
Method Article
人诱导多能干细胞体外生成体细胞衍生物的研究
摘要
我们在这里提出了一个方案, 分化为每个体细胞衍生物 (肌细胞, 硬化体, 皮肤和合成) 在化学定义的条件下, 这在未来的疾病建模和骨科手术中基于细胞的治疗。
摘要
为了响应 Wnt 等信号, 骨形态发生蛋白 (Bmp) 和从周围组织分泌的声波刺猬 (SHH), 体细胞 (Sonic) 产生多种细胞类型, 包括肌瘤 (MYO)、菌核 (SCL)、皮肤病体 (d) 和合成体 (SYN), 依次发展为骨骼肌、轴向骨骼、背真皮和轴向肌腱韧带。因此, 从人类诱导的多能干细胞 (Ipsc) 中生成 Sm 及其衍生物对于获得多能干细胞 (Psc) 用于再生医学和骨科外科领域的疾病研究至关重要。尽管一些研究人员以前曾报告过来自 Psc 的 MYO 和 SCL 诱导协议, 但尚未有研究表明从 Ipsc 中引入了 SYN 和 D。因此, 高效诱导完全胜任的 Sm 仍然是一项重大挑战。在这里, 我们通过模仿鸡鼠 SM 发展过程中的信号环境, 在体外总结人类 Sm 模式, 并报告了在化学条件下从人类 Ipsc 系统诱导 sm 衍生物 (MYO、SCL、D 和 SYN) 的方法。通过预压中胚层 (PSM) 和 SM 状态定义的条件。成功地将有关鸡鼠 SM 发展的知识应用于人 Ipsc 对 Sm 的诱导。该方法可作为研究人体体细胞生成和模式的新工具, 不使用胚胎, 并用于基于细胞的治疗和疾病建模。
引言
从 Psc 开发所需细胞类型的定向分化方法是将 psc 衍生细胞的研究转化为临床应用的必要步骤。关键基因的强迫表达是一种很有希望的从 Psc 分化的细胞, 并提高了我们对胚胎发生过程中细胞命运测定、器官形态发生和组织的遗传调控的认识.此外, 以小鼠和小鸡胚胎的发育为路线图, 对内源性信号环境进行概述, 被认为是 Psc 定向分化的关键。然而, 考虑到 psc 衍生细胞在基于细胞的疗法等临床研究中的应用, 后一种策略更合适, 因为它不需要基因操纵。
几项研究报告了在化学定义条件下从人类和小鼠 Psc 中诱导的中胚层。通常情况下, 这些方法依赖于活性因子β (TGFβ) 信号和骨形态发生蛋白 (BMP) 信号, 被认为是进行中胚层和中胚层分化, 导致低诱导效率副轴中胚层 (约 20%)2。换句话说, 这些信号通路诱导的 psc 衍生的中胚层主要是侧板中胚层, 而不是后轴中胚层。最近, 一些研究证明了基于不同策略3、4、5、6、7、8的psc 衍生的副轴中胚层的高效生产.在这些研究中, 用相对较高浓度的糖原合成酶激酶激酶 3 (wsk3) 抑制剂 (wnt 信号激活剂) 培养 psc, 使副轴间胚层的诱导效率达到 70%-95%6,7.
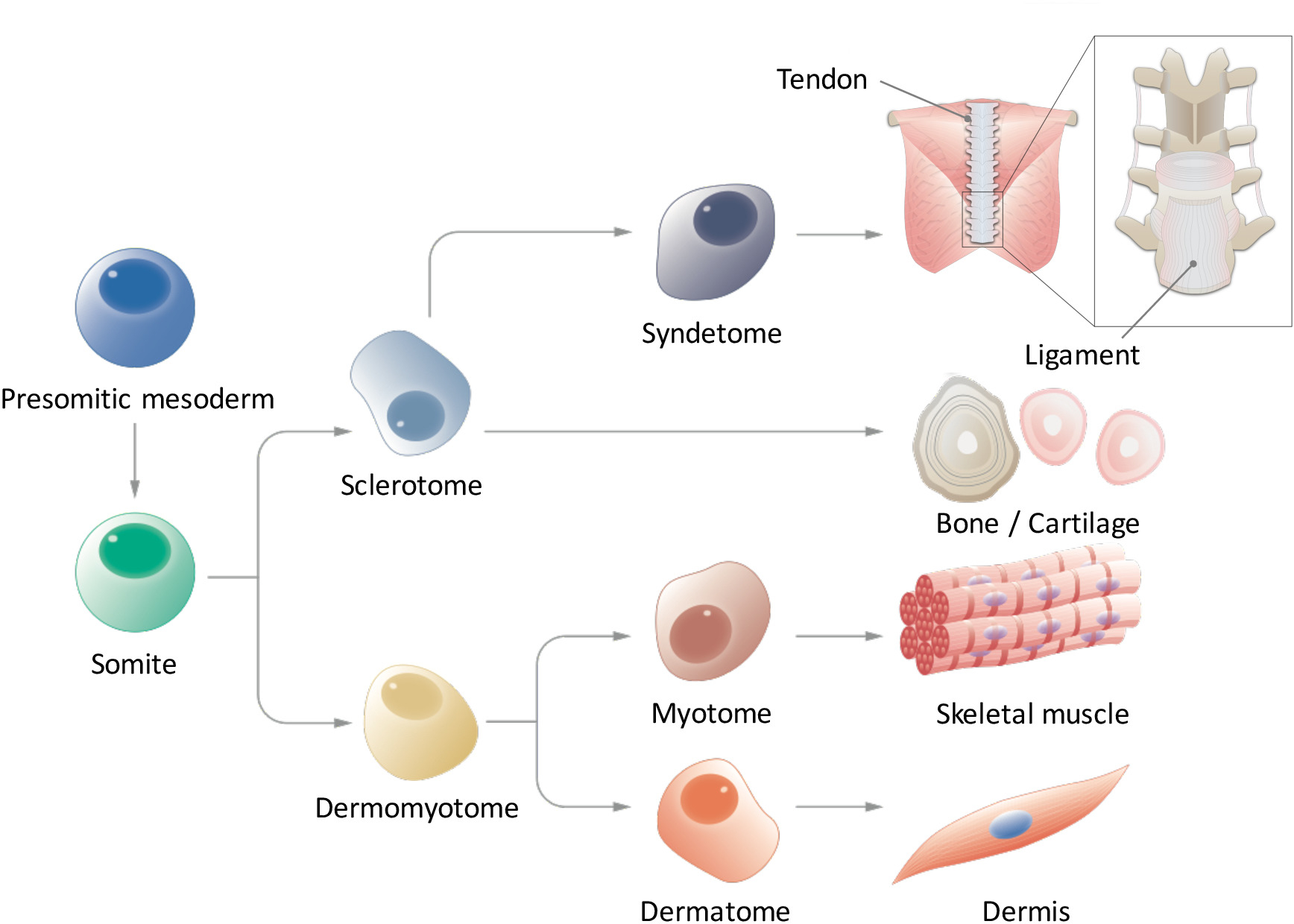
在体细胞发生过程中, 后轴间胚层首先形成前部中胚层 (psm), 然后通过中膜至上皮过渡9,10在前部形成体细胞 (sm)。缺口配体 delta 样 1 (DLL1) 已知在体体发生过程中起着关键作用, 因为在 mRNA 和蛋白质水平上对 DLL1 表达的振荡控制, 调节 SM 分割。SMs 最终细分为两部分, 导致皮肤内和腹侧硬膜体 (SCL).随后, DM 区分为皮肤科 (D), 真皮的前体, 肌母细胞 (MYO), 骨骼肌的前体;另外, SCL 的腹侧部分形成突触 (SYN), 肌腱和韧带 12的前体 (图 1)。一些研究人员报告了 psc 衍生 sm 衍生物的诱导, 如 myo4、13和 scl14;然而, 这些研究有几个局限性。值得注意的是, 由于我们对 D 和 SYN 信号环境的了解是零碎的, 因此还没有系统地建立 D 和 SYN 的归纳协议。为了证明从 Psc 诱导的 Sm 的充分能力, 有必要显示诱导的 Sm 对所有四种衍生物 (D、MYO、SCL 和 SYN) 的多重分化能力, 而以前的研究只集中在特定的 SM 衍生物。在这里, 我们报告如何通过 PSM 和 SM 命运从人类 Ipsc 15 生成所有四个 SM 导数, 包括 D 和 SYN.我们相信, 建立一个体外逐步方法, 模型 SM 发展过程可以有助于研究人类 SM 如何发展在胚胎发生过程中, 而不使用胚胎。
研究方案
京都大学医学系和医学研究生院伦理委员会批准了所有涉及人类 Ipsc 的实验规程。
1. 人工 Ipsc 的制备
请注意:用灵长类 es 细胞培养基在 SNL 供体细胞 16上培养人的 Ipsc (201b7-pax3-gfp), 辅以 4 ngml 重组人基本成纤维细胞生长因子 (fgf2) 和0.5% 青霉素和链霉素 (以下简称 hESC 培养基, 见表 1)。当融合比达到 70%-80% 时, 通过细胞如前面所述的17。
-
人的 Ipsc 在 SNL 馈线单元上的传递17
- 为了进行传代, 在细胞培养盘中加入 PBS 并冲洗细胞。然后, 删除 PBS (以下, 此过程将被称为使用 PBS 清洗)。
- 在室温 (rt) 下添加1毫升 CTK 溶液 (见表 1), 并等待 snl 馈线单元开始从盘子底部分离。
- 取出 CTK 溶液, 然后使用 PBS 清洗两次。
- 在盘子中加入1毫升的 hESC 培养基 (见表 1), 然后用刮刀刮刮细胞, 收集到15毫升的锥形管中。
- 用 1, 000Μl 尖端轻轻移滤五次, 然后转移到装满 hESC 介质的新菜中。使用1:4 到1:4 的拆分比率, 具体取决于传递前的汇合比。此外, 根据菜的大小变化 hESC 介质的体积 (例如, 6 厘米的菜品为3毫升, 10 厘米的菜品为8毫升)。
- 在37°c 和 5% CO 2 下培养人的 Ipsc.
- 每天更换培养基 (传代后的第二天除外), 并培养细胞, 直到下一次传代过程。
-
人 IPM 诱导前人工 Ipsc 的无 fed 培养
注:为了最大限度地减少从 SNL 给料细胞分泌的生长因子的影响, 在无微弱的条件下, 用无微弱的细胞培养基 (见表 1) 对细胞外基质 (ECM) 溶液进行培养 (见表 1) (见表 1)在 PSM 诱导前3天涂盘。- 第4天 (开始 PSM 诱导前 4天)
- 要准备 ECM 溶液涂层盘, 在4°C 下一夜添加4毫升的 ECM 溶液到10厘米的盘。
请注意:在准备时将 ECM 解决方案放在冰上。
- 要准备 ECM 溶液涂层盘, 在4°C 下一夜添加4毫升的 ECM 溶液到10厘米的盘。
- 第3天
- 首先, 从盘中取出 ECM 溶液, 加入8毫升无质细胞培养基。
- 要开始无虚弱的培养, 用 PBS 清洗一次, 冲洗培养的细胞。
- 在 RT 处添加1毫升 CTK 溶液, 直到 SNL 馈送器单元开始从盘底分离。
请注意:使用显微镜来确认所有进纸器单元都已从底部分离。 - 取出 CTK 溶液, 用 PBS 清洗两次, 以便完全卸下所有 SNL 进纸器单元。
- 在盘中加入1毫升无感觉细胞培养基, 然后用刮刀刮刮细胞, 收集到15毫升的锥形管中。
- 使用 1, 000Μl 尖端轻轻移液三次, 然后转移到新的 ECM 溶液涂层10厘米的盘子 (在步骤1.2.1 准备)。使用大约1:2 到 1: 4 的拆分比率, 具体取决于无力热培养前的汇流比。
- 在37°c 和 5% co 2 下培养人的 Ipsc 3天,在-1 日更换介质。
- 第4天 (开始 PSM 诱导前 4天)
2. 荧光活化细胞分选 (流式细胞仪) 的 PSM 鉴别和分离
-
PSM 分化 (第4天 0–Day)
- 吸入无微弱细胞培养基, 并添加8毫升的 PSM 诱导培养基 (CDM 基部培养基, 辅以 10Μm SB431542、10μm CHIR99021、2μm DMH1 和 20 ngml FGF2, 见表 1)。
请注意:PSM 分化开始时的细胞融合是诱导效率的关键。使用显微镜来确认融合率约为30%。 - 在 37°c (5% CO2)下培养细胞 4天, 在第3天改变培养基。
- 第4天的流式细胞 (下文第2.2 节)。
- 吸入无微弱细胞培养基, 并添加8毫升的 PSM 诱导培养基 (CDM 基部培养基, 辅以 10Μm SB431542、10μm CHIR99021、2μm DMH1 和 20 ngml FGF2, 见表 1)。
-
荧光活化细胞分选法分离 DLL1 阳性 PSM 细胞
注:以下是在 facs 对 DLL1 阳性细胞进行分类之前的细胞制备过程。根据制造商的协议, 使用流式细胞仪进行流式细胞仪的流式细胞仪分类。- 吸气的介质, 然后用 PBS 清洗。随后, 添加1毫升细胞离解试剂, 并在 RT 离开3分钟。
- 加入4毫升的 CDM 基部培养基, 用刮刀刮刮细胞, 并将其收集到15毫升的锥形管中。
- 使用自动单元格计数器计数电池数, 然后在 280 x g离心器3分钟。
- 通过吸入小心去除上清液, 并将细胞重新悬浮在流式细胞仪缓冲液中 (见表 1), 浓度为 1.0 x10 7 Cellss/ml。对于负对照样品 (同型对照, 或常规无抗体), 将50μl 转移到15-mL 锥形管中, 然后用450μl 的流式细胞仪缓冲液悬浮。
- 以1200的比例添加 DLL1 抗体 (见材料表)。保护管道不受光线的影响, 并保持在冰上30分钟。
- 离心机以 280 x克3分钟。
- 小心吸气, 并在流式细胞仪缓冲液 (1.0 x10 7 cells/ml) 中重新悬浮, 辅以 1 Mg/ML dapi。
- 转移到收集管, 结合在一个35μm 尼龙网的盖子过滤, 然后把管在冰上, 直到分类完成。对负控制样本执行相同的过程 (步骤 2.2.4)。
- 根据制造商的协议, 使用流式细胞仪进行分类。
- 将分类的 DLL1 阳性细胞收集到15毫升的锥形管中, 其中含有4毫升的 CDM 基部培养基, 辅以10Μm 的 Y27632。对于总 RNA 提取, 离心机在 280 x g下 3分钟, 然后重新悬浮在 rna 裂解缓冲液中, 并储存在-30°c。有关更多详细信息, 请参阅 RNA 提取、逆转录酶和 RT-qPCR 程序 (5.1 节)。
- 根据下面的协议 (第3节) 使用排序的细胞执行 SM 分化。
3. SM 与 PSM 的区别
-
从分类 DLL1 阳性 PSM 细胞中的 SM 分化 (第8天 4–Day)
注:在流式细胞仪分选的前一天准备 ECM 溶液涂层的12孔板。要制备 ECM 溶液涂层的12孔板, 在4°C 下将1毫升的 ECM 溶液加入每口井, 然后离开一夜。在准备时, 将 ECM 解决方案放在冰上。- 2.2.10 步骤后, 离心机在 280 x g处3分钟。
- 仔细吸吸上清液并在1毫升 SM 诱导介质中重新悬浮 (CDM 基部培养基, 辅以 10Μm SB431542 和5μm 智利 99021, 见表 1)。
- 使用自动单元格计数器计数单元格数。
- 种子 1.0 x 105细胞在每个井的 ecm 溶液涂层12孔板含有1毫升 sm 感应介质补充 10Μm Y27632。
- 在37°c 下与 5% CO2 一起孵化 4天 , 直到第8天。在第5天 (流式细胞仪排序后的第二天) 和第7天更改不包含 Y27632 的介质。
- 根据下面的协议, 使用诱导 SM 细胞进行 SM 衍生物分化。对于从诱导的 SM 细胞中提取的全部 RNA, 将细胞收集到一个15-mL 锥形管和离心机在 280 x g 3分钟, 然后重新悬浮在 rna 裂解缓冲液中, 并储存在-30°c。
4. SM 衍生物 (DM、MYO、D、SCL、SYN) 与 SM 的区别
请注意:为了证明 SM 细胞的充分能力, 首先使用 ipsc 衍生的 SM 细胞进行 DM (真皮细胞) 和 SCL (硬化症) 诱导。随后, 利用 DM 细胞进行肌电 (肌瘤) 和 D (皮肤) 诱导, 并使用 SCL 细胞进行 SYN (合成体) 诱导。以下是诱导 SM 细胞在体外诱导的每种衍生物 (DM、MYO、D、SCL 和 SYN) 的诱导协议。
-
SM 细胞 DM 分化 (第11天 8–Day)
- 吸气培养基, 然后加入1毫升的 DM 诱导培养基 (CDM 基部培养基, 辅以 5μm CHIR99021 和 10ngml BMP4, 见表 1)。
- 在37°c 下, 用5% 的 CO2 将细胞培养 3天, 直到第11天。更改第10天 (DM 归纳的第2天) 的介质。
- 根据下面的协议, 使用诱导 DM 细胞进行 MYO 和 D 分化。
-
从 DM 细胞分化的 MYO (11–Day 41 天)
- 吸收培养基, 然后加入1毫升的 MYO 诱导培养基 (CDM 基部培养基, 辅以 5μm CHR99021, 见表 1)。
- 在37°c 下将细胞培养, 用5% 的 CO2,持续 30天, 直到第41天。每3天更换一次介质。
-
D 与 DM 细胞的分化 (第20天 11–Day)
- 吸气介质, 然后加入1毫升的 D 诱导培养基 (CDM 基部培养基, 辅以 5μm CHIR99021 和 10ngml BMP4, 见表 1)。
- 在37°c 下, 用5% 的 CO2 将细胞培养 9天, 直到第20天.每3天更换一次介质。
-
SM 细胞的 SCL 分化 (第11天 8–Day)
- 吸收培养基, 然后加入1毫升的 SCL 诱导培养基 (CDM 基部培养基, 辅以 100 nM sag 和 0.6μm LDN19189, 见表 1)14。
- 用5% 的 Co2 在37°c 下将细胞培养 3天.更改第10天 (SCL 归纳的第2天) 的媒体。
- 根据下面的协议, 使用诱导的 SCL 细胞进行 SYN 分化。
-
从 SCL 细胞的 SYN 分化 (天 11–Day 32)
注:在启动 SYN 感应的前一天准备 ECM 溶液涂层的24孔板。要制备 ECM 溶液涂层的24孔板, 在4°C 下将 0.5 mL 的 ECM 溶液加入每口井中, 然后过夜。在准备时, 将 ECM 解决方案放在冰上。- 将培养基吸气, 然后用 PBS 清洗, 然后在每口井中加入0.2 毫升细胞离解试剂, 然后在 RT 离开3分钟。
- 在每口井中加入 0.8 mL 的 Cdm 基部培养基, 然后刮擦所有细胞, 并将所有细胞收集到一个 15 mL 的锥形管中。
- 离心机以 280 x克3分钟。
- 仔细吸气上清液并重新悬浮在1毫升的 SYN 诱导中 1 (CDM 基部培养基补充 20 ng/mL FGF8, 见表 1), 然后使用自动细胞计数器计数细胞的数量。
- 种子 5.0 x 104 细胞进入每个井的 ecm 溶液涂层24孔板含有1毫升 syn 诱导中1。
- 在37°c 下, 用5% 的 CO2 进行 2013年生.
- 第14天 (SYN 感应第3天), 将培养基替换为 SYN 感应中 2 (CDM 基部培养基, 辅以 10 ngml BMP7 和 10ngml TGFβ3, 见表 1)。
- 在37°c 下, 用5% 的 CO2 进行孵化, 为期 18天, 直到第32天。每3天更换一次介质。
5. ipscs 衍生产品的特性
请注意:在分化时, 使用定量实时 PCR (RT-qPCR)、免疫细胞化学 (ICC)、酶联免疫吸附试验 (ELISA) 和机械拉伸刺激检测来表征人的 Ipsc 衍生物。
-
细胞采集、总 RNA 提取、逆转录酶和定量实时 PCR (RT-qPCR) 分析
- 将细胞样品 (程序2.2.10、3.1.6、5.4.3) 收集到 1.5 mL 管中, 然后以 280 x克的速度离心3分钟。
- 取出上清液, 然后在350μl 的 RNA 裂解缓冲液中重新悬浮, 由适当的总 RNA 提取试剂盒提供。
- 根据制造商的协议, 使用该试剂盒提取总 RNA。
- 根据制造商的协议, 将分离出的1微克总 RNA 反转到 cDNA。
- 根据制造商的协议, 使用合适的酶、试剂和引物执行 RT-qPCR。本研究中使用的引物序列见表 2。
-
免疫细胞化学 (ICC)
- 在使用抗体进行免疫细胞化学之前, 将2% 的甲醛与细胞固定在4°c 下 10分钟, 并用 PBS 清洗两次。
- 对于渗透性, 用0.2% 甲醇或0.2% 聚山梨酸 20/pbs (以下简称 PBS-B) 在4°c 下孵育15分钟。
- 去除渗透剂, 用适当的阻滞剂或1% 的牛血清蛋白在4°c 下对细胞进行60分钟的治疗。
- 在 PBS-T 中加入用10% 阻断缓冲剂稀释的第一抗体, 并在4°c 的振动机上放置一夜。
- 用 PBS-T 清洗三次 (添加 PBS-T 并放置在 RT 的摇摇机上 10分钟)。
- 加入第二个抗体, 在 PBS-T 中稀释10% 的阻滞缓冲液, 并放置在 RT 的摇摇机上60分钟。本研究中使用的 ICC 第一和第二抗体见表 3。
请注意:从这一步开始, 用铝箔包裹平台, 以保护平台不受光线的照射。 - 用 PBS-T 清洗两次。
- 对于计数器染色, 添加半 5000 DAPI 稀释 PBS, 并放置在 RT 的摇摇机上5分钟。
- 删除 DAPI 解决方案, 并将 PBS 添加到每个井中。
- 用荧光显微镜观察细胞染色。或者, 将平台在4°c 下存放长达1个月。
-
酶联免疫吸附法 (ELISA) 对 ipsc 衍生 d 的功能分析
注:人真皮成纤维细胞 (hdf) 可在市场上获得。在 DMEM 中培养 HDF, 辅以10% 的胎牛血清 (见表 1)。- 种子 1.0 x 10 5 细胞的 ipscs 衍生 d 和 hdf 到24孔板含有1毫升的每个培养基 (d:d 诱导培养基, HDF:DMER 补充10% 的胎儿牛血清)。
- 经过3天的细胞培养后, 收集每种培养基 100Μl, 放入 1.5 mL 管中, 并在4°c 下储存。
- 根据制造商的指示, 执行一系列程序, 如添加检测抗体和二级抗体, 并通过生成针对控制浓度的标准曲线来量化目标数量分析样品。
-
Ipsc 衍生 SYN 功能分析中的机械拉伸刺激试验
注:成年人类肌腱细胞可在商业上获得 (见材料表)。培养人 ipsc 衍生的 syn 和成人人的肌腱细胞在细胞拉伸装置上进行机械拉伸刺激试验如下:18,19。分别使用 SYN 诱导中2和肌腱细胞生长培养基 (见材料表) 作为每个 ipsc 衍生的 syn 和成人人的肌腱细胞的培养培养基。- 在拉伸前 24小时, ipsc 衍生 SYN 和人的肌腱细胞的 1.0 x10 5 细胞进入 ecm 溶液涂层的多井型硅橡胶室, 每个室的培养表面为1.5 厘米 x 1.5 厘米 (见材料表)。
- 设置设备上的室, 用于电池拉伸, 并强制单轴循环应变 (0.5 Hz, 5%)12小时。
- 对于总 RNA 提取, 添加350μl 的 RNA 裂解缓冲液, 然后刮并将细胞收集到 1.5 mL 管中, 用于总 RNA 提取和随后的 RT-qPCR 分析 (见过程 5.1)。
结果
本报告中的所有数字都是通过 201B7-pax3-gfp Ipsc 获得的, 其中 EGFP 取代了 exon 1 中 PAX3 编码序列的一个等位基因。将在其他地方 (H. SAKURAI, 个人通讯) 介绍建立 201B7-pex3-gfp Ipsc 的情况。利用统计软件对统计意义进行了评价。低于0.05 的 p 值被认为是显著的。
人 ipsc 衍生 PSM 和 SM 细胞的表征
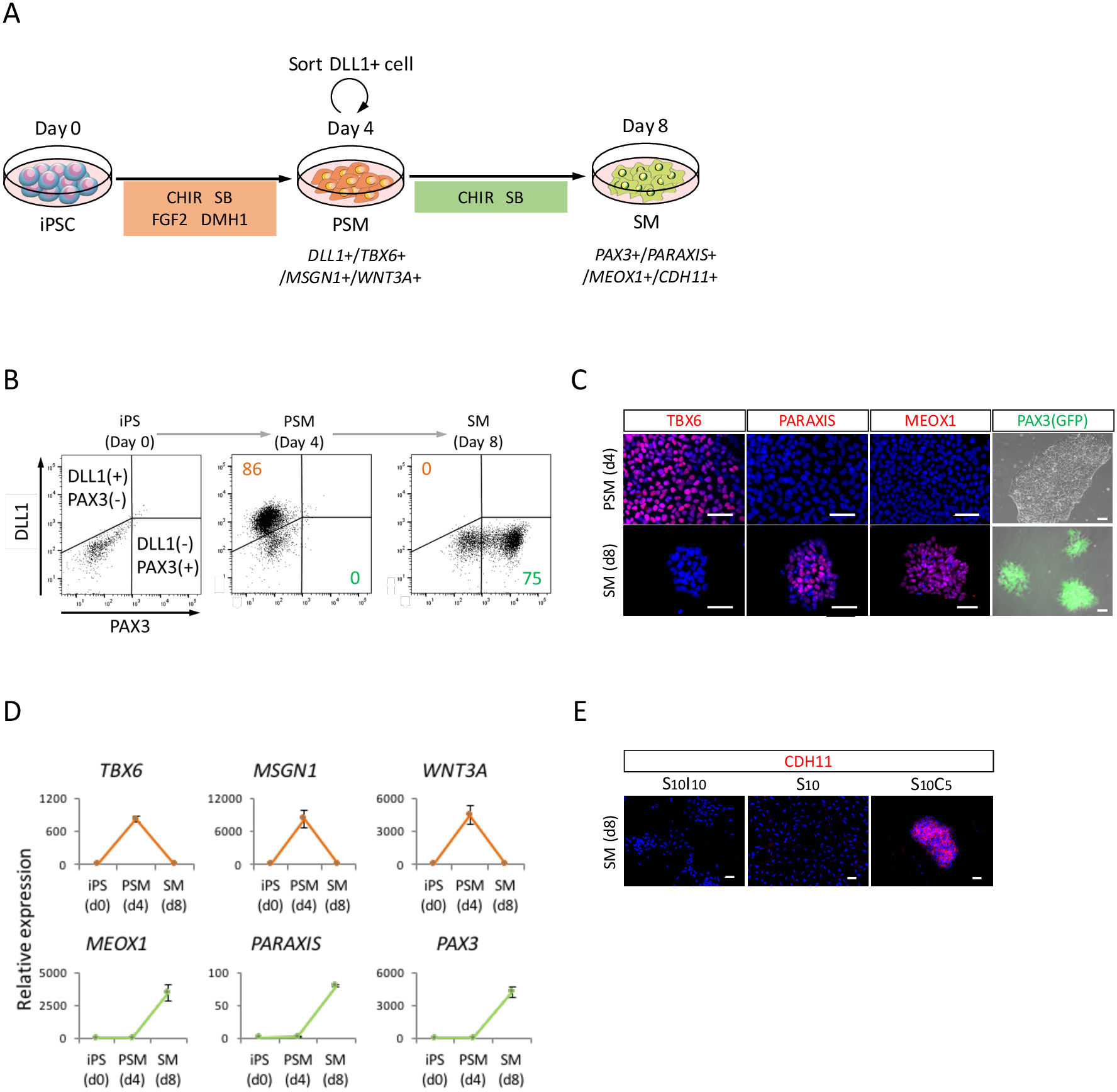
为了评估人类 Ipsc 通过 PSM 状态 (图 2a) 向 sm 的分化, 进行了流式细胞仪分析、icc 分析和 RT-qPCR 分析。如图2B 所示, 85% 以上的细胞对 psm 标记 DLL1 呈阳性, 但在使用人 ipsc 诱导 4天 psm 后, 对 sm 标记 PEX3 呈阴性反应。随后, 该人群在4天的 SM 诱导后成为 PAX3 阳性 sm 细胞。ICC (图 2c) 和 RT-qPCR (图 2d) 也证实了 psm-sm 过渡。TBX6、MSGN1 和 WNT3A、PSM 标记在 PSM 状态 (第4天) 表示, 但在 SM 状态 (第8天) 没有表示。PARAXIS、MEOX1 和 PAX3、SM 标记在 SM 表达, 但未在 PSM 中表示。此外, CDH11, 上皮化 sm 的标记染色, 只在细胞结合智利 99021 (图 2e) 添加 SB431542 后, 在细胞结合部积累。
人 ipsc 衍生 sm 细胞诱导 SM 衍生物的表征
为了评价人 ipsc 衍生 SM 的分化能力, 采用 ICC 分析和 PAC3 (GFP) 荧光法对 DM、MYO、D、SCL 和 SYN (图 3 a) 进行了分化 (图 3a) 进行了评价。如图 3B所示, ALX4 和 en1 染色以及 POX3 (gfp) 荧光证实了 dm 分化;MYOD、MYOG 和肌球蛋白重链 (MHC) 染色证实了 MYO 分化;EN1 和 PDGFRa 染色证实了 d 分化;PEX1、PAX9 和 NKX3.2 染色证实了 SCL 分化;通过 SCX、MKX、COL1A1 和 COL1A2 染色证实了 SYN 分化。
诱导 D 和 SYN 的表征
1. 酶联免疫吸附法 (ELISA) 对 ipsc 衍生 d 的功能分析
在人体中, 真皮成纤维细胞的主要功能之一是分泌细胞外基质 (ECM) 蛋白, 如胶原蛋白和透明质酸, 该蛋白滋润皮肤, 帮助维持皮肤结构。为了证明在 ipsc 衍生 d 和 HDF 培养基中分泌了相当数量的胶原蛋白1和透明质酸蛋白, 进行了 ELISA 检测, 如图 4 a所示。
2. ipsc 衍生 SYN 功能分析的机械拉伸刺激试验
正如一些研究已经报道的那样, 机械刺激会影响出生前后肌腱的发育, 并促进肌腱细胞与前体细胞的分化 18, 19。因此, 众所周知, 对机械应力的反应是肌腱细胞的特征之一。为了证明人 ipsc 衍生的 SYN 和人类成人肌腱细胞的可比反应性, 进行了机械拉伸刺激试验, 如图 4 b 所示。

图 1: 后轴中胚层分层分化示意图.前体中胚层是在早期胚胎发生过程中瞬时出现并经过分割形成体细胞的细胞群。体细胞是一种短暂的干细胞群, 可产生多种细胞类型, 如硬化症、真皮细胞、合生体、皮肤和肌细胞, 最终分化为肌腱韧带、骨软骨、骨骼肌和真皮细胞。请点击这里查看此图的较大版本.
{kind=link}

图 2: 流式细胞仪、RT-qPCR 和对人类 ipsc 衍生 psm 的 ICC 分析.(A) 通过 PSM 进行 sm 微分协议的示意图。(B) psm 诱导第4天和 sm 诱导第4天 (来自 ipsc 的第8天) DLL1 染色和 pax3 (Gfp) 荧光的代表性点图。(C) 具有代表性的免疫细胞化学图像和 PS3 (gfp) 荧光在 psm 诱导第4天和 sm 诱导第4天 (第8天从 ipsc)。细胞用抗 TBX6、PARAXIS 和 MEOX1 抗体 (红色) 染色, 用 DAPI (蓝色) 共同染色, 或用 PAX3 (GFP) 荧光 (绿色) 检测。(D) 在 ipsc、psm 和 SM 对 psm 和 sm 标记进行 rt-qpcr 分析。给出了三组实验的均值±标准误差 (s. e.)。(E) sm 第4天 (ipsc 第8天) 的代表性免疫细胞化学图像, 在 s10i10 (sb431542 和 wiwr1 的组合, wnt 信号的抑制剂)、S10 (sb431542) 和 S10I10 (SB431542 和 chir99021 的组合) 条件下培养。细胞用抗 CDH11 抗体 (红色) 染色, 并与 DAPI (蓝色) 共同染色。iPS, 诱导多能干细胞;PSM, 预压中胚层;SM, somite;S10, SB431542 10Μm;C5, CHIR99021 5Μm;I10, IWR1 10Μm;刻度柱 = 50μm。这一数字已从 Nakajima 等人 (2018年)修改15。请点击这里查看此图的较大版本.
{kind=link}

图 3: dm、myo、d、SCL 和 SYN 与人类 ipsc 派生 sm 的 icc 分析.(A) sm 衍生物微分协议的示意图。(B) dm 诱导第3天 (ipsc 第11天) 具有代表性的免疫细胞化学图像和 pax3 (Gfp) 荧光, myo 感应第30天 (第41天从 iPSC), d 诱导的第9天 (第20天从 ipsc), scl 感应的第3天 (第11天从 iPSC) 和第21天 (第32天)从 iPSC) 的 SYN 感应。DM, 细胞被染成抗 ALX4 和 EN1 抗体 (红色), 并与 DAPI (蓝色) 共染色或用 PAX3 (GFP) 荧光 (绿色) 检测;MYO, 细胞被染红抗 myod, MYOG (红色) 和 MHC (青色) 抗体, 也与 DAPI (蓝色) 共染色;D, 细胞被染成抗 EN1 (红色) 和 PDGFRa (青色) 抗体, 并与 DAPI (蓝色) 共同染色;SCL, 细胞被抗 PAX1、PAX9 和 NKX3.2 (红色) 抗体染色, 并与 DAPI (蓝色) 共同染色;SYN, 细胞被染红抗 scx, MKX, COL1A1 和 COL1A2 (红色) 抗体, 并与 DAPI (蓝色) 共染色。DM, 真皮肌瘤;MYO, 肌瘤;D、皮肤科;SCL, 硬化剂;SYN, sydeome;刻度柱 = 50μm。这一数字已从 Nakajima 等人 (2018年)修改15。请点击这里查看此图的较大版本.
{kind=link}

图 4: 诱导 d 和 SYN 的功能测定.(A) 采用 elisa 法分析了培养基中胶原蛋白1和透明质酸蛋白的含量。(B) 用 RT-qPCR 技术评价机械拉伸刺激对诱导 syn 和人成体细胞的影响。给出了三组实验的均值±标准误差 (s. e.)。* p < 0.05;* * p < 0.01;p < 0.001 通过 dunnett 的多重比较 t 检验与拉伸 (-) 进行比较;没有意义, HDF, 成人真皮成纤维细胞。这一数字已从 Nakajima 等人 (2018年)修改15。请点击这里查看此图的较大版本.
{kind=link}
| 中/解决方案 | 里根 | 浓度 |
| 清洁发展机制基础介质 | Iscove 修改了 Dulbecco 的 medium/Ham 的 F12 | 1: 1 |
| 青霉素/链霉素 | 0.5% | |
| 化学定义的脂质精矿 | 1% | |
| Apo-转铁蛋白 | 15 Mg/ml | |
| 单氯甘油酯 | 450 mM | |
| 牛血清白蛋白 | 5 Mg/ml | |
| 胰岛素 | 7 mgml | |
| CTK 解决方案 | 水 | - |
| 胰蛋白酶 | 0.25% | |
| 胶原酶 IV | 0.1 Mg/ml | |
| 氯化钙 | 1米 | |
| 击倒 SR | 20% | |
| D 感应介质 | 清洁发展机制基础介质 | - |
| 智利99021 | 5μm | |
| BMP4 | 10 ngml | |
| DM 感应介质 | 清洁发展机制基础介质 | - |
| 智利99021 | 5μm | |
| BMP4 | 10 ngml | |
| ECM 解决方案 | 人工细胞外基质 | 0.3 Mg/ml |
| DMEM/F12 | - | |
| 流式细胞仪缓冲 | Pbs | - |
| 牛血清白蛋白 | 0.1% | |
| 无饲料细胞培养培养基 | mTeSR1 | - |
| 青霉素/链霉素 | 0.5% | |
| HDF 培养基 | DMEM | - |
| 胎儿牛血清 | 10% | |
| hESC 媒体 | 灵长类 es 细胞培养基 | - |
| 青霉素/链霉素 | 0.5% | |
| FGF2 | 4 ngml | |
| MYO 诱导介质 | 清洁发展机制基础介质 | - |
| 智利99021 | 5μm | |
| PSM 感应介质 | 清洁发展机制基础介质 | - |
| SB431542 | 10μm | |
| 智利99021 | 10μm | |
| DMH1 | 2微米 | |
| FGF2 | 20 ngml | |
| SCL 感应介质 | 清洁发展机制基础介质 | - |
| 凹陷 | 100 nM | |
| LDN19189 | 0.6μm | |
| SM 感应介质 | 清洁发展机制基础介质 | - |
| SB431542 | 10μm | |
| 智利99021 | 5μm | |
| SYN 感应中1 | 清洁发展机制基础介质 | - |
| FGF8 | 20 ngml | |
| SYN 感应中2 | 清洁发展机制基础介质 | - |
| BMP7 | 10 ngml | |
| TGFβ3 | 10 ngml |
表 1: 媒体和解决方案配方。
| 名字 | 向前 | 反向 |
| ACTB | CACATTGGACAGGGGTTC | AGGTTGGGGGGCCCCAGT |
| COL1A1 | GGACAGAGGTTCAFGT | GCCATCATCCAGGAGC |
| MEOX1 | GAGATTGGTAACCGGA | GACTGGAGGCTGGA |
| MGN1 | GGAGAGCCAGGAGGGGA | GCTGGGGTTCCGAGGGT |
| PARAXIS | TCCTGAGGTGGAGGAT | CCACCCCACACACACACACAGT |
| POX3 | AGGAGGAGGAGGAGGAAG | CAGTTGTGTGGGAGG |
| SCX | CCCADAGCTGCCTC | GCGAATCGCTCTTCTGTC |
| TBX6 | AGCCTGTTTCCATCGT | AGGCACCAGAGAGAGGAAT |
| TNMD | CCCTCATGAGCCCCTT | CCTCTCTCACAGATTGG |
| WNT3A | AGATTGCACCAGAGIT | ATGGGGCACTGCAAAG |
表 2: 用于 RT-qPCR 分析的引物序列。
| 浓度 | ||
| 第一 抗体 | ALX4_Goat | -50 |
| CDH11_Mouse | -1000 | |
| COL1A1_Rabbit | 1.5 100 | |
| COL2A1_Mouse | 1-2 微克/升 | |
| EN1_Rabbit | -50 | |
| MEOX1_Rabbit | -50 | |
| MHC _ 兔 | 半200 | |
| MKX _ 兔 | -50 | |
| MYOD _ 兔 | -500 | |
| 鼠标 | 1.5 400 | |
| NKX3.2_Rabbit | -50 | |
| PARAXIS _ 兔 | -50 | |
| PAX1_Rabbit | -50 | |
| PAX9_Rabbit | -50 | |
| PDGFRa _ 山羊 | 1.5 100 | |
| SCX _ 兔 | -50 | |
| TBX6_Goat | -50 | |
| 第二 抗体 | 驴抗山羊 IgG (H + L) 二次抗体555 | -500 |
| 驴抗山羊 IgG (H + L) 二次抗体647 | -500 | |
| 山羊抗鼠 IgG (H + L) 二次抗体555 | -500 | |
| 山羊抗兔 IgG (H + L) 二次抗体555 | -500 | |
| 山羊抗兔 IgG (H + L) 二次抗体647 | -500 |
表 3: ICC 的第一和第二抗体。
讨论
通过 PSM 诱导 psc 衍生 sm 的一种众所周知的方法是在 PSC 诱导 psm 期间, 而不是在 PSM 成熟过程6中, 将 chr99021 + A83-01 (tgfβ抑制剂) 结合起来。在本研究中, 用 C59 诱导 sm 抑制 Wnt/β-catenin 信号转导。然而, 我们引入了使用 SIR99021 激活 wnt 途径在 SM 分化过程中。这一决定是根据以下调查结果作出的: 在 SM 的周围组织中表达了几个 Wnt, 并考虑到 WNT 记者在sm 20 中处于活跃状态。因此, 我们观察到上皮化, 这是 SM 在体内的一个特征, 只有在 SIR99021 的条件下, 基于 CDH11 在细胞连接中的积累 (图 2e)。这一观察表明, WNT 信号在 PSM 分化和 SM 上皮化过程中的关键参与, 因此我们的协议可以更好地重述内源性信号环境。然而, 这也意味着在分化过程中微调 Wntyβ-catenin 信号通路的进一步可能性, 因为分化的鲁棒性和效率可能因细胞类型、细胞系和各种不同而有显著差异每个研究人员使用的 wnt 诱导剂的化合物。
这种方法还允许我们从人类 Ipsc 生成所有四个 SM 导数, MYO、D、SCL 和 SYN。我们使用 CDM 的逐步协议可用于识别人体体细胞模型中的信号需求, 并为人类 SM 的发展提供重要的见解。例如, 我们的方法可以用于研究分割时钟机制, 一个分子振荡系统, 调节 SM 的形成。它已在小鼠、小鸡和斑马鱼身上进行了彻底的研究, 但由于缺乏适当的实验工具, 在人类身上没有进行过彻底的研究。
此外, 我们的方法还可以适用于未来的临床细胞疗法。例如, 人类 ipsc 衍生的 D 或 SYN 可以移植到严重受伤的皮肤或肌腱断裂进行再生和治疗。但是, 在实际应用此方法之前, 需要解决几个限制。虽然在本研究中, 我们使用 SNL 馈线细胞进行 iPSC 维护和 ECM 解决方案, 这是从恩格尔布雷斯-霍尔姆-热小鼠肉瘤提取, 作为在诱导过程中在盘子表面涂层, 这些非人类动物衍生试剂应该是以提高临床质量。此外, 还必须提高细胞数量和质量, 包括所需细胞的纯度和成熟度。此外, 不仅细胞数量, 而且细胞强度也是肌腱韧带再生的重要特征。此外, 开发用于纯化的表面标记和一种新的三维重建方法对于推进我们的临床细胞疗法方案是必不可少的。
披露声明
作者没有什么可透露的。
致谢
我们要感谢博士. Junya Toguchida (CiRA) 协助项目管理和资金收购, Mitsuaki Shibata 先生 (CiRA) 和 Mei Terashima 女士 (CiRA) 提供技术援助, Yayoi Toyooka 博士 (CiRA) 和 Daisuke Kamiya 博士 (CiRA)他们校对了手稿, 并为 Masaya Todani 先生 (CiRA) 提供了一个插图 (图 1)。我们还感谢池亚和托古奇达实验室 (CiRA) 的所有成员在本研究期间给予的支持。这项工作得到了日本科学促进会 (26670661) 科学研究赠款的支持, 该项目是利用日本科学和技术局针对疾病的 ips 细胞进行的难治性疾病研究方案机构 (JST) 和日本医学研究与发展机构 (AMED)、实现再生医学研究中心网络的 iPS 细胞研究核心中心以及 iPS 细胞研究基金 (部分前往 Makoto Ikeya 和Junya Toguchida)。Makoto Ikeya 还得到了科学研究赠款 (JSPS) (16H05447) 和利用特定疾病 iPS 细胞进行难治性疾病研究加速方案的支持。
材料
| Name | Company | Catalog Number | Comments |
| ALX4_Goat antibody | Santacruz | sc-22066 | |
| Apo-transferrin | Sigma | T1147 | |
| BMP4 | R&D | 314-BP-010 | |
| BMP7 | R&D | 354-BP-010 | |
| Bovine serum albumin | Sigma | A8806 | |
| Calcium chloride | Nacalai tesque | 067730-15 | |
| CDH11_Mouse antibody | Cell signaling | 13577 | |
| Cell streching device | Strex | STB-140 | |
| Chemically defined lipid concentrate | Gibco | 11905-031 | |
| CHIR99021 | Axon | 1386 | |
| COL1A1_Rabbit antibody | Abcam | ab34710 | |
| COL2A1_Mouse antibody | Thermo scientific | MS-235 | |
| Collagenase IV | Thermofisher | 17104019 | |
| DLL1 APC-conjugated_Mouse antibody | R&D | FAB1818A | For FACS |
| DMEM | Sigma | D6046 | |
| DMEM/F12 | Gibco | 11320-082 | |
| DMH1 | Tocris | 4126 | |
| EN1_Rabbit antibody | Abcam | ab70993 | |
| Fetal bovine serum | Nichirei | 171012 | |
| FGF2 | Wako | 060-04543 | |
| FGF8 | Peprotech | 100-25 | |
| Human dermal fibroblast | Cell applications | 160-05a | |
| Human tenocyte | Angio proteomie | cAP-0041 | |
| Insulin | Wako | 090-06474 | |
| Iscove’s modified Dulbecco’s medium/Ham’s F12 | Gibco | 21056023 | |
| Knockout SR | Gibco | 10828028 | |
| LDN193189 | Axon | 1509 | |
| Matrigel | BD bioscience | 354230 | Artificial extracellular matrix |
| MEOX1_Rabbit antibody | Abcam | ab75895 | |
| MHC_Rabbit antibody | Santacruz | sc-20641 | |
| MKX_Rabbit antibody | Atlas antibodies | A83377 | |
| Monothioglycerol | Sigma | M6145 | |
| mTeSR1 | Stemcell tech | 85850 | |
| Multi well-type silicon rubber chamber | Strex | STB-CH-4W | |
| MYOD_Rabbit antibody | Abcam | ab133627 | |
| MYOG_Mouse antibody | Santacruz | sc-12732 | |
| NKX3.2_Rabbit antibody | Sigma | HPA027564 | |
| Novex Donkey anti Goat IgG(H+L) secondary antibody555 | Invitrogen | A21432 | |
| Novex Donkey anti Goat IgG(H+L) secondary antibody647 | Invitrogen | A21447 | |
| Novex Goat anti Mouse IgG(H+L) secondary antibody555 | Invitrogen | A21422 | |
| Novex Goat anti Rabbit IgG(H+L) secondary antibody555 | Invitrogen | A21428 | |
| Novex Goat anti Rabbit IgG(H+L) secondary antibody647 | Invitrogen | A21245 | |
| PARAXIS_Rabbit antibody | Santacruz | sc-98796 | |
| PAX1_Rabbit antibody | Abcam | ab95227 | |
| PAX9_Rabbit antibody | Gene tex | GTX104454 | |
| PBS | - | - | |
| PDGFRa_Goat | R&D | AF307 | |
| Penicillin/Streptomycin | Invitrogen | 15140-122 | |
| Primate ES cell medium | Reprocell | RCHEMD001 | |
| SAG | Calbiochem | 566661 | |
| SB431542 | Selleckchem | SEL-S1067-10 | |
| SCX_Rabbit antibody | Abcam | ab58655 | |
| TBX6_Goat antibody | R&D | AF4744 | |
| Tendon cell growth medium | Angio-proteomie | cAP-40 | Tenocytes growth medium |
| TGFβ3 | R&D | 243-B3-200 | |
| Trypsin | Gibco | 15090046 |
参考文献
- Tanaka, A., et al. Efficient and reproducible myogenic differentiation from human iPS cells: prospects for modeling Miyoshi Myopathy in vitro. PLoS One. 8 (4), e61540 (2013).
- Sakurai, H., et al. In vitro modeling of paraxial mesodermal progenitors derived from induced pluripotent stem cells. PLoS One. 7 (10), e47078 (2012).
- Chal, J., et al. Generation of human muscle fibers and satellite-like cells from human pluripotent stem cells in vitro. Nature Protocl. 11 (10), 1833-1850 (2016).
- Chal, J., et al. Differentiation of pluripotent stem cells to muscle fiber to model Duchenne muscular dystrophy. Nature Biotechnology. 33 (9), 962-969 (2015).
- Umeda, K., et al. Human chondrogenic paraxial mesoderm, directed specification and prospective isolation from pluripotent stem cells. Scientific Reports. 2, 455 (2012).
- Loh, K. M., et al. Mapping the Pairwise Choices Leading from Pluripotency to Human Bone, Heart, and Other Mesoderm Cell Types. Cell. 166 (2), 451-467 (2016).
- Xi, H., et al. In Vivo Human Somitogenesis Guides Somite Development from hPSCs. Cell Reports. 18 (6), 1573-1585 (2017).
- Chal, J., Pourquie, O. Making muscle: skeletal myogenesis in vivo and in vitro. Development. 144 (12), 2104-2122 (2017).
- Tam, P. P., Beddington, R. S. The formation of mesodermal tissues in the mouse embryo during gastrulation and early organogenesis. Development. 99 (1), 109-126 (1987).
- Aulehla, A., Pourquie, O. Signaling gradients during paraxial mesoderm development. Cold Spring Harbor Perspectives in Biology. 2 (2), a000869 (2010).
- Christ, B., Scaal, M. Formation and differentiation of avian somite derivatives. Advances in Experimental Medicine and Biology. 638, 1-41 (2008).
- Brent, A. E., Schweitzer, R., Tabin, C. J. A somitic compartment of tendon progenitors. Cell. 113 (2), 235-248 (2003).
- Sakurai, H., et al. Bidirectional induction toward paraxial mesodermal derivatives from mouse ES cells in chemically defined medium. Stem Cell Research. 3 (2-3), 157-169 (2009).
- Zhao, J., et al. Small molecule-directed specification of sclerotome-like chondroprogenitors and induction of a somitic chondrogenesis program from embryonic stem cells. Development. 141 (20), 3848-3858 (2014).
- Nakajima, T., et al. Modeling human somite development and fibrodysplasia ossificans progressiva with induced pluripotent stem cells. Development. 145 (16), (2018).
- McMahon, A. P., Bradley, A. The Wnt-1 (int-1) proto-oncogene is required for development of a large region of the mouse brain. Cell. 62 (6), 1073-1085 (1990).
- Takahashi, K., et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 131 (5), 861-872 (2007).
- Suzuki, H., et al. targeting of the transcription factor Mohawk in rats causes heterotopic ossification of Achilles tendon via failed tenogenesis. Proceedings of the National Academy of Sciences of the United States of America. 113 (28), 7840-7845 (2016).
- Marturano, J. E., Arena, J. D., Schiller, Z. A., Georgakoudi, I., Kuo, C. K. Characterization of mechanical and biochemical properties of developing embryonic tendon. Proceedings of the National Academy of Sciences of the United States of America. 110 (16), 6370-6375 (2013).
- Maretto, S., et al. Mapping Wnt/beta-catenin signaling during mouse development and in colorectal tumors. Proceedings of the National Academy of Sciences of the United States of America. 100 (6), 3299-3304 (2003).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。