
Для просмотра этого контента требуется подписка на Jove Войдите в систему или начните бесплатную пробную версию.
Method Article
Разработка противовирусных агентов с помощью поверхностного плазмонного резонанса
В этой статье
Резюме
Настоящий протокол описывает новые инструменты для анализов связывания SPR для изучения связывания CV-N с ГК, гликопротеином S, родственными гликанами гибридного типа и олигосахаридами с высоким содержанием маннозы. SPR используется для определения KD для связывания димерного или мономерного CV-N с этими гликанами.
Аннотация
Поверхностный плазмонный резонанс (SPR) используется для измерения связывания гемагглютинина (HA) с димером циановирина-N (CV-N) с заменой домена и для мониторинга взаимодействия между маннозилированными пептидами и сайтом связывания CV-N с высоким сродством. Сообщалось, что всплески оболочки вируса gp120, HA и гликопротеин Эболы (GP) 1,2 связывают как высоко-, так и низкоаффинные сайты связывания на димерном CVN2. Диманнозилированный пептид HA также связывается в двух низкоаффинных сайтах связывания с инженерной молекулой CVN2, которая несет высокоаффинный сайт для соответствующего лиганда и мутирует, чтобы заменить стабилизирующую дисульфидную связь в углеводсвязывающем кармане, тем самым подтверждая поливалентное связывание. Показано связывание ГК с одним высокоаффинным сайтом связывания псевдоантител CVN2 при константе диссоциации (KD) 275 нМ, что дополнительно нейтрализует вирус иммунодефицита человека типа 1 (ВИЧ-1) посредством олигомеризации. Корреляция числа дисульфидных мостиков в CVN2 с заменой домена, которые уменьшаются с 4 до 2 путем подстановки цистинов в полярные пары остатков глутаминовой кислоты и аргинина, приводит к снижению сродства связывания с ГК. Среди самых сильных взаимодействий Ebola GP1,2 связан CVN2 с двумя высокоаффинными сайтами связывания в нижнем наномолярном диапазоне с использованием гликана оболочки без трансмембранного домена. В настоящем исследовании связывание мультиспецифического мономерного CV-N с тяжелым острым респираторным синдромом коронавируса 2 (SARS-CoV-2) спайка (S) гликопротеина измеряется при KD = 18,6 мкМ по сравнению с наномолярным KD с этими другими вирусными всплесками и через его рецептор-связывающий домен в среднем μ-молярном диапазоне.
Введение
Тетерин-ассоциированная противовирусная активность индуцируется интерферон-α, и она содержит тросы на белковой основе, что приводит к удержанию полностью сформированных вирионов на инфицированных клеточных поверхностях1. Необходимость гликозилирования тетерина в ингибировании высвобождения вируса остается неопределенной, что подразумевает важность паттернов гликозилирования на рекомбинантно выраженных гликанах для исследований in vitro 1,2, которая зависит от конформации (в случае вируса гриппа) поверхностно экспрессированного гемагглютинина гриппа HA 3,4 . Отмечено, что модификации олигосахарида, привязанного к N-сцепленному гликозилированию, достаточно для тетерин-опосредованного ограничения высвобождения ВИЧ типа 12, в то время как димеризация играет существенную роль в предотвращении высвобождения вируса, тем самым вовлекая трансмембранный домен или гликозил-фосфатидил-инозитол (GPI)-якорь для привязки почковых вирионов5 . Описаны уникальные особенности человеческого и мышиного тетерина, блокирующего множественные оболочки вирусов, ретровирусов и филовирусов. BST-2/tetherin представляет собой интерферон-индуцируемый противовирусный белок врожденного иммунитета 1,6, действующий с широкой противовирусной активностью и антагонизированный оболочкой гликопротеинов5 с целью либо транслоцировать тетерин, либо нарушить структуру тетерина6. Например, поверхностно-экспрессированный гликопротеин оболочки ГК и нейраминидаза на вирусе гриппа А хорошо известны антагонизмом тетерина специфическим для штамма способом7, облегчающим распознавание сайтов связывания рецепторов хозяина8. Антитела, нацеленные на гликаны, изучаются в стехиометрии их взаимодействия с быстро настраиваемыми гликкановыми щитами на ГК, что приводит к сродству связывания с подтипами гриппа A H3N2 и H1N14.
Для выяснения механизмов связывания между противовирусными агентами и шипами оболочки вируса, т.е. углеводными лигандами, и комплементарными иммунологическими и спектроскопическими методами химически синтезируют моно-, ди- и триманнозные фрагменты. Маннозилированные пептиды создаются путем азидо-гликозилирования гликозил {бета}-перацетатов до трансформации 1,2-транс-гликозилазидов9, имитируя обычно встречающиеся N-ацетилглюкозамин и олигосахариды с высоким содержанием маннозы на поверхности опасных для жизни вирусов. Биоизостеры триазола используются для имитации связей, которые образуют маннозилированный остаток пептида ГК10 и облегчают сайт-специфические взаимодействия с противовирусными производными CV-N вокруг второго N-связанного пятна гликозилирования на головном домене ГК (вершина ГК с 4 N-связанными гликанами N54, N97, N181, N301)8,11,12 . Взаимодействия между глутаминовой кислотой (Glu) и аргинином (Arg) и полученным в результате диполем спирали проявили хорошую стабильность как модельных пептидов, так и белков, но визуализируются с использованием SPR. Если сравнивать с распознаванием одного химически синтезированного участка гликозилирования наГК-10 путем прямого ингибирования связывания рецепторов на фрагментах гликана, более высокое сродство четырехсайтовой мутированной структуры Fc к его рецептору, как показано, вызывает эффекторные функции in vivo, выявляя несвязанный состав N-связанных гликанов, прикрепленных к мутанту Fc, который механически детерминирован13.
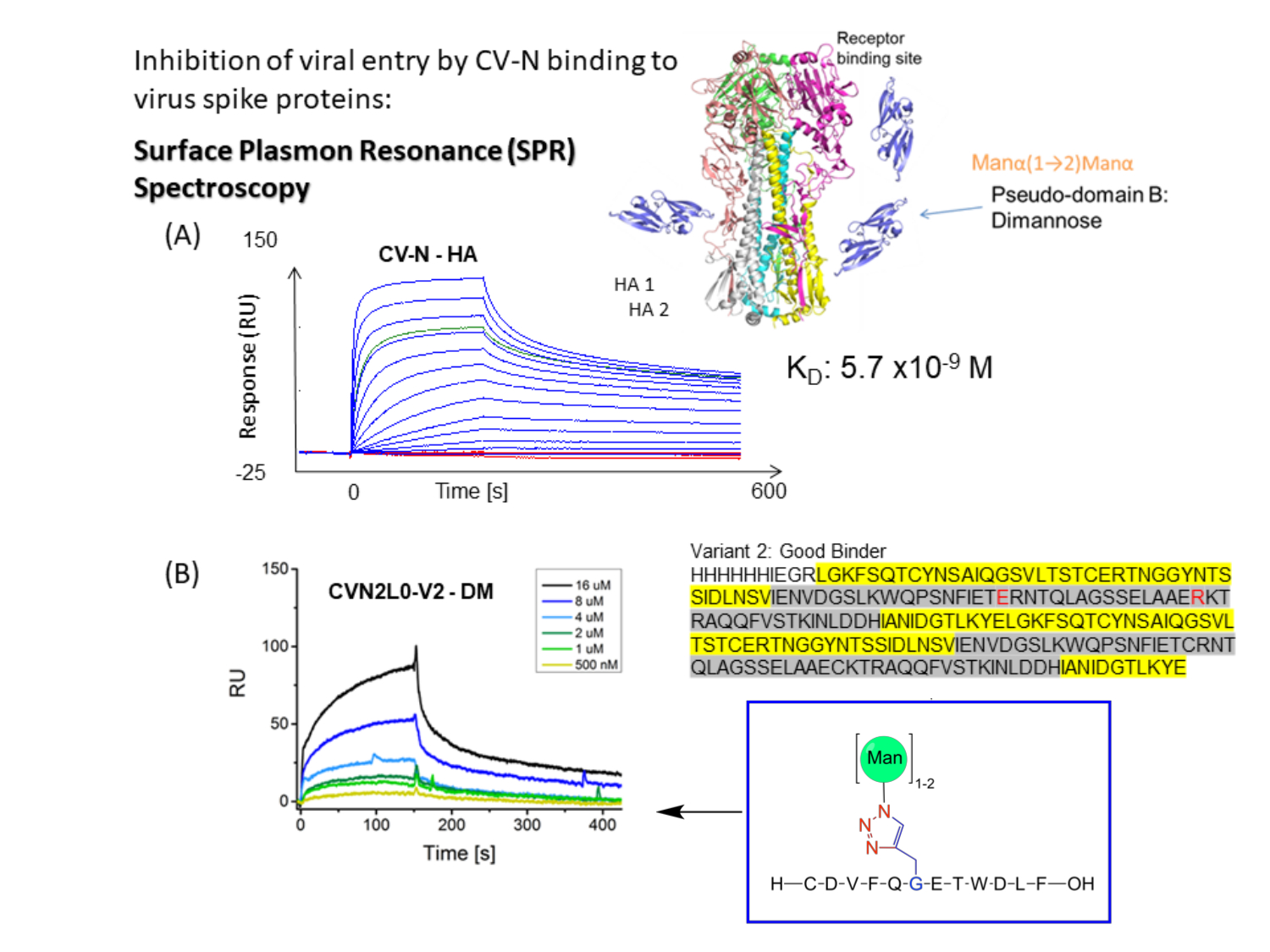
CV-N проявляет противовирусную активность в отношении ВИЧ14,15, гриппа16 и вируса Эбола, которая опосредована наномолярным связыванием с высокоманнозными олигосахаридными модификациями на белках оболочки 12,17,18,19. Связывание ГК гриппа с одним высокоаффинным углеводсвязывающим сайтом (H) в CV-N или двумя Hs в ковалентно связанном димерном CVN2 определяется как имеющее равновесные константы диссоциации (KD) = 5,7 нМ (рисунок 1A) и KD = 2,7 нМ соответственно. И CV-N, и CVN2 содержат еще один или два низкоаффинных сайта связывания углеводов (L) 12,17,20,21. Эбола GP1,2 связывается с 2H CVN2 со сродством в нижнем наномолярном диапазоне (KD = 26 нМ). Связывание CV-N WT с Эболой GP1,2 и HA проявляет сродство от KD = 34 нМ к KD = 5,7 нМ (A/New York/55/04)12. Лектины, такие как CV-N, которые специально нацелены на гликаны с высоким содержанием маннозы на вирусных оболочках, дополнительно ингибируют репликацию вируса гепатита С, SARS-CoV, герпесвируса, вируса Марбурга и вируса кори22.
Небольшая молекула CV-N тщательно изучается более 20 лет, поскольку она функционализируется для связывания широкого спектра вирусов для ингибирования проникновения вируса 16,18. Структурный анализ и анализы сродства связывания указывают на сшивание двух Ls в димере CVN2 с заменой домена путем двухвалентного связывания в микромолярном диапазоне для повышения авидности к гликопротеинам вирусной оболочки10,19. Селективное связывание Manα1-2Manα на руках Man(8) D1D3 и Man(9) содержит два сайта связывания различного сродства, расположенных на противоположных белковых протомерах20, тем самым достигая сродства к наномолярному связыванию (фиг.1B). Таким образом, CVN2 считается псевдоантителом относительно его применения для связывания эпитопов на ВИЧ gp120, аналогично вируснейтрализующим антителам17. При этом автор заинтересован в исследовании потенциального связывания CVN2 с всплеском SARS-CoV-2 через его рецептор-связывающий домен (RBD). Кривые связывания иммобилизованного человеческого ангиотензинпревращающего фермента (АПФ)-2 с RBD SARS-CoV-2 приводят к KD = 4,7 нМ для этого биологически значимого связывающего взаимодействия23.
Напротив, выбранные классы иммуноглобулинов распознают специфические и последовательные структурные белковые паттерны, которые придают субстрат для созревания аффинности в мембранно-анкерных областяхГК 24. CV-N проявляет высокую мощную активность почти у всех вирусов гриппа А и В16 и является широко нейтрализующим противовирусным средством. Наши знания являются неполными о расположении целевых эпитопов на стволе HA1 и HA2, которые, возможно, включают эпитопические структуры для нацеливания на гликаны с помощью высоконейтрализующих антител и по сравнению с связыванием лектина25.

Рисунок 1: Схематическое представление анализа связывания SPR для CV-N с всплесками оболочки вируса. (A) SPR Анализ для связывания CV-N с лигандом: полноразмерный белок HA (90 кДа). Набор кинетических данных (5120, 2560, 1280, 640, 320, 160, 80, 40, 20, 10, 5, 2,5, 0 нМ), показывающий двойное связывание в реальном времени с гриппом HA A/New-York/55/04 (H3N2). (B) CVN2L0 вариант V2 связывание с иммобилизованным лигандом DM в диапазоне концентраций от 500 нМ до 16 мкМ. Последовательность: L остатки выделены желтым цветом. Остатки H выделены серым цветом. E58 и R73 являются заменой цистеинов в белке дикого типа и делают V2 стабильной белковой складкой с тремя вместо четырех дисульфидных связей Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
В то время как гликановый щит на мембранно-дистальной верхней части ГК индуцирует высокоаффинное связывание с CV-N12, связывание CVN2 с ГК, прилегающее к дисульфидному мосту верхней части ГК, также наблюдалось на его участках с низким сродством 10,12. Различные полярные взаимодействия и сайты взаимодействия идентифицируются в углеводном связывании CV-N. Эти взаимодействия проверяются путем генерации нокаутирующих вариантов в месте связывания для корреляции сродства связывания с in silico предсказанным гликозилированием12. Таким образом, проект направлен на сравнение ранее протестированных химически маннозилированных пептидов ГК в сродстве связывания и специфичности с короткими пептидными последовательностями из связанных с SARS всплесков 2019-nCoV и SARS-CoV-2, встречающихся в природе, модифицированных небольшим количеством различных N-связанных сайтов гликозилирования и O-связанного гликозилирования. Используя криоэлектронную микроскопию и связывающие анализы, Пинто и его коллеги сообщают о моноклональном антителе S309, которое потенциально распознает эпитоп на спайковом белке SARS-CoV-2, содержащем сохраненный гликан в подроде sarbecovirus, не конкурируя с рецепторным присоединением26. Протокол этого исследования описывает, как проектирование, экспрессия и характеристика вариантов CV-N важны для изучения того, как CV-N и CVN2 связываются с гликозилированными белками и синтетическими маннозилированными пептидами с использованием технологии SPR10,12.
Тандем-связанный димер CVN2L027 и варианты сайта связывания (V2-V5) рекомбинантно экспрессированы, а варианты с заменами дисульфидных связей (C58E и C73R) (рисунок 2A). Кроме того, мутант с одноточечной мутацией E41A получают, потому что это положение рассматривалось как межмолекулярный перекрестно-контактирующий остаток. Этот мутант является еще одной интересной молекулой для измерений связывания SPR между лектином и высокоманнозными олигосахаридами, расшифровывающими связывающие домены, и позволяет сравнивать с димерной формой. Кристаллическая структура CVN2 с заменой домена показывает гибкий компоновщик, который простирается от 49 до 54 остатков. Два домена могут продолжать двигаться вокруг шарнира в виде твердых тел, развивая либо мономер через внутримолекулярные доменные взаимодействия (домен A -остатки 1-39;90-101- с доменом B -остатки 40-89), либо димер путем межмолекулярной замены домена [домен A (первого мономера) с доменом B (второго) и домен B (первого мономера) с доменом A (второй копии)]. Между доменами A и B двух протомеров нет тесных взаимодействий, за исключением Glu4128. Ген CV-N может быть разработан с использованием повторяющегося метода ПЦР с 40-мерным синтезированным олигосом29 и затем субклонирован в NdeI и BamHI участки pET11a для преобразования (электропорации) в электрокомпетентные клетки, как описано Keeffe, J.R.27. Белок, который используется для достижения соответствующей кристаллической структуры (PDB ID 3S3Y), включает N-концевую метку очистки 6-гистидина, за которой следует участок расщепления протеазы фактора Xa. Сайт-направленный мутагенез используется для создания точечных мутаций, переключения кодонов и вставки или удаления одного или нескольких оснований или кодонов для обмена аминокислотами. Эти превращения дают бесценное представление о функции и структуре белка. Рекомбинантно экспрессированные и очищенные CV-N, CVN2 и CVN3 были биофизически хорошо изучены 20,21,27, дешевы в производстве и, следовательно, используются для характеристики анализов связывания с гликанами, иммобилизованными на чипах датчиков SPR. Обычный иммуноферментный анализ (ИФА) обеспечивает меньшую воспроизводимость в отношении метода иммобилизации лигандов гликана и преобразует связывание в реальном времени различных вариантов сайта связывания, что показано для SPR, в анализы конечных точек.
Связывающе-аффинный вариант CVN2L0-V2 (неповрежденная складка гомодимерного CV-N с замещением дисульфидного моста10) экспрессируется His-меткой в Escherichia coli (E. coli), очищенной над колонкой Ni-NTA с применением аффинной хроматографии и проверенной на связывание с HA (H3N2), мономаннозилированным HA-пептидом и диманнозилированным HA-пептидом с использованием SPR. Химически маннозилированные пептиды, или белок HA и S, все они являются лигандами и амином, связанными с гидрофильной поверхностью чипа через реактивные сложные эфиры или биотин-стрептавидиновую белковую инженерию. Та же процедура последовательных запусков применяется к этим лигандам, вводя различные разведения CV-N и варианты CV-N (и CVN2) для получения кинетической информации для анализа молекулярного взаимодействия, как описано ниже30. RBD-иммобилизованный сенсорный чип SPR используется для исследований связывания пептидов CV-N в S, а сродство сравнивается с связыванием SARS-CoV-2 с ЧЕЛОВЕЧЕСКИМ ACE2.
протокол
Для настоящего исследования CVN-малый убиквитин-подобный модификатор (SUMO) слитый белок был использован в иммуноферментных анализах вместо CV-N и подходит для клеточных анализов. Рекомбинантный полноразмерный белок вируса гриппа А HA H3 получают коммерчески (см. Таблицу материалов) или экспрессируют в клеточных линиях HEK293 млекопитающих и инфицированных бакуловирусом клетках насекомых в соответствии со стандартными протоколами12. Спайковый белок Wuhan-1 экспрессируется в клетках HEK293 млекопитающих. Синтез мономаннозилированного пептида (ММ) и диманнозилированного пептида (СД) позволяет обнаруживать гомогенные лиганды к CVN2 и мономаннозилированной малоймолекуле 10.
1. Создание конструкций CV-N
- Для каждого из вариантов CVN2 и белка CVN2L0 (PDB ID 3S3Y) получите генную конструкцию с N-концевой ведущей последовательностью pelB и His-tag в векторе pET27b(+) из коммерческих источников (см. Таблицу материалов, дополнительный файл 1).
- Получите CVN2L0 и его варианты (V2, V3, V4 и V5; Рисунок 2A,C) на фоне гена шаблона CVN2L0, состоящего из двух различных последовательностей ДНК для каждого повтора CV-N.
- Растворить лиофилизированную плазмидную ДНК в стерильной деионизированной дистиллированной воде (ddH2O) до конечной концентрации 100 нг/мкл.
2. Получение пластин LB-агара с плазмидными ДНК-трансформированными клетками
- Приготовьте культуральную среду LB-Lennox, растворив 10 г/л пептона, 5 г/л дрожжевого экстракта и 5 г/л NaCl в ddH2O (см. Таблицу материалов), и отрегулируйте рН до 7,4. Выполняют преобразование в компетентную Кишечную палочку BL21 (DE3) для каждого варианта (V2-V5) химическим методом в соответствии с ранее опубликованным отчетом10.
- Разделите раствор (900 мкл и 100 мкл), переложите 100 мкл на пластины LB-агара (50 мкг/мл канамицина) и осторожно используйте стерильный клеточный распределитель. Инкубировать агаровые пластины в течение ночи при температуре 37 °C.
3. Клонирование
- Субклонируйте ген CV-N в NdeI и BamHI участки pET11a (см. Таблицу материалов) для преобразования (электропорации) в электрокомпетентные клетки по ссылке27.
4. Мутагенез, направленный на сайт
- Генерировать CVN2L029 и мутантный CVN-E41A на фоне гена шаблона CVN2L0, содержащего две различающиеся последовательности ДНК для каждого CV-Nповтора 27.
- Производят мутации с помощью набора мутагенеза, направленного на сайт (см. Таблицу материалов) и специфических мутагенных праймеров 5'-gagaaccgtcaacgtttgcgataacagagttcagg-3' и 5'-cctgaactctgttatcgcaaacgttgacggttctc-3' для запуска ПЦР31.
- Начните серию пробных реакций с использованием многократных концентраций шаблона двухцепочечной ДНК (дцДНК) в диапазоне от 5 до 50 нг (например, 5, 10, 20 и 50 нг шаблона дцДНК). Держите концентрацию грунтовки постоянной.
ПРИМЕЧАНИЕ: Обычно используется протокол ПЦР-смеси и термоциклирования, как описано в руководстве по эксплуатации для набора32 для мутагенеза, направленного на сайт.
- Начните серию пробных реакций с использованием многократных концентраций шаблона двухцепочечной ДНК (дцДНК) в диапазоне от 5 до 50 нг (например, 5, 10, 20 и 50 нг шаблона дцДНК). Держите концентрацию грунтовки постоянной.
- Добавьте фермент рестрикции Dpn I (1 мкл, 10 Ед/мкл, см. Таблицу материалов) под наложением минерального масла. Тщательно и осторожно перемешайте реакции, открутите в настольной микроцентрифуге в течение 1 мин и сразу же инкубируйте при 37 °C в течение 1 ч для переваривания родительской суперспиральной дцДНК.
5. Трансформация бактериальных клеток
- Аккуратно разморозьте сверхкомпетентные ячейки XL1-Blue (см. Таблицу материалов) на льду. Для преобразования каждой контрольной и пробной реакции аликвотируют сверхкомпетентные клетки (50 мкл) в предварительно охлажденную полипропиленовую трубку круглого дна (14 мл).
- Перенос 1 мкл обработанной Dpn I одноцепочечной ДНК (ssDNA) из каждой контрольной реакции и реакции образца (мутированной ssDNA) в отдельные аликвоты сверхкомпетентных клеток, которые синтезируют комплементарную цепь. Тщательно закрутите реакции трансформации, чтобы перемешать и инкубировать реакции на льду в течение 30 мин.
ПРИМЕЧАНИЕ: Перед переносом ДНК, обработанной Dpn I, в реакцию трансформации рекомендуется осторожно удалить оставшееся минеральное масло из кончика пипетки. В качестве дополнительного элемента управления эффективность трансформации суперкомпетентных клеток XL1-Blue необходимо проверить путем смешивания 0,1 нг/мкл управляющей плазмиды pUC18 (1 мкл) с аликвотой 50 мкл суперкомпетентных клеток.
- Перенос 1 мкл обработанной Dpn I одноцепочечной ДНК (ssDNA) из каждой контрольной реакции и реакции образца (мутированной ssDNA) в отдельные аликвоты сверхкомпетентных клеток, которые синтезируют комплементарную цепь. Тщательно закрутите реакции трансформации, чтобы перемешать и инкубировать реакции на льду в течение 30 мин.
- Применяют тепловой импульс к реакциям превращения при 42 °С в течение 45 с, а затем помещают реакции на лед на 2 мин.
ПРИМЕЧАНИЕ: Применяемый тепловой импульс уже оптимизирован для указанных условий в полипропиленовых трубках круглого дна (14 мл). - Добавить 0,5 мл бульона NZY+ (содержащего на литр: 10 г амина NZ (гидролизат казеина), 5 г дрожжевого экстракта, 5 г NaCl, 12,5 мл 1 M MgCl2, 12,5 мл 1 M MgSO4, 10 мл 2 М глюкозы, рН 7,5 и предварительно нагретый до 42 °C) и инкубировать реакции трансформации при 37 °C с встряхиванием при 225-250 об/мин в течение 1 ч. На пластинах LB-ампициллина агар на пластинах LB-ампициллина агара наносится правильный объем каждой реакции трансформации (5 мкл от контрольной плазмиды; 250 мкл от трансформации образца).
ПРИМЕЧАНИЕ: Для контроля трансформации и мутагенеза распространяются клетки на пластины агарового агарового LB-ампициллина, имеющие 5-бром-4-хлор-3-индолил-β-D-галактопиранозид (X-галлон, 80 мкг/мл) и изопропил-1-тио-β-D-галактопиранозид (IPTG, 20 мМ) (см. Таблицу материалов). Инокулируют 50 мл клеточных культур одной колонией трансформированного Е. клетки кишечной палочки для очистки мутировавшей плазмидной ДНК для анализов. Мутагенез подтверждается секвенированием ДНК на внешнем объекте.
6. Экспрессия и очистка белка
- Для крупномасштабной культуры инокулируют небольшое количество LB (содержащего ампициллин) одной колонией из преобразованной пластины.
- Используя культуру на ночь, инокулируют экспрессионную культуру добавками, такими как 10 мМ MgCl2, 10 мМ MgSO4 и 20 мМ глюкозы, разбавляя семенную культуру до 1/100.
- Выращивайте клетки с энергичным встряхиванием при 37 °C. Вырастите клетки до ABS 600 нм между 0,4-0,6 (средняя фаза log) перед охлаждением клеток до 20 °C. Индуцировать с 1 мМ IPTG и расти в течение ночи.
- Затем собирают клетки путем центрифугирования при 4000 х г в течение 15 мин при 4 °C и выбрасывают супернатант с пипеткой.
- Повторное суспендирование клеточной гранулы в фосфат-буферном солевом (PBS) буфере и повторной центрифуге при 4000 х г в течение 15 мин при 4 °C. Затем выбросьте супернатант с помощью пипетки. Повторно суспендируют оставшуюся гранулу в 10 мл лизисного буфера и инкубируют суспензию в течение 1 ч при 37 °С.
ПРИМЕЧАНИЕ: Состав лизисного буфера: (50 мМ2PO4, 300 мМ NaCl, 2% тритон-X100, 500 нг/мл лизоцима, 1 мМ фенилметилсульфонилфторида (PMSF), 1 мМ дитиотрейтола, 1 мМMgCl2, рН 8, см. Таблицу материалов).- Подвергайте смесь двум циклам замораживания-оттаивания (-80 °C). Разделяйте растворимые и нерастворимые фракции центрифугированием при 4000 х г в течение 15 мин при 4 °C и анализируйте их с помощью электрофореза полиакриламидного геля (PAGE), в частности, додецилсульфата натрия (SDS)-PAGE33 (рисунок 2B, C).
ПРИМЕЧАНИЕ: Клетки могут быть лизированы несколькими способами, такими как замораживание-оттаивание, обработка ультразвуком, гомогенизация, ферментативный лизис или комбинация этих методов. Очищение от органов включения рекомендуется для сбора высоких белковых выходов26.
- Подвергайте смесь двум циклам замораживания-оттаивания (-80 °C). Разделяйте растворимые и нерастворимые фракции центрифугированием при 4000 х г в течение 15 мин при 4 °C и анализируйте их с помощью электрофореза полиакриламидного геля (PAGE), в частности, додецилсульфата натрия (SDS)-PAGE33 (рисунок 2B, C).
- Очищайте белки с помощью колоночной хроматографии Ni-NTA (см. Таблицу материалов). Загрузите растворимую фракцию в регенерированную колонку ni-NTA (1 мл/мин). Промывайте систему буфером TBS (50 мМ Tris, 150 мМ хлорида натрия, рН 7,5) перед запуском градиента (0-100% 500 мМ имидазола в TBS в течение 60 мин) и сбора фракций (1 мл/мин). Диализируйте очищенные белки для биохимической характеристики против 100 мМ PBS (рисунок 2C,D).
- Альтернативно, используйте гегель сродства His-Select Ni2+ (см. Таблицу материалов) в пробирках по 14 мл для связывания и повторной приостановки его меченого рекомбинантно экспрессированного CV-N в буферных растворах с имидазолом 20 мМ и имидазолом 250 мМ соответственно. Инкубировать порционно не менее 30 мин.
ПРИМЕЧАНИЕ: Применяют эти полуочищенные белки к одноразовой предварительно упакованной колонке для буферного обмена и очистки биологических образцов, например, углеводов и белков, которые могут загружать элюат 1-1,5 мл из иммобилизованной аффинной хроматографии металлов. - Переложите белковые растворы в центрифугирующие трубки с отсечным фильтром 10 кДа (см. Таблицу материалов) и концентрируйте их центрифугированием в течение 10 мин при 4 500 х г и 4 °C. Для измерений SPR замените растворы анализируемого вещества на 10 мМ HEPES, 150 мМ хлорида натрия, 3 мМ этилендиаминтетрауксусной кислоты (ЭДТА) и 0,05% Tween20, рН 7,4 (HBS-EP(+), см. Таблицу материалов).
- Добавьте этот рабочий буфер SPR к коэффициенту разбавления 1:10 и центрифугу четыре раза к исходному объему в течение 10 мин при 4 500 х г и 4 °C.
- Определить концентрацию белка при 280 нм с помощью спектрофотометра NanoDrop UV-Vis (см. Таблицу материалов) на основе расчетного коэффициента вымирания (20 440 М-1 см-1) для основного белка CVN2L0, показывающего размер 23 474 Da34. Используйте PBS (100 мМ, pH 7,0) или буфер SPR в качестве заготовки и измерьте концентрацию белка на трех этапах разведения (1:1, 1:10 и 1:100).

Рисунок 2: Последовательности и экспрессия CV-N. (A) CVN2 без связующего звена между каждым cv-N-повтором (по 101 аминокислоте в каждом) и четырьмя дисульфидными мостиками выражается в векторе pET11a в E. кишечная палочка. (B) Выражения двух независимых колоний для CV-N (мономер) и CVN2 (димер). (C) Варианты дисульфидных связей очищаются и анализируются на SDS-PAGE. В качестве эталона используется низкомолекулярный маркер (6 мкл). WT = CVN2L0, несущий четыре дисульфидных моста с маркировкой (A). V2 представляет собой вариант с заменой дисульфидной связи полярными остатками в положениях 58 и 73. V3-V5 представляют собой варианты с двумя оставшимися S-S связями и либо полярными (C58E-C73R), либо неполярными (C58W-C73M), заменяющими аминокислоты, либо комбинацией этих замен этих остаточных пар. (D) ВЭЖХ-хроматограммы очищенного CVN2L0 элюируют со скоростью потока 1 мл/мин с линейным градиентом от 5%-65% буфера B в буфере A в течение 30 мин. Буфер A составляет: 0,1% (v/v) трифторуксусной кислоты в ddH2O, буфер B составляет: 0,08% (v/v) трифторуксусной кислоты в ацетонитриле. Белок анализируют на высокоэффективном силикагеле 300-5-С4 (150 х 4,6 мм) колонке при 214 нм и 280 нм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
7. SPR-спектроскопия
- Используйте двухканальную систему SPR (см. Таблицу материалов) с работающим буфером HBS-EP(+) и 10 мМ глицина HCl pH 1,5-1,6 в качестве буфера регенерации. Включите прибор, дегазатор, автопробоотборник, а также перекачайте и промывайте всю систему с ddH20 в течение 1 ч. Поместите готовый к использованию рабочий буфер в отдельную бутылку.
- Капните эмерсионное масло на детектор и установите стеклянный сенсорный чип (см. Таблицу материалов), покрытый тонкой золотой пленкой и на верхней стороне функционализированный карбоксиметилдекстрановым гидрогелем, непосредственно на детектор под трехпортовой проточной ячейкой. Исправьте настройку, потянув вниз обработку.
ПРИМЕЧАНИЕ: C19RBDHC30M 200 нм стрептавидин дериватизированный карбоксиметилдекстран гидрогель со средней плотностью биотинилированного тяжелого острого респираторного синдрома коронавируса-2 RBD белка, представляет собой готовый к применению сенсорный чип с предварительно иммобилизованным лигандом.
8. SPR-связывающий анализ для связывания CV-N с HA, S-белком и RBD
- Обездвижьте белковые лиганды для сенсорных чипов, следуя приведенным ниже шагам.
- Откройте таблицу выполнения, щелкнув Форма в строке меню и Запустить редактор таблиц в интегрированном программном обеспечении SPRAutoLink (см. Таблица материалов). Выберите и нажмите на BASIC_Immobilization из списка доступных таблиц запуска и следуйте шагам экспериментальной процедуры на экране компьютера. Соответствующий используемый редактор образцов отображается в правом верхнем углу.
- Нажмите на Редактор наборов образцов в разделе Форма, чтобы заполнить список реагентов для двух стоек, размещенных в автосэмплере для дальнейшего анализа. Нажмите на Autosampler Direct Control в качестве «Инструмента» в строке меню, чтобы переместить стойки вперед или домой. Выберите 4 °C в качестве рабочей температуры.
ПРИМЕЧАНИЕ: Программное обеспечение SPR позволяет использовать "SPR Instrument Direct Control" и "Pump Direct Control" путем выбора соответствующих инструментов, а также автоматической обработки и нажатия на Форму; Кроме того, для выполнения анализа данных можно выбрать редактор таблиц, диаграмму данных или постобработку . Файлы сохраняются непосредственно в каталоге по умолчанию и экспортируются как scrubber.files из окна постобработки.
- Запустите насос для вливания ddH20, щелкнув Инструменты и прямое управление насосом и записывая данные, нажимая на SPR Instrument Direct Control, и каждый раз запускайте в новых окнах. Поместите реагенты связи (шаг 8.3) во флаконы емкостью 300 мкл, поместите их в стойки автопробоотборника и запустите таблицу запуска, нажав кнопку Выполнить.
ПРИМЕЧАНИЕ: Поверхности стружки либо кондиционируются глициновым буфером pH 9,0 мМ, либо могут быть промыты 1 М хлорида натрия, 0,1 М боратного буфера натрия pH 9,0 для кондиционирования карбоксильной дериватизированной поверхности чипа для активации EDC/NHS смеси30. - Для этого простого белково-белкового взаимодействия используйте чип CMD500D (см. Таблицу материалов) для генерации микропреломляющих показателей единиц (μRIU) = 2500 - 3000 проточных клеток с иммобилизованным ГК и μRIU = 400 проточных клеток с спайковым белком. При скорости потока вливания 15 мкл/мин вводят водную и равную смесь 0,4 М N-этил-N'-(диметиламинопропил) карбодиимида гидрохлорида (EDC*HCl) и 0,1 М N-гидроксисукцинимида (NHS), применяя следующие последовательные этапы.
- Заправка насоса со скоростью 25 000 мкл/мин, выполнение базовой регулировки в течение 30 с, введение 90 мкл раствора для активации образца (EDC/NHS) в течение 6 мин времени контакта, а затем удержание еще 5 мин.
- Повторите этот цикл после исходного запуска в течение 1 мин при 10 мкл/мин только на левой проточной клетке (синем) для введения и иммобилизации химически синтезированных пептидов10, ГК и спайкового белка при 20 мкг/мл и допускайте последующую базовую корректировку с ddH2O в течение 1,5 мин перед закалкой активированной поверхности чипа 1 М этаноламином HCl pH 8,5.
- Переключите трубки с жидкого пробоотборника на дегазатор из ddH20 во флакон с HBS-EP(+) (дополнительный рисунок 1).
- Проанализируйте сенсорограммы SPR.
- Для выполнения кинетических исследований используют различные концентрации анализируемых веществ (10-5-10-8 М), с шагом регенерации после каждой инъекции и пустыми измерениями после различных аналитов. Измените скорость потока до 10 мкл/мин и начните инъекции в течение 4 мин контактного времени, затем 5 мин базовой генерации и двух ступеней регенерации по 2 мин каждая с интервалом 30 с.
- Вводят буферный раствор для пустых измерений, сенсорграммы которого вычитаются из пробных прогонов для нормализации различных концентраций белка.
- Нажмите « Форма», прокрутите вниз и перейдите на «Постобработка», щелкнув этот режим работы. Нажмите кнопку Добавить , чтобы выбрать кривые привязки, созданные с течением времени в форме печати данных для каждой ячейки потока, и экспортируйте наложение в виде файла скруббера (.ovr). Нажмите «Файл», чтобы открыть параметры сохранения файлов. Получение кривых отклика путем выравнивания левой и правой кривых и вычитания сигналов второго опорного канала из сигналов лигандного канала.
ПРИМЕЧАНИЕ: Данные обрабатываются в "Постобработке" путем вычислительного определения сенсорграмм. Он помещается в наложение сенсорграмм из левой и правой проточных ячеек или представляется в виде сенсорграмм от разницы обоих каналов. - Очистите всю жидкость 50-100 мМ глицинового буфера рН 9,5, воду и 20% этанол до и после измерений связывания, чтобы удалить следы соли или любого белкового загрязнения, или более строгие, с 0,5% SDS и глицином.
ПРИМЕЧАНИЕ: Для предотвращения повреждения прибора рекомендуется проверить механическую стабильность стеклянного чипа перед повторной вставкой картриджа чипа в прибор, если чипы хранились в буфере или при 100% влажности.
Результаты
Молекула CVN2L0 с димерной заменой домена тестируется на связывание с верхней областью ГК в трех отдельных экспериментах SPR, и сродство связывания представлено в значениях KD. Предполагается, что домен B содержит Н-связывающие участки, на которые воздействует замена дисульфидной свя?...
Обсуждение
Сродство привязки CV-N коррелирует с количеством функциональных сайтов привязки [2H на доменах B и 2L на домене (доменах) A при проектировании как димер с заменой домена]. Вариант с измененным сродством связывания (CVN2L0-V2, гомодимерная стабильная складка CV-N, содержащая нокаут дисульфидного мо...
Раскрытие информации
Автору нечего раскрывать.
Благодарности
Автор благодарит доктора Кристиана Дернтля из Департамента биотехнологии и микробиологии в Венском университете и Департамента медицины III, Отделения нефрологии и диализа в Медицинском университете Вены, особенно доктора Маркуса Варманна за техническую и научную поддержку. Экспрессия белка в клетках млекопитающих была поддержана кафедрой биотехнологии в Университете природных ресурсов и наук о жизни (BOKU) в Вене. Автор хочет выразить свое глубокое признание доктору Нико Данкбару из XanTec bioanalytics в Дюссельдорфе, Германия, за полезные научные дискуссии по выполнению анализов связывания SPR.
Материалы
| Name | Company | Catalog Number | Comments |
| Äkta primeplus | Cytiva | ||
| Amicon tubes | Merck | C7715 | |
| Ampillicin | Sigma-Aldrich | A5354 | |
| Beckmann Coulter Cooler Allegra X-30R centrifuge | Beckman Coulter | B06320 | |
| Cell spreader | Sigma-Aldrich | HS86655 | silver stainless steel, bar L 33 mm |
| Custom DNA Oligos | Sigma-Aldrich | OLIGO | |
| Custom Gensynthesis | GenScript | #1390661 | cloning vector: pET27b(+) |
| Cytiva HBS-EP+ Buffer 10, 4x50mL | Thermo Scientific | 50-105-5354 | |
| Dionex UlitMate 3000 | Thermo Scientific | IQLAAAGABHFAPBMBFB | |
| Dpn I restriction enzyme (10 U/μL) | Fisher Scientific | ER1701 | |
| DTT | Merck | DTT-RO | |
| EDC | Merck | 39391 | |
| EDTA | Merck | E9884 | |
| Eppendorf Safe-Lock Tubes | Eppendorf | 30120086 | |
| Eppendorf Safe-Lock Tubes | Eppendorf | 30120094 | |
| Eppendorf Minispin and MiniSpin Plus personal microcentrifuge | Sigma-Aldrich | Z606235 | |
| Ethanol | Merck | 51976 | |
| Ethanolamine HCl | Merck | E6133 | |
| Falcon 50mL Conical Centrifuge Tubes | Fisher Scientific | 14-432-22 | |
| Falcon 14 mL Round Bottom Polystyrene Test Tube, with Snap Cap, Sterile, 25/Pack | Corning | 352057 | |
| Glucose | Merck | G8270 | |
| Glycine HCl | Merck | 55097 | |
| HA H3 protein | Abcam | ab69751 | |
| HEPES | Merck | H3375 | |
| His-select Ni2+ | Merck | H0537 | |
| Imidazole | Merck | I2399 | |
| IPTG | Merck | I6758 | |
| Kanamycin A | Sigma-Aldrich | K1377 | |
| Kromasil 300-5-C4 | Nouryon | ||
| LB agar | Merck | 52062 | |
| LB agar | Merck | 19344 | |
| LB Lennox | Merck | L3022 | |
| Lysozyme | Merck | 10837059001 | |
| Magnesium chloride | Merck | M8266 | |
| Magnesium sulfate | Merck | M7506 | |
| NaH2P04 | Merck | S0751 | |
| NanoDrop UV-Vis2000c spectrophotometer | Thermo Scientific | ND2000CLAPTOP | |
| NaOH | Merck | S5881 | |
| NHS | Merck | 130672 | |
| NZ amine (casein hydrolysate) | Merck | C0626 | |
| PBS | Merck | 806552 | |
| PD MidiTrap G-10 | Sigma-Aldrich | GE28-9180-11 | |
| Peptone | Merck | 70171 | |
| pET11a | Merck Millipore (Novagen) | 69436 | |
| PMSF | Merck | PMSF-RO | |
| QIAprep Spin Miniprep Kit (1000) | Qiagen | 27106X4 | |
| Reichert Software Package Autolink1-1-9 | Reichert | ||
| Reichert SPR SR7500DC Dual Channel System | Reichert | ||
| Scrubber2-2012-09-04 for data analysis | Reichert | ||
| SDS | Merck | 11667289001 | |
| Site-directed mutagenesis kit incl pUC18 control plasmid | Stratagene | #200518 | |
| Sodim chloride | Merck | S9888 | |
| Sodium acetate.Trihydrate | Merck | 236500 | |
| SPR sensor chip C19RBDHC30M | XanTec bioanalytics | SCR C19RBDHC30M | |
| SPR sensor chip CMD500D | XanTec bioanalytics | SCR CMD500D | |
| Sterilin Standard 90mm Petri Dishes | Thermo Scientific | 101R20 | |
| TBS | Merck | T5912 | 10x, solution |
| Triton-X100 | Merck | T8787 | |
| Tryptone | Merck | 93657 | |
| Tween20 | Merck | P1379 | |
| Vortex-Genie 2 Mixer | Merck | Z258423 | |
| X-gal | Merck | XGAL-RO | |
| XL1-Blue Supercompetent Cells | Stratagene | #200236 | |
| Yeast extract | Merck | Y1625 |
Ссылки
- Perez-Caballero, D., et al. Tetherin inhibits HIV-1 release by directly tethering virions to cells. Cell. 139 (3), 499-511 (2009).
- Waheed, A. A., Gitzen, A., Swiderski, M., Freed, E. O. High-mannose but not complex-type glycosylation of tetherin is required for restriction of HIV-1 release. Viruses. 10 (1), 26 (2018).
- Wilson, I. A., et al. Structure of the haemagglutinin membrane glycoprotein of influenza virus at 3 A resolution. Nature. 289 (5796), 366-373 (1981).
- Otterstrom, J. J., et al. Relating influenza virus membrane fusion kinetics to stoichiometry of neutralizing antibodies at the single-particle level. Proceedings of the National Academy of Sciences of the United States of America. 111 (48), 5143-5148 (2014).
- Kaletsky, R. L., Francica, J. R., Agrawal-Gamse, C., Bates, P. Tetherin-mediated restriction of filovirus budding is antagonized by the Ebola glycoprotein. Proceedings of the National Academy of Sciences of the United States of America. 106 (8), 2886-2891 (2009).
- Tokarev, A., Skasko, M., Fitzpatrick, K., Guatelli, J. Antiviral activity of the interferon-induced cellular protein BST-2/tetherin. AIDS Research and Human Retroviruses. 25 (12), 1197-1210 (2009).
- Gnirss, K., et al. Tetherin sensitivity of influenza A viruses is strain specific: Role of hemagglutinin and neuraminidase. Journal of Virology. 89 (18), 9178-9188 (2015).
- Fleury, D., et al. A complex of influenza hemagglutinin with a neutralizing antibody that binds outside the virus receptor binding site. Nature Structural Biology. 6 (6), 530-534 (1999).
- Salunke, S. B., et al. Iron(III) chloride as an efficient catalyst for stereoselective synthesis of glycosyl azides and a cocatalyst with Cu(0) for the subsequent click chemistry. Chemical Communication. (Camb). 47 (37), 10440-10442 (2011).
- Schilling, P. E., et al. Mannosylated hemagglutinin peptides bind cyanovirin-N independent of disulfide-bonds in complementary binding sites. RSC Advances. 10 (19), 11079-11087 (2020).
- Fleury, D., Wharton, S. A., Skehel, J. J., Knossow, M., Bizebard, T. Antigen distortion allows influenza virus to escape neutralization. Nature Structural Biology. 5 (2), 119-123 (1998).
- Maier, I., Schiestl, R. H., Kontaxis, G. Cyanovirin-N binds viral envelope proteins at the low-affinity carbohydrate binding site without direct virus neutralization ability. Molecules. 26 (12), 3621 (2021).
- Ahmed, A. J., Keremane, S. R., Vielmetter, J., Bjorkman, P. J. Structural characterization of GASDALIE Fc bound to the activating Fc receptor FcγRIIIa. Journal of Structural Biology. 194 (1), 78-89 (2016).
- Boyd, R. Discovery of cyanovirin-N, a novel human immunodeficiency virus-inactivating protein that binds viral surface envelope glycoprotein gp120: potential applications to microbicide development. Antimicrobial Agents and Chemotherapy. 41 (7), 1521-1530 (1997).
- Bolmstedt, A. J., O'Keefe, B. R., Shenoy, S. R., McMahon, J. B., Boyd, M. R. Cyanovirin-N defines a new class of antiviral agent targeting N-linked, high-mannose glycans in an oligosaccharide-specific manner. Molecular Pharmacology. 59 (5), 949-954 (2001).
- O'Keefe, B. R., et al. Potent anti-influenza activity of cyanovirin-N and interactions with viral hemagglutinin. Antimicrobial Agents and Chemotherapy. 47 (8), 2518-2525 (2003).
- Shenoy, S. R., et al. Multisite and multivalent binding between cyanovirin-N and branched oligomannosides: calorimetric and NMR characterization. Chemical Biology. 9 (10), 1109-1118 (2002).
- Bewley, C. A., Kiyonaka, S., Hamachi, I. Site-specific discrimination by cyanovirin-N for alpha-linked trisaccharides comprising the three arms of Man(8) and Man(9). Journal of Molecular Biology. 322 (4), 881-889 (2002).
- Barrientos, L. G., Matei, E., Lasala, F., Delgado, R., Gronenborn, A. M. Dissecting carbohydrate-Cyanovirin-N binding by structure-guided mutagenesis: functional implications for viral entry inhibition. Protein Engineering Design & Selection. 19 (12), 525-535 (2006).
- Bewley, C. A., Otero-Quintero, S. The potent anti-HIV protein cyanovirin-N contains two novel carbohydrate binding sites that selectively bind to Man(8) D1D3 and Man(9) with nanomolar affinity: implications for binding to the HIV envelope protein gp120. Journal of the American Chemical Society. 123 (17), 3892-3902 (2001).
- Bewley, C. A. Solution structure of a cyanovirin-N:Man alpha 1-2Man alpha complex: structural basis for high-affinity carbohydrate-mediated binding to gp120. Structure. 9 (10), 931-940 (2001).
- Jensen, S. M. R., et al. Differential inhibitory effects of Cyanovirin-N, Griffithsin, and Scytovirin on entry mediated by envelopes of gammaretroviruses and deltaretroviruses. Journal of Virology. 88 (4), 2327-2332 (2014).
- Lan, J., et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature. 581 (7807), 215-220 (2020).
- Lingwood, D., et al. Structural and genetic basis for development of broadly neutralizing influenza antibodies. Nature. 489 (7417), 566-570 (2012).
- Ekiert, D. C., et al. Antibody recognition of a highly conserved influenza virus epitope. Science. 324 (5924), 246-251 (2009).
- Pinto, D., et al. Cross-neutralization of SARS-CoV-2 by a human monoclonal SARS-CoV antibody. Nature. 583 (7815), 290-295 (2020).
- Keeffe, J. R., et al. Designed oligomers of cyanovirin-N show enhanced HIV neutralization. Proceedings of the National Academy of Sciences of the United States of America. 108 (34), 14079-14084 (2011).
- Yang, F., et al. Crystal structure of cyanovirin-N, a potent HIV-inactivating protein, shows unexpected domain swapping. Journal of Molecular Biology. 288 (3), 403-412 (1999).
- Stemmer, W. P., Crameri, A., Ha, K. D., Brennan, T. M., Heyneker, H. L. Single-step assembly of a gene and entire plasmid from large numbers of oligodeoxyribonucleotides. Gene. 164 (1), 49-53 (1995).
- Fischer, M. J. E., Mol, N., Fischer, M. Amine coupling through EDC/NHS: A practical approach. Surface Plasmon Resonance. Methods in Molecular Biology (Methods and Protocols). 627, (2010).
- Novoradovsky, A., et al. Computational principles of primer design for site directed mutagenesis. Technical Proceedings of 2005 NSTI Nanotechnology Conference and Trade Show. , 532-535 (2005).
- . QuikChange Site-Directed MutagenesisKit, User Manual Available from: https://users.drew.edu/jliu3/Docs/Stratagene%20Quikchange%20mutagenesis.pdf#:~:text=The%20QuikChange%20sitedirected%20mutagenesis%20kit%20is%20used%20to (2005)
- Laemmli, U. K. Cleavage of structural proteins during the assembly of the head bacteriophage T4. Nature. 227 (5259), 680-685 (1970).
- . Expasy.org Available from: https://web.expasy.org/cgi-bin/protparam/protparam (2022)
- Karlsson, R. Real-time competitive kinetic analysis of interactions between low-molecular-weight ligands in solution and surface-immobilized receptors. Analytical Biochemistry. 221 (1), 142-151 (1994).
- Schuck, P., Zhao, H. The role of mass transport limitation and surface heterogeneity in the biophysical characterization of macromolecular binding processes by SPR biosensing. Methods Molecular Biology. 627, 15-54 (2020).
- Barnes, O., et al. SARS-CoV-2 neutralizing antibody structures inform therapeutic strategies. Nature. 588 (7839), 682-687 (2020).
- . Using reichert Surface Plasmon Resonance (SPR) for Antiviral Development, Application Note 28 Available from: https://www.reichertspr.com/clientuploads/directory/application_notes/Application_Note_28__Using_Reichert_Surface_Plasmon_Resonance_for_Antiviral_Testing.pdf (2022)
- Sundberg, E. J., Andersen, P. S., Gorshkova, I. I., Schuck, P., Schuck, P. Surface plasmon resonance biosensing in the study of ternary systems of interacting proteins. Protein Interactions: Biophysical Approaches for the Study of Complex Reversible Systems. 5, 97-141 (2007).
- . Method Development Notes Available from: https://www.reichertspr.com/applications/method-development-notes/ (2022)
- Angulo, J., Enríquez-Navas, P. M., Nieto, P. M. Ligand-receptor binding affinities from saturation transfer difference (STD)-NMR spectroscopy: the binding Isotherm of STD initial growth rates. Chemistry. 16 (26), 7803-7812 (2010).
- Goldflam, M., Tarragó, T., Gairí, M., Giralt, E. NMR studies of protein-ligand interactions. Methods in Molecular Biology. 831, 233-259 (2012).
- Kumar, S., Maurya, V. K., Prasad, A. K., Bhatt, M. L. B., Saxena, S. K. Structural, glycosylation and antigenic variation between 2019 novel coronavirus (2019-nCoV) and SARS coronavirus (SARS-CoV). Virusdisease. 31 (1), 13-21 (2020).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеThis article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены