
Method Article
C. 예르간에서 큰 Exopher 소포에서 생체 신경 골집 및 세포구 압출에서 득점에 대한 정량적 접근
요약
이 프로토콜은 막 에바인딩 exophers의 형태로 C. elegans 세포에 의해 생성된 큰 골체 및/또는 세포기관 압출(~4 μm)의 검출 및 수량에 대한 접근 법을 설명합니다. 우리는 이 파편 추방 기계장치의 해부를 용이하게 하는 데 필요한 긴장, 성장 조건, 채점 기준, 타이밍 및 현미경 고려 사항을 설명합니다.
초록
잘못 접힌 단백질과 미토콘드리아 기능 장애의 독성은 종에 걸쳐 노화와 관련된 기능성 신경 감소 및 신경 퇴행성 질환을 촉진하는 중추적 인 요인입니다. 이러한 신경 독성 문제는 오랫동안 세포 내재로 간주되었지만, 상당한 증거는 이제 한 신경에서 유래된 잘못된 인간 질병 단백질이 이웃 세포에 나타날 수 있음을 뒷받침하며, 이는 인간의 신경 퇴행성 질환에 대한 병리학 확산을 촉진하기 위해 제안된 현상입니다.
C. 집계 단백질을 표현하는 성인 뉴런은 골거된 단백질, 미토콘드리아 및 리소좀을 포함할 수 있는 대형(~4 μm) 막으로 둘러싸인 소포를 돌출할 수 있습니다. 이 큰 소포는 "exophers"에게 불리고 엑소좀 (약 100 배 더 작고 다른 생물 발생이 있는)와 구별됩니다. exophers에 세포 파편을 던지는 것은 근본적이지만 이전에는 인식할 수 없는 신경 프로테오스타증 및 미토콘드리아 품질 관리의 분지인 보존된 메커니즘에 의해 발생할 수 있으며, 이는 골재가 인간 신경 퇴행성 질환에 퍼지는 과정과 관련이 있습니다.
exophers터치 뉴런 내에서 높은 복사 형질형 mCherry를 표현하는 동물에서 주로 연구되었지만, 이러한 프로토콜은 형광 태그 된 세포기관 또는 뉴런의 다양한 클래스에 관심있는 다른 단백질을 사용하여 exophergenis의 연구에서 동등하게 유용합니다.
여기에 설명된 C. elegans exophers의 물리적 특징, 탐지 전략, 식별 기준, 수량에 대한 최적의 타이밍 및 exopher 생산 수준을 조절할 수 있는 스트레스를 제어하는 동물 성장 프로토콜이 있습니다. 여기에 설명된 프로토콜의 세부 사항은 실험실 전체의 퇴출물의 정량적 분석을 위한 표준을 수립하는 역할을 해야 합니다. 이 문서는 exophers가 생성되고 이웃과 먼 세포에 의해 반응되는 분자 메커니즘을 정교하게 하고자하는 실험실의 자원역할을하고자합니다.
서문
골재 및 기능 장애 미토콘드리아의 신경독성 문제는 오랫동안 세포 내재된 것으로 간주되어 왔지만, 최근에는 한 뉴런에서 발생하는 잘못된 인간 질환 단백질이 이웃 세포로 확산되어 병리학1을촉진할 수 있다는 것이 분명해졌습니다. 마찬가지로, 포유류 미토콘드리아는 세포간 분해2 또는 도전된 이웃 세포3에서미토콘드리아 집단의 구출을 위해 원래 생산의 세포에서 보내질 수 있다. 다양한 크기의 소포는 일반적으로 이웃 세포 또는 유체 주변4로 세포 물질을4전송하는 것으로 관찰되었다. 일부 압출 소포는 평균 신경 소마 (평균 터치 뉴런 소마 ~ 6 μm)의 크기에 접근하고 큰 골재 및 세포기관을 수용 할 수 있습니다.
단백질 응집체및 세포기관을 운반할 수 있는 큰 소포 압출의 눈에 띄는 예는 C. elegans 터치 수용체 뉴런에서 발생하며, 이는 유해 응집경향이 있는, 분해 저항성 mCherry5를인코딩하는 높은 카피 번호 기자 구성을 시현하는 수용체 뉴런을 터치한다. exophers에게 불린 터치 뉴런에서 압출은 , ~4 μm 평균 직경, 선택적으로 mCherry 또는 그밖 응재를 포함하고, 일반적으로 접촉 수용체 뉴런을 포위하는 이웃 피질로 직접 전달됩니다. 피질계는 리소좀 기반 의 분해를 시도하지만, mCherry 골재와 같은 일부 비소화성 내용들은 피하구체에 의해 동물의 유체 채워진 의사콜롬으로 재압될 수 있으며, 이 과정에서 mCherry는 장기간 저장을 위해 coelomocytes라고 불리는5원격 청소세포에 의해 채택될 수 있다(그림1, 그림 2)
큰 압출 제거 소포는 터치 수용체 혈장 막에 둘러싸여 세포를 떠나 고집계 된 인간 질환 단백질을 포함 할 수 있습니다, 미토콘드리아, 및 리소좀. exopher 생산의 과정은 잠재적으로 독성 종의 정렬을 포함 하는 것으로 보인다 (예를 들어 집계 하기 쉬운 표현 mCherry는 수용성에서 분리, 신경 소마에 주로 남아 GFP 같은 불쾌 한 단백질). 이러한 방식으로, 위협적 개체의 지시추방은 뉴런5에의해 수행된다. 발성기 대책은 자가식 녹다운, MG132 매개 프로테아솜 억제, 또는 헌팅턴병 관련 확장 폴리글루타민 Q128 또는 알츠하이머병-병-성 단편 Aβ1-42와같은 인간 질환 단백질의 형질 발현에 의해 유도된 스트레스와 같은,5-42,엑스포퍼를 생성하는 뉴런의 수를 증가시킬 수 있다.
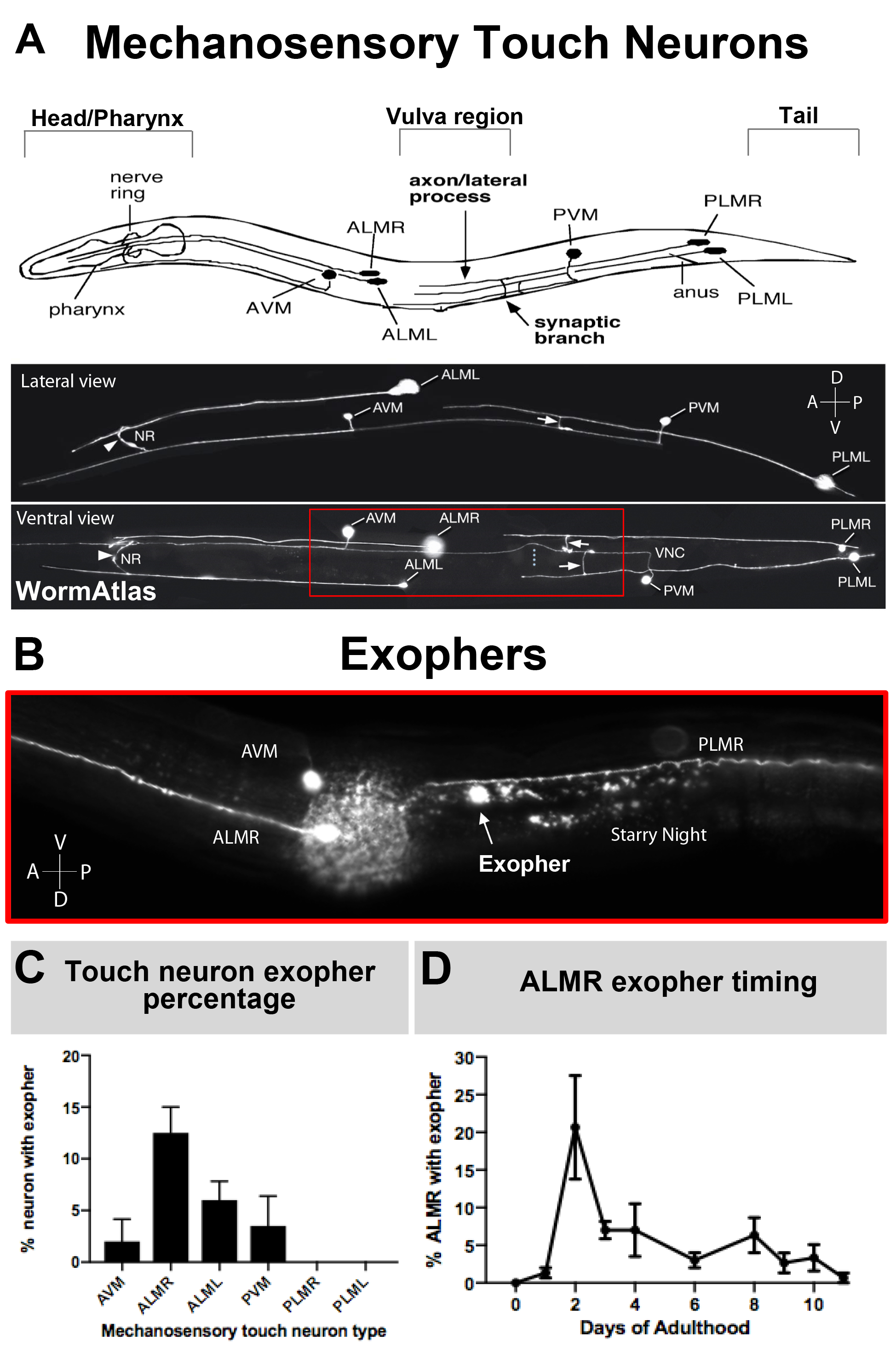
엑소퍼는 최근에 문서화되었기 때문에 생물학 장점 설명으로 알려져 있습니다. Exophers에서 발견 되었다, 그리고 가장 잘 공부, C. elegans 터치 수용 체 뉴런. 6개의 C. elegans 메카노감각 터치 뉴런이 있는데, 세포 체가 몸 주위에 분포되어있으며(그림 3A)라고불리며, 그들의 초구조가 독특한 15 개의 프로토필라멘트 마이크로투블러를 특징으로하기 때문에 마이크로 튜블러 세포라고 합니다. 터치 수용체 뉴런은 전방 AVM (전방 복부 마이크로 투툴 뉴런), ALMR 및 ALML (전방 측면 마이크로 투덜뉴런 오른쪽과 왼쪽), 더 중앙 PVM (후방 복부 마이크로 투덜뉴런), 후방 PLMR 및 PLML (후방 측면 미세 투불 뉴런 및 오른쪽 왼쪽)입니다. 흥미롭게도, 6개의 터치 수용체 뉴런은 동일한 공격적인 트랜스진(도3C)을표현했음에도 불구하고 다른 속도로 엑소퍼를 생성한다. 6개의 메카노감각 터치 수용체 뉴런의, ALMR 뉴런은 그밖 터치 뉴런 보다는 더 자주 exophergenesis를 겪습니다. 따라서 터치 뉴런에서 엑소퍼 번호의 양은 일반적으로 ALMR에 초점을 맞추어 확립된다.
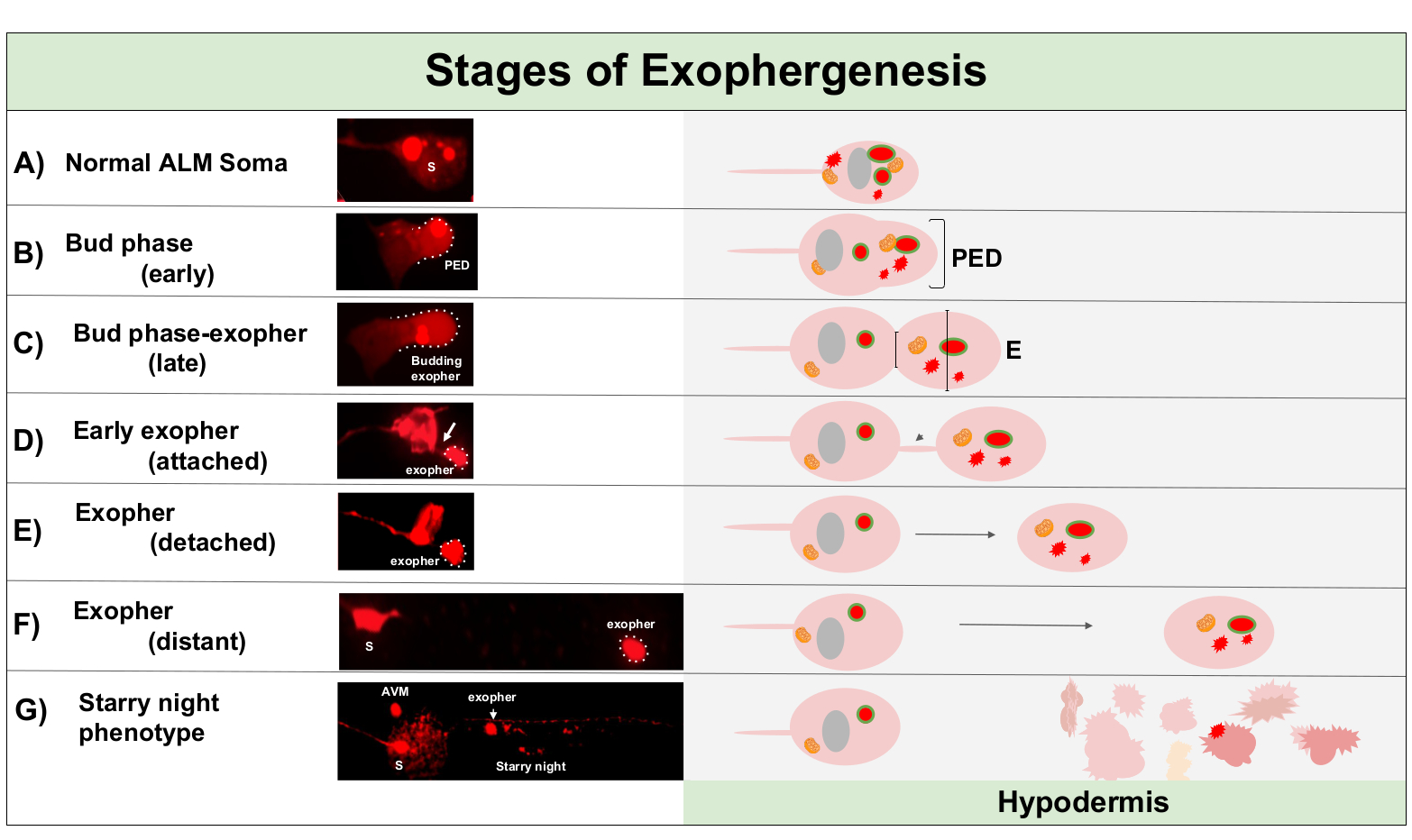
엑소페네시스는 전형적으로 뉴런 세포질의 부종으로 시작되는 동적 과정이다(도1A-B). 세포 내용, 세포기관 또는 단백질 응집체는 뉴런 소마의 한쪽으로 수집되며, 가장 일반적으로 ALMR 뉴런의 후방 끝(투영 중성염에서 멀리 떨어져), 사전 외향 도메인(PED)을 형성한다(도1B). PED가 바깥쪽으로 투영되기 시작하면 초기 돌출이 관찰되어 인식 가능한 돌출된 싹을 형성합니다. 후기 싹은 전구 도메인의 가장 넓은 직경이 소마 엑소퍼넥(도 1C)의수축직경보다 약 1/3 더 큰 경우 정의된다. Exophers는 소마에서 거의 모든 방향으로 배출 될 수 있지만 대부분의 exophers는 세포 체에서 후방으로 빠져 기원하는 소마와 거의 동일한 초점 평면에 남아 있습니다.
싹의 목이 얇은 필라멘트로 좁아지면서 엑소퍼는 원래의 소마에서 멀리 이동할 수 있습니다. 이 필라멘트(도1D, 화살표)를통해 소마에 부착된 상태로 유지될 수 있으며 나중에 분리될 수 있다. 칼슘, 골재 및 미토콘드리아와 같은 세포 내용물을 부착된 exopher5로이 필라멘트를 통해 전송할 수 있지만, 압출된 물질의 대부분은 대규모 신진 이벤트에 의해 이출 구획에 투입된다. 엑소퍼는 눈에 보이는 연결 튜브 또는 얇은 필라멘트가 없고 엑소퍼가 송신 소마(도1E)로부터완전히 분리될 때 성숙한 것으로 간주됩니다.
C. elegans에 의해 생성된 Exophers는 신경세포를 터치하는 즉시 터치 뉴런을 둘러싸고 있는 조직인 피하와 마주치게 됩니다. 가장 일반적으로, exopher 소포는 꼬리를 향해 후면피 내에서 이동하는 것처럼 보이며, exopher 내용이 저하를 대상으로 나타나기 전에 소마에서 상당히 먼 것일 수 있습니다 (예를 들어, 거리는 소마(도 1F)에서100 μm 떨어져 있을 수 있습니다). 형광 발산 소포는 "별이 빛나는 밤"(도1G 및 도 2)이라고불리는 외관을 복용하여 피더피 내의 많은 작은 소포로 분해됩니다. "별이 빛나는 밤" 단계에서, 천시테이트 형광 물질은 원래 독방 exopher에 비해 형광의 많은 작은 지점으로 피하 싱크티움을 가로 질러 흩어져 관찰 될 수있다. 별이 빛나는 밤은 낮은 배율에서 천점을 볼 수 있으며 배율이 높을수록 피시테이트 및/또는 피서피 내에서 네트워크를 형성할 수 있습니다. 별이 빛나는 밤의 형광 신호는 전형적으로 엑소퍼및 뉴런 발현 형광(도2B-C)보다희미하다. 많은 천막 소포로 mCherry의 분산은 피하 세포의 내피 / 리소솜 네트워크와 phagosome 성숙 및 융합을 수반하는 것으로 생각된다. 일부 외신 물질은 피하 리소솜 네트워크에서 저하될 가능성이 있지만, 분해에 강한 잔류 종(예: mCherry 골집)은 피질에서 세포 이물질을 함유할 수 있는 유체 구획인 슈도콜롬으로 버려집니다. 형광 물질은 나중에 coelomocytes(도 2C)에게불린 원격 청소구 세포에 의해 취취해지며, 이는 mCherry의 농축, 저장 및 다시 저하를 시도할 수 있습니다.
,골재 압출 및 전달 현상은 C. elegans5,6,67 및 D. melanogaster8,,9와 같은 유전 모델뿐만 아니라 다중 포유류 모델에서 보고된 필라를 통해 보존된 것으로 나타난다. 엑소퍼와 같은 압출은 포유류 세포10에대해 보고되었으며, 보존된 메커니즘이 골재 및 세포세포 추방의 근간이 될 수 있음을 시사하는 관측이다. 이와 같이 Exopher 생산은 근본적이지만 이전에는 인식할 수 없는 세포 이물질 관리의 보존된 메커니즘일 수 있으며, 이는 불균형이 발생할 때 신경 퇴행성 질환에 적극적으로 기여할 수 있는 신경 프로테오스타증 및 미토콘드리아 품질 관리의 분기입니다. 파편 차별 및 분류에 관여하는 분자의 식별, 뚜렷한 아세포 로케일로의 수송, 압출, 소마와 늦은 추방을 연결하는 관 연결의 형성/가위, 그리고 이웃 세포에 의한 원격 분해를 위한 대형 압출 소포의 인식은 미래의 작업을 위해 남아 있다. 선충 및 비행 모델에 있는 연구 결과는 생리적인 맥락에서 참여 분자를 확인하기 위하여 이 모형에 의해 제안된 편견유전 접근 및 강력한 세포 생물학 공구를 사용하여, 집계 및 세포기관 수집 및 전송의 기계장치를 정의하는 데 매우 중요합니다.
exopher 생물학에서 작동 하는 메커니즘을 해독에 중요 한 첫 번째 단계 생체 내에서 재현 에 대 한 프로토콜을 정의 포함. C. elegans 모델은 신체가 투명하고 형광태그 단백질 이나 세포기관을 포함 할 때 쉽게 관찰 할 수 있기 때문에 이러한 노력에 대한 특별한 이점을 제공합니다. Exophers는 C. elegans 도파민성 뉴런 PDE 및 CEP, ASE 및 ASER 감각 뉴런, 염료 충진 증폭 뉴런5에의해 생성되는 것으로 보고되었다. 터치 수용체 뉴런에 의해 생성 된 exophers가 가장 잘 특성화되기 때문에, 여기에 초점은 exopher 분석을위한 터치 뉴런의 사용에 있습니다. 그러나 기본적인 접근 법은 모든 셀에서 exopher 생산을 측정하기 위하여 적용될 수 있습니다. C. elegans에 의해 생성 된 exophers를 감지 하고 양을 검출 하는 프로토콜 은 mCherry 단백질을 형질적으로 표현하는 수용체 뉴런을 터치, 모니터링 할 수있는 화물에 중점을 두고 점수에 시간적 제약. 이 문서는 생체 내 외신 식별, 그리고 이포 생산을 조절하는 환경 및 유전 적 조건의 양을 향한 접근 방식을 정의합니다. 프로토콜은 기준선 외향성 생산의 측정및 유전자형 에 걸친 비교를 위한 일정한 비응력 조건에 중요한 주의를 강조합니다.
프로토콜
1. exopher 검출에 유용한 균주
- C. elegans의 뉴런 내에서 형광화물을 표현하는 균주를 선택하여 exophers를 쉽게 시각화하십시오.
참고: 표 1은 터치 수용체 뉴런5,11,,12에서생성된 엑소셔를 시각화하는 데 사용된 균주를 나열합니다., 원칙적으로, 임의의 세포 또는 뉴런 타입은 세포 또는 조직 특정 프로모터를 사용하여 압출을 위해 응집되거나 그렇지 않으면 선택되는 형광 단백질의 발현을 유도함으로써 외출 생산을 위해 시험될 수 있다. - 또는, 염료 충진 분석기를 사용하여 앰피드 헤드 뉴런의 엑소퍼를 시각화하여 환경에 개방되어5,,13을백필할 수 있다.
2. 성장 미디어
- 표준방법(14,,15)에따른 배양균에 표준 선충 성장 미디어(NGM)를 준비한다.
참고: 음식의 부족, 또는 플루오로 데옥시리딘 (FuDR), 자손 생산을 차단하기 위해 일반적으로 사용, 극적으로 exopher 생산에 영향을 미칠 수 있습니다. 지속적으로 먹이를 주는 인구를 유지하고 (세균성 음식 피로의 짧은 기간도 피하십시오) 동물을 일정한 온도로 유지하십시오.
3. 일관된 외향적 생산에 중요한 동물 축산
- 일관된 미디어와 일관된 세균성 식품 공급원으로 동물을 키우게 합니다. 동물은 음식 제한이 크게 exopher 생산 수준을 변경할 수 있기 때문에 시간의 짧은 기간 동안, 세균성 음식이 부족해서는 안됩니다.
- 연구 전반에 걸쳐 미디어 레시피와 준비 유니폼을 유지합니다.
참고: 미디어를 변경하면 기저 생산 수준에 영향을 줄 수 있습니다. Agar 배치는 기준선 exopher 수준에 영향을 미칠 수 있으므로 공급 로트 변경 시 날짜를 기록합니다. 건강한 세균성 음식을 보장하고 건조한 천을 방지하기 위해 2 주 후에 스톡 플레이트를 던져 서래 수준에 영향을 미치는 천막 발진의 변화를 일으킵니다. - 기저 조건의 경우 동물을 20 °C의 일정한 온도로 유지하십시오. 가변 온도(온도의 일시적인 변화)에서 동물을 사육하면 최대 외향성 생산 의 타이밍에 변화가 발생할 수 있습니다.
참고: 온도 변동성은 문화 조건에 만국한되지 않습니다. 실험 중이나 실험실 벤치에서의 온도 변화는 영향을 미칠 수 있습니다. 예를 들어 현미경 실 내의 온도는 배양 인큐베이터 또는 실험실 벤치와 크게 다르지 않아야합니다. - 수정 된 계란은 exophers의 초기 성인 생산에 중요 하기 때문에 약리학 안티 불 임 내정간섭을 사용 하지 마십시오.
참고: 플루오로 데옥시리딘(FuDR)16 또는 C2217의사용은 피해야 합니다. 수명 또는 노인 동물 실험을 수행할 때, 연령 동기화 된 인구는 일반적인 약리학 항 불임 개입을 사용하는 대신 박테리아로 퍼지는 신선한 접시에 그들을 따기로 성인을 물리적으로 제거하여 유지되어야합니다. - 오염된 배양을 사용하지 마십시오. 인구 또는 판의 생물학적 타협의 경우에 실험을 다시 시작합니다. 세균성 또는 곰팡이 오염은 동물의 스트레스와 대사 변화를 유발할 수 있으며 실험 인구에서 결석해야합니다.
- 재현 가능한 결과를 극대화하기 위해, 잠재적인 환경 유발 후성 유전학 변경을 피하기 위하여 실험하기 전에 20°C에서 적어도 2개의 건강하고 잘 먹이는, 오염 없는 세대에 대한 배양을 유지하십시오.
4. 표백, 자당 부양 또는 L4 애벌레 따기로 엑서셔 점수를 위한 연령 동기화
- exopher 탐지 패턴은 성인 나이에 따라 다르고 혼합 연령 인구의 동물의 비교가 결과를 혼동 할 수 있기 때문에 실험 인구를 동일한 생물학적 연령으로 유지합니다. 항상 L4 단계에서 "백색 초승달"외음부 형태를 확인하여 실험 동물 인구의 성공적인 동기화를 보장합니다.
참고: 일반적으로, C. elegans mechanosensory ALMR 뉴런에 대한 피크 exopher 생산은 성인 일 2-3(그림 3D)에서발생하며, L4 단계 이후일로부터 측정됩니다. 성인 일 1은 "백색 초승달"외음부 형태(그림 5E)로구별되는 L4 애벌레 단계 후 24 시간입니다. - 중력 성인을 표백하여 동기화 된 계란 인구를 준비합니다.
- NGM 접시에서 자라는 동물을 세척하여 계란으로 가득 한 중력 성인을 수집합니다. 세척하려면 1mL의 M9 버퍼로 접시를 범람하고, 파이프를 위아래로 파이프하여 부유 한 동물과 파이프로 액체를 1.5 mL 마이크로 센트리슈지 튜브로 수집하십시오. 작은 원심분리기로 중력 침전 또는 부드러운 원심분리로 동물을 펠렛하고 상퍼를 제거합니다.
- H 2 O에서 5M NaOH 150 μL과 6% 나트륨 hypochlorite(표백제)의 150 μL을 H2O에 1mL에 넣고 반전으로 약 5분간 섞습니다.
참고: 신선한 표백 액면은 동물 큐티클이 계란 수확을 위해 중단될 수 있도록 합니다. 큐티클을 방해하는 진행은 해부 현미경으로 모니터링 될 수있다; 성인은 표백을 중단해야 하는 시점에서 계란을 깨고 놓아야 합니다. - 20초 동안 소형 원심분리기 튜브가 있는 부드럽게 원심분리기를 제거하고 상체를 제거합니다. M9 버퍼와 원심분리기 1mL을 다시 추가하여 펠릿 위에 약 100 μL을 남깁니다.
- 표백제 용액의 흔적을 제거하기 위해 4.2.3 단계를 두 번 반복하십시오.
- 남은 부피에서 계란을 다시 중단하고 신선한 시드 NGM 플레이트로 옮김합니다. 성인은 lysed 것입니다,하지만 많은 실행 가능한 계란준비에 있어야합니다.
- 시간 시간 별 달걀 레이로 동기화 된 인구를 준비합니다.
- 표준 전송프로토콜(14)을사용하여 시드 NGM 플레이트에 20명의 중력 성인을 선택합니다.
- 동물이 자유롭게 크롤링하고 1.5 시간 동안 계란을 낳을 수 있도록 (낮은 무리 크기의 돌연변이 균주는 더 많은 성인 동물의 도입을 필요로 할 수 있습니다).
- 동기화된 계란 인구를 남겨두고 따기로 접시에서 모든 성인 동물을 제거하십시오. 몇 시간 후에 접시를 확인하여 성인 제거 중에 생존 가능한 성인이 누락되지 않았는지 확인하십시오.
- 계란의 자당 부양 선택에 의해 동기화 된 계란 인구를 준비합니다.
- 중력 동물이 0.1 %의 세제 (예 : Tween 20 또는 Triton X-100)로 M9 용액을 가진 플레이트를 범람하고 15 mL 튜브로 수집하여 적어도 24 시간 동안 계란을 낳은 5 개의 NGM 플레이트에서 동물과 계란을 수집하십시오. 실온에서 부드러운 원심분리에 의해 펠릿 성인 (30 초 2,000 x g).
- 신선한 M9의 15mL에서 수퍼네티드를 제거하고 씻어 내고, 세척 후 상체를 버리고, 동물과 계란에 펠릿을 풍부하게 유지하십시오.
- 2mL의 상체를 유지하고 펠릿을 다시 중단하십시오. 볼륨 자당에 의해 60 % 무게의 2 mL을 추가합니다.
- 2000 x g의 원심분리기는 5분 동안. 이제 용액은 계란이 풍부하게 농축된 상층을 표시합니다.
- 상층의 약 2.5mL를 새로운 15mL 튜브로 옮기고 10mL의 M9을 추가한다.
- 반전으로 1분 간 섞은 다음 원심분리기 2000 x g를 1분 간 섞습니다.
- 상체를 제거하고 M9에서 계란이 풍부한 펠릿을 씻으십시오. 달걀 펠릿의 10-15 μL은 신선한 OP50 시드 NGM 플레이트에 분배될 수 있다.
참고 : 이 방법은 계란의 큰 숫자를 준비; 수집된 동물이 OP50 대장균 사료가 부족하도록 허용하지 않습니다.
- 개발의 L4 단계에서 동물을 따기하여 동기화 된 인구를 준비합니다.
- 위에서 설명한 대로 시드 NGM 플레이트에서 동물을 성장시다.
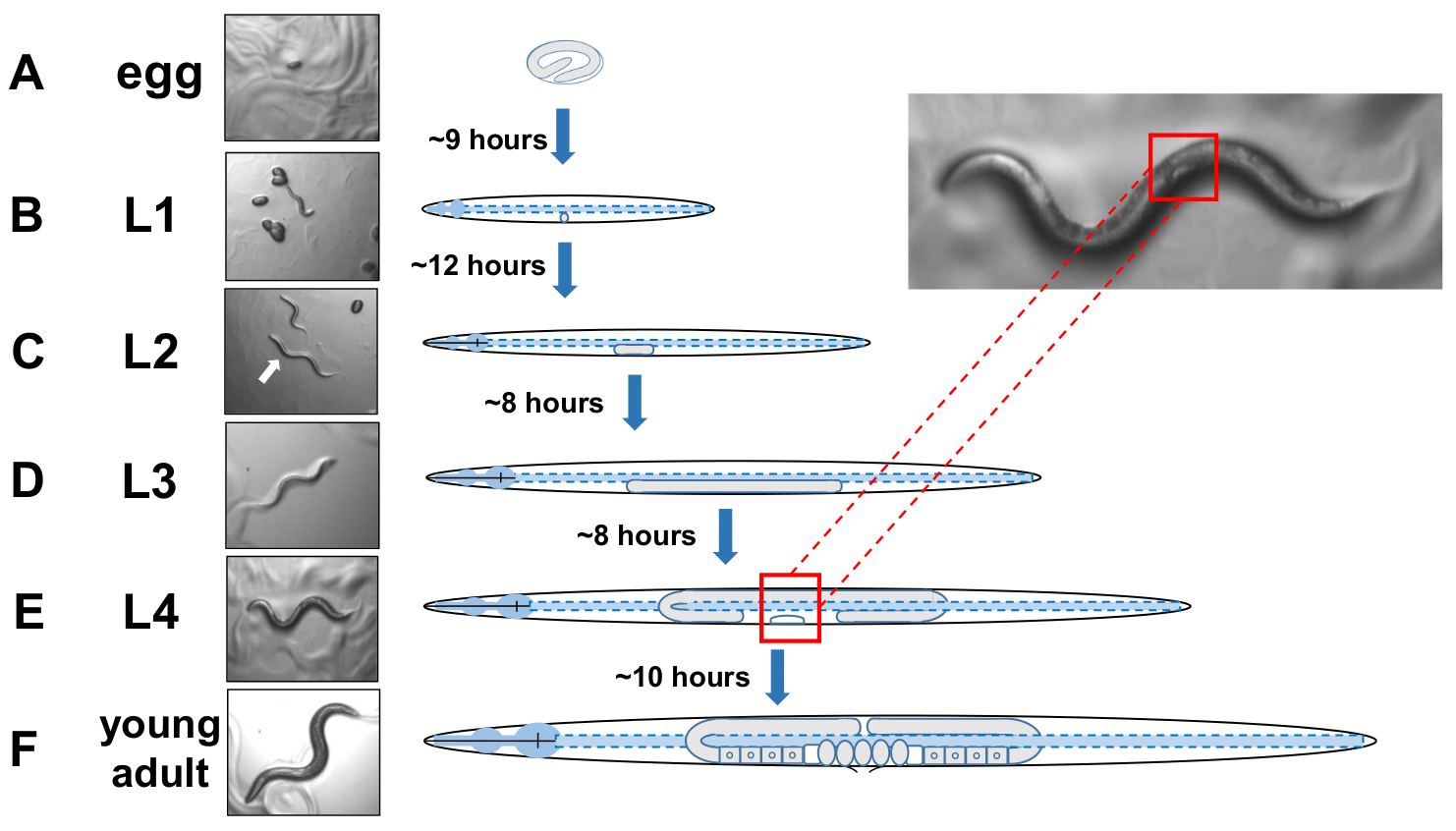
참고: C. 예르간은 네 단계의 개별 단계로 발전합니다. 20°C에서 새로 놓은 알은 부화하는 데 약 9시간이걸립니다(그림 5A). 포스트 해치, 동물은 애벌레 단계 1 (L1)을 통과 하여 애벌레 단계 4 (L4)로 통과하며 각 단계는 각 몰트(그림 5A-F)사이에8-12 시간 동안 지속됩니다. 따라서 계란을 접종하여 제조 한 접시에는 계란이 도입 된 후 약 40 시간을 선택하는 많은 L4 동물이 있어야합니다. - 개발 외음부의 흰색 반달 초승달 모양을 찾아 L4 단계의 동물을 식별(그림 5E).
참고: L4 단계의 동물은 크기와 신체 색소 침착이 균일합니다. 단계적 동물의 검사를 위해 신선한 성장 접시에 흰색 초승달 동물을 선택합니다. 다음 날(~ 24시간 후)은 성인 1일로 계산해야 합니다. - 성인 2 일째에 매일 동물의 개체수를 기록합니다.
참고: Exophers는 일반적으로 기저 조건하에서 피크 exopher 생산인 성인기의 2일째에 득점됩니다. 그러나, 엑소페네스피크와 시기는 연구되고 있는 환경적 또는 유전적 변화에 의해 이동될 수 있기 때문에, 가장 포괄적인 그림을 생성하기 위해 4일 동안 매일 성인 동물의 인구를 채점하는 것이좋습니다(그림 3D).
- 위에서 설명한 대로 시드 NGM 플레이트에서 동물을 성장시다.
5. 형광 현미경을 사용하여 exophers의 검출
- 형광 현미경에 대 한 장착 된 높은 배율 의사-스테레오 해부 현미경을 사용 하 여 exophers를 관찰.
- NGM agar 플레이트 표면에 10-100 mM 레바미솔/테트라미솔 용액의 100-200 μL을 파이프팅하여 NGM 플레이트에 동물을 고정합니다. 2-4 분 후, 동물은 마비되고 천판에서 직접 관찰 할 수 있습니다.
참고: 고정 처리는 절대적으로 요구되지 않으며, 훈련된 눈, 신경 식별 및 exopher 존재는 exopher가 생성되었는지 여부를 결정할 때 플레이트의 현미경으로 동물을 크롤링하여 시각적으로 다음하여 득점될 수 있습니다. - 100x의 총 배율을 사용하여 형광 뉴런을 관찰하여 exophers의 해부 현미경 검출을 수행하십시오.
참고: 해부 현미경 검사를 사용하여 exopher 이벤트를 채점하면 사육되는 한천 판에서 직접 비교적 쉽게 많은 수의 동물을 관찰할 수 있습니다. - 공초점 현미경 검사를 사용하여 exopher 연구를 위한 살아있는 화상 진찰 및 장착 기자 긴장
- 공초점 현미경을 사용하여 세포 내 역학 및 엑소더니시스의 특성을 살수 있습니다.
참고: 라이브 이미징은 exopher 생산이 동적 공정이기 때문에 exopher 생산의 미묘한 세부 사항을 관찰하는 유리한 방법입니다. - 10-100mMMMM에서 레바미솔 또는 테트라미솔의 사용 또는 하이드로겔 폴리스티렌 마이크로비드(직경 15μm, 30 μm 또는 40 μm)의 적용을 포함하여 편리한 방법을 사용하여 고해상도 라이브 이미징을 위한 동물 이동을 제한한다18.
- 공초점 현미경을 사용하여 세포 내 역학 및 엑소더니시스의 특성을 살수 있습니다.
- 화합물 및 공초점 현미경 검사법을 위한 슬라이드 준비
- 현미경 슬라이드에 고정 에이전트에 20-50 동물을 마운트. 13mm 직경의 제기 링으로 그려진 재사용 가능한 고리 세포론 슬라이드는 장착에 유용합니다.
- 살아있는 동물을 10-100 mM 레바미솔 또는 테트라미솔과 같은 마비의 5-20 μL로 선택하십시오.
- 마비를 위해 4 분을 기다린 다음 커버 슬립 (11/2 호 (0.16 - 0.19 mm) 또는 2 호 (0.17 - 0.25 mm)로 슬라이드를 덮습니다.
- 소량의 동물 장착
- 장착 된 동물을 분쇄하지 마십시오; 슬라이드 당 몇 마리(20미만) 동물만 관찰할 때, 커버슬립의 불평등한 압력으로 인해 일부 동물을 분쇄할 위험이 있습니다. 이 위험은 장착에 대한 낮은 비율의 아가로즈 패드를 사용하여 최소화 될 수있다.
- 2-4% 아가로즈 패드 슬라이드를 한 다음 패드에 2-15 μL의 마비 용액을 추가합니다. 레파미솔과 테트라미솔이 패드에 확산되어 유효 농도를 줄입니다.
- 한천 패드에 놓인 마비 용액 또는 마이크로비드의 2-15 μL 방울로 동물을 선택하여 마운트하십시오. 커버슬립을 위에 놓고 동물이18을그대로 유지되어 있는지 확인하십시오.
- 한천 패드 준비
- 2% 화마 패드를 준비하려면, 아가로즈가 균일하고 용융 된 상태에 될 때까지 M9 용액과 전자 레인지에 2 % 아가로즈를 가열합니다.
- 충분한 품질의 한천 패드를 얻으려면 20 초 이내에 저전력에서 대체 믹싱 및 마이크로 웨이브를 사용합니다. 가열 블록에 끓는 천을 배치하고 거품이 표면으로 상승 할 수 있도록하여 패드 내에 기포가 포함되지 않도록하십시오.
- 파스퇴르 파이펫을 사용하여 상승 된 거품 아래용액 깊은 곳에서 한천을 그립니다.
- 두 개의 테이프 슬라이드를 준비하고 평평한 표면에 깨끗한 유리 현미경 슬라이드의 양쪽에 배치합니다. 테이핑 된 슬라이드를 만들기 위해 각 슬라이드(그림 6A)에실험실 테이프 2 개를 배치합니다.
- 파스퇴르 파이펫을 사용하여, 테이핑 된 슬라이드(그림 6B)사이에 끼워 깨끗한 현미경 슬라이드에 한 방울의 천지를 놓습니다.
- 테이프슬라이드(그림 6Cc)를가로질러 네 번째 깨끗한 슬라이드로 용융 된 한천방울을 조심스럽게 빠르게 덮습니다.
참고 : 슬라이드는 용융 된 한천을 약 0.4mm 두께 (테이프두께)에 평평한 원으로 부드럽게 눌러야합니다(그림 6D). 한천은 빠르게 식어야 합니다. - 상단 슬라이드를 미끄러져 제거합니다(그림6E). 한천 패드는 빠르게 건조하고 몇 분 이내에 가장 잘 사용됩니다. 상단 슬라이드가 제거되면, 동물을 장착즉시 젤 패드를 사용합니다. 기포가 있는 패드를 사용하지 마십시오.
- 두 유리 슬라이드 사이에 최대 30분 동안 한천 패드를 보관합니다. 말린 한천은 동물이 뭉쳐서 담정합니다. 마비 용액 또는 마이크로 비드의 2-15 μL 이내에 동물을 마운트하고 커버 슬립으로 커버; 마비 및 장착 20 분 이내에 슬라이드를 화면(그림 6).
참고: 스트레스 조건은 exopher 비율을 변경할 수 있기 때문에, 외향성 에 대한 검사 시 산화 스트레스를 유발할 수 있는 마비를 피하십시오(예: 아지드 나트륨).
- 회전 디스크 공초점 현미경을 사용하여 이모의 검출
- 63x 및 100x에서 1.4 수치 조리개 목표를 가진 세포 기관 및 기타 내용과 같은 세포 생물학적 특징을 관찰한다.
- 다차원 인수를 활용하여 단계 제어 및 이미지 수집이 가능한 소프트웨어를 사용합니다. 현미경 및 이미지 처리 소프트웨어는 또한 이 단계가 표준 화상 진찰 접근을 관련시키는 때 화상 진찰 및 데이터 수집에 적합해야 합니다.
6. 터치 뉴런을 식별하고 장착 된 동물과 함께 exophers에 대한 점수
- 산 마비 성인 동물(그림 6).
- 원하는 Z 평면을 식별합니다. 낮은 배율 밝은 필드(10-40x)를 사용하여 동물의 적합한 Z-평면을 식별하여 동물의 위치, 머리 꼬리 방향 및 외음부의 위치를 기록합니다 - 이는 후기 뉴런 및 엑시퍼 식별을 위한 랜드마크입니다(그림3A & 도 5E).
- 선택한 기자의 형광 신호에 초점을 맞춥니다. 같은 Z-비행기에 머무르고 선택한 사이토솔릭 리포터를 위해 10-40x에서 와이드필드 형광으로 전환하십시오.
참고: 이 예에서 형광발현은 메크-4 메카노감각 터치 뉴런 특이적 프로모터에 의해 구동된다. 높은 복사 배열 및 다른 형광은 발현의 가변성을 가지므로 변액 형광 강도가 있습니다. 필요한 경우 조정합니다. - Z 축 내에서 스크롤하여 초점 평면에서 동물의 깊이와 형광식을 관찰합니다. 이렇게 하는 동안 머리 꼬리 방향을 확인합니다. 머리/인두는 형광 신경 링을 가지며,이 경우 꼬리에는 1-2 개의 눈에 보이는 PLM 소마(그림 3A)가포함됩니다.
- 터치 뉴런 식별
- 동물이 왼쪽 또는 오른쪽에 장착되었는지 여부를 식별합니다(그림3A).
참고: 동물의 3차원성을 고려할 때, 광학에 가장 가까운 측면에서 최고의 이미징 해상도가 수행됩니다. - 관찰하여 소마 (ALM, ALMR, AVM)를 식별 - 신경 반지 및 측면 신경 과정을 식별하기 위해 머리에서 시작합니다.
- 10-40배율에서 Z축을 천천히 스크롤하여 연결된 프로세스를 식별합니다.
- 프로세스가 확인되면, 소마가 명백해질 외음부를 향한 후부 방향으로 측면으로 따라, 과정의 끝에 둥근 세포 본문에 의해 표시. 일단 가장 초점 신경 세포 소마 발견, 그것은 다음과 같이 다른 신경 랜드 마크를 사용 하 여 확인 될 수 있습니다.:
- 근처 복부 뉴런인 AVM을 사용하여 동물 오리엔테이션을 할당합니다. AVM 뉴런이 ALM과 동일한 평면에 있는 경우 동물은 옆으로 쉬고 그 비행기 외부의 뉴런은 ALMR입니다. AVM 뉴런이 문제의 ALM과 동일한 평면에 없는 경우 초점 평면에 가장 가까운 터치 뉴런은 ALML입니다.
- 전방 터치 뉴런이 동일한 평면에 있는지 여부를 나타내기 위해 꼬리 근처에 위치한 또 다른 복부 터치 뉴런인 PVM 뉴런을 식별합니다. 그렇다면 관찰된 터치 뉴런은 ALML입니다.
- 관심 영역 근처의 다른 소마 몸체의 위치(소마의 양쪽에 위치한 형광 뉴런) 및 모든 Z-평면에서 명확한 초점으로 가장 깊은 뉴런을 설정할 수 없더라도 감각을 얻으십시오.
참고: 모든 터치 뉴런 솜마의 식별은 초점이 닿지 않는 소마가 엑소셔스로 오인 될 수 있기 때문에 중요합니다.
- 동물이 왼쪽 또는 오른쪽에 장착되었는지 여부를 식별합니다(그림3A).
7. 추방자 식별 및 점수 매기기
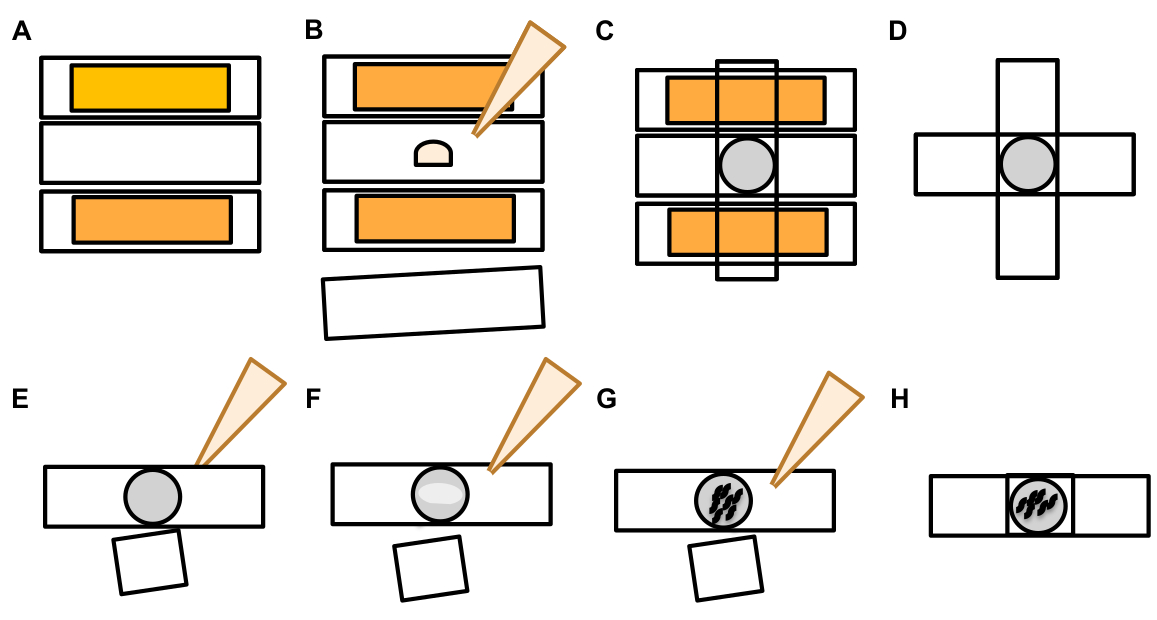
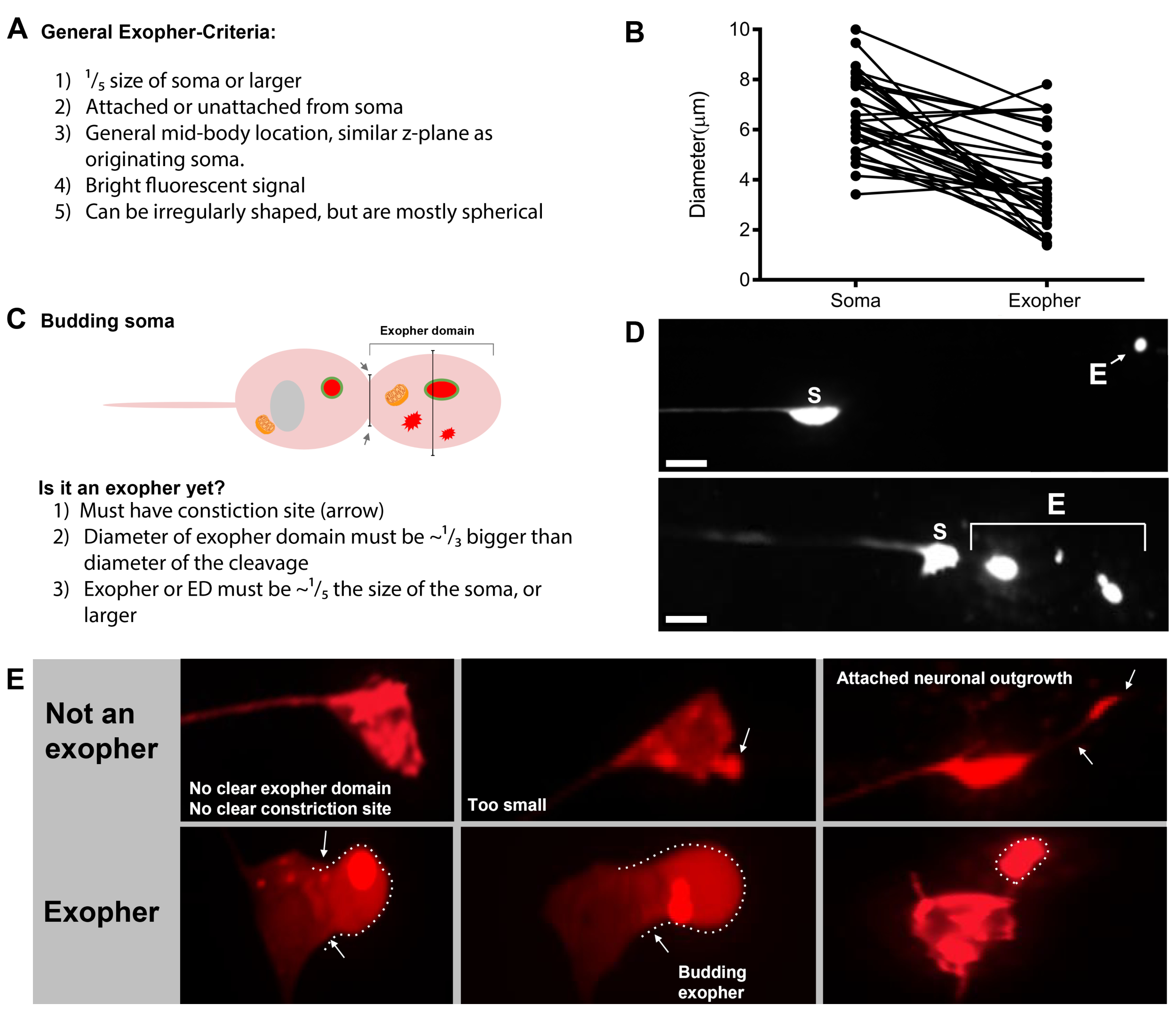
- 터치 뉴런이 발견되면, 큰 돌출 (exopher 도메인)에 대해 검사하여 싹 이압으로 간주 될 만큼 충분히 큰 (기원 된 소마의 크기의 적어도 1/5에 도달)(도1C).
참고: 평균 외음은 직경 이 2-8 μm 정도를 측정하고, a(ZB4065 bzIs166[Pmec-4:mCherry])의평균 소마는 2일째 성인 2일째에 6-10 μm을 측정합니다(그림7B). - 싹이나 외향적 도메인이 관찰되지 않으면, 소마에서 방출되는 부착 된 얇은 필라멘트에 대한 신경 소마를 검사합니다. 부착된 exophers는 유래 소마와 유사한 Z 평면에 더 가깝게 위치하는 경향이 있습니다.
참고 : Exophers는 항상 소마에 부착 된 상태로 유지되지 않습니다. 연결된 필라멘트의 검출은 개체가 exopher임을 나타내는 확실한 표시입니다. - 첨부되지 않은 exopher를 식별하려면 exopher의 내용을 찾습니다. Exophers는 추방된 형광 단백질을 농축할 수 있고 그러므로 수시로 소마 보다는 밝습니다.
참고: 엑소퍼의 내용은 이질적이고 가변적입니다. 리소좀 및 미토콘드리아와 같은 세포 세포기관은 또한 엑소퍼(도 4Figure 4C-E)내에서압출될 수 있다. - 원래 소마가 발견된 비행기와는 다른 초점 평면에서 부착되지 않은 외향적 외피를 찾습니다. 엑소퍼는 어떤 방향으로든 ALM 소마에서 돌출하는 것으로 보였지만, 외향체가 신경 과정에서 후방 방향으로 소마에서 튀어나온 것이 일반적입니다.
- 뉴런 소마로 위치및 식별되지 않은 크고 구형 물체를 확인합니다. Exophers불규칙하게 형성될 수 있지만 일반적으로 구형 구조입니다. Exophers는 시간이 지남에 따라 저하되기 때문에 나이가 많은 exophers는 더 불규칙한 모양을 갖는 경향이 있습니다.
참고 : 성숙하거나 오래된 exophers는 exophers의 밝은 형광 강도와 구형 모양을 통해 분산 된 "별이 빛나는 밤"단계와 구별됩니다. - 이전 엑소페네스의 증거로 "별이 빛나는 밤"표현형을 조사하십시오. 엑소퍼는 더 작은 소포로 분해되고 주변 의 피규어가 exopher 내용(그림 1G,2B, 3B 및 7A)을저하시키려는 시도로 "별이빛나는 밤"단계로 진행됩니다.
참고: 별이 빛나는 야간 스테이지는 구조적 무결성을 잃고 터치 뉴런 및 엑시퍼 구조에 비해 희미한 형광을 표시하는 단편화및 산란(때로는 네트워크) 형광 엔티티로 표시됩니다. - "여러 exopher 이벤트"의 인스턴스를 찾습니다. Exophers는 일반적으로 단수 발생(1soma에서 발산되는 1 엑서)으로 생성되지만, 어떤 상황에서는 하나 이상의 엑소퍼가 단일 소마(도7D)로부터방출될 수 있다.
참고: 성숙한 exophers 는 피더피에서 저하될 때 여러 소포로 저하될 수 있습니다. 각 exopherher가 독립적 인 exophergenesis 이벤트에 의해 생성되었는지 또는 추가 소포를 만들기 위해 하나의 원래 exopher 분할 여부를 구별하는 것은 시간 경과 관찰에 의해서만 결정될 수 있습니다. - 모든 형태학적 이상이 exophers로 성숙하지는 않는다는 것을 명심하십시오.
- 비하된 소마를 exopher로 점수를 매기지 마십시오. 확장 또는 뾰족한 소마는 때때로 관찰 할 수 있습니다 (특히 나이 또는 스트레스 미만), 하지만 명확한 수축 사이트없이 확장은 exopher로 득점되지 않습니다.
- exopher 이벤트 정량화에서 소마의 크기1/5를 달성하지 않는 작은 해결 된 싹을 거부합니다.
- 신경질적인 성장을 exophers로 계산하지 마십시오. 성숙한 중성염은 나이 (일반적으로 신경 과정의 반대 방향으로) 극적으로 확장될 수 있고 형광 단백질은 그 같은 구조물의 말단으로 이동할 수 있습니다19.
참고 : 이러한 중성자 아웃 성장은 며칠 및 몇 주 동안 뚜렷한 발달 패턴을 가지고 싹을 형성하지 않으며 분리하지 않기 때문에 exophers가 아닙니다(그림 7E).
- exophers가 아닌 형광 실체를 식별합니다.
참고: 압출형 형광실과 자동형광의 정확한 식별을 보장하기 위해 배경 형광에 대한 아이디어를 얻는 것이 중요합니다.-
트랜스제닉 형광 발현 대 자동 형광. 형질 전환식에 대한 자동 형광을 착사하지 마십시오. 진정한 exopher 신호는 장이나 창자에 있지 않을 것입니다 (DIC 확인은 이 조직을 식별하는 데 사용될 수 있습니다) 및 exopher 신호는 배경 자동 형광보다 상당히 밝아질 것입니다.
참고: 자동 형광은 창자 과립 장 형광 색소 침착에 기인하고 나이와 함께 축적됩니다. 그것은 이종성, 특히 다른 파장으로 볼 수 있습니다. - 배아로부터의 신호. 엑소더스 발생에 대한 배아 신호를 착사하지 마십시오. 형광에서 밝은 필드 조명으로 전환하고 자궁에서 계란과 신호의 연관성을 확인하여 배아 신호의 의심을 확인합니다.
-
비행기에서 또는 근처의 소마 시체에서. 관측의 시작 시 근처의 모든 소마 시체, 심지어 초점이 벗어난 소마를 식별하여 exopher에 대한 비행기 소마의 오해를 피하십시오.
참고: ALMR의 exophers에 대한 점수를 매기는 경우 AVM 및 ALMR 소마의 위치를 식별하고 설명합니다. 소마 신체 식별에 대한 자세한 내용은 도 3A에설명되어 있습니다.
-
트랜스제닉 형광 발현 대 자동 형광. 형질 전환식에 대한 자동 형광을 착사하지 마십시오. 진정한 exopher 신호는 장이나 창자에 있지 않을 것입니다 (DIC 확인은 이 조직을 식별하는 데 사용될 수 있습니다) 및 exopher 신호는 배경 자동 형광보다 상당히 밝아질 것입니다.
8. 채점 및 통계
- 바이너리로 점수 exophers (예, exopher / 아니오, exopher가 없습니다).
- 주어진 뉴런에 대한 "exopher 이벤트"로 exopher 검출을 고려하십시오. 엑소퍼 이벤트는 소마 또는 여러 엑소퍼 근처의 단일 엑소퍼를 관찰할 수 있습니다.
참고: 개별 엑소처생성 이벤트의 수를 정량화하려면 시간 경과 관찰을 사용합니다. - 다른 세포가 동일한 속도로 엑소퍼를 생성하지 않기 때문에 특정 식별 된 셀 당 exopher 이벤트를 계산합니다 (예 : 그림 3C참조). ALMR 뉴런은 본원에 기재된 균주에서 가장 많은 기준선 엑소퍼를 생성하므로 종종 터치 수용체 뉴런으로부터 의정화를 위해 선택된 세포이다.
- 통계의 경우, 일반적으로, 적어도 3개의 생물학적 시험을 실시, 적어도 30 동물은 중단의 분석에 필요한 관측의 해당 수와 시험 당 득점.
- 대조군과 비교하여 하나 또는 두 개의 돌연변이/처리를 관련시키는 다중 예심을 위해, Cochran-Mantel-Haenszel 시험은 p 값을 결정하기 위하여 적정합니다.
- 대조군과 비교하여 두 개 이상의 돌연변이를 포함하는 시험의 경우, 이진 물류 회귀 분석을 사용하여 모든 수의 범주 예측 변수에 대한 중요성을 평가하는 것이 적절합니다.
결과
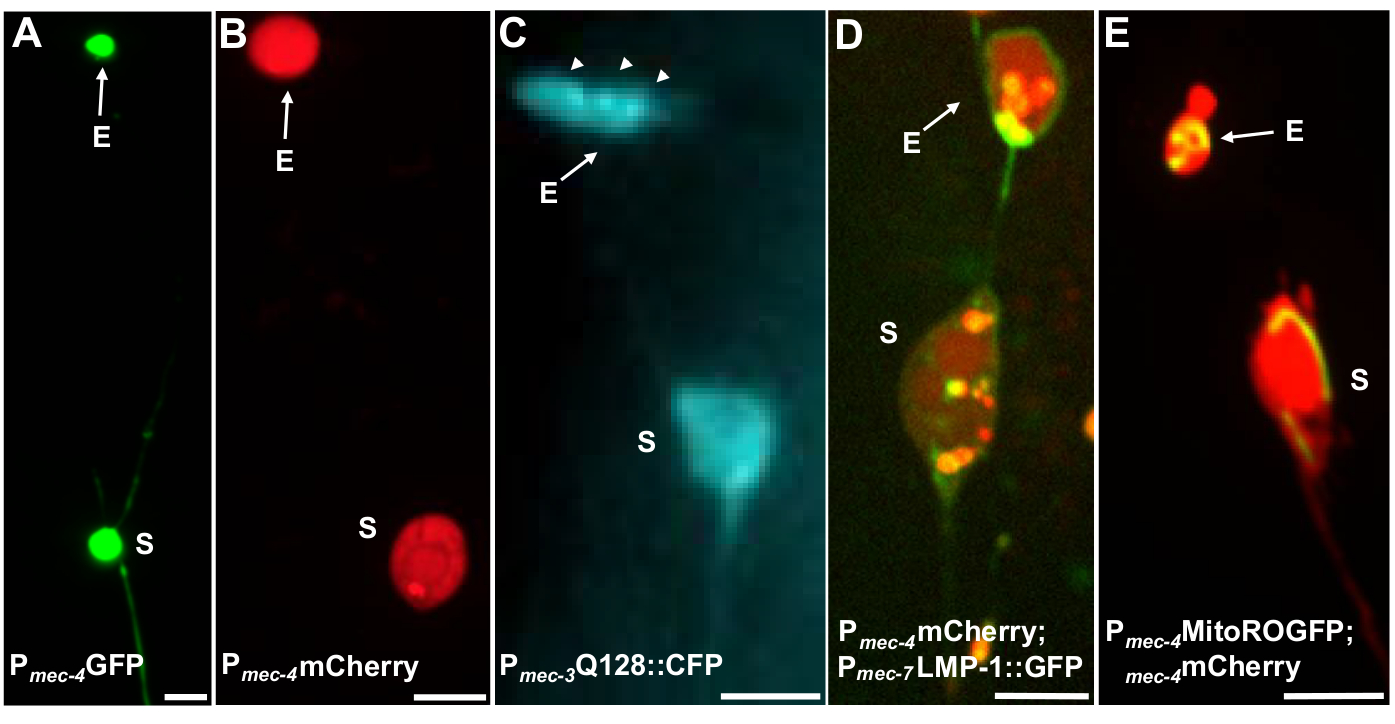
여러 형광 기자는 엑소셔를 측정하는 데 사용할 수 있습니다. 터치 뉴런 엑소퍼는 압출을 위해 선택할 수 있는 단백질의 형광 태깅, 압출될 수 있는 세포기관을 표시하거나 세포막을 태그하여 생체 내에서 쉽게 시각화됩니다. 표 1은 피규어 4에포함된 대표적인 예와 함께 엑소셔를 모니터링하는 데 사용된 터치 뉴런 발현 형광 기자를 식별한다. 외피로 압출되는 것으로 알려진 화물은 인간 헌팅틴의 N단말 도메인을 확장 된 폴리 글루타민 (Q128)(도4B)으로융합하고, 리소솜은 리소솜 관련 막 단백질 (LMP-1)(도4C)및 미토콘드 타그리아(그림4C)를포함한다. 세포질 GFP는 강하게 추방되지 않으며 GFP가 약하게 엑소퍼(그림 4A)를시각화 할 수 있지만, 소마5에서우선적으로 유지됩니다. GFP가 추방되는 단백질에 융합되면 이 태그를 사용하여 exophers를 시각화할 수 있습니다. 중요한 점은 다른 단백질을 태그함으로써 특정 화물및 세포기관의 추방뿐만 아니라 외향체를 구성하는 단백질과 막에 대한 많은 질문을 해결할 수 있다는 것입니다.
의사 스테레오현미경 설정은 천 판에 동물의 외피를 볼 수있는 효과적인 도구입니다. 이 설정은 각 배율에 높은 숫자 조리개 광학, 의사 스테레오 기술 (입체 베이스에 대한 이산 목표), 설치 된 목표에 배율 중간에서 볼 수있는 줌 작동 스위치를 포함하는 화합물 및 입체 기술의 하이브리드입니다. 이와 같은 현미경은 10배 의 안구와 고처리량 점수에 대한 신경 형태 및 발산 생산을 관찰할 수 있을 만큼 강력한 목표를 장착해야 합니다(스캔/따기에 사용되는 2배 목표, 식별 및 채점에 사용되는 10배 목표).
표준 스테레오현미경의 배율 기능은 일반적으로 형광 단백질을 표현하는 터치 뉴런의 네트워크를 볼 수 있을 만큼 충분한 해상도를 가지고 있지만, 표준 해부 현미경은 soma의 관 연결과 같은 외형의 세포 이하 세부 사항을 관찰하기에 충분하지 않습니다. 이러한 관측은 공초점 현미경 검사를 필요로한다 (장비 세부 사항에 대한 재료의 표 참조).
Exopher 수량 연구는 실험적 스트레스를 제거하기 위해 엄격한 제어가 필요합니다. 재현 가능한 exopher 생산을 위해서는 일관된 성장 조건의 세심한 유지 보수가 필요합니다. 보다 구체적으로, exopher 생산은 일관된 공급, 일정한 온도 및 수 세대에 걸친 오염없는 성장이 재현성에 매우 중요합니다. mCherry의 높은 신경 발현을 가진 기저 성장 조건하에서, exopher 생산은 상대적으로 낮습니다 (ALM의 5-25%는 exophers를 생성합니다) 그러나 몇몇 스트레스는, 삼투성 및 산화 스트레스를 포함하여, exopher 비율을 증가시킬 수 있습니다. mCherry 발현은 스트레스로 생각할 수 있지만, exopher 수준의 스트레스 민감도의 조합은, 제대로 제어하면, 실험적 스트레스 도입이 보다 쉽게 유도하고 외신을 관찰하는 전략이 될 수 있다는 것입니다.
타이밍과 예상 exopher 생산 수준. 추방자는 애벌레 발달 중에 사실상 결석합니다. 젊은 성인 생활에서 피크 exopher 생산의 기간은 성인 일 동안 높게 제한 될 것으로 보인다 1-4, 가장 일반적으로 성인 일 2 또는 3에서 분명 되 고. 피크가 조금 앞서 또는 뒤로 이동할 수 있기 때문에, exopher 생산 프로필의 가장 완벽한 평가는 성인 일 1-4에 매일 여러 시험을 득점하는 것입니다. 일반적으로 ALMR은 소포가 적어도 24 시간 동안 지속되는 하나의 주요 exopher를 생산합니다. exopher는 상당히 빠르게 생산 할 수 있습니다 (가장 빠른 분 순서에). 가장 일반적으로, 단지 하나의 주요 exopher 초기 성인 생활에서 뉴런 당 생산, 하지만 여러 exophers의 생산 가능.
기초 조건하에서 mCherry를 표현하는 AlMRs에 의한 일반적인 exopher 생산은 성인 일 2-3(그림 3D)의최적 기간 내에 조사 된 ALM의 5-25 %의 범위입니다. 프로테오스타증 위기5뿐만아니라 다른 응력에 노출하면 엑시셔 레벨을 조절할 수 있습니다. 스트레스 또는 유전 적 혼란은 외출압을 생성하는 ALMR 뉴런의 90 %의 높은 검출 속도로 exopher 생산을 증가시킬 수 있습니다.
exophergenesis에 있는 특정 유전자의 역할을 시험하기 위한 공급 기지를 둔 RNAi. 선충 C. 엘레간은 일반적으로 관심있는 유전자를 대상으로 이중 좌초 RNA (dsRNA)를 표현하는 대장균 변형 HT115 변형 동물을 공급하여 RNAi 노크를실시한다 20. HT115 박테리아는 RNAi5를먹이는 데 에 있는 exophers를 위해 득점할 때 사용될 수 있습니다. 대부분의 조직에 있는 성적 증명서는 이 기술을 사용하여 RNAi에 의해 표적으로 할 수 있는 동안, 뉴런은 더 내화입니다. RNAi에 대한 감도는 뉴런 특이적 프로모터하에서 형질전환 dsRNA 수송기 SID-1을 발현하는 동물을 사용하여 교정될 수 있다. 이런 식으로 신경 조직은 RNAi21에민감하게 될 수 있다.
관심 있는 유전자의 조직 특이적 녹다운은 그 성분에서 결핍되는 돌연변이 내내 내인성 RNAi 대사의 성분을 표현함으로써 달성될 수 있다. 예를 들면: Argonaute 단백질 RDE-1은 동물이 그 유전자를 표적으로 하는 RNAi 내정간섭에 드러낼 때 만 신경에 있는 관심있는 유전자의 녹다운을 달성하기 위하여 rde-1 돌연변이 동물의 뉴런에서 구체적으로 표현될 수 있다.
표준 선충 RNAi 프로토콜 을 사용하여20,,22,RNAi에 L4 단계에서 부모의 노출과 성인이 강한 유전 적 노크를 생성할 때까지 변형 된 HT115 박테리아를 소비하는 그들의 자손을 허용하지만 실험 동물이 빈 대조군 벡터와 다르게 성장할 수 있으므로 RNAi에 의해 유도 된 잠재적 인 발달 지연에 세심한. 항상 음수 제어 비교를 위한 빈 벡터 컨트롤을 포함하는 것이 중요합니다. HT115 박테리아는 RNAi를 먹이기에서 exophers를 위해 득점할 때 사용될 수 있습니다. 그러나, 몇몇 유전자는 RNAi 노출5의짧은 기간 도중조차 exophergenesis 비율을 변경에 효과적이다는 것을 주의합니다. 특정 유전자의 표적이 발달 실패로 이끌어 내는 경우에, 일생 녹다운에 동물을 노출하는 것을 피하기 위하여, 동물은 단순히 L4에서 성인 D2 또는 D3에 노출을 위해 RNAi 접시에 L4 단계에서 포착될 수 있습니다.
| 스트레인 이름 | 유전자 형 | 설명 | Exopher 백분율 | 참조 |
| SK4005 | zdIs5[Pmec-4GFP] | 터치 뉴런에서 GFP의 세포경식 발현. | 1-8% ALM | 그림 4A, 멜렌티예비치 2017 |
| ZB4065 | bzIs166[Pmec-4::mCherry] | 터치 뉴런에서 mCherry (bzIs166)의 과발현은 세포경 신호와 mCherry 골재를 모두 생성합니다. bzIs166은 엑소퍼 유도제입니다. mCherry 집계는 엑소퍼넥시스의 예측변수이며, 외향적으로 압출됩니다. | 3-20% ALM (정상 조건). 20-80% ALM (금식 조건). | 그림 4B, 멜렌티예비치 2017 |
| ZB4067 | bzIs167[Pmec-4미토그fp Pmec-4mCherry4]; igIs1[Pmec-7YFP Pmec-3htt57Q128::cfp lin-15+]; | YFP 세포학적 으로 메크-7 터치 뉴런을 라벨. 공동 발현 된 Q128::CFP 집계 및 엑소퍼를 유도합니다. CFP는 우선적으로 침묵합니다. | ~25% | 그림 4C, 멜레티예비치 2017 |
| ZB4509 | bzIs166[Pmec-4mCherry]; bzIs168[Pmec-7LMP-1:::GFP] | bzIs168 LMP-1::GFP 라벨 플라즈마 멤브레인 및 리소경 멤브레인. bzIs168신경막, 엑소셔(막 바운드) 및 리소경 막 구조를 식별하는 데 사용할 수 있습니다. | 3-20% ALM | 그림 4D, 멜렌티예비치 2017 |
| ZB4528 | bzIs166[Pmec-4mCherry]; zhsEx17 [Pmec-4미톨::ROGFP] | Allele zhsEx17은 국소 산화 환경에 따라 피크 흥분 파장을 405nm(산화)에서 476nm(감소)로 변경하는 미토콘드리아로 국소화된 기자입니다. 그것은 터치 뉴런에서 발현되고 터치 뉴런과 미토 엑소셔에서 미토콘드리아를 식별하기 위해 자체적으로 사용할 수 있습니다. | 3-20% ALM 프로테오 엑소퍼. % ALM 미토 -exopher 수량 진행중입니다. | 그림 4E, 멜렌티예비치 2017, 캐논 2008, 고스 2013 |
표 1. 터치 뉴런의 시각화에 사용 된 균주, 터치 뉴런 엑소셔, 및 exopher 내용.

그림 1: 엑소퍼제네시스의 단계. 굴레를 만들고 배출하는 과정은 'exopher-genesis'라고 합니다. 엑소퍼 형성의 동적 과정은 몇 분에서 몇 시간 정도 걸릴 수 있습니다. 묘사된 것은 높은 엑소퍼 생성 균주, ZB4065 bzIs166[Pmec-4mCherry]에서 동적 엑소페네시스 과정 동안 특정 단계에서 소마 및 exopher 형태의 예입니다. 모든 이미지는 100x 목표로 촬영한 2일 성인 ALM 뉴런입니다. (A)정상 소마. 성인 메카노 감각 터치 뉴런 ALM 형질적으로 표현 Pmec-4mCherry. 묘사 된 소마 형태는이 균주에 있는 젊은 성인 뉴런의 전형입니다, 세포질에 있는 mCherry 농도와. (B)조기 싹 단계. 외습제의 첫 번째 관찰 단계는 소마 막의 가장자리에 선택된 세포질 물질의 편광을 포함한다. 이 단계는 종종 소마의 팽창 또는 붓기를 동반합니다. 터치 뉴런의 경우, 사전 exopher 도메인 (PED)은 주변 피더니 (여기에 표시되지 않음)로 확장됩니다. 초기 싹 도메인에 mCherry 재료의 더 큰 농도를 참고하십시오. (C)늦은 싹 단계. 추가 세포 편광과 사전 exopher 도메인의 확장시, 소마와 exopher (화살표) 사이의 수축이 분명해진다. 이 이벤트는 후반 새싹 단계로의 전환을 나타냅니다. 늦은 싹 단계에서 세포는 명확한 수축 부위와 별도의 소마 및 엑소셔 도메인을 나타내지만 아직 소마에서 완전히 꼬집지 않습니다. 신진 엑소퍼는 두꺼운 줄기(화살표)에 의해 부착될 수 있다. 신진 도메인은 문제의 exopher 도메인의 직경이 건설 현장/줄기의 직경보다 약 1/3 더 큰 경우 초기 exopher로 간주됩니다. (D)조기 퇴출 단계. 초기 엑소퍼는 출발 소마에서 줄기에 의해 부착 될 수 있습니다 -이 연결의 직경은 엑소퍼가 소마에서 멀리 이동으로 얇을 수 있습니다. 세포질 물질은 대부분의 물질이 신진 하는 과정에서 로드 되지만,이 튜브를 통해 exopher에 소마에서 전송될 수 있습니다. 엑소퍼는(E)에묘사된 바와 같이 소마에서 분리할 수 있으며, 분리된 이포는 성숙한 엑소퍼(F)로 간주됩니다.F 성숙한 exopher는 떠나는 소마에서 멀리 이동, 주변 피하 조직을 통해 통과 할 수 있습니다. (G)mCherry 라벨이 부착된 exopher를 피성부 내에 작은 소포로 분해하면 mCherry 재료의 산발성 천막 모양이 발생하며, 이는 피하 내막증 네트워크에 들어갈 때 가장 가능성이 높습니다. 분산된 천점 신호는 "별이 빛나는 밤" 단계라고 합니다. 일부 exopher 내용의 저하는 피하 리소좀에 의해 달성 될 가능성이 있지만, 일부 물질은 완전히 저하되지 않고 종종 의사 에 피감에 의해 다시 압출된다. 사후 발생 mCherry 전송은 도 2에서더 자세히 설명되어 있습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

도 2: mCherry는 외향적인 피하 리소소말 네트워크에서 터치 뉴런으로부터 압출되었지만 나중에 coelomocytes가 mCherry를 저장/ 저하시킬 수 있는 의사 코일롬으로 압출될 수 있습니다. (A)만화 요약mCherry 는 뉴런에 의해 추방 후 몸을 유출 추방에 압출 하는 방법에 대한 요약. exophergenesis 동안 선택된 세포 내용물 인 mCherry는 뉴런 및 피하 혈장 막으로 둘러싸인 독립적 인 소포에서 보내는 신경 소마에서 국화되고 싹이 나아집니다. 터치 뉴런은 피하 조직에 내장되어 있기 때문에, exopher 도메인 싹바깥쪽으로 그것은 피하로 더 이동으로. exopher는 피증을 통과 할 수 있으며, 몇 시간에서 며칠 후에, exopher 내용체는 피하의 내분비 네트워크 내에서 단편화 할 수 있습니다. mCherry는 "별이 빛나는 밤"이라는 무대인 피더피 전체에 흩어져 있는 말장난으로 나타날 수 있습니다. 며칠 후, 일부 mCherry는 코로모세포라고 불리는 스캐빈저 세포가 접근할 수 있는 주변 의사콜로피에서 피하구를 통과할 수 있으며, 이를 수용할 수 있습니다. (B)별이 빛나는 밤 mCherry 소포의 출현의 예. 큰 exopher 조각과 별이 빛나는 밤 소포와 mCherry 태그 ALM 소마의 이미지. 스트레인은 ZB4065 bzIs166[Pmec-4mCherry]입니다. (C)먼 코로모세포에서 mCherry 농도의 예. 10일 종의 원소ZB4065 bzIs166[Pmec-4mCherry]의 사이드뷰는 코로모세포(화살표)에 집중된 mCherry를 보여준다. 일부 별이 빛나는 밤 소포도 분명합니다. 일반적으로 coelomocyte 농도는 성인 일 5 에 대 한 후 분명 해진다. (B하단) (B)의B만화 재생, 터치 뉴런과 빨간색으로 설명 된 프로세스, 밝은 exopher 조각으로; 다른 Z 깊이의 흩어져 작은 소포는 밝은 분홍색으로 표시됩니다. (C하단) (C)의이미지의 만화 버전은 녹색 분홍색과 coelomocytes에 빨간색, 별이 빛나는 밤에 신경 과정을 보여주는. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 3: 메카노 감각 터치 뉴런은 정확한 측두프로파일을 가진 다른 수준에서 발산을 생성합니다. (A)(상단) 동물의 머리에 펌핑 인두와 신경 조밀 한 신경 링을 포함하여 C. 엘레건의 주요 해부학 적 랜드 마크와 공간 관계에서 메카노 감각 터치 뉴런의 만화 묘사, 중간 몸의 외음부, 그리고 테이퍼 테일. (아래쪽) 위쪽과 왼쪽에서 볼 수 있듯이 GFP를 표현하는 형광 표시 터치 뉴런 (WormAtlas에서 적응 된 이미지). 빨간색 상자는 ALM 엑소퍼가 일반적으로 있는 영역을 묘사합니다. (B)ALM 유래 엑엑소퍼가 [Pmec-4mCherry]를 발현하는 균주에서 생산되는 중간 체지방의 높은 배율 보기. AVM과 ALMR 뉴런이 묘사되고, mCherry 별이 빛나는 밤과 함께 ALMR 엑소퍼가 표시됩니다. ALMR 뉴런은 가장 쉽게 exophers를 생산하고 있습니다. (C)ALMR 메카노감각 터치 뉴런은 기초 조건 하에서 헤르마프로디테이트에 있는 다른 터치 뉴런에 비해 더 쉽게 exophers를 생성한다. 개별 터치 수용체 뉴런에 대한 점수로, 성인 의 날에 Mechanosensory 터치 뉴런 exopher 생산 2, 표시. 스트레인: ZB4065 bzIs166[Pmec-4mCherry], N>150, 오류 막대는 SEM.(D)ALMR 터치 뉴런은 사춘기 L4 단계 또는 고령의 동물에 비해 성인기의 일 2 및 3 일 동안 더 많은 exophers를 생산하고 있습니다. 스트레인: ZB4065 bzIs166[Pmec-4mCherry], N>150, 오류 막대는 SEM입니다. Please click here to view a larger version of this figure.
{kind=link}

그림 4: 일부 형광 기자의 예로, 이글비글 내용의 태그를 붙인 다. exophers를 관찰 하는 간단한 방법은 신경 프로모터에서 형광을 표현 하는 형질 전환 동물을 만드는 것입니다. 형광은 외향성 및 형질 전환 식의 시각화를 허용하여 외신을 증가시키는 응집 및 / 또는 프로테오스트레스를 유도합니다. 증폭 된 뉴런에 의해 생성 된 Exophers는 또한 기본 조건하에서 관찰 될 수있다, 시각화를위한 염료 충전을 사용하여. 도시 된 exophers, (E) exopher, (S) 소마를 관찰 하는 데 사용할 수 있는 일반적인 균주의 예. (A)Soma 및 변형 SK4005 zdIs5[Pmec-4GFP],100x 목적사진, 스케일 바 3μm의 성인의 ALM에서 엑소퍼. 이 균주에서는 수용성 GFP를 포함하는 외래가 측정되지만 외래 생산은 드물게 발생합니다. GFP를 다른 연구에서 exophers에서 우선적으로 압출할 수 있는 단백질에 융합하면 GFP 융합이 성숙한 외신에서 검출될 수 있음을확인합니다. (B)변형 ZB4065 bzIs166[Pmec-4mCherry]의 성인의 ALM 소마와 엑소퍼, 이는 mCherry를 표현하고 터치 뉴런 exopher 생산을 유도. 사진, 스케일 바 5 μm에 사용되는 100배 목표. (C)변형 ZB4067 bzIs167[Pmec-4미토그fp Pmec-4mCherry4]의 성인의 ALM 소마 및 엑소셔; igIs1[Pmec-7YFP Pmec-3htt57Q128::cfp lin-15+]; htt57Q128::CFP 의 이미지에 사용되는 선택적 파란색 채널. exopher는 htt57Q128::CFP 집계(화살표)를 포함하며, 이는 소마보다 exopher에 더 집중되어 있는 것처럼 보입니다. 사진, 스케일 바 5μm에 사용되는 40배 목표. (D-E) Exophers는 세포기관을 포함할 수 있고 형광 단백질을 가진 세포기관 특정 태깅을 포함할 수 있어 세포외 압출을 모니터링할 수 있습니다. (D)리소소말 막 태그 LMP-1::GFP는 소마및 엑시퍼 멤브레인을 설명하고 플라즈마 멤브레인을 약하게 태그 (플라즈마 멤브레인 국소화는 리소소말 타겟팅하는 방법에 대한 인신 매매 단계입니다) 및 리소소말 세포기관을 강하게 라벨. 도시된 성인 ALM 소마는 Pmec-4mCherry와 멤브레인과 리소좀에 국한되는 Pmec-7LMP-1::GFP를 공동 발현한다. 소마는 외래 파편 (화살표)될 가능성이 다른 작은 압출과 부착 된 exopher가 있습니다. GFP 양성 구조는 소마에 포함되며 큰엑소퍼, 변형에 존재합니다: ZB4509 bzIs166[Pmec-4mCherry]; bzIs168[Pmec-7LMP-1:GFP]. 사진, 스케일 바 5 μm에 사용되는 100배 목표. E)미토콘드리아 GFP 마커는 소마및 엑소퍼에서 미토콘드리아를 식별하는 데 사용될 수 있다. 미토콘드리아 매트릭스에 국소화되는 Pmec-4mCherry 및 미토:ROGFP를 발현하는 성인 ALM 소마가 도시되어 있다. mito:ROGFP 혼자 표현, mCherry 없이, 또한 쉽게 뉴런을 식별 하 고 미토 콘 드리 아를 포함 하는 exophers에 대 한 점수를 사용할 수 있습니다. 변형: ZB4528 bzIs166[Pmec-4mCherry]; zhsEx17 [Pmec-4미톨::ROGFP]. 사진에 사용되는 100배 목표; 스케일 바 5μm. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 5: C. 예인대 및 L4 식별의 발달 주기. (A)20°C에서 알은 어머니가 한 번 부화하는 데 약 9시간이 걸립니다. (B)새로 부화한 동물은 12시간 후에 라발 1단계(L1)와 몰트에 L2 애벌레로 들어간다. (C)동물은 L2 및L3애벌레 단계에 각각 약 8시간 동안 남아 있다. (E)사춘기 동물은 네 번째 애벌레 단계 (L4)로 간주되며 중간 몸 근처의 흰색 초승달로 나타나는 눈에 띄는 개발 외음부에 의해 표시됩니다. 초승달 이 존재는 L4 단계의 동물을 쉽게 식별하고 따기 를 통해 나중에 exophers에 대한 점수를 용이하게하는 동기화 된 문화를 확립 할 수 있습니다. 동물은 알, 눈에 보이는 정자, 계란 누워의 개시에 의해 확인 된 중력 성인, F)에그들의 마지막 몰트 전에 약 10 시간 동안 L4 단계에 남아 있습니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 6: 현미경 슬라이드 한천 패드의 준비. (A)상단에 걸쳐 세로로 배치 된 실험실 테이프의 단일 스트립으로 두 개의 슬라이드를 준비합니다. 그림대로 테이프가 없는 현미경 슬라이드를 촬영할 사이에 놓습니다. B)용융 아가로즈 한 방울을 슬라이드 위에 놓습니다. (C)낙하 위에 깨끗한 슬라이드를 부드럽게 놓고 아가로즈를 수축 된 원형 패드로 누릅니다. (D)테이핑된 슬라이드를 제거하여 짝수 패드를 만드는 데 필요한 천의 균일한 평탄을 수행합니다. (E)아가로즈 패드가 건조되면 상단 슬라이드를 제거합니다. (F)파이펫은 한천 패드 위에 마비용 용액(레바미솔 또는 테트라미솔)을 한다. (G)적절하게 준비된 동물을 마비동물로 선택합니다. (H)동물을 커버슬립으로 부드럽게 덮고 동물이 살아 있는지 확인합니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}

그림 7: 엑소퍼및 엑소셔 식별 기준의 문자입니다. (A)엑소퍼를 식별하는 일반적인 기준. (b)μm으로 측정된 송신소와 압출된 외출자 사이의 직경 비교. 성인 ALM somas, N=35, 변형: ZB4065 bzIs166[Pmec-4mCherry] - 6.53 μm 평균 크기의 소마 와 3.83 μm 평균 크기의 엑셔. (C)엑소퍼 도메인과 신진 엑소퍼 사이의 차별화기준을 정의한다. (D)가장 일반적으로 개별 뉴런은 하나의 큰 외향성을 만들어 나중에 피더가 그 내용을 저하시키려고 시도함에 따라 갈라지거나 조각됩니다. 여전히, 여러 exophers 한 신경에서 여러 exopher 이벤트 또는 대안으로 파생 될 수 있는 한 터치 뉴런 옆에 관찰 될 수 있습니다., exophers 또한 싹 또는 조각 자체 수 있습니다. 여러 exopher 와 같은 개체의 기원은 시간 경과 현미경 검사를 사용하여 결정될 수 있습니다. 상단은 하나의 먼 exopher와 ALMR 터치 뉴런 소마를 묘사한다. 바닥은 여러 발췌와 같은 압출을 가진 ALMR 터치 뉴런 소마를 묘사합니다. (E)exopher 이벤트로 오인 될 수 있습니다 성인 ALM 터치 뉴런 소마에서 일반적인 형태학적 특징. 왼쪽 위 - 명확한 exopher 도메인 또는 수축 사이트가없는 DISDALM 소마. 상단 중간 - 뉴런은 exophers와 유사 할 수있는 작은 세포 외 돌출을 가질 수 있지만, exopher로 간주될 크기 요구 기준을 충족하지 않습니다. 오른쪽 상단 – 나이와 함께, 터치 뉴런은 그들의 사소한 neurite를 따라 성장을 개발할 수 있습니다. 종종 mCherry 재료는 중성염 아웃 성장의 끝에 수집 할 수 있습니다. 수집된 mCherry가 exopher-to-soma 크기 요구 사항을 충족하지 않는 경우 이 점은 exopher로 채점되지 않습니다. 바닥은 exopher 도메인 또는 exopher에 대 한 정의 기준을 가진 성인 ALM 뉴런을 묘사. Botom 왼쪽 - 선택적으로 mCherry 사이토솔과 mCherry 태그 집계를 포함하는 눈에 띄는 exopher 도메인을 가지고 ALM 소마. exopher 도메인 수축 사이트는 화살표로 표시되고 크기 기준(소마 의 크기 의 1/5분의 1 이상)을 충족합니다. exopher 도메인의 가장 큰 직경은 수축 사이트의 직경보다 거의 1/3 더 크며 exopher 이벤트에 대한 기준을 충족합니다. 하단 중간 - 크기 기준을 충족하는 눈에 띄는 신진 엑소퍼가 있는 ALM 소마. 명확한 수축 사이트가 있습니다. 오른쪽 아래 - EXopher 크기 요구 사항을 충족하는 mCherry 채워진 exopher가 부착 된 ALM 소마. 엑소퍼는 얇은 연결 필라멘트에 의해 부착됩니다. 모든 이미지는 변형 ZB4065 bzIs166[Pmec-4mCherry]에서 입니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
토론
큰 exophers의 형태로 골재 및 세포구 제거의 생체 내 분자 메커니즘의 특성은 초기 단계에 있다. 추방을 위한 화물의 지정, 셀 내의 이 화물의 편광 수집, 외출을 생성하는 결정의 조절, 압출을 중재하는 기계, 그리고 이웃 세포에서 분해 기계와의 외설의 상호 작용에 대한 질문은 모두 해결되어야 합니다. 더욱이 칼슘, 골재 및 미토콘드리아를 포함하는 생물학적 물질을 통과할 수 있는 관 연결의 생체 내 시각화는 흥미롭고 연구되지 않은 생물학입니다. 특정 세포가 다른 세포보다 생산을 추방하는 경향이 있는 이유에 대한 질문은 해결되지 않았지만 이 프로토콜에 설명된 접근법으로 유전적으로 해부되기 시작할 수 있습니다.
이 프로토콜에 자세히 설명된 것은 인근 세포 소마와 외래를 구별하는 데 주의를 기울이고, 엑시셔 생산의 피크를 포착하기 위한 분석 타이밍, 그리고 이포 수준을 조절할 수 있는 의도하지 않은 응력을 제거하기 위한 성장 조건의 엄격한 제어와 함께, exopher 생산의 재현 가능한 점수를 달성하는 접근법입니다. 큰 초기 exopher의 구별, 또는 주변 피더니에 "별이 빛나는 밤'분산은 exopher 생산의 증거로 수량 될 수있다. 즉, 기저 조건 하에서 mCherry를 표현하는 뉴런은 가장 자주 exopher를 생산하는 특정 유형의 뉴런의 5-25 %와 관련이 있습니다. 스트레스 조건의 통제된 소개는 압출을 생성하는 뉴런의 90%만큼 높은 검출에 exopher 생산을 증가시키기 위하여 적용될 수 있었습니다, modifior를 위한 유전 또는 약리학 스크린에 특히 유용.
인간 신경 퇴행성 질환에서, 큰 골재는 병리학 확산을 촉진하기 위하여 이웃 세포로 병들린 뉴런에서 전송할 수 있습니다. exopher 메커니즘은 필라 를 통해 집계 압출에 사용되는 보존 된 메커니즘을 통해 침투 할 수 있습니다. 이 과정의 효율성을 향상시키는 생체 내 분자를 정의 (더 효과적인 proteostasis 제어로 간주) 또는 여러 신경 퇴행성 질환을 퇴치하기위한 새로운 전략의 디자인에 영향을 미칠 수 있습니다 차단. 이와 같이, 여기에서 기술된 프로토콜은 고전적인 유전 돌연변이 스크린, 강화제 및 억제제를 확인하기 위하여 유전자를 체계적으로 노크하는 게놈 넓은 RNAi 스크린, 또는 이 과정의 후보 약리학 수정자를 확인하는 약 내정간섭 연구 결과를 위해 이용될 수 있었습니다. 접근 방식은 간단하지만 다소 힘들다. Exophers는 너무 커서 고배율 해부 현미경으로 볼 수 있습니다. 여전히, C. elegans 뉴런은 상대적으로 작고 그들의 세포기관 또는 그들의 막을 보고 더 높은 힘 공초점 심상을 필요로 하고 느린 프로세스입니다. 더 높은 처리량을 위한 선택권은 다중 잘 플레이트 형식으로 높은 콘텐츠 화상 진찰 접근을 관련시킬 수 있었습니다.
퇴출 점수에 표준화 된 접근 방식의 응용 프로그램은 뉴런이 구성하고 세포 이물질을 제거 할 수있는 과정의 공동 유전 해부를 기초해야한다.
공개
없음
감사의 말
R01AG047101 및 R37AG56510: 다음과 같은 NIH 보조금을 인정합니다. Driscoll 및 Grant 연구소의 구성원은 엄격한 실험과 강력한 의사 소통을 통해 설명된 프로토콜의 개발 및 미세 조정에 광범위하게 기여했습니다.
자료
| Name | Company | Catalog Number | Comments |
| 95B Scientific CMOS camera | Photometrics Prime | ||
| 1,000 μL low retention tips | Sarstedt | ||
| 10 mL serological pipette | Appleton Woods | CC214 | |
| 10 μL low retention tips | Sarstedt | 70.1130.105 | |
| 13% sodium hypochlorite | Acros Organics | AC219255000 | |
| 15 mL centrifuge tubes | Fisher Scientific | 05-539-12 | |
| 2 L erlenmeyer flasks | Scientific Laboratory Supplies | FLA4036 | |
| 25 mL serological pipette | Appleton Woods | CC216 | |
| 300 μL low retention tips | Sarstedt | 70.765.105 | |
| 50 mL serological pipette | Appleton Woods | CC117 | |
| 5-Fluoro-2'-deoxyuridine 98% | Alfa Aesar | L16497.ME | |
| 9 cm sterile Petri dishes | Fisher Scientific | 11309283 | |
| absolute ethanol | Vwr | 20821.33 | |
| Agar | Sigma Aldrich | A1296 | |
| C. elegans strain wild type | Supplied by CGC | N2 | C. elegans strain |
| calcium chloride dihydrate | Sigma Aldrich | C3881 | |
| cholesterol | Acros | 110190250 | |
| dibasic sodium phosphate | Sigma Aldrich | S3264 | |
| E. coli strain OP50 | Supplied by CGC | Op50 | E coli strain |
| FBS10 Standard microscope | Meyer Instruments | KSC 410-1-100-1 | FBS10 Standard with Plate Base, 100/100 Trinocular Head and Flip zoom |
| glass pipette 270 mm | Fisherbrand | FB50255 | |
| Heraeus Multifuge X3R | Thermofisher scientific | 75004515 | |
| Inoculating Spreaders | Fisher Scientific | 11821741 | |
| LB medium capsules | MP biomedicals | 3002-031 | |
| LDI – Laser Diode Illuminator | 89 North | ||
| levamisole | Sigma Aldrich | 16595-80-5 | |
| M4 multipette | Eppendorf | 4982000012 | |
| magnesium sulphate | Sigma Aldrich | M7506 | |
| monobasic potassium phosphate | Sigma Aldrich | P0662 | |
| Multitron Standard shaking incubator | Infors HT | INFO28573 | |
| Nalgene 1 L Centrifuge pots | Fisher Scientific | 3120-1000 | |
| P10 pipette | Eppendorf Research Plus | 3123000020 | |
| P1000 pipette | Eppendorf Research Plus | ||
| P200 pipette | Eppendorf Research Plus | 3123000055 | |
| pipeteboy 2 | VWR | 612-0927 | |
| Polystyrene microbeads | Sigma Aldrich | MFCD00131491 | |
| RC5C plus floor mounted centrifuge | Sorvall | 9900884 | |
| Reusable ringed cytology slides | ThermoFisher Scientific | 22037242 | |
| SK4005 zdIs5[Pmec-4GFP] | contract Driscoll lab | GFP expressed in touch neurons | |
| sodium chloride | Sigma Aldrich | 13422 | |
| Sodium hydroxide | Fisher Chemical | S/4880/53 | |
| Tactrol 2 Autoclave | Priorclave | ||
| Triton-X | Thermofisher scientific | 28313 | |
| Tween 20 | Sigma Aldrich | 9005-64-5 | |
| X-Light V2 Spinning Disk Confocal Unit | CrestOptics | ||
| ZB4065 bzIs166[Pmec-4mCherry] | contract Driscoll lab | mCherry expressed in touch neurons | |
| ZB4067 bzIs167[Pmec-4mitogfp Pmec-4mCherry4]; igIs1[Pmec-7YFP Pmec-3htt57Q128::cfp lin-15+] | contract Driscoll lab | Q128 expressed in touch neurons | |
| ZB4509 bzIs166[Pmec-4mCherry]; bzIs168[Pmec-7LMP-1::GFP] | contract Driscoll lab | mitoROGFP expressed in touch neurons | |
| ZB4528 bzIs166[Pmec-4mCherry]; zhsEx17 [Pmec-4mitoLS::ROGFP] | contract Driscoll lab | autophagy marker expressed in touch neurons | |
| ZEISS Axio Vert.A1 | Zeiss |
참고문헌
- Davis, A. A., Leyns, C. E. G., Holtzman, D. M. Intercellular Spread of Protein Aggregates in Neurodegenerative Disease. Annual Review of Cell and Developmental Biology. 34, 545-568 (2018).
- Davis, C. H., et al. Transcellular degradation of axonal mitochondria. Proceedings of the National Academy of Sciences of the United States of America. 111 (26), 9633-9638 (2014).
- Torralba, D., Baixauli, F., Sanchez-Madrid, F. Mitochondria Know No Boundaries: Mechanisms and Functions of Intercellular Mitochondrial Transfer. Frontiers in Cell and Developmental Biology. 4, 107 (2016).
- Stahl, P. D., Raposo, G. Extracellular Vesicles: Exosomes and Microvesicles Integrators of Homeostasis. Physiology (Bethesda, Md.). 34 (3), 169-177 (2019).
- Melentijevic, I., et al. C-elegans neurons jettison protein aggregates and mitochondria under neurotoxic stress. Nature. 542 (7641), 367 (2017).
- Nussbaum-Krammer, C. I., Park, K. W., Li, L., Melki, R., Morimoto, R. I. Spreading of a prion domain from cell-to-cell by vesicular transport in Caenorhabditis elegans. PLoS Genetics. 9 (3), 1003351 (2013).
- Tyson, T., et al. Novel animal model defines genetic contributions for neuron-to-neuron transfer of alpha-synuclein. Scientific Reports. 7, (2017).
- Babcock, D. T., Ganetzky, B. Transcellular spreading of huntingtin aggregates in the Drosophila brain. Proceedings of the National Academy of Sciences of the United States of America. 112 (39), 5427-5433 (2015).
- Pearce, M. M. P., Spartz, E. J., Hong, W., Luo, L., Kopito, R. R. Prion-like transmission of neuronal huntingtin aggregates to phagocytic glia in the Drosophila brain. Nature Communications. 6, 6768 (2015).
- Fu, H., Li, J., Du, P., Jin, W., Cui, D. Metabolic wastes are extracellularly disposed by excretosomes, nanotubes and exophers in mouse HT22 cells through an autophagic vesicle clustering mechanism. bioRxiv. 10 (1), (2019).
- Ghose, P., Park, E. C., Tabakin, A., Salazar-Vasquez, N., Rongo, C. Anoxia-reoxygenation regulates mitochondrial dynamics through the hypoxia response pathway, SKN-1/Nrf, and stomatin-like protein STL-1/SLP-2. PLoS Genetics. 9 (12), 1004063 (2013).
- Cannon, M. B., Remington, S. J. Redox-sensitive green fluorescent protein: probes for dynamic intracellular redox responses. A review. Methods in Molecular Biology. 476, 51-65 (2008).
- Perkins, L. A., Hedgecock, E. M., Thomson, J. N., Culotti, J. G. Mutant sensory cilia in the nematode Caenorhabditis elegans. Developmental Biology. 117 (2), 456-487 (1986).
- Stiernagle, T. Maintenance of C. elegans. WormBook: The Online Review of C. Elegans Biology. , 1-11 (2006).
- Sutphin, G. L., Kaeberlein, M. Measuring Caenorhabditis elegans life span on solid media. Journal of Visualized Experiments. (27), (2009).
- Mitchell, D. H., Stiles, J. W., Santelli, J., Sanadi, D. R. Synchronous growth and aging of Caenorhabditis elegans in the presence of fluorodeoxyuridine. Journal of Gerontology. 34 (1), 28-36 (1979).
- Weicksel, S. E., et al. A novel small molecule that disrupts a key event during the oocyte-to-embryo transition in C. elegans. Development. 143 (19), 3540-3548 (2016).
- Dong, L., et al. Reversible and long-term immobilization in a hydrogel-microbead matrix for high-resolution imaging of Caenorhabditis elegans and other small organisms. PloS One. 13 (3), 0193989 (2018).
- Toth, M. L., et al. Neurite sprouting and synapse deterioration in the aging Caenorhabditis elegans nervous system. Journal of Neuroscience. 32 (26), 8778-8790 (2012).
- Conte, D., MacNeil, L. T., Walhout, A. J. M., Mello, C. C. RNA Interference in Caenorhabditis elegans. Current Protocols in Molecular Biology. 109, (2015).
- Calixto, A., Chelur, D., Topalidou, I., Chen, X., Chalfie, M. Enhanced neuronal RNAi in C. elegans using SID-1. Nature Methods. 7 (7), 554-559 (2010).
- Maher, K. N., Catanese, M., Chase, D. L. Large-scale gene knockdown in C. elegans using dsRNA feeding libraries to generate robust loss-of-function phenotypes. Journal of Visualized Experiments. (79), e50693 (2013).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기더 많은 기사 탐색
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유