
Method Article
Microscopia a forza di trazione per studiare l'attivazione dei linfociti B
In questo articolo
Riepilogo
Qui presentiamo un protocollo utilizzato per eseguire esperimenti di microscopia a forza di trazione su cellule B. Descriviamo la preparazione di gel morbidi in poliacrilammide e la loro funzionalizzazione, nonché l'acquisizione dei dati al microscopio e un riepilogo dell'analisi dei dati.
Abstract
La microscopia a forza di trazione (TFM) consente la misurazione delle forze prodotte da una cellula su un substrato. Questa tecnica deduce le misurazioni della forza di trazione da un campo di spostamento osservato sperimentalmente prodotto da una cellula che tira su un substrato elastico. Qui, abbiamo adattato TFM per indagare la struttura spaziale e temporale del campo di forza esercitato dalle cellule B quando attivato dall'innesto dell'antigene del recettore cellulare B. La rigidità del gel, la densità delle perline e la funzionalizzazione delle proteine devono essere ottimizzate per lo studio di cellule relativamente piccole (~ 6 μm) che interagiscono con, e rispondono specificamente ai ligandi per i recettori della superficie cellulare.
Introduzione
Le cellule B sono le cellule produttrici di anticorpi del sistema immunitario. Per attivare la risposta immunitaria adattiva, acquisiscono prima l'antigene in una forma nativa (cioè non elaborata) attraverso uno specifico recettore chiamato recettore delle cellule B (BCR)1. Questo processo si verifica nella zona cellulare del linfonodo B. Anche se alcuni antigeni possono raggiungere la cellula B attraverso fluidi linfatici, la maggior parte degli antigeni, specialmente con un elevato peso molecolare (>70 kDa, che è la dimensione limite per i condotti linfatici) sono infatti presentati nella loro forma nativa sulla superficie di una cellula di presentazione dell'antigene (APC), tipicamente un macrofago del seno sottocapsulare o una cellula dendritica follicolare, attraverso la lectina o i recettori Fc (non specifici). Il contatto con questa cellula porta alla formazione di una sinapsi immunitaria in cui il BCR esercita forza sugli antigeni associati all'APC. L'associazione di un antigene al BCR avvia la segnalazione BCR, che può attivare meccanismi di generazione della forza. Queste forze potrebbero essere importanti per amplificare la segnalazione BCR, ma sono anche essenziali per le cellule B per estrarre e quindi internalizzare l'antigene.
Recenti studi hanno dimostrato che il BCR è effettivamente meccanosensibile2. Ad esempio, i substrati più rigidi suscitano una segnalazione BCR migliorata3. Inoltre, la forza generata alla sinapsi immunitaria tira su singoli BCR per sondare la sua affinità con l'antigene e quindi garantire la discriminazione di affinità4. È quindi interessante studiare la risposta meccanica delle cellule B alla presentazione dell'antigene e sezionare questa risposta in termini di tipo di recettori implicati (IgG/IgM)5, molecole di adesione (ligandi integrina) o in cellule farmacologicamente e geneticamente modificate (cioè silenziamento di una proteina a valle della segnalazione BCR o dinamica del citoscheletro)6.
Un metodo semplice per osservare la risposta di una cellula a un substrato di rigidità fisiologica e, allo stesso tempo, forze di studio esercitate sul substrato è la microscopia a forza di trazione (TFM). TFM consiste nell'osservare il campo di spostamento prodotto dalla cellula tirando su un substrato elastico. Originariamente la deformazione del gel è stata osservata attraverso le rughe dell'elastomero stesso mediante microscopia a contrasto di fase7, ma l'inserimento di microperline di fluorescenza come marcatori fiduciari ha permesso una migliore risoluzione e da allora è diventato lo standard8. Questo metodo è stato utilizzato per studiare la forza di trazione esercitata da cellule aderenti, tessuti e persino organoidi incorporati nei gel. Diverse varianti di TFM sono statesviluppate 9 tra cui, combinazione con microscopia a supersoluzione (cioè, STED10 o SRRF11),modifica dell'indice di rifrazione del gel per consentire la microscopia TIRF12,sostituzione delle perline con motivi nanostampati13e utilizzo di nanopillari al posto della superficie piatta14. Per una revisione completa di queste variazioni, vedere Colin-York etal.
Il protocollo qui presentato descrive una procedura per misurare le forze esercitate dalle cellule B su un substrato rivestito di antigene. Queste forze sono applicate sui ligandi (antigene) al fine di raggrupparli e successivamente estrarli dal substrato che presenta l'antigene. Abbiamo adattato il protocollo TFM standard per imitare la rigidità dei substrati fisiologici che presentano antigeni, le dimensioni e il rivestimento pertinente per le cellule B. Questo protocollo consente lo studio di più cellule contemporaneamente e può essere utilizzato in combinazione con tecniche di microscopia a fluorescenza e trattamenti chimici. Tuttavia, non mira a sondare le misurazioni della forza di una singola molecola, per le quali le pinzette ottiche16,le sonde ditensione molecolare 17,18,le sonde di forza biomembrana19e la microscopia a forza atomica20 sono tecniche più adatte. Rispetto ad altri metodi di misurazione della forza a singola cella (ad esempio, micropipette21 o micropiatte22)TFM consente la ricostruzione di una mappa completa delle forze esercitate alla sinapsi con una risoluzione di ~ 300 nm. Ciò è utile per identificare i modelli spazio-temporali nelle forze esercitate sulla superficie e, poiché il gel è compatibile con l'imaging confocale, per correlarli con il reclutamento di proteine specifiche (ad esempio, citoscheletro e proteine di segnalazione).
Sebbene TFM 3D sia possibile, non è compatibile con la rigidità e la configurazione che abbiamo utilizzato. Le deformazioni in 3D sono ottenibili da altre configurazioni più complesse come la microscopia a forza di protrusione (afm scansione di una membrana deformabile in cui le cellule sono placcate)23,24 e microscopia a sollecitazione di interferenza del risuonatore elastico (ERISM, un gel che funge da cavità di risinazione per la luce ed evidenziazione di deformazioni del substrato con precisione di pochi nanometri)25. Sebbene queste tecniche siano molto promettenti, non sono ancora state utilizzate nelle cellule B. Altri tipi di TFM, come sui nanopillari14, potrebbero essere usati per avere substrati più riproducibili. Tuttavia, questa geometria non è adattata alle cellule morbide poiché la cellula interpenetra i pilastri, il che complica l'analisi. Questo approccio è stato infatti utilizzato nelle cellule T per osservare la capacità della cellula di costruire strutture attorno ai pilastri26.
Nonostante la sua semplicità, TFM utilizzando gel di poliacrilammide consente l'osservazione simultanea di molte cellule e può essere implementato facilmente ed economicamente in qualsiasi laboratorio dotato di banco e un microscopio a epifluorescenza (anche se si consiglia un disco confocale / rotante).
Per imitare la rigidità fisiologica di un APC, abbiamo usato gel di poliacrilammide con una rigidità di ~ 500 Pa27 e funzionalizzato il gel con antigeni attivanti. In questo protocollo, abbiamo funzionalizzato la superficie del gel di poliacrilammide con lisozima all'uovo di gallina (HEL). Ciò consente la misurazione delle forze generate dalla stimolazione del BCR attraverso l'impegno del sito di legame dell'antigene. L'uso di questo antigene e delle cellule B specifiche hel dei topi MD4 garantisce una generazione di forza relativamente uniforme in risposta alla legatura dell'antigene28. Tuttavia, altre molecole (come l'anti-IgM per i topi B6) possono essere innesate sul gel, ma le forze generate in questi casi potrebbero essere più eterogenee e meno intense. Poiché le cellule B sono piccole celle (diametro ~6 μm), il numero di perline è stato ottimizzato per essere massimo ma ancora tracciabile. Per le celle di grandi dimensioni che esercitano forze ~kPa sui loro substrati, si possono ottenere risultati soddisfacenti usando perline relativamente sparse o eseguendo una semplice velocimetria dell'immagine delle particelle (PIV) per ricostruire il campo di deformazione. Tuttavia, per piccole cellule come i linfociti B che esercitano stress fino a ~50 Pa, è necessario l'uso del tracciamento di singole particelle (velocimetria di tracciamento delle particelle, PTV) per ottenere la precisione desiderata durante la ricostruzione del campo di deformazione. Per tracciare in modo affidabile le perline singolarmente, l'ingrandimento dell'obiettivo deve essere almeno 60x e la sua apertura numerica intorno a 1,3. Pertanto, i gel devono essere relativamente sottili (<50 μm), altrimenti le perline non sono visibili in quanto sono al di sopra della distanza di lavoro dell'obiettivo.
Il protocollo principale si compone di tre sezioni: preparazione del gel, funzionalizzazione del gel e imaging; altre due sezioni sono opzionali e sono dedicate alla quantificazione dell'estrazione dell'antigene e all'imaging delle cellule fluorescenti.
Protocollo
1. Preparazione del gel
- Sillanizzazione del supporto gel
- Attivare la coverlip o la piastra di Petri con fondo di vetro (che verrà utilizzata come supporto gel) con una lampada UV per 2 min (attendere 30 s prima dell'esposizione alla lampada UV per evitare l'esposizione all'ozono residuo).
- Silanizzare il piatto coverlip/glass-bottom utilizzando 200 μL amminopropiltrimetossisilano (APTMS) per 5 min. Questo preparerà il supporto per il legame covalente del gel.
- Lavare accuratamente il coperchio/piatto con fondo di vetro con acqua ultra pura.
- Asciugare il coperchio/piatto con fondo di vetro utilizzando l'aspirazione sottovuoto.
- Preparazione della coverslip da 18 mm utilizzata per appiattire il gel
- Per preparare i copripavimenti, prima mettili in un portascipelli in ceramica. Quindi mettere il portapavigli in un piccolo becher (50 mL) e versare il reagente siliconizzante (conservato a 4 °C, riutilizzabile) sulle copertine, assicurandosi di coprirli completamente.
- Coprire il becher con un foglio di alluminio e incubare per 3 minuti a temperatura ambiente. Durante l'attesa, riempire un grande becher (500 mL) con acqua ultra pura. Dopo 3 minuti di incubazione nel reagente siliconizzante, trasferire il portapazzole con coverlips al becher dell'acqua.
- Risciacquare accuratamente i copricapo con acqua ultra pura, asciugarli bene e tenere le salviette di carta. Per ottenere i migliori risultati, procedere immediatamente alla sezione successiva.
- Polimerizzazione del gel
- Per gel di 0,5 kPa, mescolare 75 μL di acrilammide al 40% con 30 μL di bisacrilammide (reticolare) al 2% e 895 μL di soluzione salina tamponata da fosfati (PBS). Questa premiscele può essere conservata fino a un mese a 4 °C.
- A 167 μL di premix in gel da 0,5 kPa, aggiungere l'1% (1,67 μL) di perline, vortice e sonicato per 5 minuti in un sonicatore da bagno (pulitore ad ultrasuoni da banco standard con potenza di 50-100 W e frequenza 40 kHz). Mantenere la miscela protetta dalla luce utilizzando un foglio di alluminio.
NOTA: La premisce non si polimerizza fino all'aggiunta dell'iniziatore (TEMED). - Per catalizzare la polimerizzazione, aggiungere l'1% (1,67 μL) del 10% w/v persolfato di ammonio (APS).
- Per avviare la polimerizzazione, aggiungere lo 0,1% (0,2 μL) N,N,N′,N′-Tetrametiletilenediammina (TEMED). Mescolare con una pipetta. Una volta aggiunti APS e TEMED, il gel si polimerizza rapidamente, quindi procedi rapidamente alla fusione del gel.
- Fusione gel
- Pipetta 9 μL di miscela di gel su ogni coverlip/piatto con fondo di vetro (goccia al centro, Figura 1A)
- Posizionare il coverslip silanizzato/idrofobico e appiattire il gel (Figura 1B). Utilizzando le forcep, premere il coverslip per assicurarsi che il gel si diffonda su tutta l'area del coverslip (Figura 1C) fino a quando non inizia a fuoriuscire.
- Invertire la piastra di coverlip/glass-bottom in una grande piastra di Petri e toccarla sulla panca per forzare le perline che vanno verso la superficie del gel (Figura 1D).
- Coprire con un foglio di alluminio e lasciare per 1 h per polimerizzare a temperatura ambiente in una camera umida (cioè, mettere un tessuto umido sopra il piatto per evitare l'evaporazione).
- Dopo 1 h, aggiungere PBS al campione per facilitare il rilascio di coverlip. Con attenzione, rimuovere il coverslip utilizzando un ago (il rivestimento con silani diversi dovrebbe consentire un facile peeling del coverslip dal gel, Figura 1E).
- Lasciare il gel in PBS.
NOTA: I gel possono ora essere conservati in PBS a 4 °C per 5-7 giorni, ma si consiglia di utilizzarli entro 48 ore.
2. Funzionalizzazione del gel
- Preparare la soluzione di esanoato sulfosuccinimidil 6-(4'-azido-2'-nitrofenilamino) (Sulfo SANPAH) a 0,5 mg/mL in tampone HEPES da 10 mM. Questo può essere conservato a 4 °C coperto con foglio di alluminio per un massimo di una settimana.
- Aspirare il PBS dai gel.
- Aggiungere 150 μL di Sulfo SANPAH al gel a temperatura ambiente (Figura 1F).
- Esporre il gel al trattamento UV per 2 minuti per fotoattivare i siti di Sulfo SANPAH e farlo attaccare alla superficie del gel.
- Lavare con PBS tre volte(Figura 1G).
- Ripetere i passaggi da 2,2 a 2,5.
- Aggiungere 250 μL di HEL (100 μg/mL) a ciascun gel e incubare durante la notte in una camera umida a 4 °C durante la notte mantenendosi coperti con un fogliodi alluminio (Figura 1H).
- Rimuovere l'antigene HEL e lavare con PBS tre volte.
NOTA: HEL agisce sia come antigene che come molecola di adesione. Può essere sostituito da altre molecole che si legano al recettore (ad esempio, un IgM anti-topo, albumina sierica bovina, ovalbumina) o mescolato con ligandi integrina (ad esempio, ICAM1 legato a LFA1). Se necessario, l'estrazione dell'antigene può essere osservata con una versione fluorescente dell'HEL (ottenuta macchiando la molecola con un kit di etichettatura proteica, vedere il passaggio 4). Si noti che una data concentrazione alla rinfusa potrebbe non produrre la stessa concentrazione superficiale sul gel del vetro: questo deve essere quantificato con colorazione secondaria se è necessario un confronto diretto.
3. Caricamento cellulare e imaging
- Prima dell'imaging, rimuovere il PBS dai gel e aggiungere 500 μL di supporto cellulare B (RPMI 1640, 10% siero fetale di vitello decomplementato, 1% di penicillina-streptomicina, 2% piruvato di sodio, 50uM mercaptoetanolo e 1X aminoacidi non essenziali) e lasciarli equilibrare a RT.
- Preparazione cellulare
- Purificare le cellule B primarie dalla milza secondo un protocollo di selezione negativo (vedere Tabella dei materiali). La resa tipica finale delle cellule B è di circa 1 x 107 cellule. Concentratelo a 3 x 106 cellule/mL in mezzo cellulare B (RPMI-1640 integrato con siero fetale di vitello al 10%, 1% penicillina-streptomicina, 0,1% mercaptoetanolo e 2% piruvato di sodio).
- Conservare le celle in base alle esigenze fino a 6 ore a 4 °C.
- Mantenere le celle a 37 °C per 30 minuti prima dell'acquisizione dell'immagine.
- Imaging
- Utilizzare un microscopio confocale con controllo termico e (possibilmente)CO 2.
NOTA: Indipendentemente dal fatto che venga utilizzato un microscopio confocale o a disco rotante, è importante utilizzare un obiettivo/foro stenopeica che consenta a un pixel size <200 nm di tracciare comodamente le perline in fase di analisi (ad esempio, 60x, NA 1.3). La microscopia a epifluorescenza può anche essere utilizzata, tuttavia fornisce un rapporto segnale/rumore inferiore e può rendere più difficile il tracciamento delle perline individuali. - Due strati principali di perline appariranno sul fondo e sulla parte superiore del gel. Concentrati sul piano gel.
NOTA: Un bel gel apparirà come un cielo stellato, con perline distribuite approssimativamente uniformemente sullo stesso piano. - Programmare l'acquisizione per 30 min con una frequenza fotogrammi di 5 s (questo è adattabile alle esigenze dell'esperimento, ad esempio, acquisire altri colori, acquisire stack z, ecc.)
- Aspirare il supporto dal gel, lasciando circa 200 μL di mezzi sul gel. Posizionare il gel al microscopio e trovare lo strato superficiale di perline e una bella area uniforme sul gel.
- Aggiungere 80 μL di cellule (evitare di toccare il gel per mantenere la messa a fuoco).
- Assicurarsi che la messa a fuoco sia ancora corretta e che le cellule possano essere viste scendere nell'area (sotto la luce trasmessa). Avviare l'acquisizione prima che le cellule raggiungano il gel.
- In caso di contatto accidentale con gel, vibrazioni o deriva di messa a fuoco, regolare la messa a fuoco.
NOTA: È fondamentale raccogliere un'immagine del gel rilassato e questa può essere qualsiasi immagine scattata prima dell'arrivo delle cellule sul gel.
- Utilizzare un microscopio confocale con controllo termico e (possibilmente)CO 2.
4. Esperimento di estrazione HEL fluorescente
- Preparare l'HEL fluorescente legando un colorante fluorescente (di un colore diverso da quello delle perline come Alexa 555), vedere il Tavolo dei Materiali.
- Nel passaggio 2.7, sostituire l'HEL convenzionale con l'HEL fluorescente.
- Acquisire immagini con impostazioni di illuminazione basse o bassa frequenza fotogrammi (ad esempio, 2 fotogrammi al minuto) per evitare lo sbiancamento delle foto.
- Per quantificare l'estrazione HEL, calcolare l'intensità integrata sull'area cellulare per ogni fotogramma I(t) corretto e normalizzato dall'intensità I(0) del fotogramma 0 secondo la formula:

NOTA: L'antigene coniugato con un fluorofore non è visibile (probabilmente a causa della tempra del fluorofore sulla superficie del gel), ma la sua presenza sul gel può essere verificata con un anticorpo secondario anti-HEL e fluorescente. Si può verificare che il fluorofo sia effettivamente fluorescente quando staccato togliendolo dal gel con un coverslip rivestito con anti-HEL e rivelandolo con un anticorpo fluorescente secondario (sul coverslip)6. Il segnale dell'antigene estratto è molto fioco e talvolta viene mascherato dalla perdita delle perline. Se si è interessati solo all'estrazione dell'antigene, si consiglia di preparare il gel senza perline (saltare i passaggi 1.3.2 e 1.4.3).
5. Imaging a fluorescenza
- Ottenere cellule B fluorescenti purificando la cellula B dalle milza dei topi geneticamente modificati come fatto per il tipo selvatico (ad esempio, dai topi Lifeact-GFP o Myosin II GFP).
- Per l'imaging di celle fluorescenti, utilizzare (se possibile) un microscopio a disco rotante con un obiettivo di immersione in acqua a lunga distanza 40x-100x.
- Mantenere bassa la durata dell'esposizione e la frequenza dei fotogrammi per evitare lo sbiancamento.
NOTA: La funzione di diffusione del punto in Z è altamente degradata dalla presenza del gel, quindi suggeriamo di utilizzare un obiettivo di immersione in acqua. La microscopia verticale viva con obiettivi di immersione in acqua soffre di forti aberrazioni sferiche indotte dalla presenza della cellula (sferica) (e del nucleo cellulare) nel percorso di emissione.
6. Analisi
NOTA: L'analisi dei dati viene in generale eseguita correggendo prima l'intera pila per la deriva, trovando le perline in ogni fotogramma, monitorandone i movimenti rispetto a un frame di riferimento (preso in assenza di celle), interpolando il campo di spostamento e invertendo il problema per ottenere la sollecitazione utilizzando la trasformata di Fourier29. A tal fine, si consiglia di utilizzare una combinazione di programmi ImageJ Macro e MATLAB scaricabili da un repository online30.
- Aprire il filmato in ImageJ come pila di immagini
- Eseguire la macro "Crop_and_save.ijm"
- Selezionare le aree di interesse (ROI) con lo strumento "Rettangolo" e aggiungerle all'elenco del ROI utilizzando la chiave 't'.
- Quando ritaglia la cella, assicurati di includere un'area di almeno 5-10 pixel di perline immobili. Escludere dall'analisi le celle troppo vicine ai limiti o ad altre celle. Al termine fare clic su 'OK'.
- La macro propone una maschera della cella: se questo è soddisfacente clicca su "OK". Se non soddisfacente, fare clic su "Non ok" e quindi selezionare manualmente una regione chiusa con qualsiasi strumento di selezione (ad esempio "Mano libera" o "Ovale") e fare clic su "Continua".
- Aprire MATLAB ed eseguire "TFM_v1.m".
- Inserire i parametri richiesti: in particolare controllare le proprietà dell'immagine (dimensione dei pixel, intervallo di tempo di acquisizione) e le proprietà del gel (young modulus E, rapporto Poisson).
- L'immagine di riferimento è impostata per essere la prima per impostazione predefinita. Impostarlo su un altro frame, se necessario, o impostarlo su "0" per caricare un file esterno.
- Individuare gli output del software nella stessa directory del file originale (per una descrizione vedere la User_notice.pdf file). Ciò include una traccia preliminare delle perline ("FILENAME.fig"), un grafico dell'energia contrattile nel tempo ("FILENAME_energy.fig"), una tabella di diverse quantità integrate sopra la cella (energia, area, momenti, ecc.) "FILENAME_finaltable.mat", una struttura contenente il campo di spostamento e forza, film del tallone, campo di spostamento, stress ed energia (che può essere aperto con qualsiasi lettore avi).
NOTA: Nei parametri di input, la "dimensione della finestra" è la finestra su cui viene interpolato lo spostamento, da cui la risoluzione finale del campo di sollecitazione e spostamento. Questo è impostato su alcuni (per impostazione predefinita quattro) pixel. Non è consigliabile ridurlo in quanto aumenterebbe artificialmente la risoluzione interpolando le regioni in cui non ci sono perline.
Risultati
Date le dimensioni delle cellule, gli algoritmi che estraggono la mappa di spostamento delle perline attraverso tecniche correlative (come la velocimetria dell'immagine delle particelle) non sono in generale molto precisi. Tuttavia, a seconda del grado di risoluzione richiesto, si possono facilmente ottenere risultati qualitativi utilizzando un plug-in Fiji / ImageJgratuito 31,32. Sebbene questo approccio sia sufficiente per confrontare condizioni stimolanti rispetto a quelle non stimolanti, per un'analisi approfondita si consiglia di utilizzare il nostro software scaricabile da un repository online30, che tiene traccia delle perline singolarmente e fornisce la mappa del campo di spostamento in un dato momento come interpolazione dei singoli spostamenti di perline33. A questo punto sono possibili diverse quantificazioni. Ad esempio (supponendo che lo spostamento sia causato solo da sollecitazioni tangenziali alla superficie del gel) il software fornisce anche la sollecitazione in ogni punto causando quella specifica mappa di spostamento. Questo è un tipo di "problema di inversione": lo spostamento in un certo punto dipende dalla somma di tutte le forze applicate in tutti gli altri punti. L'"algoritmo di inversione" tiene conto dei parametri fisici del substrato: la sua rigidità (modulo giovane) e il rapporto di Poisson. Gli algoritmi diretti sono in genere molto accurati ma computazalmente costosi. Gli algoritmi basati sulla trasformata di Fourier, come il nostro, eseguono essenzialmente una deconvoluzione nello spazio di Fourier e sono più efficienti ma soggetti ad alcuni errori (principalmente dovuti alla fase di interpolazione). Questi algoritmi richiedono generalmente l'ottimizzazione di un parametro che impedisce che piccoli spostamenti locali (e potenzialmente artefatti) diventino troppo rilevanti nel calcolo del campo di sollecitazione (parametro di regolarizzazione di Tikhonov8,29; variabile "Regolarizzazione" nella finestra di dialogo; qui in genere abbiamo impostato uguale a 5 x 10-19). Per interpretazioni e analisi più avanzate, come correlazioni spazio-temporali, movimenti locali, correlazioni con canali fluorescenti, si consiglia di collaborare con esperti del settore. Per una revisione dei metodi computazionali si veda Schwarz etal.
Come accennato in precedenza, le immagini corrette delle perline sembrano un "cielo stellato", una distribuzione uniforme e casuale dei punti luminosi (Figura 2A). I dati e l'analisi non sono affidabili quando il numero di perline è troppo basso (Figura 2B) o l'immagine è fuori fuoco (Figura 2C). Una volta che le cellule B si sono depositate sulla superficie del gel, le perline sotto le cellule iniziano a muoversi a causa della forza di trazione esercitata dalla cellula sul gel. I telai per i quali le perline non sono tracciabili devono essere scartati.
Come controllo, è possibile osservare a occhio il movimento delle perline confrontando il "quadro di riferimento", tipicamente quello che precede il primo contatto della cellula con il substrato. I risultati approssimativi possono essere ottenuti dal tracciamento delle singole particelle (ad esempio, Trackmate, Fiji 34) come fatto nella figura 3A. L'analisi fornisce una segmentazione delle perline nell'immagine di riferimento ("FILENAME.fig") come controllo.
Con il software che proponiamo, si possono ottenere lo spostamento (Figura 3B) e il campo di sollecitazione (il vettore della sollecitazione locale ad ogni pixel e ogni punto di tempo ottenuto per inversione dal campo di spostamento, Figura 3C). Il prodotto scalare dei campi di spostamento e forza integrati nell'area della cella fornisce il lavoro totale esercitato dalla cellula sul substrato (Figura 4A). Questo calcolo richiede la maschera della cella introdotta nel passaggio 6.2 del protocollo.
Per confrontare due condizioni biologiche (come l'attivazione del substrato HEL rispetto al substrato non attivante BSA, o il tipo selvaggio rispetto al knock-out) è utile calcolare la curva media (Figura 4B) o, ancora più sinteticamente, un valore medio negli ultimi punti di tempo (20 minuti) in cui l'energia raggiunge un plateau(Figura 4C). Quando le informazioni spaziali delle forze sono rilevanti è possibile confrontare singoli punti di tempo di ciascuna condizione (Figura 4D). Fare riferimento a Kumari etal.
Un esempio di lapse del tempo di estrazione dell'antigene a fluorescenza è mostrato nella figura 5A:l'aspetto progressivo dei segnali di fluorescenza al distacco di antigene indicato dalla sinapsi dal gel. La curva di estrazione media con il suo intervallo di confidenza (errore standard della media) su 15 celle è illustrata nella figura 5B.

Figura 1: Schema della preparazione del gel e della sua funzionalizzazione. I passaggi sono descritti nel protocollo. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 2: Tre esempi di immagini di perline di diverse qualità. (A) Esempio di immagine del tallone con il corretto rapporto segnale/rumore e la densità corretta. (B) Esempi di immagini con un numero troppo insufficiente di perline e (C) fuori dal piano di messa a fuoco. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 3: Elaborazione delle immagini per estrarre il campo di forza. (A) Esempio di immagine delle perline (contorno della cella in bianco, estratta dall'immagine di trasmissione), tracciamento delle perline al tempo t = 5 min (sovrapposizione rossa) e spostamento (frecce) rispetto al tempo t = 0 min (barra di scala 5 μm). (B) Campo di spostamento interpolato (rappresentato come faretra vettoriale e mappa di magnitudine, le frecce sono proporzionali allo spostamento [nm]; vedi la barra dei colori a destra); in basso: un'immagine più liscia della magnitudine (ottenuta per interpolazione con una funzione bicubica). (C) Campo di sollecitazione dal campo di spostamento nel pannello B (rappresentato come faretra vettoriale e mappa di magnitudine; le frecce sono proporzionali alla sollecitazione di taglio [Pa]; vedere la barra di colore a destra); in basso: un'immagine più liscia della magnitudine (ottenuta per interpolazione con una funzione bicubica). Clicca qui per visualizzare una versione più grande di questa figura.
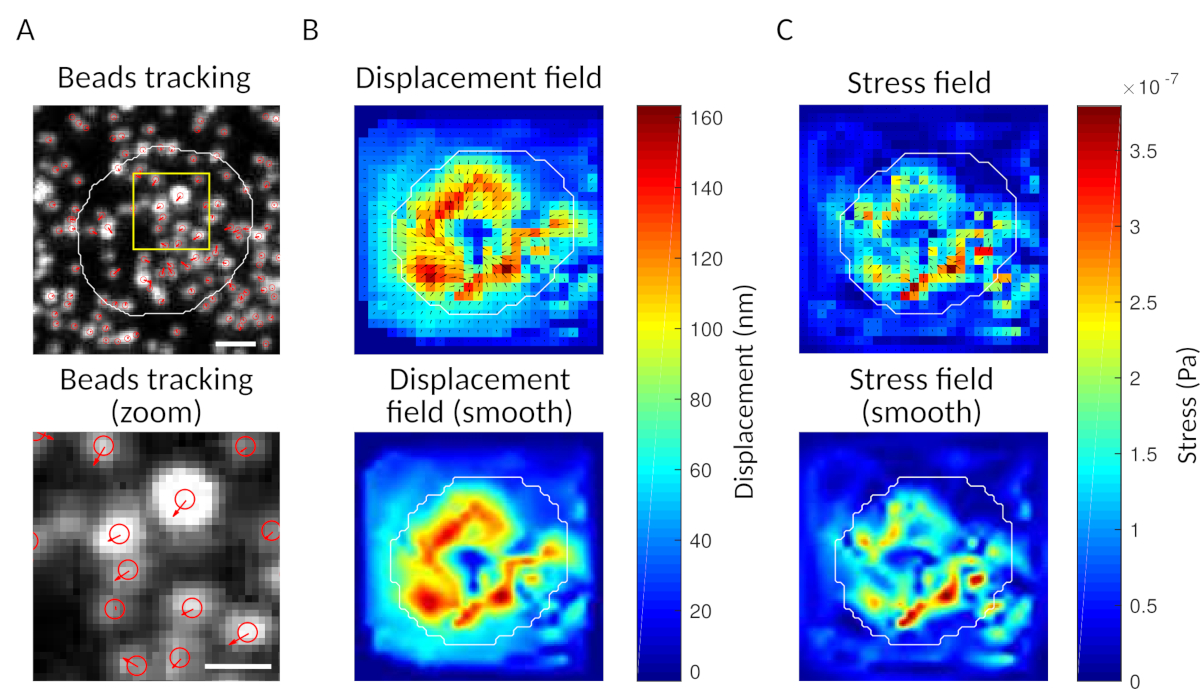{kind=link}

Figura 4: Esempio di informazioni che possono essere estratte dai campi di forza e spostamento. (A) Esempio di evoluzione dell'energia nel tempo per una singola cellula: una fase di plateau (evidenziata in grigio) si presenta dopo circa 10 min. (B) Confronto delle curve energetiche medie e (C) dei livelli relativi di plateau per 65 celle placcate su gel rivestito HEL (attivante) e 35 celle su gel rivestito BSA (non attivante) (sono mostrate gamme interquartili ± mediane, il test Mann-Whitney è stato utilizzato per il significato statistico). (D) Mappe di colore time-lapse di sollecitazione per hel e controllo condizione BSA; vengono mostrate sia le trame di magnitudo che di faretra. Queste immagini sono state adattate da Kumari etal. Clicca qui per visualizzare una versione più grande di questa figura.
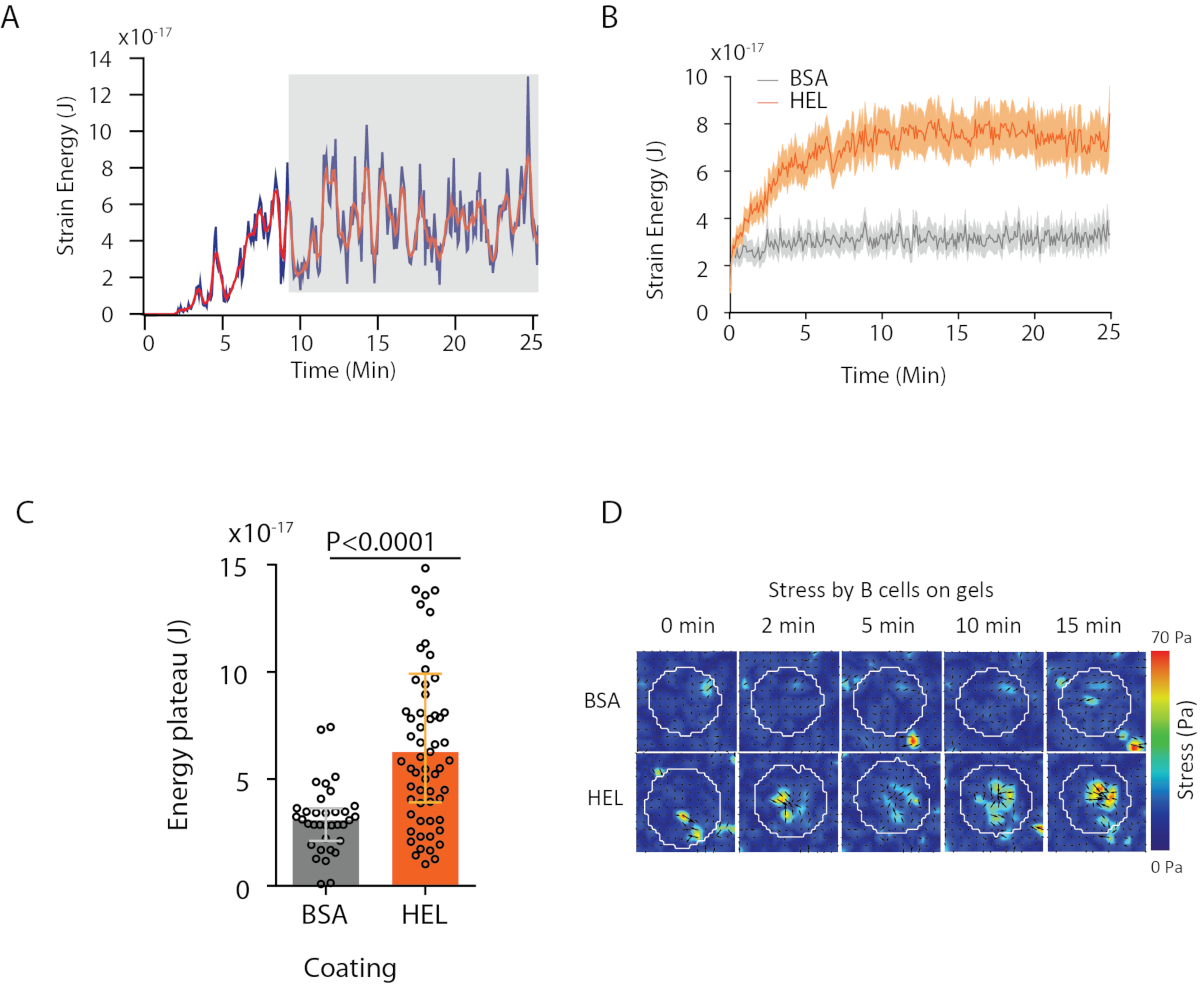{kind=link}

Figura 5: Esempio di esperimenti con antigene fluorescente. (A) Time lapse dell'estrazione di HEL fluorescente (sotto: percentuale del massimo, barra di scala = 3μm). (B) Raccolta dell'antigene nel tempo (± SEM, n = 15). Queste immagini sono state adattate da Kumari etal. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}
Discussione
Il metodo TFM qui descritto consente lo studio sistematico delle capacità meccaniche attive delle cellule B. Nel contesto delle cellule B, questo è correlato alla capacità di estrarre e internalizzare l'antigene. Rispetto ad altri metodi TFM, il protocollo qui presentato è semplice e piuttosto riproducibile: la rigidità, misurata per indentazione di una microsfera di vetro e utilizzando il modello Hertz, è compresa tra 400 e 600 Pa. Protocolli simili sono stati utilizzati con successo non solo per le celle B35 ma anche per le celle T36. Rispetto ai nanopillari (usati anche per i linfociti T37) fornisce una superficie piatta omogenea, quindi i risultati sono più facili da interpretare poiché l'interazione del gel è principalmente vincolata ad essere tangenziale alla superficie.
Il protocollo che abbiamo descritto dà accesso alla dinamica spaziotemporale delle forze esercitate dalle cellule B su substrati che presentano antigeni. A livello spaziale questo fornisce informazioni sulla localizzazione delle forze, e in combinazione con la microscopia a fluorescenza, consente allo sperimentatore di correlare le forze locali con la presenza di molecole specifiche (cioè componenti del citoscheletro o della cascata di segnalazione BCR). A livello temporale, è possibile integrare quantità (come energia totale o sollecitazione totale) per fornire un valore per punto di tempo e ridurre il rumore. Ciò consente l'osservazione dell'evoluzione della forza di trazione nel tempo (crescita e plateau) e della presenza di modelli pulsatili.
Gli aspetti sperimentali critici per l'analisi sono descritti come segue. — densità cellulare: per eseguire un'analisi corretta, le cellule devono essere sufficientemente separate. Consideriamo una cellula aalizzabile se ha intorno ad essa una regione vuota delle sue dimensioni. — immagine di trasmissione: è consigliabile raccogliere almeno un'immagine di trasmissione delle cellule durante l'esperimento da utilizzare come maschera nell'analisi. (iii) Numero di perline nell'immagine: suggeriamo di analizzare solo le immagini in cui il numero di perline nella sinapsi è compreso tra 30 e 200 (cioè 1-8 perline/μm²). Densità più basse non consentono un'adeguata ricostruzione dello spostamento della mappa. Le alte densità di perline rendono inaffidabile il tracciamento di singole particelle. — il numero di perline deve essere costante durante l'esperimento; tuttavia, le fluttuazioni possono verificarsi a causa della piccola variabilità nelle condizioni di imaging (specialmente nelle perline troppo vicine l'una all'altra). La deriva di messa a fuoco, se si verifica, deve essere corretta e i fotogrammi problematici devono essere scartati. ( v) Qualità del gel: i gel con troppe crepe, la variabilità nella distribuzione delle perline o i gel troppo spessi devono essere scartati. (vi) A seconda del tipo di cella, dopo esposizioni ripetute, le cellule in ritardo (>300 fotogrammi) possono subire effetti fototossici. Si consiglia di eseguire il programma su una maschera priva di celle come "linea di base" da confrontare con i dati. Ciò fornisce una grandezza del livello di rumore esclusivamente a causa delle condizioni sperimentali.
I gel utilizzati per misurare la forza di trazione nell'adesione classica consentono lo studio dei processi che si verificano all'adesione focale (flussi di actina e reclutamento di molecole di segnalazione) - i punti in cui le forzevengono applicate 38,39. Tuttavia, le forze alla sinapsi non vengono applicate attraverso adesioni focali. Il modello spaziotemporale di generazione della forza presso la sinapsi immunitaria della cellula B non è stato studiato quantitativamente utilizzando questo metodo fino a poco tempo fa. Utilizzando TFM, abbiamo osservato per la prima volta, il patterning della forza alla sinapsi immunitaria della cellula B, come presentato nel nostro recentestudio 6,aprendo prospettive incoraggianti nello studio dei linfociti.
In particolare, questo metodo utilizza un'immagine scattata prima dell'arrivo delle cellule sul gel come immagine di riferimento per il calcolo della forza. I soliti protocolli TFM suggeriscono di prendere l'immagine di riferimento alla fine dell'esperimento, dopo aver staccato le cellule con tripina; questo permette allo sperimentatore di cercare una regione ricca di cellule. Sebbene ciò sia possibile anche qui, la tripsiderina è piuttosto inefficiente nel staccare le cellule B dal gel rivestito di antigene, è necessario attendere a lungo il distacco e il rischio di modifica e movimenti del gel (che rendono l'intero set di dati inspiegabile) è più alto.
Il metodo qui presentato è flessibile e può essere applicato per studiare l'effetto di altri segnali alla sinapsi immunitaria in quanto consente di innestare altre proteine sulla superficie del gel (ad esempio, sono stati testati ligandi integrina e immunoglobuline) e persino antigene fluorescente (vedi sezione 4). Inoltre, le cellule rimangono accessibili allo sperimentatore per il trattamento farmacologico e le perturbazioni locali. Infine, il metodo è anche compatibile con l'imaging di celle fisse. Per queste osservazioni, si consiglia di fare il gel su un coverslip, macchiare le cellule, incollare il coverslip su una diapositiva e solo allora aggiungere supporti di montaggio e un altro coverslip. L'osservazione verrà quindi eseguita con il gel in cima per evitare la degradazione dell'immagine attraverso il gel.
Possibili insidie sono la variabilità del gel nella polimerizzazione e nel rivestimento. I problemi di polimerizzazione sono dovuti principalmente alla qualità dell'iniziatore/catalizzatore. Inoltre, il gel può gonfiarsi, soprattutto se non utilizzato subito dopo il montaggio. Questo problema non sembra influenzare drammaticamente le proprietà meccaniche del gel, ma può rendere lo strato di perline irraggiungibile per l'obiettivo, rendendo efficacemente il gel inutile. Si consiglia di preparare gel extra per ogni condizione quando appare questo problema. Potrebbe esserci anche una certa variabilità nel rivestimento, ed è fondamentale avere sanpah Sulfo appena diluito.
In conclusione, abbiamo descritto un metodo semplice, economico e riproducibile per misurare le forze esercitate dalle cellule B alla sinapsi immunologica quando attivate dal ligando BCR. Può essere adattato per studiare la reazione ad altri ligandi e altri tipi di linfociti (cellule B di memoria, cellule T, ecc.) con l'uso del ligando del recettore corretto.
Divulgazioni
Gli autori non hanno nulla da rivelare.
Riconoscimenti
Gli autori ringraziano M. Bolger-Munro per la lettura critica e riconoscono il Nikon Imaging Center@CNRS-InstitutCurie e PICT-IBiSA, Institut Curie, Parigi, membro dell'infrastruttura di ricerca nazionale France-BioImaging, per il supporto nell'acquisizione di immagini e nel Curie Animal Facility. PP è stato supportato dal CNRS. AK e JP sono stati supportati dalla borsa di dottorato Paris Descartes e dall'Ecole Doctorale FIRE- Programme Bettencourt. Questo progetto è stato finanziato con sovvenzioni a PP (ANR-10-JCJC-1504-Immuphy) e AMLD (ANR-PoLyBex-12-BSV3-0014-001, ERC-Strapacemi-GA 243103).
Materiali
| Name | Company | Catalog Number | Comments |
| 3-aminopropyltrimethoxysilane (APTMS) | Sigma-Aldrich | 281778 | Store aliquoted, protected from humidity |
| 40% Acrylamide Solution | Biorad | 1610140 | |
| Alexa555 microscale protein labeling kit | Molecular Probes | A30007 | |
| Ammonium Persulfate (APS) | Sigma-Aldrich | A3678 | |
| B cell Isolation Kit, Mouse | Miltenyi Biotec | 130-090-862 | |
| B-mercaptoethanol | Gibco | 31350-010 | |
| 2% Bis Solution | Biorad | 161-0142 | |
| Bovine Serum Albumin (BSA) | Euromedex | 04-100-812-C | |
| Coverslip 18mm | VWR | 631-1580 | |
| Fetal calf serum | PAA | A15-151 | Decomplemented (40min @56°C) |
| Fluorodishes FD35 | World Precision Instruments, Inc | FD35100 | |
| Fluosphere: carboxylate-modified, 0.2um, dark red | Molecular Probes | F8807 | |
| Hen Egg Lysozyme | Sigma-Aldrich | L6876 | Stocked in aliquote 100mg/ml |
| MEM Non-Essential Amino Acids Solution (100X) | Thermofisher/Gibco | 11140035 | |
| N,N,N',N'-tetrametiletilendiammine (TEMED) | Euromedex | 50406-B | |
| PBS (Phosfate Buffer Saline) | Gibco | 10010-015 | |
| Penicillin–streptomycin | Gibco | 15140-010 | |
| RMPI 1640 – Glutamax I | Thermofisher | 61870-010 | |
| Sigmacote | Sigma-Aldrich | SL2 | |
| Sodium pyruvate | Gibco | 11360-039 | |
| sulfosuccinimidyl 6-(4'-azido-2'-nitrophenylamino)hexanoate (Sulfo-SANPAH) | Thermo Scientific | 22589 |
Riferimenti
- Yuseff, M. -I., Pierobon, P., Reversat, A., Lennon-Duménil, A. -M. How B cells capture, process and present antigens: a crucial role for cell polarity. Nature Reviews. Immunology. 13 (7), 475-486 (2013).
- Spillane, K. M., Tolar, P. B cell antigen extraction is regulated by physical properties of antigen-presenting cells. The Journal of Cell Biology. 216 (1), 217-230 (2017).
- Shaheen, S., Wan, Z., et al. Substrate stiffness governs the initiation of B cell activation by the concerted signaling of PKCβ and focal adhesion kinase. eLife. 6, (2017).
- Natkanski, E., et al. B cells use mechanical energy to discriminate antigen affinities. Science. 340 (6140), 1587-1590 (2013).
- Wan, Z., Chen, X., et al. The activation of IgM- or isotype-switched IgG- and IgE-BCR exhibits distinct mechanical force sensitivity and threshold. eLife. 4, (2015).
- Kumari, A., Pineau, J., et al. Actomyosin-driven force patterning controls endocytosis at the immune synapse. Nature Communications. 10 (1), 2870(2019).
- Dembo, M., Wang, Y. L. Stresses at the cell-to-substrate interface during locomotion of fibroblasts. Biophysical Journal. 76 (4), 2307-2316 (1999).
- Sabass, B., Gardel, M. L., Waterman, C. M., Schwarz, U. S. High resolution traction force microscopy based on experimental and computational advances. Biophysical Journal. 94 (1), 207-220 (2008).
- Schwarz, U. S., Soiné, J. R. D. Traction force microscopy on soft elastic substrates: A guide to recent computational advances. Biochimica et Biophysica Acta. 1853 (11), Pt B 3095-3104 (2015).
- Colin-York, H., Shrestha, D., et al. Super-Resolved Traction Force Microscopy (STFM). Nano Letters. 16 (4), 2633-2638 (2016).
- Stubb, A., Laine, R. F., Guzmán, C., Henriques, R., Jacquemet, G., Ivaska, J. Fluctuation-Based Super-Resolution Traction Force Microscopy. BioRxiv. , (2019).
- Gutierrez, E., Tkachenko, E., et al. High refractive index silicone gels for simultaneous total internal reflection fluorescence and traction force microscopy of adherent cells. Plos One. 6 (9), 23807(2011).
- Bergert, M., Lendenmann, T., et al. Confocal reference free traction force microscopy. Nature Communications. 7, 12814(2016).
- Schoen, I., Hu, W., Klotzsch, E., Vogel, V. Probing cellular traction forces by micropillar arrays: contribution of substrate warping to pillar deflection. Nano Letters. 10 (5), 1823-1830 (2010).
- Colin-York, H., Fritzsche, M. The future of traction force microscopy. Current Opinion in Biomedical Engineering. 5, 1-5 (2018).
- Feng, Y., et al. Mechanosensing drives acuity of αβ T-cell recognition. Proceedings of the National Academy of Sciences of the United States of America. 114 (39), 8204-8213 (2017).
- Spillane, K. M., Tolar, P. DNA-Based Probes for Measuring Mechanical Forces in Cell-Cell Contacts: Application to B Cell Antigen Extraction from Immune Synapses. Methods in Molecular Biology. 1707, 69-80 (2018).
- Stabley, D. R., Jurchenko, C., Marshall, S. S., Salaita, K. S. Visualizing mechanical tension across membrane receptors with a fluorescent sensor. Nature Methods. 9 (1), 64-67 (2011).
- Merkel, R., Nassoy, P., Leung, A., Ritchie, K., Evans, E. Energy landscapes of receptor-ligand bonds explored with dynamic force spectroscopy. Nature. 397 (6714), 50-53 (1999).
- Hinterdorfer, P., Dufrêne, Y. F. Detection and localization of single molecular recognition events using atomic force microscopy. Nature Methods. 3 (5), 347-355 (2006).
- Sawicka, A., Babataheri, A., et al. Micropipette force probe to quantify single-cell force generation: application to T-cell activation. Molecular Biology of the Cell. 28 (23), 3229-3239 (2017).
- Desprat, N., Guiroy, A., Asnacios, A. Microplates-based rheometer for a single living cell. Review of Scientific Instruments. 77 (5), 055111(2006).
- Labernadie, A., Bouissou, A., et al. Protrusion force microscopy reveals oscillatory force generation and mechanosensing activity of human macrophage podosomes. Nature Communications. 5, 5343(2014).
- Bouissou, A., Proag, A., et al. Protrusion force microscopy: A method to quantify forces developed by cell protrusions. Journal of Visualized Experiments. (136), 57636(2018).
- Kronenberg, N. M., Liehm, P., et al. Long-term imaging of cellular forces with high precision by elastic resonator interference stress microscopy. Nature Cell Biology. 19 (7), 864-872 (2017).
- Basu, R., Whitlock, B. M., et al. Cytotoxic T cells use mechanical force to potentiate target cell killing. Cell. 165 (1), 100-110 (2016).
- Bufi, N., Saitakis, M., et al. Human Primary Immune Cells Exhibit Distinct Mechanical Properties that Are Modified by Inflammation. Biophysical Journal. 108 (9), 2181-2190 (2015).
- Goodnow, C. C., Crosbie, J., et al. Altered immunoglobulin expression and functional silencing of self-reactive B lymphocytes in transgenic mice. Nature. 334 (6184), 676-682 (1988).
- Butler, J. P., Tolić-Nørrelykke, I. M., Fabry, B., Fredberg, J. J. Traction fields, moments, and strain energy that cells exert on their surroundings. American Journal of Physiology. Cell Physiology. 282 (3), 595-605 (2002).
- MBPPlab/TFM_v1: Software for Time dependent Traction Force Microscopy. , Available from: https://github.com/MBPPlab/TFM_v1 (2019).
- Tseng, Q., Duchemin-Pelletier, E., et al. Spatial organization of the extracellular matrix regulates cell-cell junction positioning. Proceedings of the National Academy of Sciences of the United States of America. 109 (5), 1506-1511 (2012).
- ImageJ plugins by Qingzong TSENG. , Available from: https://sites.google.com/site/qingzongtseng/ (2019).
- Plotnikov, S. V., Sabass, B., Schwarz, U. S., Waterman, C. M. High-resolution traction force microscopy. Methods in Cell Biology. 123, 367-394 (2014).
- Schindelin, J., Arganda-Carreras, I., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Wang, J., Lin, F., et al. Profiling the origin, dynamics, and function of traction force in B cell activation. Science Signaling. 11 (542), (2018).
- Hui, K. L., Balagopalan, L., Samelson, L. E., Upadhyaya, A. Cytoskeletal forces during signaling activation in Jurkat T-cells. Molecular Biology of the Cell. 26 (4), 685-695 (2015).
- Bashour, K. T., Gondarenko, A., et al. CD28 and CD3 have complementary roles in T-cell traction forces. Proceedings of the National Academy of Sciences of the United States of America. 111 (6), 2241-2246 (2014).
- Gardel, M. L., Sabass, B., Ji, L., Danuser, G., Schwarz, U. S., Waterman, C. M. Traction stress in focal adhesions correlates biphasically with actin retrograde flow speed. The Journal of Cell Biology. 183 (6), 999-1005 (2008).
- Stricker, J., Sabass, B., Schwarz, U. S., Gardel, M. L. Optimization of traction force microscopy for micron-sized focal adhesions. Journal of Physics. Condensed Matter. 22 (19), 194104(2010).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati