
Method Article
Somite türevleri indüklenen insan nesil Vitro Pluripotent kök hücreler
Bu Makalede
Özet
Gelecekteki hastalığı modelleme uygulamaları olan kimyasal olarak tanımlanan koşullarda, burada İnsan İndüklenmiş pluripotent kök hücre farklılaşma içine her somite türev (myotome, sclerotome, dermatome ve syndetome) için bir iletişim kuralı mevcut ve Ortopedik cerrahi hücre tabanlı terapiler.
Özet
Buna karşılık sinyalleri WNTs gibi kemik morfogenetik proteinler (BMP) ve dokuları çevreleyen salgılanan sonic kirpi (SHH), somites (SMs) myotome (MYO), sclerotome (SCL), dermatome (D) ve syndetome (SYN) de dahil olmak üzere birden çok hücre tipleri ortaya çıkmasına neden , hangi sırayla geliştirmek kas iskelet, axial iskelet, dorsal dermis ve Aksiyel tendon/bağ, sırasıyla. Bu nedenle, SMs ve onların türevleri İnsan İndüklenmiş pluripotent kök hücreler (iPSCs) üzerinden nesil pluripotent kök hücreler (PSC) ortopedik cerrahi alanında hastalığı araştırma ve rejeneratif tıp uygulamasında için elde etmek için önemlidir. Her ne kadar daha önce çeşitli araştırmacılar tarafından rapor MYO ve SCL PSC'ler üzerinden indüksiyon protokollerde, hiçbir çalışma henüz iPSCs SYN ve D indüksiyon göstermiştir. Bu nedenle, tam yetkili SMs verimli indüksiyon büyük bir meydan okuma olarak kalır. Burada, sinyal çevre piliç/fare SM geliştirme ve rapor yöntemleri SM türevlerinin (MYO, SCL, D ve SYN) altında insan iPSCs sistematik indüksiyon sırasında kimyasal olarak taklit ederek insan SM desenlendirme insan iPSCs tüp bebek ile özetlemek tanımlanmış koşulları presomitic mesoderm (PSM) ve SM Birleşik. Civciv/fare SM geliştirme ile ilgili bilgi için SMs indüksiyon ile insan iPSCs başarıyla uygulandı. Bu yöntem insan somitogenesis okuyan ve hücre tabanlı tedavi ve hastalık modelleme için embriyo ve kullanımı olmadan desenlendirme için bir roman araç olabilir.
Giriş
Yönlendirilmiş farklılaşma yöntemi PSC'ler istenen hücre türünden için geliştirmek PSC kaynaklı hücreler çalışma klinik uygulamalara çeviri için gerekli bir adımdır. Zorla ifade anahtar genlerin organ-hücre farklılaşması PSC'ler üzerinden için umut verici bir stratejidir ve genetik düzenleme hücre kader tayini, organ morfogenez ve organizasyon embriyogenez1sırasında bizim anlayış geliştirdi. Ayrıca, fare ve civciv embriyo gelişimi bir yol haritası kullanarak endojen sinyal ortamlar recapitulating PSC'ler yönlendirilmiş farklılaşma için temel olarak kabul edilir. Bu gen düzenleme gerektirmez çünkü ancak, uygulama PSC türetilmiş hücre hücre tabanlı terapiler gibi klinik çalışmalarda göz önüne alındığında, ikinci strateji daha uygundur.
Çeşitli çalışmalarda insan dan mesoderm indüksiyon bildirdin ve fare PSC'ler kimyasal koşullar tanımlanan. Genellikle, bu yöntemler aktivin/nodal/dönüştürme büyüme faktörü β (TGFβ) sinyal üzerinde yararlanmıştır ve kemik morfogenetik protein (BMP) sinyal, Mezo-endoderm ve mesoderm farklılaşması, içinde bir düşük indüksiyon verimliliğini kaynaklanan gerçekleştirmek için inanıyordu paraxial mesoderm (yaklaşık % 20)2. Başka bir deyişle, bu sinyal yolları tarafından indüklenen PSC kaynaklı mesoderm esas olarak yan plaka mesoderm ve paraxial mesoderm oldu. Son zamanlarda, farklı stratejiler3,4,5,6,7,8 dayalı PSC kaynaklı paraxial mesoderm verimli üretim birkaç çalışmalar göstermiştir . Bu çalışmalarda PSC'ler glikojen sentaz kinaz 3 (GSK3) nispeten yüksek konsantrasyonları ile kültürlü inhibitörleri (WNT sinyal harekete geçirmek), sonuç olarak paraxial mesoderm indüksiyon verimliliğini %70-%956,7 ulaştı .
Somitogenesis, paraxial mesoderm ilk presomitic mesoderm (PSM) özafagusu oluşturur ve sonra ön bölümü ile Mezenşim epitel geçiş9,10somites (SMs) oluşturur. SM segmentasyon salınım DLL1 ifade, mRNA ve protein seviyesinde, her ikisi de kontrolünü düzenleyen gibi çentik ligand Delta benzeri 1 (DLL1) bir rol somitogenesis sırasında sahip olduğu bilinmektedir. SMs sonunda alt bölümlere iki parça, dorsally dermomyotome (DM) sebebiyet veren ve sclerotome (SCL) ventrally11. Daha sonra DM dermatome (D), dermis ve myotome (MYO), kas iskelet habercisi habercisi ayırır; Ayrıca, syndetome (SYN), tendon ve bağ12 (şekil 1) habercisi SCL ventral bir bölümünü oluşturur. Bazı araştırmacılar PSC kaynaklı SM türevleri MYO4,13 ve SCL14gibi indüksiyon bildirdin; Ancak, bu çalışmalarda bazı sınırlamaları vardır. D ve SYN sinyal ortamları ile ilgili bilgilerimizi parçalar halinde olduğu için özellikle, D ve SYN indüksiyon protokollerde henüz sistematik olarak kurulmuş değil. SMs tam yetki indüklenen PSC'ler göstermek için çoklu farklılaşma göstermek için esastır kapasite tüm dört türevleri (D, MYO, SCL ve SYN) içine indüklenen SMS'nin önceki çalışmalarda sadece belirli SM türevleri üzerinde odaklanmış iken. Burada, biz nasıl D ve SYN, insan iPSCs15PSM ve SM kader dahil olmak üzere, tüm dört SM türevleri oluşturmak için rapor. İnandığımız bir tüp bebek kademeli yöntemi bu modelleri SM geliştirme süreci nasıl insan SM çalışma odasına katkıda bulunabileceğini kurulması embriyogenez sırasında embriyolar kullanmadan gelişir.
Protokol
İnsan iPSCs içeren tüm deneysel protokoller tıp bölümü ve lisansüstü tıp Fak., Kyoto Üniversitesi Etik Kurulu tarafından kabul edildi.
1. insan iPSCs önce hazırlık indüksiyon
Not: Kültür insan iPSCs (201B7-PAX3-GFP) SNL Besleyici üzerinde 4 ng/mL rekombinant insan temel fibroblast büyüme faktörü (FGF2) ve % 0,5 penisilin ve streptomisin primat ES hücre orta ile16 hücreleri (bundan sonra da adlandırılır hESC aracı olarak görmek için Tablo 1). Ne zaman izdiham oranı % 70-%80, geçiş hücreleri yukarıda açıklanan17olarak ulaşır.
-
SNL besleyici hücreler17 insan iPSCs, passaging
- Passaging için PBS hücre kodlamayla çanak içine ekleyin ve hücreleri durulayın. Sonra PBS kaldırın (bundan sonra bu işlem için PBS ile yıkama olarak ifade edilecektir).
- CTK çözüm 1 mL ekleyin (bkz. Tablo 1) oda sıcaklığında (RT) ve çanak alttan ayırma SNL besleyici hücreler başlatmak kadar bekleyin.
- CTK çözüm kaldırın ve sonra iki kez PBS ile yıkayın.
- HESC orta 1 mL ekleyin (bkz. Tablo 1) amacına, sonra bir kazıyıcı kullanarak hücreleri scrape ve 15 mL konik tüp içine toplamak.
- Yavaşça beş kez 1.000 µL bahşiş belirtilen içeriğini pipette ve sonra hESC orta ile dolu yeni bir tabak için transfer. 1:4-1:10, passaging önce izdiham oranı bağlı olarak bölünmüş oranında kullanın. Ayrıca, ses düzeyini hESC orta çanağı (örneğin, 3 mL 6 cm çanak, 10 cm yemek için 8 mL için) ölçeğini bağlı olarak değişir.
- 37 ° C ve % 5 CO2insan iPSCs kuluçkaya.
- (Hariç passaging sonra gün) orta her gün değiştirin ve hücreleri passaging yordam kadar kültür.
-
Besleyici-Alerjik PSM indüksiyon önce insan iPSCs, kültür
Not: SNL besleyici hücrelerden salgılanan büyüme faktörlerinin etkisi en aza indirmek için bir besleyici-Alerjik hücre kültür orta ile Besleyici-Alerjik koşullarda insan iPSCs kültür (bkz. Tablo 1) (bkz. Tablo 1) hücre dışı Matriks (ECM) çözümleri boyalı çanak PSM indüksiyon öncesi 3 gündür.- Gün -4 (4 gün PSM indüksiyon başlamadan önce)
- ECM çözümü kaplı yemek hazırlamak için 4 ° C'de 10 cm çanak üzerine ECM çözümünün 4 mL gecede ekleyin.
Not: ECM çözümü hazırlarken Buza koyun.
- ECM çözümü kaplı yemek hazırlamak için 4 ° C'de 10 cm çanak üzerine ECM çözümünün 4 mL gecede ekleyin.
- Gün -3
- İlk olarak, yemek ECM çözümü kaldırın ve besleyici-Alerjik hücre kültür ortamının 8 mL ekleyin.
- Besleyici-Alerjik kültür başlatmak için bir kez kültürlü hücreleri durulama için PBS ile yıkayın.
- SNL besleyici hücreler başlayana kadar yemek alttan ayırma RT 1 mL CTK çözeltisi ekleyin.
Not: Mikroskopi tüm besleyici hücreler alttan ayrılır onaylamak için kullanın. - CTK çözümü kaldırmak ve böylece tüm SNL besleyici hücreler tamamen kaldırılır PBS ile iki kez yıkayın.
- 1 mL besleyici-Alerjik hücre kültür orta çanağı içine ekleyin sonra bir kazıyıcı kullanarak hücreleri scrape ve onları 15 mL konik tüp içine toplamak.
- Hafifçe üç kez 1.000 µL ipucu belirtilen içeriğini pipette sonra (1.2.1 adımı sırasında hazır) yeni bir ECM çözümü kaplı 10 cm tabak transfer. Kabaca 1:2 1:4, besleyici-Alerjik kültür önce izdiham oranı bağlı olarak bölünmüş oranında kullanın.
- İnsan iPSCs 37 ° C ve % 5 CO2 gün -1 orta değişen 3 gün kuluçkaya.
- Gün -4 (4 gün PSM indüksiyon başlamadan önce)
2. PSM farklılaşma ve yalıtım Floresans aktif hücre (FACS) sıralama
-
PSM farklılaşma (gün 0-gün 4)
- Besleyici-Alerjik hücre kültür orta Aspire edin ve PSM indüksiyon Orta 8 mL ekleyin (CDM Bazal orta 10 µM ile takıma SB431542, 10 µM CHIR99021, 2 µM DMH1 ve 20 ng/mL FGF2, bkz: Tablo 1).
Not: Hücre izdiham PSM farklılaşma inisiyasyon, indüksiyon verimlilik için önemlidir. Mikroskopi Konfluent oranı yaklaşık % 30 olduğunu onaylamak için kullanın. - Orta gün 3 değiştirme 4 gün için % 5 CO2, 37 ° C'de hücreler kuluçkaya.
- Hücreleri (2.2, aşağıda bölüm) günde 4 FACS için hasat.
- Besleyici-Alerjik hücre kültür orta Aspire edin ve PSM indüksiyon Orta 8 mL ekleyin (CDM Bazal orta 10 µM ile takıma SB431542, 10 µM CHIR99021, 2 µM DMH1 ve 20 ng/mL FGF2, bkz: Tablo 1).
-
Yalıtım DLL1 olumlu PSM hücre Floresans aktif hücre (FACS) sıralama
Not: FACS DLL1 pozitif hücrelerinin sıralama önce hücre hazırlık için bir yordam aşağıdadır. Akış Sitometresi kullanarak FACS sıralama üreticinin protokole göre gerçekleştirin.- Orta Aspire edin sonra PBS ile yıkayın. Daha sonra hücre ayrılma reaktif 1 mL ekleyin ve 3 dk RT. az için bırakın
- CDM Bazal orta 4 mL ekleyin, bir kazıyıcı kullanarak hücreleri scrape ve onları 15 mL konik tüp içine toplamak.
- Bir otomatik hücre sayaç kullanarak hücreleri saydırmak sonra vasıl 280 x g 3 dk santrifüj kapasitesi.
- Dikkatle süpernatant aspirasyon tarafından kaldırmak ve FACS arabellek hücrelerde resuspend (bkz. Tablo 1) 1.0 x 107 hücre/mL bir konsantrasyon. Bir negatif kontrol örneği için (izotip kontrol, veya antikor olmadan Sözleşmesi), 50 µL 15 mL konik tüp içine aktarmak ve FACS arabellek 450 µL ile askıya alma.
- DLL1 antikor ekleyin ( Tablo malzemelerigörmek) 1/200 oranında. Tüp ışıktan korumak ve 30 dk için buz üzerinde tutun.
- 280 x g , santrifüj 3 dk için.
- Dikkatle süpernatant Aspire edin ve FACS arabelleği (1.0 x 107 hücre/mL) 1 mg/mL ile DAPI takıma resuspend.
- Aktarım süzme, Şapkalı 35 µm naylon fileli dahil bir koleksiyon tüp içine görüntülenmemesini tüp buz üzerinde sıralama tamamlanıncaya kadar. Negatif kontrol örneği (Adım 2.2.4) ile aynı yordamı gerçekleştirin.
- Akış Sitometresi üreticinin protokolüne göre kullanarak sıralama gerçekleştirir.
- CDM Bazal orta Y27632 10 µM ile desteklenmiş 4 mL içeren 15 mL konik tüp içine sıralanmış DLL1 pozitif hücrelerinin toplamak. Toplam RNA ayıklama, 280 x g 3 dk de santrifüj kapasitesi sonra RNA lizis arabellekte resuspend ve -30 ° C'de saklarız RNA ekstraksiyon, ters transkripsiyon ve RT-qPCR yordamları (Bölüm 5.1) daha fazla bilgi için bkz:.
- SM farklılaşma göre sıralanmış hücreleri kullanarak gerçekleştirmek Protokolü (Bölüm 3) aşağıda.
3. SM farklılaşma PSM üzerinden
-
SM farklılaşma sıralanmış DLL1 olumlu PSM hücreleri (günde 4-Day 8)
Not: bir gün önce FACS sıralama ECM çözümü kaplı 12-şey tabak hazırlamak. Bir ECM çözümü kaplı 12-şey plaka hazırlamak için 4 ° C'de her kuyuya ECM çözümü 1 mL ekleyin ve gece bırakın. Buzda ECM çözümü hazırlarken tutun.- Adım 2.2.10, santrifüj 280 x g de 3 min için takip.
- Dikkatle süpernatant Aspire edin ve 1 mL SM indüksiyon orta resuspend (CDM Bazal orta 10 µM ile desteklenmiş SB431542 ve 5 mikron CHIR99021, bkz. Tablo 1).
- Bir otomatik hücre sayaç kullanarak hücreleri saydırmak.
- Tohum 1.0 x 105 hücreleri üzerine her şey plakaların Y27632 10 µM ile desteklenmiş SM indüksiyon orta 1 mL içeren ECM çözümü kaplı 12-şey.
- 37 ° c % 5 CO2 gün 8 kadar 4 gün kuluçkaya. Y27632 gün 5 (FACS sıralama sonra gün) ve gün 7 içeren değil orta değiştirin.
- SM türevleri farklılaşma indüklenen SM hücre göre aşağıdaki protokolleri kullanarak gerçekleştirin. Toplam RNA ayıklama indüklenen SM hücrelerden 15 mL konik tüp ve 3 dk, 280 x g , santrifüj hücreleri toplar sonra RNA lizis arabellekte resuspend ve -30 ° C'de depolayın
4. SM türevleri (DM, MYO, D, SCL, SYN) farklılaşma SM den
Not: Tam yetki SM hücreleri göstermek için ilk DM (dermomyotome) ve buna göre IPSC kaynaklı SM hücreleri kullanarak SCL (sclerotome) indüksiyon gerçekleştirmek. Daha sonra MYO (myotome) ve D (dermatome) indüksiyon DM hücreleri kullanarak gerçekleştirmek ve SCL hücreleri kullanarak SYN (syndetome) indüksiyon kuralları. Aşağıdaki iletişim kuralları her türev (DM, MYO, D, SCL ve SYN) indüklenen SM hücrelerden indüksiyon için tüp bebek vardır.
-
(Gün 8-gün 11) SM hücrelerden DM farklılaşma
- Orta Aspire edin, sonra DM indüksiyon orta 1 mL ekleyin (CDM Bazal orta 5 mikron ile desteklenmiş CHIR99021 ve 10 ng/mL BMP4, bkz: Tablo 1).
- Gün 11 kadar 3 gün için % 5 CO2, 37 ° C'de hücreler kuluçkaya. Orta gün 10 (gün 2 DM indüksiyon) değiştirin.
- MYO gerçekleştirmek ve farklılaşma D kullanarak aşağıdaki protokolleri göre DM hücreleri indüklenen.
-
(Gün 11-gün 41) DM hücrelerden MYO farklılaşma
- Orta Aspire edin, sonra MYO indüksiyon orta 1 mL ekleyin (CDM Bazal orta 5 mikron ile takıma CHIR99021, bkz. Tablo 1).
- Gün 41 kadar 30 gün için % 5 CO2, 37 ° C'de hücreler kuluçkaya. Orta 3 günde değiştirin.
-
(Gün 11-gün 20) DM hücrelerden D farklılaşma
- Orta Aspire edin, sonra D indüksiyon orta 1 mL ekleyin (CDM Bazal orta 5 mikron ile desteklenmiş CHIR99021 ve 10 ng/mL BMP4, bkz: Tablo 1).
- %5 CO2, 37 ° C'de hücreler için gün 20 kadar 9 gün kuluçkaya. Orta 3 günde değiştirin.
-
SCL farklılaşma SM hücrelerden (8-gün 11. gün)
- Orta Aspire edin, sonra SCL indüksiyon orta 1 mL ekleyin (CDM Bazal orta 100 ile desteklenmiş nM SAG ve 0.6 µM LDN193189, bkz. Tablo 1)14.
- %5 CO2, 37 ° C'de hücreler için 3 gün kuluçkaya. Orta gün 10 (gün 2 SCL indüksiyon) değiştirin.
- SYN farklılaşma protokolüne göre indüklenen SCL hücreleri kullanarak gerçekleştirin.
-
SYN farklılaşma SCL hücrelerden (gün 11-gün 32)
Not: bir gün önce SYN indüksiyon başlatılıyor ECM çözümü kaplı 24-şey tabak hazırlamak. Bir ECM çözümü kaplı 24-şey plaka hazırlamak için 4 ° C'de her kuyuya ECM çözümü 0.5 mL ekleyin ve gece bırakın. Buzda ECM çözümü hazırlarken tutun.- Orta Aspire edin sonra PBS ile yıkayın sonra her şey için hücre ayrılma reaktif 0.2 mL eklenir ve RT. adlı 3 dakika bırakılır
- Her şey için CDM Bazal Orta 0.8 mL ekleyin sonra scrape ve 15 mL konik tüp içine tüm hücreleri toplamak.
- 280 x g , santrifüj 3 dk için.
- Dikkatle süpernatant Aspire edin ve SYN indüksiyon orta-1'in 1 mL resuspend (CDM Bazal orta 20 ng/mL ile FGF8, desteklenen bakınız Tablo 1), bir otomatik hücre sayaç kullanarak hücreleri saydırmak.
- Tohum 5.0 x 104 hücreleri plakaların SYN indüksiyon orta-1'in 1 mL içeren ECM çözümü kaplı 24-şey her kuyuya.
- %5 CO2, 37 ° C'de 3 gün kuluçkaya.
- Gün 14 (SYN gün 3 indüksiyon), orta SYN indüksiyon orta-2 ile değiştirin (CDM Bazal orta 10 ng/mL BMP7 ve 10 ng/mL TGFβ3, bkz: Tablo 1).
- %5 CO2, 37 ° C'de gün 32 kadar 18 gün kuluçkaya. Orta 3 günde değiştirin.
5. IPSC türetilen ürünleri karakterizasyonu
Not: Farklılaşma nicel gerçek zamanlı PCR (RT-qPCR), immunocytochemistry (ICC), enzim bağlı immunosorbent deneyleri (ELISA) ve mekanik streç stimülasyon deneyleri, buna göre kullanarak insan iPSCs türevleri karakterize.
-
Hücre hasat, toplam RNA çıkarma, ters transkripsiyon ve nicel gerçek zamanlı PCR (RT-qPCR) analiz
- Hücre örnekleri (yordamlar 2.2.10, 3.1.6, 5.4.3) 1,5 mL tüp içine toplamak sonra vasıl 280 x g 3 dk santrifüj kapasitesi.
- Süpernatant kaldırın, sonra bir uygun toplam RNA ekstraksiyon kiti tarafından sağlanan RNA lizis tamponunun 350 µL içinde resuspend.
- Toplam RNA üreticinin protokolüne göre kit kullanarak ayıklayın.
- Ters cDNA, toplam RNA izole 1 µg üreticinin protokole göre uyarlamak.
- RT-qPCR astar üreticinin protokolüne göre uygun enzimler ve reaktifler kullanarak gerçekleştirin. Bu çalışmada kullanılan astar dizileri Tablo 2' de listelenmiştir.
-
Immunocytochemistry (ICC)
- İmmunocytochemistry antikorlar ile gerçekleştirmeden önce 10 dk 4 ° C'de % 2 paraformaldehyde ile hücreleri tamir ve iki kez PBS ile yıkayın.
- 15 dakika 4 ° C'de % 0,2 metanol veya % 0,2 polysorbate 20/PBS (aşağıda PBS-T anılacaktır) ile permeabilization için kuluçkaya.
- Permeabilization reaktifler kaldırmak ve bir uygun engelleme arabellek veya % 1 sığır serum albümin/PBS için 60 dk 4 ° C'de hücrelerle tedavi.
- Gecede 4 ° C'de titreyen bir makinede yeri ve PBS-T % 10 engelleme arabelleği ile seyreltilmiş ilk antikor ekleyin.
- Üç kez PBS-T ile yıkama (PBS-T ekleyin ve yer üstünde RT sallayarak makinede 10 dk).
- PBS-T ve yer RT, titreyen makineye 60 dk için % 10 engelleme arabelleği ile seyreltilmiş İkinci antikor ekleyin. Bu çalışmada kullanılan ICC için birinci ve ikinci antikor Tablo 3' te listelenir.
Not: Bu adıma itibaren folyo sarma tarafından plaka/çanak ışık korumak. - İki kez PBS-T. ile yıkayın
- Sayaç boyama için PBS ve 5 min için RT, makine sallayarak üzerinde yer ile seyreltilmiş 1/5000 DAPI ekleyin.
- DAPI çözüm kaldırın ve PBS her kuyunun içine ekleyin.
- Hücre boyama floresan mikroskop kullanarak gözlemlemek. Alternatif olarak, plaka/çanak 4 ° c ilâ 1 ay için saklayın.
-
Enzim bağlı Immunosorbent assay (ELISA) IPSC türetilmiş d fonksiyonel analiz için
Not: insan dermal fibroblast hücreleri (HDF) ticari olarak kullanılabilir. Kültür DMEM % 10 fetal sığır serum ile desteklenmiş HDF (bkz. Tablo 1).- Her kültür ortamı 1 mL içeren 24-şey plakalar üzerine IPSC türetilmiş D ve HDF tohum 1.0 x 105 hücreleri (D: D indüksiyon orta, HDF: % 10 fetal sığır serum ile desteklenmiş DMEM).
- Hücre kültürü çalışmalarının, 3 gün topladıktan sonra her orta 100 µL 1,5 mL tüp içine yerleştirin ve 4 ° C'de depolayın
- Yordamlar, algılama antikorlar ve ikincil antikorlar, eklenmesi gibi dizi üreticinin yönerge göre gerçekleştirmek ve konsantrasyon denetimin karşı standart bir eğri üreterek hedef sayısını ölçmek analit örnekleri.
-
Mekanik streç stimülasyon tahlil IPSC kaynaklı SYN fonksiyonel analizi için
Not: yetişkin insan tenocytes piyasada bulunan (bakınız Tablo reçetesi) vardır. Kültür insan IPSC kaynaklı SYN ve aygıt Mekanik streç stimülasyon tahlil için18,19açıklandığı gibi uzanan bir hücreyi yetişkin insan tenocytes. Kullanım SYN indüksiyon orta-2 ve tenocytes büyüme orta ( Tablo malzemelerigörmek) kodlamayla orta her IPSC kaynaklı SYN ve yetişkin insan tenocytes olarak anılan sıraya göre.- 24 h germe önce plaka 1.0 x 105 hücreleri IPSC kaynaklı SYN ve ECM çözümü kaplı çok iyi tipi silikon kauçuk üzerine insan tenocytes Odaları, her biri bir kültür yüzey 1,5 cm x 1,5 cm ( Tablo malzemelerigörmek).
- Hücre germe için cihazın üzerindeki odaları koymak ve monoaxial çevrimsel gerilme (0.5 Hz, % 5) güç 12 h için.
- Toplam RNA çıkarılması için RNA lizis arabelleği, 350 µL eklemek scrape ve toplam RNA çıkarma ve daha sonraki RT-qPCR analiz (5.1 yordama bakın) için 1,5 mL tüp içine hücreleri toplamak.
Sonuçlar
Bu rapordaki tüm rakamlar 201B7-PAX3-GFP iPSCs, EGFP PAX3 kodlama dizisi exon 1 içinde bir gen yerini alır ile elde edilmiştir. 201B7-PAX3-GFP iPSCs kurulması-ecek var olmak başka bir bölümünde (H. Sakurai, kişisel iletişim). İstatistiksel anlamlılık istatistiksel yazılım kullanılarak değerlendirilmiştir. 0,05 önemli kabul edildi daha P değerleri daha düşük.
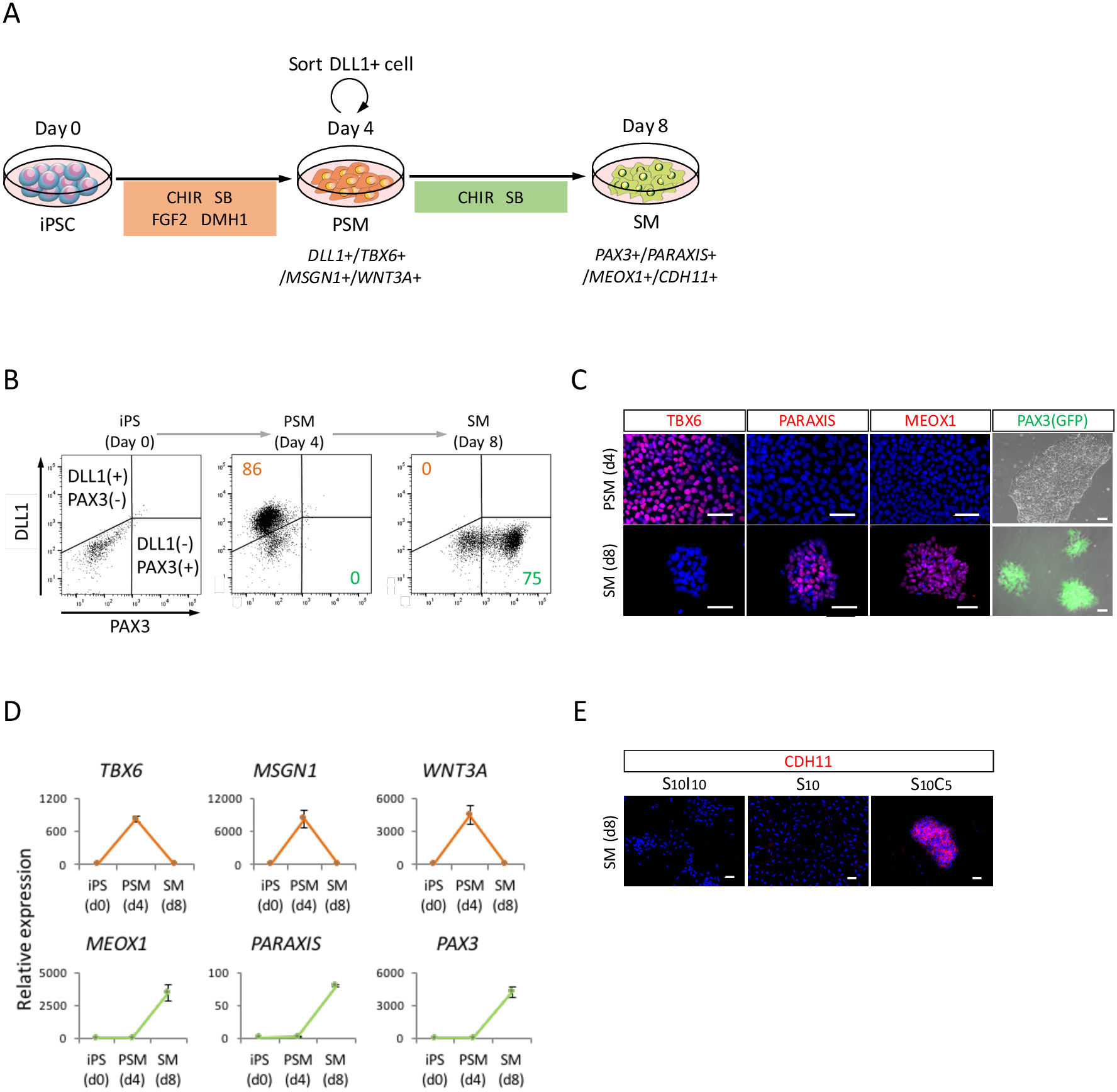
İnsan IPSC kaynaklı PSM ve SM hücrelerin karakterizasyonu
SM doğru insan iPSCs PSM devlet (şekil 2A), FACS analizi, aracılığıyla farklılaşma değerlendirmek için ICC analiz ve RT-qPCR analizi gerçekleştirilmiştir. Şekil 2Biçinde gösterildiği gibi üzerinde hücreleri % 85'i PSM indüksiyon insan iPSCs ile 4 gün sonra olumlu DLL1, PSM, bir işaretleyici ama negatif çıktı PAX3, SM, bir işaret için idi. Daha sonra bu nüfusun PAX3 pozitif SM hücrelerinin SM indüksiyon 4 gün sonra oldu. PSM-SM geçiş ICC (şekil 2C) ve RT-qPCR (şekil 2B) tarafından da doğrulandı. TBX6, MSGN1 ve WNT3A, PSM işaretleri PSM devlet (4 gün) ifade ancak SM devlet (günde 8) ifade değil. SM işaretleri, PARAXIS, MEOX1 ve PAX3, SM ifade, ama PSM ifade değil. Ayrıca, CDH11, epithelialized SM, bir marker boyama sadece SB431542 CHIR99021 ile eklenmesi aşağıdaki hücre-hücre kavşağında birikmiş (şekil 2E).
İnsan IPSC kaynaklı SM hücreleri indüklenen SM türevleri karakterizasyonu
İnsan IPSC kaynaklı SM farklılaşma potens değerlendirmek için DM, MYO, D, SCL ve SYN (şekil 3A) yönünde farklılaşma ICC analizi ve PAX3 (GFP) tarafından değerlendirildi-floresan. Şekil 3B' de gösterildiği DM farklılaşma ALX4 ve EN1 tarafından teyit edildi boyama ve PAX3 (GFP)-floresan; MYO farklılaşma boyama Tanrım, MYOG ve Myosin ağır zinciri tarafından (MHC) doğrulandı; D farklılaşma EN1 ve boyama PDGFRa tarafından doğrulandı; SCL farklılaşma, PAX1, PAX9 ve NKX3.2 Boyama tarafından doğrulandı; ve SYN farklılaşma olduğunu doğruladı SCX, MKX, COL1A1 ve COL1A2 tarafından boyama.
İndüklenen D ve SYN karakterizasyonu
1. enzim bağlı immunosorbent assay (ELISA) IPSC türetilmiş d fonksiyonel analiz için
İnsan vücudunda hidrat cilt ve cilt yapısı sürdürmek yardımcı kolajen ve hyaluronik asit gibi hücre dışı Matriks (ECM) proteinler salgılaması için dermal fibroblastlar birincil işlevlerinden biridir. Kollajen-tip 1 ve hyaluronik asit proteinlerin karşılaştırılabilir bir miktar IPSC türetilmiş D ve HDF kültür ortamında salgılanan göstermek için ELISA, şekil 4Agörüldüğü gibi gerçekleştirildi.
2. Mekanik streç stimülasyon tahlil IPSC kaynaklı SYN fonksiyonel analizi için
Çeşitli çalışmalarda zaten bildirdin gibi mekanik stimülasyon tendon geliştirme öncesi ve doğum sonrası etkiler ve tenocytes farklılaşma öncül hücreleri18,19destekler. Bu nedenle, bu iyi reaktivite mekanik stres tenocytes özelliklerinden biri olduğu bilinmektedir. İnsan IPSC kaynaklı SYN ve insan yetişkin tenocytes karşılaştırılabilir reaktivite göstermek için bir Mekanik streç stimülasyon tahlil şekil 4B' gösterildiği gibi gerçekleştirildi.

Şekil 1: paraxial mesoderm hiyerarşik farklılaşma şematik. Presomitic mesoderm geçici erken embriyogenez sırasında ortaya çıkar ve segmentasyon formu somites için uğrar bir hücre nüfus var. Somites sonunda tendon/bağ, kemik/kıkırdak, iskelet kası ve DermIS ayırmak sclerotome, dermomyotome, syndetome, dermatome ve myotome hücreleri gibi birden çok hücre tipleri için yükselişi verir bir geçici kök hücre nüfus vardır hücreleri. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
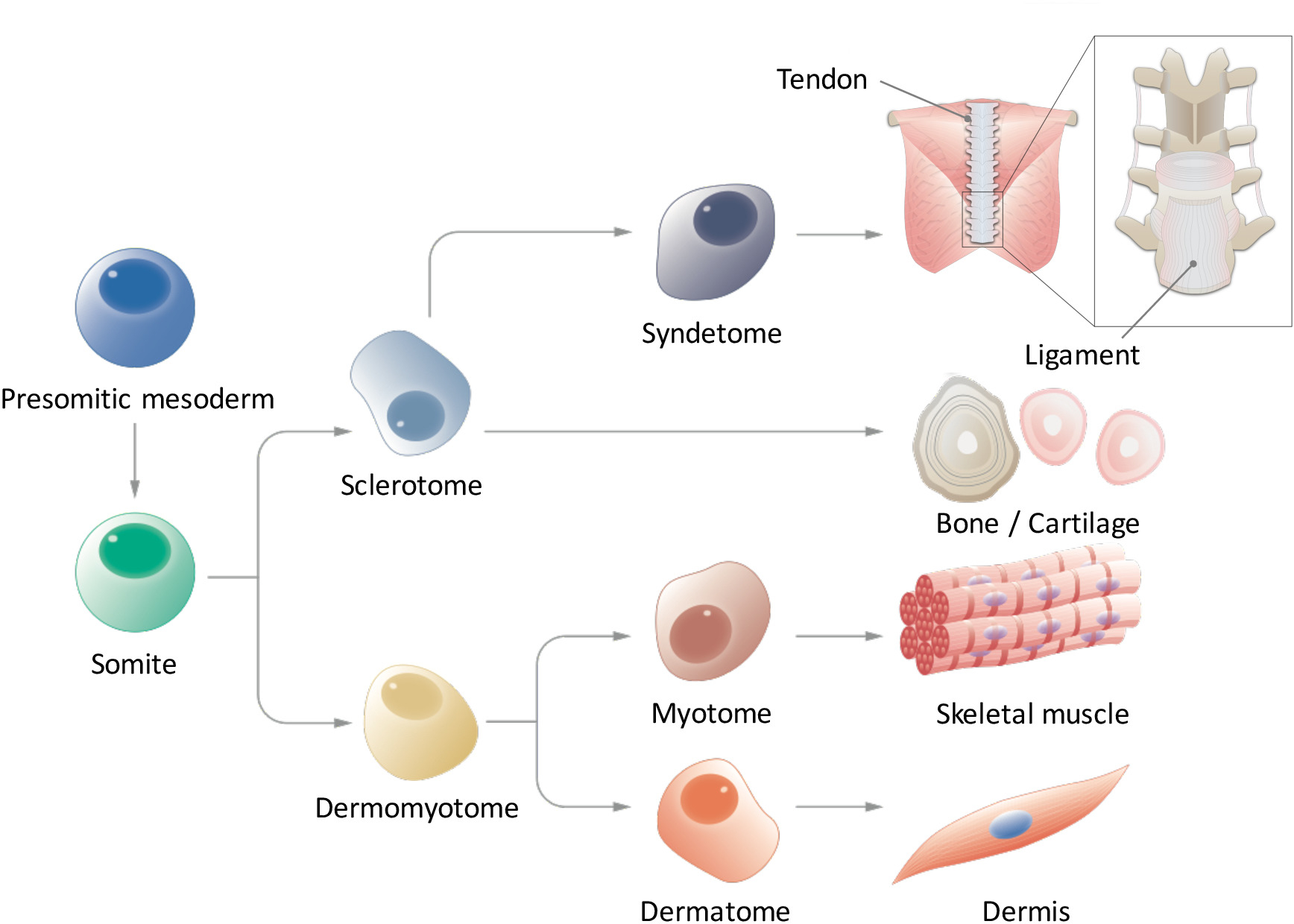{kind=link}

Şekil 2: insan IPSC kaynaklı PSM ve SM. FACS, RT-qPCR hem de ICC analizi (A)şematik SM farklılaşma PSM aracılığıyla için bir iletişim kuralı. (B) temsilcisi nokta arsa DLL1 boyama ve PAX3 (GFP)-PSM indüksiyon 4 gün ve gün 4 (gün 8 IPSC) SM indüksiyon floresan. (C) temsilcisi immunocytochemical görüntüleri ve PAX3 (GFP)-PSM indüksiyon 4 gün ve gün 4 (gün 8 IPSC) SM indüksiyon floresan. Hücreleri lekeli anti-TBX6, PARAXIS ve MEOX1 ile antikor (kırmızı) ve DAPI (mavi) ile birlikte lekeli veya PAX3 (GFP) ile tespit-floresan (yeşil). (D) için işaretleri PSM ve SM IPSC, adlı PSM ve SM RT-qPCR analizi. Deneyler üç kümelerinden anlamına gelir ± standart hata (S.E.) gösterilir. (E) temsilcisi immunocytochemical S10I10 içinde kültürlü SM, günde 4 (gün 8 IPSC) görüntüleri (SB431542 ve IWR1, WNT sinyal bir inhibitörü kombinasyonu), S10 (SB431542) ve S10C5 (SB431542 ve CHIR99021 birleşimi) koşulları. Hücreleri ile anti-CDH11 antikor (kırmızı) lekeli ve DAPI (mavi) ile birlikte lekeli. IPS, İndüklenmiş pluripotent kök hücre; PSM, presomitic mesoderm; SM, somite; S10, SB431542 10 MİKRON; C5, CHIR99021 5 MİKRON; I10, IWR1 10 MİKRON; Ölçek çubukları 50 mikron =. Bu rakam Nakajima et al. (2018)15değiştirildi. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
{kind=link}

Şekil 3: DM, MYO, D, SCL ve SYN ICC analizi ayrıştırılan insan IPSC kaynaklı SM. (A)iletişim kuralları için SM türevleri farklılaşma şematik. (B) temsilcisi immunocytochemical görüntüleri ve PAX3 (GFP)-gün 3 (gün 11 IPSC) DM indüksiyon, gün 30 (gün 41 IPSC) MYO indüksiyon, gün 9 (günden 20 IPSC) D indüksiyon, gün 3 (gün 11 IPSC) SCL indüksiyon ve gün 21 (gün 32 floresan IPSC) gelen SYN indüksiyon. DM, hücreleri anti-ALX4 ve EN1 antikor (kırmızı) ile lekeli ve lekeli DAPI (mavi) ile birlikte veya ile PAX3 (GFP) tespit-floresan (yeşil); MYO, hücreleri anti-Tanrım, MYOG (kırmızı) ve MHC (mavi) antikorları da DAPI (mavi); ile birlikte lekeli, lekeli D, hücreleri anti-EN1 (kırmızı) ve PDGFRa (mavi) antikorları ile lekeli ve DAPI (mavi); ile birlikte lekeli SCL, hücreleri edildi anti-PAX1, PAX9 ve NKX3.2 ile (kırmızı) antikorları lekeli ve DAPI (mavi); ile birlikte lekeli SYN, hücreleri edildi anti-SCX, MKX, COL1A1 ve COL1A2 ile (kırmızı) antikorları lekeli ve DAPI (mavi) ile birlikte lekeli. DM, dermomyotome; MYO, myotome; D, dermatome; SCL, sclerotome; SYN syndetome; Ölçek çubukları 50 mikron =. Bu rakam Nakajima et al. (2018)15değiştirildi. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
{kind=link}

Şekil 4: indüklenen D ve eşanlamlı fonksiyonel tahlil (A) proteinler kültür orta ELISA tarafından analiz edildi kollajen-tip 1 ve hyaluronik asit miktarı. (B) Mekanik streç etkisini uyarılma indüklenen SYN ve insan yetişkin tenocytes RT-qPCR tarafından değerlendirildi. Deneyler üç kümelerinden anlamına gelir ± standart hata (S.E.) gösterilir. * p < 0,05; ** p < 0,01; p < 0,001 Dunnett'ın en birden fazla karşılaştırmalar t-Testi karşılaştırıldığında streç (-); n.s, önemli değil, HDF, insan yetişkin dermal fibroblast. Bu rakam Nakajima et al. (2018)15değiştirildi. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
{kind=link}
| Orta/çözüm | Reagant | Konsantrasyon |
| CDM Bazal orta | Dulbecco'nın orta/Ham'ın F12 Iscove'nın modifiye | 1:1 |
| Penisilin/streptomisin | %0.5 | |
| Kimyasal olarak tanımlanan lipid konsantresi | % 1 | |
| APO-transferrin | 15 mg/mL | |
| Monothioglycerol | 450 mM | |
| Sığır serum albümin | 5 mg/mL | |
| İnsülin | 7 mg/mL | |
| CTK çözüm | Su | - |
| Tripsin | % 0.25 | |
| Collagenase IV | 0.1 mg/mL | |
| Kalsiyum klorür | 1 mM | |
| Nakavt SR | % 20 | |
| D indüksiyon orta | CDM Bazal orta | - |
| CHIR99021 | 5 MİKRON | |
| BMP4 | 10 ng/mL | |
| DM indüksiyon orta | CDM Bazal orta | - |
| CHIR99021 | 5 MİKRON | |
| BMP4 | 10 ng/mL | |
| ECM çözümü | Yapay hücre dışı matriks | 0.3 mg/mL |
| DMEM/F12 | - | |
| FACS arabellek | PBS | - |
| Sığır serum albümin | % 0.1 | |
| Besleyici-Alerjik hücre kültür orta | mTeSR1 | - |
| Penisilin/streptomisin | %0.5 | |
| HDF kültür orta | DMEM | - |
| Fetal sığır serum | % 10 | |
| hESC orta | Primat ES cep orta | - |
| Penisilin/streptomisin | %0.5 | |
| FGF2 | 4 ng/mL | |
| MYO indüksiyon orta | CDM Bazal orta | - |
| CHIR99021 | 5 MİKRON | |
| PSM indüksiyon orta | CDM Bazal orta | - |
| SB431542 | 10 ΜM | |
| CHIR99021 | 10 ΜM | |
| DMH1 | 2 ΜM | |
| FGF2 | 20 ng/mL | |
| SCL indüksiyon orta | CDM Bazal orta | - |
| SAG | 100 nM | |
| LDN193189 | 0.6 ΜM | |
| SM indüksiyon orta | CDM Bazal orta | - |
| SB431542 | 10 ΜM | |
| CHIR99021 | 5 MİKRON | |
| SYN indüksiyon orta-1 | CDM Bazal orta | - |
| FGF8 | 20 ng/mL | |
| SYN indüksiyon orta-2 | CDM Bazal orta | - |
| BMP7 | 10 ng/mL | |
| TGFβ3 | 10 ng/mL |
Tablo 1: Medya ve çözüm tarifleri.
| ADI | İleri | Ters |
| ACTB | CACCATTGGCAATGAGCGGTTC | AGGTCTTTGCGGATGTCCACGT |
| COL1A1 | GGACACAGAGGTTTCAGTGGT | GCACCATCATTTCCACGAGC |
| MEOX1 | GAGATTGCGGTAAACCTGGA | GAACTTGGAGAGGCTGTGGA |
| MSGN1 | GGAGAAGCTCAGGATGAGGA | GTCTGTGAGTTCCCCGATGT |
| PARAXIS | TCCTGGAGAGCTGTGAGGAT | CACACCCTGTCACCAACAGT |
| PAX3 | AGGAAGGAGGCAGAGGAAAG | CAGCTGTTCTGCTGTGAAGG |
| SCX | CCCAAACAGATCTGCACCTTC | GCGAATCGCTGTCTTTCTGTC |
| TBX6 | AGCCTGTGTCTTTCCATCGT | AGGCTGTCACGGAGATGAAT |
| TNMD | CCCTTCATGCTGAAGCCACTT | CTCACTTTCAGCAGAATTGGGG |
| WNT3A | CAAGATTGGCATCCAGGAGT | ATGAGCGTGTCACTGCAAAG |
Tablo 2: Primer RT-qPCR analiz dizileri.
| Konsantrasyon | ||
| 1. Antikor | ALX4_Goat | 1/50 |
| CDH11_Mouse | 1/1000 | |
| COL1A1_Rabbit | 1/100 | |
| COL2A1_Mouse | 1-2 μg/mL | |
| EN1_Rabbit | 1/50 | |
| MEOX1_Rabbit | 1/50 | |
| MHC_Rabbit | 1/200 | |
| MKX_Rabbit | 1/50 | |
| MYOD_Rabbit | 1/500 | |
| MYOG_Mouse | 1/400 | |
| NKX3.2_Rabbit | 1/50 | |
| PARAXIS_Rabbit | 1/50 | |
| PAX1_Rabbit | 1/50 | |
| PAX9_Rabbit | 1/50 | |
| PDGFRa_Goat | 1/100 | |
| SCX_Rabbit | 1/50 | |
| TBX6_Goat | 1/50 | |
| 2. Antikor | Eşek keçi IgG(H+L) ikincil antibody555 anti | 1/500 |
| Eşek keçi IgG(H+L) ikincil antibody647 anti | 1/500 | |
| Fare IgG(H+L) ikincil antibody555 anti keçi | 1/500 | |
| Keçi tavşan IgG(H+L) ikincil antibody555 anti | 1/500 | |
| Keçi tavşan IgG(H+L) ikincil antibody647 anti | 1/500 |
Tablo 3: İlk ve ikinci antikor ICC için.
Tartışmalar
PSC kaynaklı SM PSM aracılığıyla indüksiyon için iyi bilinen bir yöntem CHIR99021 + A83-01 (TGFβ inhibitörü) PSC üzerinden PSM indüksiyon sırasında ama PSM olgunlaşma süreci6sırasında birleşimidir. Bu da çalışmanın, WNT/beta-catenin sinyal, SM PSM üzerinden ikna etmek için C59 kullanarak inhibe. Ancak, WNT yolu SM farklılaşma sırasında etkinleştirmek için CHIR99021 kullanımı tanıtıldı. Bu kararı birkaç WNTs SM çevre dokular ifade ve WNT gazetecilere SM20dakika sonra etkin olduğu gerçeği göz önüne alındığında bulgu üzerinde temel alınmıştır. Sonuç olarak, biz epithelialization, SM içinde vivo, yalnızca koşulu ile CHIR99021, altında bir karakteristik CDH11 birikimi hücre-hücre kavşak (şekil 2E) temel alarak gözlenen. Bu gözlem sırasında PSM farklılaşma ve SM epithelialization, bu nedenle bizim iletişim kuralı daha iyi endojen sinyal çevre özetlemek sinyal WNT kritik tutulumu gösterir. Ancak, sağlamlık ve verimliliği farklılaşma hücre tipleri, hücre satırları ve çeşitli bağlı olarak önemli ölçüde değişebilir çünkü WNT/beta-catenin yol ayrımında sırasında sinyal ince ayar başka bir olasılık da ima WNT-indükleyicileri her araştırmacı tarafından kullanılan kimyasal bileşikler.
Bu yöntem aynı zamanda tüm dört SM türevleri, MYO, D, SCL ve SYN, insan iPSCs oluşturmak bize izin verir. Kademeli protokolümüze CDM kullanarak insan somitogenesis/biçimlenme somite sırasında sinyal gereksinimleri tanımlamak için kullanılan ve insan SM gelişme içine önemli bilgiler sağlar. Örneğin, bizim yöntemleri segmentasyon saat mekanizmaları, SM oluşumunu düzenleyen moleküler salınım sistemi eğitim için yararlı olabilir. Bunu iyice fareler, civciv ve Zebra balığı ama uygun deneysel araçları eksikliği nedeniyle değil insanlarda araştırmış.
Ayrıca, bizim yöntemi gelecekteki klinik hücre tabanlı terapiler için geçerli olabilir. Örneğin, insan IPSC türetilmiş D veya SYN ağır yaralı deri içine nakledilen veya rejenerasyon ve tedavi için tendonlar yırtıldı. Ancak, bazı sınırlamaları bu yöntem hemen hemen uygulanabilir önce giderilmesi gerekiyor. Bu da çalışmanın, SNL besleyici hücreler IPSC bakım ve Engelbreth-Holm-Swarm fare sarkomu indüksiyon sırasında çanak üzerinde yüzey bir kat olarak ayıklanır ECM çözümü için kullandığımız rağmen bu insan hayvan kaynaklı reaktifler olmalıdır Klinik kalitesini artırmak için kaldırıldı. Buna ek olarak, hücre miktarı ve kalitesi, saflık ve olgunlaşma istediğiniz hücreleri içeren aynı zamanda iyileştirilmesi gerektiği. Ayrıca, sadece cep numarası aynı zamanda hücre gücü tendon/bağ rejenerasyon için önemli bir özelliği değildir. Ayrıca, arıtma için yüzey işaretleyicileri ve 3D sulandırma için yeni bir yöntem geliştirilmesi amacıyla protokolümüze klinik hücre tabanlı terapiler için ilerlemek için vazgeçilmez.
Açıklamalar
Yazarlar ifşa gerek yok.
Teşekkürler
Biz Dr Junya Toguchida (CiRA) için proje yönetimi ve satın alma, Bay Mitsuaki Shibata (CiRA) ve Bayan Mei Terashima (CiRA) onların teknik destek için Dr Yayoi Toyooka (CiRA) ve Dr. Daisuke Kamiya (CiRA) finansman ile onun yardım için teşekkür etmek istiyorum onların el yazması ve Bay Masaya Todani (CiRA) bir örnek (şekil 1) sağlamak için yazım denetleme. Biz de bu çalışma sırasında Ikeya ve Toguchida Laboratuvarları (CiRA) desteklerinden dolayı tüm üyeleri teşekkür ederiz. Bu eser Grants-in-aid tarafından Japonya Derneği bilimsel araştırma için promosyon, bilim (JSP'ler) (26670661), dirençli hastalıklar araştırma kullanan hastalığa özgü IP'leri hücrelerden Japonya bilim ve teknoloji için Program desteklenmiştir Ajansı (JST) ve tıbbi araştırma ve geliştirme (AMED) Japonya ajansı, çekirdek Merkezi IP'leri hücre araştırma Araştırma Merkezi ağ, rejeneratif tıp gerçekleşme (JST/AMED) için ve IPS Hücre Araştırma Fonu (kısmen Makoto Ikeya için ve JUNYA Toguchida). Makoto Ikeya Ayrıca Grants-in-aid tarafından bilimsel araştırma (JSP'ler) (16 H 05447) ve Hızlandırma programı için dirençli hastalıklar hastalığa özgü IP'leri hücreleri (AMED) kullanan araştırma için desteklenen bir durumdu.
Malzemeler
| Name | Company | Catalog Number | Comments |
| ALX4_Goat antibody | Santacruz | sc-22066 | |
| Apo-transferrin | Sigma | T1147 | |
| BMP4 | R&D | 314-BP-010 | |
| BMP7 | R&D | 354-BP-010 | |
| Bovine serum albumin | Sigma | A8806 | |
| Calcium chloride | Nacalai tesque | 067730-15 | |
| CDH11_Mouse antibody | Cell signaling | 13577 | |
| Cell streching device | Strex | STB-140 | |
| Chemically defined lipid concentrate | Gibco | 11905-031 | |
| CHIR99021 | Axon | 1386 | |
| COL1A1_Rabbit antibody | Abcam | ab34710 | |
| COL2A1_Mouse antibody | Thermo scientific | MS-235 | |
| Collagenase IV | Thermofisher | 17104019 | |
| DLL1 APC-conjugated_Mouse antibody | R&D | FAB1818A | For FACS |
| DMEM | Sigma | D6046 | |
| DMEM/F12 | Gibco | 11320-082 | |
| DMH1 | Tocris | 4126 | |
| EN1_Rabbit antibody | Abcam | ab70993 | |
| Fetal bovine serum | Nichirei | 171012 | |
| FGF2 | Wako | 060-04543 | |
| FGF8 | Peprotech | 100-25 | |
| Human dermal fibroblast | Cell applications | 160-05a | |
| Human tenocyte | Angio proteomie | cAP-0041 | |
| Insulin | Wako | 090-06474 | |
| Iscove’s modified Dulbecco’s medium/Ham’s F12 | Gibco | 21056023 | |
| Knockout SR | Gibco | 10828028 | |
| LDN193189 | Axon | 1509 | |
| Matrigel | BD bioscience | 354230 | Artificial extracellular matrix |
| MEOX1_Rabbit antibody | Abcam | ab75895 | |
| MHC_Rabbit antibody | Santacruz | sc-20641 | |
| MKX_Rabbit antibody | Atlas antibodies | A83377 | |
| Monothioglycerol | Sigma | M6145 | |
| mTeSR1 | Stemcell tech | 85850 | |
| Multi well-type silicon rubber chamber | Strex | STB-CH-4W | |
| MYOD_Rabbit antibody | Abcam | ab133627 | |
| MYOG_Mouse antibody | Santacruz | sc-12732 | |
| NKX3.2_Rabbit antibody | Sigma | HPA027564 | |
| Novex Donkey anti Goat IgG(H+L) secondary antibody555 | Invitrogen | A21432 | |
| Novex Donkey anti Goat IgG(H+L) secondary antibody647 | Invitrogen | A21447 | |
| Novex Goat anti Mouse IgG(H+L) secondary antibody555 | Invitrogen | A21422 | |
| Novex Goat anti Rabbit IgG(H+L) secondary antibody555 | Invitrogen | A21428 | |
| Novex Goat anti Rabbit IgG(H+L) secondary antibody647 | Invitrogen | A21245 | |
| PARAXIS_Rabbit antibody | Santacruz | sc-98796 | |
| PAX1_Rabbit antibody | Abcam | ab95227 | |
| PAX9_Rabbit antibody | Gene tex | GTX104454 | |
| PBS | - | - | |
| PDGFRa_Goat | R&D | AF307 | |
| Penicillin/Streptomycin | Invitrogen | 15140-122 | |
| Primate ES cell medium | Reprocell | RCHEMD001 | |
| SAG | Calbiochem | 566661 | |
| SB431542 | Selleckchem | SEL-S1067-10 | |
| SCX_Rabbit antibody | Abcam | ab58655 | |
| TBX6_Goat antibody | R&D | AF4744 | |
| Tendon cell growth medium | Angio-proteomie | cAP-40 | Tenocytes growth medium |
| TGFβ3 | R&D | 243-B3-200 | |
| Trypsin | Gibco | 15090046 |
Referanslar
- Tanaka, A., et al. Efficient and reproducible myogenic differentiation from human iPS cells: prospects for modeling Miyoshi Myopathy in vitro. PLoS One. 8 (4), e61540 (2013).
- Sakurai, H., et al. In vitro modeling of paraxial mesodermal progenitors derived from induced pluripotent stem cells. PLoS One. 7 (10), e47078 (2012).
- Chal, J., et al. Generation of human muscle fibers and satellite-like cells from human pluripotent stem cells in vitro. Nature Protocl. 11 (10), 1833-1850 (2016).
- Chal, J., et al. Differentiation of pluripotent stem cells to muscle fiber to model Duchenne muscular dystrophy. Nature Biotechnology. 33 (9), 962-969 (2015).
- Umeda, K., et al. Human chondrogenic paraxial mesoderm, directed specification and prospective isolation from pluripotent stem cells. Scientific Reports. 2, 455 (2012).
- Loh, K. M., et al. Mapping the Pairwise Choices Leading from Pluripotency to Human Bone, Heart, and Other Mesoderm Cell Types. Cell. 166 (2), 451-467 (2016).
- Xi, H., et al. In Vivo Human Somitogenesis Guides Somite Development from hPSCs. Cell Reports. 18 (6), 1573-1585 (2017).
- Chal, J., Pourquie, O. Making muscle: skeletal myogenesis in vivo and in vitro. Development. 144 (12), 2104-2122 (2017).
- Tam, P. P., Beddington, R. S. The formation of mesodermal tissues in the mouse embryo during gastrulation and early organogenesis. Development. 99 (1), 109-126 (1987).
- Aulehla, A., Pourquie, O. Signaling gradients during paraxial mesoderm development. Cold Spring Harbor Perspectives in Biology. 2 (2), a000869 (2010).
- Christ, B., Scaal, M. Formation and differentiation of avian somite derivatives. Advances in Experimental Medicine and Biology. 638, 1-41 (2008).
- Brent, A. E., Schweitzer, R., Tabin, C. J. A somitic compartment of tendon progenitors. Cell. 113 (2), 235-248 (2003).
- Sakurai, H., et al. Bidirectional induction toward paraxial mesodermal derivatives from mouse ES cells in chemically defined medium. Stem Cell Research. 3 (2-3), 157-169 (2009).
- Zhao, J., et al. Small molecule-directed specification of sclerotome-like chondroprogenitors and induction of a somitic chondrogenesis program from embryonic stem cells. Development. 141 (20), 3848-3858 (2014).
- Nakajima, T., et al. Modeling human somite development and fibrodysplasia ossificans progressiva with induced pluripotent stem cells. Development. 145 (16), (2018).
- McMahon, A. P., Bradley, A. The Wnt-1 (int-1) proto-oncogene is required for development of a large region of the mouse brain. Cell. 62 (6), 1073-1085 (1990).
- Takahashi, K., et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 131 (5), 861-872 (2007).
- Suzuki, H., et al. targeting of the transcription factor Mohawk in rats causes heterotopic ossification of Achilles tendon via failed tenogenesis. Proceedings of the National Academy of Sciences of the United States of America. 113 (28), 7840-7845 (2016).
- Marturano, J. E., Arena, J. D., Schiller, Z. A., Georgakoudi, I., Kuo, C. K. Characterization of mechanical and biochemical properties of developing embryonic tendon. Proceedings of the National Academy of Sciences of the United States of America. 110 (16), 6370-6375 (2013).
- Maretto, S., et al. Mapping Wnt/beta-catenin signaling during mouse development and in colorectal tumors. Proceedings of the National Academy of Sciences of the United States of America. 100 (6), 3299-3304 (2003).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır