
Method Article
الجر قوة المجهر لدراسة B تنشيط الخلايا الليمفاوية
In This Article
Summary
هنا نقدم بروتوكول يستخدم لإجراء تجارب المجهر قوة الجر على الخلايا B. نحن نصف إعداد المواد الهلامية البولي أكريلاميد لينة ووظيفية لها، فضلا عن الحصول على البيانات في المجهر وملخص لتحليل البيانات.
Abstract
يتيح التحليل المجهري لقوة الجر قياس القوات التي تنتجها خلية على ركيزة. هذه التقنية يستدل على قياسات قوة الجر من حقل إزاحة لوحظ تجريبياً ينتجه خلية تسحب على ركيزة مرنة. هنا، قمنا بتكييف TFM للتحقيق في البنية المكانية والزمانية لحقل القوة الذي تمارسه خلايا B عند تنشيطه عن طريق مشاركة مستضد مستقبلات الخلية B. يجب أن يكون الحدي هلام، وكثافة حبة، والبروتين وظيفية الأمثل لدراسة الخلايا الصغيرة نسبيا (~ 6 ميكرومتر) التي تتفاعل مع، والاستجابة على وجه التحديد ليغاندس لمستقبلات سطح الخلية.
Introduction
خلايا ب هي الخلايا المنتجة للأجسام المضادة في الجهاز المناعي. لتنشيط الاستجابة المناعية التكيفية، فإنها أول الحصول على المستضد في شكل الأصلي (أي، غير المجهزة) من خلال مستقبلات محددة تسمى مستقبلات الخلية B (BCR)1. تحدث هذه العملية في منطقة الخلايا العقدة الليمفاوية B. حتى لو كان بعض المستضدات يمكن أن تصل إلى الخلية B من خلال السوائل اللمفاوية، فإن معظم المستضدات، وخاصة مع الوزن الجزيئي العالي (> 70 كيلوإدا، وهو الحد الأقصى لحجم القنوات اللمفاوية) يتم تقديمها بالفعل في شكلها الأصلي على سطح خلية تقديم مستضد (APC)، عادة ما تكون ماكرو جيبية فرعية أو خلية متشعبة متجولة، من خلال مستقبلات الليكين أو Fc (غير محددة). الاتصال مع هذه الخلية يؤدي إلى تشكيل المشبك المناعي حيث يمارس BCR القوة على المستضدات المرتبطة APC. ربط مستضد إلى BCR يبدأ إشارات BCR، والتي قد تنشيط آليات توليد القوة. هذه القوى يمكن أن تكون مهمة لتضخيم إشارات BCR، ولكن أيضا ضرورية للخلايا B لاستخراج ومن ثم استيعاب المستضد.
وقد أظهرت الدراسات الحديثة أن BCR هو في الواقع الحساسية2. على سبيل المثال، ركائز أكثر صلابة تثير تعزيز BCR الإشارة3. وعلاوة على ذلك، القوة المتولدة في المشبك المناعي تسحب على BCRs واحدة للتحقيق في تقاربها إلى مستضد وبالتالي ضمان التمييز تقارب4. ولذلك فمن المثير للاهتمام للتحقيق في الاستجابة الميكانيكية لخلايا B لعرض مستضد وتشريح هذا الرد من حيث نوع المستقبلات المتورطة (IgG/IgM)5، جزيئات التصاق (ligaز) أو في الخلايا المعدلة وراثيا و الدوائية (أي، إسكات البروتين المصب من BCR إشارة أو ديناميات cytoskeleton)6.
طريقة بسيطة لمراقبة استجابة خلية لركيزة من صلابة الفسيولوجية، وفي الوقت نفسه، والقوى دراسة تمارس على الركيزة هو الجر قوة المجهر (TFM). TFM يتكون من مراقبة حقل التشرد التي تنتجها الخلية سحب على الركيزة مرنة. في الأصل لوحظ تشوه الجل من خلال التجاعيد من الثني نفسه عن طريق المجهر المرحلة النقيض7، ولكن إدراج ميكرودسيس الفلوراس كعلامات fiducial سمح لقرار أفضل ومنذ ذلك الحين أصبح المعيار8. وقد استخدمت هذه الطريقة للتحقيق في قوة الجر التي تمارسها الخلايا والأنسجة والأنسجة، وحتى organoids جزءا لا يتجزأ من المواد الهلامية. وقد تم تطوير عدة أشكال مختلفة من TFM9 بما في ذلك، تركيبة مع المجهر الفائقة البلورية (أي، STED10 أو SRRF11)،تعديل مؤشر الانكسار من الجل للسماح لTIRF المجهر12، استبدال الخرز بواسطة نانو طبع أنماط13، واستخدام nanopils بدلا من سطحمسطحة 14. ولاطلاع على استعراض كامل لهذه الاختلافات، انظر كولين - يورك وآخرون15.
يصف البروتوكول المعروض هنا إجراءً لقياس القوى التي تمارسها الخلايا B على ركيزة مغلفة بمستضد. يتم تطبيق هذه القوات على اليغاندس (مستضد) من أجل تجميعها واستخراجها في وقت لاحق من الركيزة التي تقدم المستضد. لقد قمنا بتكييف بروتوكول TFM القياسي لمحاكاة صلابة ركائز تقديم المستضد الفسيولوجية ، والحجم والطلاء ذي الصلة للخلايا B. يسمح هذا البروتوكول بدراسة العديد من الخلايا في وقت واحد ويمكن استخدامه بالاقتران مع تقنيات المجهر الفلوري والعلاجات الكيميائية. ومع ذلك، فإنه لا يهدف إلى التحقيق واحد جزيء القوة القياسات، والتي ملاقطبصرية 16، جزيئية17،18، biomembrane قوة تحقيقات19، والقوة الذريةالمجهرية 20 هي أكثر ملاءمة التقنيات. بالمقارنة مع غيرها من أساليب قياس قوة الخلية واحدة (على سبيل المثال، micropipettes21 أو microplates22) TFM يسمح لإعادة بناء خريطة كاملة للقوات التي تمارس في المشبك مع قرار من ~ 300 نانومتر. وهذا مفيد لتحديد أنماط الزقت الزقت في القوى التي تمارس على السطح، وكما الجل متوافق مع التصوير confocal، لربط لهم مع تجنيد بروتينات محددة (على سبيل المثال، cytoskeleton وإشارات البروتينات).
على الرغم من أن TFM 3D ممكن ، إلا أنه غير متوافق مع الصلابة والإعداد الذي استخدمناه. التشوهات في 3D قابلة للتحقيق من قبل غيرها من الاجهزة أكثر تعقيدا مثل المجهر قوة نتوء (AFM مسح غشاء تشوه حيث مطلي الخلايا)23,24 ومرنة المجهر الإجهاد تداخل رناناتورس (ERISM, هلام بمثابة تجويف صدى للضوء وتسليط الضوء على التشوهات من الركيزة مع دقة من عدد قليل من نانومتر)25. وعلى الرغم من أن هذه التقنيات واعدة جدا، إلا أنها لم تستخدم بعد في الخلايا باء. ويمكن استخدام أنواع أخرى من TFM، مثل على14نانوبيلاروس، للحصول على مزيد من ركائز قابلة للاستنساخ. ومع ذلك، لا يتم تكييف هذه الهندسة إلى الخلايا الناعمة كما interpenetrates الخلية الركائز، مما يعقد التحليل. وقد استخدم هذا النهج بالفعل في الخلايا التائية لمراقبة قدرة الخلية على بناء هياكل حول الأعمدة26.
على الرغم من بساطته، TFM باستخدام المواد الهلامية polyacrylamide يسمح للمراقبة في وقت واحد من العديد من الخلايا ويمكن بسهولة وبتكلفة غير مكلفة تنفيذها في أي مختبر مجهزة مقاعد البدلاء والمجهر epifluorescence (على الرغم من أننا نوصي confocal / الغزل القرص).
لمحاكاة صلابة الفسيولوجية من APC، استخدمنا المواد الهلامية polyacrylamide مع صلابة من ~ 500 السلطة الفلسطينية27 ووظيفية الجل مع مولدات تفعيل. في هذا البروتوكول ، قمنا بوظيل سطح هلام البولي أكريلاميد مع lysozyme بيضة الحنكة (HEL). وهذا يسمح لقياس القوى التي تولدها تحفيز BCR من خلال الاشتباك من موقع الربط المستضد. استخدام هذا المستضد والخلايا B HEL محددة من الفئران MD4 يضمن موحدة نسبيا توليد القوة ردا على ربط مستضد28. ومع ذلك، يمكن تطعيم جزيئات أخرى (مثل مضادة IgM ماوس B6) على الجل، ولكن القوى المتولدة في هذه الحالات يمكن أن تكون أكثر التجانس وأقل كثافة. لأن الخلايا B هي خلايا صغيرة (قطرها ~ 6 ميكرومتر)، وقد تم تحسين عدد الخرز لتكون القصوى ولكن لا تزال قابلة للتتبع. بالنسبة للخلايا الكبيرة التي تبذل قوى كيلو باسكال على ركائزها ، يمكن للمرء تحقيق نتائج مرضية باستخدام الخرز المتفرق نسبيًا أو تنفيذ عملية تجميل صور الجسيمات البسيطة (PIV) لإعادة بناء حقل التشوه. ومع ذلك، بالنسبة للخلايا الصغيرة مثل الخلايا الليمفاوية B التي تمارس الإجهاد صغيرة مثل ~ 50 باسكال، مطلوب استخدام تتبع الجسيمات واحدة (قياس تتبع الجسيمات، PTV) لتحقيق الدقة المطلوبة عند إعادة بناء مجال التشوه. من أجل تتبع موثوق به الخرز بشكل فردي ، والتكبير من عدسة الهدف يجب أن يكون على الأقل 60x والفتحة العددية لها حول 1.3. وهكذا، يجب أن تكون المواد الهلامية رقيقة نسبيا (<50 ميكرومتر)، وإلا فإن الخرز غير مرئية لأنها فوق مسافة العمل للهدف.
ويتألف البروتوكول الرئيسي من ثلاثة أقسام: إعداد الهلام، ووظيفية الهلام والتصوير؛ واللبابية واللائق؛ واللبابية؛ واللائق قسمين أكثر اختيارية ومكرسة لاستخراج المستضد الكمي والتصوير للخلايا الفلورية.
Protocol
1. هلام التحضير
- سيلاني من دعم هلام
- قم بتنشيط طبق الأغطية أو طبق بيتري الزجاجي السفلي (والذي سيتم استخدامه كدعم هلامي) مع مصباح الأشعة فوق البنفسجية لمدة 2 دقيقة (انتظر 30 s قبل التعرض لمصباح الأشعة فوق البنفسجية لتجنب التعرض للأوزون المتبقي).
- Silanize طبق الأغطية / الزجاج القاع باستخدام 200 ميكرولتر أمينوبريلتري ميثوكسيلين (APTMS) لمدة 5 دقائق. هذا سوف يعد الدعم للربط التساهمي للجل.
- غسل جيدا الأغطية / طبق الزجاج القاع مع المياه النقية جدا.
- جفف الطبق الأغطية/ الزجاجي السفلي باستخدام التطلع الفراغي.
- إعداد غطاء 18mm المستخدمة لتسطيح هلام
- لإعداد الأغطية، وضعت أولا لهم في حامل الأغطية السيراميك. ثم وضع حامل غطاء في الكوابر الصغيرة (50 مل) وصب سيليكونيسير الكاشف (المخزنة في 4 درجة مئوية، قابلة لإعادة الاستخدام) على الأغطية، ويجري التأكد من تغطيتها تماما.
- تغطية كوب مع رقائق الألومنيوم واحتضان لمدة 3 دقائق في درجة حرارة الغرفة. أثناء الانتظار، قم بتعبئة قاريء كبيرة (500 مل) بماء نقي للغاية. بعد 3 دقائق من الحضانة في الكواشف السيليكون، نقل حامل غطاء مع الأغطية إلى منقار الماء.
- شطف جيدا الأغطية مع المياه النقية جدا، وتجفيفها جيدا والحفاظ على مناديل الورق. للحصول على أفضل النتائج، انتقل مباشرةً إلى القسم التالي.
- جل البلمرة
- بالنسبة للجل من 0.5 كيلو باسكال، اخلط 75 ميكرولتر من 40٪ أكريلاميد مع 30 ميكرولتر من 2٪ بيسكريلاميد (crosslinker) و 895 ميكرولتر من محلول ملحي الفوسفات (PBS). يمكن تخزين هذا الـ premix لمدة تصل إلى شهر واحد عند 4 درجة مئوية.
- إلى 167 ميكرولتر من 0.5 كيلو باسكال هلام premix، إضافة 1٪ (1.67 ميكرولتر) من الخرز، دوامة و sonicate لمدة 5 دقائق في 2000000 100 100 100 100 100 100 10000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000 الحفاظ على المزيج المحمية من الضوء باستخدام رقائق الألومنيوم.
ملاحظة: لا يتم بالشمش premix حتى تتم إضافة البادئ (TEMED). - لتحفيز البلمرة، أضف 1٪ (1.67 ميكرولتر) من 10٪ ث / v persulfate الأمونيوم (APS).
- لبدء البلمرة، إضافة 0.1٪ (0.2 ميكرولتر) N،N′N′، N′Tetramethylethylenediamine (TEMED). اخلطي مع ماصة. مرة واحدة وقد تم إضافة وكالة الأنباء الجزائرية وTEMED، هلام بسرعة البلمرة حتى المضي قدما بسرعة إلى صب هلام.
- جل الصب
- Pipet 9 ميكرولتر من مزيج جل على كل طبق coverlip / الزجاج القاع (قطرة في المركز، الشكل 1A)
- ضع غطاء silanized / hydrophobic وتسوية الجل (الشكل 1B). باستخدام ملقط، اضغط على غطاء لضمان ينتشر هلام عبر كامل مساحة الغطاء(الشكل 1C)حتى يبدأ تسرب.
- عكس الأغطية / طبق القاع الزجاجي في طبق بيتري كبير والاستفادة منه على مقاعد البدلاء لإجبار الخرز تسير نحو سطح هلام(الشكل 1D).
- يُغطّى مع رقائق الألومنيوم ويُترك لمدة ساعة واحدة ليُبلمر في درجة حرارة الغرفة في غرفة رطبة (أي، ضع نسيجًا رطبًا فوق الطبق لمنع التبخر).
- بعد 1 ساعة، إضافة برنامج تلفزيوني إلى العينة لتسهيل الافراج عن الأغطية. بعناية، وإزالة غطاء باستخدام إبرة (طلاء مع سيلانيس مختلفة ينبغي أن تسمح سهلة تقشير الخروج من غطاء من هلام، الشكل 1E).
- اترك الجل في برنامج تلفزيوني.
ملاحظة: يمكن الآن تخزين المواد الهلامية في برنامج تلفزيوني عند 4 درجة مئوية لمدة 5-7 أيام، ولكن من المستحسن استخدامها في غضون 48 ساعة.
2. جل وظيفية
- إعداد sulfosuccinimidyl 6-(4'-azido-2'-nitrophenylamino) hexanoate (Sulfo SANPAH) حل في 0.5 ملغ / مل في 10 mM HEPES العازلة. ويمكن تخزين هذا في 4 درجة مئوية مغطاة رقائق الألومنيوم لمدة تصل إلى أسبوع واحد.
- استلهم برنامج تلفزيوني من المواد الهلامية.
- إضافة 150 μL من السولفو سانباه إلى هلام في درجة حرارة الغرفة (الشكل 1F).
- يعرض الجل لعلاج الأشعة فوق البنفسجية لمدة 2 دقيقة لphotoactivate مواقع SANPAH Sulfo وجعلها التمسك سطح هلام.
- غسل مع برنامج تلفزيوني ثلاث مرات (الشكل 1G).
- كرر الخطوات 2.2-2.5.
- إضافة 250 ميكرولتر من HEL (100 ميكروغرام / مل) إلى كل هلام واحتضان بين عشية وضحاها في غرفة رطبة في 4 درجة مئوية بين عشية وضحاها مع الحفاظ على تغطية مع رقائق الألومنيوم(الشكل 1H).
- إزالة مستضد HEL وغسل مع برنامج تلفزيوني ثلاث مرات.
ملاحظة: يعمل HEL على حد سواء كمستضد وكجزيء التصاق. ويمكن استبداله بجزيئات أخرى ترتبط المستقبلات (على سبيل المثال، IgM المضادة للماوس، البوم الدم البقري، أوفالبومين) أو مختلطة مع يغاندات integrin (على سبيل المثال، ICAM1 ملزمة لLFA1). إذا لزم الأمر، يمكن ملاحظة استخراج المستضد مع نسخة الفلورسنت من HEL (التي تم الحصول عليها عن طريق تلطيخ الجزيء مع مجموعة من البروتين وضع العلامات، انظر الخطوة 4). لاحظ أن التركيز في الجزء الأكبر قد لا ينتج نفس تركيز السطح على الجل كما هو الحال على الزجاج: وهذا يحتاج إلى أن يكون كميا مع تلطيخ الثانوية إذا كان هناك حاجة إلى المقارنة المباشرة.
3. تحميل الخلايا والتصوير
- قبل التصوير، وإزالة برنامج تلفزيوني من المواد الهلامية وإضافة 500 ميكرولتر من وسائل الإعلام خلية B (دورة في الدقيقة 1640، 10٪ decomented مصل العجل الجنين، 1٪ البنسلين-الستربتوميسين، 2٪ بيروفات الصوديوم، 50uM ميركاتوبوتاينول و1X غير أمينو الأحماض الأساسية أمينو) والسماح لهم لتكعيب إلى RT.
- إعداد الخلية
- تنقية الخلايا B الأولية من الطحال وفقا لبروتوكول تحديد سلبي (انظر جدول المواد). نموذجي النهائي الخلية العائد هو حوالي 1 × 107 الخلايا. التركيز على هذا إلى 3 × 106 خلايا / مل في الخلية B المتوسطة (RPMI-1640 تستكمل مع مصل عجل الجنين 10٪ ، 1 ٪ البنسلين - ستريبتوميسين ، 0.1 ٪ mercaptoethanol و 2 ٪ بيروفات الصوديوم).
- تخزين الخلايا حسب الحاجة حتى 6 ساعات عند 4 درجات مئوية.
- إبقاء الخلايا عند 37 درجة مئوية لمدة 30 دقيقة قبل الحصول على الصورة.
- التصوير
- استخدام المجهر confocal مع الحرارية و (ربما) CO2 السيطرة.
ملاحظة: بغض النظر عن ما إذا كان يتم استخدام مجهر القرص confocal أو الغزل، من المهم استخدام هدف/ الثقب الذي يسمح لحجم بكسل < 200 نانومتر لتتبع بشكل مريح الخرز في مرحلة التحليل (على سبيل المثال، 60x، NA 1.3). يمكن أيضًا استخدام المجهر الضوئي Epifluorescence ، ومع ذلك فإنه يوفر إشارة أقل إلى نسبة الضوضاء وقد يجعل تتبع حبة الفردية أكثر صعوبة. - سوف تظهر طبقتين رئيسيتين من الخرز في الجزء السفلي والجزء العلوي من الجل. ركز على مستوى الجل
ملاحظة: سوف يظهر هلام لطيف كسماء مرصع بالنجوم، مع حبات موزعة بشكل موحد تقريبا على نفس الطائرة. - برنامج اقتناء لمدة 30 دقيقة مع معدل الإطار من 5 s (وهذا هو قابل للتكيف مع احتياجات التجربة، على سبيل المثال، الحصول على الألوان الأخرى، والحصول على z المكدس، الخ.)
- التعرق وسائل الإعلام من هلام، وترك حوالي 200 ميكرولتر من وسائل الإعلام على هلام. ضع الجل على المجهر والعثور على الطبقة السطحية من الخرز ومنطقة لطيفة حتى على الجل.
- إضافة 80 ميكرولتر من الخلايا (تجنب لمس هلام للحفاظ على التركيز).
- تأكد من أن التركيز لا يزال صحيحاً وأنه يمكن رؤية الخلايا تنازلية في المنطقة (تحت الضوء المنقول). إطلاق اقتناء قبل الخلايا تصل إلى هلام.
- في حالة ملامسة عرضي مع هلام، والاهتزازات، أو الانجراف التركيز، وضبط التركيز.
ملاحظة: من المهم جمع صورة من هلام استرخاء وهذا يمكن أن يكون أي صورة اتخذت قبل وصول الخلايا على هلام.
- استخدام المجهر confocal مع الحرارية و (ربما) CO2 السيطرة.
4. تجربة استخراج الفلوري هيل
- إعداد الفلورسنت HEL عن طريق ربط صبغة الفلورسنت (من لون مختلف عن الخرز واحد مثل اليكسا 555)، انظر جدول المواد.
- في الخطوة 2.7، استبدل HEL التقليدية مع HEL الفلورية.
- الحصول على الصور مع إعدادات الإضاءة منخفضة أو انخفاض معدل الإطار (على سبيل المثال، 2 الإطارات في الدقيقة الواحدة) لتجنب تبييض الصور.
- لتحديد كمية استخراج HEL، حساب كثافة متكاملة على مساحة الخلية لكل إطار I(t) تصحيح وتطبيعها من قبل كثافة I(0)من الإطار 0 وفقا للصيغة:

ملاحظة: المضادات المقترنة مع الفلوروفور غير مرئية (ربما بسبب إخماد الفلوروهور على سطح الجل)، ولكن يمكن التحقق من وجودها على الجل مع المضادة HEL والأجسام المضادة الثانوية الفلورية. ويمكن التحقق من أن الفلوروفور هو في الواقع الفلورسنت عندما فصلت عن طريق تجريده من هلام مع غطاء المغلفة مع المضادة HEL والكشف عنه مع الأجسام المضادة الفلورسنت الثانوية (على غطاء)6. إشارة المستضد المستخرجة قاتمة جداً وأحياناً تكون ملثمة بتسريب الخرز. إذا كان أحد مهتم فقط في استخراج المستضد، فمن المستحسن لإعداد هلام دون الخرز (تخطي الخطوات 1.3.2 و 1.4.3).
5. التصوير الفلوري
- الحصول على الخلايا الفلورية B عن طريق تنقية الخلية B من طحال الفئران المعدلة وراثيا كما فعلت لنوع البرية (على سبيل المثال، من Lifeact-GFP أو Myosin الثاني فئران GFP).
- لتصوير الخلايا الفلورية، استخدم (إن أمكن) مجهر قرص دوار مع غمر مائي لمسافات طويلة 40x-100x الهدف.
- الحفاظ على مدة التعرض ومعدل الإطار منخفضة لتجنب التبييض.
ملاحظة: وظيفة انتشار النقطة في Z متدهورة للغاية بسبب وجود الجل ، وبالتالي نقترح استخدام هدف غمر الماء. المجهرية المصطبة مع أهداف غمس الماء تعاني من انحرافات كروية قوية ناجمة عن وجود الخلية (الكروية) (ونواة الخلية) في مسار الانبعاثات.
6- التحليل
ملاحظة: يتم إجراء تحليل البيانات بشكل عام عن طريق تصحيح أول المكدس كله للانجراف، وإيجاد الخرز في كل إطار، وتتبع تحركاتها فيما يتعلق الإطار المرجعي (التي اتخذت في غياب الخلايا)، واستيفاء حقل التشرد وعكس المشكلة للحصول على الضغط باستخدام تحويل فورييه29. وتحقيقا لهذه الغاية، نقترح استخدام مزيج من برامج ImageJ ماكرو و MATLAB للتحميل من مستودع على الانترنت30.
- فتح الفيلم في ImageJ ككومة من الصور
- تشغيل الماكرو "Crop_and_save.ijm"
- حدد المناطق ذات الاهتمام (ROI) باستخدام أداة "المستطيل" وإضافتها إلى قائمة العائد على الاستثمار باستخدام مفتاح "t".
- عند اقتصاص الخلية، تأكد من تضمين منطقة لا تقل عن 5-10 بكسل من الخرز غير المُجَلّل. استبعاد الخلايا القريبة جدًا من الحدود أو إلى خلايا أخرى من التحليل. عند الانتهاء انقر على 'موافق'.
- الماكرو يقترح قناع الخلية: إذا كان هذا هو ما يرضي انقر على "موافق". إن لم تكن مرضية، انقر على "غير موافق" ثم حدد يدويا منطقة مغلقة مع أي أداة اختيار (على سبيل المثال"حر" أو "البيضاوي") وانقر على "متابعة".
- فتح MATLAB وتشغيل "TFM_v1.m".
- إدخال المعلمات المطلوبة: على وجه الخصوص تحقق من خصائص الصورة (حجم البكسل، الفاصل الزمني للاستحواذ) وخصائص الجل (معامل الشباب E، نسبة بواسون).
- تم تعيين الصورة المرجعية لتكون الأولى بشكل افتراضي. قم بتعيينه إلى إطار آخر إذا لزم الأمر أو تعيينه إلى "0" لتحميل ملف خارجي.
- حدد موقع مخرجات البرنامج في نفس الدليل مثل الملف الأصلي (للحصول على وصف راجع ملف User_notice.pdf). وهذا يشمل مسارا أوليا من الخرز ("FILENAME.fig")، مؤامرة من الطاقة المتعاقدة على مر الزمن ("FILENAME_energy.فيغ")، وجدول من عدة كميات متكاملة على الخلية (الطاقة، المنطقة، لحظات، الخ) "FILENAME_finaltable.mat"، هيكل يحتوي على حقل النزوح والقوة، وأفلام من الخرزة، حقل النزوح، والإجهاد والطاقة (التي يمكن فتحها مع أي قارئ افي).
ملاحظة: في معلمات الإدخال " حجم النافذة " هو الإطار الذي displacementated الاحتياج ومن ثم الحل النهائي من حقل التحمّل والتشريد. يتم تعيين هذا إلى عدد قليل (افتراضياً أربعة) بكسل. ليس من المستحسن للحد من هذا لأنه سيزيد بشكل مصطنع القرار من قبل الاستيفاء المناطق التي لا توجد فيها الخرز.
النتائج
ونظرا لحجم الخلايا، فإن الخوارزميات التي تستخرج خريطة إزاحة الخرز عن طريق تقنيات مترابطة (مثل قياس حجم صورة الجسيمات) ليست دقيقة جدا بشكل عام. ولكن اعتمادا على درجة القرار المطلوبة، يمكن للمرء بسهولة الحصول على النتائج النوعية باستخدام مجاني فيجي / ImageJ المساعد31،32. في حين أن هذا النهج يكفي لمقارنة تحفيز مقابل الظروف غير المحفزة، لتحليل شامل نوصي باستخدام برنامجنا للتحميل من مستودع على الانترنت30، التي تتعقب الخرز بشكل فردي ويوفر خريطة الحقل النزوح في وقت معين كما الاستيفاء الفردية الخرز التشريد33. العديد من القياسات الكمية ممكنة في هذه المرحلة. على سبيل المثال (بافتراض أن النزوح هو سبب فقط عن طريق الإجهاد عرضية لسطح هلام) كما يوفر البرنامج الإجهاد في كل نقطة مما تسبب في أن خريطة التشريد محددة. هذا هو نوع من "مشكلة الانقلاب": النزوح في نقطة معينة يعتمد على مجموع جميع القوى المطبقة في جميع أنحاء النقاط الأخرى. "خوارزمية الانقلاب" تأخذ في الاعتبار المعلمات المادية للركيزة: جمودها (معامل الشباب) ونسبة بواسون. الخوارزميات المباشرة عادة ما تكون دقيقة جدا ولكن مكلفة حسابيا. خوارزميات على أساس فورييه تحويل ، مثل لنا ، وأداء أساسا deconvolution في الفضاء فورييه وأكثر كفاءة ولكنها عرضة لبعض الأخطاء (ويرجع ذلك أساسا إلى خطوة الاستيفاء). هذه الخوارزميات تتطلب عموما ضبط المعلمة التي تمنع الصغيرة المحلية (وربما artifactual) التشريد لتصبح ذات الصلة جدا في حساب حقل الإجهاد (Tikhonov الانتظام المعلمة 8 ،29 ؛ 29؛ 10000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000 متغير "الانتظام" في إطار الحوار؛ هنا نحن عادة تعيين يساوي 5 × 10-19). وللمزيد من التفسير والتحليل المتقدمين، مثل العلاقات المتبادلة بين الزواك والحركات المحلية والارتباطات مع القنوات الفلورية، نوصي بالتعاون مع الخبراء في هذا المجال. لمراجعة الطرق الحسابية انظر شوارز وآخرون9.
كما ذكر أعلاه ، تبدو صور حبة الصحيح مثل "السماء المرصعة بالنجوم" ، توزيع موحد وعشوائي للبقع المضيئة(الشكل 2A). البيانات والتحليل لا يمكن الاعتماد عليها عندما يكون عدد الخرز منخفض جداً (الشكل 2B) أو الصورة خارج التركيز (الشكل 2C). مرة واحدة وقد استقرت الخلايا B على سطح الجل، الخرز تحت الخلايا تبدأ في التحرك بسبب قوة الجر التي تمارسها الخلية على هلام. يجب تجاهل الإطارات التي لا يمكن تتبعها.
كفحص، فمن الممكن أن نلاحظ عن طريق العين حركة الخرز مقارنة "الإطار المرجعي"، وعادة ما يسبق أول اتصال من الخلية مع الركيزة. ويمكن الحصول على النتائج التقريبية من تتبع الجسيمات المفردة (مثل Trackmate، فيجي 34)كما هو الحال في الشكل 3A. يوفر التحليل تجزئة الخرز في الصورة المرجعية ("FILENAME.fig") كـ عنصر تحكم.
مع البرنامج نقترح، يمكن للمرء الحصول على النزوح (الشكل 3B) والضغط الميداني (متجه من الإجهاد المحلي في كل بكسل وكل نقطة زمنية الحصول عليها من قبل عكس من حقل التشرد، الشكل 3C). يوفر المنتج العددي من حقول النزوح والقوة المتكاملة على مساحة الخلية مجموع العمل الذي تمارسه الخلية على الركيزة (الشكل 4A). يتطلب هذا الحساب قناع الخلية التي تم تقديمها في الخطوة 6.2 من البروتوكول.
لمقارنة شرطين البيولوجية (كما تفعيل HEL مقابل غير تفعيل الركيزة BSA، أو نوع البرية مقابل خروج المغلوب) فمن المفيد لحساب منحنى متوسط(الشكل 4B)أو، حتى أكثر تركيبيا، متوسط قيمة خلال آخر نقطة الوقت (20 دقيقة) حيث تصل الطاقة إلى هضبة(الشكل 4C). عندما المعلومات المكانية للقوات ذات الصلة فمن الممكن لمقارنة نقطة وقت واحد من كل شرط(الشكل 4D). راجع كوماري وآخرون6 لإجراء تحليل أعمق.
مثال على الفلوريسين استخراج الاجناين الفاصل الزمني مبين في الشكل 5A: المظهر التدريجي لإشارات الفلوريسين في المفرزة المُستضدية المُشار إليها من الجل. يظهر متوسط منحنى الاستخراج مع فاصل الثقة (خطأ قياسي من الوسط) أكثر من 15 خلية في الشكل 5B.

الشكل 1: عرض تخطيطي لإعداد الجل ووظيفيته. يتم وصف الخطوات في البروتوكول. الرجاء النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}

الشكل 2: ثلاثة أمثلة من صور حبة من الصفات المختلفة. (A) مثال على صورة حبة مع إشارة الصحيح إلى نسبة الضوضاء والكثافة الصحيحة. (B) أمثلة من الصور مع عدد غير كاف جدا من الخرز و (C) من الطائرة التركيز. الرجاء النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}

الشكل 3: معالجة الصور لاستخراج حقل القوة. (أ) مثال على صورة الخرز (مخطط للخرز باللون الأبيض، مستخرج من صورة الإرسال)، تتبع الخرز في الوقت t = 5 دقائق (تراكب أحمر) ونزوح (الأسهم) نسبة إلى الوقت t = 0 دقيقة (شريط مقياس 5 ميكرومتر). (B) حقل النزوح المستدَف (الممثَّل كرعبة متجه وخريطة حجم، تكون الأسهم متناسبة مع النزوح [nm]؛ انظر شريط الألوان على اليمين)؛ أسفل: صورة أكثر سلاسة من حجم (التي تم الحصول عليها من قبل الاستيفاء مع وظيفة bicubic). (C)حقل الإجهاد من حقل النزوح في اللوحة B (ممثلة كرعبة متجه وخريطة حجم; الأسهم تتناسب مع الإجهاد القص [Pa]; انظر شريط الألوان على اليمين); أسفل: صورة أكثر سلاسة من حجم (التي تم الحصول عليها من قبل الاستيفاء مع وظيفة bicubic). الرجاء النقر هنا لعرض نسخة أكبر من هذا الرقم.
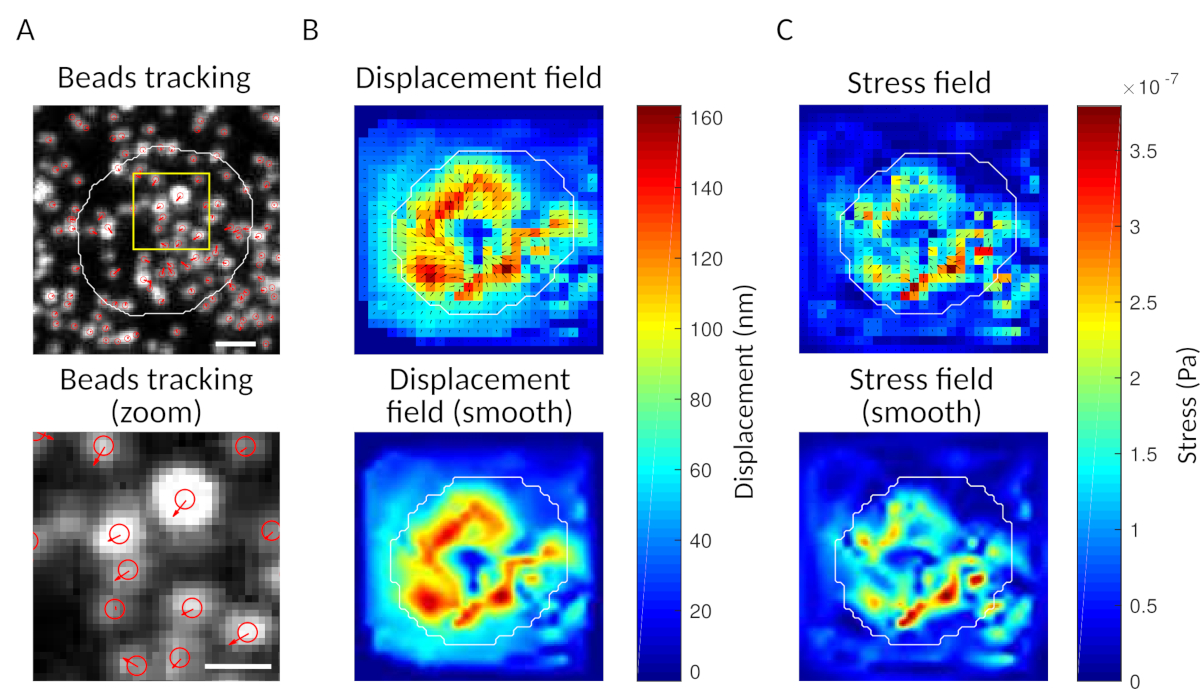{kind=link}

الشكل 4: مثال على المعلومات التي يمكن استخلاصها من حقول القوة والتشريد. (أ) مثال لتطور الطاقة في الوقت المناسب لخلية واحدة: تظهر مرحلة الهضبة (المميزة باللون الرمادي) بعد حوالي 10 دقائق. (B) مقارنة منحنيات الطاقة المتوسطة و (C) من مستويات الهضبة النسبية ل 65 خلايا مطلية على هلام HEL (تفعيل) المغلفة و 35 خلية على جل مغطي BSA (غير تفعيل) (متوسط ± بين الربع تظهر، تم استخدام اختبار مان ويتني للأهمية الإحصائية). (D) الوقت الفاصل خرائط اللون من الإجهاد ل HEL والسيطرة على شرط BSA؛ يتم عرض كل من حجم وجععبة المؤامرات. وقد تم تكييف هذه الصور من كوماري وآخرون6. الرجاء النقر هنا لعرض نسخة أكبر من هذا الرقم.
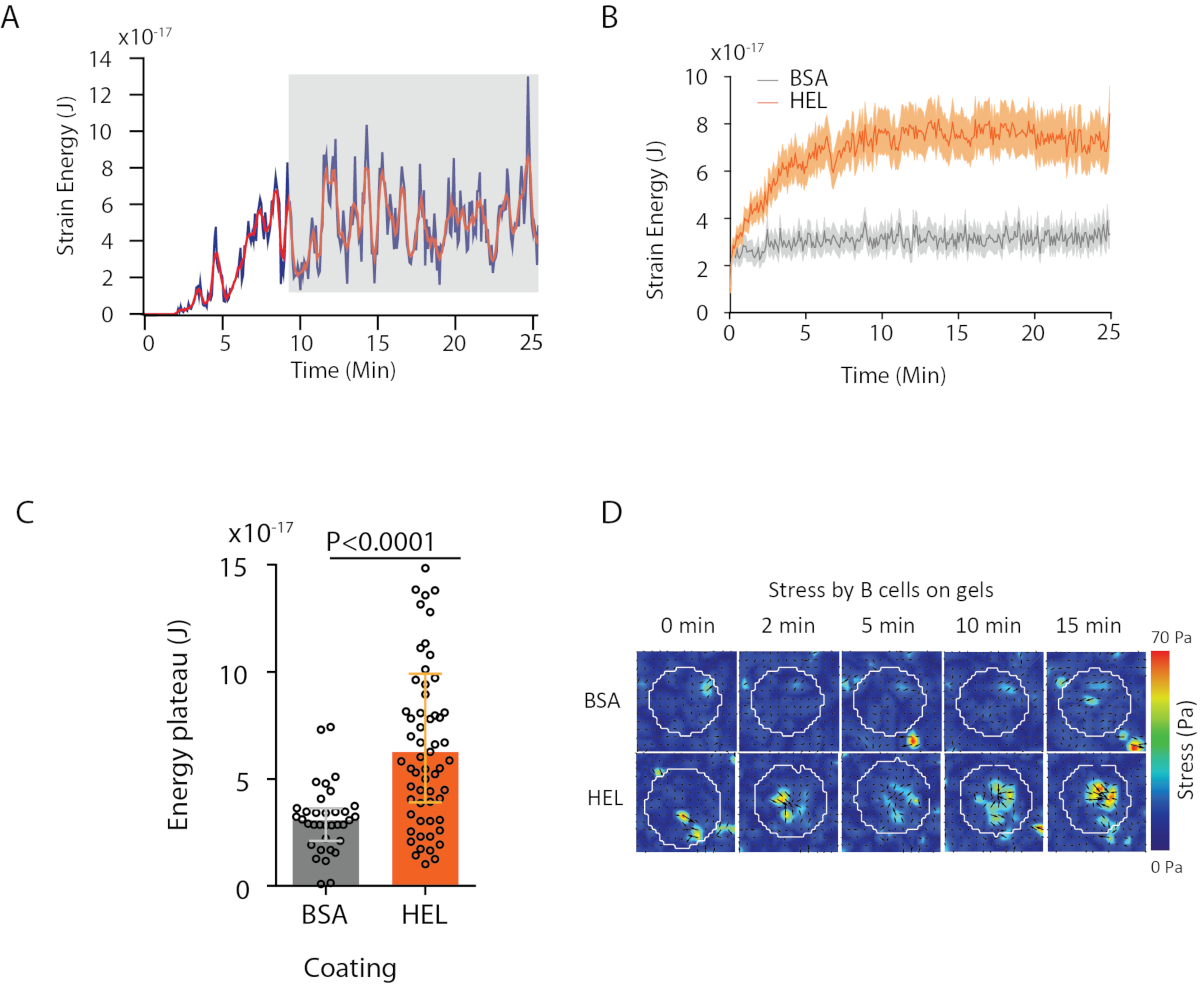{kind=link}

الشكل 5: مثال على التجارب مع مستضد الفلورسنت. (A) الفاصل الزمني لاستخراج الفلورسنت هيل (أدناه: النسبة المئوية للحد الأقصى، شريط مقياس = 3μm). (B) جمع المستضدات بمرور الوقت (متوسط ± SEM، n = 15). وقد تم تكييف هذه الصور من كوماري وآخرون6. الرجاء النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}
Discussion
طريقة TFM الموصوفة هنا تسمح للدراسة المنهجية للقدرات الميكانيكية النشطة للخلايا B. في سياق الخلايا B، وهذا يرتبط بالقدرة على استخراج واستيعاب المستضد. بالمقارنة مع أساليب TFM الأخرى ، فإن البروتوكول المقدم هنا بسيط ويمكن استنساخه إلى حد ما: الصلابة ، التي تقاس بالمسافات البادئة للميكروسفير الزجاجي واستخدام نموذج Hertz ، تتراوح بين 400 و 600 باسكال. وقد تم استخدام بروتوكولات مماثلة بنجاح ليس فقط للخلايا B35 ولكن أيضًا للخلايا T36. بالمقارنة مع nanopillars (تستخدم أيضا لتي اللمفاويات37)فإنه يوفر سطحا مستوًا متجانسًا ، وبالتالي تكون النتائج أسهل في تفسير تفاعل الجل مقيدة بشكل رئيسي لتكون عرضية على السطح.
البروتوكول الذي وصفناه يتيح الوصول إلى الديناميات الزمانية الزقتية للقوات التي تمارسها الخلايا B على ركائز تقديم المستضد. على المستوى المكاني، يوفر هذا معلومات عن توطين القوات، وبالاقتران مع المجهر الفلوري، يمكّن المجرب من ربط القوى المحلية بوجود جزيئات محددة (أي مكونات الهيكل الكيسي أو BCR الإشارات تتالي). على المستوى الزمني، من الممكن دمج الكميات (مثل الطاقة الإجمالية أو الإجهاد الكلي) لتوفير قيمة واحدة لكل نقطة زمنية وتقليل الضوضاء. هذا يسمح لمراقبة تطور قوة الجر في الوقت (النمو والهضبة) ووجود أنماط النبض.
وتوصف الجوانب التجريبية الحرجة للتحليل بأنها التالية. '1' كثافة الخلايا: لإجراء تحليل صحيح، ينبغي فصل الخلايا بشكل كاف. نحن نعتبر الخلية قابلة للتحليل إذا كان لديها منطقة فارغة من حجمها الخاص حولها. '2' صورة الإرسال: من المستحسن جمع صورة انتقال للخلايا على الأقل أثناء التجربة لاستخدامها كقناع في التحليل. '3' عدد الخرزات في الصورة: نقترح تحليل الصور فقط التي يتراوح فيها عدد الخرزات في المشبك بين 30 و200 (أي 1-8 حبات/ميكرومتر). ولا تسمح الكثافة المنخفضة بإعادة بناء التشريد على الخرائط على نحو كاف. تجعل كثافات الخرز العالية تتبع الجسيمات المفردة غير موثوق بها. '4' ينبغي أن يكون عدد الخرز ثابتا أثناء التجربة؛ ومع ذلك، يمكن أن تحدث التقلبات بسبب التغيرية الصغيرة في ظروف التصوير (وخاصة في الخرز التي هي قريبة جدا من بعضها البعض). يجب تصحيح الانجراف التركيز، إذا حدث، وينبغي تجاهل الأطر إشكالية. '5' نوعية الجل: ينبغي التخلص من المواد الهلامية التي بها الكثير من الشقوق أو التباين في توزيع الخرز أو المواد الهلامية السميكة جدا. '6' تبعا لنوع الخلية، بعد التعرض المتكرر، قد تعاني الخلايا في وقت متأخر من الوقت (> 300 إطار) من آثار تكسية ضوئية. من المستحسن تشغيل البرنامج على قناع يخلو من الخلايا كـ "خط أساس" للمقارنة مع البيانات. وهذا يوفر حجم مستوى الضوضاء فقط بسبب الظروف التجريبية.
المواد الهلامية المستخدمة لقياس قوة الجر في الالتصاق الكلاسيكية تسمح للتحقيق في العمليات التي تحدث في الالتصاق البؤري (التدفقات actin وتجنيد جزيئات الإشارة)—النقاط حيث يتم تطبيق القوات38,39. ومع ذلك، لا يتم تطبيق القوات في المشبك من خلال الالتصاقات البؤرية. لم يتم التحقيق كميا نمط ازن الزمني من توليد القوة في الخلية B المشبك المناعي باستخدام هذه الطريقة حتى وقت قريب. باستخدام TFM، لاحظنا للمرة الأولى، نقش القوة في المناشع الخلية B، كما هو معروض في دراستنا الأخيرة6، وفتح وجهات النظر المشجعة في دراسة الخلايا الليمفاوية.
بشكل ملحوظ، هذه الطريقة توظف صورة مأخوذة قبل وصول الخلايا على الجل كصورة مرجعية لحساب القوة. تشير بروتوكولات TFM المعتادة إلى أخذ الصورة المرجعية في نهاية التجربة ، بعد فصل الخلايا باستخدام التربسين ؛ وهذا يسمح للمجرب للبحث عن منطقة غنية في الخلايا. على الرغم من أن هذا ممكن هنا أيضا، التربسين غير فعالة بدلا في فصل الخلايا B من هلام مستضد المغلفة، يحتاج المرء إلى الانتظار طويلا فصيفة وخطر تعديل هلام والحركات (التي تجعل مجموعة البيانات بأكملها غير قابل للتسؤ) هو أعلى.
الطريقة المعروضة هنا مرنة ويمكن تطبيقها لدراسة تأثير الإشارات الأخرى في المشبك المناعي لأنها تسمح بتطعيم بروتينات أخرى على سطح الجل (على سبيل المثال، تم اختبار اللوينتينتين والغلوبولين المناعي) وحتى مستضد الفلورسنت (انظر القسم 4). وعلاوة على ذلك، تبقى الخلايا في متناول المجرّب للعلاج من المخدرات والانزعاجات المحلية. وأخيراً، فإن الطريقة متوافقة أيضاً مع التصوير بالخلايا الثابتة. لهذه الملاحظات، فمن المستحسن لجعل هلام على غطاء، وصمة عار الخلايا، الغراء غطاء على شريحة وفقط ثم إضافة وسائل الإعلام المتصاعدة وآخر coverslip. ثم سيتم القيام بالملاحظة مع الجل على القمة لتجنب تدهور الصورة من خلال هلام.
المزالق المحتملة هي التباين في الجل في البلمرة والطلاء. مشاكل البلمرة ويرجع ذلك أساسا إلى نوعية البادئ / محفز. أيضا، يمكن أن تضخيم هلام، وخاصة إذا لم تستخدم الحق بعد التجمع. لا يبدو أن هذه المشكلة تؤثر بشكل كبير على الخصائص الميكانيكية للجل ، ولكنها يمكن أن تجعل طبقة الخرز غير قابلة للوصول للهدف ، مما يجعل الجل عديم الفائدة بشكل فعال. نوصي بإعداد مواد هلامية إضافية لكل حالة عند ظهور هذه المشكلة. قد يكون هناك أيضا بعض التباين في الطلاء، وأنه من الأهمية بمكان أن يكون المخفف حديثا Sulfo SANPAH.
في الختام، وصفنا طريقة بسيطة ورخيصة وقابلة للاستنساخ لقياس القوى التي تمارسها الخلايا B في المشبك المناعي عندما يتم تنشيطها من قبل BCR ligand. ويمكن تكييفها لدراسة رد فعل ليغاندس أخرى وأنواع أخرى من الخلايا الليمفاوية (الذاكرة الخلايا B, الخلايا التائية, إلخ) مع استخدام يغاند مستقبلات السليم.
Disclosures
ليس لدى أصحاب البلاغ ما يكشفون عنه.
Acknowledgements
يشكر المؤلفون م. بولجر مونرو على القراءة النقدية ويعترفون Center@CNRS تصوير نيكون ومعهد البحوث الطبية PICT-IBiSA، ومعهد كوري، باريس، عضو البنية التحتية الوطنية للبحوث في فرنسا- BioImaging، لدعمها في الحصول على الصور ومرفق كوري الحيواني. وقد حظي هذا الحزب بتأييد المركز الوطني للبحوث العلمية. وقد تم دعم AK و JP من قبل زمالة باريس ديكارت دكتوراه وإركوب دكتوريل فاير - برنامج Bettencourt. تم تمويل هذا المشروع من خلال منح PP (ANR-10-JCJC-1504-Immuphy) و AMLD (ANR-PoLyBex-12-BSV3-0014-001، ERC-Strapacemi-GA 243103).
Materials
| Name | Company | Catalog Number | Comments |
| 3-aminopropyltrimethoxysilane (APTMS) | Sigma-Aldrich | 281778 | Store aliquoted, protected from humidity |
| 40% Acrylamide Solution | Biorad | 1610140 | |
| Alexa555 microscale protein labeling kit | Molecular Probes | A30007 | |
| Ammonium Persulfate (APS) | Sigma-Aldrich | A3678 | |
| B cell Isolation Kit, Mouse | Miltenyi Biotec | 130-090-862 | |
| B-mercaptoethanol | Gibco | 31350-010 | |
| 2% Bis Solution | Biorad | 161-0142 | |
| Bovine Serum Albumin (BSA) | Euromedex | 04-100-812-C | |
| Coverslip 18mm | VWR | 631-1580 | |
| Fetal calf serum | PAA | A15-151 | Decomplemented (40min @56°C) |
| Fluorodishes FD35 | World Precision Instruments, Inc | FD35100 | |
| Fluosphere: carboxylate-modified, 0.2um, dark red | Molecular Probes | F8807 | |
| Hen Egg Lysozyme | Sigma-Aldrich | L6876 | Stocked in aliquote 100mg/ml |
| MEM Non-Essential Amino Acids Solution (100X) | Thermofisher/Gibco | 11140035 | |
| N,N,N',N'-tetrametiletilendiammine (TEMED) | Euromedex | 50406-B | |
| PBS (Phosfate Buffer Saline) | Gibco | 10010-015 | |
| Penicillin–streptomycin | Gibco | 15140-010 | |
| RMPI 1640 – Glutamax I | Thermofisher | 61870-010 | |
| Sigmacote | Sigma-Aldrich | SL2 | |
| Sodium pyruvate | Gibco | 11360-039 | |
| sulfosuccinimidyl 6-(4'-azido-2'-nitrophenylamino)hexanoate (Sulfo-SANPAH) | Thermo Scientific | 22589 |
References
- Yuseff, M. -. I., Pierobon, P., Reversat, A., Lennon-Duménil, A. -. M. How B cells capture, process and present antigens: a crucial role for cell polarity. Nature Reviews. Immunology. 13 (7), 475-486 (2013).
- Spillane, K. M., Tolar, P. B cell antigen extraction is regulated by physical properties of antigen-presenting cells. The Journal of Cell Biology. 216 (1), 217-230 (2017).
- Shaheen, S., Wan, Z., et al. Substrate stiffness governs the initiation of B cell activation by the concerted signaling of PKCβ and focal adhesion kinase. eLife. 6, (2017).
- Natkanski, E., et al. B cells use mechanical energy to discriminate antigen affinities. Science. 340 (6140), 1587-1590 (2013).
- Wan, Z., Chen, X., et al. The activation of IgM- or isotype-switched IgG- and IgE-BCR exhibits distinct mechanical force sensitivity and threshold. eLife. 4, (2015).
- Kumari, A., Pineau, J., et al. Actomyosin-driven force patterning controls endocytosis at the immune synapse. Nature Communications. 10 (1), 2870 (2019).
- Dembo, M., Wang, Y. L. Stresses at the cell-to-substrate interface during locomotion of fibroblasts. Biophysical Journal. 76 (4), 2307-2316 (1999).
- Sabass, B., Gardel, M. L., Waterman, C. M., Schwarz, U. S. High resolution traction force microscopy based on experimental and computational advances. Biophysical Journal. 94 (1), 207-220 (2008).
- Schwarz, U. S., Soiné, J. R. D. Traction force microscopy on soft elastic substrates: A guide to recent computational advances. Biochimica et Biophysica Acta. 1853 (11), 3095-3104 (2015).
- Colin-York, H., Shrestha, D., et al. Super-Resolved Traction Force Microscopy (STFM). Nano Letters. 16 (4), 2633-2638 (2016).
- Stubb, A., Laine, R. F., Guzmán, C., Henriques, R., Jacquemet, G., Ivaska, J. Fluctuation-Based Super-Resolution Traction Force Microscopy. BioRxiv. , (2019).
- Gutierrez, E., Tkachenko, E., et al. High refractive index silicone gels for simultaneous total internal reflection fluorescence and traction force microscopy of adherent cells. Plos One. 6 (9), 23807 (2011).
- Bergert, M., Lendenmann, T., et al. Confocal reference free traction force microscopy. Nature Communications. 7, 12814 (2016).
- Schoen, I., Hu, W., Klotzsch, E., Vogel, V. Probing cellular traction forces by micropillar arrays: contribution of substrate warping to pillar deflection. Nano Letters. 10 (5), 1823-1830 (2010).
- Colin-York, H., Fritzsche, M. The future of traction force microscopy. Current Opinion in Biomedical Engineering. 5, 1-5 (2018).
- Feng, Y., et al. Mechanosensing drives acuity of αβ T-cell recognition. Proceedings of the National Academy of Sciences of the United States of America. 114 (39), 8204-8213 (2017).
- Spillane, K. M., Tolar, P. DNA-Based Probes for Measuring Mechanical Forces in Cell-Cell Contacts: Application to B Cell Antigen Extraction from Immune Synapses. Methods in Molecular Biology. 1707, 69-80 (2018).
- Stabley, D. R., Jurchenko, C., Marshall, S. S., Salaita, K. S. Visualizing mechanical tension across membrane receptors with a fluorescent sensor. Nature Methods. 9 (1), 64-67 (2011).
- Merkel, R., Nassoy, P., Leung, A., Ritchie, K., Evans, E. Energy landscapes of receptor-ligand bonds explored with dynamic force spectroscopy. Nature. 397 (6714), 50-53 (1999).
- Hinterdorfer, P., Dufrêne, Y. F. Detection and localization of single molecular recognition events using atomic force microscopy. Nature Methods. 3 (5), 347-355 (2006).
- Sawicka, A., Babataheri, A., et al. Micropipette force probe to quantify single-cell force generation: application to T-cell activation. Molecular Biology of the Cell. 28 (23), 3229-3239 (2017).
- Desprat, N., Guiroy, A., Asnacios, A. Microplates-based rheometer for a single living cell. Review of Scientific Instruments. 77 (5), 055111 (2006).
- Labernadie, A., Bouissou, A., et al. Protrusion force microscopy reveals oscillatory force generation and mechanosensing activity of human macrophage podosomes. Nature Communications. 5, 5343 (2014).
- Bouissou, A., Proag, A., et al. Protrusion force microscopy: A method to quantify forces developed by cell protrusions. Journal of Visualized Experiments. (136), 57636 (2018).
- Kronenberg, N. M., Liehm, P., et al. Long-term imaging of cellular forces with high precision by elastic resonator interference stress microscopy. Nature Cell Biology. 19 (7), 864-872 (2017).
- Basu, R., Whitlock, B. M., et al. Cytotoxic T cells use mechanical force to potentiate target cell killing. Cell. 165 (1), 100-110 (2016).
- Bufi, N., Saitakis, M., et al. Human Primary Immune Cells Exhibit Distinct Mechanical Properties that Are Modified by Inflammation. Biophysical Journal. 108 (9), 2181-2190 (2015).
- Goodnow, C. C., Crosbie, J., et al. Altered immunoglobulin expression and functional silencing of self-reactive B lymphocytes in transgenic mice. Nature. 334 (6184), 676-682 (1988).
- Butler, J. P., Tolić-Nørrelykke, I. M., Fabry, B., Fredberg, J. J. Traction fields, moments, and strain energy that cells exert on their surroundings. American Journal of Physiology. Cell Physiology. 282 (3), 595-605 (2002).
- . MBPPlab/TFM_v1: Software for Time dependent Traction Force Microscopy Available from: https://github.com/MBPPlab/TFM_v1 (2019)
- Tseng, Q., Duchemin-Pelletier, E., et al. Spatial organization of the extracellular matrix regulates cell-cell junction positioning. Proceedings of the National Academy of Sciences of the United States of America. 109 (5), 1506-1511 (2012).
- . ImageJ plugins by Qingzong TSENG Available from: https://sites.google.com/site/qingzongtseng/ (2019)
- Plotnikov, S. V., Sabass, B., Schwarz, U. S., Waterman, C. M. High-resolution traction force microscopy. Methods in Cell Biology. 123, 367-394 (2014).
- Schindelin, J., Arganda-Carreras, I., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Wang, J., Lin, F., et al. Profiling the origin, dynamics, and function of traction force in B cell activation. Science Signaling. 11 (542), (2018).
- Hui, K. L., Balagopalan, L., Samelson, L. E., Upadhyaya, A. Cytoskeletal forces during signaling activation in Jurkat T-cells. Molecular Biology of the Cell. 26 (4), 685-695 (2015).
- Bashour, K. T., Gondarenko, A., et al. CD28 and CD3 have complementary roles in T-cell traction forces. Proceedings of the National Academy of Sciences of the United States of America. 111 (6), 2241-2246 (2014).
- Gardel, M. L., Sabass, B., Ji, L., Danuser, G., Schwarz, U. S., Waterman, C. M. Traction stress in focal adhesions correlates biphasically with actin retrograde flow speed. The Journal of Cell Biology. 183 (6), 999-1005 (2008).
- Stricker, J., Sabass, B., Schwarz, U. S., Gardel, M. L. Optimization of traction force microscopy for micron-sized focal adhesions. Journal of Physics. Condensed Matter. 22 (19), 194104 (2010).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved