
Method Article
간질의 제브라피시 모델의 행동 및 생리학적 분석
요약
여기서, 우리는 DEPDC5 유전자의 일시적인 억제로 인한 간질의 제브라피시 모델의 개발 및 특성화를 위한 프로토콜을 제시한다.
초록
간질은 전 세계적으로 약 5천만 명의 사람들에게 영향을 미치는 가장 흔한 신경 장애 중 하나입니다. 유전 연구에 있는 최근 어드밴스는 간질의 각종 양식에 연루된 유전자의 큰 스펙트럼을, 이 무질서의 이질적인 본질을 강조하. 적절한 동물 모델은 간질에 연루된 유전 적 돌연변이에 의해 유발되는 병리학 적 메커니즘을 조사하고 전문적이고 표적화된 치료법을 개발하는 데 필수적입니다. 최근 몇 년 동안, 제브라피쉬는 새로운 항간질 치료제를 확인하기 위해 펜티레네타졸(PTZ)과 같은 알려진 간질 약물에 대한 유전적 조작과 노출을 모두 사용하여 간질을 모델링하는 귀중한 척추동물 유기체로 부상하고 있다. mTOR 레귤레이터 DEPDC5의 해로운 돌연변이는 제브라피시 정형소의 다양한 형태의 초점 간질 및 노크 다운과 관련이 있어 자발적인 발작과 같은 에피소드와 관련된 과잉 행동을 유발할 뿐만 아니라 향상된 전기 촬영 활동 및 특징적인 턴 휠 수영과 관련이 있습니다. 여기서, 우리는 DEPDC5 기능 손실 모델을 생성하는 데 관여하는 방법을 설명하고 28 및 48 hpost 수정(hpf)에서 모터 활성을 평가하기 위한 프로토콜뿐만 아니라 제브라피시 광학 텍텀에서 현장 활성을 기록하는 방법을 설명했다. 시간이 지남에 따라 신경 활동에 간질 성 약물 PTZ의 효과의 그림도 제공됩니다.
서문
개발 초기 단계에서 의작은 크기, oviparous 개발 및 투명성으로 인해, 제브라피쉬는 심혈관, 암 또는 신경 장애1,2등다양한 인간 질환을 모델링하기위한 귀중한 척추 동물 유기체로 부상하고있다. Zebrafish는 장기 건축 및 유전 코드의 높은 보존을 포함하여 척추 동물의 장점을 간단한 모델 유기체의 작은 크기와 유전 조작의 용이성과 결합하여 기본 연구와 번역 응용 프로그램을 모두 촉진합니다. 특히, 세포 과정의 동작 및 형광 마커의 고처리량 자동 선별에 대한 편의성은 제브라피시를 간질 연구를 위한 특히 매력적인 모델로 만들었습니다. 이는 최근 간질3,4,5의 화학적으로 유도된 및/또는 유전적 모델을 특징으로 하는 간행물의 수가 증가하여 입증되었으며, 최근에는 이들 모델에서 화학스크린에서 얻은 유망한 치료법에 대한보고6,7,8이입증되었다.
DEPDC5는 GATOR1 컴플렉스의 구성원이며 mTOR 시그널링9의 네거티브 레귤레이터입니다. DEPDC5 유전자의 돌연변이는 2013년에 자가우성 지배적인 초점간질(10,11)으로 고통받는 대장에서 처음 발견되었으며, 이후 초점 간질 증상 및 초점 피질 이형성증(12)과 관련된 다수의 임상 조건에서 보고되었다. 보고된 돌연변이의 대다수는유전자(12)의기능 상실을 야기할 것으로 예상되며, 이는 말도 안되는 매개 mRNA 붕괴12,13에의해 표적으로 하는 다수의 DEPDC5 돌연변이 성적증명서에 대해 공식적으로 입증되었다. 이에 따라, 항센스 모르폴리노 올리고뉴클레오티드(AmOs)를 이용한 제브라피시의 유전자 정형소그를 노크하면 과잉 행동, 회전 휠 형 수영, 자발적인 발작 및 향상된 뉴런 활동14,15,16,17,18을포함하여 이 유기체에서 간질 모델에 공통적인 여러 가지 특징이 생성된다. 흥미롭게도, mTOR 신호의 억제제인 라파마이신을 사용하여 치료하여 이모델(18)의행동 특징을 반전시켜, DEPDC5 기능 상실이 mTOR 통로9,19의오용으로 인해 간질을 유발할 수 있다는 가설을 뒷받침한다.
모르폴리노 변형을 운반하는 안티센스 올리고뉴클레오티드를 이용한 생체 내 유전자 발현의 일시적인 노크다운은 si/shRNA 기반 기술과 동등한 특정 유전자의 역할을 연구하는 데 매우 중요한 도구였습니다. 최근 AMO 기반 전략은 2016년2016년듀첸 근위축 치료에 대한 FDA 승인을 받은 최초의 AMO 요법으로 임상 응용 분야도 발견했습니다. 제브라피시에서 급성 AMO 계 유전자 노크다운의 표현형이 항상 구성적인 녹아웃모델(21)과상관관계가 있는 것은 아니라고 보고되었지만, 이는 적어도 어떤 경우에는 구성적인 유전자변형(22)에의해 야기된 보상 메커니즘이 있기 때문일 수 있다. 그러나, AMO 유도 표현형의 특이성 문제는 이 기술을 이용한 연구에서 부지런히 다루어야 할 명백한관심사이다(23). AMO 기반 녹다운 표현형의 특이성을 보장하기 위해서는 몇 가지 주요 컨트롤이 필요합니다. 이들은 유전 물질의 과잉의 소개 때문에 전반적인 독성을 피하는 유전자 노크 다운에 효과적인 AMO의 가장 낮은 복용량의 선택을 허용하는 복용량 반응 곡선을 포함합니다. 게놈에 있는 어떤 특정 영역도 표적으로 하지 않는 불일치 AMO의 사용은 또한 적당한 복용량을 확립하고 특정 표현형을 확인하기 위해 요구됩니다. 스플라이스 차단 AMO와 같은 동일한 유전자의 상이한 영역을 표적으로 하는 제2 AMO는 표현형이 표적 유전자의 노크 다운에 기인한다는 것을 확인하는 데 필요하다. 유전자의 cDNA를 가진 노크 다운 표현형의 구조, 인간 정형 소 또는 AMO에 의해 표적으로 할 수없는 제브라피시 유전자의 코돈 변형 버전, 표현형 특이성에 찬성 강한 인수를 제공합니다. 기능 상실 돌연변이(예: 조기 정지 코돈의 도입)를 포함하는 동일한 cDNA를 가진 구조의 부족은 이 방향으로 추가 증거입니다.
여기서, 우리는 제브라피시 DEPDC5 기능 손실 모델및 행동 페노티핑을 위한 프로토콜을 생성하는 방법을 제시한다 28 및 48 h 포스트 수정 (hpf). 28 hpf에서 DEPDC5 기능 손실은 초리온 내의 배아의 향상된 코일링 및 경련 운동에 의해 입증된 바와 같이 전반적인 과잉 행동을 일으킵니다. 이 단계에서는 배아당 전체 활성을 정량화하기 위해 자동화된 모션 감지 시스템을 사용할 수 있습니다. 48hpf에서, 제브라피쉬는 터치에 대한 응답으로 수영 고정 관념 탈출을 전시한다. DEPDC5의하향 조절된 발현을 가진 제브라피어에서, 수영 궤도는 대조군보다 훨씬 더 고통스럽고, 이 유기체3,4에서보고된 다른 간질 모델과 유사한 패턴과 같은 "코르크 나사" 또는 "턴휠"을 나타내는 물고기. 전기생리학적 기록은 4-6일 간 제브라피시 애벌레의 광학 텍텀에서 수득되었고, DEPDC5 녹다운 동물에서 뉴런 활성의 기준증증가를 보여주었다. 이 모델의 장점은 개발 중 약물 치료의 효능을 모니터링하고 평가하는 데 유용 할 수있는 다른 시점에서 여러 가지 현상제 기능을 제시한다는 것입니다.
프로토콜
실험 절차는 국가 및 기관 윤리위원회의 승인을 받았습니다.
1. 제브라피시 배아에서 DEPDC5 유전자의 일시적인 노크 다운
- 도구 준비:
- 실리콘 엘라스토머 코팅 주입 페트리 접시 준비: 키트의 베이스 및 경화제(재료 표참조)를 10:1 비율로 섞습니다. 혼합물로 35mm 페트리 접시를 중간에 채웁니다. 실리콘을 사용하기 전에 굳어질 때까지 기다립니다 (이것은 며칠이 걸릴 수 있습니다).
- 안티센스 모르폴리노 올리고뉴클레오티드의 1.2 mmol/L 스톡 솔루션을 준비합니다(AMO; 재료표참조). 멸균물 250μL을 300nmole lyophilized AMO에 추가하여 1.2 mmol/L 스톡 솔루션을 얻습니다. 완전한 용해를 위해 바이알을 65°C에서 5분 동안 가열합니다. 소용돌이가 짧게. 플라스틱 필름으로 튜브 캡을 밀봉합니다(재료 표참조).
- 제어 구조 실험을 위해, PCS2 백본 또는 유사한 제브라피시 호환 식 플라스미드에 복제된 DEPDC5의 인간 cDNA를 포함하는 발현 플라스미드(table of Materials)및 유사한 제브라피시 호환 식 플라스미드를 준비한다. 부정적인 대조군으로서, 조기 정지 코돈(p.Arg487*)을 유발하는 돌연변이가 cDNA에 도입되었다.
- 배아 물 준비: 0.06 g/L 수족관 소금 (재료의 표참조) 역 삼투압 물 + 0.5 mg/L 메틸렌 블루.
- 주사의 날, 풀러를 사용하여 미세 주입 보로 실리 케이트 유리 바늘을 준비 (재료의 표참조). 바늘 풀러에 적절한 온도 설정을 설정합니다. 길이 10cm, 1/0.5 OD/ID mm 보로실리케이트 유리 모세관을 사용하여 길이가 약 1cm인 얇은 팁으로 2~5cm의 모세혈관을 생성합니다.
- 바늘의 끝이 매우 미세한 경우, 용액의 배출을 방지, 현미경에서 집게를 사용하여 테이퍼 팁의 맨 끝을 깰.
- 주입 직전에 AmOs의 작업 솔루션을 준비하십시오. 항상 결과의 재현성을 보장하기 위해 신선한 솔루션을 준비하십시오. AMO 스톡 바이알을 65°C에서 5분 동안 가열합니다. 빠른 녹색 염료 (0.02 % 최종 농도, 재료 의 표참조)를 포함하는 5 μL 사출 샘플을 준비하고 물에서 작업 농도에서 희석 AMO.
- 용량 반응 곡선을 사용하여 각 유전자에 대한 AMO의 작업 농도를 경험적으로 결정합니다. 작업 농도는 AMO가 총 형태학적 결함과 같은 일반적인 독성을 일으키지 않고 유전자를 쓰러뜨리는 데 효과적인 농도를 나타낸다. 전형적으로, AMO 작업 농도는 0.2 mmol/L에서 1 mmol/L(0.4 mmol/L의 범위에서 이 연구18에대한 효과적인 농도로 결정되었다). 효과적인 AMO와 동일한 농도로 제어 불일치 모르몰리노를 주입합니다.
- 튜브와 원심분리기를 잠시 소용돌이쳐 방울을 튜브 바닥으로 가져옵니다.
- 구조 실험을 위해, 작업 농도에서 희석된 AMO를 사용하여 5 μL 사출 시료를 준비하고, cDNA 발현 플라스미드는 최종 농도로 질적으로 결정된다. DEPDC5 및 음극식 플라스미드의 발현을 위해, 100 ng/μL은 표현형 구조에 효과적이었다.
- 배아 준비:
- 미세 주입 전날, 제브라피시 결합 탱크를 설정합니다. 주사의 아침, 산란을 가능하게 하는 칸막이를 제거합니다. 미세한 체로 배아 물로 채워진 100mm 페트리 접시에 계란을 채집합니다. 계란은 한 세포 단계에있는 동안, 수집에서 20-30 분 이내에 주입.
- 플라스틱 파스퇴르 파이펫으로 60-80개의 달걀을 선택하고 실리콘 코팅 페트리 접시에 담아 주입합니다. 실리콘 표면은 주사 중에 계란이 미끄러지는 것을 방지합니다. 배아 물의 대부분을 제거, 중간 계란을 커버하기에 충분한 떠나.
- 미세 주입:
- 유리 바늘을 사출 용액으로 채웁니다. 주사 용액을 포함하는 튜브 중 하나에 바늘을 수직으로 배치하여 바늘의 바닥 끝이 용액을 만지고 있는지 확인합니다. 유색 주입 용액이 모세 혈관에 의해 상승하고 바늘의 끝에서 볼 때까지 몇 분 기다립니다.
- 마이크로 인젝터의 주입 핸들에 채워진 바늘을 장착하십시오 (재료 표참조).
- 공기 압축기를 켜고 압력 설정을 조정하여 ~2 nL의 사출 부피를 생성합니다.
- 주입된 용액의 부피를 계산하려면 마이크로톤 슬라이드에 미네랄 오일 한 방울을 놓습니다. 설정된 압력 및 시간 매개 변수를 사용하여 염료 함유 솔루션을 주입합니다. 주입된 유체 구의 직경을 측정하고 주입된 볼루스의 측정 직경을 d=로 측정한 수식 볼륨=4/3*π*(d/2)3을사용하여 총 부피를 계산합니다.
- 4배 배율을 가진 해부 쌍안경 현미경을 사용하여, 초리온과 노른자를 통과하여 단일 세포 단계에서 계란을 주입하고, 세포 내의 용액을 직접 투영한다.
- 배아 물로 100mm 페트리 접시에 주입된 배아를 수집하고, 접시에 라벨을 부착하고, 28°C에서 배양합니다.
- 배아 개발 속도가 열민감성이기 때문에 인큐베이터 온도가 시간이 지남에 따라 안정되어 있는지 확인하십시오. 예를 들어, 더 높은 온도에서 성장이 가속화되고 개발 단계는 표현형24를적절히 평가하는 데 중요합니다.
- 주입 후 계란6-8 h의 품질을 확인하고 플라스틱 파스퇴르 파이펫을 사용하여 죽은 및 수정되지 않은 배아를 제거하십시오.
- 다음 날 아침, 플라스틱 파스퇴르 파이펫으로 각 접시에 죽은 배아를 계산하고 제거합니다.
2. 동작 분석
- 28hpf의 글로벌 활동 분석:
- 미세 주입 후 하루의 오후에 테스트를 실시 (28 hpf), 테스트가 수행되는 하루 중 시간이 배아 개발이 매우 빠른 로 유효한 통계 분석을 수행하기 위해 실험을 통해 일관된 것을 보장.
- 35mm 접시(test dish)를 배아 물로 채우고 인큐베이터(28°C)에서 테스트를 시작하기 전에 최소 15분 동안 따뜻하게 할 수 있도록 합니다.
- 플라스틱 메쉬 그리드(1.2x1.2 mm)를 크기로 잘라 테스트 접시 의 바닥에 놓습니다.
- 또 다른 실험자가 배아의 시험 순서를 무작위로 하고 시험할 조건의 이름을 가리게 한다.
- 사망률이 조건 사이에서 변경되지 않고 표현형의 특이성을 보장하기 위해 비 주입 된 배아와 비교하여 변하지 않도록하십시오. 모든 조건에서 죽은 배아의 퍼센트는 10-13 %18을초과하지 않아야합니다.
- 플라스틱 파스퇴르 파이펫을 사용하여 플라스틱 메쉬에 여전히 10-12 개의 배아를 놓습니다. 배아를 물에 잠그지만 떠다니지 않도록 충분한 배아 물로 검사 접시를 채웁니다. 필요한 경우 플라스틱 팁을 사용하여 배아를 주의하여 이동하여 그리드에 배치하십시오.
- 해부 현미경에 부착된 비디오 카메라(재료표참조)를 사용하여 정의된 시간 동안 자발적인 코일 활동을 기록합니다(10-20분 길이의 비디오는 일반적으로 정량화를 위한 활동 버스트의 대표적인 샘플을 얻기에 충분)
- 배아를 각 접시에 넣고 배아를 인큐베이터에 다시 넣습니다. 각 조건에 필요한 만큼의 배아를 실험하십시오(90%의 전력 분석에 의해 결정된).
- 총 자발적인 움직임을 분석하려면 ZebraLab 시스템(재료 표 참조)을사용합니다. 활동 정량화 모듈을 사용하여, 녹화 된 비디오를 업로드하고 적절하게 각 배아 주위에 추적 경기장을 설계. 동결 및 버스트 임계값을 각각 10 및 50으로 설정합니다.
- 정의된 각 경기장 내에서 총 활동을 정량화하는 자동화된 비디오 분석을 실행한 다음 데이터 집합을 스프레드시트로 복구하고 데이터 분석 소프트웨어를 사용하여 분석을 수행합니다.
- 48hpf에서 터치 유화 탈출 응답 (TEER) :
- 주사 후 2일 아침에 시험을 실시한다(수정 후 48시간).
- 테스트 하기 전에 적어도 2 시간, 미세 한 집게를 사용 하 여 배아를 비초. 비반향과 행동 테스트의 하루 중 시간이 실험에 비해 일관되도록 보장합니다.
- 배아 물로 130mm 접시(test dish)를 채우고 시험 시작 전에 최소 15분 동안 인큐베이터(28°C)에서 워밍업할 수 있도록 합니다.
- 죽은 유충을 계산하고 제거합니다. 각 조건에 대한 숫자를 기록합니다.
- 다른 실험자가 순서를 무작위로 지정하고 테스트할 조건의 이름을 명문화하게 합니다.
- 테스트 접시 위에 카메라를 장착(재료 표참조)을 탑재하여 테스트 접시 전체가 시야 내에 있는지 확인합니다. 뷰 필드 내에 눈자를 배치하면 거리에 대한 내부 보정이 제공됩니다.
- 플라스틱 파스퇴르 파이펫으로 배아를 테스트 접시의 중앙에 놓고 30 fps의 획득 률을 사용하여 기록을 시작합니다.
- 미세한 플라스틱 팁으로 배아의 꼬리를 가볍게 터치합니다.
- 애벌레가 움직임을 종료했을 때 녹음을 중지합니다.
- 시험 접시에서 배아를 제거하고 배아 물로 채워진 새로운 접시에 놓습니다. 각 조건에 필요한 만큼의 배아로 테스트를 반복하십시오(90%의 전력 분석에 의해 결정된).
- 배아를 원래 요리로 되돌리고 배양기안에 다시 넣습니다.
- 수영 동작의 매개 변수를 분석하려면 녹화된 비디오를 ImageJ 분석 소프트웨어에 로드합니다. ImageJ의 수동 추적 플러그인을 다운로드하고 설치합니다(자료 표참조). 도구를 선택하여 플러그인을 | 플러그인 | 메뉴에서 수동 추적.
- 대화 상자 창에서 이미지의 보정된 배율을 소개합니다. 카메라 필드에 눈자를 포함하면 cm를 픽셀로 변환할 수 있습니다.
- 트랙 추가를 선택하고 첫 번째 프레임에서 제브라피시 애벌레의 이미지를 클릭하여 궤적 추적을 시작합니다. 트레이스에 추가된 각 점을 사용하여 프레임이 자동으로 진행됩니다.
- 수영 에피소드가 끝날 때까지 움직임을 추적계속하십시오.
- 추적 창에서 끝 트랙을 선택하고 X-Y 좌표를 검색하고 총 거리, 속도 및 회전 각도를 계산합니다.
3. 전기 생리분석
- 시약 및 공구 준비:
- 배아물에서 아가로즈 1%를 준비한다(섹션 1.1.4 참조). 액체 아가로즈를 미세 원심분리기 튜브에 알리쿼트와 42°C의 가열 블록에 보관하여 아가로즈가 경화되는 것을 방지합니다.
- 레코딩 용액 준비(mmol/L): NaCl 134, KCl 2.9, CaCl2 2.1, MgCl2 1.2, 포도당 10, HEPES 10, pH 7.8.
- 1.5-2 μm (5-6mm2 ·kg·s·s-3) 팁 개구부로 유리 마이크로파이펫을 당기기 A-2 저항) 광택되지 않습니다.
- 전기 생리학을위한 얼룩말 물고기 애벌레의 준비 :
- 생선을 유리 바닥 페트리 접시에 넣고(재료 표참조) 물고기가 가능한 한 뚜껑 에 가깝게 지나갈 수 있도록 과도한 세포 외 미디어를 제거하십시오.
- 플라스틱 파스퇴르 파이펫을 사용하여 애벌레 주변에 따뜻한 액체 아가로즈를 넣습니다. 물고기를 덮기 위해 충분한 아가로즈를 사용하십시오. 아가로즈가 굳어지는 동안, 미세한 집게를 사용하여 생선을 직선 자세로 방향을 조정하고, 복부 쪽을 아래로, 접시의 중앙에 두어 주세요.
- 신경 근육 전염을 차단하기 위해 10 μM 판큐로늄 브로마이드 (재료표 참조)를포함하는 레코딩 용액 2mL을 추가합니다. 녹화 중 작은 움직임으로 인해 유물을 제거하기 위해 마비기의 추가가 필요합니다.
- 전기 생리학적 기록
- 마이크로파이프를 기록 용액으로 채웁니다.
- 전압 클램프 구성의 패치 클램프 증폭기(재료 표참조)를 사용하여 배스의 전극 저항을 측정하여 올바른 값을 확인합니다.
- 20배 의 목표를 사용하여, 관의 중앙 필드에 애벌레의 머리를 배치하고 광학 텍텀 내에서 뇌의 기록 위치에 도달하기 위해 마이크로 파이프를 낮춥니다.
- 패치 클램프 증폭기를 현재 클램프로 전환하고 유지 전류를 0mA로 수정합니다.
- 1kHz의 로우 패스 필터, 1kHz의 획득 률 및 10의 디지털 게인을 사용하여 기준 활동 수준을 결정하기 위해 60분 동안 자발적인 활동을 기록합니다.
- 기준선 레코딩 1시간 후, 200 μL 펜티네트라졸(PTZ, 재료표참조) 용액 300mmol/L을 욕조에 추가하여 20mmol/L PTZ의 최종 농도를 제공합니다.
- PTZ에서 120분 동안 신경 활동을 기록합니다.
- 탈극화 이벤트 결정
- 필드 레코딩 이벤트는 매우 느린 역학을 가지고 있습니다 (관심의 주파수는 0.005-0.2 s-1의범위에 있습니다). 따라서 별칭을 피하기 위해 낮은 패스(버터워스 5차 LPF 100s-1)로신호를 필터링합니다. 획득 프레임 속도(이 경우 1ks-1)에서250s-1(RAW SIGNAL)까지 기록된 전압 데이터를 하위 샘플링합니다.
- 각 탈극화 이벤트에 대한 타임스탬프를 식별하려면 기록된 신호의 하이패스 필터링 버전인 검출 신호를 사용합니다(버터워스 1차 HPF 0.01s-1).
- 저주파 성분을 제거함으로써 간단한 임계값 을 사용하여 탈극화 이벤트를 감지할 수 있습니다. 노이즈 제거 및 이벤트 감지에 고정 된 임계값을 사용합니다 (0.3 mV가이 연구에 사용되었습니다).
- 4s보다 작은 시간 간격으로 발생하는 일련의 임계값 횡단에 의해 탈극화 이벤트를 특성화합니다. 임계값 횡단의 연속 시퀀스에서 결정된 대로 탈극화 이벤트의 시작과 끝을 계산합니다. 40ms 미만의 이벤트는 소음으로 삭제할 수 있습니다.
- 필터링되지 않은(RAW SIGNAL)에서 이벤트의 진폭을 계산하여 이벤트의 피크에 대한 로우 패스 필터링의 영향으로 인한 오류를 제거합니다. 여과된 신호에서 결정된 타임스탬프를 사용하여 원시 신호에서 탈극화 웨이블렛을 선택합니다. 진폭을 원시 신호에서 선택한 웨이블릿의 최대 값과 최소 값의 차이로 측정합니다.
참고: 단계 3.4 - 탈극화 이벤트 결정-그림 1을 가져오는 스크립트 파일은 이 문서에 첨부된 보충 파일로 제공됩니다.
결과
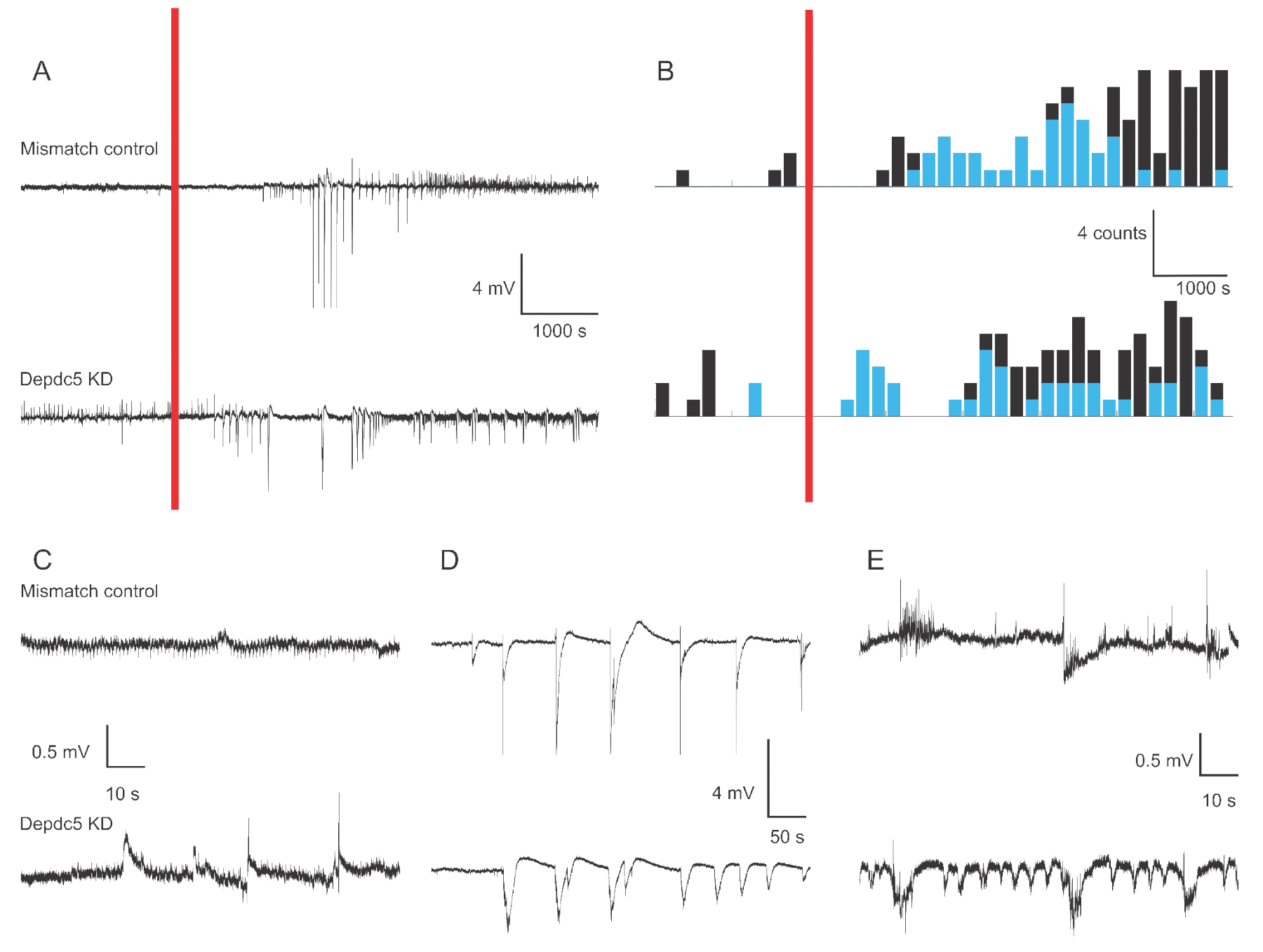
도 1은 두 가지 유전 조건의 경우 4-6 dpf 제브라피쉬 유충 의 대표적인 전압 흔적을 보여줍니다: 불일치 제어 및 DEPDC5 노크 다운. 레코딩의 기준 기간동안 DEPDC5 노크다운은 자발적인 이벤트의 발생이 더 높으며, 불일치 컨트롤은 변동이 거의 나타나지 않습니다. 이러한 활동 패턴은 DEPDC5의기능 상실로 인한 뉴런 활동의 현저한 증가를 대표하며, 이전에18을보고한 바와 같이. PTZ 응용 프로그램 후, 불일치 제어와 DEPDC5 노크 다운 모두 탈극화 이벤트의 증가를 보여줍니다. PTZ 적용 후 첫 번째 피리어드(10~60분)에서는 대부분의 이벤트가 높은 진폭(>1mV)인 미스매치 컨트롤과 DEPDC5 노크다운 모두에서 분당 0.8개의 이벤트가 관찰됩니다. 후자의 응답 기간(PTZ 적용 후 60~120분)에서는 탈극화 이벤트 속도가 분당 약 1이벤트로 증가하고 대부분의 이벤트는 진폭(≤1mV)이 적다.

그림 1: 제브라피시 애벌레 뇌의 필드 레코딩의 예 추적. (A)미스매치 컨트롤 애벌레 및 DEPDC5 녹다운에 대한 180분 기록개요. 먼저 자발적인 기준선 활동이 기록된 다음 PTZ를 욕조(red bar)에 적용하였다. (B)불일치 제어 및 DEPDC5 노크 다운에 대한 탈극화 이벤트의 페리 자극 시간 히스토그램. 이벤트는 높은 진폭 (>1 mV - 파란색) 및 낮은 진폭 (≤1 mV - 검은 색)으로 분류되었다. (C-E) 레코딩의 상이한 기간의 예추적:(C)자발적활성,(D)PTZ 적용 후 1주기 동안 높은 진폭 이벤트,(E)PTZ 적용 후 후기 기간 동안 낮은 진폭 이벤트. 이러한 수치를 얻기 위한 스크립트 파일은 보충 파일로제공됩니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
보충 파일: 3.4 단계에 대한 스크립트 파일. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
토론
간질은 유전염 시퀀싱 기술의 출현으로 해명되기 시작하는 다양한 식신을 특징으로 하는 복잡한 신경질환으로,25,26,27. 다목적 동물 모델은 유전적으로 연결된 간질의 병리학 적 메커니즘에 대한 통찰력뿐만 아니라이 조건의 뚜렷한 형태에 대한 표적 치료법에 대한 통찰력을 모두 얻을 효율적인 번역 전략에 필수적입니다. Zebrafish 모델은 간질의 주요 특징을 재현하고 항 간질 약물 스크리닝5,28에대한 신뢰할 수있는 판독을 제공하는 데 매우 효과적이었다. 자발적인 발작은 유전자 변형 제브라피시(15,29,30,31) 및 이들모델(28)에서 신경생리학적 분석에서 검출될 수 있으며 간질과 같은 행동의 뉴런 기초를확인하였으며(32,33). 소형 제브라피쉬 애벌레는 자발적인 수영과 같은 간단한 동작의 자동 검출을 사용하여 96웰 형식으로 화학 스크린에 사용가능하므로 잠재적 인 치료법을 신속하게 감지 할 수 있습니다.
여기서 제시된 DEPDC5 노크다운 모델은 개발 중 유전자 발현을 차단하기 위해 제브라피시 배아에 AMO를 주입함으로써 얻어진다. 이 모델은 화학 적 또는 유전 적 선별 프로토콜 동안 치료 효율의 지표로 사용할 수있는 애벌레 발달의 다른 시간 지점 동안 여러 키스톤 현상 기능을 제공합니다. AMO 매개 유전자 노크 다운은 화학적으로 유도된 발작 모델에 비해 이점을 표시하는 강력한 기술로, 특히 관심 유전자의 발현을 표적으로 하여 유전 적 돌연변이에 의해 유발되는 근본적인 병원성 메커니즘의 식별을 허용합니다. 화학 유도제, 약물 검열을 위한 그럼에도 불구하고 강력한 공구는, 연구 하에 유전 돌연변이와 항상 관련되지 않을 지도 모르다 다중 세포 통로를 통해 행동할 수 있습니다. AMO 주입 자체가 실험자에 의해 마스터 할 때 간단한 기술이지만, 또한 한계의 번호를 제시한다. 주사는 1 세포 단계 배아에서 수행되어야합니다. 우리의 손에, 나중에 주사 크게 표현형의 가변성을 증가. 이것은 주입에 사용할 수있는 시간을 제한; 따라서 시간 순서로 주사를 위해 계란을 생성하는 전략이 유용하다. 우리는 정기적으로 우리가 15-20 분 간격으로 열리는 4-5 크로스를 사용하여 다음 클러치를 얻기 전에 하나의 클러치를 주입 할 수 있습니다. 또한, 고정관념이 있는 행동이 개발 의 첫 날 동안 급속히 진화함에 따라 서로 다른 실험 사이의 동시에 표현형을 평가하기 위해 주의를 기울여야 합니다. 과도한 양을 주입하여 인한 일반적인 독성이 특정 표현형을 가리기 때문에 AmOs의 부피 및 농도도 신중하게 제어되어야 합니다. 소개에 제시된 다른 컨트롤은 올바른 주사 용량 및 해당 표현형을 결정하는 데 필수적입니다.
애벌레 얼룩말 뇌의 필드 기록은 글로벌 신경 활동에 다른 뇌 질환에 관여하는 유전 적 돌연변이의 해로운 효과를 조사하기위한 유용한 도구입니다(34). 이러한 실험 조건하에서 볼 수 있는 탈극성 이벤트는 상이한 간질 조건에서 약물의 전기생리학적 효과를 평가하기 위한 확립된방법이다(15,35). 그러나 이러한 효과에 대한 평가는 대부분 정량적으로 보다는 질적으로 수행되었으며, 분석에서 배우로서 주관적인 관찰자를 갖는 것입니다. 여기서, 우리는 탈극화의 비율, 진폭 및 지속 시간을 객관적으로 정량화할 수 있는 자동 검출 전략을 개발하고, 시간 마다, 또는 다른 유전 또는 약리학적 내정간섭으로 이 매개변수의 진행상황을 평가할 수 있습니다.
여기에 제시된 대표적인 결과는 간질형과 같은 전기촬영 활성을 소개하기 위하여 PTZ의 적용 전후에 4-6 dpf 제브라피시의 불일치 대조군과 비교하여 DEPDC5 녹다운 유전 모형의 예상필드 활성을 보여줍니다. 이전에는 DEPDC5 녹다운조건(18)의기저 활성이 현저한 증가를 보였다. 여기서, 우리는 이러한 두 가지 조건의 반응이 화학 간질 활성 유도제인 PTZ에 대한 반응으로, 비교적 낮은 주파수, 고진폭 탈극성 발생의 기간으로 시작하여 더 높은 주파수, 낮은 진폭 탈극성 이벤트의 기간으로 계속되는 시간에 유사한 궤적을 가지고 있음을 보여준다. 필드 레코딩 이벤트는 느린 역학(관심 의 주파수는 0.005-0.2s-1의범위에 있음)을 가지므로 이 프로토콜에서 로우 패스 및 하이패스 필터가 모두 사용되어 관심 있는 이벤트를 격리합니다. 저주파 노이즈를 제거한 후 간단한 임계값을 사용하여 탈극화 이벤트를 감지합니다. 신호의 통계는 탈극화 이벤트의 존재에 의해 크게 영향을 받고 있기 때문에, 우리는이 임계 값을 결정하기 위해 총 신호의 표준 편차를 사용할 수 없습니다. 데이터 집합 전반에 걸친 표준 편차 값의 가변성은 관찰된 기록 노이즈 수준보다 컸습니다. 따라서, 추적의 육각 검사 후, 우리는 탈극화 활성의 상이한 수준에 의해 유도된 편견을 피하기 위해 0.3 mV의 임계값의 고정값을 사용했다.
상기 기질프로토콜은 제브라피시 모델에서 간질편형 표현형을 특성화하기 위해, 시광텍투석에서 의극화 이벤트의 자동 검출과 결합된 세포외 전류 클램프 전압 레코딩을 통해 모터 동작 및 뉴런 필드 활성을 평가하는 표준화되고 간단한 방법을 제공한다.
공개
저자는 공개 할 것이 없습니다.
감사의 말
신경생리학 실험이 수행된 ICM 전기생리학 플랫폼의 직원에게 감사드립니다. 우리는 또한 기술적 인 도움을 앙카 마리안에게 감사드립니다. SC는 트램폴린 그랜트 #21488에 의해 지원되었다. EK는 AFM 그랜트 #18469 및 ERC 통합자 그랜트 (ALS-네트워크)에 의해 지원되었다. HC는 퐁디오 라 레체르체 메디칼(PLP20141031462)과 ARSLA에서 박사 학위를 받았습니다. AD 및 RM의 경우, 이 작품은 루마니아 국립 과학 연구 및 혁신 당국의 CNCS-UEFISCDI(프로젝트 번호 PN-III-P4-ID-PCE-2016-0010)의 3건의 보조금으로 지원되었습니다. PN-III-P2-2.1-PED-2016-0007, 그리고 COFUND-NEURON-NMDAR-PSY), 유럽 연합의 호라이즌 2020 연구 및 혁신 프로그램에 의해 보조금 - 보조금 계약 번호 668863-SyBil-AA, 국립 과학 재단은 NSF-IOS-16560에 의해 자금을 지원했다.
자료
| Name | Company | Catalog Number | Comments |
| Agarose | Sigma-Aldrich, France | A9539 | |
| Aquarium salt | Instant Ocean, Blacksburg, VA | SS15-10 | |
| Borosilicate glass with filament | Sutter Instruments | BF100-50-10 | OD: 1.5mm, ID: 0.5 mm |
| CaCl2 | Sigma-Aldrich, France | C1016 | |
| Depdc5-atg antisense morpholino | GeneTools, OR, USA | N/A | sequence 5’- TGCCTTCATGGTGACCGTCATTTTA -3’ |
| Depdc5-mis antisense morpholino | GeneTools, OR, USA | N/A | sequence 5’- TGCgTTgATcGTGACCcTgATTTTA -3’ |
| Depdc5-splice antisense morpholino | GeneTools, OR, USA | N/A | sequence 5’- ACATTCCTGTTTCACCATAGATGAT -3’ |
| Digitizer | Molecular Devices, CA, USA | Digidata 1550 | |
| Fast Green Dye | Sigma-Aldrich, France | F7258 | Stock solution of 0.2% |
| Glass-bottom petri dishes | Ibidi, Germany | 81218 | |
| Glucose | Sigma-Aldrich, France | 68270 | |
| Grasshopper 2 camera | FLIR, BC, Canada | GRAS-03K2M-C | formerly Point Grey Research |
| HEPES | Sigma-Aldrich, France | H3375 | |
| Human wild-type DEPDC5 cDNA | Dharmacon, France | NM_001242897.1 | Accession: BC144291 Clone ID 905 |
| ImageJ software | NIH, USA | N/A | |
| KCl | Sigma-Aldrich, France | P9333 | |
| Matlab software | MathWorks, MA, USA | N/A | |
| MgCl2 | Sigma-Aldrich, France | M2670 | |
| NaCl | Sigma-Aldrich, France | S7653 | |
| NaOH | Sigma-Aldrich, France | 71687 | |
| Pancuronium bromide | Alomone Labs | P-130 | Stock solution of 60 mM in water |
| Parafilm | Sigma-Aldrich, France | P7793 | |
| Patch clamp amplifier | Molecular Devices, CA, USA | MultiClamp 700B | Computer-controled patch clamp amplifier |
| pClamp10 acquisition software | Molecular Devices | N/A | |
| Pentylenetetrazol (PTZ) | Sigma-Aldrich, France | P6500 | Stock solution of 300 mM (dissolved in recording solution) |
| Pipette puller | Narishige, Japan | PC-10 | |
| Pneumatic PicoPump | WPI, France | PV 820 | |
| Sylgard 184 kit | Sigma-Aldrich Intl. | 761036 | |
| Transfer plastic pipettes | Sigma-Aldrich, France | Z350605 | |
| Zebralab | Viewpoint, France | N/A |
참고문헌
- Kabashi, E., Champagne, N., Brustein, E., Drapeau, P. In the swim of things: Recent insights to neurogenetic disorders from zebrafish. Trends in Genetics. 26 (8), 373-381 (2010).
- Baxendale, S., van Eeden, F., Wilkinson, R. The Power of Zebrafish in personalised medicine. Advances in Experimental Medicine and Biology. , (2017).
- Baraban, S. C., Taylor, M. R., Castro, P. A., Baier, H. Pentylenetetrazole induced changes in zebrafish behavior, neural activity and c-fos expression. Neuroscience. , (2005).
- Cunliffe, V. T. Building a zebrafish toolkit for investigating the pathobiology of epilepsy and identifying new treatments for epileptic seizures. Journal of Neuroscience Methods. , (2016).
- Griffin, A., Krasniak, C., Baraban, S. C. Advancing epilepsy treatment through personalized genetic zebrafish models. Progress in Brain Research. , (2016).
- Griffin, A., Hamling, K. R., Knupp, K., Hong, S. G., Lee, L. P., Baraban, S. C. Clemizole and modulators of serotonin signalling suppress seizures in Dravet syndrome. Brain. , (2017).
- Orellana-Paucar, A. M., et al. Insights from zebrafish and mouse models on the activity and safety of ar-turmerone as a potential drug candidate for the treatment of epilepsy. PLoS ONE. , (2013).
- Baxendale, S., et al. Identification of compounds with anti-convulsant properties in a zebrafish model of epileptic seizures. Disease Models & Mechanisms. , (2012).
- Bar-Peled, L., et al. A tumor suppressor complex with GAP activity for the Rag GTPases that signal amino acid sufficiency to mTORC1. Science. 340 (6136), 1100-1106 (2013).
- Ishida, S., et al. Mutations of DEPDC5 cause autosomal dominant focal epilepsies. Nature Genetics. , (2013).
- Dibbens, L. M., et al. Mutations in DEPDC5 cause familial focal epilepsy with variable foci. Nature Genetics. , (2013).
- Baulac, S., Weckhuysen, S. . DEPDC5-Related Epilepsy. GeneReviews®. , (1993).
- Picard, F., et al. DEPDC5 mutations in families presenting as autosomal dominant nocturnal frontal lobe epilepsy. Neurology. , (2014).
- Teng, Y., et al. Knockdown of zebrafish lgi1a results in abnormal development, brain defects and a seizure-like behavioral phenotype. Human Molecular Genetics. , (2010).
- Baraban, S. C., Dinday, M. T., Hortopan, G. A. Drug screening in Scn1a zebrafish mutant identifies clemizole as a potential Dravet syndrome treatment. Nature Communications. , (2013).
- Suls, A., et al. De novo loss-of-function mutations in CHD2 cause a fever-sensitive myoclonic epileptic encephalopathy sharing features with dravet syndrome. American Journal of Human Genetics. , (2013).
- Grone, B. P., et al. Epilepsy, behavioral abnormalities, and physiological comorbidities in syntaxin-binding protein 1 (STXBP1) mutant zebrafish. PLoS ONE. , (2016).
- de Calbiac, H., et al. DEPDC5 knockdown causes mTOR-dependent motor hyperactivity in zebrafish. Annals of Clinical and Translational Neurology. , (2018).
- Panchaud, N., Péli-Gulli, M. P., De Virgilio, C. Amino acid deprivation inhibits TORC1 through a GTPase-activating protein complex for the Rag family GTPase Gtr1. Science Signaling. , (2013).
- Lim, K. R. Q., Maruyama, R., Yokota, T. Eteplirsen in the treatment of Duchenne muscular dystrophy. Drug Design, Development and Therapy. , (2017).
- Kok, F. O., et al. Reverse genetic screening reveals poor correlation between morpholino-induced and mutant phenotypes in zebrafish. Developmental Cell. , (2015).
- Rossi, A., et al. Genetic compensation induced by deleterious mutations but not gene knockdowns. Nature. , (2015).
- Stainier, D. Y. R., et al. Guidelines for morpholino use in zebrafish. PLoS Genetics. , (2017).
- Kimmel, C. B., Ballard, W. W., Kimmel, S. R., Ullmann, B., Schilling, T. F. Stages of embryonic development of the zebrafish. Developmental dynamics an official public. 203 (3), 253-310 (1995).
- Møller, R. S., Dahl, H. A., Helbig, I. The contribution of next generation sequencing to epilepsy genetics. Expert Review of Molecular Diagnostics. , (2015).
- Allen, A. S., et al. De novo mutations in epileptic encephalopathies. Nature. , (2013).
- Dunn, P., et al. Next generation sequencing methods for diagnosis of epilepsy syndromes. Frontiers in Genetics. , (2018).
- Grone, B. P., Baraban, S. C. Animal models in epilepsy research: Legacies and new directions. Nature Neuroscience. , (2015).
- Zhang, Y., et al. Pharmacological characterization of an antisense knockdown zebrafish model of Dravet syndrome: Inhibition of epileptic seizures by the serotonin agonist fenfluramine. PLoS ONE. , (2015).
- Swaminathan, A., et al. Non-canonical mTOR-Independent Role of DEPDC5 in Regulating GABAergic Network Development. Current Biology. , (2018).
- Samarut, &. #. 2. 0. 1. ;., et al. γ-Aminobutyric acid receptor alpha 1 subunit loss of function causes genetic generalized epilepsy by impairing inhibitory network neurodevelopment. Epilepsia. , 2061-2074 (2018).
- Turrini, L., et al. Optical mapping of neuronal activity during seizures in zebrafish. Scientific Reports. , (2017).
- Rosch, R. E., Hunter, P. R., Baldeweg, T., Friston, K. J., Meyer, M. P. Calcium imaging and dynamic causal modelling reveal brain-wide changes in effective connectivity and synaptic dynamics during epileptic seizures. PLoS Computational Biology. , (2018).
- Baraban, S. C. Forebrain Electrophysiological Recording in Larval Zebrafish. Journal of Visualized Experiments. , (2013).
- Afrikanova, T., et al. Validation of the Zebrafish Pentylenetetrazol Seizure Model: Locomotor versus Electrographic Responses to Antiepileptic Drugs. PLoS ONE. , (2013).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유