для скрининга ингибиторов ДНК-метилтрансферазы. Непрерывный связанный кинетический анализ позволяет определить начальные скорости метилирования ДНК в присутствии и отсутствии потенциальных ингибиторов малых молекул. Анализ использует метил-чувствительную эндонуклеазу Gla I для соединения метилирования гемиметилированного субстрата ДНК с генерацией флуоресценции.
Этот непрерывный анализ позволяет контролировать активность ферментов в режиме реального времени. Проведение анализа в небольших объемах в микротитрных пластинах снижает стоимость реагентов. Используя этот анализ, был проведен небольшой пример скрининга для ингибиторов DNMT1, наиболее распространенного изофермента DNMT у людей. Высокозамещенный натуральный продукт антрахинона, лаккаевая кислота А, является мощным, конкурентоспособным по ДНК ингибитором DNMT1. Здесь мы исследуем три потенциальных ингибитора малых молекул — антрахиноны или антрахиноподобные молекулы с одним-тремя заместителями — в двух концентрациях, чтобы описать протокол анализа. Начальные скорости используются для расчета процентной активности, наблюдаемой в присутствии каждой молекулы. Одно из трех исследованных соединений демонстрирует концентрационно-зависимое ингибирование активности DNMT1, что указывает на то, что оно является потенциальным ингибитором DNMT1.
Введение
Метилирование ДНК является важным эпигенетическим маркером, который регулирует экспрессию генов и структуру хроматина. Метилирование происходит преимущественно в cpG динуклеотидах — цитозине, за которым следует гуанозин; метильную группу добавляют к 5-позиции цитозина. Правильные паттерны метилирования ДНК и, следовательно, правильная экспрессия генов необходимы для соответствующего клеточного развития и функционирования. Многие болезненные состояния были связаны с изменениями нормальной картины метилирования 1,2,3. Например, существует связь между инициацией и прогрессированием рака и изменениями в паттерне метилирования ДНК. Как правило, раковые клетки демонстрируют более низкие общие уровни метилцитозина, что способствует нестабильности генома. При этом метилцитозин, присутствующий в геноме, концентрируется в промоторных областях генов-супрессоров опухолей, что приводит к глушению генов этих важных белков. Примечательно, что эпигенетические изменения являются динамичными и обратимыми, в отличие от мутаций ДНК, связанных с опухолевым генезом. Это сделало белки, участвующие в эпигенетической регуляции генов, интересными лекарственными мишенями 2,4.
ДНК-метилтрансферазы (DDNMT) являются белками, ответственными за генерацию и поддержание паттернов метилирования ДНК. Три каталитически активных изофермента, DNMT1, DNMT3a и DNMT3b, существуют у людей. Во время разработки и дифференцировки de novo метилтрансферазы, DNMT3a и DNMT3b, устанавливают паттерны метилирования. Оба фермента могут связывать каталитически неактивный белок DNMT3L с образованием комплексов, которые проявляют повышенную активность 1,5. После деления клеток дочерние клетки содержат гемиметилированную ДНК — ДНК, содержащую метилцитозин только в одной нити дуплекса — потому что вновь синтезированная ДНК лишена меток метилирования. Основная функция DNMT1 заключается в метилировании этой гемиметилированной ДНК, тем самым восстанавливая полную картину метилирования 1,5.
Связи между активностью DNMT и раком хорошо известны. Сверхэкспрессия DNMT1, либо транскрипционными, либо посттрансляционными механизмами, является следствием нескольких общих онкогенных путей 6,7,8,9. Генетические подходы к снижению активности DNMT1 с использованием гипоморфных аллелей приводят к уменьшению образования опухоли у мышей Apc(Min)10. Антисмысловые олигонуклеотиды, которые нокдают DNMT1, ингибируют неоплазию в клеточной культуре и моделях опухолей мышей 11,12. Таким образом, ингибирование активности DNMT1 кажется многообещающим подходом к терапии рака. Однако роли, которые играют изоферменты DNMT3, не так просты. Мутации DNMT3a обнаруживаются при остром миелоидном лейкозе13 и миелодиспластическом синдроме14. Было показано, что по меньшей мере одна из идентифицированных мутаций снижает активность метилирования ДНК фермента15. Тем не менее, DNMT3b чрезмерно экспрессируется при раке молочной железы16 и колоректальном раке17. Поскольку различные изоферменты DNMT играют различную роль в канцерогенезе, идентификация изофермент-специфических ингибиторов будет иметь решающее значение. Мало того, что эти соединения будут полезны для разработки терапевтических средств, но изофермент-специфические ингибиторы также будут бесценным инструментом для анализа роли каждого изофермента DNMT в этиологии рака.
В литературе сообщалось о нескольких ингибиторах DNMT. Известные ингибиторы DNMT можно разделить на два класса: нуклеозидные и ненуклеозидные. Нуклеозидные ингибиторы обычно являются аналогами цитидина. Эти соединения включены в ДНК и ковалентно улавливают DNMTs. 5-азацитидин и 5-аза-2'-дезоксицитидин были одобрены для лечения миелодиспластического синдрома и острого миелоидного лейкоза 4,18. Высокая токсичность, низкая биодоступность и химическая нестабильность этих соединений представляют проблемы. Текущая работа изучает эффективность следующего поколения нуклеозидных ингибиторов; SGI-110, полученный из 5-аза-2'-дезоксицитидина, является одним из примеров19,20. Нуклеозидные ингибиторы не являются изофермент-специфическими и инактивируют любой встречающийся изофермент DNMT. Таким образом, обработка нуклеозид-деметилирующим агентом приводит к истощению всех изоферментов DNMT 4,18. Ненуклеозидные ингибиторы не нужно включать в ДНК, чтобы оказывать их ингибирующее действие. Вместо этого эти молекулы связываются непосредственно с DDNMT, вводя возможность для изофермент-специфического ингибирования. На сегодняшний день было обнаружено несколько ненуклеозидных ингибиторов, включая SGI-102721, гидралазин22, прокаинамид23, RG108 и производные24, а также натуральные продукты, (-)-эпигаллокатехин 3-галлат (EGCG)25 и лаккаевую кислоту A26,27. Большинство ненуклеозидных ингибиторов, обнаруженных на сегодняшний день, не являются изофермент-селективными или демонстрируют слабые предпочтения для одного изофермента DNMT. Кроме того, необходимо улучшить эффективность этих молекул, особенно в клетках 4,18. Таким образом, существует необходимость в открытии или разработке более мощных, изофермент-селективных ингибиторов DNMT.
Препятствием для открытия новых низкомолекулярных ингибиторов DNMT являются трудоемкие анализы, традиционно используемые для изучения активности DNMT28. Анализы обычно прерывистые с несколькими шагами. Ферментативная активность DNMT по-прежнему регулярно анализируется с использованием радиоактивного S-аденозилметионина (SAM)29,30,31,32,33,34. Также были разработаны нерадиоактивные анализы метилирования ДНК. Например, описаны анализы с использованием метилчувствительных эндонуклеаз рестрикции и электрофореза для разделения продуктов пищеварения35,36. Эти типы прерывистых, многоступенчатых анализов не поддаются открытию лекарств. С середины 2000-х годов было разработано несколько анализов метилирования ДНК с более высокой пропускной способностью28. Сцинтилляционный бесконтактный анализ использовался для скрининга ингибиторов DNMT137. Другой анализ с использованием метил-чувствительной рестрикционной эндонуклеазы был использован для скрининга ингибиторов DNMT3a25,38. Хотя оба анализа позволили получить более высокую пропускную способность, чем традиционные анализы метилирования ДНК, анализы требуют нескольких этапов и не позволяют наблюдать активность метилирования в режиме реального времени. Совсем недавно был описан непрерывный кинетический анализ, который связывает образование S-аденозилгомоцистеина (SAH), одного из продуктов реакции метилирования, со спектроскопическим изменением при 340 нм, связанным с окислением NADPH39. Этот анализ использует три фермента связи для генерации спектроскопического сигнала.
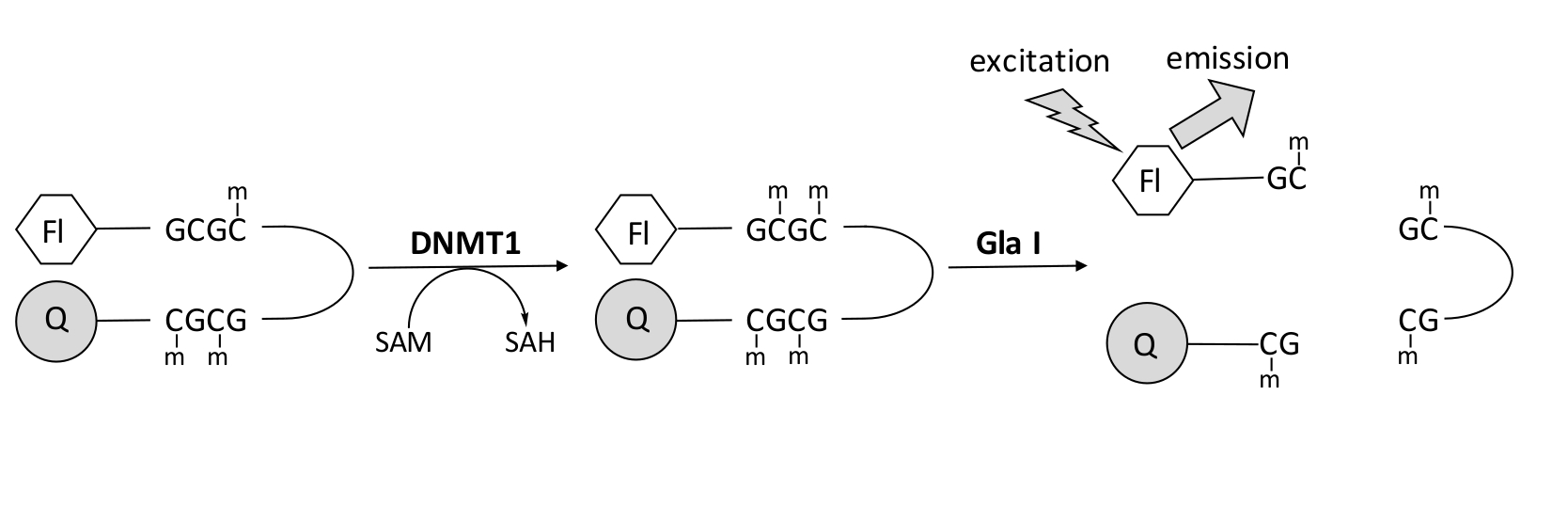
Мы разработали анализ метилирования ДНК на основе флуоресценции, связанный с эндонуклеазой, который использует один коммерчески доступный фермент связи и может генерировать данные в режиме реального времени (рисунок 1). В качестве субстрата используется олигонуклеотид шпильки, содержащий три метилцитозина. СУБСТРАТНАЯ ДНК содержит флуорофор на 5' конце и гаситель на 3' конце. Метилирование гемиметилированного сайта CpG генерирует участок расщепления для эндонуклеазы Gla I — полностью метилированного GCGC. Расщепление продукта олигонуклеотид gla I высвобождает флуорофор из гасителя и генерирует флуоресценцию в режиме реального времени. Анализ может быть использован для изучения активности любой изоформы DNMT; однако более высокая активность наблюдается при DNMT1, так как этот изофермент преимущественно метилирует гемиметилированную ДНК 1,5. Еще более устойчивая активность наблюдается, если домен автоингибаторной последовательности таргетирования фокусов репликации (RFTS) удаляется из DNMT1. Этот домен, обнаруженный в N-концевой регуляторной области, связывается с каталитическим сайтом и предотвращает связывание ДНК. Удаление первых ~ 600 аминокислот приводит к усеченному ферменту, который значительно более активен, чем фермент полной длины (~ 640-кратное увеличение kcat/Km)40. Эта активированная форма фермента, называемая RFTS-отсутствующим DNMT1 (аминокислоты 621–1616), позволяет легче идентифицировать ингибиторы из-за его повышенной каталитической мощности. В этой статье представлен протокол для использования RFTS-отсутствующего DNMT1 в анализах для скрининга потенциальных ингибиторов малых молекул. С помощью непрерывного анализа, связанного с эндонуклеазой, начальную скорость определяют в присутствии и отсутствии нескольких малых молекул. Каждый потенциальный ингибитор исследуется в двух концентрациях для поиска концентрационно-зависимого ингибирования DNMT1. Процент активности, наблюдаемый в присутствии малых молекул, был рассчитан в каждом случае.

Рисунок 1: Анализ метилирования ДНК. В качестве субстрата используется гемиметилированная шпилька ДНК с флуорофором на 5' конце и гасителем на 3' конце. DNMT1 катализирует перенос метильной группы из S-аденозилметионина в неметилированный сайт CpG, генерируя S-аденозилгомоцистеин и полностью метилированную ДНК. Продукт ДНК содержит участок расщепления эндонуклеазы Gla I, которая расщепляет полностью метилированные участки GCGC. Расщепление ДНК продукта высвобождает 5' флуорофор из 3' гасителя, генерируя флуоресценцию. Сокращения: Fl = флуорофор; Q = quencher; DNMT1 = ДНК-метилтрансфераза 1; SAM = S-аденозилметионин; SAH = S-аденозилгомоцистеин. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
протокол
1. Подготовьте пробирные решения для экрана
ПРИМЕЧАНИЕ: Концентрации субстратов, используемых в этом анализе, могут быть адаптированы. Для RFTS-отсутствующего DNMT1 экспериментальноопределенные значения K m для субстрата ДНК шпильки и SAM составляют 1–2 нМ и 2 мкМ соответственно26,40.
- Подготовьте 600 мкл каждого условия анализа, тестируемого в микроцентрифужных пробирках на льду.
ПРИМЕЧАНИЕ: Общий объем необходимого аналитического раствора зависит от количества проводимых реплик. Каждое условие анализа выполнялось в трех экземплярах для этого протокола.- Добавьте ddH2O и 5x буфер метилирования (250 мМ Tris pH 7,5, 5 мМ MgCl2, 500 мМ глутамата калия, 5 мМ дитиотрейтола (DTT), 25% глицерина) в каждую трубку для достижения конечной концентрации 1x буфера. Для анализов, содержащих соединение 20 мкМ, добавляют 472 мкл ddH2O и 120 мкл 5x буфера. Для анализов, содержащих соединение 50 мкМ, добавляют 470 мкл ddH2Oи 120 мкл 5x буфера.
- Добавьте 3,15 мкл 20 мг/мл бычьего сывороточного альбумина (BSA) к каждому образцу.
- Добавьте 2 мкл субстрата ДНК шпильки 3,15 мкМ и 1,33 мкл 4,75 мМ SAM к каждому образцу.
ПРИМЕЧАНИЕ: Концентрация субстратов в смеси пробирного раствора в 1,05 раза превышает желаемые конечные концентрации. - Добавьте ингибитор к конечной концентрации 21 мкМ (1,26 мкл 10 мМ) или 52,5 мкМ (3,15 мкл 10 мМ) к соответствующим образцам. Добавьте эквивалентное количество диметилсульфоксида (ДМСО) в контрольный образец.
ПРИМЕЧАНИЕ: Концентрация ингибитора, используемого в экране, может варьироваться. Максимальная конечная концентрация ДМСО должна составлять ≤5% (v/v). Концентрации ДМСО до 5% (v/v) не влияют на активность, наблюдаемую в анализе26.
- Перемешайте растворы путем вихря (3 000 об/мин) в течение 3 с. Вращайте образцы в течение нескольких секунд в настольной мини-центрифуге (1 200 × г), чтобы убедиться, что раствор собран в нижней части трубки.
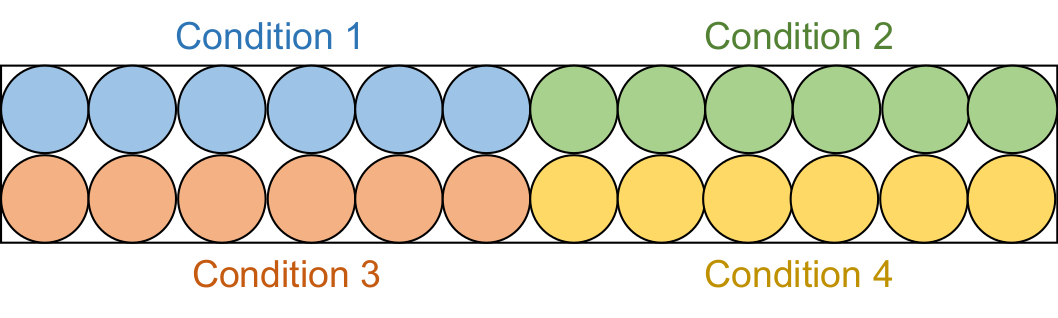
- Аликвота 95 мкл каждого пробирного раствора в 6 последовательных скважин в черной половинной площади 96-луночной пластины (рисунок 2).
ПРИМЕЧАНИЕ: Эти скрининговые анализы проводились в двух партиях. Сначала были исследованы образцы ингибиторов 20 мкМ (4 общих условия), а затем образцы ингибиторов 50 мкМ (4 общих условия).

Рисунок 2: Установка пробирной пластины. Каждый пробирный раствор распределяется на шесть скважин в черной 96-луночной пластине: контроль DMSO (синий), соединение 1 (зеленый), соединение 2 (красный) и соединение 3 (желтый). Оба DNMT1 и Gla I, не имеющие RFTS, будут добавлены к трем скважинам. В качестве контроля, Gla I один будет добавлен к другим трем скважинам. Сокращения: RFTS = последовательность таргетинга фокусов репликации; DNMT1 = ДНК-метилтрансфераза 1; ДМСО = диметилсульфоксид. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
2. Подготовьте ферментные растворы для экрана
ПРИМЕЧАНИЕ: RFTS-отсутствующий DNMT1 может быть экспрессирован в E. coli и очищен до однородности. Процедуры экспрессии и очистки для RFTS-отсутствующего DNMT1 были описаны ранее41. Объем необходимого фермента зависит от количества проводимых анализов. Здесь в каждом наборе выполняются четыре различных анализа; каждый анализ выполняется в трех экземплярах.
- Готовят по 75 мкл каждого раствора фермента в микроцентрифужных пробирках на льду.
ПРИМЕЧАНИЕ: Потребуется два раствора ферментов. Одно решение будет содержать DNMT1 и Gla I; другой будет содержать только Gla I.- Добавьте ddH2O и 5x буфер метилирования (250 мМ Tris pH 7,5, 5 мМ MgCl2, 500 мМ глутамата калия, 5 мМ DTT, 25% глицерина) в каждую пробирку для достижения конечной концентрации 1x буфера. Для раствора фермента Gla I добавьте 15 мкл 5x буфера и 58,8 мкл ddH2O. Для раствора фермента DNMT1+Gla I добавьте 15 мкл 5x буфера и 58,2 мкл ddH2O.
- Добавьте 1,2 мкл 10 Ед/мкл Gla I к каждому раствору для конечной концентрации 0,16 ЕД/мкл.
- Добавьте 0,6 мкл 5 мкМ RFTS-без DNMT1 для конечной концентрации 40 нМ в раствор DNMT1+Gla I.
- Осторожно нажмите, чтобы перемешать растворы. Вращайте образцы в течение нескольких секунд в настольной мини-центрифуге (1 200 × г), чтобы убедиться, что раствор собран в нижней части трубки.
- Аликвота 12 мкл каждого раствора фермента в 6 лунок в коническом дне 96-луночной пластины (рисунок 3).

Рисунок 3: Настройка ферментной пластины. Растворы Gla I (серый) и DNMT1+Gla I (синий) распределены по шести скважинам в 96-луночной плите. Используя многоканальный пипет, фермент можно добавлять в ряд пробирных растворов одновременно. Для каждого условия анализа (шесть скважин) три скважины получат DNMT1 + Gla I, а три скважины получат Gla I в одиночку. Аббревиатура: DNMT1 = ДНК метилтрансфераза 1. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
3. Запустите анализ и проанализируйте данные.
- Разогрейте пластинчатый считыватель до 37 °C. Вставьте черную пластину, содержащую пробирные растворы. Встряхните пластину (двойную орбиталь при 425 cpm в течение 5 с) и считывайте флуоресценцию с длиной волны возбуждения 485 нм и длиной волны излучения 528 нм. Инкубировать пластину при 37 °C в течение 5 мин.
ПРИМЕЧАНИЕ: В качестве альтернативы для флуоресцентных показаний могут использоваться фильтры с надлежащими отсечками длины волны. - Снимите пробирную пластину. Добавьте 5 мкл раствора фермента в каждую лунку и перемешайте путем пипетки вверх и вниз.
ПРИМЕЧАНИЕ: Используйте многоканальные пипетки, чтобы ряд образцов мог быть инициирован одновременно. - Вставьте пластину в устройство считывания пластин. Записывайте флуоресценцию каждые 53 с в течение 30 мин. Встряхивайте пластину (двойную орбиталь при 425 cpm в течение 4 с) перед каждым чтением.
- Среднее утроение контрольных анализов Gla I для каждого состояния. Вычтите средний след управляющей реакции Gla I, содержащий из тройных следов, полученных в присутствии RFTS-отсутствующего DNMT1. Определите среднюю и стандартную погрешность среднего значения для исправленных реплик.
- Постройте среднюю скорректированную след реакции и поднесите начальную линейную часть к линии, чтобы определить начальную скорость.
- Определить процент активности, разделив скорость, наблюдаемую в присутствии соединения, на скорость, наблюдаемую в ДМСО-содержащей управляющей реакции, и умножив на 100.
4. Дополнительный контроль анализа — последовательное добавление ферментов
- Приготовьте 550 мкл пробирного раствора в микроцентрифужной трубке на льду. Добавьте 110 мкл 5-кратного метилирующего буфера, 3,88 мкл 3,15 мкМ субстрата ДНК шпильки, 6,43 мкл 47,5 мМ SAM, 3,06 мкл 20 мг/мл BSA и 426,6 мкл ddH2O.
- Перемешайте раствор вихрем (3 000 об/мин) в течение 3 с. Вращайте образцы в течение нескольких секунд в настольной мини-центрифуге (1 200 × г), чтобы убедиться, что раствор собран в нижней части трубки.
- Аликвота 90 мкл пробирного раствора в 6 последовательных скважин в черной полупространственной 96-луночной пластине.
- Приготовьте растворы ферментов DNMT1 на льду. Добавьте 4 мкл 5-кратного метилирующего буфера, 0,76 мкл 5 мкМ RFTS-отсутствия DNMT1 и 15,24 мкл ddH2Oв одну трубку. Добавьте 4 мкл 5-кратного буфера метилирования и 16 мкл ddH2Oв другую трубку.
- Осторожно нажмите, чтобы перемешать растворы. Вращайте образцы в течение нескольких секунд в настольной мини-центрифуге (1 200 × г), чтобы убедиться, что раствор собран в нижней части трубки.
- Добавьте 5 мкл раствора, содержащего DNMT1, в три скважины в черной половинной площади 96-луночной пластины. Добавьте 5 мкл буферного контрольного раствора в остальные три скважины.
- Поместите микротитровую пластину в считыватель пластин, предварительно нагретый до 37 °C. Встряхните пластину (двойную орбиталь при 425 cpm в течение 3 с) и считывайте флуоресценцию с длиной волны возбуждения 485 нм и длиной волны излучения 528 нм. Повторяйте встряхивание и читайте каждые 60 с в течение 30 минут.
- Пока пробирная пластина находится в считывателе пластин, приготовьте раствор Gla I на льду. Добавьте 8 мкл 5-кратного метилирующего буфера, 0,64 мкл 10 Ед/мкл Gla I и 31,4 мкл ddH2Oв микроцентрифужную трубку. Осторожно постучите, чтобы перемешать раствор. Вращайте образец в течение нескольких секунд в настольной мини-центрифуге (1 200 × г), чтобы убедиться, что раствор собран в нижней части трубки.
- Аликвота 6 мкл раствора Gla I в 6 скважинах в коническом дне 96-луночной плиты.
- После начального 30-минутного считывания снимите черную пластину со считывателя пластин. Добавьте 5 мкл раствора Gla I во все 6 лунок и перемешайте путем пипетки вверх и вниз.
ПРИМЕЧАНИЕ: Используйте многоканальную пипетку, чтобы все скважины могли быть инициированы одновременно. - Поместите черную пластину обратно в устройство считывания пластин. Записывайте флуоресценцию каждые 35 с в течение 35 мин. Встряхивайте пластину (двойную орбиталь при 425 cpm в течение 3 с) перед каждым считыванием.
Результаты
Активный DNMT1 является обязательным условием для этого анализа. RFTS-отсутствующий DNMT1 экспрессировался в E.coli и очищался до однородности в соответствии с ранее опубликованными процедурами41. Чтобы убедиться, что очищенный фермент был активным, для изучения активности ме?...
Обсуждение
Для выявления и характеристики ингибиторов ДНК-метилтрансфераз необходимо измерить активность фермента. Существует несколько методов изучения активности ДНК-метилтрансферазы. Активность обычно контролируется с помощью радиоактивности; перенос меченой метильной группы SAM может быт...
Раскрытие информации
У авторов нет конфликта интересов для раскрытия.
Благодарности
Авторы благодарят Университет Бакнелла и химический факультет за поддержку этой работы.
Материалы
| Name | Company | Catalog Number | Comments |
| 96-well Half Area Black Flat Bottom Polystyrene Not Treated Microplate | Corning | 3694 | |
| 96-Well Polystyrene Conical Bottom Plates | ThermoFisher | 249570 | |
| Bovine Serum Albumin | NEB | B9000S | |
| compound 1 | ChemBridge | 5812086 | screening compound; resuspended in DMSO to 10 mM |
| compound 2 | ChemBridge | 6722175 | screening compound; resuspended in DMSO to 10 mM |
| compound 3 | ChemBridge | 5249376 | screening compound; resuspended in DMSO to 10 mM |
| Dithiothreitol | Sigma | D0632 | |
| Gla I | SibEnzyme | E494 | methyl-sensitive endonuclease |
| Glycerol | RPI | G22025 | |
| Magnesium Chloride | Sigma | M0250 | |
| Oligonucleotide (5'-FAM-CCTATGCGmCATCAGTTTTCTGATGmCGmCATAGG-3'-Iowa Black Quencher) | IDT | custom synthesized | internally quenched hairpin DNA (substrate) |
| Potassium Glutamate | Sigma | G1501 | |
| S-adenosylmethionine | Sigma | A4377 | methyl-donating co-factor (substrate) |
| Tris Base | RPI | T60040 |
Ссылки
- Jurkowska, R. Z., Jurkowski, T. P., Jeltsch, A. Structure and function of mammalian DNA methyltransferases. Chembiochem. 12 (2), 206-222 (2011).
- Hamidi, T., Singh, A. K., Chen, T. Genetic alterations of DNA methylation machinery in human diseases. Epigenomics. 7 (2), 247-265 (2015).
- Norvil, A. B., Saha, D., Dar, M. S., Gowher, H. Effect of disease-associated germline mutations on structure function relationship of DNA methyltransferases. Genes. 10 (5), 369 (2019).
- Foulks, J. M., et al. Epigenetic drug discovery: targeting DNA methyltransferases. Journal of Biomolecular Screening. 17 (1), 2-17 (2012).
- Goll, M. G., Bestor, T. H. Eukaryotic cytosine methyltransferases. Annual Review of Biochemistry. 74, 481-514 (2005).
- Bigey, P., Ramchandani, S., Theberge, J., Araujo, F. D., Szyf, M. Transcriptional regulation of the human DNA Methyltransferase (dnmt1) gene. Gene. 242 (1-2), 407-418 (2000).
- Detich, N., Ramchandani, S., Szyf, M. A conserved 3'-untranslated element mediates growth regulation of DNA methyltransferase 1 and inhibits its transforming activity. Journal of Biological Chemistry. 276 (27), 24881-24890 (2001).
- MacLeod, A. R., Rouleau, J., Szyf, M. Regulation of DNA methylation by the Ras signaling pathway. Journal of Biological Chemistry. 270 (19), 11327-11337 (1995).
- Slack, A., Cervoni, N., Pinard, M., Szyf, M. DNA methyltransferase is a downstream effector of cellular transformation triggered by simian virus 40 large T antigen. Journal of Biological Chemistry. 274 (15), 10105-10112 (1999).
- Eads, C. A., Nickel, A. E., Laird, P. W. Complete genetic suppression of polyp formation and reduction of CpG-island hypermethylation in Apc(Min/+) Dnmt1-hypomorphic mice. Cancer Research. 62, 1296-1299 (2002).
- MacLeod, A. R., Szyf, M. Expression of antisense to DNA methyltransferase mRNA induces DNA demethylation and inhibits tumorigenesis. Journal of Biological Chemistry. 270 (14), 8037-8043 (1995).
- Ramchandani, S., MacLeod, A. R., Pinard, M., von Hofe, E., Szyf, M. Inhibition of tumorigenesis by a cytosine-DNA, methyltransferase, antisense oligodeoxynucleotide. Proceedings of the National Academy Sciences of the United States of America. 94 (2), 684-689 (1997).
- Ley, T. J., et al. DNMT3A mutations in acute myeloid leukemia. New England Journal of Medicine. 363, 2424-2433 (2010).
- Walter, M. J., et al. Recurrent DNMT3A mutations in patients with myelodysplastic syndromes. Leukemia. 25 (7), 1153-1158 (2011).
- Russler-Germain, D. A., et al. The R882H DNMT3A mutation associated with AML dominantly inhibits wild-type DNMT3A by blocking its ability to form active tetramers. Cancer Cell. 25 (4), 442-454 (2014).
- Roll, J. D., Rivenbark, A. G., Jones, W. D., Coleman, W. B. DNMT3b overexpression contributes to a hypermethylator phenotype in human breast cancer cell lines. Molecular Cancer. 7, 15 (2008).
- Nosho, K., et al. DNMT3B expression might contribute to CpG island methylator phenotype in colorectal cancer. Clinical Cancer Research. 15 (11), 3663-3671 (2009).
- Erdmann, A., Halby, L., Fahy, J., Arimondo, P. B. Targeting DNA methylation with small molecules: what's next. Journal of Medicinal Chemistry. 58 (6), 2569-2583 (2015).
- Chuang, J. C., et al. S110, a 5-Aza-2'-deoxycytidine-containing dinucleotide, is an effective DNA methylation inhibitor in vivo and can reduce tumor growth. Molecular Cancer Therapeutics. 9 (5), 1443-1450 (2010).
- Issa, J. J., et al. Safety and tolerability of guadecitabine (SGI-110) in patients with myelodysplastic syndrome and acute myeloid leukaemia: a multicentre, randomised, dose-escalation phase 1 study. Lancet Oncology. 16 (9), 1099-1110 (2015).
- Datta, J., et al. A new class of quinoline-based DNA hypomethylating agents reactivates tumor suppressor genes by blocking DNA methyltransferase 1 activity and inducing its degradation. Cancer Research. 69 (10), 4277-4285 (2009).
- Zambrano, P., et al. A phase I study of hydralazine to demethylate and reactivate the expression of tumor suppressor genes. BMC Cancer. 5, 44 (2005).
- Lee, B. H., Yegnasubramanian, S., Lin, X., Nelson, W. G. Procainamide is a specific inhibitor of DNA methyltransferase 1. Journal of Biological Chemistry. 280 (49), 40749-40756 (2005).
- Asgatay, S., et al. Synthesis and evaluation of analogues of N-phthaloyl-l-tryptophan (RG108) as inhibitors of DNA methyltransferase 1. Journal of Medicinal Chemstry. 57 (2), 421-434 (2014).
- Ceccaldi, A., et al. C5-DNA methyltransferase inhibitors: from screening to effects on zebrafish embryo development. Chembiochem. 12 (9), 1337-1345 (2011).
- Fagan, R. L., Wu, M., Chédin, F., Brenner, C. An ultrasensitive high throughput screen for DNA methyltransferase 1-targeted molecular probes. PLoS One. 8 (11), 78752 (2013).
- Fagan, R. L., Cryderman, D. E., Kopelovich, L., Wallrath, L. L., Brenner, C. Laccaic acid A is a direct, DNA-competitive inhibitor of DNA methyltransferase 1. Journal of Biological Chemistry. 288 (33), 23858-23867 (2013).
- Eglen, R. M., Reisine, T. Screening for compounds that modulate epigenetic regulation of the transcriptome: an overview. Journal of Biomolecular Screening. 16 (10), 1137-1152 (2011).
- Holz-Schietinger, C., Matje, D. M., Reich, N. O. Mutations in DNA methyltransferase (DNMT3A) observed in acute myeloid leukemia patients disrupt processive methylation. Journal of Biological Chemistry. 287 (37), 30941-30951 (2012).
- Norvil, A. B., et al. Dnmt3b methylates DNA by a noncooperative mechanism, and its activity is unaffected by manipulations at the predicted dimer interface. Biochemistry. 57 (29), 4312-4324 (2018).
- Bashtrykov, P., Ragozin, S., Jeltsch, A. Mechanistic details of the DNA recognition by the Dnmt1 DNA methyltransferase. FEBS Letters. 586 (13), 1821-1823 (2012).
- Bashtrykov, P., et al. Targeted mutagenesis results in an activation of DNA methyltransferase 1 and confirms an autoinhibitory role of its RFTS domain. Chembiochem. 15 (5), 743-748 (2014).
- Berkyurek, A. C., et al. The DNA methyltransferase Dnmt1 directly interacts with the SET and RING finger-associated (SRA) domain of the multifunctional protein Uhrf1 to facilitate accession of the catalytic center to hemi-methylated DNA. Journal of Biological Chemistry. 289 (1), 379-386 (2014).
- Kanada, K., Takeshita, K., Suetake, I., Tajima, S., Nakagawa, A. Conserved threonine 1505 in the catalytic domain stabilizes mouse DNA methyltransferase 1. Journal of Biochemistry. 162 (4), 271-278 (2017).
- Bashtrykov, P., et al. Specificity of Dnmt1 for methylation of hemimethylated CpG sites resides in its catalytic domain. Chemistry & Biology. 19 (5), 572-578 (2012).
- Dolen, E. K., McGinnis, J. H., Tavory, R. N., Weiss, J. A., Switzer, R. L. Disease-associated mutations G589A and V590F relieve replication focus targeting sequence-mediated autoinhibition of DNA methyltransferase 1. Biochemistry. 58 (51), 5151-5159 (2019).
- Kilgore, J. A., et al. Identification of DNMT1 selective antagonists using a novel scintillation proximity assay. Journal of Biological Chemistry. 288 (27), 19673-19684 (2013).
- Ceccaldi, A., et al. Identification of novel inhibitors of DNA methylation by screening of a chemical library. ACS Chemical Biology. 8 (3), 543-548 (2013).
- Duchin, S., Vershinin, Z., Levy, D., Aharoni, A. A continuous kinetic assay for protein and DNA methyltransferase enzymatic activities. Epigenetics & Chromatin. 8, 56 (2015).
- Syeda, F., et al. The replication focus targeting sequence (RFTS) domain is a DNA-competitive inhibitor of Dnmt1. Journal of Biological Chemistry. 286 (17), 15344-15351 (2011).
- Switzer, R. L., Medrano, J., Reedel, D. A., Weiss, J. Substituted anthraquinones represent a potential scaffold for DNA methyltransferase 1-specific inhibitors. PLoS One. 14 (7), 0219830 (2019).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены