
Method Article
Hematopoetik Kök ve Progenitör Hücreler için Akış Sitometrisi ile Akrabalık, Bölünme Sayısı ve Fenotipin Eşzamanlı Değerlendirilmesi
Bu Makalede
Özet
Burada sunulan, hücre bölünmelerinin sayısını, yüzey hücresi fenotipini ve hücresel akrabalığı aynı anda ölçmeye izin veren akış sitometrisine dayalı bir tekniktir. Bu özellikler, permütasyon tabanlı bir çerçeve kullanılarak istatistiksel olarak test edilebilir.
Özet
Çok az teknik, aynı hücre için fenotip ve kaderi aynı anda değerlendirebilir. Fenotipi karakterize etmek için kullanılan mevcut protokollerin çoğu, büyük veri kümeleri üretebilmelerine rağmen, ilgili hücrenin yok edilmesini gerektirmekte ve işlevsel kaderini değerlendirmeyi imkansız kılmaktadır. Bu nedenle hematopoez gibi heterojen biyolojik farklılaştırıcı sistemleri tanımlamak zordur. Hücre bölünmesi izleme boyalarına dayanarak, birçok tek hematopoetik progenitör için akrabalık, bölünme numarası ve farklılaşma durumunu aynı anda belirlemek için bir protokol geliştirdik. Bu protokol, çeşitli biyolojik kaynaklardan izole edilen murin ve insan hematopoetik progenitörlerinin ex vivo farklılaşma potansiyelinin değerlendirilmesine olanak tanır. Dahası, akış sitometrisine ve sınırlı sayıda reaktife dayandığından, tek hücreli seviyede, nispeten ucuz bir şekilde hızlı bir şekilde büyük miktarda veri üretebilir. Ayrıca, sağlam bir istatistiksel çerçeve ile birlikte tek hücreli analiz için analitik boru hattı sağlıyoruz. Bu protokol, hücre bölünmesi ve farklılaşmasının tek hücre düzeyinde bağlanmasına izin verdiğinden, simetrik ve asimetrik kader taahhüdünü, kendini yenileme ve farklılaşma arasındaki dengeyi ve belirli bir taahhüt kaderi için bölünme sayısını nicel olarak değerlendirmek için kullanılabilir. Toplamda, bu protokol, hematopoetik progenitörler arasındaki biyolojik farklılıkları tek hücreli bir bakış açısıyla çözmeyi amaçlayan deneysel tasarımlarda kullanılabilir.
Giriş
Son on yıl, hücresel ve moleküler biyolojiye tek hücreli yaklaşımların dünya çapında yayılmasıyla işaretlendi. Tek hücreli genomik 1,2'nin adımlarını izleyerek, günümüzde her yıl gelişen yeni tek hücreli -omik tekniklerle tek bir hücrenin birçok bileşenini (örneğin, DNA, RNA, proteinler) incelemek mümkündür. Bu teknikler, hem insan hem de model organizma hücrelerini kullanarak immünoloji, nörobiyoloji, onkoloji ve diğerleri için eski ve yeni sorulara ışık tutmuştur3. Tek tek hücreler arasındaki farklılıkları vurgulayarak, tek hücreli -omikler, hematopoetik kök ve progenitör hücrelerin (HSPC'ler) heterojenliğine odaklanan ve klasik ayrık homojen popülasyon modelinden uzaklaşan yeni bir hematopoez modelinin tanımlanmasını sağlamıştır 4,5.
Tüm -omik tekniklerin birkaç dezavantajından biri, işlevselliğini değerlendirme olasılığını ortadan kaldırarak, ilgili hücrenin tahrip edilmesidir. Tersine, tek hücreli transplantasyon testi ve soy izleme teknolojileri gibi diğer tek hücreli yöntemler, bireysel hücrelerin in vivo 6,7 kaderini değerlendirerek ata hücrenin işlevselliğinin okunmasını sağlar. Soy izleme teknolojileri, ilgilenilen hücrenin kalıtsal bir genetik7 veya floresan etiket 8,9 ile etiketlenmesini içerir ve aynı anda birden fazla tek hücrenin kaderinin izlenmesini sağlar. Bununla birlikte, başlangıç hücrelerinin karakterizasyonu tipik olarak, akış sitometrisi10 tarafından değerlendirilen birkaç yüzey proteininin ekspresyonu gibi sınırlı sayıda parametre ile sınırlıdır. Ek olarak, tek hücreli soy izleme teknolojileri, tipik olarak DNA / RNA dizilimi veya görüntüleme yoluyla hücresel etiketin zahmetli bir şekilde algılanmasını gerektirir. Özellikle bu son nokta, tek bir deneyde test edilebilecek koşulların sayısını sınırlar.
Tek hücrelerin işlevselliğini incelemek için kullanılan bir başka yöntem sınıfı, tek HSPC'lerin ex vivo hücre kültürleme sistemleridir. Gerçekleştirilmesi kolay, bu altın standart testler, bireysel hücrelerin 96 kuyucuklu hücre kültürü damarlarına ayrılmasını ve kültürden sonra, hücre soy fenotipini, tipik olarak akış sitometrisi veya morfolojik analiz ile karakterize etmeyi içerir. Bu testler çoğunlukla, HSPC'lerin olgun hücrelere uzun vadeli farklılaşmasını karakterize etmek için, tipik olarak 2-3 haftalık kültür11,12'den sonra kullanılmıştır. Alternatif olarak, insan kök hücre nakli için tıbbi fayda vaadi ile ex vivo HSPC'leri 13,14,15,16,17,18'i korumaya ve genişletmeye çalışmak için kullanılmıştır 19. Son olarak, HSPC'lerin kısa süreli kültür20 kullanarak erken taahhüdünü incelemek için kullanılmıştır, bu kültürde üretilen düşük hücre sayısı ana sınırlayıcı faktördür. Bu farklı ex vivo tahlil türlerinin bir dezavantajı, in vivo karmaşıklığı sadece kısmen yansıtmalarıdır; Yine de, insan HSPC farklılaşmasını incelemenin nadir yollarından biridir.
Mevcut tek hücreli yöntemlerden (tek hücreli omikler, soy izleme ve ex vivo kültür) eksik bir bilgi parçası, HSPC dinamikleri21'i incelerken göz önünde bulundurulması gereken önemli bir parametre olan hücre bölünmelerinin doğru tespitidir. Akış sitometrisi yoluyla bölünme sayısını değerlendirmenin basit bir yolu, 5-(ve 6)-karboksifloresein diasetat süksinimidil ester (CFSE)22 gibi çözünür "protein boyalarının" kullanılmasıdır. Bu bölünme boyaları, boyanmış hücrelerin sitoplazması içinde yayılır ve yarı yarıya seyreltilir ve her hücre bölünmesinde iki yavru hücreye geçerek 10 bölüme kadar numaralandırmaya izin verir. Birkaç bölme boyasını birleştirerek, her bir boya farklı torunların ayrılmasına izin verdiği için aynı kuyuda birden fazla bireysel progenitörün tohumlanması mümkündür. Bu, ilk olarak murin lenfositler23,24 için tanıtılan multipleks klonal ve bölünme takibi için hücre boyalarının kullanılmasının arkasındaki prensiptir.
Burada, murin ve insan HSPC'leri ile kullanılmak üzere MultiGen testinin geliştirilmesini sunuyoruz. Birçok tek hücrenin farklılaşma, bölünme ve ex vivo akrabalık özellikleri için aynı anda test edilmesine izin verir. Bu yüksek verimli, gerçekleştirilmesi kolay ve ucuz tahlil, hücresel fenotipi, gerçekleştirilen bölünme sayısını ve kuyudaki diğer hücrelerle hücre akrabalığını ve klonal ilişkiyi aynı anda ölçmeye izin verir. Simetrik ve asimetrik kader taahhüdünü, kendini yenileme ve farklılaşma arasındaki dengeyi ve belirli bir taahhüt kaderi için gerekli olan bölünme sayısını nicel olarak değerlendirmek için kullanılabilir. Protokol, bir floresan aktif hücre sıralayıcısı (FACS) ve bir plaka okuyuculu bir akış sitometresi ve ayrıca hücre kültürünü gerçekleştirmek için gerekli ekipmanı gerektirir. Tahlilin insan HSPC'leri üzerinde yürütülmesi için teknik protokole ek olarak, hücre ailesi25 kavramıyla ilgili hücresel özellikleri değerlendirmek için gerekli istatistiksel testler de dahil olmak üzere ayrıntılı analiz çerçevesini de sağlıyoruz. Bu protokol, murin HSPC bölmesi26,27'yi tanımlamak için başarıyla kullanılmıştır.
Aşağıdaki protokol, başlangıç malzemesi28 olarak manyetik olarak zenginleştirilmiş CD34+ hücrelerini kullanır. Bu şekilde, insan HSPC'lerini farklı kan kaynaklarından (örneğin, kordon kanı, kemik iliği, periferik kan) etkili bir şekilde boyamak ve izole etmek mümkündür. CD34 fraksiyonunu atmamak önemlidir, çünkü farklı deneysel kontrol türlerini ayarlamak için protokolün bir parçası olarak kullanılacaktır. Söz konusu hücre miktarları ve hacimleri, deneysel iş akışına ve gereksinimlere göre yukarı veya aşağı ölçeklendirilebilir. Benzer şekilde, protokol, hücre sıralama ve akış sitometrisi adımları için kullanılan antikorları değiştirerek, farklı tipte progenitörlerin çalışmasına uyarlanabilir.
Protokol
Aşağıdaki protokol için, HSPC kaynağı olarak tanımlanmamış göbek kordon kanı kullanılmış ve Saint-Louis Hastanesi kordon kanı biyobankası (AC-2016-2759 yetkilendirmesi) ve Helsinki Deklarasyonu tarafından tanımlanan kılavuzlara uygun olarak toplanmıştır.
NOT: Başlamadan önce, bu protokol için gerekli tüm reaktiflerin ve ekipmanın, Malzeme Tablosunda listelendiği ve protokolde belirtildiği gibi mevcut olduğundan emin olun. İlgili reaktifleri taze hazırlayın ve açıkça belirtilmedikçe saklamayın.
1. Hücre boyası boyama
NOT: Bu bölümde, CFSE ve mor boya (CTV) hücre bölmeli boyaların dört kombinasyonu ile boyama açıklanmaktadır. Hiçbir hücre boyası çözeltisi eklenmemiş olsa bile, tüm tüpleri aynı anda işleyin. Tüm adımlar, aşağıdaki hücre kültürü adımına izin vermek için steril koşullarda gerçekleştirilir. Gereken süre: yaklaşık 100 dk.
- Kordon kanı ünitesini manyetik bir ayıklama protokolüne göre işleyin29. İki fraksiyonun mevcut olduğundan emin olun: büyük bir CD34 fraksiyonu ve daha küçük bir CD34 + fraksiyonu. Her iki tüpü de 300 x g'de 5 dakika döndürün. Pelet bozmadan süpernatantı aspire edin.
- CD34 + fraksiyonu için, fetal sığır serumu (FBS) olmadan Dulbecco'nun modifiye Eagle besiyerinin (DMEM) 1 mL'sinde yeniden askıya alın. Bir hemositometre kullanarak hücreleri sayın; hücre yoğunluğu 3 x 106 hücre/mL'den yüksek olmamalıdır. Bu durumda, ses seviyesini buna göre ayarlayın. CD34 fraksiyonu için, FBS olmadan DMEM'de yeniden askıya alın ve ses seviyesini maksimum 6 x 106 hücre / mL'ye ayarlayın.
- CD34+ fraksiyonunun 250 μL'sini dört adet 15 mL polipropilen tüpe alikotlayın. Tüpleri aşağıdaki gibi etiketleyin: CD34+/CF (CFSE_only), CD34+/CV (CFSE_high CTV_low), CD34+/VC (CFSE_low CTV_high) ve CD34+/VI (CTV_high). CD34 fraksiyonunun 250 μL'sini başka bir dört adet 15 mL polipropilen tüpe alikotlayın. Tüpleri aşağıdaki gibi etiketleyin: CD34-/CF (CFSE_only), CD34-/CV (CFSE_high CTV_low), CD34-/VC (CFSE_low CTV_high) ve CD34-/VI (CTV_high). CD34 fraksiyonundan kalan hücreler atılabilir.
- CFSE_high ve CFSE_low adlı iki CFSE çözümü hazırlayın. CFSE_high (10 μM) için, FBS olmadan 1,1 mL DMEM'i 2,2 μL CFSE stok (5 mM) çözeltisiyle karıştırın. CFSE_low (5 μM) için, FBS olmadan 550 μL DMEM ve 0,55 μL CFSE stok (5 mM) çözeltisini karıştırın.
- CF ve CV tüplerine 250 μL CFSE_high çözeltisi, VC tüplerine 250 μL CFSE_low çözeltisi ve VI tüpüne FBS olmadan 250 μL DMEM ekleyin. Hücre süspansiyonu ve hücre boyasının verimli bir karışımını sağlamak için, tüpü neredeyse 90 derece eğin ve CFSE çözeltilerini tüp duvarına yatırın. Ardından, iki çözeltiyi karıştırmak için tüpü dikey olarak tutun ve CFSE çözeltilerinin askıya alınmış hücrelerle hızlı bir şekilde karıştırılmasını sağlamak için üç veya dört kez pipet yapın. 37 ° C'de tam olarak 8 dakika boyunca inkübe edin.
- Kuluçkadan sonra, 5 mL DMEM +% 10 FBS ekleyin. Tüpleri 5 dakika boyunca 37 ° C'de tutun.
- Tüpleri 300 x g'de 5 dakika döndürün. Peletleri rahatsız etmeden süpernatantı aspirasyon yoluyla çıkarın ve peleti 5 mL fosfat tamponlu salin 1x / etilendiamintetraasetik asit (PBS 1x / EDTA) ile yıkayın. 300 x g'de 5 dakika boyunca tekrar döndürün. Peletleri rahatsız etmeden süpernatantı atın ve hücre peletini 250 μL 1x PBS / EDTA'da yeniden askıya alın.
- CTV_high ve CTV_low adlı iki CTV çözümü hazırlayın. CTV_high (10 μM) için 1,1 mL PBS 1x/EDTA ve 2,2 μL CTV stoğunu (5 mM) karıştırın. CTV_low (5 μM) için 550 μL PBS 1x/EDTA'yı 0,55 μL CTV stoğu (5 mM) ile karıştırın.
- VC ve VI tüplerine 250 μL CTV_high çözeltisi, CV tüpüne 250 μL CTV_low çözeltisi ve CF tüpüne 250 μL 1x PBS / EDTA ekleyin. Adım 1.5'te açıklanan tekniğin aynısını kullanın. 37 ° C'de tam olarak 8 dakika boyunca inkübe edin.
- Kuluçkadan sonra, 5 mL DMEM +% 10 FBS ekleyin. 5 dakika boyunca 37 ° C'de tutun.
- Tüpleri 300 x g'de 5 dakika döndürün, süpernatantı peleti rahatsız etmeden atın ve ardından peleti 5 mL 1x PBS / EDTA ile yıkayın. 300 x g'de 5 dakika boyunca tekrar döndürün.
- Peletleri rahatsız etmeden süpernatantı atın ve CD34- fraksiyonlarını 1x PBS / EDTA'da 1.5 x 106 hücre / mL'lik son konsantrasyon için yeniden askıya alın. CD34+ fraksiyonlarını 40 μL boyama tamponunda tekrar askıya alın ve hücreleri 1,5 mL tüplere aktarın.
2. Antikor boyama
NOT: Antikor boyama deneysel ihtiyaçlara göre özelleştirilebilir. Sadece CD34 + fraksiyonları antikor boyamasına maruz kalır; CD34- fraksiyonları, hücre bölünmesi boya kombinasyonları (CV, VC, CF ve VI fraksiyonları) için tek bir boyama kontrolü olarak kullanılır. Aşağıdaki panel, dört tip HSPC'nin tespiti için uyarlanmıştır: hematopoetik kök hücreler (HSC'ler), multipotent progenitörler (MPP'ler), lenfoid astarlı multipotent progenitörler (LMPP'ler) ve hematopoetik progenitör hücreler (HPC'ler)12. Bununla birlikte, HSC'lerin ve MPP'lerin tanımlanması sunulmaktadır. Gereken süre: 75 dk.
- Kompanzasyon boncukları kullanarak yüzey boyama için tek boyamayı hazırlayın. Negatif boncukları ve İmmünoglobulin G (IgG) boncuklarını 1:1 oranında, 20 μL x yüzey belirteci sayısına eşdeğer toplam hacim için karıştırın (örneğin, boyama paneli altı antikor içeriyorsa 120 μL).
- Her bir işaretleyici için ayrı ayrı 1,5 mL tüplere 20 μL boncuk gönderin. İlgili tüpteki her antikor için seyreltme faktörüne karşılık gelen hacmi ekleyin (örneğin, seyreltme faktörü 1:20 ise, 1 μL ekleyin).
- CD34+ hücrelerini boyamak için, Tablo 1'e dayanan bir ana antikor12 karışımı hazırlayın. Antikorları tek bir 0.5 mL tüpe karıştırın. Dört CD34+ koşulunun her birine antikor ana karışımından 7 μL ekleyin.
- Kompanzasyon boncuklarını ve CD34+ numunelerini 4°C'de en az 30 dakika boyunca inkübe edin.
NOT: Kuluçka süresi, boyama için kullanılan antikorların teknik detaylarına uyarlanmalıdır. - İnkübasyon sırasında, sıralama için kullanılacak 96 delikli yuvarlak tabanlı plakayı hazırlayın ve çok kanallı bir pipet kullanarak her bir kuyucuğa 100 μL hücre kültürü ortamı ekleyin.
NOT: H8-H12 kuyularını boş bırakın. - Yüzey boyama kontrolleri (5, boncuklar kullanılarak), hücre bölme boyaları kontrolleri (4, CD34- fraksiyonları kullanılarak) ve CD34+ numuneleri (4) için 5 mL polipropilen tüpleri etiketleyin.
- İnkübasyonun sonunda, hücreleri ve boncukları 1 mL boyama tamponu ile yıkayın. Toplam hacmi 5 mL polipropilen tüplere aktarın. Tüpleri 300 x g'de 5 dakika boyunca santrifüjleyin, ardından peleti rahatsız etmeden süpernatantı aspire edin.
- Boncuklar ve CD34+ hücreleri için her biri yaklaşık 500 μL ve CD34 tüpleri için 1 mL kullanarak hücreleri boyama tamponunda yeniden askıya alın.
Tablo 1: Bir hücre sıralama deneyi için antikor mastermix'ini hazırlamak için şablon. Bu Tabloyu indirmek için lütfen tıklayınız.
3. Hücre sıralama
NOT: Sıralanmış hücre numaraları, kullanılabilir toplam hücre miktarına göre değişebilir. Protokolde, her denetim için minimum bir hücre numarası sağlanır. Gereken süre (tek bir plaka için): 100 dk.
- Şablon denemesini açın veya yeni bir deneme ayarlayın. Her koşulda bir tane olmak üzere tek bir örnek ve birden fazla tüp oluşturun.
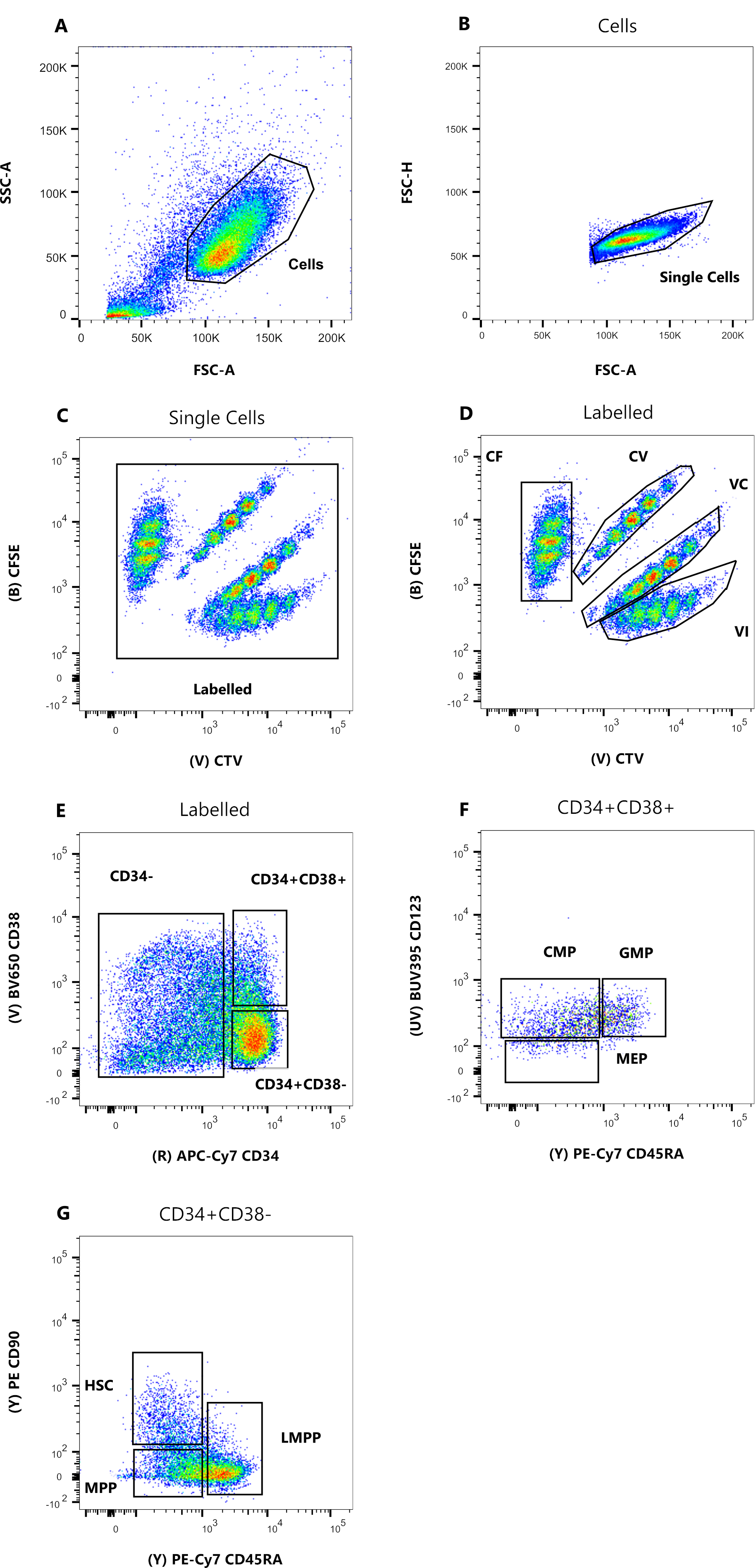
- Altı nokta çizim diyagramı oluşturarak Şekil 1'de ayrıntılı olarak açıklanan geçit stratejisini ayarlayın. İlk olarak, hücreleri bir FSC-A/SSC-A nokta grafiğinde görselleştirin ve düşük kenar saçılımına sahip bir popülasyon seçmek için çokgen geçit aracına çift tıklayın (Şekil 1A). Aşağıdaki nokta grafiğinde (FSC-A / FSC-H ), grafiğe sağ tıklayın ve üzerine tıklayarak açılır menüden "Hücreler" kapısını seçin. İki eksen arasındaki köşegen üzerinde sıkı bir popülasyon seçmek için aynı geçit aracını kullanın (Şekil 1B).
- Üçüncü nokta grafiğinde (APC vs. FSH-H), "Tek Hücreler" popülasyonunu görüntüleyin ve APC Soyu (Lin) ifadesi için negatif hücreleri geçitleyin (Şekil 1C). Dördüncü grafikte (CFSE ve CTV), "Lin-" popülasyonunu görüntüleyin ve her boya kombinasyonu için bir tane olmak üzere dört ayrı kapı oluşturun (Şekil 1D).
NOT: Bu kapılar, homojen olarak boyanmış hücrelerin sadece küçük bir kısmını seçmek için sıkı olmalıdır. - İlgilenilen öncülleri tanımlamak için beşinci ve altıncı grafiği (APC-Cy7'ye karşı BV650 ve PE-Cy7'ye karşı PE) kullanın. Beşinci grafikte CD34+CD38- popülasyonunu ve CD34+CD38+'ı cömertçe geçitleyin (Şekil 1E). Ardından, altıncı grafikte CD34 + CD38- popülasyonunu seçin ve Şekil 1F'ye göre üç kapı çizin.
- Kompanzasyon boncuklarını içeren tek boyama tüplerini çalıştırın, Al düğmesine tıklayın. Fotoçoğaltıcı tüp (PMT) voltajlarını, özellikle hücre bölme boyaları için (iki üstel ölçekte 104 ile 105 arasında) Parametreler açılır menüsünden ayarlayın.
- Ücret matrisini, Ücret sekmesini kullanarak sıralama için kullanılan panele göre hassaslaştırın. Kaydet düğmesine tıklayarak boncuk kapısına en az 5.000 olay kaydedin.
- CD34 kesirlerini çalıştırın ve ücret matrisini tekrar kontrol edin. Tek hücreli geçitte en az 10.000 olay kaydedin.
- CD34+ kesirlerini çalıştırarak tek hücreli geçitte en az 5.000 olay kaydedin. Her boya kombinasyonu için kapıyı ayarlayın, homojen bir popülasyon seçmek için sıkı bir kapı ayarlayın (Şekil 1D). Benzer şekilde, HSC'leri ve MPP'leri seçmek için geçidi ayarlayın.
- Analiz tamamlandıktan ve tüm tüpler kaydedildikten sonra, 96 delikli plakalarda sıralama için standart Aria kalibrasyonunu gerçekleştirdikten sonra plakayı uygun tutucuya yerleştirin. Plakanın soğutulması önerilir.
- Plaka sıralama şablonunu, deneme sıralama düzenini kullanarak Tablo 2'de sunulan şemaya göre hazırlayın. "CD34-" olarak adlandırılan kuyular, CF / CV / VC / VI kapısında sıralanmış 5.000-10.000 hücre içerir. "Toplu" kuyular, CD34 + CD38- kapısında sıralanmış en az 500 hücre içerir. Son olarak, tek hücreli kuyular, kuyu başına hücre bölünmesi boya kombinasyonu başına yalnızca bir olay içerir, bu nedenle toplamda kuyu başına dört olay içerir.
NOT: "Toplu" popülasyonlar, belirli progenitörlerin alt kümesine uyarlanabilir; 500'den az hücreyi sıralamayın. - Sıralama için, bir sonrakine geçmeden önce her hücre bölme boya kombinasyonunu tamamlayarak sırayla devam edin. Örneğin, CD34-CF'yi verim saflığı modunda sıralamaya başlayın. Al düğmesine, ardından sıralama düğmesine tıklayın.
- CD34- sıralamanın sonunda, CF CD34+ tüpünü takın. Edinin, ardından saflık derecesi olarak 0/ 16/0 işaretlediğinizden emin olarak sıralama düğmesine tıklayın. Son olarak, ilgilenilen hücreleri, kuyu başına bir hücre, tek hücreli saflıkta sıralayın ve Dizin Sıralama seçeneğini işaretlediğinizden emin olun.
- Aynı sırayı tekrarlayarak aşağıdaki hücre bölmeli boya kombinasyonuna geçin. Referans olarak, Tablo 2 sıralanmış bir plaka örneği sunmaktadır.
NOT: Dizin sıralama işlevi, sıralanan her koşul için ayrı dosyalar oluşturur. - Sıralamanın sonunda, dosyaları .fcs 3.0 dosyaları olarak dışa aktarın. Hücreleri 37 ° C,% 5 CO2 inkübatöre koyun. Hücreler, deneysel tasarıma göre, en az 24 saat26 gün boyunca kültürlenir.
Tablo 2: Ardışık akış sitometrisi analizi için özel gereksinimlere dayanan 96 delikli plakayı sıralayan bir hücre şablonu şablonu. Bu Tabloyu indirmek için lütfen tıklayınız.
4. Hücre sıralama veri analizi
NOT: Hücre sıralamanın kalitesini doğrulamak için, ilerlemeden önce FACS veri analizi gereklidir. Bu adımın ana çıktısı, sıralanan her bir hücrenin işaretleyici yoğunluklarını içeren bir elektronik tablonun oluşturulmasıdır.
- .fcs 3.0 dosyalarını analiz yazılımına yükleyin.
- Gerçek sıralamadan önce kaydedilen tek boyama dosyalarını kullanarak hücre sıralama sırasında kullanılan telafi ayarını doğrulayın.
- Farklı yığına karşılık gelen dosyaları kullanarak gözden geçirilmiş geçit stratejisini ayarlayın. Bu geçitleri kopyalayıp dizin sıralama dosyalarına yapıştırın.
- Dizin sıralanmış hücrelerin ayarlanan kapıya düşüp düşmediğini kontrol edin. Yanlış bir şekilde geçilen bazı sıralanmış hücreler varsa, bunlar dizin sıralama sırasında kaydedilen plaka koordinatları dışa aktarılarak tanımlanabilir ve analizde daha sonra kaldırılabilir.
- Olayı, dizin sıralama dosyalarından telafi edilmiş parametreler olarak dışa aktarın. Bunları "ölçek değerleri" ve "telafi edilmiş parametreler" seçeneklerini işaretleyerek .csv dosyaları olarak dışa aktarın. Bu dosyalar "Dışa aktarılan dosyalar" adlı bir klasöre verilmelidir.
- Ek Dosya 1'deki komut dosyasını kullanarak tüm dosyaları tek bir .csv dosyasında birleştirin. "setwd" işleviyle doğru yolu ayarlayın. Bu komut dosyasının çıktısı, tüm farklı geçitli olayları ve tüm parametrelerin göreli yoğunluklarını içeren bir elektronik tablodur.
5. Kültür sonrası antikor boyama
NOT: Protokolün bu bölümünü steril koşullarda gerçekleştirin; Birkaç reaktif önceki adımlarla paylaşılır ve steril kalması gerekir. Akış sitometrisi analizi için, plaka okuyuculu bir akış sitometresi kullanın. Bu, boyamanın doğrudan doku kültürü plakasında yapılmasına izin verir, pipetleme ve eğirme miktarını sınırlayarak hücre kaybını minimuma indirir. CF/CV/VC/VI renkleri için tek boyamayı temsil eden ve 96 delikli plakada zaten mevcut olan A1-A4 kuyucukları hariç, kompanzasyon boncuklarını kullanarak yüzey işaretleyicisini tek renk boyama hazırlayın. Hücre boyasına göre sıralanan toplu popülasyonlar, bölünme sayısı ve genel geçit için geçit stratejisinin belirlenmesine yardımcı olur. Gereken süre: 120 dk.
- Protokole başlamadan önce, plakayı ters çevrilmiş bir mikroskop altında kontrol ederek en az bir hücre içeren kuyucukları işaretleyin. Bu adım, boyama için kullanılan antikor miktarını optimize etmeyi sağlar ve prosedürü hızlandırır.
- Antikor mastermix'ini Tablo 3'e göre hazırlayın. Önemli miktarda pipetleme olduğundan, tabloda pipetlemeden kaynaklanan teknik hata dikkate alınır ve buna fazladan %5'lik bir hacim de dahildir. Tabloda açıklanan antikorlar, insan kordon kanı örneklerinden bir dizi HSPC'yi karakterize etmeyi sağlar12.
- Plakayı 300 x g'de 5 dakika boyunca santrifüjleyin. Süpernatanı çıkarmak için plakayı kaputun altında ve bir kağıt havlunun üzerinde hızla ters çevirin.
- A1-A4 kuyucuklarına 8 μL boyama tamponu ekleyin. Karışımın 8 μL'sini diğer kuyucuklara ekleyin.
- Negatif boncukları ve IgG kompanzasyon boncuklarını 1:1 oranında, 120 μL'ye eşdeğer toplam hacim için karıştırın. Seyreltme faktörüne karşılık gelen antikor hacmini ekleyin (örneğin, seyreltme faktörü 1:20 ise, 1 μL ekleyin).
NOT: Toplam hacmi, boyama için kullanılan işaretleyici sayısına uyarlayın (örneğin, boyama paneli beş antikor içeriyorsa 100 μL). - Plakayı ve tek boyama telafisi kontrollerini +4 °C'de en az 30 dakika boyunca inkübe edin.
NOT: Kuluçka süresi, boyama için kullanılan antikorların teknik detaylarına uyarlanmalıdır. - Boncukları 1 mL boyama tamponu ile yıkayın. Toplam hacmi daha önce etiketlenmiş 5 mL polipropilen tüplere aktarın. Tüpleri 300 x g'de 5 dakika boyunca santrifüj edin, ardından süpernatantı aspirasyon yoluyla çıkarın.
- Çok kanallı bir pipet kullanarak kuyucuk başına 100 μL boyama tamponu ekleyerek plakadaki hücreleri yıkayın. Plakayı 5 dakika boyunca 300 x g'de santrifüjleyin, ardından süpernatanı çıkarmak için plakayı kaputun altına ve bir kağıt havlunun üzerine hızla ters çevirin.
- Çok kanallı bir pipet kullanarak hücreleri 85 μL boyama tamponunda yeniden askıya alın.
- Analize akış sitometresinde (Edinme modu), özel şablonu kullanarak ve özele tıklayarak başlayın. Bu özelleştirilmiş şablon, 96 delikli yuvarlak tabanlı plakanın teknik özelliklerini, özellikle de her bir kuyucuğun boyutlarını (çap, derinlik ve kalınlık) dikkate alır. Prob kuyunun dibine ulaşmalıdır, bu nedenle A1 ve H12 kuyularının tam merkezine yerleştirin.
- Yazılım tarafından önerilen listeden ilgilenilen floroforları seçtikten sonra, en az bir hücre içeren kuyucuk sayısı için düzeltilmiş Tablo 2'nin plaka şablonunu izleyerek plaka kurulumunu ayarlayın.
- Edinme hacmi sınırı olarak 100 μL'yi seçin. Ajitasyon seçeneğini işaretleyin. Düşük hız, kuyu başına analiz edilen toplam hacmi iyileştirdiği için edinme hızını maksimum 1 μL/sn olarak ayarlayın.
- H8-H12 kuyularına uygun temizleme ve yıkama solüsyonlarını ekleyin. Tablo 2'deki şablon özellikle H8-H12 kuyularını boş bırakır, çünkü akış sitometresinin analizin sonunda bir dizi yıkama koşulunu çalıştırması gerekir.
NOT: Bu adım, kullanılan akış sitometresinin özelliklerine uyarlanmıştır. - Grafikler ve kapılar bölümünde, önce FSC-A/SSC-A dağılım grafiğini, ardından FSC-H/FSC-A dağılım grafiğini kullanarak tek hücreli kapıyı ayarlayın. İlgilendiğiniz her işaretçi için bir histogram oluşturun.
- Ayarlar onaylandıktan sonra Analiz bölümüne geçin. Önce tek boyamayı analiz edin, hem kompanzasyon boncukları hem de CD34 boyalı fraksiyonlar için en az 5.000 olay (optimum aralık: 5.000-15.000 olay) kaydedin . Gerekirse voltajları ayarlayın.
- Tek tek boyamaların tümü kaydedildikten sonra, Edinme işlevine tıklayarak gerçek edinimi başlatmak mümkündür.
Tablo 3: Bir akış sitometri deneyi için, özellikle HSPC'lerin insan kordon kanından tanımlanması için antikor mastermix. Bu Tabloyu indirmek için lütfen tıklayınız.
6. Kültür sonrası akım sitometrisi veri analizi
NOT: Açıklanan veri analizi, Malzeme Tablosunda belirtilen yazılımlara özgüdür. Ana çıktı, analiz edilen her hücre başına yüzey işaretleyici yoğunluğu, bölünme sayısı ve akrabalık için bilgi içeren bir elektronik tablonun oluşturulmasıdır. Protokolün bu bölümünde, son analiz elektronik tablosunu oluşturmak için bu iş akışı için gerekli olan R ile yazılmış bir komut dosyası bulunur.
- Dosyaları akış sitometresinden .fcs dosyaları olarak dışa aktarın. Bunları analiz yazılımına yükleyin, "tek boyama", "toplu" ve "tek hücreli" olarak gruplandırın.
- Tek boyama dosyalarını kullanarak bir ücret matrisi hazırlayın ve sürükleyip bırakarak diğer iki gruba uygulayın.
NOT: Otomatik bir kompanzasyon aracı kullanılıyorsa, devam etmeden önce kaliteyi elle kontrol edin. - Temsili bir geçişe sahip olmak için, farklı toplu kuyuları tek bir dosyada birleştirin. Bu adım, iki rengin çakışıp çakışmadığını (tipik olarak CV ve VC) veya diğer anormallikleri hızla vurgular ve bu nedenle hariç tutulması gerekir. Popülasyonları birleştir seçeneğine tıkladıktan sonra, "parametreler" menüsünden telafi edilmemiş tüm parametreleri seçin ve ardından birleştir'e tıklayın.
- Birleştirilmiş dosyayı çalışma alanına yükleyin, ardından sürükleyip bırakarak ücret matrisini uygulayın.
- Birleştirilmiş dosyayı kullanarak Şekil 2'de tanımlanan geçit stratejisini hazırlayın. Tek hücreli kapıda, olayları CFSE ve CTV ile bir dağılım grafiğinde görüntüleyin. Dört rengin tümünü içeren ve olası otomatik floresanı hariç tutan Etiketli adlı ilk kapıyı oluşturun (Şekil 2C). Ardından, her rengi ayrı ayrı açın.
- CV ve VC ile etiketlenmiş hücreler, rengin CFSE ve CTV sinyallerinin sonucu olduğu göz önüne alındığında, dönüştürülmüş bir değere ihtiyaç duyar. Bu nedenle, koordine edilen iki sinyal, bölünme seyreltmesinin x eksenine paralel olarak ilerlemesine izin vermek için logaritmik bir ölçekte 45 ° döndürülür. Bu dönüştürülen değer, Araçlar'a ve ardından Türev Parametresi'ne tıklanarak manuel olarak türetilir. Aşağıdaki formülü formül kutusuna yapıştırın:

NOT:Denklem 26 , CFSE ve CTV parametrelerinin 03 ve 17 olduğunu varsayar. - Türetilmiş Parametre adlı bu yeni parametreyi doğru bir şekilde görselleştirmek için, ~ 3-7 arasında değişen doğrusal bir eksen ayarlayın, Eksen parametresi seçeneğine tıklayın ve Ekseni Özelleştir'i seçin.
- Geçidi her renge ayrı ayrı histogram grafiği olarak uygulayın: CF ve VI için, sırasıyla x ekseninde CFSE-A ve CTV-A'yı ayarlayın. CV ve VC için, x ekseninde yeni Türetilmiş Parametreyi ayarlayın. Şekil 3'te gösterildiği gibi her tepe noktasına karşılık gelen kapıları ayarlayın.
- Geçidi her bir tek hücreli kuyucuğa uygulayın. Türetilmiş parametreyi analiz edilen her kuyuya eklediğinizden emin olun. Belirli bir zirveye yanlış atanmış olayları algılamak için her bir kuyu için her renk kapısını el ile doğrulayın. Geçit örnekleri Şekil 4'te gösterilmiştir.
- Analiz tamamlandıktan ve tüm kuyular doğrulandıktan sonra, en az bir hücre içeren tüm CF/CV/VC/VI kapılarını seçin. "Ölçek değerleri" ve "telafi edilmiş parametreler" seçeneklerini işaretleyerek bunları .csv dosyaları olarak dışa aktarın. Bu dosyalar "Dışa aktarılan dosyalar" adlı bir klasöre verilir.
- Ek Dosya 1'deki R komut dosyasını kullanarak tüm dosyaları tek bir .csv birleştirin. "Setwd" işleviyle doğru yolu ayarlamayı unutmayın. Bu komut dosyasının çıktısı, tüm farklı geçitli olayları ve tüm parametrelerin göreli yoğunluklarını içeren bir elektronik tablodur.
- Elektronik tabloyu açın ve her parametre için sütunları yeniden adlandırın, örneğin aşağıdaki adları kullanın: CFSE, CTV, CD90, CD123, CD45RA, CD34, CD38. Bu adlar, her hücreye kimliklerini doğru bir şekilde atamak üzere geçiş eşiğini tanımlamak için kullanılacaktır.
- "Peki", "Durum", "Renk", "Nesil", "Original_cell" ve "Culture_time" adlı altı sütun ekleyin. Bu değişkenler deneysel olarak tanımlanan değişkenlerdir ve her satırdan çıkarılır:
CD34 + PBS_CV_Peak 1.csv.1 = A10 (kuyu), CD34+ (Original_cell), PBS (Durum), CV (Renkli), Peak_1 (Nesil) export_A10. - Geçiş için eşik değerleri belirlemek üzere toplu kuyuları dışa aktarın: "ölçek değerleri" ve "telafi edilen parametreler" seçeneklerini işaretleyerek telafi edilen ilgili popülasyonu (örneğin, CD34+CD38-) .csv dosyaları olarak dışa aktarın. Bu dosyaları "Dışa aktarılan dosyalar" adlı bir klasöre dışa aktarın.
- CD38 eşiğini bulmak için, bu parametrenin en büyük sayısal değerini belirleyin. Tersine, CD34 eşiğini bulmak için, bu parametrenin en küçük sayısal değerini tanımlayın. İlgilendiğiniz tüm parametreler için bu işlemi tekrarlayın.
NOT: Protokolde sunulan analiz için, CD45RA işaretleyicisi hem CD34+CD38- kapısındaki LMPP'leri hem de CD34+CD38+ kapısındaki CMP/GMP'yi tanımlamak için kullanılır. Bu, bu işaretleyici için iki farklı eşik değerinin çıkarılması gerektiği anlamına gelir. - Eşik değerlerini kopyalayıp "gating_matrix" adlı bir excel dosyasına yapıştırın. Bu dosya Tablo 4'e göre düzenlenmiştir ve birden fazla bağımsız deneyin analizine izin verir. Her sütunu tam olarak bu şemayla adlandırmak çok önemlidir: XXYYMMDD_xxh, burada XX operatörün iki baş harfini, YY yılın son iki sayısını, ay için MM'yi, gün için DD'yi ve analiz zaman noktasını xx temsil eder.
Tablo 4: İstatistiksel analizden önce hücre kaderi ataması için geçit matrisi. CD45h, HPC alt kümesi geçişi için CD45RA'nın yoğunluğunu, CD45l, CD34 + CD38- alt kümeleri için CD45RA'nın yoğunluğunu ifade eder. Bu Tabloyu indirmek için lütfen tıklayınız.
7. İstatistiksel analiz
NOT: Oluşturulan verilerin istatistiksel testi, Python programlama dili (Ek Dosya 2, Ek Dosya 3 ve Ek Dosya 4) kullanılarak kodlanmış özel yapım bir analiz işlem hattını içerir. Komut dosyası üç blok halinde düzenlenmiştir: birincisi elektronik tabloyu işlemek için, ikinci blok veri görselleştirme için ısı haritası oluşturmak için ve son blok farklılaşma ve bölme özelliklerini analiz etmek ve test etmek için birden fazla histogram oluşturmak için.
- "0_process_data" bloğundan (Ek Dosya 2) başlayarak, gating_matrix ve veri elektronik tablosu yollarının kodda doğru tanımlandığından emin olun.
- Her hücreye ilgili hücre kaderlerini atamak için "cell_cols" sözlüğünü tanımlayın. Spesifik durumda, kaderler HSC'ler, MPP'ler, LMPP'ler, ortak miyeloid progenitörler (CMP'ler), granülo-monositik progenitörler (GMP'ler), megakaryositik-eritroid progenitörler (MEP'ler) ve CD34-'tür.
- Toplu kuyulardan tanımlanan eşik değerlerini kullanarak (adım 6.16), "cell_class_exp_time" işlevini tanımlayın. Adım 6.12'de her sütunu tanımlamak için kullanılan adı kullanarak sütun adlandırmasında tutarlı olmak, bu eşikleri doğru bir şekilde tanımlamak önemlidir.
- Hücre fenotipleri, akış sitometri analizi sırasında tespit edilen eşiklere dayanan bir dizi "if-else" ifadesi kullanılarak kodda tanımlanır.
NOT: Bu ifadeler diğer işaretleyici kombinasyonlarına uyacak şekilde değiştirilerek farklı fenotipler görüntülenebilir. - "cond_rule" fonksiyonunu kullanarak deneysel spesifik koşulları belirtin (örneğin, farklı deneysel tedaviler). Sağlanan veri kümesi için koşullar "GT" ve "Diff" olarak adlandırılır. Hücreleri kültürlemek için kullanılan iki farklı hücre kültürü ortamını tanımlayın. Bu bilgi, ısı haritasını çizmek için "1_dot_plot" bloğu (Ek Dosya 3) tarafından kullanılacaktır.
- "2_bar_plot" bloğunda (Ek Dosya 4), ilgilenilen ayrık hücre kaderleri de dahil olmak üzere "class_dct" sözlüğünü tanımlayın. Sağlanan veri kümesi için, ilgilenilen hücre kaderleri "cell_cols" sözlüğünde açıklananla aynıdır.
- "Conds" (koşullar), "or_cells" (orijinal hücre), "sym_labs" (simetri etiketleri) ve "zamanları" (deneysel zaman noktası) tanımlayın. Bunlar, çizim için gerekli olan yinelenen filtrelerdir. "Conds" yine "cond_rule" da tanımlanan koşulları alır, "or_cells" HSC'ler ve MPP'lerdir ve "sym_labs" bölümlerin türünü tanımlar.
- "2_bar_plot" bloğunda, bölüm 6'ya kadar ilerleyen hücreleri çizmek mümkündür.
NOT: Sağlanan veri kümesi yalnızca bölüm 4'e kadar olan hücreleri içerir, bu nedenle bir hata iletisi açılır, ancak bu komut dosyasının çalışmasını engellemez. - Script tarafından oluşturulan şekiller pdf dosyaları olarak "şekiller" adlı klasöre alınabilir. "Test" adlı dosyalar, karşılık gelen histogram için gerçekleştirilen farklı istatistiksel testleri temsil eder.
Sonuçlar
FACS sıralama
Bu protokolde sunulan sıralama geçit stratejileri yaygın olarak kabul gören stratejilere dayanmaktadır 12,30,31. Şekil 1'de sunulan geçit stratejisi için, başlangıç materyali daha önce CD34 + manyetik zenginleştirme yoluyla saflaştırılan kordon kanı progenitörleridir, bu da Lineage pozitif hücrelerinin ihmal edilebilir yüzdesini açıklar. Dört hücre içi boya kombinasyonu (örneğin, şekildeki CTV için) için sıkı kapılar kullanmak, aşağıdaki analiz sırasında piklerin çözünürlüğünü artırmak ve doğru hücre popülasyonunu geçmek için gereklidir (Şekil 1D). Şekilde gösterilen durumda, kapılar en büyük ve daha iyi tanımlanmış popülasyonu seçer. Her hücre bölünmesi boya kombinasyonu için çoklu, yakın popülasyonların varlığı, deneyimlerimize göre, biyolojik farklılıkları temsil etmemektedir. Bunun yerine, a) optimal olmayan bir boyama prosedürünü veya b) hücrelerin başlangıç havuzunda büyük bir heterojenliği (özellikle boyut olarak) gösterebilir. Kordon kanı veya diğer karmaşık biyolojik kaynaklardan (örneğin, kemik iliği aspiratları, periferik kan) başlarken bu beklenmedik bir durum değildir. Kapı sıkı bir şekilde tanımlanmamışsa, farklı boya kombinasyonlarının aşamalı seyreltilmesi, özellikle CV ve VC koşulları için daha sonraki zirvelerin birleştirilmesine yol açabilir (Şekil 2D). Optimal olmayan bir kapının bir diğer olumsuz sonucu, hücre kültüründen sonra farklı zirveleri etkili bir şekilde ayırt edememektir, çünkü heterojen bir başlangıç popülasyonu sığ zirvelere yol açabilir.

Şekil 1: Hücre sıralama için geçit stratejisi. (A) FSC-A'ya karşı SSC-A, enkaz ve kirletici hücreleri dışlamak için. (B) FSC-A'ya karşı FSC-H, çiftleri ve hücre kümelerini hariç tutmak için. (C) Lin+ olan hücreleri hariç tutmak için Lin'e karşı FSC-H. (D) CF, CV, VC ve VI boya kombinasyonları ile boyanmış hücreleri tek sesli olarak tanımlamak için CTV ve CFSE. Kapılar homojen bir popülasyonu içerecek kadar katı olmalıdır. (E) CD34 + CD38 + (HPC'ler olarak da adlandırılır) sınırlı progenitörleri CD34 + CD38 - çoklu potent bölmesinden ayırmak için CD34'e karşı CD38. (F) CD34+CD38- popülasyonundan CD45RA'ya karşı CD90, HSC'de zenginleştirilmiş en olgunlaşmamış progenitörler (CD90+CD45RA-), LMPP (CD90ortaCD45RA+) ve daha kararlı MPP (CD90-CD45RA-) arasında ayrım yapmak için. (G) Hücre boyası kombinasyon boyamaları için burada temsil edilen indeks sıralanmış olaylar ve (H) CD90 ve CD45RA yüzey belirteçlerinin ekspresyonu. Bu şeklin daha büyük bir versiyonunu görmek için lütfen buraya tıklayın.
{kind=link}
Hücre kültürü sonrası akım sitometri analizi
Şekil 2'deki veriler, bir dizi miyeloid progenitör ve öncülü destekleyebilen çoklu sitokinlerin varlığında, 72 saat boyunca kültürde tutulan insan kordon kanı HSC'lerini temsil etmektedir. 2A ila 2D panelleri, bireysel hücrelerin her birinin akrabalığını kurmak için gerekli olan geçidi temsil ederken, 2E ila 2G panelleri hücresel fenotiplemeye izin verir. Şekildeki MEP'lerin varlığının azalması muhtemelen bu temsili deney için kullanılan kültür koşullarının sonucudur (Şekil 2F). Farklı sitokinler ve kültür koşulları kullanmak, deney için farklı başlangıç hücrelerinin seçilmesine benzer şekilde, her bir alt kümenin göreceli yüzdesini değiştirir.

Şekil 2: Akış sitometrisi analizi için geçit stratejisi. (A) FSC-A'ya karşı SSC-A, enkaz ve kirletici hücreleri dışlamak için. (B) FSC-A'ya karşı FSC-H, çiftleri ve hücre kümelerini hariç tutmak için. (C) CTV ve CFSE karşılaştırması, Etiketli kapı, veri çözünürlüğünü etkileyebilecek herhangi bir otomatik floresan olayının hariç tutulmasına izin verir. (D) CTV ve CFSE. Hücre bölünmesi boya seyreltmelerine dayanarak dört popülasyonu sıkı bir şekilde bağlamak son derece önemlidir. (E) CD34'e karşı CD38, kararlı öncüller (CD34-), kısıtlı progenitörler (HPC) (CD34+CD38+) ve olgunlaşmamış progenitörler (CD34+CD38-) arasında ayrım yapmak için. (F) CD45RA'ya karşı CD123, üç tip kısıtlı progenitörleri ayırt etmek için: CMP (CD123 + CD45RA-), MEP (CD123-CD45RA-) ve GMP (CD123 + CD45RA+). (G) HSC'leri, LMPP'leri ve MPP'leri tanımlamak için CD34+CD38-'den CD45RA ve CD90. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}
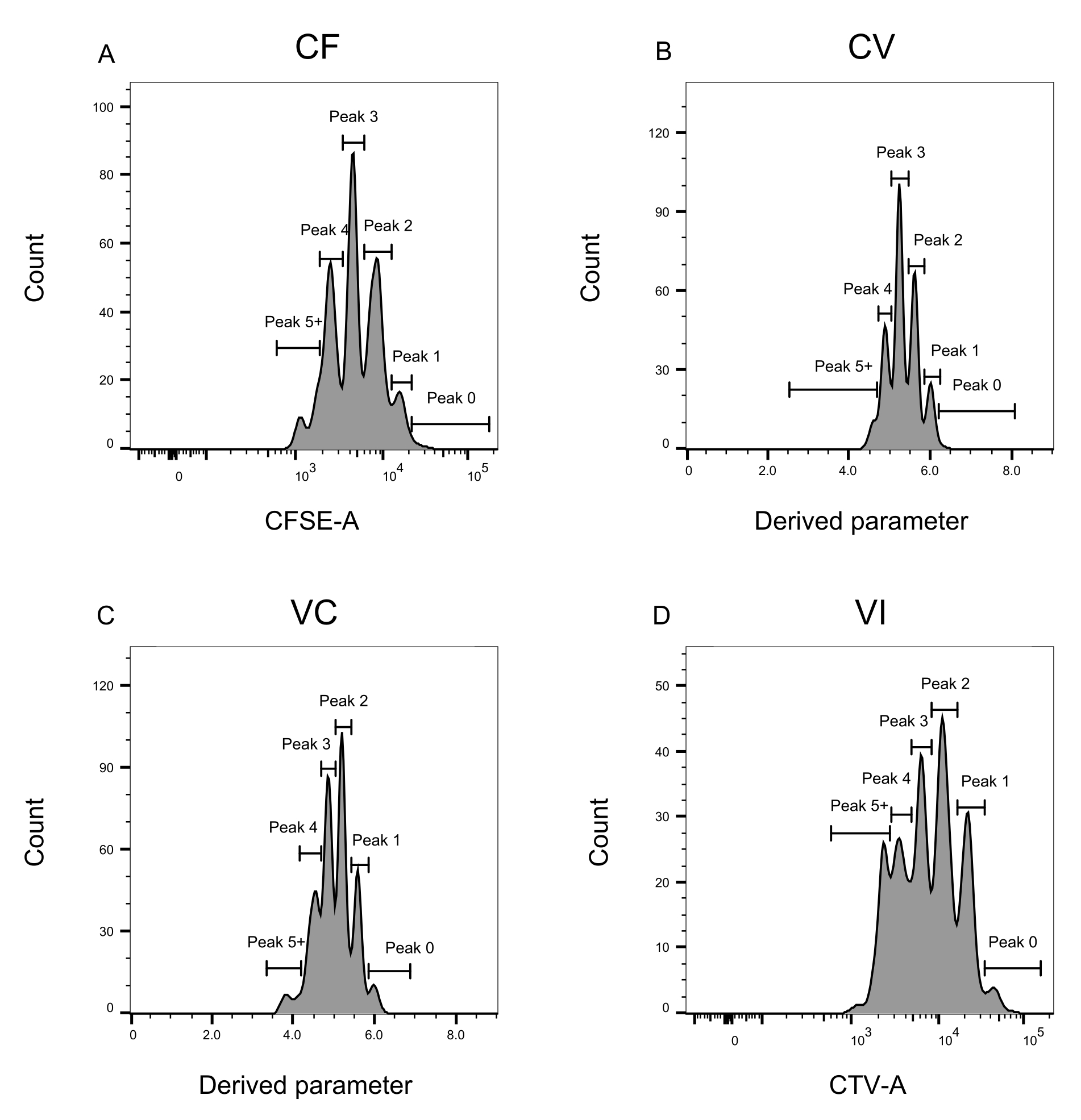
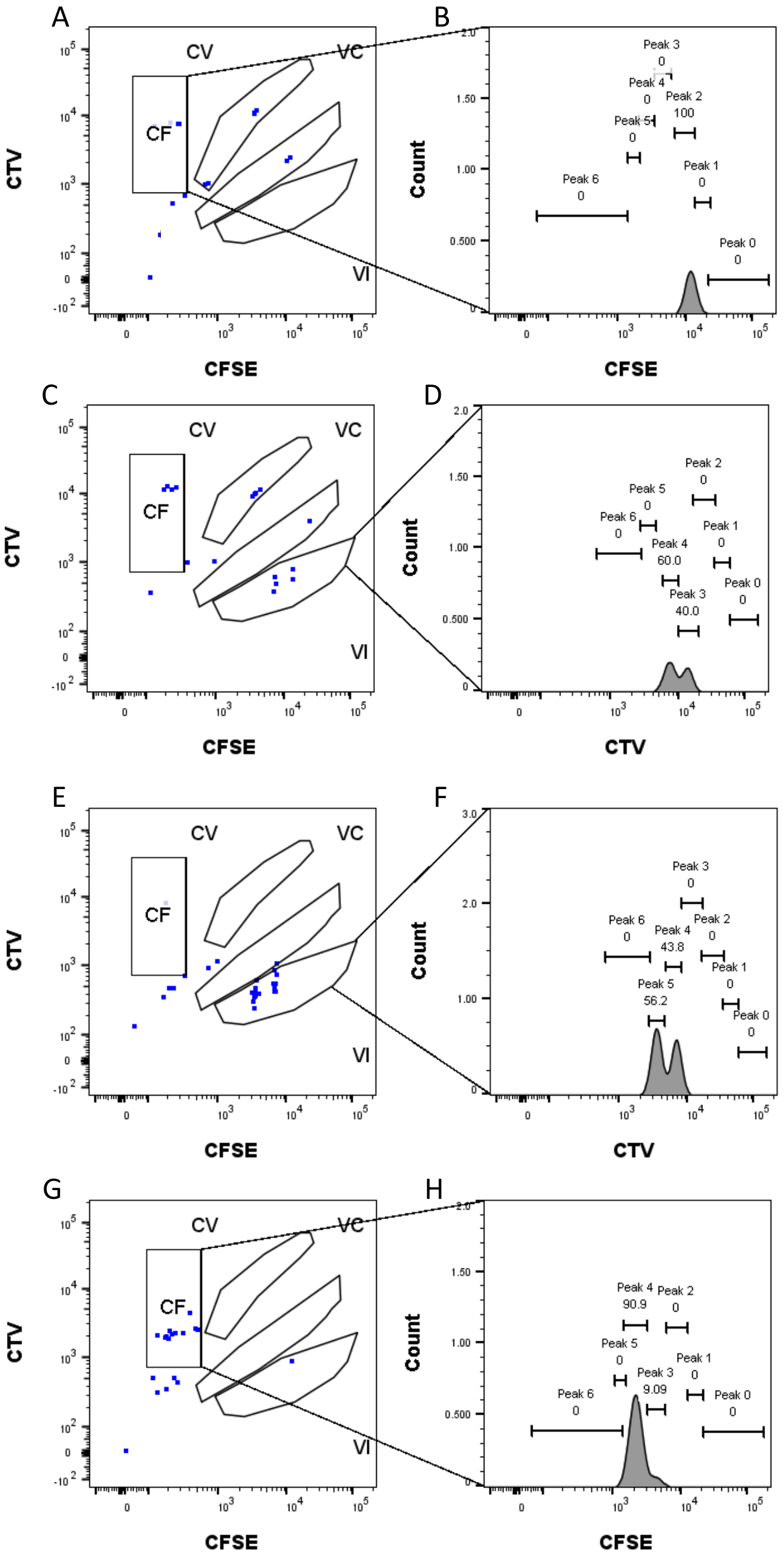
Tepe tanımı ve atama adımları (Şekil 3 ve Şekil 4) protokolün çok önemli yönleridir ve katı kapıların tanımlanmasını gerektirir. Tepe tanımı için (Şekil 3), güvenilir bir tanımlama için en az 1.000 olay gereklidir. Bu anlamda, "Toplu" kuyucuklar için hücre sıralama adımı sırasında daha fazla hücreyi izole etmek faydalı olabilir. Şekil 4'te, birden fazla aileyi içeren dört tek kuyu örneği açıklanmaktadır. Bu şekil, özellikle her ailenin ve her zirvenin tanımlanması için Şekil 2D ve Şekil 3 geçişinin önemini açıklığa kavuşturmaktadır. Şekil 4A, CF kapısındaki tüm hücreler birbirine çok yakın olduğundan ve tek bir tepe noktasına kolayca atanabildiğinden, basit bir örnek göstermektedir. Şekil 4C, Şekil 4D'nin histogramında açıkça gösterildiği gibi, iyi ayrılmış iki tepe noktasına tek sesli olarak dağılmış bir ailenin başka bir örneğini göstermektedir. Şekil 4E, G, çok sayıda olaya dayanan katı geçitlemenin önemini ortaya koymaktadır; Her ikisi de yakın olan, ancak boya kombinasyonu kapılarının dışında olan birkaç olay gösterir. Bu olaylar, yalnızca tek kuyulu analize dayanarak, VI ve CF kapılarına yanlış bir şekilde dahil edilebilir. Son olarak, Şekil 4F,H, iki benzer yoğunluk zirvesine (Şekil 4F) ve iki eşit olmayan yoğunluk zirvesine (Şekil 4H) bir örnek ile birden fazla zirveye yayılmış iki farklı aile örneğini göstermektedir.

Şekil 3: Akış sitometrisi analizi için tepe tanımı. (A-D) Zirveler, her bir pik için iyi bir temsil sağlamak için en az 500 olayı kaydedecek şekilde tanımlanmalıdır. (A) CFSE-A yoğunluğu için histogram. Her biri farklı bir bölünen hücre popülasyonuna karşılık gelen birkaç tepe noktası tanımlanabilir. (B,C) Türetilmiş parametrenin yoğunluğu için histogramlar, sırasıyla CFSE-CTV karışımını, CV (B) ve VC (C) temsil eder. (D) CTV-A yoğunluğu için histogram. Bu şeklin daha büyük bir versiyonunu görmek için lütfen buraya tıklayın.
{kind=link}

Şekil 4: Tepe ataması . (A,B) Bu kuyu için sadece CF kapısında bir tepe tespit edilebilir. (C,D) Bu kuyuda, VI kapısında neredeyse eşit yoğunlukta iki tepe noktası tespit edilebilir. Zirveler iyi çözülmüştür. (E,F) Bu kuyuda, VI kapısında karşılaştırılabilir yoğunlukta iki tepe noktası tespit edilebilir. Toplu kuyular kullanılarak belirlenen stratejiye dayanarak sadece kapıdaki olaylar dikkate alınmıştır. (G-H) Bu kuyuda, CF kapısında eşit olmayan yoğunlukta iki tepe noktası tespit edilebilir. bu rakamın daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}
Veri gösterimi ve istatistiksel testler
Şekil 5, her ikisi de 72 saatlik hücre kültüründen sonra gerçekleştirilen iki ayrı deneyin farklı veri temsili türlerini göstermektedir. HSC'ler ve MPP'ler, hücre bölünmesini ve farklılaşma özelliklerini değiştirmesi beklenen iki farklı hücre kültürü ortamında kültürlenmiştir. Bu ortamlar "Diff" (Farklılaşma)32 ve "GT"33; birincisi, eritropoietin (EPO) ve granülo-monositik koloni uyarıcı faktör (GM-CSF) içerdiğinden miyeloid ve eritroid farklılaşmasını teşvik ederken, ikincisi, HSPC'lerin yüksek bir yüzdesini korumak ve yükseltmek amacıyla gen terapisi klinik çalışmaları bağlamında geliştirilmiştir. Şekil 5A , çeşitli hücre ailelerini temsil eden "Diff" durumu için temsili bir ısı haritasıdır, hem hücre kaderlerinde hem de bölünmelerde. Bu ısı haritasında, her satır ayrı bir aileyi, her kare ayrı bir hücreyi temsil eder ve sütunlar aynı nesildeki tüm hücreleri gruplandırır (örneğin, 2. nesildeki hücreler en az iki kez bölünür). Tek bir hücre tipinden oluşan ve aynı sayıda bölünme gösteren (örneğin, aile #63) ve iki nesil boyunca üç hücre tipi (örneğin, aile # 84) içeren heterojen aileleri ayırt etmek mümkündür. Bu analiz için hücresel iyileşme oranı yaklaşık% 70 olduğundan, tüm hücrelerinin muhtemelen farklı nesillerde (örneğin, 1. nesilde bir hücrenin bir ailesi ve 2. nesilde iki hücrenin bir ailesi) geri kazanılmasıyla tanımlanan tam aileler nadiren gözlenir ( Şekil 5A'da kimlik numaralarının yanında bir hashtag görüntülenir). Teknik (boyama sorunu, protokole bağlı hücre kaybı) veya biyolojik (hücre ölümü ve / veya apoptoz) olabilecek eksik tespiti açıklayan birden fazla açıklama vardır. Teknik sınırlamalar, tek tek numuneyle ilişkili ölü hacmi azaltmak için tasarlanmış bir analizör kullanılarak ve hacim pipetlemesini azaltmak için hücre boyama işlemini doğrudan hücre kültürü plakasında gerçekleştirerek aşılabilir. Tersine, hücre ölümünün miktarını belirlemek için ortogonal yöntemler (örneğin, canlı hücre görüntüleme deneyleri yoluyla), eksik tespitle sonuçlanan teknik ve biyolojik faktörleri ayırt etmeye yardımcı olabilir.
Şekil 5Bi , kültür durumunun hücre tipi bileşimi üzerindeki etkisinin, sanki toplu bir tahlil yapılmış gibi nasıl görselleştirileceğini göstermektedir. Burada, Diff koşulu daha fazla sayıda kaderi ve CD34 + hücrelerinin daha yüksek bir yüzdesini (CD34- dışındaki tüm hücre tipleri olarak tanımlanır) teşvik eder. Güven aralıkları , komut dosyasında temel önyüklemeyle hesaplanır ve 250.000 önyüklemeli veri kümesi34 bulunur. Şekil 5'teki diğer tüm histogramların aynı şekilde hesaplanan güven aralıklarını gösterdiğini belirtmek gerekir. Tablo 5 , her nesildeki aile sayısı ve hücre sayısı hakkındaki tüm bilgileri özetlemektedir.
Şekil 5Bii , "2_bar_plot" senaryosunda gerçekleştirilen istatistiksel testlerin çıktısını grafiksel olarak temsil eder. İstatistiksel çerçevenin resmi bir açıklaması mevcuttur26. Kısacası, bu çerçeve, gözlemlenen tüm hücreler arasında bağımsızlık gerektiren klasik istatistiklerin aksine, aynı aileden hücrelerin bağımlı olduğunu varsayarken istatistiksel hipotez testine olanak tanır (kendisi test edilebilir bir varsayım). Şekilde sunulan özel durumda, istatistiksel test, kültürde bulunan farklı hücre tiplerinin frekansları olarak ölçülen MPP'lerin hücre kaderi seçimlerinin, kullanılan hücre kültürü koşullarından bağımsız olduğu hipotezine meydan okumaktadır. İlk olarak, G-testi istatistiği, farklı hücre ortamlarından hücre tipi frekansları arasındaki tutarsızlığı değerlendirmek için kullanılır (örneğin, Bii'de, bu istatistik kırmızı çubukla temsil edilir). Daha sonra, verilerin randomizasyonu permütasyon yoluyla gerçekleştirilir ve iki hücre kültürü koşulu arasında tüm hücre ailelerini değiştirir. Bu, aileyle ilgili hücreler arasındaki bağımlılığı korurken, her kümedeki aile sayısını orijinal verilerle tutarlı tutmaktır. G-testi istatistiği rastgele veri kümesinden hesaplanır. 5Bii ile temsil edilen mavi değerler, 250.000 permütasyon için G testi istatistiğidir. Son olarak, p değeri, orijinal veri kümesinin permute olanların dağılımından ne ölçüde saptığını değerlendirmek için hesaplanır. Örnekte, orijinal istatistik büyük ölçüde dağılımdan saparak küçük bir p değeri ile sonuçlanır ve böylece MPP'lerin hücre kaderinin kültür koşullarından bağımsız olduğu hipotezini reddeder.
Şekil 5C , farklı koşulların hücre ailesi başına hücre bölünmesini nasıl değiştirdiğini keşfetmek için maksimum nesil başına hücre ailelerinin yüzdesini temsil eder. Bu veri grafiği, 72 saatte, Diff koşulunda kültürlenen hücrelerin, GT koşulundaki hücrelerden daha fazla sayıda bölünmeyi tamamladığını göstermektedir. Temsil edilen, her aile başına maksimum nesil sayısıdır, bu nedenle 1. ve 2. nesillerdeki hücreleri görüntüleyen bir aile, 2. nesil olarak kabul edilir. Şekil 5B için kullanılan aynı istatistiksel çerçeve, hücresel bölünme ve kültür durumu arasındaki bağımsızlığı istatistiksel olarak test etmek için kullanılabilir.
Şekil 5D , farklı ata tipleri (HSC'ler veya MPP'ler) için ilk bölümün simetri / asimetri tipini araştırmaktadır. 1. nesildeki tüm hücre aileleri için - iki yavru hücreyi kardeş hücreler olarak kesin olarak kurmanın mümkün olduğu tek nesil - dört farklı simetri / asimetri türü tanımlanabilir: "Sym Undiff" etiketi, her iki kızın da menşe hücresinin fenotipini koruduğu aileleri tanımlar. "Sym Diff", her iki kızın da aynı fenotipe sahip olduğu ve menşe hücresinden farklı olduğu anlamına gelir. "Asym Undiff", bir kızın sadece menşe hücresinin fenotipini koruduğu anlamına gelir. Son olarak, "Asym Diff", her iki kızın da farklı fenotiplere sahip olduğu ve hiçbirinin menşe hücresiyle aynı olmadığı aileleri tanımlar. Bu simetrik / asimetrik kaderleri değerlendirmede istatistiksel güç kazanmak için, 1. nesilde yavruları bulunan daha fazla aileyi gözlemlemek için MultiGen analizini erken zaman noktalarında yapmak arzu edilir.
Son olarak, Şekil 5E , bölümler arasındaki farklılaşma paterni ilerlemesi hakkında fikir edinmek için bölme sayısının bir fonksiyonu olarak hücre tiplerinin yüzdelerini temsil eder. Örneğin, şekilde görüntülenen veriler, hücrelerin CD34 durumuna ilerlediğini ve yalnızca üç bölümden sonra bu sınıftaki algılanan hücrelerin% 50'sinden fazlasının olduğunu göstermektedir. Dahası, MPP'lerin kendi kendini yenileme bölünmesini desteklemediği sonucuna varmak mümkündür, çünkü hücrelerin küçük bir yüzdesi orijinal fenotipi korur. Bu sonuçlardan bazıları daha sonra önceki şekillerde sunulan istatistiksel çerçeve kullanılarak test edilebilir.

Şekil 5: Kordon kanı HSPC'lerini kullanan 72 saatlik bir deney için veri gösterimi örneği. (A) Seçilen bir veri kümesi için ısı haritaları (HSC, "Diff" ortamında, 72 saatlik kültürden sonra). Grafikler, akrabalıklarına (satırlarına), gerçekleştirilen bölünme sayısına (sütunlar, nesil olarak adlandırılır) ve fenotiplerine (renkler) göre tüm bireysel hücreleri (kareleri) temsil eder. (Bi) HSC'lerin ve MPP'lerin hücre soylarının hücre tiplerinin oranlarını, GT koşulu ile Diff. (Bii) koşulu arasındaki oranlarını karşılaştıran histogram. (Bii) Grafik, "Diff" ve "GT" sitokin kokteylleri arasında karşılaştırma yaparak, 72 saatlik kültürde MPP'ler için "2_bar_plot" senaryosunda gerçekleştirilen istatistiksel testleri temsil eder. Deneysel değer kırmızı renkte , 250.000 permütasyonla üretilen değerler ise mavi renkte görüntülenir. G-testinin p. değeri, test için kullanılan aile sayısı ile birlikte sağ üst köşede belirtilir . (C) Her nesildeki ailelerin yüzdesini (renk kodlu) karşılaştıran histogram, kültür koşulu başına HSC'ler ve MPP'ler için. Güven aralıkları, 250.000 önyüklenmiş veri kümesiyle hesaplanır. (D) 1. nesilde iki hücreli aileler için kızı hücrelerin kaderi arasındaki simetri/asimetri tipini temsil eden histogram: Sym Undiff (her iki kız da menşe hücrenin fenotipini korur), Sym Diff (her iki kız da aynı fenotipe sahiptir ve menşe hücresinden farklıdır), Asym Undiff (sadece bir kızı menşe hücresinin fenotipini korur), ve Asym Diff (her iki kızın da farklı fenotipleri vardır ve hiçbiri köken hücresine benzemez). (E) "Diff" kokteyli ile kültürlenen MPP'ler için nesil başına sınıflandırılan hücre tiplerinin katkısının histogramları; n = 204 hücre ve 97 aile. Bu şeklin daha büyük bir versiyonunu görmek için lütfen buraya tıklayın.
{kind=link}
Tablo 5: Her deneysel koşul başına analiz edilen ailelerin ve hücrelerin sayısının tanımı (orijin hücre ve hücre kültürü ortamı). Bu Tabloyu indirmek için lütfen tıklayınız.
Ek Dosya 1: Bu Dosyayı indirmek için lütfen buraya tıklayın.
Ek Dosya 2: Bu Dosyayı indirmek için lütfen buraya tıklayın.
Ek Dosya 3: Bu Dosyayı indirmek için lütfen buraya tıklayın.
Ek Dosya 4: Bu Dosyayı indirmek için lütfen buraya tıklayın.
Tartışmalar
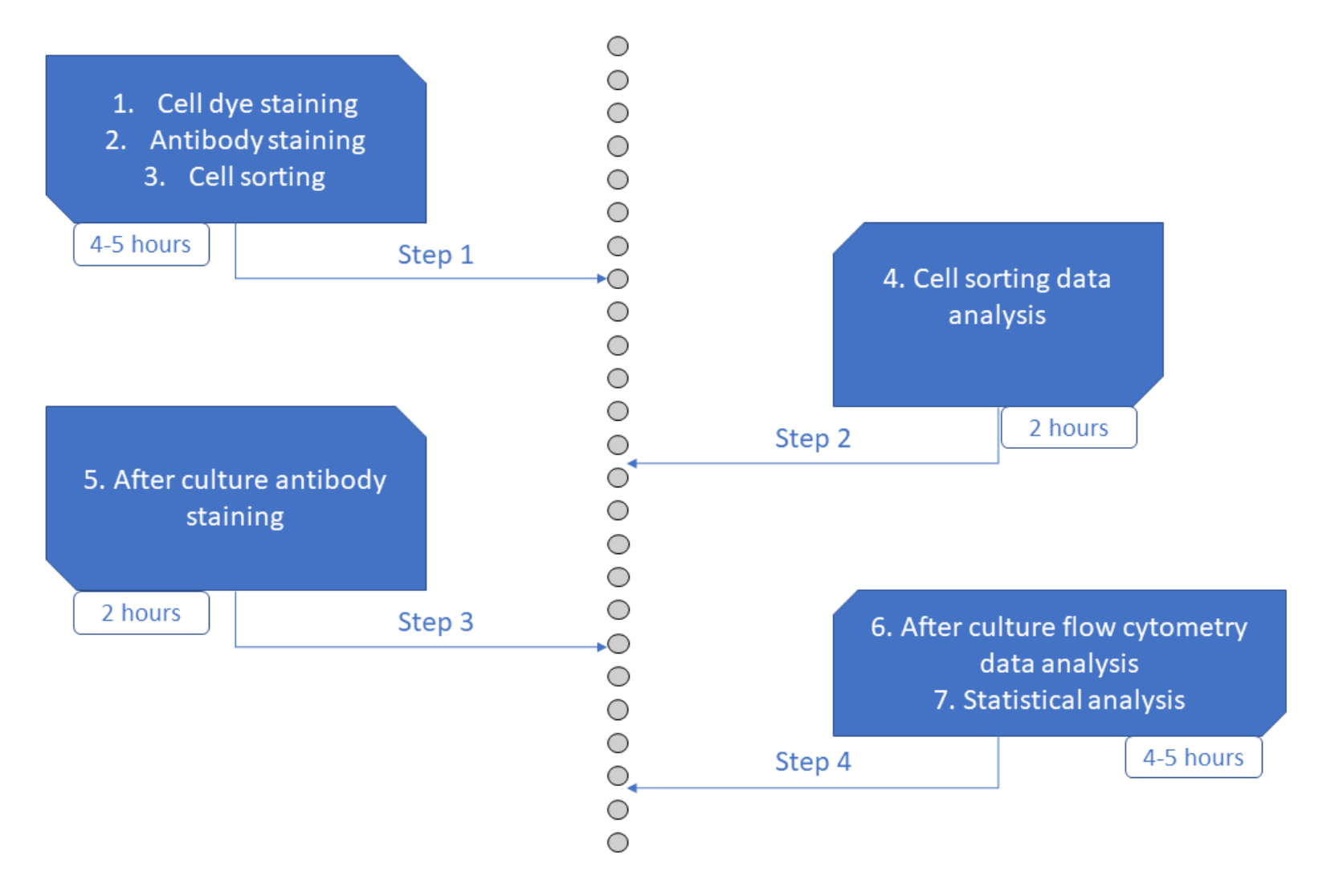
MultiGen testi, lenfosit 23,24,35 ve murin hematopoetik hücreleri26,27'yi incelemek için etkili olan yüksek verimli, gerçekleştirilmesi kolay ve ucuz olmayan bir tahlildir. Burada, kısa süreli kültür kullanarak tek hücreli düzeyde, insan HSPC taahhüdünün erken evresini ex vivo olarak deşifre etmeyi sağlayan yaklaşımın yeni bir gelişimini sunuyoruz (Şekil 6). Tek hücreli ex vivo kültür sistemleri tipik olarak HSPC'lerin olgun hücrelere uzun vadeli kaderini değerlendirmek için kullanılır, ancak bazı kaderler diğerlerinden daha erken ortaya çıkar36, potansiyel olarak analizi daha az kadere doğru önyargılı hale getirir. Ek olarak, bu kültür sistemleri genellikle kader taahhüdü sırasında bölünmeler hakkındaki bilgileri kaçırır. İlk taahhüt adımlarının, kültürün başlangıcında olduğu kadar erken bir zamanda, bazen 26,37'lik bölünme olmadan, kısa vadeli kültürü ve izleme bölünmesini erken kader taahhüdünü incelemek için gerekli kıldığı gösterilmiştir. Kader, bölünme ve akrabalığı eşzamanlı olarak takip ederek, bu tahlil insan HSPC'lerinde ilk bölünme ve kader kararının rolünü anlamayı sağlar. Testi kullanarak, taahhüt sürecinin kaç bölünmeden sonra, bu erken atalar için kendini yenileme ve farklılaşma arasındaki dengeyi ve bu özelliklerin nesiller boyunca nasıl miras alındığını çıkarmak mümkündür. Bildiğimiz kadarıyla, bu, tek hücreli çözünürlükte insan HSPC'leri için bu tür ölçümlere izin veren tek tahlildir. Ek olarak, hücre bölme boyalarının farklı kombinasyonlarını kullanarak, analizin verimini artırdık ve bu tahlili hızlı bir şekilde büyük veri kümeleri oluşturmak için değerli bir araç haline getirdik. Boya kombinasyonları, aynı kuyucuktaki birkaç aileyi takip etmeyi sağlar ve kısa süreli kültürde analiz için mevcut hücre sayısını arttırır. Kombinasyonların sayısı, diğer boyaların (örneğin, sarı boya) eklenmesi veya CFSE ve CTV oranının değiştirilmesi yoluyla potansiyel olarak daha da arttırılabilir. Ancak bu, analiz edilebilecek diğer parametrelerin sayısını azaltır.

Şekil 6: Protokolün şematik gösterimi. Bu şeklin daha büyük bir versiyonunu görmek için lütfen buraya tıklayın.
{kind=link}
Analizi başarılı bir şekilde gerçekleştirmek için, çok sayıda kuyucuk ve analiz edilecek hücre sayısının azalması nedeniyle, akış sitometri analizini bir plaka okuyucu ile donatılmış bir analizörde çalıştırmak gerekir. Yeni nesil tezgah analizörleri özellikle bu tahlile uyarlanmıştır, çünkü çoğu hücre kaybı yüzdesini azaltmak için daha küçük bir ölü hacme sahiptir. Bu da, her bir kuyunun tamamının geri kazanılmasında daha yüksek bir verimliliği garanti eder ve bu da% 70 aralığında tahmin edilen bir verimliliği tetikler26. Akış sitometrisi kazanımı sırasında hücre kaybını tahmin etmek, her bir ailenin analizi için çok önemlidir. Örneğin, hücre ölümü olmadığını varsayarsak ve bölünme sayısını sayarsak, her aile başına hücre sayısını tahmin etmek mümkündür. Bununla birlikte, özellikle test edilen kültür koşullarında hücre ölümünün tahmin edilmesinde ve belirli sayıda hücre kullanılarak deneysel olarak iyileşme oranının ölçülmesinde bazı doğrulayıcı deneylerin yapılması arzu edilir.
Bu protokolün en önemli adımlarından biri zirve atamasıdır. Daha önce de belirtildiği gibi, kaliteli bir tepe dağılımı, hücre sıralamada çok dar piklerin izolasyonuna büyük ölçüde bağlıdır. Bununla birlikte, dağıtıma benzersiz bir şekilde dayanarak doğru sayıda bölüm atamak hala zordur. Hücre sıralama ve akış sitometrisi analizi iki farklı makinede yapıldığından, her sinyalin yoğunluğunu doğrudan karşılaştırmak mümkün değildir, bu nedenle histogramın sağ ucunda gözlenen ilk pikin tepe 0 mı yoksa tepe 1 mi olduğunu bilmek zor olabilir. Bu bağlamda, çok az çözüm mümkündür; Bunun bir yolu, bu hücreler tarafından gerçekleştirilen bölünmelerin sayısını doğru bir şekilde ölçmek için ortogonal bir deney yapmaktır (örneğin, canlı hücre görüntüleme). Diğer bir olasılık, akış sitometri analizini çalıştırmadan önce, ters çevrilmiş bir parlak alan mikroskobu altında kuyudaki hücre sayısını saymaktır. Bu, ortalama bir bölünme sayısı çıkaracaktır (hücre ölümü olmadığı varsayılarak). Son olarak, zirve ataması için post-hoc bir çözüm, alışılmadık sayıda "imkansız ailenin" tespitidir; Bu aileler, nesil başına mümkün olandan daha fazla sayıda hücreden oluşur (örneğin, nesil 2'de beş hücre veya nesil 1'de iki hücre ve nesil 2'de bir hücre). İmkansız aileleri dışlama olasılığı istatistiksel analiz adımında kodlanır ve imkansız aileyi işaretler. Bu hataların ortaya çıkması çok yüksekse, tepe atamasının gözden geçirilmesi gerektiğini varsaymak mantıklıdır.
Bu protokolde, büyük veri kümelerinin oluşturulmasında ve yorumlanmasında önemli bir adım haline geldiğinden, test için birkaç veri gösterimi ve analizi örneği ekledik38. İlk örnek, aile başına organize edilmiş analiz edilen tüm hücrelerin toplamını gösteren ısı haritasıdır. Bu, verilerin genel özelliklerini ve potansiyel sonuçları araştırmak için etkili bir araçtır: aileler birden fazla hücre tipinden mi oluşuyor yoksa bileşimde homojen olma eğiliminde mi? Aileler birden fazla nesle yayılmış mı, yoksa çoğunlukla aynı sayıda bölünüyorlar mı? Bu keşifsel analizin daha sonra daha spesifik grafikler ve istatistiksel testlerle tamamlanması gerekir. Simetrik ve asimetrik kader taahhüdünü, bölünmeden farklılaşmayı, kendini yenileme ve farklılaşma arasındaki dengeyi ve belirli bir taahhüt kaderi için bölünme sayısını nicel olarak değerlendirmek için kullanılabilir. Deneysel planlama sırasında, hücre kültürü uzunluğunu sorulan sorunun türüne göre ayarlamak esastır; Örneğin, ilk iki soru için (simetrik/asimetrik denge ve bölünmeden farklılaşma), çok kısa kültür adımlarının planlanması,26'da sadece bir bölünme gerçekleştiren veya hiç yapmayan çok sayıda ailenin izole edilmesini sağlar. Tersine, daha uzun deneyler, farklılaşmanın farklı aşamalarındaki aileleri örnekledikleri için, belirli bir hücre taahhüdü için gereken bölünme sayısının araştırılmasına izin verir. Bununla birlikte, bu yöntem uzun süreli kültürler (2-3 hafta) için tasarlanmamıştır, çünkü hücre boyası seyreltmesi yedi veya sekizden fazla bölümü doğru bir şekilde izleyememektedir22. Sonuç olarak, bu araç çoğunlukla hematopoetik progenitörlerin erken bağlılığını incelemek için uyarlanmıştır ve bu hücrelerin uzun vadeli farklılaşma özellikleri hakkında sağlam sonuçlar çıkarmak için tasarlanmamıştır.
İstatistiksel çerçeve, özellikle bu tür verilerin analizi için geliştirilmiştir ve permütasyonlarkavramına dayanmaktadır 26. Bu, hücre tipi dağılımına ve yapılan bölünme sayısına ailesel bir bağımlılığın gözlemlenmesi nedeniyle gerekliydi. Başka bir deyişle, aynı ailenin parçası olan hücrelerin de benzer fenotipler gösterme ve aynı sayıda bölünme olasılığı daha yüksektir. Derinlemesine bir analiz bu çalışmanın kapsamı dışında olsa da, farklı koşulları değerlendirirken sağlanan istatistiksel testler seti yeterli olmalıdır.
Sonuç olarak, bu protokol hematopoetik kök ve progenitör hücrelerin hücresel dinamiklerini ex vivo olarak hızlı ve ucuz bir şekilde değerlendirmek için değerli bir araçtır. Zaman noktası, kültür koşulları ve analiz edilen HSPC'lerin türü ile ilgili esnekliği ve çok yönlülüğü nedeniyle, çeşitli deneysel koşulların test edilmesine izin verir. Akış sitometrisine dayalı bir tahlil olarak, çoğu laboratuvarda uygulanabilir ve kapsamlı bir ön bilgi gerektirmez, bu da onu taramalar ve pilot deneyler için iyi bir aday haline getirir.
Açıklamalar
Yazarlar bu çalışma ile ilgili çıkar çatışması olmadığını beyan ederler. Fon verenlerin çalışma tasarımı, veri toplama ve yorumlamada veya çalışmayı yayınlanmak üzere gönderme kararında hiçbir rolü yoktu.
Teşekkürler
Institut Curie Flow Facility üyelerine, akış sitometrisi deneylerinin kurulmasındaki yardımları için teşekkür ederiz. Ayrıca, Perié Takımı'nın diğer üyelerinin birden fazla tartışma sırasındaki katkılarını da kabul etmek istiyoruz. Dr. Julia Marchingo ve Prof. Phil Hodgkin'e (Walter end Eliza Hall Tıbbi Araştırma Enstitüsü) lenfositler üzerindeki hücre bölünmesi boyalarının çoğullanması protokolünü paylaştıkları için teşekkür ederiz. Saint Louis hastanesi kordon kanı biyobankasına bu protokolün geliştirilmesi için gerekli biyolojik kaynakları sağladığı için teşekkür ederiz. Çalışma, CNRS ve Bettencourt-Schueller Vakfı'ndan (L.P.'ye) bir ATIP-Avenir hibesi, Labex CelTisPhyBio (ANR-10-LBX-0038) (L.P. ve A.D.'ye), Idex Paris-Science-Lettres Programı'ndan (ANR-10-IDEX-0001-02 PSL'ye) (L.P.'ye), Canceropole INCA Emergence (2021-1-EMERG-54b-ICR-1, L.P.'ye) ve ITMO MIIC hibesinden (21CM044, L.P.'ye) hibeler ile desteklenmiştir. Avrupa Birliği'nin Horizon 2020 araştırma ve inovasyon programı ERC StG 758170-Microbar (L.P.'ye) kapsamında Avrupa Araştırma Konseyi'nin (ERC) finansmanının yanı sıra A.D., Fondation de France'dan bir burs ile desteklendi.
Malzemeler
| Name | Company | Catalog Number | Comments |
| 1.5 mL polypropylene microcentrifuge tubes | vWR | 87003-294 | |
| 15-mL polypropylene tubes | vWR | 734-0451 | |
| 50-mL polypropylene tubes | vWR | 734-0448 | |
| 96-well U-bottom culture plate | Falcon | 353077 | |
| Anti-human Lin APC | Thermo Fisher | 22-7776-72 | Dilution 1/40 |
| ARIA III | BD | Can be replaced with any FACS sorter able to sort individual cells in 96-wells plate | |
| Carboxyfluorescein succinimidyl ester (CFSE) | Life Technologies | C34570 | |
| Cell Trace Violet (CTV) | Life Technologies | C34571 | |
| Compensation beads | BD | 552843 | |
| Dulbecco’s modified Eagle medium (DMEM) | Life Technologies | 11320033 | |
| Ethylenediaminetetraacetic acid (EDTA) | Thermo Scientific | J62948-36 | Prepare a solution 0.5 M, in sterilised water |
| FC block Fc1.3216 | BD | 564220 | Dilution 1/50 |
| Fetal Bovine Serum (FBS) | Dutscher | S1900-500C | Batch S00CH |
| FlowJo v10.8.1 | BD | ||
| Mouse anti-human CD10 PerCP-5.5, clone HI10a | Biolegend | 312216 | Dilution 1/20 |
| Mouse anti-human CD123 BUV395, clone 7G3 | BD | 564195 | Dilution 1/15 |
| Mouse anti-human CD34 APC-Cy7, clone 581 | Biolegend | 343513 | Dilution 1/40 |
| Mouse anti-human CD38 BV650, clone HB7 | Biolegend | 356620 | Dilution 1/40 |
| Mouse anti-human CD45RA AF700, clone HI100 | BD | 560673 | Dilution 1/20 |
| Mouse anti-human CD45RA PE-Cy7, clone HI100 | BD | 560675 | Dilution 1/20 |
| Mouse anti-human CD90 PE, clone 5E10 | Biolegend | 328110 | Dilution 1/20 |
| Phosphate Buffered Saline (PBS) 1X | Life Technologies | 10010001 | |
| Python | |||
| R | |||
| Sterile 12x75 mm conical polypropylene tubes | Falcon | ||
| ZE5 | Biorad | Can be replaced with any flow cytometry analyzer equipped with a plate reader | |
| Laboratory prepared | |||
| Cell culture media | Depends from the specific experiment. Prepare fresh daily and store at +4 °C until use | ||
| DMEM + 10% FBS | Can be stored in sterile conditions, at +4 °C up to 1 year. To prepare 500 mL, add 50 mL of FBS to 450 mL DMEM | ||
| PBS 1X + EDTA 0.1% | Can be stored in sterile conditions, at room temperature, up to 1 year. To prepare 500 mL, add 3.42 mL of EDTA 0.5 M to 500 mL PBS 1X | ||
| Staining buffer | Can be stored in sterile conditions, at +4 °C up to 1 year. To prepare 500 mL, add 2 mL of EDTA 0.5 M and 1 mL FBS to 500 mL PBS 1X |
Referanslar
- Ginhoux, F., Yalin, A., Dutertre, C. A., Amit, I. Single-cell immunology: Past, present, and future. Immunity. 55 (3), 393-404 (2022).
- Ke, M., Elshenawy, B., Sheldon, H., Arora, A., Buffa, F. M. Single cell RNA-sequencing: A powerful yet still challenging technology to study cellular heterogeneity. Bioessays. 44 (11), 2200084 (2022).
- Regev, A., et al. The human cell atlas. Elife. 6, 27041 (2017).
- Laurenti, E., Göttgens, B. From haematopoietic stem cells to complex differentiation landscapes. Nature. 553 (7689), 418-426 (2018).
- Haas, S., Trumpp, A., Milsom, M. D. Causes and consequences of hematopoietic stem cell heterogeneity. Cell Stem Cell. 22 (5), 627-638 (2018).
- Loughran, S. J., Haas, S., Wilkinson, A. C., Klein, A. M., Brand, M. Lineage commitment of hematopoietic stem cells and progenitors: insights from recent single cell and lineage tracing technologies. Experimental Hematology. 88, 1-6 (2020).
- Perié, L., Duffy, K. R. Retracing the in vivo haematopoietic tree using single-cell methods. FEBS Letters. 590 (22), 4068-4083 (2016).
- Yu, V. W. C., et al. Epigenetic memory underlies cell-autonomous heterogeneous behavior of hematopoietic stem cells. Cell. 167 (5), 1310-1322 (2016).
- Ganuza, M., et al. Lifelong haematopoiesis is established by hundreds of precursors throughout mammalian ontogeny. Nature Cell Biology. 19 (10), 1153-1163 (2017).
- Naik, S. H., Schumacher, T. N., Perié, L. Cellular barcoding: A technical appraisal. Experimental Hematology. 42 (8), 598-608 (2014).
- Quek, L., et al. Genetically distinct leukemic stem cells in human CD34 − acute myeloid leukemia are arrested at a hemopoietic precursor-like stage. The Journal of Experimental Medicine. 213 (8), 1513-1535 (2016).
- Karamitros, D., et al. Single-cell analysis reveals the continuum of human lympho-myeloid progenitor cells. Nature Immunology. 19 (1), 85-97 (2018).
- Boitano, A. E., et al. Aryl hydrocarbon receptor antagonists promote the expansion of human hematopoietic stem cells. Science. 329 (5997), 1345-1348 (2010).
- Delaney, C., et al. Notch-mediated expansion of human cord blood progenitor cells capable of rapid myeloid reconstitution. Nature Medicine. 16 (2), 232-236 (2010).
- Fares, I., et al. Cord blood expansion. Pyrimidoindole derivatives are agonists of human hematopoietic stem cell self-renewal. Science. 345 (6203), 1509-1512 (2014).
- Guo, B., Huang, X., Lee, M. R., Lee, S. A., Broxmeyer, H. E. Antagonism of PPAR-γ 3 signaling expands human hematopoietic stem and progenitor cells by enhancing glycolysis. Nature Medicine. 24 (3), 360-367 (2018).
- Vannini, N., et al. The NAD-booster nicotinamide riboside potently stimulates hematopoiesis through increased mitochondrial clearance. Cell Stem Cell. 24 (3), 405-418 (2019).
- Gupta, R., et al. Nov/CCN3 enhances cord blood engraftment by rapidly recruiting latent human stem cell activity. Cell Stem Cell. 26 (4), 527-541 (2020).
- Horwitz, M. E., et al. Omidubicel vs standard myeloablative umbilical cord blood transplantation: results of a phase 3 randomized study. Blood. 138 (16), 1429-1440 (2021).
- Weinreb, C., Rodriguez-Fraticelli, A., Camargo, F. D., Klein, A. M. Lineage tracing on transcriptional landscapes links state to fate during differentiation. Science. 367 (6479), 3381 (2020).
- Loeffler, D., Schroeder, T. Understanding cell fate control by continuous single-cell quantification. Blood. 133 (13), 1406-1414 (2019).
- Tario, J. D., et al. Optimized staining and proliferation modeling methods for cell division monitoring using cell tracking dyes. Journal of Visualized Experiments. (70), e4287 (2012).
- Marchingo, J. M., et al. T-cell stimuli independently sum to regulate an inherited clonal division fate. Nature Communications. 7, 13540 (2016).
- Horton, M. B., et al. Multiplexed division tracking dyes for proliferation-based clonal lineage tracing. Journal of Immunology. 201 (3), 1097-1103 (2018).
- Lehmann, E. L., Romano, J. P., Casella, G. . Testing statistical hypotheses. , 784 (2005).
- Tak, T., et al. HSPCs display within-family homogeneity in differentiation and proliferation despite population heterogeneity. Elife. 10, 360624 (2021).
- Sommerkamp, P., et al. Mouse multipotent progenitor 5 cells are located at the interphase between hematopoietic stem and progenitor cells. Blood. 137 (23), 3218-3224 (2021).
- Kato, K., Radbruch, A. Isolation and characterization of CD34+ hematopoietic stem cells from human peripheral blood by high-gradient magnetic cell sorting. Cytometry. 14 (4), 384-392 (1993).
- Miltenyi, S., Müller, W., Weichel, W., Radbruch, A. High gradient magnetic cell separation with MACS. Cytometry. 11 (2), 231-238 (1990).
- Doulatov, S., et al. Revised map of the human progenitor hierarchy shows the origin of macrophages and dendritic cells in early lymphoid development. Nature Immunology. 11 (7), 585-593 (2010).
- Goardon, N., et al. Coexistence of LMPP-like and GMP-like leukemia stem cells in acute myeloid leukemia. Cancer Cell. 19 (1), 138-152 (2011).
- Laurenti, E., et al. CDK6 levels regulate quiescence exit in human hematopoietic stem cells. Cell Stem Cell. 16 (3), 302-313 (2015).
- Aiuti, A., et al. Lentiviral hematopoietic stem cell gene therapy in patients with Wiskott-Aldrich syndrome. Science. 341 (6148), 1233151 (2013).
- Davison, A. C., Hinkley, D. V. . Bootstrap Methods and their Application. , (1997).
- Horton, M. B., et al. Lineage tracing reveals B cell antibody class switching is stochastic, cell-autonomous, and tuneable. Immunity. 55 (10), 1843-1855 (2022).
- Notta, F., et al. Distinct routes of lineage development reshape the human blood hierarchy across ontogeny. Science. 351 (6269), 2116 (2016).
- Grinenko, T., et al. Hematopoietic stem cells can differentiate into restricted myeloid progenitors before cell division in mice. Nature Communications. 9 (1), 1898 (2018).
- Saeys, Y., Van Gassen, S., Lambrecht, B. N. Computational flow cytometry: Helping to make sense of high-dimensional immunology data. Nature Reviews Immunology. 16 (7), 449-462 (2016).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır