
Method Article
C. elegans 中大型外层囊中体内神经聚合和有机体挤出评分的定量方法
摘要
该协议描述了检测和定量由 C.elegans细胞以膜结合 外外层形式产生的大型聚集和/或细胞器挤出(±4 μm)的方法。我们描述了菌株、生长条件、评分标准、时间和显微镜注意事项,以便于解剖这种碎片驱逐机制。
摘要
错误折叠的蛋白质和线粒体功能障碍的毒性是促进年龄相关的功能神经元衰退和跨物种神经退行性疾病的关键因素。虽然这些神经毒性挑战长期以来一直被认为是细胞内在的,现在有相当多的证据表明,源自一个神经元的误折叠的人类疾病蛋白质可能出现在邻近细胞中,这种现象被提出来促进人类神经退行性疾病的病理学传播。
C. elegans 表示聚合蛋白质的成年神经元可以挤出大(±4 μm)膜包围的囊泡,其中可包括聚合蛋白、线粒体和卵母体。这些大囊泡被称为"外层",不同于外体(其体积约100倍,具有不同的生物发生)。将细胞碎片抛出外层可能由一种保存机制发生,该机制构成神经元蛋白位突虫病和线粒体质量控制的一个基本但以前未被识别的分支,与聚合在人类神经退行性疾病中传播的过程相关。
虽然外显子主要在接触神经元中表达高拷贝转基因mCherry的动物中进行了研究,但这些协议在研究使用荧光标记的细胞器或不同神经元类别中感兴趣的其他蛋白质的外生基因时同样有用。
这里描述的是 C.elegans 外外层的物理特征,其检测策略,识别标准,定量的最佳时间,以及动物生长协议,控制压力,可以调节外层生产水平。此处概述的协议细节应有助于建立跨实验室外外层定量分析的标准。本文件旨在为实验室提供该领域的资源,以寻求分析产生外外层和外外pher的分子机制,以及外外pher由邻近和远距离细胞作出反应的机制。
引言
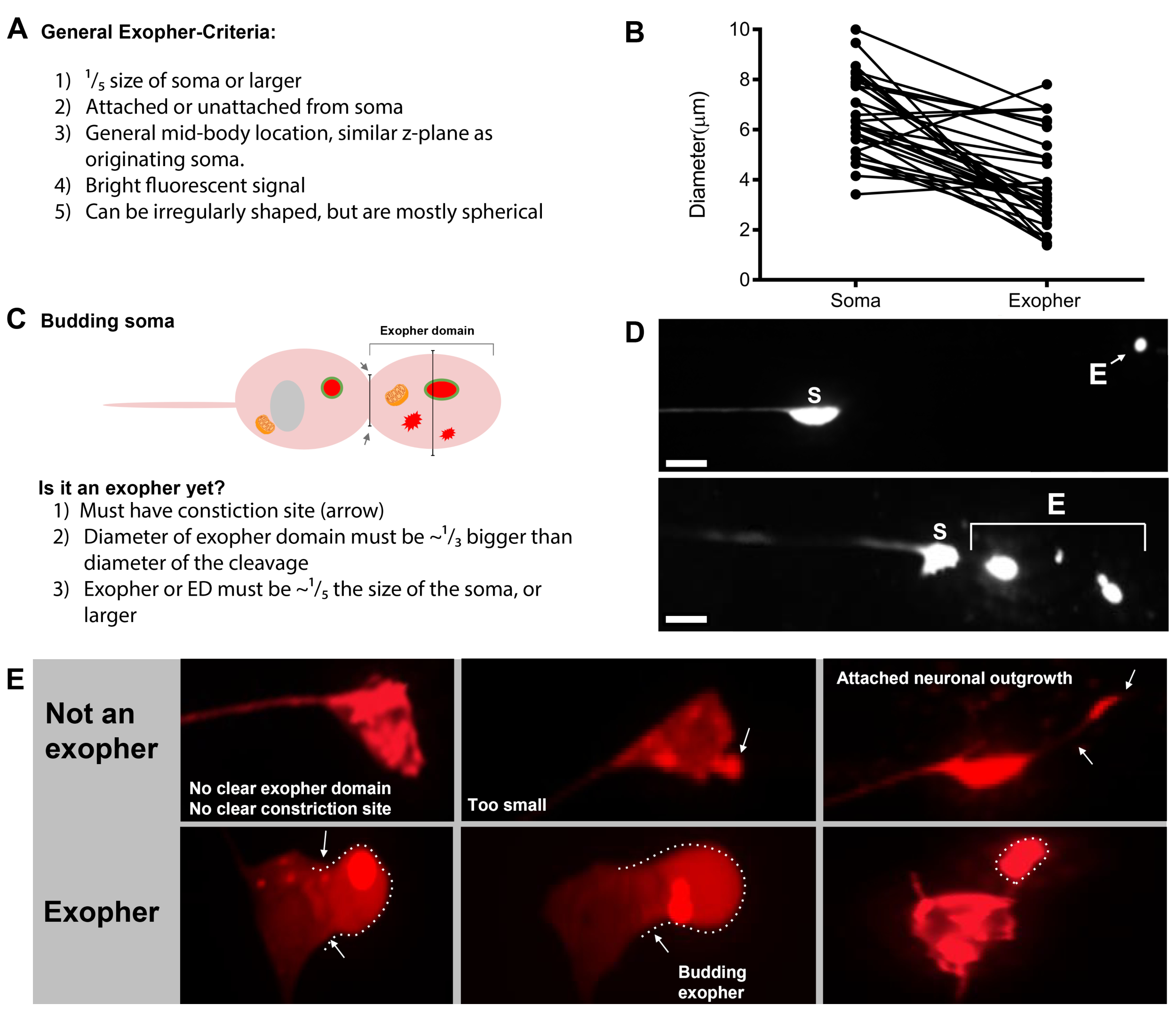
聚合和功能失调的线粒体的神经毒性挑战长期以来一直被认为是细胞内在的,但最近很明显,源自一个神经元的误折叠的人类疾病蛋白质也可以传播到邻近的细胞,促进病理学1。同样,哺乳动物线粒体可以被送出其原始生产的细胞,用于变性降解2 或拯救受挑战相邻细胞3中的线粒体种群。一般观察到各种大小的囊泡将细胞物质转移到邻近的细胞或流体周围4。一些挤出的囊泡接近平均神经元母体的大小(平均接触神经元索玛 = 6 μm),可以容纳大型聚合体和细胞器。
一个可以携带蛋白质聚合物和细胞器的大囊泡挤压的显著例子出现在C.elegans触摸受体神经元中,这些神经元表示一个高拷贝数报告器构造编码一种有毒聚集易发、抗降解的mCherry5。来自触摸神经元的挤出,称为外向,平均直径为±4μm,选择性地包括mCherry或其他聚合体,并直接传递到相邻的皮下,通常围绕触摸受体神经元。皮下试图以碱体为基础的降解,但一些不可消化的内容,如mCherry聚合物,可以重新挤出到动物的液体填充伪细胞,从中,mCherry可以通过远程清除细胞称为共糖细胞长期储存(图1,图2)5。5
大型挤出外层囊泡离开细胞被触摸受体血浆膜包围,可以含有聚合的人类疾病蛋白质、线粒体和利索索姆。外外层生产的过程似乎涉及对潜在有毒物种的分类(例如,容易聚集的表达mCherry与可溶性、无攻击性的蛋白质(如GP)分离,这些蛋白质大多留在神经元索马中)。这样,定向驱逐威胁实体是由神经元5完成的。一种蛋白质组虫的挑战,如自噬敲击引起的应激,MG132中显蛋白体抑制,或人类疾病蛋白质的转基因表达,如亨廷顿舞蹈症相关扩大多糖胺Q128或阿尔茨海默氏症牵连片段 A+1-42,可以增加产生外向球的神经元的数量。
由于外外phers最近才被记录,关于它们的生物学已知情况值得描述。外层在 C.elegans触摸受体神经元中被发现,并且是研究 最良好的。有六 个C.elegans 机械感官触摸神经元,有细胞体分布在身体周围(图3A),被称为微管细胞,因为它们的超结构特征独特的15个原丝微管。触摸受体神经元为前前AVM(前前前前前前微管神经元)、ALMR和ALML(右前侧微管神经元)和后部PVM(后侧前前前侧微管神经元)和尾部PLMR和PLML(右后侧显微管神经元)。有趣的是,六个触摸受体神经元产生不同的速率外向,尽管表达相同的攻击性转基因(图3C)。在六个机械感官触摸受体神经元中,ALMR神经元比其它触摸神经元更频繁地经历外向生成。因此,从触摸神经元中对外层数字的定量通常是通过聚焦ALMR建立的。
外向发生是一个动态过程,通常从神经元细胞质肿胀开始(图1A-B)。细胞内,细胞器或蛋白质聚集物被收集到神经元的一侧,最常见的是ALMR神经元的后端(远离投射的神经),形成一个前外向域(PED)(图1B)。当 PED 开始向外投射时观察到早期突起,形成可识别的突出芽。当前外驱域的最宽直径比索马外传颈部收缩的直径大约1/3时,定义晚芽(图1C)。外外层可以从索马向几乎任何方向喷射,但大多数外显子从细胞体后部退出,并留在与原体大致相同的焦点平面中。
外外pher可以远离原发的索玛,因为芽的脖子变窄成细丝。外层可以通过这种灯丝保持附着在索玛上(图1D,箭头),以后可以分离。钙、骨和线粒体的细胞含量可以通过这种灯丝转移到附着的外层5中,尽管大部分挤出材料被巨大的萌芽事件放入外层隔间。当没有明显的连接管或细丝和外显子与发送的索马完全分离时,外显子被认为是成熟的(图1E)。
C. elegans 产生的外向器触摸神经元会立即遇到皮下,即触摸神经元周围的组织。最常见的情况是,外皮囊泡似乎在皮下后向尾部运动,在外皮物含量出现降解目标之前,它们可能离母细胞相当远(例如,距离母细胞的距离可以约100μm(图1F)。荧光外层囊泡在皮下分解成许多较小的囊泡,外观称为"星夜"(图1G和图2)。在"星夜"阶段,与原始的单独外显器相比,可以观察到分散在皮下同步的荧光材料分散到许多较小的荧光点中。星夜可以在低放大倍率下看起来更刺耳,放大率更高,可以看起来在皮下放大和/或网络化。星夜的荧光信号通常比外音和神经元表达的荧光更暗(图2B-C)。mCherry 的分散到许多双核囊泡被认为涉及噬菌体成熟和与皮下细胞的内索体/乳糖细胞的融合。一些外皮物质很可能在皮下体利索索马网络中降解,但抗降解的残余物种(如mCherry集料)被从皮下物质中扔进伪细胞,这是一种液体舱,可以含有细胞碎片。荧光材料后来被称为细胞核糖的远程清除细胞(图2C)所利用,这种细胞可以浓缩、储存并再次尝试降解mCherry。
聚合挤压和转移现象似乎在植物之间得到保存,,在基因模型中,如C.elegans5、6、7,6和D.黑色素78,9,9以及多个哺乳动物模型中都报告了。据报道,哺乳动物细胞10的外向状挤出,这一观察表明,保存的机制可能是聚合和细胞器驱逐的根据。因此,外向器生产可能是细胞碎片管理的一种保存机制,构成神经元蛋白位和线粒体质量控制的一个基本但以前未被识别的分支,当不平衡时,可能积极促进神经退行性疾病。识别碎片识别和分拣所涉及的分子,将物质运送到一个独特的亚细胞区域,挤压,形成/剪刀状的管状连接连接的索马和晚期外外层,并识别大挤出的囊泡,供邻近细胞远程降解。线虫和苍蝇模型的研究对于确定聚合和细胞器收集和转移的机制至关重要,利用这些模型提供的无偏遗传方法和强大的细胞生物工具来识别生理环境中的参与分子。
在外层生物学中操作的破译机制的关键第一步涉及定义在体内外 外层定量可 重复的协议。 C. elegans 模型 为此类工作提供了特殊优势,因为身体是透明的,当外显体含有荧光标记的蛋白质或细胞器时,可以很容易地观察到。据报道,外显细胞是由 C.elegans 多巴胺神经元PDE和CEP,ASE和ASER感觉神经元,和染料填充的安非他明神经元5生成。由于触摸受体神经元产生的外显子是最佳特征,因此这里的重点是使用触摸神经元进行外外层分析。然而,基本方法可以应用于测量来自任何细胞的外层产生。对 C.elegans 接触受体神经元产生的外向进行检测和定量的规程,从基因上表达mCherry蛋白,并概述可监测的货物和评分时的时间限制。本文定义了体内外 外层识别 的方法,以及调节外外外外层生产的环境和遗传条件的定量方法。议定书强调对恒定的非应力条件给予关键关注,以便确定基线外外层生产,并进行比较基因型。
研究方案
1. 可用于外外层检测的应变
- 选择一种在 C.elegans神经元内表达荧光货物的菌株 ,以便于可视化外显子。
注:表1列出了用于可视化接触受体神经元5、11、12,中产生的,外显器的菌株。原则上,任何细胞或神经元类型都可以通过使用细胞或组织特异性启动子来驱动荧光蛋白的表达,该蛋白聚合或以其他方式被选为挤出。 - 或者,使用染料填充测定来可视化安非他明头神经元中的外显器,这些外向神经元对环境开放,并且可以回填5,5,13。
2. 增长介质
- 根据标准方法14、15,为培养菌株准备标准线虫14生长介质(NGM)。
注:缺乏食物,或氟-脱氧核糖核酸(FuDR),通常用于阻止后代生产,并可以显著影响外层生产。保持种群持续喂养(避免短时间细菌性食物耗尽),并保持动物在恒定的温度。
3. 畜牧业对于一致的外外层生产至关重要
- 在一致的介质和一致的细菌食物来源下饲养动物。动物 不得 耗尽细菌性食物,即使是很短的时间,因为食物限制可以显著改变外在的产生水平。
- 在整个研究中保持媒体配方和准备统一。
注:更换介质会影响外层生产的基底水平。Agar 批次会影响基线外向值水平,因此当供应批次发生变化时,请记下日期。两周后扔掉库存盘,以确保健康的细菌性食物,并防止干性阿加,这会导致影响外外体水平的果糖渗透性变化。 - 对于基础条件,将动物保持在20°C的恒定温度下。 在可变温度下饲养动物(即使是温度的暂时变化)会导致最大外外外层生产时间的变化。
注:温度变异性不限于培养条件。实验期间或实验室工作台上的温度变化可能很有影响。例如,显微镜室内的温度不应与培养箱或实验室工作台有显著差异。 - 不要使用药理性抗生育干预措施,因为受精卵对早期成人产生外向体至关重要。
注:必须避免使用氟-脱氧核糖核酸(FuDR)16 或C22 17。在进行寿命延长或老年动物实验时,应通过将成年人从较小的后代身上取出,将成年人从较小的后代身上取出,从而维持年龄同步种群,将它们采摘到用细菌传播的新鲜盘子上,而不是使用常见的药理学抗生育干预措施。 - 不要使用受污染的培养物;在种群或板块发生生物损害时重新启动实验。细菌或真菌污染可诱发动物的应力和代谢变化,实验人群必须不存在。
- 为了最大限度地提高可重复的结果,在实验前,在20°C下至少保持两个健康、喂养良好、无污染的世代培养,以避免潜在的环境诱发表观遗传变化。
4. 通过漂白、蔗糖浮选或 L4 幼虫采摘进行外向评分的年龄同步
- 保持实验种群相同的生物年龄,因为外在检测模式随成年年龄而变化,对混合年龄动物的比较可能会混淆结果。始终通过检查L4阶段的"白新月"外阴形态,确保实验动物种群的成功同步。
注:一般来说,C.elegans机械性ALMR神经元的峰值外向产生发生在成人第2-3天(图3D),从L4阶段后, 几天开始测量。成人第1天是L4幼虫阶段后24小时,该阶段以"白色新月"外阴形态区分(图5E)。 - 通过漂白成年的成年准备同步的卵子种群。
- 通过清洗生长在NGM盘子上的动物来收集装满鸡蛋的格拉维德成年人。要清洗,用1 mL的M9缓冲液充水板,上下移液收集悬浮动物的液体,并移入1.5 mL微离心管。通过重力沉降或用迷你离心机轻轻离心,去除上清液。
- 在 H2O 中的 1 mL 中加入 150 μL 的 5M NaOH 和 150 μL 的 6% 次氯酸钠(漂白剂),通过反转混合约 5 分钟。
注:新鲜的漂白溶液确保动物角质层可以被破坏的卵子收获。可以在解剖显微镜下监测破坏角质层的进展;成年人应该打破和释放鸡蛋点, 应该停止漂白。 - 用迷你离心管轻轻离心 20 s,然后取出上流水。加入 1 mL 的 M9 缓冲液并再次离心,在颗粒上留下约 100 μL。
- 重复步骤 4.2.3 两次,以去除漂白剂溶液的痕迹。
- 将鸡蛋在剩余体积中重新暂停,并转移到新鲜种子的NGM板。成人将被冷冻,但许多可行的鸡蛋应该在准备。
- 通过时间煮蛋准备同步种群。
- 使用标准转移协议14挑选20名成年成人到种子NGM板。
- 允许动物自由爬行和产卵1.5小时(低育雏大小的突变菌株可能需要引入更多的成年动物)。
- 通过采摘将所有成年动物从盘子里取出,将同步的蛋组抛在脑后。几个小时后检查车牌,以确认在成人搬迁期间没有遗漏任何可行的成人。
- 通过蔗糖浮选选择准备同步的蛋群。
- 从五个NGM盘子中收集动物和卵子,在5个NGM板上,格拉维德动物已经产卵至少24小时,用0.1%的洗涤剂(如Tween 20或Triton X-100)将盘子淹没在M9溶液中,并收集到15 mL管中。在室温下温和离心(30 s 2,000 x g)使 成人小气。
- 去除上经液,在15 mL的新鲜M9中清洗动物三次,每次洗涤后丢弃上经剂,确保保持富含动物和鸡蛋的颗粒。
- 保留 2 mL 的上重物,然后重新暂停颗粒。加入2 mL的60%的重量体积蔗糖。
- 在2000 x g下离心 5分钟。该解决方案现在将显示一个上相高度丰富的鸡蛋。
- 将上相约2.5 mL转移到新的15 mL管中,并加入10 mL的M9。
- 通过反转混合1分钟,然后离心2000xg1分钟。 g
- 取出上清液,在M9中清洗富含鸡蛋的颗粒。10-15 μL的蛋丸可以分发到新鲜的OP50种子NGM板。
注:这种方法准备大量的鸡蛋;不允许收集的动物用完OP50 大肠杆菌食物 。
- 在L4发展阶段采摘动物,为同步种群做好准备。
- 如上文所述,在种子的NGM板上生长动物。
注: C. elegans 在 四个离散阶段发展。在20°C下,新产卵孵化大约需要9个小时(图5A)。舱口后,动物通过幼虫阶段1(L1)到幼虫阶段4(L4),每个阶段在每个摩尔之间持续8-12小时(图5A-F)。因此,用鸡蛋接种准备的盘子在引入卵子后40小时内应有许多L4动物来采摘。 - 通过定位正在发育的外阴的白色半月牙形状来识别L4级动物(图5E)。
注:L4阶段的动物在大小和身体色素沉着方面是均匀的。挑选白色新月的动物到新的生长板,以检查分阶段的动物。第二天(24小时后)应计为成人第1天。 - 在成人日第 2 天每天对动物数量进行评分。
注:外显子通常在成年后的第2天得分,这是基底条件下的外外层产生高峰。然而,由于外生动物的产生高峰和时间可能因正在研究的环境或基因变化而改变,建议在四天内每天对成年动物进行评分,以生成最全面的画面(图3D)。
- 如上文所述,在种子的NGM板上生长动物。
5. 使用荧光显微镜检测外外层
- 使用高倍率伪立体解剖显微镜观察外显器,该显微镜适合进行荧光显微镜。
- 通过将 10-200 μL 的 10-100 mM levamisole/四米索溶液移液到 NGM 加糖板表面,将动物固定在 NGM 板上。2-4分钟后,动物瘫痪,可以直接在阿加盘上观察。
注:固定化治疗并非绝对需要,因此,在确定是否产生外外层时,可以通过在板的显微镜下目视跟踪爬行动物来评分,这样,对于训练有素的眼睛,神经元识别和外外层的存在可以进行评分。 - 使用总放大倍数 100 倍观察荧光神经元,完成对外外层进行解剖显微镜检测。
注:使用解剖显微镜对外层事件进行评分,可以相对容易地观察大量动物,直接在它们饲养的阿加盘子里。 - 使用共声显微镜进行外外层研究的实时成像和安装记者应变
- 使用共生显微镜对细胞内动态和外显性特征进行活图像成像。
注:实时成像是观察外外层生产的微妙细节的有利方法,因为外外影像生产是一个动态过程。 - 使用方便的方法限制动物移动进行高分辨率活成像,包括利用 10-100 mM 的利卡米索或四米索或水凝胶聚苯乙烯微珠(直径为 15 μm、30 μm 或 40 μm)18。
- 使用共生显微镜对细胞内动态和外显性特征进行活图像成像。
- 复合显微镜和共共体显微镜的滑动准备
- 将 20-50 只动物安装到显微镜幻灯片上的固定剂中。可重复使用的环形细胞学幻灯片,涂有 13 mm 直径的凸起环,可用于安装。
- 将活的动物挑选成 5-20 μL 的麻麻麻膜,如 10-100 mM levamisole 或四米索在彩绘圆圈内或在阿加垫上。
- 等待 4 分钟,否则请稍等 4 分钟,然后用盖玻片(建议 11+2 号(0.16 × 0.19 mm)或 2 号(0.17 - 0.25 毫米)盖住幻灯片。
- 安装少量动物
- 不要压碎安装过的动物;当每张幻灯片只观察少数(少于 20 种)动物时,由于盖玻片的不等压力,可能会压碎一些动物。通过使用低百分比的 agarose 垫进行安装,可以最大限度地降低此风险。
- 制作 2-4% 的 agarose 垫幻灯片,然后向垫中加入 2-15 μL 的麻麻溶液。请记住,利瓦米索和四米索扩散到垫,降低其有效浓度。
- 通过将动物采摘到 2-15 μL 滴的麻麻溶液或微珠放在阿加垫上进行安装。将盖玻片放在上面,检查动物是否完好无损。
- 阿加垫准备
- 为了准备2%的加糖垫,在M9溶液和微波中加热2%的加糖,直到加糖处于均匀和熔融状态。
- 为了实现足够质量的搅拌垫,在低功耗下交替混合和微摇,时间少于 20 秒。通过将沸腾的 agar 放在加热块上,让气泡上升到表面,避免在垫中加入气泡。
- 使用巴斯德移液器从上升气泡下方熔融溶液深处的熔融溶液中绘制糖。
- 准备两张带胶带的幻灯片,放在干净的玻璃显微镜幻灯片的两侧,放在平坦的表面上。使带状的幻灯片在每张幻灯片上放置两条 5 厘米的实验室胶带(图 6A)。
- 使用巴斯德移液器,将一滴糖放在夹在胶带滑梯之间的清洁显微镜幻灯片上(图 6B)。
- 小心而快速地,通过放置在胶带的幻灯片上,用第四张干净的幻灯片盖住熔融的阿加的掉落(图6Cc)。
注:幻灯片应轻轻地将熔融的搅拌压入约 0.4 mm 厚(胶带厚度)的扁平圆(图6D)。阿加应该快点冷却。 - 滑动顶部幻灯片(图6E)将其拆下。Agar 垫干燥迅速,最好在几分钟内使用。拆下顶部滑轨后,立即使用凝胶垫安装动物。避免使用带气泡的垫。
- 将阿加垫存放在两个玻璃滑梯之间长达 30 分钟。干的阿加使动物聚集在一起干燥。将动物安装在2-15μL的麻麻溶液或微珠内,并盖上盖板;在瘫痪和安装后 20 分钟内筛选幻灯片(图 6)。
注:由于应激条件可以改变外向率,避免在筛选外外层时引起氧化应激(例如:阿齐德钠)的麻石。
- 使用旋转盘共体显微镜检测外泌体
- 观察细胞生物特征,如细胞器和其他内容物,在63倍和100倍时具有1.4个数值孔径目标。
- 使用能够利用多维采集进行舞台控制和图像采集的软件。显微镜和图像处理软件还应适合成像和数据收集,因为这些步骤涉及标准成像方法。
6. 识别触摸神经元,并评分外向动物与安装动物
- 山瘫痪的成年动物(图6)。
- 确定所需的 Z 平面。使用低放大倍率亮场(10-40 倍)来识别动物的合适 Z 平面,注意动物的定位、头尾方向和外阴的位置 - 这是后期神经元和外外显子识别的地标(图3A 和图 5E)。
- 关注所选记者的荧光信号。在同一个 Z 平面上,切换到广场荧光观察 10-40 倍为选定的细胞细胞学记者。
注:在此示例中,荧光表达由 mec-4 机械感官触摸神经元特异性启动子驱动。高拷贝阵列和不同的荧光团在表达上具有变异性,因此荧光强度也可变。如果需要,请进行调整。 - 在 Z 轴内滚动,观察焦点平面中动物和荧光表达的深度。这样做时,确认头尾方向;头部/咽部将具有荧光神经环,在这种情况下,尾部将包含1-2个可见的PLM索马(图3A)。
- 识别触摸神经元
- 确定动物是安装在左侧还是右侧(图3A)。
注:考虑到动物的三维性,最佳成像分辨率在最接近光学元件的一侧完成。 - 通过观察识别索玛(ALM、ALMR、AVM)- 从头部开始识别神经环和横向神经元过程。
- 在 10-40 倍放大倍率下,缓慢滚动 Z 轴以识别所连接的过程。
- 一旦确定过程,横向跟随它朝后向外阴,在那里索马将明显,在过程结束时以圆形细胞体为标志。一旦发现最聚焦的神经元母细胞,可以使用其他神经元地标进行识别,如下所示:
- 使用 AVM, 附近的腹神经元,以帮助分配动物方向。如果AVM神经元与ALM位于同一平面上,则动物将侧靠,而平面外的神经元是ALMR。如果 AVM 神经元与该问题 ALM 不在同一平面上,则最接近焦点平面的触摸神经元是 ALML。
- 识别位于尾部附近的另一个腹体触摸神经元 PVM 神经元,以指示前触摸神经元是否在同一平面中。如果是这样,观察到的触摸神经元是ALML。
- 了解其他索马体的位置,靠近感兴趣的区域(位于索马的两侧的荧光神经元),并在所有 Z 平面中,即使不可能将最深的神经元设置为清晰的焦点。
注意:识别所有接触神经元索马斯很重要,因为对焦外索玛可能被误认为是外在的。
- 确定动物是安装在左侧还是右侧(图3A)。
7. 识别和评分外在人
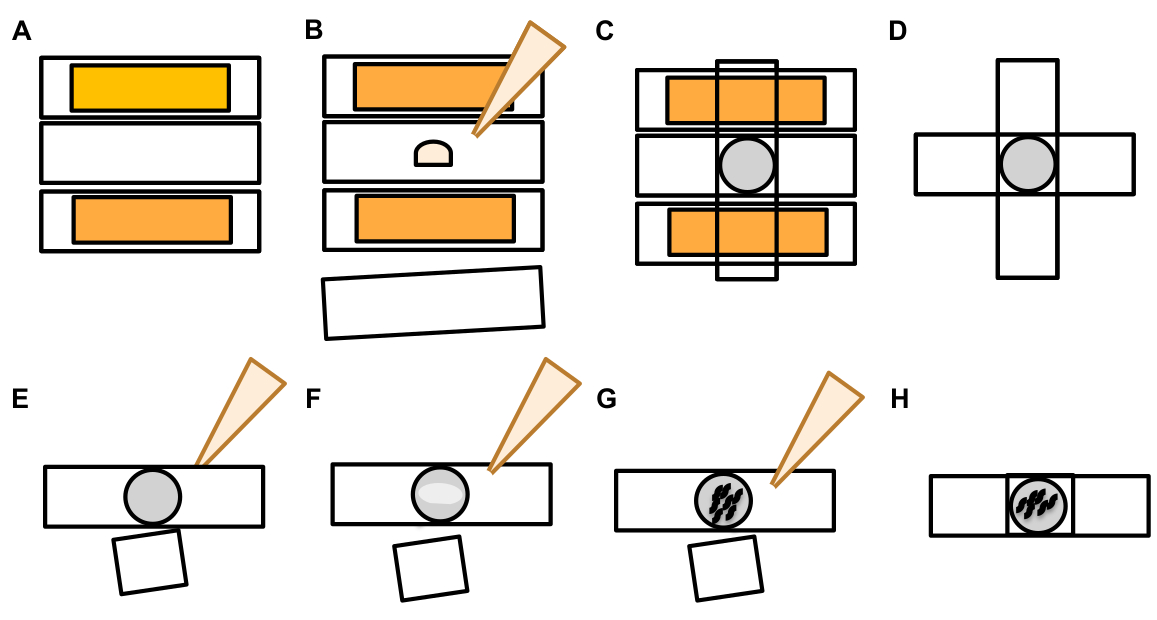
- 一旦发现触摸神经元,检查它大突起(外显域)大到足以被视为芽外泌物,(达到至少1/5的原生索玛的大小)(图1C)。
注:平均外外pher测量直径约2-8μm,而平均母性(ZB4065 bzIs166 [Pmec-4:::mCherry])动物测量6-10μm在第2天成人(图7B)。 - 如果没有观察到芽或外显域,检查神经元母细胞是否从索玛发出的附着细丝。附着的外层往往位于更接近原发索马和类似的Z平面。
注: 外向器并不总是与索玛保持连接。连接的灯丝的检测是物体是外在的明确标志。 - 要识别未连接的外外pher,请查找外外pher的内容。外层可以浓缩排出的荧光蛋白,因此通常比索马更亮。
注:外层的内容是异构的和可变的。细胞细胞器,如利索索姆和线粒体,也可以在外显体内挤出(图4C-E)。 - 在不同焦平面上查找与发现原始索玛的平面不同的未连接外在器。虽然外外层已经看到从 ALM soma 向任何方向突出,但通常的外驱从母细胞中伸出,从神经元过程向后方向突出。
- 检查未定位并标识为神经元的球形物体。外层可以不规则的形状,但通常是球形结构。外外层会随着时间的流逝而退化,因此较老的外外层往往具有更不规则的形状。
注:成熟或较老的外外pher通过外显子的更亮荧光强度及其球形,与分散的"星夜"阶段区分开来。 - 调查"星夜"表型作为早期外生形成的证据。外外pher进入一个"星夜"阶段,外显器分解成较小的囊泡,周围的皮下试图降解外显子的内容(图1G,2B,3B和7A)。
注:星空夜台的特点是零碎和分散(有时是网络化)荧光实体,与触摸神经元和外外层结构相比,这些实体失去了结构完整性,并表现出暗淡的荧光。 - 查找"多个外层事件"的实例。外层通常产生为奇异现象(1个外显子来自1个声马),但在某些情况下,可以从单个声子释放多个外向(图7D)。
注:成熟的外层可以降解成多个囊泡,因为它们在皮下降解。区分每个外热球是否由独立的外向发生事件生成,或者一个原始的外外pher分裂以产生额外的囊泡只能通过延时观测来确定。 - 请记住,并非所有的形态异常都成熟成外在。
- 不要将分散的索玛作为外在器评分。有时可以观察到延长或尖的索玛(特别是随着年龄的增长或压力),但没有明确收缩站点的扩展不会作为外在获得评分。
- 拒绝在外在外层事件定量中未达到 somath大小的小解析芽。
- 不要把神经素的外白算作外白。成熟的中性可以随着年龄的增长而急剧延长(通常与神经元过程相反的方向),荧光蛋白可以迁移到这种结构19的端端。
注:这些神经素生长不是外显子,因为它们在数天和数周内有明显的发育模式,不形成芽,不分离(图7E)。
- 识别非外 流子 的荧光实体。
注:获得背景荧光的想法,以确保正确识别挤出荧光实体与自动荧光,这一点很重要。-
转基因荧光表达与自荧光。 不要将自荧光误认为是转基因表达。真正的外外音信号不会在肠道或肠道中(DIC确认可用于识别这些组织),外外音信号将明显比背景自荧光更亮。
注:自荧光是由肠道颗粒肠道荧光色素沉着引起的,并随着年龄的增长而积累。它是异质的,尤其是不同波长的。 - 来自胚胎的信号。 不要误认为胚胎信号是外生。通过从荧光切换到亮场照明,并检查信号与子宫内卵子的关联,确认对胚胎信号的怀疑。
-
出飞机或附近的索马尸体。 通过识别附近所有索马尸体,甚至观察开始时的对焦外索玛,避免误用飞机外索马作为外向体。
注:如果对来自ALMR的外层进行评分,请识别和说明AVM和ALMR索马斯的位置。有关索马体识别的更多详细信息,请如图 3A所述。
-
转基因荧光表达与自荧光。 不要将自荧光误认为是转基因表达。真正的外外音信号不会在肠道或肠道中(DIC确认可用于识别这些组织),外外音信号将明显比背景自荧光更亮。
8. 评分和统计
- 得分外在作为二进制(是的,有一个外在器/否,有没有外在)。
- 将外层检测视为给定神经元的"外层事件"。外外pher事件可以构成对一个外在外在器附近的单个外在的观察。
注:要量化单个外向发生事件的数量,请使用延时观察。 - 计算每个特定标识的细胞的外向事件数,因为不同的细胞不会以相同的速率产生外向(参见图 3C)。ALMR神经元产生本文所述菌株中最基线的外显子,因此,这通常是从触摸受体神经元中选择用于外显细胞定量的细胞。
- 据统计,一般来说,至少进行3次生物试验, 每项试验至少有30 只动物得分,并须进行相应数量的观察,以分析干扰。
- 与对照相比,涉及一个或两个突变体/治疗的多个试验,科克伦-曼特尔-汉塞尔试验是适当的,以确定 p 值。
- 对于涉及两个多突变体的治疗与控制相比的试验,也适合使用二进制逻辑回归分析来评估任何数量的分类预测变量的显著性。
结果
多个荧光测量器可用于测量外显器。触摸神经元外显体很容易在体内通过荧光in vivo标记蛋白质,可能选择挤出,通过标签的细胞器,可以挤出,或通过标记细胞膜。表1列出了用于监测外显子的触摸神经元表示荧光报告器,图4中包括了具有代表性的例子。已知在外层中挤出的货物包括将人类亨廷顿蛋白的N终端域融合到扩展的多聚葡萄糖胺(Q128),用糖体相关膜蛋白(LMP-1)标记的体糖(LMP-1)和用基质局部GP标记的线粒体(图4D)。Figure 4BFigure 4C细胞质GP并没有被强烈排出,并优先保留在索马5,虽然GP可以弱地可视化外显体(图4A)。当GP被融合到被排出的蛋白质时,这个标签可以用来可视化外显子。一个重要的一点是,通过标记不同的蛋白质,可以解决关于驱逐特定货物和细胞器以及组成外层蛋白质和膜的一系列问题。
伪立体显微镜设置是观察动物在阿加盘上的外热动物的有效工具。此设置是复合和立体技术的混合体,包括每个放大倍率上的高数值孔径光学、伪立体技术(立体底座上的离散目标)以及用于在放大倍率中到已安装目标的变焦操作开关。这样的显微镜应配备10倍的目镜和目标,足以观察神经元形态和外层生产,实现高通量评分(2倍目标用于扫描/拾取,10倍目标用于识别和评分)。
虽然标准立体显微镜的放大能力通常具有足够高的分辨率,能够看到表达荧光蛋白的触摸神经元网络,但标准解剖显微镜不足以观察外显子的亚细胞细节,如索马与外显子的管状连接。这种观测需要共和显微镜(有关设备详情 ,请参见 材料表)。
外层定量研究需要严格的控制来消除实验应力。 对于可重复的外层生产,需要对一致的生长条件进行周密的维护。更具体地说,外层生产具有压力响应能力,因此,一致的喂养、恒定的温度和跨代的无污染增长对于可重复性至关重要。在具有高神经元表达的基底生长条件下,外显子的产生相对较低(5-25% 的ALMAR产生外显子),但一些压力,包括渗透和氧化应激,可以增加外外音率。虽然mCherry表达可以被认为是压力,但外外层压力敏感性的推论是,如果控制得当,实验压力引入可以更容易地诱导和观察外向发生。
时机和预期的外层生产水平。 在幼虫发育过程中,几乎不存在外层。年轻成人生活中外外外层产生高峰期似乎高度局限于成人日1-4,最常见的是在成人第2天或第3天。由于峰值可以提前或回移一点,因此对外外pher生产配置文件的最完整评估是每天在成人日 1-4 中进行多次试验。一般来说,ALMR产生一个主要的外层,囊泡至少持续24小时。外外pher可以相当快地产生(以最快的速度在几分钟内产生)。 最常见的情况是,在成年早期,每个神经元只产生一个主要外驱,但可以产生多个外外层。
一般来说,ALMRs在基础条件下表示mCherry的外向生产范围为5-25%的ALMAR在成人日2-3的最佳时间范围内检查(图3D)。蛋白位虫危机5,以及暴露于其他应力可以调节外层水平。压力或遗传扰动可以增加外外泌物的产生,检测率高达90%的ALMR神经元产生外外层挤出。
基于喂养的RNAi,用于测试特定基因在外生代的作用。 线虫 C.elegans 通常受到RNAi击倒通过喂养 动物转化的 大肠杆菌菌株HT115,表示双搁浅RNA(dsRNA)针对感兴趣的基因20。HT115细菌可用于在喂养RNAi 5时对外外层进行评分。虽然大多数组织的转录本可以由RNAi使用这种技术作为靶向,但神经元更难耐。对RNAi的敏感性可以使用在神经元特异性启动子下表达转基因dsRNA运输机SID-1的动物进行校准。这样,神经元组织就可以对RNAi21进行敏感。
通过表达缺乏该成分的突变体内内源性RNAi代谢成分,可以实现对感兴趣的基因的组织特异性敲击。例如:Argonaute 蛋白 RDE-1 可以专门在rde-1突变动物的神经元中表达,以便仅在动物接触针对该only基因的 RNAi 干预时,才能在神经元中敲掉感兴趣的基因。
使用标准线虫RNAi协议20,22,,22在L4阶段的父母暴露在RNAi,并允许他们的后代发展消耗转化的HT115细菌,直到成年产生强大的基因击倒,但注意RNAi诱发的潜在发育迟缓,因为实验动物可能生长不同于空病媒控制。始终包括空矢量控件以进行负控制比较非常重要。HT115 细菌可用于在喂养 RNAi 时对外层进行评分。然而,请注意,一些基因是有效的改变外生率,即使在较短的期间RNAi暴露5。如果针对某些基因导致发育失败,避免让动物终生被击倒,动物只需在L4阶段被采摘到RNAi板上,从L4接触成人D2或D3。
| 应变名称 | 基因 | 描述 | 外向百分比 | 参考 |
| SK4005 | zdis5[Pmec-4Gfp] | 在接触神经元中GP的细胞细胞表达。 | 1-8% ALM | 图 4A, 梅伦蒂耶维奇 2017 |
| ZB4065 | bzIs166[Pmec-4::mcherry] | 触摸神经元中 mCherry (bzIs166) 的过度表达,同时产生细胞细胞细胞细胞信号和 mCherry 聚合。bzIs166 是外向诱导剂。mCherry 集料是外向发生的预测变量,在外在中优先挤出。 | 3-20% ALM(正常情况)。20-80% ALM(禁食条件)。 | 图 4B, 梅伦蒂耶维奇 2017 |
| ZB4067 | bzIs167[Pmec-4mitogfp Pmec-4mcherry4];iis1[Pmec-7YFP Pmec-3htt57Q128::cfp lin-15]; | YFP 细胞体标记 mec-7 触摸神经元。共同表达 Q128::CFP 聚合和诱导外显器。CFP 优先沉默。 | ±25% | 图 4C, 梅莱蒂耶维奇 2017 |
| ZB4509 | bzIs166[Pmec-4mCherry];bzIs168[Pmec-7LMP-1::GFP] | bzIs168 LMP-1:GFP 标注血浆膜和乳糖膜。bzIs168 可用于识别神经元膜、外显层(因为它们是膜结合的)和细胞瘤膜结构。 | 3-20% ALM | 图 4D, 梅伦蒂耶维奇 2017 |
| ZB4528 | bzIs166[Pmec-4mCherry];zhsEx17 [Pmec-4米托尔斯::ROGFP] | Allele zhsEx17 是线粒体局部记者,根据局部氧化环境将其峰值激发波长从 405nm(氧化)变化为 476nm(减少)。它在触摸神经元中表达,并可用于识别触摸神经元和线粒体。 | 3-20% ALM 蛋白外向。% ALM 米托外向量正在进行中。 | 图 4E, 梅伦蒂耶维奇 2017, 大炮 2008, Ghose 2013 |
表1.用于可视化触摸神经元、触摸神经元外显层和外显子内容的菌株。

图1:外向发生阶段。 制作和弹出外外pher的过程称为"外在发生"。外层形成的动态过程可能需要几分钟到几个小时。描述的例子是索马和外外层形态在动态外外发生过程中在高外外层产生菌株,ZB4065 bzIs166[Pmec-4mCherry] 的特定步骤。所有图像都是第2天成人 ALM 神经元拍摄的 100 倍目标。(A) 正常索马.成人机械感官触摸神经元 Alm 转基因表达 Pmec - 4mcherry.所描绘的索玛形态是该菌株中年轻的成人神经元的典型特征,在细胞质中具有 mCherry 浓度。(B) 早期芽期.外外发生的第一个可观察到的步骤是将选定的细胞质材料极化到索马膜的边缘。此步骤通常伴随着索玛的膨胀或膨胀。在触摸神经元的情况下,前外向域 (PED) 延伸到周围的皮下(此处不可见)。注意将 mCherry 材料更集中到早期芽域中。(C) 晚芽阶段.随着细胞进一极化和前外向域的扩展,索马和外外层(箭头)之间的收缩变得明显。此事件表示向晚芽阶段的过渡。虽然在晚芽阶段,细胞表现出一个明确的收缩位点和单独的索马和外显子域,它还没有完全从索马捏掉;萌芽的外向可能由厚茎(箭头)连接。当有关外外pher域的直径比建筑工地/茎的直径大约1/3时,萌芽域被认为是早期外在的。(D) 早期开外层阶段。早期的外外层可以通过离境的茎连接——当外显子远离索马时,这种连接的直径会变薄。细胞质材料可以通过这种管子从索玛转移到外层,尽管大多数材料是在萌芽过程中加载的。外外pher可以脱离在(E)中描绘的索马,分离的外外层被认为是成熟的外外pher(F)。成熟的外热能穿过周围的皮下组织,远离离境的索玛。(G) 将 mCherry 标记的外层分解为皮下小囊泡,导致 mCherry 材料出现分散的双面体,很可能是在进入皮下心内体网络时。分散的庞蒂信号称为"星夜"阶段。一些外皮物的降解可能由皮下糖体完成,但有些物质没有完全降解,并且经常被下皮重新挤出到伪球体中。在图2中更详细地描述了后外生性mCherry 传输。 请单击此处查看此图的较大版本。
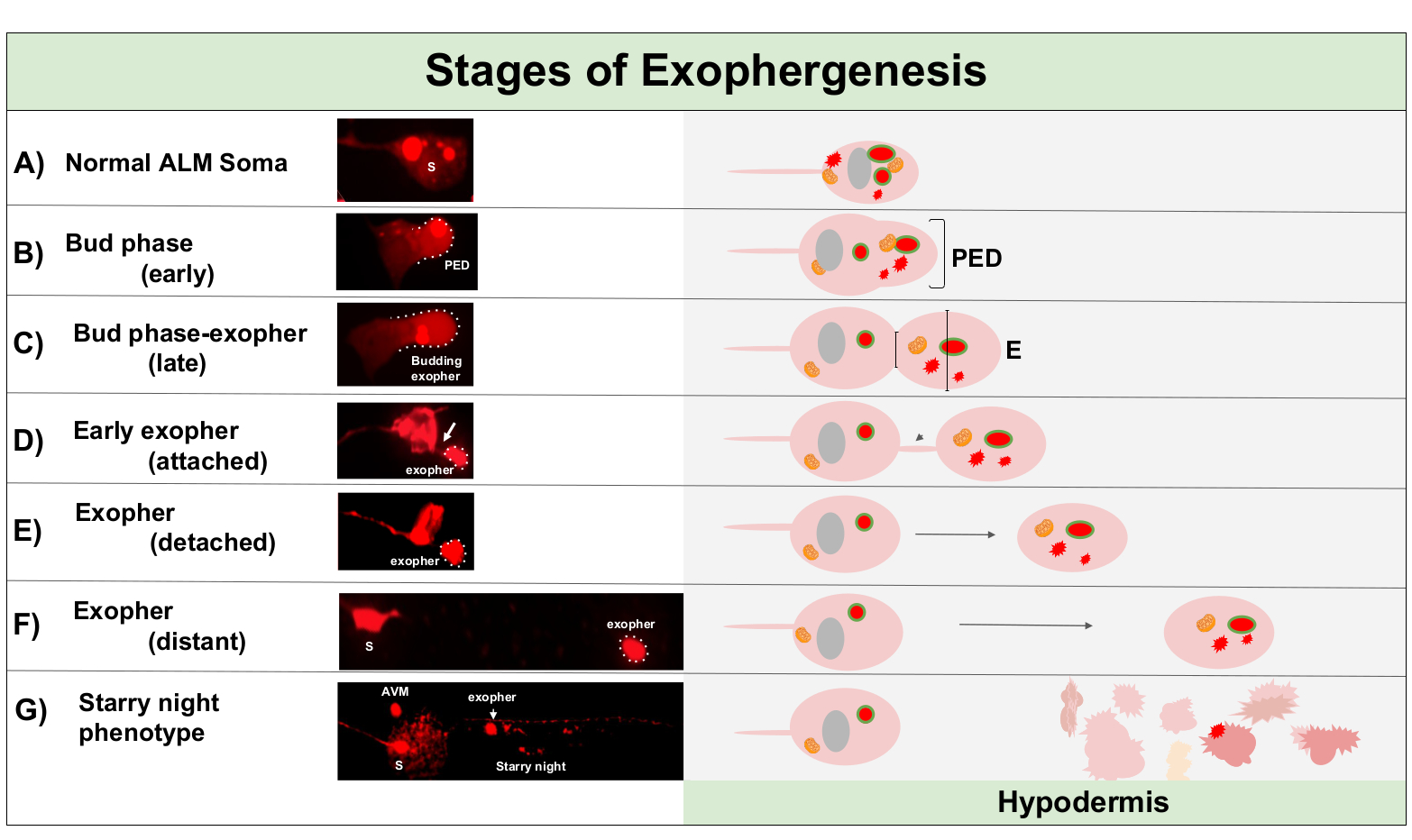{kind=link}

图2:从外层接触神经元中挤出的mCherry与周围的皮下细胞体网络接合,但随后可以挤出到伪细胞中,在那里,细胞可以储存/降解mCherry。(A) 卡通摘要,记录在外在外层中挤出的,在神经元驱逐后如何通过身体。在外生过程中,选择细胞内容,如mCherry成为局部和发芽从发送的神经元的母细胞在一个独立的囊泡包围的神经元和皮下血浆膜。由于触摸神经元嵌入在皮下组织中,当外显域向外发芽时,它进一步进入皮下。外显体可以传递皮下,数小时到数天后,外显物内容可以在皮下体内体网络内分裂。mCherry 可以在整个下皮出现分散的庞塔, 一个叫做 "星夜" 的舞台。几天后,一些mCherry可以从皮下进入周围的伪细胞,在那里,被称为细胞核细胞的清除细胞可以访问,并拿起,mCherry可以存储。(B) 星夜的毛囊外观的例子.图像的 Alm soma 标记与 mcherry 与大型外层碎片和星夜囊泡。应变是ZB4065bzIs166[Pmec-4mcherry]。(C) 遥远细胞中mCherry浓度的例子。成年动物日10的侧视图株ZB4065bzIs166[Pmec-4mcherry] 显示mCherry集中在骨髓细胞(箭头)。一些星空夜囊泡也很明显。一般来说,在成年后的第5天之后,血红细胞浓度变得很明显。(B 底部)卡通再现 (B),触摸神经元和过程以红色勾画,如最亮的外层片段;分散的不同 Z 深度的小囊泡以浅粉色显示。(C 底部)卡通版图像 (C),显示神经元过程在红色,星夜在粉红色和细胞细胞在绿色。请单击此处查看此图的较大版本。
{kind=link}

图3:机械感官触摸神经元产生不同级别的外层,具有精确的时间轮廓。(A)(顶部) 卡通描绘机械感官触摸神经元的空间关系的关键解剖 地标的C. elegans, 包括抽咽和神经元密集的神经环在动物的头部,在中体外阴,和锥形的尾巴。(底部)荧光标记的触摸神经元表示 GFP 从顶部和左侧观看(根据 WormAtlas 改编的图像)。红色框描述 ALM 外层通常位于的区域。(B) 在中体区域的高放大率视图,其中 ALM 衍生的外导球以表达 [Pmec-4mCherry] 的应变生成。AVM 和 ALMR 神经元被描绘, 显示是一个 ALMR 外显器以及 mcherry 星夜。ALMR神经元最容易产生外显子。(C) ALMR机械感官触摸神经元更容易产生外热素相比,其他触摸神经元在基础条件下的草本体。机械感官触摸神经元外层生产在成人第2天,为个别触摸受体神经元评分指示。应变: ZB4065 bzIs166[Pmec-4mCherry], N>150, 误差杆是 SEM. (D) ALMR触摸神经元在成年后的第 2 天和 3 天与青少年 L4 阶段或高龄动物相比产生更多的外向。应变: ZB4065 bzis166[Pmec-4mcherry], N>150, 误差条是 Sem. 请点击这里查看这个数字的较大版本。
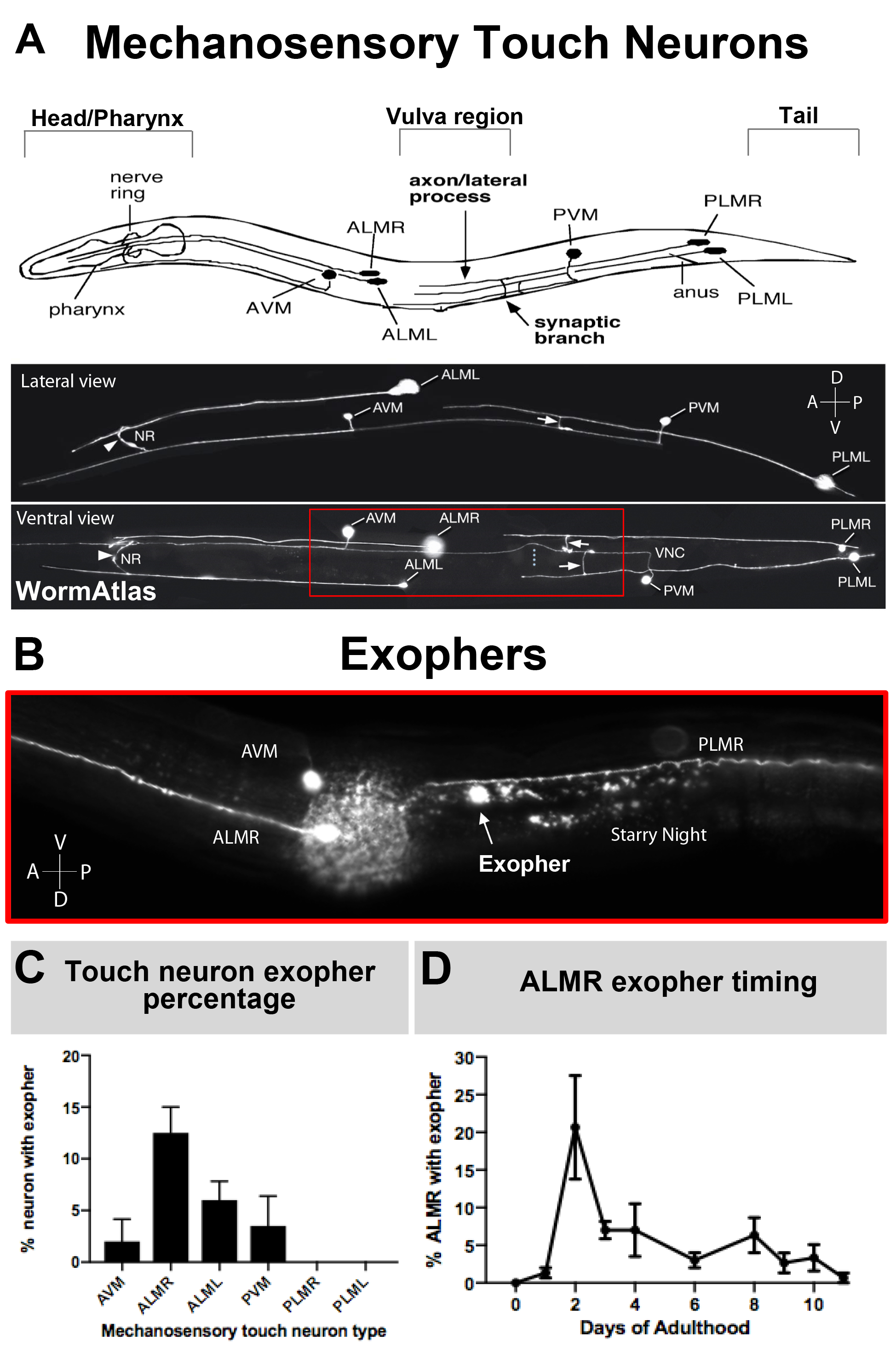{kind=link}

图4:一些荧光记者标记外在内容的示例。观察外层的直接方法就是创造转基因动物,从神经元促进剂中表达荧光。荧光团允许外显源和转基因表达的可视化诱导聚集和/或蛋白质,增加外生。在原生条件下,还可以观察到由安非他明神经元产生的外向,使用染料填充进行可视化。图中显示了可用于观察外外层(E)外显器(S)的常用菌株示例。(A) 从 ALM 的 ALM 中,一个成年的应变 SK4005 zdIs5[Pmec-4GFP],100 倍目标用于摄影,比例杆 3μm。在此菌株中,测量包括可溶性 GFP 的外显层,但外显子的产生很少发生。在其他研究中,将GP融合到可优先挤出的外外层中的蛋白质中证实,在成熟的外外层中可以检测到GP融合。(B) ALM soma 和外热菌株 ZB4065 bzIs166[Pmec-4mCherry] 的成人的 ALM soma 和外外向,表示 mCherry 并诱导触摸神经元外向产生。100倍目标用于摄影,比例杆5μm。(C) ALM soma 和外热菌株 ZB4067 bzIs167[Pmec-4mitogfp Pmec-4mcherry4];iis1[Pmec-7YFP Pmec-3htt57Q128::cfp lin-15];用于 htt57Q128 图像的选择性蓝色通道::CFP。外外pher包含htt57Q128::CFP聚合(箭头),这些聚合体在外在器中比在 soma 中更集中。40倍目标用于摄影,比例杆5μm。(D-E)外层可以包含细胞器和细胞器特异性标记与荧光蛋白,使监测细胞器挤出。(D) Lysosomal 膜标签 LMP-1::GFP 勾勒出母体和外显膜,并弱弱地标记血浆膜(血浆膜定位是迈向乳糖体靶向的贩运步骤),并强烈标注了乳糖体细胞器。显示的是一个成人 Alm soma 共同表达 Pmec-4mcherry 和 Pmec-7LMP-1::GFP 本地化到膜和利索索姆。soma 有一个附加的外向器,其他较小的挤出物可能是外在碎片(箭头)。GFP正结构包含在索马中,并存在于大外外层中,应变:ZB4509 bzIs166[Pmec-4mCherry];bzIs168[Pmec-7LMP-1:GFP]。100倍目标用于摄影,比例杆5μm。E) 线粒体GP标记可用于识别索马和外显子中的线粒体。显示的是一个成年的ALM索马表示P mec-4mcherry和米托:ROGFP,它本地化到线粒体矩阵。mito:ROGFP单独表示,没有mCherry,也可以很容易地用于识别神经元和评分的外向,包括线粒体。应变: ZB4528 bzIs166[Pmec-4mCherry];zhsEx17 [Pmec-4米托尔斯::ROGFP].用于摄影的100倍目标;比例杆 5μm。请单击此处查看此图的较大版本。
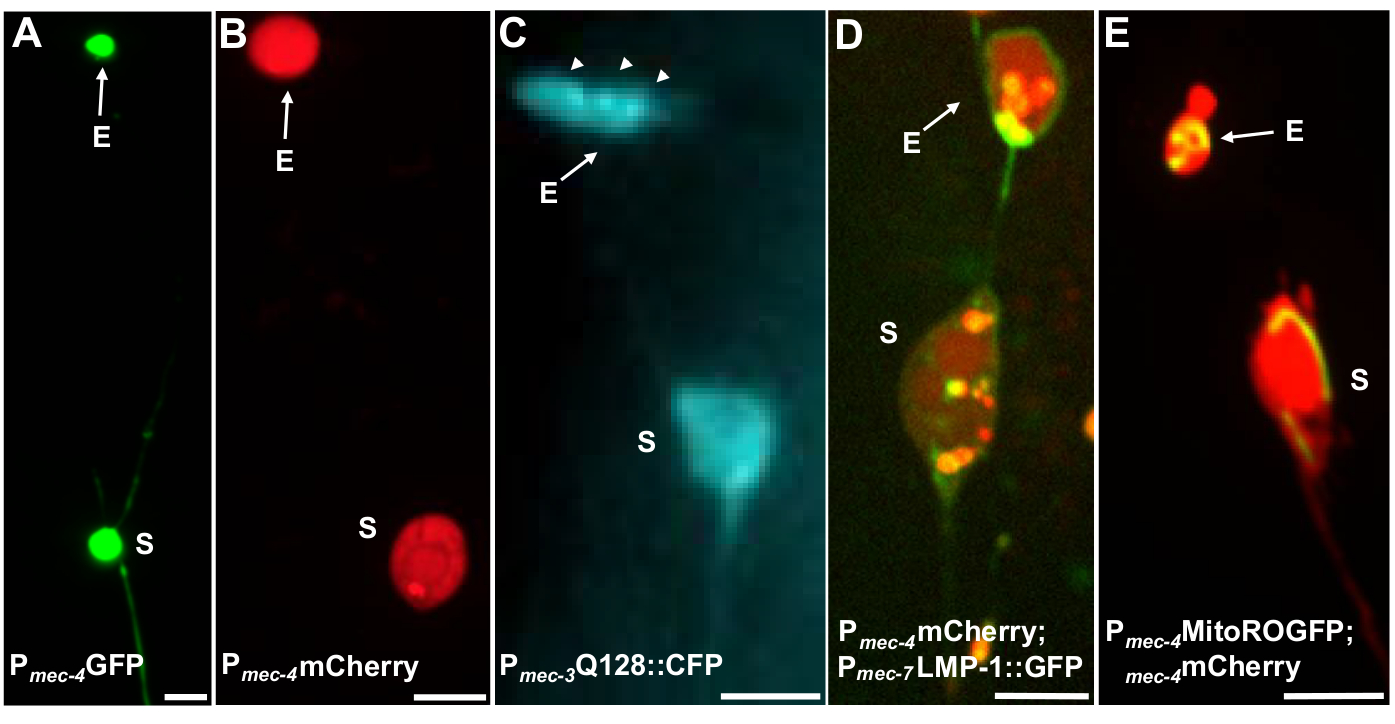{kind=link}

图5:C.elegans和L4鉴定的发育周期。(A) 在20°C下,一个卵子在母亲下完后孵化大约需要9个小时。(B) 新孵化的动物处于幼虫期1(L1),12小时后进入L2幼虫。(C) 动物在L2和(D)L3幼虫阶段各保留约8小时。(E) 青少年动物被认为是第四幼虫阶段(L4),其特点是明显发育的外阴,在中体附近显示为白色新月。这种新月的存在使得L4分阶段动物的识别和采摘变得容易,从而建立同步的培养,从而促进外在动物的评分。动物在L4阶段保持约10个小时,最后一次被活化成人,F)通过发育卵子、可见精子和开始产卵来识别。请单击此处查看此图的较大版本。
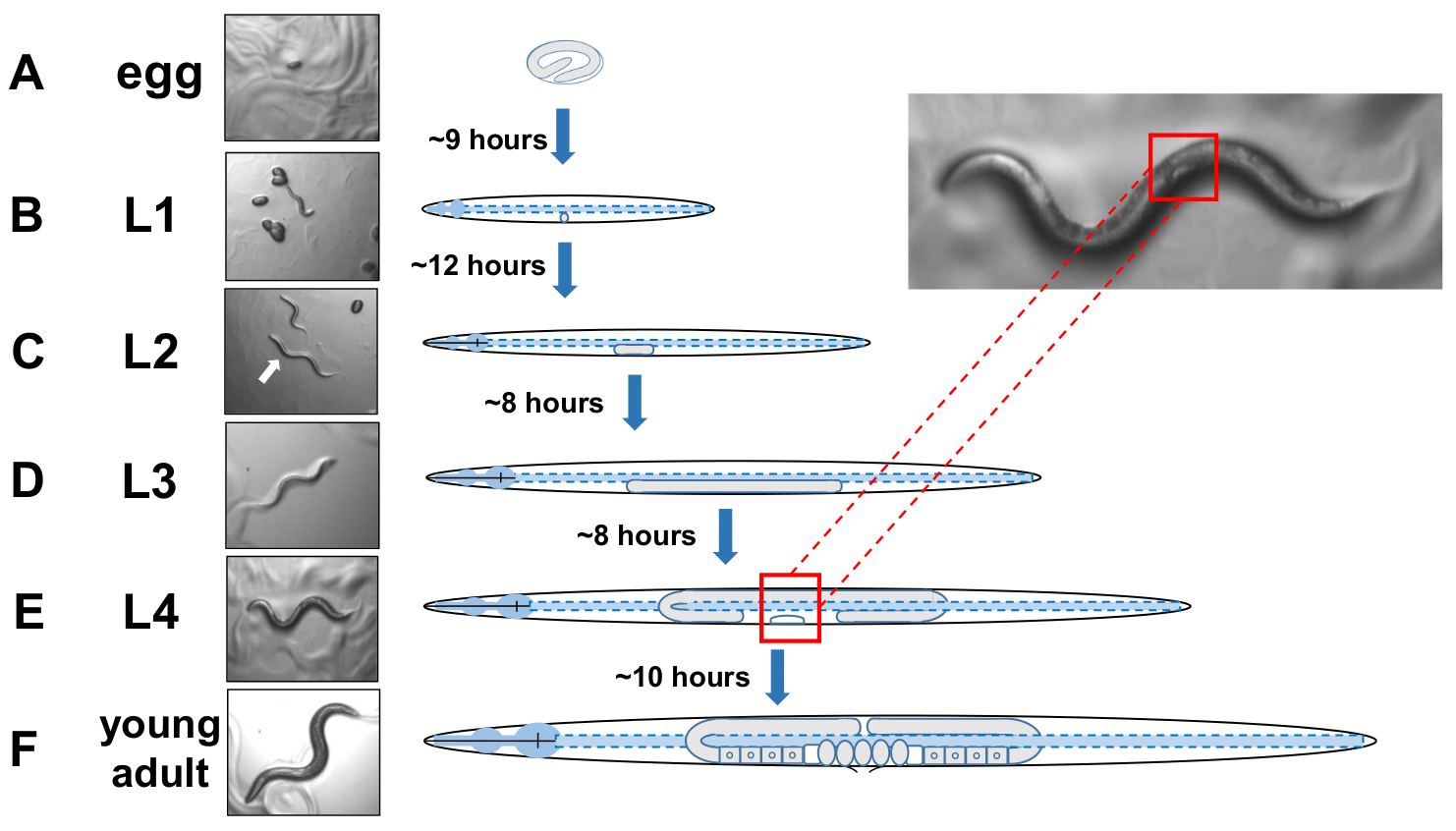{kind=link}

图6:准备显微镜幻灯片垫。 (A) 准备两张幻灯片,上面用一条实验室胶带纵向放置。如图所示,将非胶带显微镜幻灯片放在两者之间。 B) 在滑梯上放一滴熔融的阿加糖。(C) 将干净的幻灯片轻轻放在落点顶部,将气垫压入充气圆垫中。(D) 移除已录制幻灯片,用于完成创建均匀垫所需的阿加的均匀扁平化。(E) 干完加糖垫后,拆下顶部滑梯。(F) 在阿加垫上移液麻麻溶液(左西莫或四米索)。(G) 将适当分阶段的动物放入瘫痪中。(H) 用盖玻片轻轻盖住动物,确保动物活着。 请单击此处查看此图的较大版本。
{kind=link}

图7:外层特征和外外层识别标准。(A) 识别外在的一般标准。(B) 发送母索与拉伸外显器之间的直径比较,以μm为单位。成人 ALM somas, N=35, 应变: ZB4065 bzIs166[Pmec-4mCherry] - 6.53 μm 平均大小索马和 3.83 μm 平均外外膜大小。(C) 定义区分外在域和萌芽外在的外在域的标准。(D) 最常见的情况是,单个神经元产生一个大的外显子,当皮下试图降解其内容时,这些外显器会分裂或分裂。不过,在一个触摸神经元旁边可以观察到多个外驱,这些神经元可能来自一个神经元的多个外外层事件,或者外外pher也可以发芽或分裂自己。只能使用时间推移显微镜确定多个外层实体的来源。顶部描绘了一个ALMR触摸神经元索玛与一个单一的遥远的外驱。底部描绘了一个ALMR触摸神经元索马与多个外层一样挤出。(E) 成人 ALM 接触神经元的常见形态特征,可能被误认为是外在事件。左上角 - 一个分散的 ALM soma,没有明确的外传域或收缩站点。顶部中间 - 神经元可以有小的细胞外突起,可能类似于外在,但不符合大小要求标准,被认为是外在。右上– 随着年龄的增长,触摸神经元会沿着其小神经石发展出树。通常,在神经菌生长的尖端可以收集 mcherry 材料。如果收集的 mCherry 不符合外在到索马大小要求,这不能作为外在码进行评分。底部描述成年 ALM 神经元,这些神经元具有外在域或外在的定义了标准。博托姆左 - ALM soma,有一个突出的外向域,选择性地包括mCherry细胞索和mCherry标记聚合。外向域收缩站点由箭头标记,并满足大小条件(至少为 soma 大小的1/5)。外外层域的最大直径几乎比收缩场的直径大 1/3,符合外在器事件的标准。底部中间- ALM soma,有一个突出的萌芽外层,满足大小标准。有一个明确的收缩站点。右下角 -ALM soma 具有附加的 mCherry 填充外层,满足外层大小要求。外外层由细连接灯丝连接。所有图像都来自应变 ZB4065 bzIs166[Pmec-4mcherry]。请单击此处查看此图的较大版本。
{kind=link}
讨论
以大型外外 层形式 对体内分子机制的聚合和细胞器消除的表征还处于起步阶段。关于指定驱逐货物、在牢房内对这些货物进行两极分化的收集、产生外驱的决定的管制、调解挤出的机械以及外驱生物与邻近牢房中退化机械的相互作用的问题,都仍要加以解决。此外, 管状连接的 体内可视化,可以通过生物材料,包括钙,聚合体,线粒体是有趣和未研究生物学本身的权利。为什么某些细胞比另一些细胞更容易产生外层细胞的问题也未解决,但可以通过本协议中概述的方法开始进行基因解剖。
本协议中详细描述的方法是实现外外层生产可重复评分的方法,注意区分外外层与附近的细胞索马,分析时间捕捉外外pher生产的峰值,并严格控制生长条件,以消除可调节外外层水平的意外应力。大的早期外外层的区别,或"星夜"分散在周围的皮下可以量化作为外外层生产的证据。话虽如此,在基底条件下表达mCherry的神经元通常与产生外显素的特定类型的5-25%的神经元相关。控制引入应力条件可应用于增加外外泌物的产生,以检测高达90%的神经元产生挤出,尤其适用于修饰剂的遗传或药理学屏幕。
在人类神经退行性疾病中,大型聚集体可以从病变神经元转移到邻近的细胞,促进病理传播。外外pher机制可能通过用于在植物中聚合挤出的保守机制发生。定义 体内分子 ,要么提高这个过程的效率(被认为是更有效的蛋白病控制),要么阻止它可能被利用来影响对抗多种神经退行性疾病的新策略的设计。因此,此处描述的协议可用于经典基因突变体筛选、全基因组RNAi屏幕,这些屏幕系统地敲除基因来识别增强剂和抑制剂,或者用于确定这一过程的候选药理修饰剂的药物干预研究。这种方法很简单,虽然有些费力。外层是非常大的,他们可以通过高放大镜的解剖显微镜来观察。尽管如此 ,C.elegans 神经元相对较小,看他们的细胞器或膜需要更高功率的共物图像,是一个缓慢的过程。高吞吐量的选项可能涉及多井板格式的高含量成像方法。
应用标准化方法进行外外层评分应作为神经元组织和消除细胞碎片的过程的协同基因解剖基础。
披露声明
没有
致谢
我们确认以下 NIH 赠款:R01AG047101 和 R37AG56510。Driscoll 和 Grant 实验室的成员通过严格的实验和强大的沟通,为所述协议的开发和微调提供了广泛的贡献。
材料
| Name | Company | Catalog Number | Comments |
| 95B Scientific CMOS camera | Photometrics Prime | ||
| 1,000 μL low retention tips | Sarstedt | ||
| 10 mL serological pipette | Appleton Woods | CC214 | |
| 10 μL low retention tips | Sarstedt | 70.1130.105 | |
| 13% sodium hypochlorite | Acros Organics | AC219255000 | |
| 15 mL centrifuge tubes | Fisher Scientific | 05-539-12 | |
| 2 L erlenmeyer flasks | Scientific Laboratory Supplies | FLA4036 | |
| 25 mL serological pipette | Appleton Woods | CC216 | |
| 300 μL low retention tips | Sarstedt | 70.765.105 | |
| 50 mL serological pipette | Appleton Woods | CC117 | |
| 5-Fluoro-2'-deoxyuridine 98% | Alfa Aesar | L16497.ME | |
| 9 cm sterile Petri dishes | Fisher Scientific | 11309283 | |
| absolute ethanol | Vwr | 20821.33 | |
| Agar | Sigma Aldrich | A1296 | |
| C. elegans strain wild type | Supplied by CGC | N2 | C. elegans strain |
| calcium chloride dihydrate | Sigma Aldrich | C3881 | |
| cholesterol | Acros | 110190250 | |
| dibasic sodium phosphate | Sigma Aldrich | S3264 | |
| E. coli strain OP50 | Supplied by CGC | Op50 | E coli strain |
| FBS10 Standard microscope | Meyer Instruments | KSC 410-1-100-1 | FBS10 Standard with Plate Base, 100/100 Trinocular Head and Flip zoom |
| glass pipette 270 mm | Fisherbrand | FB50255 | |
| Heraeus Multifuge X3R | Thermofisher scientific | 75004515 | |
| Inoculating Spreaders | Fisher Scientific | 11821741 | |
| LB medium capsules | MP biomedicals | 3002-031 | |
| LDI – Laser Diode Illuminator | 89 North | ||
| levamisole | Sigma Aldrich | 16595-80-5 | |
| M4 multipette | Eppendorf | 4982000012 | |
| magnesium sulphate | Sigma Aldrich | M7506 | |
| monobasic potassium phosphate | Sigma Aldrich | P0662 | |
| Multitron Standard shaking incubator | Infors HT | INFO28573 | |
| Nalgene 1 L Centrifuge pots | Fisher Scientific | 3120-1000 | |
| P10 pipette | Eppendorf Research Plus | 3123000020 | |
| P1000 pipette | Eppendorf Research Plus | ||
| P200 pipette | Eppendorf Research Plus | 3123000055 | |
| pipeteboy 2 | VWR | 612-0927 | |
| Polystyrene microbeads | Sigma Aldrich | MFCD00131491 | |
| RC5C plus floor mounted centrifuge | Sorvall | 9900884 | |
| Reusable ringed cytology slides | ThermoFisher Scientific | 22037242 | |
| SK4005 zdIs5[Pmec-4GFP] | contract Driscoll lab | GFP expressed in touch neurons | |
| sodium chloride | Sigma Aldrich | 13422 | |
| Sodium hydroxide | Fisher Chemical | S/4880/53 | |
| Tactrol 2 Autoclave | Priorclave | ||
| Triton-X | Thermofisher scientific | 28313 | |
| Tween 20 | Sigma Aldrich | 9005-64-5 | |
| X-Light V2 Spinning Disk Confocal Unit | CrestOptics | ||
| ZB4065 bzIs166[Pmec-4mCherry] | contract Driscoll lab | mCherry expressed in touch neurons | |
| ZB4067 bzIs167[Pmec-4mitogfp Pmec-4mCherry4]; igIs1[Pmec-7YFP Pmec-3htt57Q128::cfp lin-15+] | contract Driscoll lab | Q128 expressed in touch neurons | |
| ZB4509 bzIs166[Pmec-4mCherry]; bzIs168[Pmec-7LMP-1::GFP] | contract Driscoll lab | mitoROGFP expressed in touch neurons | |
| ZB4528 bzIs166[Pmec-4mCherry]; zhsEx17 [Pmec-4mitoLS::ROGFP] | contract Driscoll lab | autophagy marker expressed in touch neurons | |
| ZEISS Axio Vert.A1 | Zeiss |
参考文献
- Davis, A. A., Leyns, C. E. G., Holtzman, D. M. Intercellular Spread of Protein Aggregates in Neurodegenerative Disease. Annual Review of Cell and Developmental Biology. 34, 545-568 (2018).
- Davis, C. H., et al. Transcellular degradation of axonal mitochondria. Proceedings of the National Academy of Sciences of the United States of America. 111 (26), 9633-9638 (2014).
- Torralba, D., Baixauli, F., Sanchez-Madrid, F. Mitochondria Know No Boundaries: Mechanisms and Functions of Intercellular Mitochondrial Transfer. Frontiers in Cell and Developmental Biology. 4, 107 (2016).
- Stahl, P. D., Raposo, G. Extracellular Vesicles: Exosomes and Microvesicles Integrators of Homeostasis. Physiology (Bethesda, Md.). 34 (3), 169-177 (2019).
- Melentijevic, I., et al. C-elegans neurons jettison protein aggregates and mitochondria under neurotoxic stress. Nature. 542 (7641), 367 (2017).
- Nussbaum-Krammer, C. I., Park, K. W., Li, L., Melki, R., Morimoto, R. I. Spreading of a prion domain from cell-to-cell by vesicular transport in Caenorhabditis elegans. PLoS Genetics. 9 (3), 1003351 (2013).
- Tyson, T., et al. Novel animal model defines genetic contributions for neuron-to-neuron transfer of alpha-synuclein. Scientific Reports. 7, (2017).
- Babcock, D. T., Ganetzky, B. Transcellular spreading of huntingtin aggregates in the Drosophila brain. Proceedings of the National Academy of Sciences of the United States of America. 112 (39), 5427-5433 (2015).
- Pearce, M. M. P., Spartz, E. J., Hong, W., Luo, L., Kopito, R. R. Prion-like transmission of neuronal huntingtin aggregates to phagocytic glia in the Drosophila brain. Nature Communications. 6, 6768 (2015).
- Fu, H., Li, J., Du, P., Jin, W., Cui, D. Metabolic wastes are extracellularly disposed by excretosomes, nanotubes and exophers in mouse HT22 cells through an autophagic vesicle clustering mechanism. bioRxiv. 10 (1), (2019).
- Ghose, P., Park, E. C., Tabakin, A., Salazar-Vasquez, N., Rongo, C. Anoxia-reoxygenation regulates mitochondrial dynamics through the hypoxia response pathway, SKN-1/Nrf, and stomatin-like protein STL-1/SLP-2. PLoS Genetics. 9 (12), 1004063 (2013).
- Cannon, M. B., Remington, S. J. Redox-sensitive green fluorescent protein: probes for dynamic intracellular redox responses. A review. Methods in Molecular Biology. 476, 51-65 (2008).
- Perkins, L. A., Hedgecock, E. M., Thomson, J. N., Culotti, J. G. Mutant sensory cilia in the nematode Caenorhabditis elegans. Developmental Biology. 117 (2), 456-487 (1986).
- Stiernagle, T. Maintenance of C. elegans. WormBook: The Online Review of C. Elegans Biology. , 1-11 (2006).
- Sutphin, G. L., Kaeberlein, M. Measuring Caenorhabditis elegans life span on solid media. Journal of Visualized Experiments. (27), (2009).
- Mitchell, D. H., Stiles, J. W., Santelli, J., Sanadi, D. R. Synchronous growth and aging of Caenorhabditis elegans in the presence of fluorodeoxyuridine. Journal of Gerontology. 34 (1), 28-36 (1979).
- Weicksel, S. E., et al. A novel small molecule that disrupts a key event during the oocyte-to-embryo transition in C. elegans. Development. 143 (19), 3540-3548 (2016).
- Dong, L., et al. Reversible and long-term immobilization in a hydrogel-microbead matrix for high-resolution imaging of Caenorhabditis elegans and other small organisms. PloS One. 13 (3), 0193989 (2018).
- Toth, M. L., et al. Neurite sprouting and synapse deterioration in the aging Caenorhabditis elegans nervous system. Journal of Neuroscience. 32 (26), 8778-8790 (2012).
- Conte, D., MacNeil, L. T., Walhout, A. J. M., Mello, C. C. RNA Interference in Caenorhabditis elegans. Current Protocols in Molecular Biology. 109, (2015).
- Calixto, A., Chelur, D., Topalidou, I., Chen, X., Chalfie, M. Enhanced neuronal RNAi in C. elegans using SID-1. Nature Methods. 7 (7), 554-559 (2010).
- Maher, K. N., Catanese, M., Chase, D. L. Large-scale gene knockdown in C. elegans using dsRNA feeding libraries to generate robust loss-of-function phenotypes. Journal of Visualized Experiments. (79), e50693 (2013).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。