
Method Article
מיקרוסקופ כוח מתיחה ללמוד B לימפוציטים הפעלת
In This Article
Summary
כאן אנו מציגים פרוטוקול המשמש לביצוע ניסויי מיקרוסקופ כוח גרירה על תאי B. אנו מתארים את הכנת ג'לים פוליקרילאמיד רכים ואת הפונקציונליזציה שלהם, כמו גם רכישת נתונים במיקרוסקופ וסיכום של ניתוח נתונים.
Abstract
מיקרוסקופ כוח משיכה (TFM) מאפשר מדידה של כוחות המיוצרים על ידי תא עלמצע. טכניקה זו מסיקה מדידות כוח משיכה מתוך שדה עקירה שנצפה באופן ניסיוני המיוצר על ידי תא מושך על שקוע אלסטי. כאן, התאמנו TFM לחקור את המבנה המרחבי והזמני של שדה הכוח המופעל על ידי תאי B כאשר מופעל על ידי מעורבות אנטיגן של קולטן תא B. קשיחות ג'ל, צפיפות חרוזים, ופונקציונליזציה חלבון חייב להיות אופטימיזציה למחקר של תאים קטנים יחסית (~ 6 μm) כי אינטראקציה עם, ולהגיב במיוחד ליגנדים עבור קולטני פני השטח של התא.
Introduction
תאי B הם התאים המייצרים נוגדנים של המערכת החיסונית. כדי להפעיל את התגובה החיסונית הסתגלנית, הם רוכשים תחילה את האנטיגן בצורה מקורית (כלומר, לא מעובד) באמצעות קולטן ספציפי הנקרא קולטן תא B (BCR)1. תהליך זה מתרחש באזור תא צומת הלימפה B. גם אם כמה אנטיגן יכול להגיע לתא B באמצעות נוזלים לימפה, רוב האנטיגן, במיוחד עם משקל מולקולרי גבוה (>70 kDa, שהוא גודל המגבלה עבור מוליכים לימפה) אכן מוצגים בצורתם המקורית על פני השטח של תא אנטיגן מציג (APC), בדרך כלל מקרופאג סינוס תת-קספרלי או תא דנדריטי זקיק, באמצעות קולטני לקטין או FC (לא ספציפי). הקשר עם תא זה מוביל להיווצרות סינפסה חיסונית שבה BCR מפעיל כוח על אנטיגן הקשורים APC. הכריכה של אנטיגן ל-BCR יוזמת איתות BCR, אשר עשוי להפעיל מנגנונים ליצירת כוח. כוחות אלה יכולים להיות חשובים להגברת איתות BCR, אבל הם גם חיוניים עבור תאי B לחלץ ולאחר מכן להפנים את האנטיגן.
מחקרים שנעשו לאחרונה הראו כי BCR הוא אכן mechanosensitive2. לדוגמה, מצעים נוקשים יותר מעוררים איתות BCR משופר3. יתר על כן, כוח שנוצר על סינפסה חיסונית מושך על BCRs יחיד כדי לחקור את זיקתו אנטיגן ובכך להבטיח אפליה זיקה4. לכן מעניין לחקור את התגובה המכנית של תאי B למצגת אנטיגן ול לנתח תגובה זו במונחים של סוג של קולטנים מעורב (IgG /IgM)5,מולקולות הדבקה (ליגנדים אינטגריים) או בתאים מהונדסים גנטית ורוקמטית (כלומר, השתקת חלבון במורד הזרם של דינמיקת איתות BCR או ציטוסקלטון)6.
שיטה פשוטה כדי לבחון את התגובה של תא למצע של קשיחות פיזיולוגית, באותו זמן, כוחות מחקר המופעלים על המצע הוא מיקרוסקופ כוח משיכה (TFM). TFM מורכב מהתבוננות בשדה העקירה המיוצר על ידי התא מושך עלמצע אלסטי. במקור העיוות של הג'ל נצפתה באמצעות קמטים של elastomer עצמו על ידימיקרוסקופית ניגודיות שלב 7, אבל הכניסה של microbeads פלואורסצנס כמו סמנים fiducial מותר רזולוציה טובה יותר ומאזהפך תקן 8. שיטה זו שימשה כדי לחקור את כוח המתיחה המופעל על ידי תאים דבקים, רקמות, ואפילו אורגנואידים מוטבעים ג'לים. מספר וריאציות של TFMפותחו 9 כולל, בשילוב עם מיקרוסקופית superresolution (כלומר, STED10 או SRRF11),שינוי של אינדקס שבירה של הג'ל כדי לאפשר מיקרוסקופ TIRF12, החלפת חרוזים על ידי דפוסי ננומודפסים 13, ושימוש nanopillars במקוםמשטח שטוח 14. לסקירה מלאה של וריאציות אלה, ראה קולין-יורק ואח '15.
הפרוטוקול המוצג כאן מתאר הליך למדוד כוחות המופעלים על ידי תאי B על בהצעה מצופה אנטיגן. כוחות אלה מוחלים על הליגנדים (אנטיגן) על מנת לאגבר אותם ולאחר מכן לחלץ אותם מהמצע מציג אנטיגן. התאמנו את פרוטוקול TFM הסטנדרטי כדי לחקות את הקשיחות של מצעים פיזיולוגיים להצגת אנטיגן, את הגודל ואת הציפוי הרלוונטי לתאי B. פרוטוקול זה מאפשר לחקור מספר תאים בו זמנית, ניתן להשתמש בשילוב עם טכניקות מיקרוסקופיות פלואורס הצפי וטיפולים כימיים. עם זאת, היא אינה שואפת לחקור מדידות כוח מולקולה אחת, שעבורןפינצטה אופטית 16,בדיקות מתח מולקולרי 17,18, בדיקות כוח ביומברנה19, ומיקרוסקופ כוחאטומי 20 הם טכניקות מתאימות יותר. בהשוואה לשיטות מדידה אחרות של כוח תא יחיד (לדוגמה, micropipettes21 או microplates22)TFM מאפשר שחזור של מפה מלאה של הכוחות המופעלים על הסינסה עם רזולוציה של כ-300 00 00 00 00:00.00. זה שימושי כדי לזהות דפוסי spatio-זמני בכוחות המופעלים על פני השטח, כמו הג'ל תואם עם הדמיה confocal, כדי לתאם אותם עם גיוס של חלבונים ספציפיים (למשל, cytoskeleton וחלבונים איתות).
למרות TFM 3D אפשרי, זה אינו תואם את הקשיחות ואת ההתקנה שהשתמשנו. עיוותים ב3D הם בר השגה על ידי הגדרות מורכבות יותר כגון מיקרוסקופ כוח בורה (AFM סריקת קרום מעוות שבו התאים מצופים)23,24 ו הפרעות מהדהד אלסטיות מיקרוסקופיות מתח (ERISM, ג'ל הפועל כמו חלל מהדהד לאור והדגשת עיוותים של הצוללת עם דיוק של כמה ננומטר)25. למרות טכניקות אלה מבטיחות מאוד, הם עדיין לא הועסקו בתאי B. סוגים אחרים של TFM, כגוןבננו-פיילים 14, יכולים לשמש למצעים לשחזור. עם זאת, גיאומטריה זו אינה מותאמת לתאים רכים כאשר התא מצטלב בין העמודים, מה שמסבך את הניתוח. גישה זו אכן שימשה בתאי T כדי לבחון את היכולת של התא לבנות מבנים סביב העמודים26.
למרות הפשטות שלה, TFM באמצעות ג'לים polyacrylamide מאפשר תצפית בו זמנית של תאים רבים, ניתן ליישם בקלות ובמחיר בכל מעבדה מצויד ספסל מיקרוסקופ epifluorescence (אם כי אנו ממליצים על דיסק confocal / ספינינג).
כדי לחקות את הקשיחות הפיזיולוגית של APC, השתמשנו ג'לים polyacrylamide עם קשיחות של ~ 500 Pa27 ופונקציונלי הג'ל עם הפעלת אנטיגן. בפרוטוקול זה, פונקציונלינו את פני השטח של ג'ל polyacrylamide עם לייזום ביצת תרנגולת (HEL). הדבר מאפשר מדידה של כוחות שנוצרו על ידי גירוי של BCR באמצעות מעורבות של אתר מחייב אנטיגן. השימוש אנטיגן זה ותאי B ספציפיים HEL מעכברי MD4 מבטיח ייצור כוח אחיד יחסית בתגובה לרצועות אנטיגן28. עם זאת, מולקולות אחרות (כגון אנטי-IgM לעכברי B6) ניתן להשתיל על הג'ל, אבל הכוחות שנוצרו במקרים אלה יכול להיות הטרוגני יותר ופחות אינטנסיבי. מכיוון שתאי B הם תאים קטנים (קוטר ~ 6 μm), מספר החרוזים כבר אופטימיזציה להיות מקסימלי אך עדיין למעקב. עבור תאים גדולים המפעילים ~ kPa כוחות על מצעים שלהם, אחד יכול להשיג תוצאות משביעות רצון באמצעות חרוזים ודליל יחסית או ביצוע מהירות תמונה חלקיקים פשוטה (PIV) כדי לשחזר את שדה עיוות. עם זאת, עבור תאים קטנים כגון לימפוציטים B המפעילים מתח קטן כמו ~ 50 הרשות, השימוש מעקב חלקיקים יחיד נדרש (מהירות מעקב חלקיקים, PTV) כדי להשיג את הדיוק הרצוי בעת שחזור שדה עיוות. על מנת לעקוב באופן אמין אחר חרוזים בנפרד, ההגדלה של העדשה האובייקטיבית צריכה להיות לפחות פי 60 ואת הצמצם המספרי שלה בסביבות 1.3. לכן, הג'לים חייבים להיות דקים יחסית (<50 μm), אחרת החרוזים אינם גלויים כפי שהם מעל מרחק העבודה של המטרה.
הפרוטוקול העיקרי מורכב משלושה סעיפים: הכנת ג'ל, תפקוד ג'ל והדמיה; שני חלקים נוספים הם אופציונליים ומוקדשים לכמות והדמיה של תאי פלורסנט אנטיגן.
Protocol
1. הכנת ג'ל
- סילניזציה של תמיכת הג'ל
- הפעל את כיסוי או צלחת פטרי תחתית זכוכית (אשר ישמש כתמיכה ג'ל) עם מנורת UV במשך 2 דקות (לחכות 30 s לפני חשיפה למנורת UV כדי למנוע חשיפה לאוזון שיורית).
- תסכל את המנה עם כיסוי/זכוכית-תחתית באמצעות 200 μL aminopropyltrimethoxylane (APTMS) למשך 5 דקות. פעולה זו תכין את התמיכה לכריכה הקוולנטית של הג'ל.
- שטפו ביסודיות את המנה עם כיסוי/זכוכית-תחתית עם מים טהורים במיוחד.
- יבש את מכסה / זכוכית תחתית המנה באמצעות שאיפה ואקום.
- הכנת כיסוי 18 מ"מ המשמש לשטח הג'ל
- כדי להכין את כיסויי הכיסוי, תחילה הכניסו אותם למחזיק כיסוי קרמי. לאחר מכן לשים את מחזיק כיסוי לתוך כוסות קטנה (50 מ"ל) ולשפוך סיליקון רייגנט (מאוחסן ב 4 מעלות צלזיוס, reusable) על כיסויים, להיות בטוח לכסות אותם לחלוטין.
- מכסים את הציפוי בנייר אלומיניום ותחגרו במשך 3 דקות בטמפרטורת החדר. בזמן ההמתנה, מלאו גם קערה גדולה (500 מ"ל) במים אולטרה-טהורים. לאחר 3 דקות של דגירה בסיליקון רייגנט, להעביר את מחזיק כיסוי עם כיסויים לבקת המים.
- יש לשטוף היטב את הכיסויים במים טהורים במיוחד, לייבש אותם היטב ולשמור על מגבונים מנייר. לקבלת התוצאות הטובות ביותר, המשך מיד לסעיף הבא.
- פולימר ג'ל
- עבור ג'לים של 0.5 kPa, לערבב 75 μL של 40% אקרילאמיד עם 30 μL של 2% ביסקרילמיד (crosslinker) ו 895 μL של תמיסת מלח פוספט אגירה (PBS). ניתן לאחסן את הקדם-מיקס הזה למשך עד חודש אחד ב- 4°C.
- כדי 167 μL של 0.5 kPa ג'ל premix, להוסיף 1% (1.67 μL) של חרוזים, מערבולת sonicate במשך 5 דקות בsonicator אמבטיה (מנקה אולטראסוניות ספסל סטנדרטי עם כוח של 50-100 W ותדירות 40 kHz). שמור על התערובת מוגנת מפני אור באמצעות רדיד אלומיניום.
הערה: הקדם-מיקס אינו עושה polymerize עד הוספת היוזם (TEMED). - כדי לדלל פולימר, להוסיף 1% (1.67 μL) של 10% w / v אמוניום שכנוע (APS).
- כדי ליזום פולימר, להוסיף 0.1% (0.2 μL) N,N, N′,N′-N′-Tetramethylethylenediamine (TEMED). מערבבים עם פיפטה. לאחר הוספת APS ו- TEMED, הג'ל במהירות polymerizes כל כך להמשיך במהירות יציקת ג'ל.
- יציקת ג'ל
- Pipet 9 μL של תערובת ג'ל על כל מכסה / צלחת זכוכית התחתונה (ירידה במרכז, איור 1A)
- מניחים את כיסוי silanized / הידרופובי ולשטח את הג'ל(איור 1B). בעזרת מכבשים, לחצו על הכיסוי כדי לוודא שהז'ל מתפשט על פני כל האזור של הכיסוי(איור 1C)עד שהוא מתחיל לדלוף החוצה.
- להפוך את מכסה / צלחת זכוכית התחתונה לתוך צלחת פטרי גדולה והקש אותו על הספסל כדי לכפות חרוזים הולכים לכיוון משטח הג'ל(איור 1D).
- מכסים בנייר אלומיניום ומשאירים לשעה אחת כדי לפולימר בטמפרטורת החדר בתא לח (כלומר, מניחים רקמה רטובה מעל הצלחת כדי למנוע אידוי).
- לאחר שעה אחת, הוסף PBS לדגימה כדי להקל על שחרור כיסוי. בזהירות, להסיר את כיסוי באמצעות מחט (הציפוי עם סילאן שונים צריך לאפשר קילוף קל של כיסוי מהג'ל, איור 1E).
- תשאיר את הג'ל בפי.בי.אס.
הערה: כעת ניתן לאחסן ג'לים ב- PBS ב- 4 °C למשך 5-7 ימים, אך מומלץ להשתמש בהם בתוך 48 שעות.
2. תפקוד ג'ל
- להכין sulfosuccinimidyl 6-(4'-אזידו-2'-nitrophenylamino)הקסנואט (Sulfo SANPAH) פתרון ב 0.5 מ"ג /מ"ל ב 10 mM HEPES מאגר. ניתן לאחסן זאת ב-4°C מכוסה בנייר אלומיניום למשך שבוע.
- לשאוף PBS מג'לים.
- להוסיף 150 μL של Sulfo SANPAH לג'ל בטמפרטורת החדר (איור 1F).
- לחשוף את הג'ל לטיפול UV במשך 2 דקות כדי פוטואקטיבייט האתרים של Sulfo SANPAH ו לעשות את זה להיצמד אל פני השטח ג'ל.
- לשטוף עם PBS שלוש פעמים (איור 1G).
- חזור על שלבים 2.2-2.5.
- להוסיף 250 μL של HEL (100 μg / מ"ל) לכל ג'ל דגירה לילה בתא לח ב 4 מעלות צלזיוס לילה תוך שמירה על מכוסה רדיד אלומיניום(איור 1H).
- הסר אנטיגן HEL ולשטוף עם PBS שלוש פעמים.
הערה: HEL פועל הן כאנטיגן והן כמולקולה של הדבקה. זה יכול להיות מוחלף על ידי מולקולות אחרות הנקשרות לקולטן (למשל, IgM נגד עכבר, סרום פרווין אלבומין, Ovalbumin) או מעורבב עם ליגנדים אינטגרין (למשל, ICAM1 מחייב LFA1). במידת הצורך, חילוץ אנטיגן ניתן לצפות עם גרסה פלורסנט של HEL (הושג על ידי הכתמת המולקולה עם ערכת תיוג חלבון, ראה שלב 4). שים לב כי ריכוז נתון בכמויות גדולות לא יכול להניב את אותו ריכוז פני השטח על הג'ל כמו על הזכוכית: זה צריך להיות מכמת עם כתמים משניים אם נדרשת השוואה ישירה.
3. טעינה והדמיה של תאים
- לפני הדמיה, להסיר PBS מן הג'לים ולהוסיף 500 μL של מדיה סלולרית B (RPMI 1640, 10% סרום עגל עוברי מפורק, 1% פניצילין-סטרפטומיצין, 2% נתרן Pyruvate, 50uM Mercaptoethanol ו 1X חומצות אמינו שאינן Essential) ולתת להם equilibrate ל RT.
- הכנת תא
- טהר תאי B ראשיים מהטוחול בהתאם לפרוטוקול בחירה שלילי (ראה טבלת חומרים). תפוקת תאי B סופית טיפוסית היא סביב 1 x 107 תאים. רכז את זה 3 x 106 תאים / מ"ל במדיום תא B (RPMI-1640 בתוספת עם 10% סרום עגל עוברי, 1% פניצילין-סטרפטומיצין, 0.1% mercaptoethanol ו 2% נתרן pyruvate).
- אחסן תאים כצורך למשך עד 6 שעות ב- 4°C.
- שמור את התאים ב- 37 °C למשך 30 דקות לפני רכישת תמונה.
- הדמיה
- השתמש במיקרוסקופ קונפוקאלי עם בקרת CO2 תרמית (אולי) .
הערה: בין אם נעשה שימוש במיקרוסקופ של דיסק קונמקול או מסתובב, חשוב להשתמש במטרה/חור סיכה המאפשר לגודל פיקסל <200 צפון-מז' לעקוב בנוחות אחר החרוזים בשלב הניתוח (לדוגמה, 60x, NA 1.3). מיקרוסקופית Epifluorescence ניתן להשתמש גם, עם זאת הוא מספק אות נמוך יותר יחס רעש, עשוי להפוך את מעקב חרוז בודד קשה יותר. - שתי שכבות עיקריות של חרוזים יופיעו בתחתית ובהעליון של הג'ל. תתמקדי במטוס הג'ל.
הערה: ג'ל נחמד יופיע כשמיים כוכבים, עם חרוזים מופצים באופן אחיד בערך באותו מטוס. - לתכנת את הרכישה במשך 30 דקות עם קצב מסגרות של 5 s (זה מותאם לצרכי הניסוי, למשל, לרכוש צבעים אחרים, לרכוש z מחסנית, וכו ')
- לשאוף את התקשורת מן הג'ל, עוזב על 200 μL של מדיה על הג'ל. למקם את הג'ל על המיקרוסקופ ולמצוא את שכבת פני השטח של חרוזים אזור אחיד נחמד על הג'ל.
- להוסיף 80 μL של תאים (להימנע מלגעת בג'ל כדי לשמור על מיקוד).
- ודא כי המוקד עדיין נכון ותאים זה ניתן לראות יורד באזור (תחת אור משודר). הפעל את הרכישה לפני התאים להגיע הג'ל.
- במקרה של מגע מקרי עם ג'ל, תנודות, או מיקוד להיסחף, להתאים את המוקד.
הערה: זה חיוני לאסוף תמונה של ג'ל רגוע וזה יכול להיות כל תמונה שצולמה לפני הגעת התאים על הג'ל.
- השתמש במיקרוסקופ קונפוקאלי עם בקרת CO2 תרמית (אולי) .
4. ניסוי חילוץ פלורסנט HEL
- הכן HEL פלורסנט על ידי איגוד צבע פלורסנט (של צבע שונה מהחרוזים אחד כגון Alexa 555), ראה את טבלת החומרים.
- בשלב 2.7, החלף את HEL הקונבנציונלי ב-HEL הפלורסנט.
- רכוש תמונות עם הגדרות תאורה נמוכות או קצב פריימים נמוך (לדוגמה, 2 מסגרות לדקה) כדי למנוע הלבנת תמונות.
- כדי לכמת את החילוץ של HEL, חשב את העוצמה המשולבת על אזור התא עבור כל מסגרת I(t) מתוקנת ומנורמלת לפי העוצמה I(0) של מסגרת 0 בהתאם לנוסחה:

הערה: האנטיגן הנצבר עם פלואורופור אינו נראה לעין (כנראה בשל הרוות הפלואורופור על פני השטח של הג'ל), אך נוכחותו בג'ל יכולה להיות מאומתת עם נוגדן אנטי-HEL ונוגדן משני פלורסנט. ניתן לאמת כי פלואורופור הוא אכן פלורסנט כאשר מנותק על ידי הפשטתו מן הג'ל עם כיסוי מצופה אנטי HEL וחושף אותו עם נוגדן פלורסנט משני (על הכיסוי)6. האות של האנטיגן שחולץ הוא עמום מאוד ולעתים מוסך על ידי דליפת החרוזים. אם אדם מעוניין רק בחילוץ אנטיגן, מומלץ להכין את הג'ל ללא חרוזים (דלג על שלבים 1.3.2 ו-1.4.3).
5. הדמיית פלואורסץ
- להשיג תאי פלורסנט B על ידי טיהור תא B מהטומוס של עכברים מהונדסים גנטית כפי שנעשה עבור סוג הבר (למשל, מ Lifeact-GFP או עכברי Myosin II GFP).
- עבור תאי פלורסנט הדמיה, להשתמש (אם אפשר) מיקרוסקופ דיסק מסתובב עם מים טבילה למרחקים ארוכים 40x-100x המטרה.
- שמור על משך חשיפה וקצב פריימים נמוך כדי למנוע הלבנה.
הערה: פונקציית התפשטות הנקודה ב- Z מושפלת מאוד על ידי הנוכחות של הג'ל, ולכן אנו מציעים באמצעות מטרת טבילה במים. לחיות מיקרוסקופ זקוף עם מטרות טבילה במים סובל סטייות כדוריות חזקות המושרה על ידי הנוכחות של (כדורי) התא (וגרעין התא) בנתיב הפליטה.
6. ניתוח
הערה: ניתוח נתונים מתבצע באופן כללי על ידי תיקון הראשון של הערימה כולה להיסחף, מציאת החרוזים בכל מסגרת, מעקב אחר תנועותיהם ביחס למסגרת התייחסות (נלקח בהיעדר תאים), אינטרפולציה שדה העקירה והופך את הבעיה כדי להשיג את הלחץ באמצעות פורייה להפוך29. בסוף זה, אנו מציעים באמצעות שילוב של תוכניות מאקרו ImageJ ו MATLAB להורדה ממאגר מקוון30.
- פתיחת הסרט ב- ImageJ כערמת תמונות
- הפעל את המאקרו "Crop_and_save.ijm"
- בחר את אזורי העניין (ROI) באמצעות הכלי "מלבן" והוסף אותם לרשימת החזר השקעה באמצעות מקש 't'.
- בעת חיתוך התא, הקפד לכלול אזור של לפחות 5-10 פיקסלים של חרוזים לא ניידים. אל תכלול תאים קרובים מדי לגבולות או לתאים אחרים מהניתוח. בסיום לחץ על 'אישור'.
- המאקרו מציע מסיכה של התא: אם זה משביע רצון לחץ על "אישור". אם לא משביע רצון, לחץ על "לא בסדר" ולאחר מכן בחר באופן ידני אזור סגור עם כל כלי בחירה (למשל "Freehand" או "Oval") ולחץ על "המשך".
- פתח את MATLAB והריץ את "TFM_v1.m".
- קלטו את הפרמטרים הדרושים: במיוחד בדקו את מאפייני התמונה (גודל פיקסל, מרווח זמן לרכישה) ואת מאפייני הג'ל (מודולוס E הצעיר, יחס פואסון).
- תמונת ההפניה מוגדרת להיות הראשונה כברירת מחדל. הגדר אותה למסגרת אחרת במידת הצורך או הגדר אותה כ- "0" לטעינת קובץ חיצוני.
- אתר את יציאות התוכנה באותה ספריה של הקובץ המקורי (לקבלת תיאור, עיין בקובץ User_notice.pdf). זה כולל מסלול ראשוני של החרוזים ("FILENAME.fig"), חלקה של האנרגיה ההתכווצות לאורך זמן ("FILENAME_energy.fig"), טבלה של מספר כמויות משולבות על התא (אנרגיה, אזור, רגעים, וכו ') "FILENAME_finaltable.mat", מבנה המכיל את העקירה ושדה כוח, סרטים של החרוז, שדה עקירה, מתח ואנרגיה (שניתן לפתוח עם כל קורא avi).
הערה: בפרמטרים קלט, "גודל החלון" הוא החלון שבו העקירה הוא אינטרפולציה, ומכאן הרזולוציה הסופית של שדה מתח ועקירה. אפשרות זו מוגדרת לכמה פיקסלים (כברירת מחדל ארבעה). לא מומלץ להפחית זאת כפי שהוא יגדיל באופן מלאכותי את הרזולוציה על ידי אינטרפולציה אזורים שבהם אין חרוזים.
תוצאות
בהתחשב בגודל התאים, אלגוריתמים לחלץ את מפת העקירה של החרוזים באמצעות טכניקות קורלציה (כגון velocimetry תמונה חלקיקים) הם באופן כללי לא מאוד מדויק. עם זאת, בהתאם לרמת הרזולוציה הנדרשת, ניתן להשיג בקלות תוצאות איכותיות באמצעות תוסף Fiji/ImageJחינם 31,32. בעוד גישה זו מספיקה כדי להשוות מגרה לעומת תנאים לא מגרים, עבור ניתוח יסודי אנו ממליצים באמצעות התוכנה שלנו להורדהממאגר מקוון 30, העוקב אחר החרוזים בנפרד ומספק את מפת שדה עקירה בנקודת זמן נתונה כמו האינטרפולציה של עקירותחרוז בודדים 33. בשלב זה ניתן לכמת מספר כימות. לדוגמה (בהנחה שההעקירה נגרמת רק על ידי מתח משיק על פני השטח של הג'ל) התוכנה מספקת גם את הלחץ בכל נקודה הגורמת למפת העקירה הספציפית. זהו סוג של "בעיית היפוך": העקירה בשלב מסוים תלויה בסכום של כל הכוחות המיושם בכל הנקודות האחרות. "אלגוריתם ההיפוך" לוקח בחשבון את הפרמטרים הפיזיים של המקום: הקשיחות שלו (מודולוס צעיר) ויחס פואסון. אלגוריתמים ישירים הם בדרך כלל מאוד מדויקים אבל יקרים מבחינה חישובית. אלגוריתמים המבוססים על פורייה טרנספורמטיבי, כמו שלנו, מבצעים למעשה דה-אבולוציה בחלל פורייה ויעילים יותר אך נוטים לשגיאות מסוימות (בעיקר בשל שלב האינטרפולציה). אלגוריתמים אלה דורשים בדרך כלל כוונון של פרמטר המונע עקירה מקומית קטנה (ופוטנציאלית) להיות רלוונטית מדי בחישוב שדה הלחץ (פרמטר קבוע של Tikhonov8,29; משתנה "רגיל" בחלון הדו-שיח; כאן אנו מגדירים בדרך כלל שווה ל- 5 x 10-19). לפרשנות וניתוח מתקדמים יותר, כגון מתאם ספטיו-זמני, תנועות מקומיות, התאמות עם ערוצי פלורסנט, אנו ממליצים לשתף פעולה עם מומחים בתחום. לסקירה על שיטות חישוביות ראו שוורץ ואח'9.
כפי שהוזכר לעיל, תמונות חרוז נכונות נראות כמו "שמיים כוכבים", חלוקה אחידה ואקראית של נקודות בהירות(איור 2A). נתונים וניתוח אינם אמינים כאשר מספר החרוזים נמוך מדי(איור 2B)או שהתמונה אינה בפוקוס (איור 2C). לאחר תאי B התיישבו על פני השטח של הג'ל, החרוזים מתחת לתאים מתחילים לנוע בשל כוח המתיחה המופעל על ידי התא על הג'ל. יש להשליך מסגרות שעבורן אין לבטל את החרוזים.
כבדיקה, ניתן להבחין בעין את תנועת החרוזים המשווים את "מסגרת ההפניה", בדרך כלל זו שקדמה למגע הראשון של התא עם העיתור. ניתן להשיג תוצאות משוותות ממעקב החלקיקים הבודד (לדוגמה, Trackmate, Fiji 34)כפי שנעשה באיור 3A. הניתוח מספק פילוח של החרוזים בתדמית ההפניה ("FILENAME.fig") כפקד.
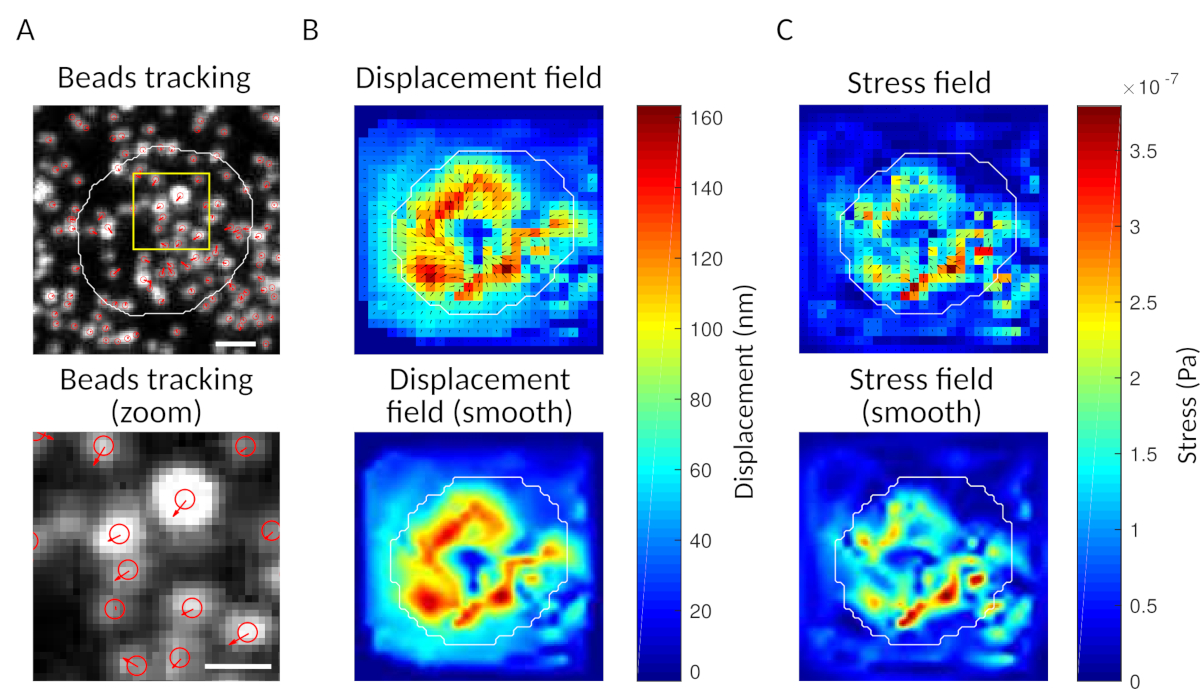
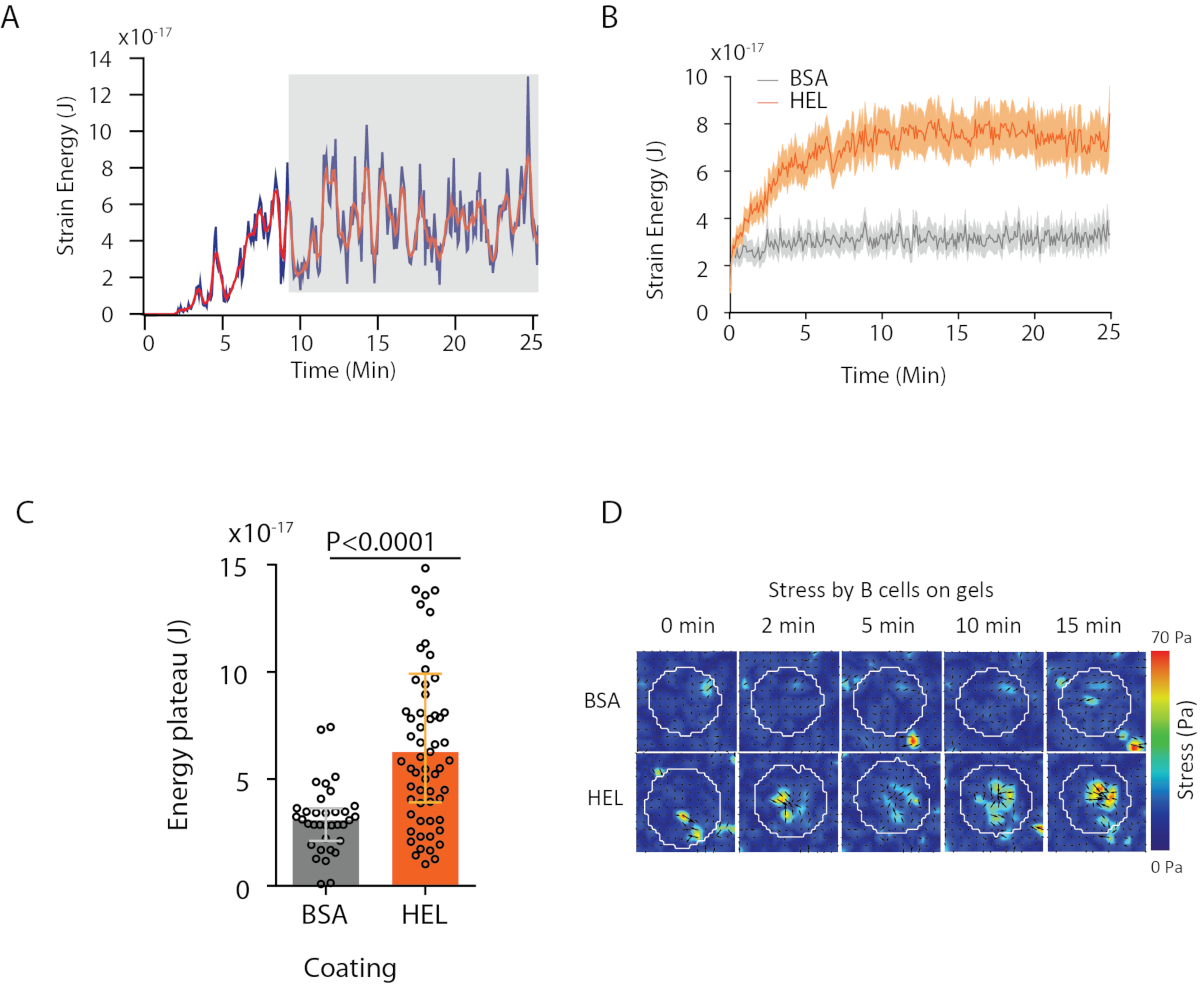
עם התוכנה שאנו מציעים, ניתן להשיג את העקירה (איור 3B) ואת שדה הלחץ (הווקטור של הלחץ המקומי בכל פיקסל ובכל נקודת זמן שהושגה על ידי היפוך מתוך שדה העקירה, איור 3C). המוצר הסקאלי של שדות העקירה והכוח המשולבים באזור התא מספק עבודה כוללת המופעלת על ידי התא בבצע(איור 4א). חישוב זה דורש את המסיכה של התא שהוכנס בשלב 6.2 של הפרוטוקול.
כדי להשוות בין שני תנאים ביולוגיים (כמו הפעלת HEL לעומת BSA שאינו מפעיל, או סוג פראי לעומת נוק-אאוט) שימושי לחשב את העקומה הממוצעת(איור 4B)או, אפילו יותר סינתטי, ערך ממוצע בנקודות הזמן האחרונות (20 דקות) שבהן האנרגיה מגיעהלרמה (איור 4C). כאשר המידע המרחבי של הכוחות רלוונטי, ניתן להשוות נקודות זמן בודדות של כל תנאי(איור 4D). עיין קומארי ואח'6 לניתוח מעמיק יותר.
דוגמה של פלואורסצנטית אנטיגן חילוץ זמן לשגות מוצג איור 5A: המראה הפרוגרסיבי של אותות פלואורסצנטיים בסינפסה הצביע הניתוק אנטיגן מן הג'ל. עקומת החילוץ הממוצעת עם מרווח הביטחון שלה (שגיאה סטנדרטית של הממוצע) מעל 15 תאים מוצגת באות 5B.

איור 1: הצגה סכמטית של הכנת הג'ל והפונקציונליזציה שלו. שלבים מתוארים בפרוטוקול. לחץ כאן כדי להציג גירסה גדולה יותר של נתון זה.
{kind=link}

איור 2: שלוש דוגמאות של תמונות חרוזים של תכונות שונות. (A) דוגמה של תמונת חרוז עם יחס האות הנכון לרעש ואת הצפיפות הנכונה. (ב)דוגמאות של תמונות עם מספר לא מספיק מדי של חרוזים ו -(C)מחוץ מישור מיקוד. לחץ כאן כדי להציג גירסה גדולה יותר של נתון זה.
{kind=link}

איור 3: עיבוד התמונות כדי לחלץ את שדה הכוח. (א)דוגמה לתמונה של החרוזים (קו מתאר של התא בלבן, שחולצו מתנופת השידור), מעקב אחר חרוזים בזמן t = 5 דקות (שכבת-על אדומה) והעקירה (חצים) ביחס ל- t זמן = 0 דקות (סרגל קנה מידה 5 μm). (ב)שדה עקירה אינטרפולציה (המיוצג כמפת רעד וקטורית ומפת סדר גודל, חצים הם פרופורציונליים לעקירה [nm]; ראה את סרגל הצבע מימין); תחתית: תמונה חלקה יותר בסדר הגודל (הושגה על ידי אינטרפולציה עם פונקציה דו-קובית). (ג)שדה מתח פילד עקירה בלוח ב' (המיוצג כמפת רעד וקטורית ומפת סדר גודל; חצים פרופורציונליים ללחץ גייה [הרשות הפלסטינית]; ראה את סרגל הצבע מימין); תחתית: תמונה חלקה יותר בסדר הגודל (הושגה על ידי אינטרפולציה עם פונקציה דו-קובית). לחץ כאן כדי להציג גירסה גדולה יותר של נתון זה.
{kind=link}

איור 4: דוגמה למידע שניתן לחלץ שדות כוח ועקירה. (א)דוגמה לאבולוציה של אנרגיה בזמן עבור תא בודד: שלב הרמה (מסומן באפור) מופיע לאחר כ- 10 דקות. (ב)השוואה של עקומות האנרגיה הממוצעות ו -(C)של רמות הרמה היחסית עבור 65 תאים מצופה HEL (הפעלת) ג'ל מצופה ו 35 תאים על BSA (לא מפעיל) ג'ל מצופה (חציון ± טווחים interquartile מוצגים, מבחן מאן-ויטני שימש משמעות סטטיסטית). (ד)מפות צבע של מתח עבור HEL ושליטה במצב BSA; הן סדר גודל וגם חלקות רעד מוצגות. תמונות אלה הותאמו מקמארי ואח'6. לחץ כאן כדי להציג גירסה גדולה יותר של נתון זה.
{kind=link}

איור 5: דוגמה לניסויים באנטיגן פלורסנט. (א)זמן לשגות החילוץ של HEL פלורסנט (להלן: אחוז המקסימום, סרגל קנה מידה = 3μm). (ב)איסוף אנטיגן לאורך זמן (± SEM, n = 15). תמונות אלה הותאמו מקמארי ואח'6. לחץ כאן כדי להציג גירסה גדולה יותר של נתון זה.
{kind=link}
Discussion
שיטת TFM המתוארת כאן מאפשרת מחקר שיטתי של היכולות המכניות הפעילות של תאי B. בהקשר של תאי B, זה קשור ליכולת לחלץ ולהפנים את האנטיגן. בהשוואה לשיטות TFM אחרות, הפרוטוקול המוצג כאן הוא פשוט ולא לשחזור: הקשיחות, נמדד על ידי חריץ של מיקרוספירה זכוכית ובשימוש מודל הרץ, הוא בין 400 ו 600 הרשות הפלסטינית. פרוטוקולים דומים שימשו בהצלחה לא רק עבור תאי B35 אלא גם עבור תאי T36. בהשוואה nanopillars (משמש גם עבור לימפוציטים T37)הוא מספק משטח הומוגני שטוח, ולכן התוצאות קל יותר לפרש כמו האינטראקציה של הג'ל מוגבלים בעיקר להיות משיק על פני השטח.
הפרוטוקול שתיארנו נותן גישה לדינמיקה של הכוחות המופעלים על ידי תאי ב' על מצעים מציגי אנטיגן. ברמה המרחבית זה מספק מידע על ההתקליזציה של כוחות, ובשילוב עם מיקרוסקופ פלואורסצנט, מאפשר לנסוי לתאם כוחות מקומיים עם נוכחות של מולקולות ספציפיות (כלומר, רכיבים של השלד ציטוסק או BCR איתות מפל). ברמת הזמן, ניתן לשלב כמויות (כגון אנרגיה כוללת או מתח כולל) כדי לספק ערך אחד לכל נקודת זמן ולהפחית את הרעש. הדבר מאפשר התבוננות באבולוציה של כוח המתיחה בזמן (צמיחה ומישור) ונוכחות של דפוסי פועם.
היבטים ניסיוניים קריטיים לניתוח מתוארים כהודעה הבאה. (i) צפיפות תאים: כדי לבצע ניתוח נכון, יש להפריד את התאים במידה מספקת. אנו מחשיבים תא שניתן לנתחו אם יש לו אזור ריק בגודלו סביבו. (2) תמונת שידור: מומלץ לאסוף לפחות תמונת שידור של התאים במהלך הניסוי שישמש כמסכה בניתוח. (iii) מספר החרוזים בתמונה: אנו מציעים לנתח רק תמונות שבהן מספר החרוזים בסינפסה הוא בין 30 ל-200 (כלומר, 1-8 חרוזים/μm²). צפיפויות נמוכות יותר אינן מאפשרות שחזור עקירת מפה הולם. צפיפויות חרוזים גבוהות הופכות מעקב אחר חלקיקים בודדים ללא אמין. (4) מספר החרוזים צריך להיות קבוע במהלך הניסוי; עם זאת, תנודות עלולות להתרחש עקב שונות קטנה בתנאי ההדמיה (במיוחד בחרוזים הקרובים מדי זה לזה). יש לתקן את הסחף של המוקד, אם מתרחש, ויש למחוק מסגרות בעייתיות. (v) איכות ג'ל: ג'לים עם סדקים רבים מדי, שונות בחלוקת חרוזים או ג'לים עבים מדי יש להשליך. (vi) בהתאם לסוג התא, לאחר חשיפות חוזרות ונשנות, תאים בנקודות זמן מאוחרות (>300 מסגרות) עלולים לסבול מאפקטים פוטוטוקסיים. מומלץ להפעיל את התוכנית על מסיכה נטולת תאים כ"בסיס" להשוואה עם הנתונים. זה מספק גודל של רמת הרעש אך ורק בשל התנאים הניסיוניים.
ג'לים המשמשים למדד כוח משיכה בהדבקה קלאסית מאפשרים חקירה של תהליכים המתרחשים בהדבקה המוקדית (זרמי א-אקטין וגיוס מולקולות איתות)– הנקודות שבהןמוחלים כוחות 38,39. עם זאת, כוחות בסינופסה אינם מיושמים באמצעות הידבקויות מוקד. דפוס spatiotemporal של דור כוח ב סינפסה חיסונית תא B לא נחקר באופן כמותי באמצעות שיטה זו עד לאחרונה. באמצעות TFM, הבחנו בפעם הראשונה, דפוס כוח בסינפסה חיסונית תא B, כפי שהוצג במחקר האחרון שלנו 6 ,פתיחת פרספקטיבות מעודדותבמחקר של לימפוציטים.
במיוחד, שיטה זו משתמשת בתמונה שצולמה לפני הגעת התאים על הג'ל כתדמית התייחסות לחישוב הכוח. פרוטוקולי TFM הרגילים מציעים לקחת את תמונת ההפניה בסוף הניסוי, לאחר נתק התאים עם trypsin; הדבר מאפשר לנסוי לחפש אזור עשיר בתאים. למרות שזה אפשרי גם כאן, trypsin הוא יעיל למדי בניתוק תאי B מג'ל מצופה אנטיגן, אחד צריך לחכות זמן רב לניתוק ואת הסיכון של שינוי ג'ל ותנועות (מה שהופך את כל הנתונים להגדיר בלתי מוסבר) הוא גבוה יותר.
השיטה המוצגת כאן היא גמישה, ניתן להחיל כדי ללמוד את ההשפעה של אותות אחרים על סינפסה חיסונית כפי שהוא מאפשר השתלת חלבונים אחרים על פני השטח של הג'ל (למשל, ליגנדים אינטגרין ו immunoglobulins נבדקו) ואפילו אנטיגן פלורסנט (ראה סעיף 4). יתר על כן, תאים נשארים נגישים לנסוי לטיפול תרופתי וturturbations מקומי. לבסוף, השיטה תואמת גם עם תאים קבועים הדמיה. עבור תצפיות אלה, מומלץ לעשות את הג'ל על כיסוי, להכתים את התאים, להדביק את כיסוי על שקופית ורק אז להוסיף מדיה הרכבה וכיסוי אחר. לאחר מכן התצפית תיעשה עם הג'ל למעלה כדי למנוע את ההשפלה של התמונה דרך הג'ל.
מלכודות אפשריות הן השונות בג'ל בפולימר וציפוי. בעיות פולימר הן בעיקר בשל איכות היוזם / זרז. כמו כן, הג'ל יכול לנפח, במיוחד אם לא נעשה שימוש מיד לאחר ההרכבה. בעיה זו לא נראה להשפיע באופן דרמטי על המאפיינים המכניים של הג'ל, אבל זה יכול להפוך את שכבת החרוז בלתי נגיש עבור המטרה, ביעילות מה שהופך את הג'ל חסר תועלת. אנו ממליצים להכין ג'לים נוספים עבור כל תנאי כאשר בעיה זו מופיעה. ייתכן שיש גם שונות מסוימת בציפוי, וזה חיוני יש מדולל טרי Sulfo SANPAH.
לסיכום, תיארנו שיטה פשוטה, זולה וניתנת לשחזור כדי למדוד את הכוחות המופעלים על ידי תאי B בסינפסה החיסונית כאשר הופעלו על ידי BCR ליגנד. זה יכול להיות מותאם ללמוד את התגובה ליגנדים אחרים סוגים אחרים של לימפוציטים (תאי זיכרון B, תאי T, וכו ') עם השימוש ליגנד קולטן הנכון.
Disclosures
לסופרים אין מה לחשוף.
Acknowledgements
המחברים מודים M. בולג'ר-מונרו על קריאה ביקורתית ולהכיר ניקון הדמיה Center@CNRS-InstitutCurie ו PICT-IBiSA, המכון קירי, פריז, חבר תשתית המחקר הלאומי צרפת-BioImaging, לתמיכה ברכישת תמונה ואת מתקן בעלי חיים קירי. PP נתמך על ידי CNRS. AK ו-JP נתמכו על ידי מלגת הדוקטורט של פריז דקארט ו-Ecole Doctorale FIRE – תוכנית בטנקורט. פרויקט זה מומן על ידי מענקים ל- PP (ANR-10-JCJC-1504-Immuphy) ו-AMLD (ANR-PoLyBex-12-BSV3-0014-001, ERC-Strapacemi-GA 243103).
Materials
| Name | Company | Catalog Number | Comments |
| 3-aminopropyltrimethoxysilane (APTMS) | Sigma-Aldrich | 281778 | Store aliquoted, protected from humidity |
| 40% Acrylamide Solution | Biorad | 1610140 | |
| Alexa555 microscale protein labeling kit | Molecular Probes | A30007 | |
| Ammonium Persulfate (APS) | Sigma-Aldrich | A3678 | |
| B cell Isolation Kit, Mouse | Miltenyi Biotec | 130-090-862 | |
| B-mercaptoethanol | Gibco | 31350-010 | |
| 2% Bis Solution | Biorad | 161-0142 | |
| Bovine Serum Albumin (BSA) | Euromedex | 04-100-812-C | |
| Coverslip 18mm | VWR | 631-1580 | |
| Fetal calf serum | PAA | A15-151 | Decomplemented (40min @56°C) |
| Fluorodishes FD35 | World Precision Instruments, Inc | FD35100 | |
| Fluosphere: carboxylate-modified, 0.2um, dark red | Molecular Probes | F8807 | |
| Hen Egg Lysozyme | Sigma-Aldrich | L6876 | Stocked in aliquote 100mg/ml |
| MEM Non-Essential Amino Acids Solution (100X) | Thermofisher/Gibco | 11140035 | |
| N,N,N',N'-tetrametiletilendiammine (TEMED) | Euromedex | 50406-B | |
| PBS (Phosfate Buffer Saline) | Gibco | 10010-015 | |
| Penicillin–streptomycin | Gibco | 15140-010 | |
| RMPI 1640 – Glutamax I | Thermofisher | 61870-010 | |
| Sigmacote | Sigma-Aldrich | SL2 | |
| Sodium pyruvate | Gibco | 11360-039 | |
| sulfosuccinimidyl 6-(4'-azido-2'-nitrophenylamino)hexanoate (Sulfo-SANPAH) | Thermo Scientific | 22589 |
References
- Yuseff, M. -. I., Pierobon, P., Reversat, A., Lennon-Duménil, A. -. M. How B cells capture, process and present antigens: a crucial role for cell polarity. Nature Reviews. Immunology. 13 (7), 475-486 (2013).
- Spillane, K. M., Tolar, P. B cell antigen extraction is regulated by physical properties of antigen-presenting cells. The Journal of Cell Biology. 216 (1), 217-230 (2017).
- Shaheen, S., Wan, Z., et al. Substrate stiffness governs the initiation of B cell activation by the concerted signaling of PKCβ and focal adhesion kinase. eLife. 6, (2017).
- Natkanski, E., et al. B cells use mechanical energy to discriminate antigen affinities. Science. 340 (6140), 1587-1590 (2013).
- Wan, Z., Chen, X., et al. The activation of IgM- or isotype-switched IgG- and IgE-BCR exhibits distinct mechanical force sensitivity and threshold. eLife. 4, (2015).
- Kumari, A., Pineau, J., et al. Actomyosin-driven force patterning controls endocytosis at the immune synapse. Nature Communications. 10 (1), 2870 (2019).
- Dembo, M., Wang, Y. L. Stresses at the cell-to-substrate interface during locomotion of fibroblasts. Biophysical Journal. 76 (4), 2307-2316 (1999).
- Sabass, B., Gardel, M. L., Waterman, C. M., Schwarz, U. S. High resolution traction force microscopy based on experimental and computational advances. Biophysical Journal. 94 (1), 207-220 (2008).
- Schwarz, U. S., Soiné, J. R. D. Traction force microscopy on soft elastic substrates: A guide to recent computational advances. Biochimica et Biophysica Acta. 1853 (11), 3095-3104 (2015).
- Colin-York, H., Shrestha, D., et al. Super-Resolved Traction Force Microscopy (STFM). Nano Letters. 16 (4), 2633-2638 (2016).
- Stubb, A., Laine, R. F., Guzmán, C., Henriques, R., Jacquemet, G., Ivaska, J. Fluctuation-Based Super-Resolution Traction Force Microscopy. BioRxiv. , (2019).
- Gutierrez, E., Tkachenko, E., et al. High refractive index silicone gels for simultaneous total internal reflection fluorescence and traction force microscopy of adherent cells. Plos One. 6 (9), 23807 (2011).
- Bergert, M., Lendenmann, T., et al. Confocal reference free traction force microscopy. Nature Communications. 7, 12814 (2016).
- Schoen, I., Hu, W., Klotzsch, E., Vogel, V. Probing cellular traction forces by micropillar arrays: contribution of substrate warping to pillar deflection. Nano Letters. 10 (5), 1823-1830 (2010).
- Colin-York, H., Fritzsche, M. The future of traction force microscopy. Current Opinion in Biomedical Engineering. 5, 1-5 (2018).
- Feng, Y., et al. Mechanosensing drives acuity of αβ T-cell recognition. Proceedings of the National Academy of Sciences of the United States of America. 114 (39), 8204-8213 (2017).
- Spillane, K. M., Tolar, P. DNA-Based Probes for Measuring Mechanical Forces in Cell-Cell Contacts: Application to B Cell Antigen Extraction from Immune Synapses. Methods in Molecular Biology. 1707, 69-80 (2018).
- Stabley, D. R., Jurchenko, C., Marshall, S. S., Salaita, K. S. Visualizing mechanical tension across membrane receptors with a fluorescent sensor. Nature Methods. 9 (1), 64-67 (2011).
- Merkel, R., Nassoy, P., Leung, A., Ritchie, K., Evans, E. Energy landscapes of receptor-ligand bonds explored with dynamic force spectroscopy. Nature. 397 (6714), 50-53 (1999).
- Hinterdorfer, P., Dufrêne, Y. F. Detection and localization of single molecular recognition events using atomic force microscopy. Nature Methods. 3 (5), 347-355 (2006).
- Sawicka, A., Babataheri, A., et al. Micropipette force probe to quantify single-cell force generation: application to T-cell activation. Molecular Biology of the Cell. 28 (23), 3229-3239 (2017).
- Desprat, N., Guiroy, A., Asnacios, A. Microplates-based rheometer for a single living cell. Review of Scientific Instruments. 77 (5), 055111 (2006).
- Labernadie, A., Bouissou, A., et al. Protrusion force microscopy reveals oscillatory force generation and mechanosensing activity of human macrophage podosomes. Nature Communications. 5, 5343 (2014).
- Bouissou, A., Proag, A., et al. Protrusion force microscopy: A method to quantify forces developed by cell protrusions. Journal of Visualized Experiments. (136), 57636 (2018).
- Kronenberg, N. M., Liehm, P., et al. Long-term imaging of cellular forces with high precision by elastic resonator interference stress microscopy. Nature Cell Biology. 19 (7), 864-872 (2017).
- Basu, R., Whitlock, B. M., et al. Cytotoxic T cells use mechanical force to potentiate target cell killing. Cell. 165 (1), 100-110 (2016).
- Bufi, N., Saitakis, M., et al. Human Primary Immune Cells Exhibit Distinct Mechanical Properties that Are Modified by Inflammation. Biophysical Journal. 108 (9), 2181-2190 (2015).
- Goodnow, C. C., Crosbie, J., et al. Altered immunoglobulin expression and functional silencing of self-reactive B lymphocytes in transgenic mice. Nature. 334 (6184), 676-682 (1988).
- Butler, J. P., Tolić-Nørrelykke, I. M., Fabry, B., Fredberg, J. J. Traction fields, moments, and strain energy that cells exert on their surroundings. American Journal of Physiology. Cell Physiology. 282 (3), 595-605 (2002).
- . MBPPlab/TFM_v1: Software for Time dependent Traction Force Microscopy Available from: https://github.com/MBPPlab/TFM_v1 (2019)
- Tseng, Q., Duchemin-Pelletier, E., et al. Spatial organization of the extracellular matrix regulates cell-cell junction positioning. Proceedings of the National Academy of Sciences of the United States of America. 109 (5), 1506-1511 (2012).
- . ImageJ plugins by Qingzong TSENG Available from: https://sites.google.com/site/qingzongtseng/ (2019)
- Plotnikov, S. V., Sabass, B., Schwarz, U. S., Waterman, C. M. High-resolution traction force microscopy. Methods in Cell Biology. 123, 367-394 (2014).
- Schindelin, J., Arganda-Carreras, I., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Wang, J., Lin, F., et al. Profiling the origin, dynamics, and function of traction force in B cell activation. Science Signaling. 11 (542), (2018).
- Hui, K. L., Balagopalan, L., Samelson, L. E., Upadhyaya, A. Cytoskeletal forces during signaling activation in Jurkat T-cells. Molecular Biology of the Cell. 26 (4), 685-695 (2015).
- Bashour, K. T., Gondarenko, A., et al. CD28 and CD3 have complementary roles in T-cell traction forces. Proceedings of the National Academy of Sciences of the United States of America. 111 (6), 2241-2246 (2014).
- Gardel, M. L., Sabass, B., Ji, L., Danuser, G., Schwarz, U. S., Waterman, C. M. Traction stress in focal adhesions correlates biphasically with actin retrograde flow speed. The Journal of Cell Biology. 183 (6), 999-1005 (2008).
- Stricker, J., Sabass, B., Schwarz, U. S., Gardel, M. L. Optimization of traction force microscopy for micron-sized focal adhesions. Journal of Physics. Condensed Matter. 22 (19), 194104 (2010).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved