
Method Article
C. elegans'ta Büyük Ekofher Veziküllerde İnvivo Nöronal Agrega ve Organel Ekstrüzyon Puanlama da Kantitatif Yaklaşımlar
* Bu yazarlar eşit katkıda bulunmuştur
Bu Makalede
Özet
Bu protokol, C. elegans hücreleri tarafından membrana bağlı ekophers şeklinde üretilen büyük agrega ve/veya organel ekstrüzyonların (~4 μm) tespiti ve niceliği için yaklaşımları açıklamaktadır. Bu enkaz kovma mekanizmasının diseksiyonunu kolaylaştırmak için gerekli suşları, büyüme koşullarını, puanlama kriterlerini, zamanlamayı ve mikroskopi hususlarını tanımlıyoruz.
Özet
Yanlış katlanmış proteinlerin toksisitesi ve mitokondriyal disfonksiyon, türler arasında yaşa bağlı fonksiyonel nöronal gerileme ve nörodejeneratif hastalığı teşvik eden önemli faktörlerdir. Bu nörotoksik sorunlar uzun hücre-içsel olarak kabul edilmiş olmasına rağmen, önemli kanıtlar şimdi bir nöron kaynaklanan yanlış katlanmış insan hastalığı proteinleri komşu hücrelerde görünebilir destekler, insan nörodejeneratif hastalık patoloji yaymak teşvik etmek için önerilen bir fenomen.
C. elegans yetişkin nöronlar bu toplama proteinleri ifade büyük ekstrüzyon olabilir (~4 μm) membran çevrili veziküller toplu protein içerebilir, mitokondri, ve lizozomlar. Bu büyük veziküller "ekophers" olarak adlandırılır ve ekzozomlardan farklıdır (yaklaşık 100 x daha küçüktür ve farklı biyogeneze sahiptir). Eksophers hücresel enkaz dışarı atma temel oluşturan korunmuş bir mekanizma tarafından oluşabilir, ama daha önce tanınmayan, nöronal proteostaz ve mitokondriyal kalite kontrolü dalı, hangi agregalar insan nörodejeneratif hastalıklarda yayıldı süreçleri ile ilgili.
Ekoferler çoğunlukla dokunma nöronların içinde yüksek kopya transgenik mCherry ifade hayvanlarda incelenmiş olsa da, bu protokoller flüoresans etiketli organeller veya nöronların çeşitli sınıflarda ilgi diğer proteinler kullanarak ekofhergenez çalışmalarında eşit derecede yararlıdır.
Burada açıklanan C. elegans ekophers fiziksel özellikleri, bunların tespiti için stratejiler, tanımlama kriterleri, nicelik için optimal zamanlama ve dışa dönük üretim düzeylerini modüle edebilir stresler için kontrol hayvan büyüme protokolleri vardır. Birlikte, burada özetlenen protokollerin ayrıntıları laboratuvarlar arasında eksophers kantitatif analizi için bir standart oluşturmak için hizmet etmelidir. Bu belge, eksopherlerin üretildiği ve eksopherlerin komşu ve uzak hücreler tarafından tepkiverilen moleküler mekanizmaları ayrıntılı olarak ortaya koyan laboratuvarlar için sahada bir kaynak olarak hizmet vermeyi amaçlamaktadır.
Giriş
Agregalar ve disfonksiyonel mitokondri nörotoksik zorluklar uzun hücre içsel olarak kabul edilmiştir, ama daha yakın zamanda bir nöron kaynaklanan yanlış katlanmış insan hastalığı proteinleri de komşu hücrelere yayılabilir açık hale gelmiştir, patoloji teşvik1. Aynı şekilde, memeli mitokondri transsellüler bozunma 2 veya meydan2 komşu hücrelerde mitokondriyal popülasyonların kurtarılması için orijinal üretim hücre dışına gönderilebilir3. Çeşitli boyutlarda veziküller genellikle komşu hücrelere veya sıvıçevresinehücresel malzeme aktarmak için gözlenmiştir 4 . Bazı ekstrüde veziküller ortalama nöronal soma boyutuyaklaşım (ortalama dokunmatik nöron soma ~ 6 μm) ve büyük agregalar ve organeller barındırabilir.
Protein agregaları ve organeller taşıyabilir büyük vezikül ekstrüzyon çarpıcı bir örnek C. elegans dokunmatik reseptör nöronlar yüksek bir kopya numarası muhabiri bir zararlı toplama eğilimli kodlama oluşturmak ifade oluşur, bozulmaya dirençli mCherry5. Dokunma nöronlarından ekstrüzyonlar, eksoshers denilen ekstrüzyonlar, ~ 4 μm ortalama çapı, seçici mCherry veya diğer agregalar içerir, ve komşu hipodermis doğrudan teslim edilir, normalde dokunma reseptör nöronları çevreleyen. Hipodermis lizom bazlı bozulmayı dener, ancak mCherry agregaları gibi bazı sindirilemeyen içerikler hipodermis tarafından hayvanın sıvı dolu pseudocoelom'una yeniden ekstrüzyon yapılabilir, bu da mCherry'nin uzun süreli depolama için coelomocyte adı verilen uzak çöpçü hücreleri tarafından alınabilir (Şekil 1, Şekil 2)5.
Büyük ekstrüzyon ekzotek vezikülleri dokunma reseptör plazma zarı ile çevrili hücre bırakın ve toplu insan hastalığı proteinleri içerebilir, mitokondri, ve lizozomlar. Eksopher üretim süreci potansiyel olarak toksik türlerin sıralanması (örneğin bir toplama eğilimli ifade mCherry çözünür, çoğunlukla nöronal soma kalır GFP gibi zararsız proteinler ayrılmış) içeren gibi görünüyor. Bu şekilde, tehdit edici varlıkların yönlendirilmiş sınırdışı nöron tarafından gerçekleştirilir5. Bir proteostaz sorun, otofaji yıkılması tarafından indüklenen stres gibi, MG132 aracılı proteozom inhibisyonu, huntington hastalığı ile ilişkili genişletilmiş poliglutamin Q128 veya Alzheimer hastalığı ile ilgili parça Aβ1-42gibi insan hastalığı proteinlerinin transgenik ekspresyonu , exhers üreten nöronların sayısını artırabilir5.
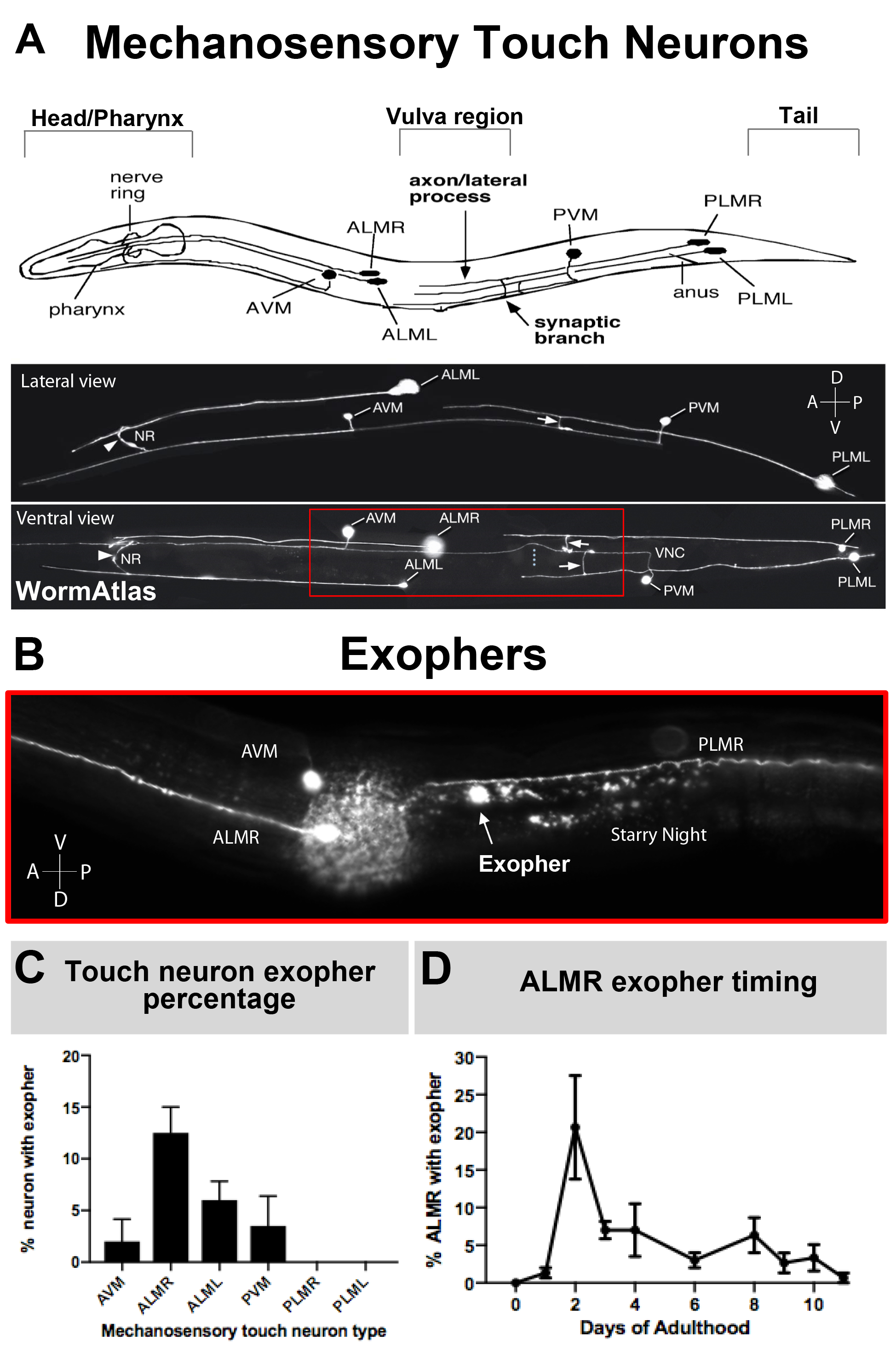
Exophers sadece son zamanlarda belgelenmiştir gibi, ne onların biyoloji yararları açıklama bilinmektedir. Eksophers keşfedildi, ve en iyi çalışılan, C. elegans dokunmatik reseptör nöronlar. Altı C. elegans mechanosensory touch nöronlar olan hücre organları vücuda dağılmış var(Şekil 3A) ve mikrotübül hücreleri denir çünkü ultrayapısı ayırt edici özellikleri 15 protofilament mikrotübüller. Dokunma reseptör nöronlar anterior AVM vardır (anterior ventral mikrotübün nöron), ALMR, ve ALML (ön lateral mikrotübün nöronlar sağ ve sol), daha merkezi PVM (posterior ventral mikrotübül nöron), ve posterior PLMR ve PLML (posterior lateral mikrotübül nöronlar sağ ve sol). İlginçtir, altı dokunmatik reseptör nöronlar farklı oranlarda ekofherler üretmek, aynı saldırgan transgen ifade rağmen (Şekil 3C). Altı mekanosensoriyel dokunma reseptör nöronlar, ALMR nöron diğer dokunma nöronlar daha sık eksozgenez uğrar. Dokunma nöronlarından eksopher numaralarının sayısallaştırılması böylece genellikle ALMR'ye odaklanılarak kurulur.
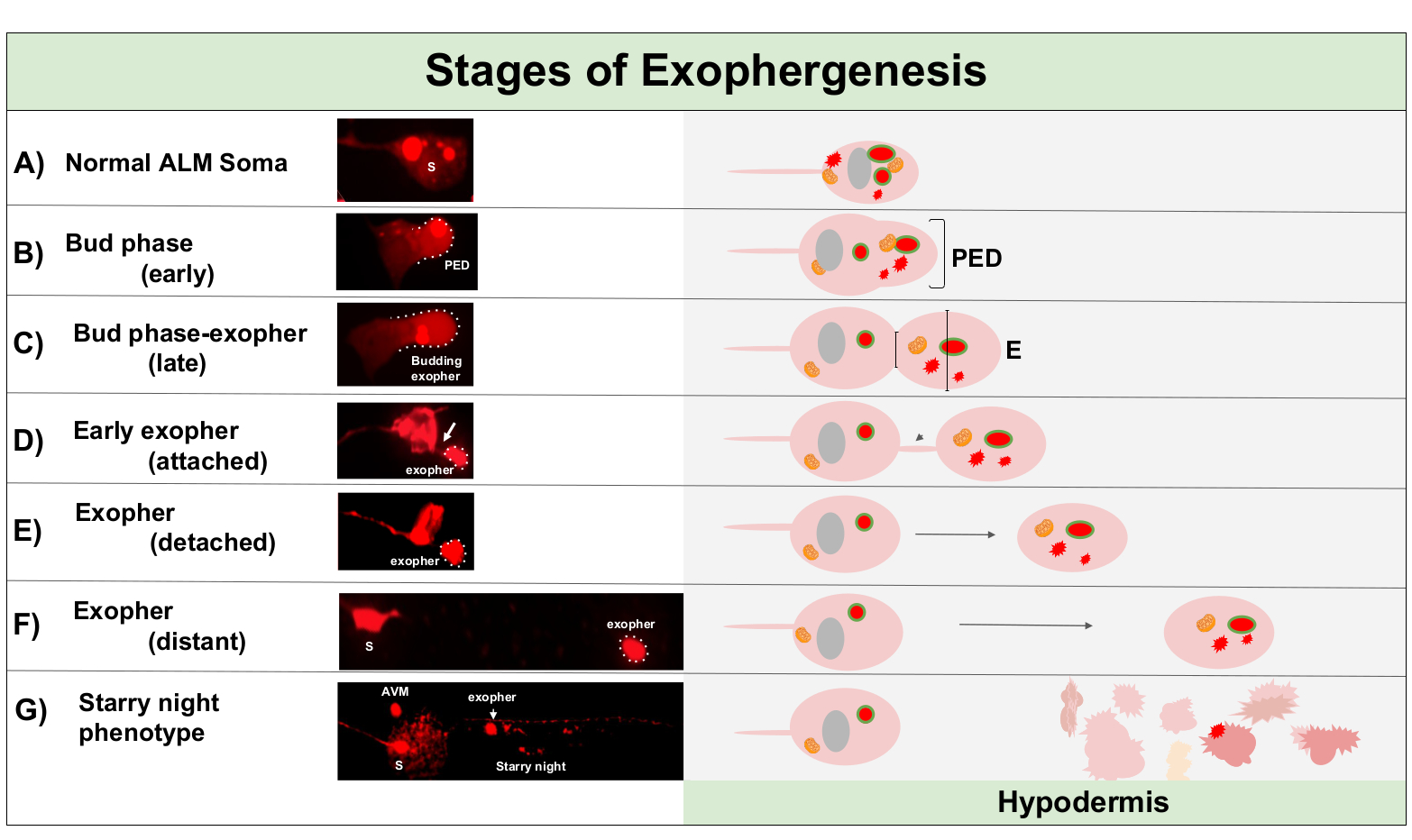
Ekofhernez genellikle nöronal sitoplazmanın şişmesi ile başlayan dinamik bir süreçtir (Şekil 1A-B). Hücresel içerikler, organeller veya protein agregaları nöronal somanın bir tarafına toplanır, en sık ALMR nöronun arka ucuna doğru (projektif netüritten uzakta), bir ön eksosetki alanı (PED) oluştururlar(Şekil 1B). PED dışa doğru yansıtmaya başladığında erken çıkıntı gözlenir ve tanınabilir bir çıkıntılı tomurcuk oluşturur. Geç tomurcuk, soma-ekopher boynunun daralması çapından yaklaşık 1/3 daha büyük olan pre-exopher etki alanının en geniş çapı nın yaklaşık olarak 1/3'ü olduğunda tanımlanır(Şekil 1C). Eksophers soma hemen hemen herhangi bir yönde atılabilir, ancak en ekofers hücre gövdesinden posteriora çıkmak ve kökeni soma olarak yaklaşık aynı odak düzleminde kalır.
Dış aparat, tomurcuk boyun ince bir filament içine daralır gibi gelen soma uzak hareket edebilir. Exophers bu filament(Şekil 1D, ok)ile soma bağlı kalabilir ve daha sonra kopuk olabilir. Kalsiyum gibi hücresel içerikler, agregalar, ve mitokondri ekli exopher içine bu filament yoluyla transfer edilebilir5, ekstrüde malzemenin toplu büyük tomurcuklanma olayı tarafından exopher bölmesi içine konulsa da. Ekophers görünür bir bağlantı tüpü veya ince filament olduğunda olgun olarak kabul edilir ve eksopher gönderme soma tamamen ayrılır(Şekil 1E).
C. elegans dokunma nöronları tarafından üretilen exophers hemen hipodermis karşılaşma, dokunma nöron çevreleyen doku. En yaygın olarak, eksopher vezikül hipodermis içinde arka arkaya kuyruk doğru seyahat görünür, ve dışa dönük içeriği bozulma için hedefli görünmeden önce soma oldukça uzak olabilir (örneğin, mesafe ~ 100 μm uzaklıkta soma olabilir(Şekil 1F)). Floresan eksopher vezikül hipodermis içinde çok daha küçük veziküller içine ayrılır, bir görünüm alarak "yıldızlı gece"(Şekil 1G ve Şekil 2). "Yıldızlı gece" aşamasında, punctate floresan malzeme orijinal yalnız exopher göre floresan birçok küçük noktalarıiçine hipodermal senkronium dağılmış görülebilir. Yıldızlı gece düşük büyütme altında delinmiş bakabilirsiniz ve yüksek büyütme ile, punctate ve / veya hipodermis içinde ağ bakabilirsiniz. Yıldızlı gecenin floresan sinyali genellikle eksopher ve nöronsal olarak ifade edilen floresandan daha sönüktür (Şekil 2B-C). Birçok punctate veziküller içine mCherry dağılımı hipodermal hücrenin endozomal / lysozomal ağ ile fagozom olgunlaşma ve füzyon içerdiği düşünülmektedir. Bazı ekopher malzemeleri muhtemelen hipodermal lisozomal ağda bozulur, ancak bozulmaya karşı dirençli artık türler (mCherry agregaları gibi) hipodermisten hücresel enkaz içerebilen bir sıvı bölmesi olan pseudocoelom'a atılır. Floresan malzeme daha sonra coelomocytes denilen uzak çöpçü hücreleri tarafından alınır(Şekil 2C),hangi konsantre olabilir, depolamak, ve tekrar mCherry bozulmagirişimi.
Agrega ekstrüzyon ve transfer olgusu phyla boyunca korunmuş olarak görünür, C. elegans5gibi genetik modellerde bildirilmiştir 5,6,7 ve D. melanogaster8,9 yanı sıra birden fazla memeli modelleri. Ekopher benzeri ekstrüzyonlar memeli hücreleri için bildirilmiştir10, korunmuş mekanizmalar agrega ve organel ihracı altında yatan olabileceğini düşündüren bir gözlem. Exopher üretimi böylece temel bir hücresel enkaz yönetimi korunmuş bir mekanizma olabilir, ama daha önce tanınmayan, nöronal proteostaz ve mitokondriyal kalite kontrolü dalı, hangi, dengesiz, aktif nörodejeneratif hastalığa katkıda bulunabilir. Enkaz ayrımcılığı ve ayrıştırma ile ilgili moleküllerin tanımlanması, ayrı bir hücre altı lokaline taşınması, ekstrüzyon, soma ve geç eksopher'i birbirine bağlayan borubağlantısının oluşumu/makaslanması ve komşu hücre tarafından uzaktan bozulması için büyük ekstrüzyon vezikülünün tanınması gelecekteki çalışmalar için kalır. Nematod ve sinek modellerinde yapılan çalışmalar, agrega ve organel toplama ve aktarma mekanizmalarının tanımlanması, bu modellerin katılımcı molekülleri fizyolojik bağlamda tanımlamak için sunduğu tarafsız genetik yaklaşımlar ve güçlü hücre biyolojik araçlarının kullanılması açısından kritik öneme sahiptir.
Eksopher biyolojisinde etkili mekanizmaların deşifre edilmesinde kritik ilk adımlar, in vivo ekopher nicelleştirmede tekrarlanabilir protokollerin tanımlanmasını içerir. C. elegans modeli, vücut şeffaf olduğundan ve floresan etiketli proteinler veya organeller içerdiklerinde kolayca gözlemlenebildiği için bu tür çabalar için özel bir avantaj sunar. Ekophers C. elegans dopaminerjik nöronlar PDE ve CEP, ASE ve ASER duyusal nöronlar tarafından oluşturulan bildirilmiştir, ve boya dolum amphid nöronlar5. Dokunma reseptör nöronları tarafından üretilen ekophers en iyi karakterize olduğundan, burada odak ekofher analizi için dokunmatik nöronların kullanımı üzerindedir. Ancak herhangi bir hücreden eksopher üretimini ölçmek için temel yaklaşım uygulanabilir. C. elegans tarafından üretilen ekofherleri algılamak ve quantitate etmek için protokoller transgenically mCherry protein ifade reseptör nöronlar dokunun, takip edilebilir yükler ve puanlama zamansal kısıtlamalar üzerinde durularak. Bu makalede, in vivo ekopher tanımlamaya yönelik yaklaşımlar ve eksopher üretimini modüle eden çevresel ve genetik koşulların niceliği tanımlanabılmaktadır. Protokoller, temel eksopher üretiminin belirlenmesi ve genotipler arasında karşılaştırmalar için sürekli gerilim dışı koşullara eleştirel dikkat ilerler.
Protokol
1. Eksopher tespiti için yararlı suşları
- Eksophers kolayca görselleştirmek için C. elegans nöronlar içinde floresan yükleri ifade eden bir suş seçin.
NOT: Tablo 1, dokunma reseptör nöronlarında üretilen eksopherleri görselleştirmek için kullanılan suşları listeler5,11,12. Prensip olarak, herhangi bir hücre veya nöronal tip agregalar veya aksi ekstrüzyon için seçilen bir floresan protein ekspresyonu sürücü için bir hücre veya doku özel organizatörü kullanılarak eksopher üretimi için test edilebilir. - Alternatif olarak, çevreye açık ve5,,13dolum için uygun amphid baş nöronlar, exophers görselleştirmek için bir boya dolum tsay kullanın.
2. Büyüme ortamı
- Standart yöntemlere göre kültür suşları için standart nematod büyüme medya (NGM) hazırlamak14,15.
NOT: Gıda eksikliği, veya floro-deoksitürin (FuDR), yaygın olarak soyundan üretimini engellemek için kullanılan ve önemli ölçüde eksopher üretimini etkileyebilir. Nüfusu sürekli beslenin (bakteriyel gıda yorgunluğunun kısa sürelerini bile önlemek) ve hayvanları sabit bir sıcaklıkta tutun.
3. Hayvancılık tutarlı eksopher üretimi için kritik
- Tutarlı medya ve tutarlı bakteriyel gıda kaynakları ile hayvanları yükseltmek. Gıda sınırlaması eksopher üretim düzeylerini önemli ölçüde değiştirebileceğinden, hayvanların bakteriyel gıdaları tükenmemelidir.
- Bir çalışma boyunca medya tarifleri ve hazırlık üniforma tutun.
NOT: Ortam değiştirme eksopher üretiminin bazal düzeylerini etkileyebilir. Agar toplu taban çizgisi ekopher düzeylerini etkileyebilir, bu nedenle tedarik çok değiştiğinde, tarih bir not yapmak. Sağlıklı bakteriyel gıda sağlamak ve kurutulmuş agar önlemek için iki hafta sonra stok plakaları atmak, hangi eksopher düzeylerini etkileyen agar ozmolarity değişikliklere neden olur. - Bazal koşullar için hayvanları 20 °C sabit sıcaklıkta tutun. Hayvanların değişken sıcaklıklarda yetiştirilmesi (sıcaklıktaki geçici değişiklikler bile) maksimal eksopher üretiminin zamanlamasında farklılıklara neden olabilir.
NOT: Sıcaklık değişkenliği kültür koşulları ile sınırlı değildir. Deneyler sırasında veya laboratuvar tezgahında sıcaklık değişimleri etkili olabilir. Örneğin, bir mikroskop odası içindeki sıcaklıklar kültür kuluçka makinesi veya laboratuvar tezgahından önemli ölçüde farklı olmamalıdır. - Döllenmiş yumurta eksophers erken yetişkin üretimi için kritik olduğu için farmakolojik anti-doğurganlık müdahalelerkullanmayın.
NOT: Floro-deoksiürin (FuDR)16 veya C2217kullanımından kaçınılmalıdır. Yaşam süresi veya yaşlı hayvan deneyleri yaparken, yaş senkronize popülasyonlar, yetişkinleri yaygın farmakolojik anti-doğurganlık müdahaleleri yerine bakterilerle yayılan taze plakalara toplayarak fiziksel olarak küçük döllerinden çıkararak muhafaza edilmelidir. - Kirlenmiş kültürler kullanmayın; popülasyonun veya plakanın biyolojik olarak tehlikeye atılsa lar deneyleri yeniden başlatın. Bakteriyel veya mantar kontaminasyonu hayvanlarda streslere ve metabolik değişikliklere neden olabilir ve deneysel popülasyonlardan yoksun olmalıdır.
- Tekrarlanabilir sonuçları en üst düzeye çıkarmak için, potansiyel çevrekaynaklı epigenetik değişiklikleri önlemek için deneylerden önce 20 °C'de en az iki sağlıklı, iyi beslenmiş, kirlenmeye nisbeten uzak nesiller için kültürleri koruyun.
4. Ağartma, sakkaroz yüzdürme veya L4 larva toplama ile eksopher skorlama için yaş senkronizasyonu
- Exopher algılama desenleri yetişkin yaşlarına göre değiştiğinden ve karışık yaş popülasyonlarına sahip hayvanların karşılaştırılması sonuçları karıştırabileceğinden, deneysel popülasyonları aynı biyolojik yaşta tutun. L4 aşamasındaki "beyaz hilal" vulva morfolojisini kontrol ederek deneysel hayvan popülasyonlarının başarılı bir şekilde senkronize edilmesini sağlayın.
NOT: Genellikle, C. elegans mechanosensory ALMR nöronlar için pik eksopher üretimi yetişkin gün 2-3 oluşur(Şekil 3D), gün sonra L4 aşamada ölçülen. Yetişkin gün 1 "beyaz hilal" vulva morfolojisi(Şekil 5E)ile ayırt edilir L4 larva aşamasından sonra 24 saattir. - Gravid yetişkinlerağdır tarafından senkronize yumurta popülasyonları hazırlayın.
- Bir NGM plaka üzerinde büyüyen hayvanları yıkayarak yumurta dolu gravid yetişkin toplamak. Yıkamak için, 1 mL M9 tampon ile plaka sel, bir 1.5 mL mikrosantrifüj tüp içine askılı hayvanlar ve pipet ile sıvı toplamak için yukarı ve aşağı pipet. Pellet hayvanlar bir mini santrifüj ile yerçekimi yerleşme veya nazik santrifüj ve supernatant kaldırın.
- 1 mL'de H2O'ya 150°L 5M NaOH ve %6 sodyum hipoklorit (çamaşır suyu) ekleyin ve yaklaşık 5 dakika ters çevrilerek karıştırın.
NOT: Taze beyazlatma solüsyonu, hayvan mığalının yumurta hasadı için bozulmasını sağlar. Miteklik bozma ilerleme bir diseksiyon mikroskop altında izlenebilir; yetişkinler beyazlatmanın durdurulması gerektiği noktasında yumurtaları kırıp serbest bırakmalıdır. - Yavaşça 20 s için bir minisantrifüj tüp ile santrifüj ve supernatant çıkarın. Peletin üzerine yaklaşık 100°L bırakarak 1 mL M9 tampon ve santrifüj ekleyin.
- Çamaşır suyu çözeltisinin izlerini gidermek için 4.2.3 adımlarını iki kez tekrarlayın.
- Kalan hacimdeki yumurtaları yeniden askıya alın ve taze tohumlu NGM plakasına aktarın. Yetişkinler lysed olacak, ancak birçok canlı yumurta hazırlık olmalıdır.
- Zamanlanmış yumurta yumurtlama ile senkronize popülasyonları hazırlayın.
- Standart transfer protokolleri14kullanarak bir tohumlu NGM plaka için 20 gravid yetişkin seçin.
- Hayvanların serbestçe sürünmesine ve 1,5 saat yumurtlamasını bekleyin (düşük kuluçka boyutlarına sahip mutant suşları daha fazla yetişkin hayvanın piyasaya sürülmesini gerektirebilir).
- Senkronize yumurta popülasyonu geride bırakarak, toplayarak plaka tüm yetişkin hayvanlar çıkarın. Yetişkinlerin yetişkin temizliği sırasında hiçbir yetişkinin kaçırılmadığını doğrulamak için birkaç saat sonra plakaları kontrol edin.
- Yumurtasakaroz yüzdürme seçimi ile senkronize yumurta popülasyonları hazırlayın.
- Gravid hayvanların en az 24 saat boyunca yumurtladıkları beş NGM plakasından hayvan ve yumurtaları toplayarak plakaları %0,1 deterjanla (Tween 20 veya Triton X-100 gibi) M9 çözeltisi ile su basarak ve 15 mL'lik bir tüpe toplayarak toplayın. Oda sıcaklığında hafif santrifüj ile pelet yetişkinler (30 s için 2.000 x g).
- Supernatant çıkarın ve taze M9 üç kez 15 mL hayvanları yıkayın, her yıkamadan sonra supernatant atarak, hayvanlar ve yumurta zenginleştirilmiş pelet tutmak için emin olmak.
- 2 mL supernatant'ı koruyun ve peleti yeniden askıya alın. Hacim sakaroza göre %60 ağırlıkta 2 mL ekleyin.
- Santrifüj 2000 x g 5 dk. Çözüm şimdi yumurta ile yüksek oranda zenginleştirilmiş bir üst faz ı sergileyecek.
- Üst fazın yaklaşık 2,5 mL'sini yeni bir 15 mL tüpe aktarın ve 10 mL M9 ekleyin.
- 1 dk için inversion tarafından karıştırın ve sonra 1 dk için 2000 x g santrifüj.
- Supernatant çıkarın ve M9 yumurta ile zenginleştirilmiş pelet yıkayın. Yumurta peletinin 10-15 μL'si taze OP50 tohumlu NGM plakasına dağıtılabilir.
NOT: Bu yöntem çok sayıda yumurta hazırlar; toplanan hayvanların OP50 E. coli gıdatü tükenmesine izin vermez.
- Gelişimin L4 aşamasında hayvanları seçerek senkronize popülasyonları hazırlayın.
- Yukarıda açıklandığı gibi tohumlu NGM plakaları üzerinde hayvan yetiştirmek.
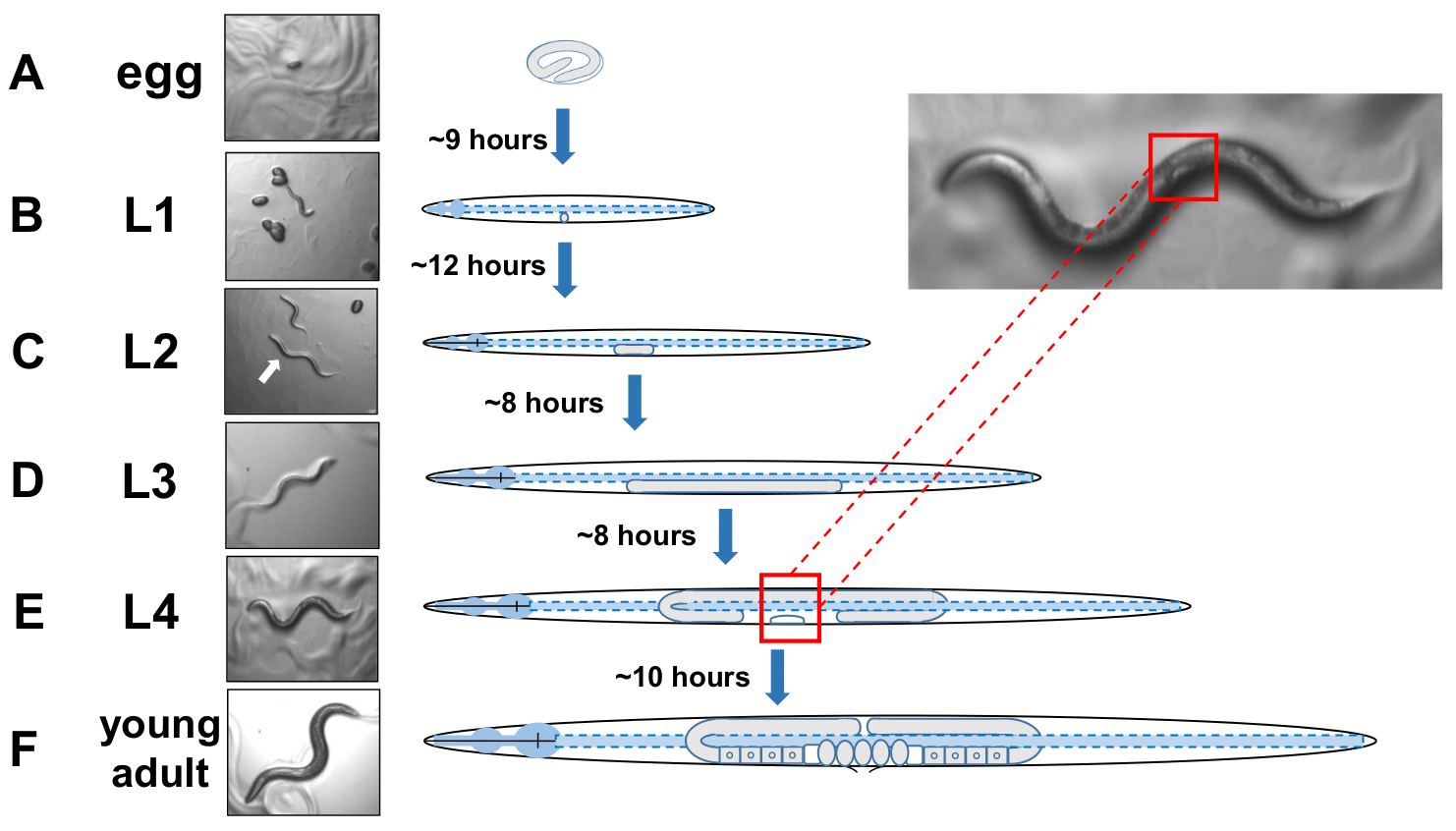
NOT: C. eleganlar dört ayrı aşamada gelişir. 20 °C'de, yeni döşenmiş bir yumurtanın yumurtadan çıkması yaklaşık 9 saat sürer (Şekil 5A). Post hatch, bir hayvan larva evre 1 geçer (L1) larva evre 4 (L4), her aşamada her molt arasında süren 8-12 saat(Şekil 5A-F). Bu nedenle, yumurta ile aşılanarak hazırlanan bir plaka yumurta tanıtıldıktan yaklaşık 40 saat sonra almak için birçok L4 hayvan olmalıdır. - Gelişmekte olan vulvanın beyaz yarım ay hilal şeklini bularak L4 aşamalı hayvanları tanımlayın(Şekil 5E).
NOT: L4 evresindeki hayvanlar boyut ve vücut pigmentasyonu nda tek tiptirler. Sahnelenen hayvanların incelenmesi için taze bir büyüme plaka beyaz hilal ile hayvanları seçin. Ertesi gün (~ 24 saat sonra) yetişkin gün 1 olarak sayılmalıdır. - Yetişkin 2 gününde günlük hayvan popülasyonu puan.
NOT: Ekoferler genellikle yetişkinliğin 2. Ancak, ekofhergenez tepe ve zamanlama çalışılmaktadır çevresel veya genetik değişiklikler tarafından kayması olabilir, çünkü en kapsamlı resim oluşturmak için dört gün boyunca yetişkin hayvanların bir nüfus puan tavsiye edilir(Şekil 3D).
- Yukarıda açıklandığı gibi tohumlu NGM plakaları üzerinde hayvan yetiştirmek.
5. Floresan mikroskop kullanarak ekoferlerin tespiti
- Floresan mikroskop için donatılmış yüksek büyütmeli sözde stereo diseksiyon mikroskobu kullanarak ekopherleri gözlemleyin.
- 100-200 μL 100-100 mM levamisole/tetramisole çözeltisini NGM agar plaka yüzeyine boru ile NGM plakaları üzerinde hareketsiz hale getirin. 2-4 dakika sonra hayvanlar felç olur ve doğrudan agar plakası üzerinde gözlemlenebilir.
NOT: İmmobilizasyon tedavileri kesinlikle gerekli değildir, eğitimli bir göz ile, nöronal tanımlama ve eksopher varlığı bir eksopher üretilip üretilmediğini belirlerken plaka üzerinde mikroskop altında sürünen hayvanları görsel olarak takip ederek puanlandırılabilir. - Eksopherlerin mikroskop tespitini gerçekleştirmek için toplam 100 x büyütme kullanarak floresan nöronları gözlemleyin.
NOT: Diseksiyon mikroskobu kullanılarak eksopher olaylarıpuanlama doğrudan yetiştirilen agar plakaları üzerinde göreceli kolaylığı ile hayvanların çok sayıda gözlem sağlar. - Konfokal mikroskopi ile eksopher çalışmaları için canlı görüntüleme ve montaj muhabiri suşları
- Hücre içi dinamikleri ve ekofhergenezözellikleri için bir konfokal mikroskop kullanın.
NOT: Canlı görüntüleme, eksopher üretiminin ince ayrıntılarını gözlemlemek için avantajlı bir yaklaşımdır, çünkü eksopher üretimi dinamik bir süreçtir. - Levamisole veya tetramisole 10-100 mM'de kullanımı veya hidrojel polistiren mikroboncukların uygulanması (çapları 15 μm, 30 μm veya 40 m)18dahil olmak üzere uygun yöntemlerle yüksek çözünürlüklü canlı görüntüleme için hayvan hareketini kısıtlayın.
- Hücre içi dinamikleri ve ekofhergenezözellikleri için bir konfokal mikroskop kullanın.
- Bileşik ve konfokal mikroskopi için slayt hazırlığı
- 20-50 hayvanı mikroskop kaydırağında hareketsiz bir maddeye monte edin. 13 mm çapında yükseltilmiş halkalarla boyanmış yeniden kullanılabilir halkalı sitoloji slaytlar montaj için yararlıdır.
- Boyalı daire içinde veya agar pedüzerinde 10-100 mM levamisole veya tetramisole gibi bir paralitik 5-20 μL içine canlı hayvanları seçin.
- Felç için 4 dakika bekleyin ve ardından kaydırağı coverslip (Tavsiye No. 16 (0,16 – 0,19 mm) veya No. 2 (0,17 - 0,25 mm) ile kapatın.
- Az sayıda hayvan ın montajı
- Atlı hayvanları ezme; slayt başına sadece birkaç (20'den az) hayvan gözlemleyerek, örtü nün eşit olmayan basıncı nedeniyle bazı hayvanların ezilmesi riski vardır. Bu risk montaj için düşük yüzdeli agarose ped kullanılarak en aza indirilebilir.
- %2-4 agarose pad slayt yapın ve sonra pede 2-15 μL paralitik çözelti ekleyin. Levamisole ve tetramisole'un etkin konsantrasyonlarını azaltarak pedin içine yayınıl olduğunu unutmayın.
- Agar pedüzerinde dinlenen 2-15 μL'lik felçli çözelti veya mikroboncuklar içine hayvanları toplayarak monte edin. Üzerine kapak yerleştirin ve hayvanların bozulmamış olup olmadığını kontrol edin18.
- Agar ped hazırlama
- % 2 agar pedleri hazırlamak için, ısı 2% agarose M9 çözeltisi ve mikrodalga agarose homojen ve erimiş durumda olana kadar.
- 20 saniyeden daha az düşük güçte yeterli kalitede, alternatif karıştırma ve mikrowaving bir agar ped elde etmek için. Bir ısıtma bloğu üzerinde kaynayan agar yerleştirerek ve kabarcıklar yüzeye yükselmeye izin vererek yastık içinde hava kabarcıkları dahil kaçının.
- Yükselen kabarcıklar altında erimiş çözelti içinde derin agar çizmek için bir Pasteur pipet kullanın.
- İki bantlı slayt hazırlayın ve düz bir yüzeye temiz bir cam mikroskop kaydırağının her iki tarafına yerleştirin. Bantlı slaytlar yapmak için her slayt(Şekil 6A)üzerine laboratuvar bandı iki 5 cm şeritler yerleştirin.
- Pasteur pipet kullanarak, bantlı slaytlar(Şekil 6B)arasına sıkışmış temiz mikroskop slayt üzerine tek bir agar damlası yerleştirin.
- Dikkatlice ve hızlı bir şekilde, bantlanmış slaytlar(Şekil 6Cc)arasında yerleştirerek dördüncü bir temiz slayt ile erimiş agar damla kapağı.
NOT: Slayt, erimiş agarı yaklaşık 0,4 mm kalınlığında (bant kalınlığı) düzleştirilmiş bir daireiçine hafifçe bastırmalıdır (Şekil 6D). Agar çabuk soğumalı. - Üst teki kaydırağı kaydırarak çıkarın (Şekil 6E). Agar pedleri hızlı bir şekilde kurur ve en iyi birkaç dakika içinde kullanılır. Üst slayt kaldırıldıktan sonra, montaj hayvanlar için hemen jel ped kullanın. Hava kabarcıkları ile pedleri kullanmaktan kaçının.
- İki cam kaydırak arasında 30 dakikaya kadar agar pedleri saklayın. Kurutulmuş agar hayvanların bir araya gelip kurutulmasıyla sonuçlanmaz. 2-15 μL'lik paralitik solüsyon veya mikroboncuklar içinde yer alan hayvanlara monte edin ve kapak lı örtüler; slama ve montaj 20 dakika içinde slayt ekran(Şekil 6).
NOT: Stres koşulları eksopher oranlarını değiştirebildiği için, eksophers için tarama yaparken oksidatif stres (ör. sodyum azit) neden olabilir paralitikler kaçının.
- İplik-disk konfokal mikroskobu kullanılarak ekoferlerin tespiti
- 63x ve 100x'te 1,4 sayısal diyafram hedefiyle organeller ve diğer içerikler gibi hücre biyolojik özelliklerini gözlemleyin.
- Çok boyutlu edinimi kullanarak sahne kontrolü ve görüntü edinimi yeteneğine sahip yazılım kullanın. Mikroskoplar ve görüntü işleme yazılımları da standart görüntüleme yaklaşımlarını içerdiğiiçin görüntüleme ve veri toplama için uygun olmalıdır.
6. Dokunma nöronların belirlenmesi ve atlı hayvanlarla ekoferler için puanlama
- Mount felçli yetişkin hayvan (Şekil 6).
- İstenilen Z-düzlemini tanımlayın. Hayvanın uygun Z düzlemini tanımlamak için düşük büyütme parlak alanını (10-40x) kullanın, hayvanın konumlandırılmasına dikkat çekerek, baş-kuyruk oryantasyonu ve vulvanın yeri - daha sonra nöronal ve ekopher tanımlamaiçin önemli yapılardır(Şekil 3A & Şekil 5E).
- Seçilen muhabirin floresan sinyaline odaklanın. Aynı Z-düzlemde kalarak, seçilen sitosolik muhabir için 10-40x geniş alan floresan görüntüleme geçiş.
NOT: Bu örnekte floresan ekspresyon mec-4 mekanosensoriyel dokunma nöron özgü organizatörü tarafından tahrik edilir. Yüksek kopya dizileri ve farklı floresan ifadeler değişkenlik ve bu nedenle değişken floresan yoğunluğu var. Gerekirse ayarlayın. - Odak düzlemindeki hayvan Ve floresan ifadenin derinliğini gözlemlemek için Z ekseni içinde ilerleyin. Bunu yaparken, baş-kuyruk yönünü onaylamak; baş/ farinks floresan sinir halkası olacak ve bu durumda, kuyruk 1-2 görünür PLM somas içerecektir(Şekil 3A).
- Dokunma nöronları belirleme
- Hayvanın sola mı yoksa sağ tarafa mı monte edilmediğini belirleyin(Şekil 3A).
NOT: Hayvanın 3 boyutluluk oranı göz önüne alındığında, en iyi görüntüleme çözünürlüğü optik lere en yakın tarafta gerçekleştirilir. - Gözlemleyerek soma (ALM, ALMR, AVM) tanımlayın - sinir halkası ve lateral nöronal süreçleri tanımlamak için baş başlar.
- 10-40x büyütme de, bağlı işlemi tanımlamak için z ekseni arasında yavaşça ilerleyin.
- İşlem tanımlandıktan sonra, soma görünür olacak vulva doğru arka yönde yanal izleyin, sürecin sonunda yuvarlak bir hücre gövdesi ile işaretlenmiş. Bir kez en odak nöronal soma bulunur, aşağıdaki gibi diğer nöronal yerler kullanılarak tespit edilebilir:
- AvM kullanın, yakındaki bir ventral nöron, hayvan oryantasyon atamaya yardımcı olmak için. AVM nöron ALM ile aynı düzlemde ise o zaman hayvan yan ve bu uçağın dışında nöron üzerine dinleniyor ALMR olduğunu . AVM nöron söz konusu ALM ile aynı düzlemde değilse, odak düzlemine en yakın dokunmatik nöron ALML'dir.
- Ön dokunma nöronunaynı düzlemde olup olmadığını belirtmek için, kuyruk yakınında bulunan başka bir ventral dokunma nöron, PVM nöron tanımlayın. Eğer öyleyse, gözlenen dokunma nöron ALML olduğunu.
- Diğer soma organlarının konumu bir fikir alın, ilgi alanı yakınında (soma her iki tarafında bulunan floresan nöronlar), ve tüm Z-uçaklarda, net odak koymak derin nöron almak mümkün olmasa bile.
NOT: Tüm dokunma nöron somalarının tanımlanması önemlidir, çünkü odak dışı soma ekofherlerle karıştırılabilir.
- Hayvanın sola mı yoksa sağ tarafa mı monte edilmediğini belirleyin(Şekil 3A).
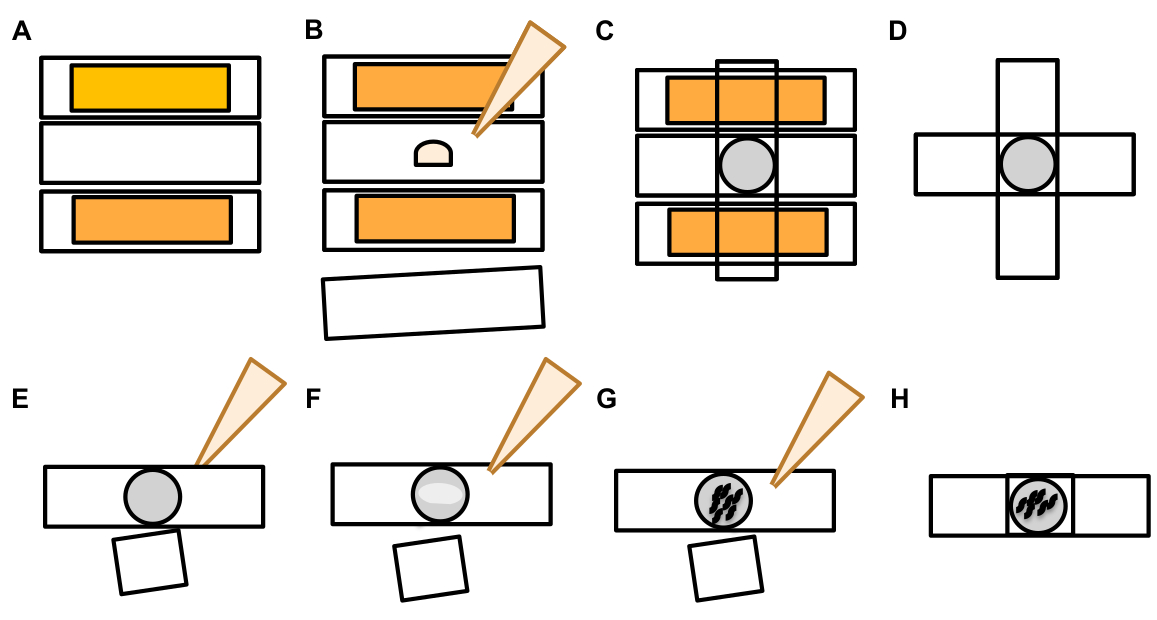
7. Eksophers için belirlenmesi ve puanlama
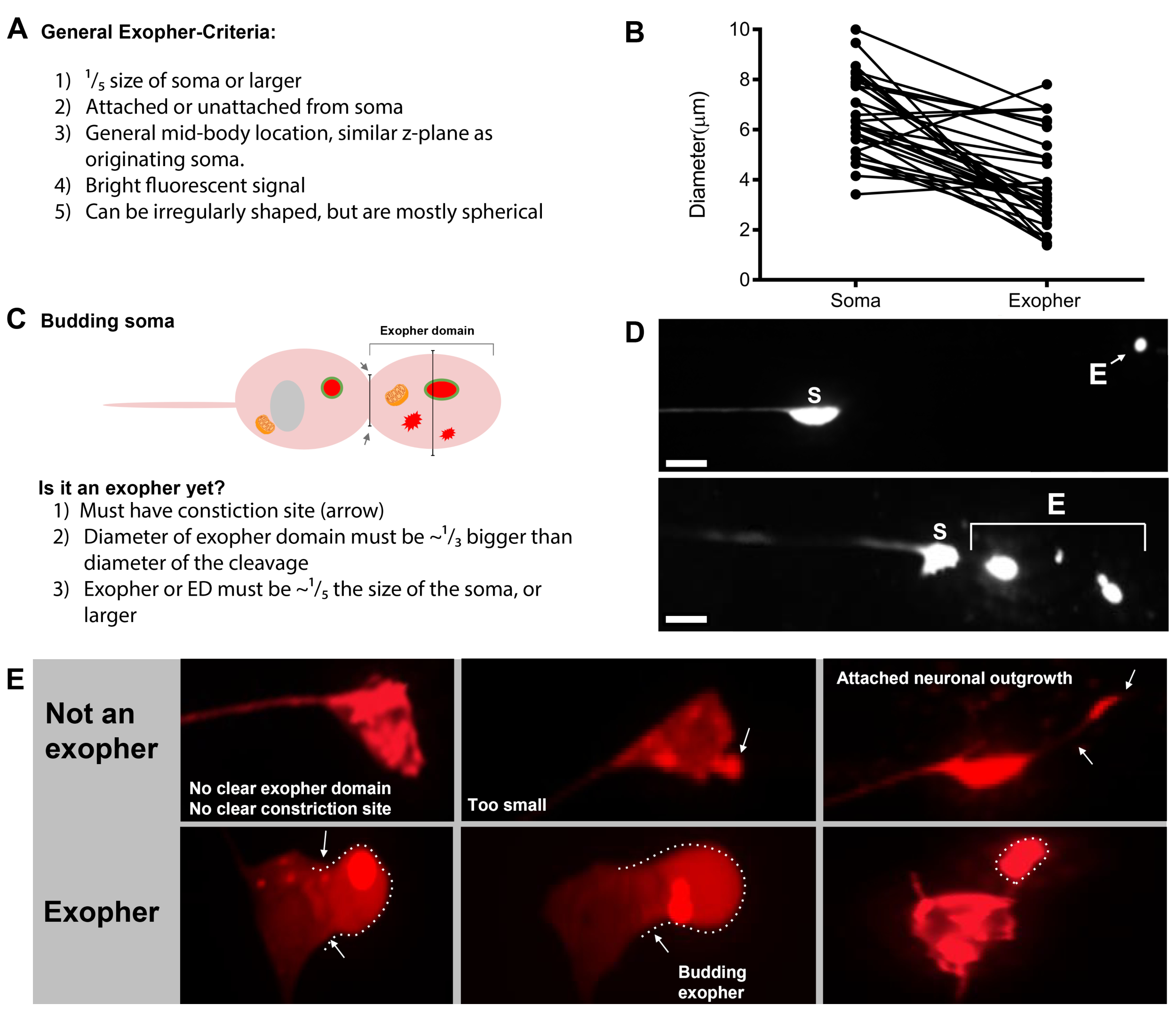
- Bir dokunma nöron bulunduktan sonra, büyük çıkıntılar için inceleyin (eksopher etki alanları) bir tomurcuk eksopher olarak kabul edilecek kadar büyük, (en az ulaşan soma boyutunu 1/5th th ulaşan)(Şekil 1C).
NOT: Ortalama eksopher çapı 2-8 μm civarında ölçerken, ortalama soma (ZB4065 bzIs166[Pmec-4::mCherry]) hayvan ölçüleri günde 6-10 μm 2 yetişkin(Şekil 7B). - Hiçbir tomurcuk veya eksoher etki gözlenirse, soma yayılan bağlı ince filaman için nöronal soma inceleyin. Ekli ekophers soma kökenli yakın ve benzer bir Z-düzlemde yer olma eğilimindedir.
NOT: Ekoferler her zaman soma bağlı kalmaz. Ekli bir filaman ın algılanması, nesnenin bir eksopher olduğunun kesin bir göstergesidir. - Eksiz bir eksopher tanımlamak için, bir eksopher içeriğini arayın. Eksophers çıkarılan floresan proteinler konsantre ve bu nedenle genellikle soma daha parlak.
NOT: Ekoferlerin içeriği heterojen ve değişkendir. Lizozomlar ve mitokondri gibi hücresel organeller de ekzotekler içinde ekstrüzyon yapılabilir (Şekil 4C-E). - Soma'nın bulunduğu düzlemden farklı odak düzlemlerinde bekar eksophers arayın. Ekophers herhangi bir yönde ALM soma çıkıntı sı görülmüş olsa da, bu eksophers uzak soma çıkıntı, nöronal süreçten bir posterior yönde tipiktir.
- Nöronal somas olarak tanımlanan ve konumlandırılmayan büyük, küresel nesneleri kontrol edin. Dışa dönükler düzensiz şekilli olabilir ama genellikle küresel yapılardır. Exophers zamanla bozulmuş olur, bu yüzden eski exophers daha düzensiz bir şekil var eğilimindedir.
NOT: Olgun veya yaşlı ekophers eksperlerin parlak floresan yoğunluğu ve küresel şekli ile dağınık "yıldızlı gece" aşamasından ayırt edilir. - Daha önceki ekofhergenezin kanıtı olarak "yıldızlı gece" fenotipini araştırın. Exophers bir "yıldızlıgece" sahne içine exopher küçük veziküller ayrılır ve çevreleyen hipodermis exopher içeriğini aşağılamak için çalışır gibi ilerleme(Şekil 1G, 2B , 3B & 7A).
NOT: Yıldızlı gece sahnesi, yapısal bütünlüğünü kaybetmiş ve dokunma nöronları ve eksopher yapılarına kıyasla loş bir floresan gösteren parçalanmış ve dağınık (bazen ağa bağlı) floresan varlıklarla işaretlenir. - Birden çok dış olayı örneklerine bakın. Ekopherler genellikle tekil bir oluşum olarak üretilir (1 somadan yayılan 1 ekopher) ancak bazı durumlarda birden fazla eksopher tek bir somadan serbest bırakılabilir(Şekil 7D).
NOT: Olgun eksophers hipodermis bozulmuş olarak birden fazla veziküller içine bozulabilir. Her eksopherin bağımsız bir eksozgenez olayı tarafından mı oluşturulduğunu veya ek bir vezikül oluşturmak için orijinal bir ekofonun bölünüp bölünmediğini ayırt etmek ancak zaman atlamalı gözlemle belirlenebilir. - Tüm morfolojik anormalliklerin eksophers olgunolduğunu unutmayın.
- Bir exopher olarak distended soma puan etmeyin. Uzun veya sivri soma zaman zaman görülebilir (özellikle yaş veya stres altında), ancak net bir daralma sitesi olmayan bir uzantısı bir eksopher olarak puanlanmaz.
- Eksopher olay nicelleştirmede somaboyutunun 1/5'ine ulaşamayan küçük çözülmüş tomurcukları reddedin.
- Neurite çıkıntıları ekofer olarak sayma. Olgun nöritler yaşla birlikte dramatik bir şekilde uzayabilir (genellikle nöronal sürecin ters yönünde) ve floresan protein gibi yapıların distal sonuna göç edebilir19.
NOT: Bu neurite outgrowths onlar gün ve hafta içinde ayrı bir gelişim deseni var gibi ekoferler değildir, tomurcukları oluşturmak değil, ve ayırmak yok (Şekil 7E).
- Ekopher olmayan floresan not varlıkları tanımlayın.
NOT: Ekstrüde floresan varlığın otofloresan ile doğru tanımlanmasını sağlamak için arka plan floresanlığı hakkında bir fikir edinmek önemlidir.-
Transgenik floresan ekspresyonuna karşı otofloresan. Otofloresansı transgenik ifadeyle karıştırmayın. Gerçek eksopher sinyali bağırsak veya bağırsakta olmayacaktır (DIC onayı bu dokuları tanımlamak için kullanılabilir) ve eksopher sinyali arka plan otofloresangöre önemli ölçüde daha parlak olacaktır.
NOT: Otofloresans bağırsak granül intestinal floresan pigmentasyon neden olur ve yaşla birlikte birikir. Heterojendir, özellikle farklı dalga boylarında incelenmiştir. - Embriyolardan sinyal. Embriyo sinyalini ekofhergenez ile karıştırmayın. Floresan'dan brightfield aydınlatmasına geçerek ve rahimdeki yumurtalarla sinyal çağrışımlarını kontrol ederek embriyo sinyali şüphelerini doğrulayın.
-
Uçaktan veya yakındaki soma gövdelerinden. Gözlemin başlangıcında, yakındaki tüm soma vücutlarını, hatta odak dışı somaları bile tanımlayarak, uçak dışı soma somaları bir eksopher için yanlış yapmaktan kaçının.
NOT: ALMR'den eksopherler için puanlama varsa, AVM ve ALMR somas'ın yerini belirleyin ve hesaplayın. Soma gövde tanımlaması ile ilgili daha fazla bilgi Şekil 3A'daaçıklanmıştır.
-
Transgenik floresan ekspresyonuna karşı otofloresan. Otofloresansı transgenik ifadeyle karıştırmayın. Gerçek eksopher sinyali bağırsak veya bağırsakta olmayacaktır (DIC onayı bu dokuları tanımlamak için kullanılabilir) ve eksopher sinyali arka plan otofloresangöre önemli ölçüde daha parlak olacaktır.
8. Puanlama ve istatistikler
- Skor ikili olarak exophers (evet, bir exopher / hayır, bir exopher yoktur).
- Belirli bir nöron için bir "eksopher-olay" olarak exopher algılama düşünün. Bir eksopher olayı, soma veya birden fazla eksopher yakınında tek bir eksopher gözlem teşkil edebilir.
NOT: Tek tek ekofhergenez olaylarının sayılarını ölçmek için zaman atlamalı gözlem kullanılır. - Farklı hücreler aynı oranda eksopher üretmediği için belirli bir tanımlanan hücre başına eksopher olaylarını sayın (örneğin Şekil 3C'yebakın). ALMR nöronlar burada açıklanan suşları en temel eksophers üretmek ve bu nedenle sık sık bu hücre dokunmatik reseptör nöronlardan ekofher nicelleştirme için seçilen.
- İstatistikler için, genel olarak, en az 3 biyolojik çalışmalar, en az 30 hayvan bozulma analizi için gerekli gözlemler karşılık gelen sayısı ile deneme başına puan ile.
- Kontrole kıyasla bir veya iki mutant/tedavi içeren çoklu çalışmalarda, Cochran-Mantel-Haenszel testi p değerlerini belirlemek için uygundur.
- Kontrole kıyasla ikiden fazla tedavi mutantı içeren çalışmalarda, herhangi bir sayıda kategorik tahmincinin önemini değerlendirmek için ikili lojistik regresyon analizi kullanmak da uygundur.
Sonuçlar
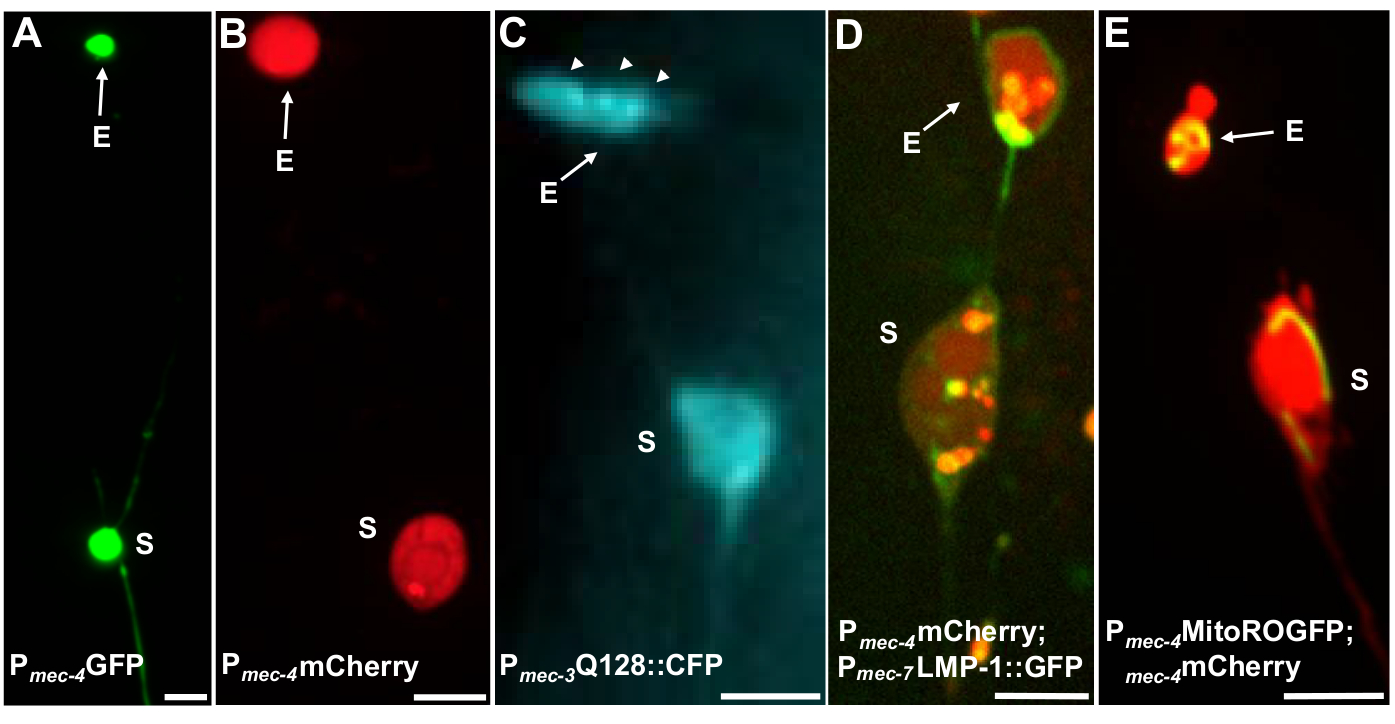
Exophers ölçmek için birden fazla floresan muhabirler kullanılabilir. Dokunma nöron eksophers kolayca ekstrüzyon için seçilebilir proteinlerin floresan etiketleme yoluyla in vivo görselleştirilmiş, ekstrüzyon olabilir organeller etiketleme, ya da hücre zarları etiketleme tarafından. Tablo 1, eksopherleri izlemek için kullanılan dokunma nöron ifade floresan muhabirleri tanımlar ve şekil 4'teyer alan temsili örneklerdir. Ekoferlerde ekstrüzyon yaptığı bilinen yükler arasında insan huntingtin'in N-terminal etki alanının genişletilmiş poliglutamin (Q128)(Şekil 4B)ile bir füzyonu , lysozomal membran proteini (LMP-1)(Şekil 4C)ile etiketlenmiş gfp etiketli lizozomlar ve mitokondri matris-lokalize GFP (Şekil 4D) ile etiketlenmiştir. Sitoplazmik GFP kuvvetle sınırdışı edilmez ve tercihen soma tutulur5, GFP zayıf ekoferler görselleştirmek rağmen(Şekil 4A). GFP çıkarılan proteinlere kaynaştığında, bu etiket eksopherleri görselleştirmek için kullanılabilir. Önemli bir nokta, farklı proteinlerin etiketlenerek, belirli yüklerin ve organellerin atılması ile ilgili çok çeşitli soruların yanı sıra dışa dönükleri oluşturan protein ve zarlarla da ele alınabileceğidir.
Sözde stereomikroskop kurulumu, hayvanlardaki eksopherleri agar plakalar üzerinde görüntülemek için etkili bir araçtır. Bu kurulum, her büyütme, sözde stereo teknoloji (stereoskopik taban üzerinde ayrık hedefler) ve yüklü hedeflere ara büyütme görüntüleme için bir zoom işletim anahtarı yüksek sayısal diyafram optik içeren bileşik ve stereoskopik teknoloji bir melez. Bunun gibi bir mikroskop, yüksek işlem liskor (tarama/toplama için kullanılan 2x hedef, tanımlama ve puanlama için kullanılan 10x hedefi) için nöronal morfoloji ve eksopher üretimini gözlemleyecek kadar güçlü 10x gözlük ve hedeflerle donatılmalıdır.
Standart stereomikroskopların büyütme yetenekleri genellikle floresan proteinleri ifade eden dokunmatik nöronların ağını görecek kadar yüksek çözünürlüğe sahip olsa da, soma'nın tübüler bağlantıları gibi dış eksilerin hücre altı detaylarını gözlemlemek için standart diseksiyon mikroskopları yeterli değildir. Bu tür gözlemler konfokal mikroskopi gerektirir (ekipman detayları için Malzeme Tablosu'na bakınız).
Exopher nicelik çalışmaları deneysel gerilmeleri ortadan kaldırmak için sıkı kontroller gerektirir. Tekrarlanabilir eksopher üretimi için tutarlı büyüme koşullarının özenli bakımı gereklidir. Daha spesifik olarak, eksopher üretimi, nesiller boyunca tutarlı besleme, sabit sıcaklık ve kontaminasyonsuz büyümenin tekrarlanabilirlik açısından kritik öneme sahip olması gibi strese duyarlıdır. mCherry yüksek nöronal ekspresyon ile bazal büyüme koşulları altında, eksopher üretimi nispeten düşük (ALM'lerin% 5-25 ekophers üretmek) ama bazı stresler, ozmotik ve oksidatif stres de dahil olmak üzere, eksopher oranlarını artırabilir. mCherry ekspresyonu stres olarak düşünülebilirken, eksopher düzeylerinin stres ekolünün bir sonucu olarak, eğer doğru şekilde kontrol edilirse, deneysel strese giriş eksophergenezin ivedine neden olmak ve gözlemlemek için bir strateji olabilir.
Zamanlama ve beklenen exopher üretim seviyeleri. Dışa dönükler larva gelişimi sırasında neredeyse yoktur. Genç erişkin yaşamda pik eksopher üretim dönemi son derece yetişkin gün 1-4 sırasında sınırlı gibi görünüyor, en sık yetişkin gün 2 veya 3 belirgin olmak. Tepe biraz ileri veya geri kayabilir çünkü, bir exopher üretim profilinin en eksiksiz değerlendirme yetişkin gün 1-4 üzerinde günlük birden fazla deneme puan etmektir. Genel olarak, bir ALMR en az 24 saat boyunca vezikül devam ile, bir büyük eksopher üretir. Eksopher oldukça hızlı bir şekilde (en hızlı dakika sırasına göre)üretilebilir. En yaygın olarak, sadece bir büyük eksopher erken erişkin yaşamında nöron başına üretilen, ancak birden fazla eksophers üretimi mümkündür.
Bazal koşullarda mCherry ifade ALM'ler tarafından genel eksopher üretimi yetişkin gün 2-3 optimal zaman dilimi içinde incelenen ALM'lerin% 5-25 arasında değişmektedir(Şekil 3D). Proteostaz krizleri5, yanı sıra diğer streslere maruz eksopher düzeyini modüle edebilirsiniz. Stres veya genetik tedirginlikler eksopher ekstrüzyonları üreten ALMR nöronların %90'ına kadar algılama oranlarına eksopher üretimini artırabilir.
Ekofhergenezde belirli genlerin rollerini test etmek için beslenme tabanlı RNAi. Nematod C. elegans genellikle rnai aşağı yem hayvanlar tarafından tabi tutulur E. coli suşu HT115 dönüştürülmüş bir çift iplikçikli RNA ifade (dsRNA) ilgi bir gen hedefleyen20. HT115 bakterileri RNAi besleme eksophers için puanlama kullanılabilir5. Çoğu dokuda transkript bu tekniği kullanarak RNAi tarafından hedef olabilir iken, nöronlar daha refrakter vardır. RNAi duyarlılığı transgenik dsRNA taşıyıcı SID-1 bir nöron özgü organizatörü altında ifade hayvanlar kullanılarak kalibre edilebilir. Bu şekilde nöronal doku RNAi21duyarlı olabilir.
İlgi bir gen doku özgü nakavt bu bileşeni eksik bir mutant içinde endojen RNAi metabolizmasının bir bileşeni ifade ederek gerçekleştirilebilir. Örneğin: Argonaute protein RDE-1 özellikle rde-1 mutant hayvanların nöronlarında, hayvanlar bu geni hedef alan bir RNAi müdahalesine maruz kaldığında sadece nöronların ilgilendiği bir genin yıkılmasını sağlamak için ifade edilebilir.
Standart nematod RNAi protokolleri kullanarak20,22, RNAi L4 aşamasında ebeveynlerin maruz kalma ve yetişkinlik güçlü genetik knock-down üretir kadar tüketen ht115 bakteri geliştirmek için onların döl izin ama deneysel hayvanlar boş bir vektör kontrolü daha farklı büyüyebilir gibi RNAi tarafından indüklenen potansiyel gelişimsel gecikmeler dikkatli olmak. Negatif kontrol karşılaştırması için boş vektör denetimini her zaman eklemek önemlidir. HT115 bakterileri rnai besleme eksophers için puanlama zaman kullanılabilir. Ancak, bazı genler RNAi maruziyeti daha kısa dönemlerde bile ekzoshergenez oranlarını değiştirmede etkili olduğunu unutmayın5. Bazı genlerin hedeflenmesi gelişimsel başarısızlığa yol açıyorsa, hayvanları ömür boyu nakavta maruz bırakmaktan kaçının, hayvanlar l4 evresinde L4'ten yetişkin D2 veya D3'e maruz kalmak için RNAi plakalarına doğru seçilebilir.
| Süzme adı | Genotip | Açıklama | Dışa dönük yüzdesi | Başvuru |
| SK4005 | zdIs5[Pmec-4GFP] | Dokunma nöronlarıngatinde GFP'nin sitosolik ekspresyonu. | 1-8% ALM | Şekil 4A, Melentijevic 2017 |
| ZB4065 | bzIs166[Pmec-4::mKiraz] | Dokunma nöronlarında vinç (bzIs166) aşırı ekspresyonu, hem sitosolik sinyal hem de mCherry agregaları üretir. bzIs166 bir eksopher indükleyicidir. mCherry agregaları ekofherenin inbelirleyicileridir ve tercihen ekofherlerde ekstrüzyonyapılır. | %3-20 ALM (normal koşullar). %20-80 ALM (açlık koşulları). | Şekil 4B, Melentijevic 2017 |
| ZB4067 | bzIs167[Pmec-4mitogfp Pmec-4mCherry4]; igIs1[Pmec-7YFP Pmec-3htt57Q128::cfp lin-15+]; | YFP sitosolik etiketler mec-7 dokunmatik nöronlar. Co-ifade Q128::CFP toplar ve ekophers neden olur. CFP tercihen sessiz. | ~25% | Şekil 4C, Meletijevic 2017 |
| ZB4509 | bzIs166[Pmec-4mCherry]; bzIs168[Pmec-7LMP-1::GFP] | bzIs168 LMP-1::GFP plazma membranlarını ve lizomal membranları etiketler. bzIs168 nöronal membranları, eksoherleri (membranbağlı oldukları için) ve lizomal membran yapılarını tanımlamak için kullanılabilir. | %3-20 ALM | Şekil 4D, Melentijevic 2017 |
| ZB4528 | bzIs166[Pmec-4mCherry]; zhsEx17 [Pmec-4mitoLS::ROGFP] | Allele zhsEx17, yerel oksidatif ortama göre en yüksek uyarma dalga boyunu 405nm 'den (okside) 476nm 'ye (azaltılmış) değiştiren mitokondrial lokalize bir muhabirdir. Bu dokunma nöronlar ifade edilir ve dokunmatik nöronlarda mitokondri tanımlamak için kendi başına kullanılabilir ve mito-exophers. | %3-20 ALM proteo-exopher. % ALM mito-ekopher quantitation devam etmektedir. | Şekil 4E, Melentijevic 2017, Top 2008, Ghose 2013 |
Tablo 1. Dokunma nöronları görselleştirme için kullanılan suşları, dokunmatik nöron ekophers, ve eksopher içeriği.

Şekil 1: Ekofhergenezin evreleri. Bir eksopher yapma ve çıkarma işlemine 'ekopher-genesis' denir. Eksopher oluşumunun dinamik süreci birkaç saat sürebilir. ZB4065 bzIs166[Pmec-4mCherry] yüksek eksoplis ürtüründe dinamik eksohergenez prosesi sırasında belirli basamaklarda soma ve ekofher morfolojiörnekleri gösterilmiştir. Tüm görüntüler gün 2 yetişkin ALM nöronlar 100x hedefi ile çekilen vardır. (A) Normal soma. Erişkin mekanosensoriyel dokunma nöron ALM transgenically Pmec-4mCherry ifade. Soma morfolojisi bu sitoplazmada mCherry konsantrasyonları ile, bu zorlanma genç yetişkin nöronlar tipiktir. (B) Erken tomurcuk fazı. Ekofhergenezin ilk gözlemlenebilir adımı, seçilen sitoplazmik materyalin soma zarının kenarına polarizasyonunu içerir. Bu adım genellikle bir genişleme veya soma şişmesi eşlik eder. Dokunma nöronları söz konusu olduğunda, pre-exopher etki alanı (PED) çevreleyen hipodermis içine uzanır (burada görünmez). Erken tomurcuk etki alanına mCherry malzemenin daha fazla konsantrasyon dikkat edin. (C) Geç tomurcuk fazı. Daha fazla hücresel polarizasyon ve pre-exopher etki alanının genişlemesi üzerine, soma ve eksopher (ok) arasında bir daralma belirgin hale gelir. Bu olay geç tomurcuk aşamasına geçiş sinyalleri. Geç tomurcuk aşamasında hücre açık bir daralma sitesi ve ayrı soma ve eksopher etki alanları sergiler rağmen, henüz soma tamamen kapalı çimdiklenmiş değil; tomurcuklanan eksopher kalın bir sap (ok) ile takılabilir. Söz konusu eksopher etki alanının çapı şantiye/sapın çapındakinden kabaca 1/3 daha büyük olduğunda tomurcuklanan etki alanı erken bir ekopher olarak kabul edilir. (D) Erken eksopher fazı. Erken dışa dönükler giden soma bir sap tarafından takılabilir-eksopher soma uzak hareket ettikçe bu bağlantının çapı inceltilebilir. Sitoplazmik malzeme soma dan eksomak için bu tüp üzerinden transfer edilebilir, çoğu malzeme tomurcuklanma işlemi sırasında yüklenir rağmen. Exophers soma dan ayrılabilirsiniz tasvir edildiği gibi (E), ayrılmış exophers olgun eksophers olarak kabul edilir (F). Olgun eksopher giden soma uzak hareket, çevreleyen hipodermal doku ile transit olabilir. (G) MCherry etiketli ekzosun hipodermis içinde daha küçük veziküllere dönüşmesi, büyük olasılıkla hipodermal endosomal ağa girerken mCherry materyalinin dağınık bir delinmiş görünümüne neden olur. Dağınık delinme sinyaline "yıldızlı gece" fazı denir. Bazı eksopher içeriğinin bozulması muhtemelen hipodermal lizozomlar tarafından gerçekleştirilir, ancak bazı malzeme tam olarak bozulmaz ve genellikle hipodermis tarafından pseudocoelom içine yeniden ekstrüzyon edilir. Post-ekofhergenez mCherry transit Şekil 2'dedaha ayrıntılı olarak açıklanmıştır. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 2: ekofherlerde dokunma nöronlarından ekstrüzyon yapan mCherry, çevredeki hipodermal lisozomal ağı devreye sokar ancak daha sonra coelomocytes'in mCherry'yi depolayabildiği/bozabileceği pseudocoelom'a ekstrüzyon edilebilir. (A) MCherry'nin dışa dönük olarak ekstrüzyonun unnöronlar tarafından sınırdışı edildikten sonra vücuda nasıl aktulu olduğunu anlatan çizgi film özeti. Ekzotermezisi sırasında mCherry gibi seçilmiş hücresel içerik lokalize olur ve nöronal ve hipodermal plazma membranları ile çevrili bağımsız bir vezikül içinde nöronal soma gönderme kapalı tomurcuk. Dokunma nöronları hipodermal dokuda gömülü olduğundan, eksopher etki alanı dışa doğru tomurcuklanır gibi hipodermisin içine daha fazla hareket eder. Eksopher hipodermisin geçişini yapabilir ve saatler sonra günlere, eksopher içeriği hipodermisin endolysomal ağı içinde parçalanabilir. mCherry hipodermis boyunca dağınık puncta olarak görünebilir, bir sahne "yıldızlı gece" denir. Birkaç gün sonra, bazı mCherry hipodermis dışarı çevreleyen pseudocoelom içine geçebilir, coelomocytes denilen çöpçü hücreleri erişebilir, ve alabilir, depolanabilir mCherry. (B) Yıldızlı gece mCherry veziküllerin görünümüne örnek. Büyük eksopher parçaları ve yıldızlı gece vezikülleri ile mCherry ile etiketlenmiş bir ALM soma Görüntü. Suşu ZB4065 bzIs166[Pmec-4mCherry]. (C) Uzak coelomocytes mCherry konsantrasyonu örneği. Yetişkin bir hayvan gün 10 suşu ZB4065 bzIs166[Pmec-4mCherry] gösteren mCherry coelomocytes (oklar) konsantre gösteren sideview. Bazı yıldızlı gece vezikülleri de belirgindir. Genel olarak coelomocyte konsantrasyonu yetişkin gün 5 yaşam hakkında sonra belirgin hale gelir. (B alt) Karikatür üreme (B), dokunmatik nöronlar ve süreçleri kırmızı özetlenen ile, parlak eksopher parçaları gibi; farklı Z-derinliklerde dağınık küçük veziküller açık pembe gösterilir. (C alt) Görüntükarikatür versiyonu(C),kırmızı nöronal süreci gösteren, pembe yıldızlı gece ve yeşil coelomocytes. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 3: Mekanosensory dokunma nöronlar hassas bir zamansal profil ile farklı düzeylerde eksostutanlar üretmek. (A) (Top) C. elegans anahtar anatomik işaretleri ile mekansal ilişkili mekanosensory touch nöronlar Karikatür tasviri pompalama farinks ve hayvanın başında nöron yoğun sinir halkası da dahil olmak üzere, orta vücut vulva, ve konik kuyruk. (Alt) Üst ve sol taraftan (WormAtlas'tan uyarlanmış görüntüler) GFP'yi ifade eden floresan olarak etiketlenmiş dokunma nöronları. Kırmızı kutu, ALM ekopherlerinin genellikle bulunduğu alanı betimlenir. (B) ALM kaynaklı eksopherlerin [Pmec-4mCherry] içeren bir gerilimde üretildiği orta gövde bölgesinin yüksek büyütme görünümü. AVM ve ALMR nöron tasvir edilir, ve gösterilen bir ALMR eksopher mCherry yıldızlı gece ile birlikte. ALMR nöronlar en kolay ekofhers üretmek. (C) ALMR mekanosensory touch nöronlar daha kolay bazal koşullar altında hermafroditler diğer dokunma nöronlar ile karşılaştırıldığında eksophers üretmek. Yetişkin gün 2 mechanosensory touch nöron eksopher üretimi, bireysel dokunma reseptör nöronları için puan olarak gösterilir. Zorlanma: ZB4065 bzIs166[Pmec-4mCherry], N>150, hata çubukları SEM. (D) ALMR dokunma nöronlar gün 2 ve 3 yetişkinlik döneminde daha fazla exophers üretmek ergen L4 evre veya ileri yaştaki hayvanlar ile karşılaştırıldığında. Strain: ZB4065 bzIs166[Pmec-4mCherry], N>150, hata çubukları SEM vardır.
{kind=link}

Şekil 4: Eksopher içeriğini etiketleyen bazı floresan muhabirlere örnekler. Ekophers gözlemlemek için basit bir yolu nöronal organizatörlerden floroforlar ifade transgenik hayvanlar yaratmaktır. Florofororlar ekoferin görselleştirilmesine izin verir ve transgenik ekspresyonun eksophergenezi artıran agregasyon ve/veya proteostrese neden olur. Amtrem nöronlar tarafından üretilen eksophers da görselleştirme için boya dolgu kullanarak, doğal koşullar altında görülebilir. Gösterilen eksophers gözlemlemek için kullanılabilecek ortak suşları örnekleridir, (E) eksopher, (S) soma. (A) Soma ve suş SK4005 zdIs5[Pmec-4GFP], 100x objektif fotoğrafçılık, ölçek çubuğu 3μm için kullanılan bir yetişkin bir ALM exopher. Bu zorlanmada, çözünür GFP içeren ekoferler ölçülür, ancak eksopher üretimi seyrek olarak gerçekleşir. GFP'nin diğer çalışmalarda ekstrüzyonlarda tercihen ekstrüzyon yapılabilen proteinlere bağlanması, GFP füzyonlarının olgun dış eksizyonlarda tespit edilebilen olduğunudoğrulamaktadır. (B) ALM soma ve zorlanma ZB4065 bzIs166[Pmec-4mCherry] bir yetişkin exopher, hangi mCherry ifade eder ve dokunma nöron ekopher üretimi neden olur. Fotoğrafçılık için kullanılan 100x objektif, ölçek çubuğu 5 μm. (C) ALM soma ve zorlanma ZB4067 bzIs167[Pmec-4mitogfp Pmec-4mCherry4]; igIs1[Pmec-7YFP Pmec-3htt57Q128::cfp lin-15+]; htt57Q128 görüntü için kullanılan seçici mavi kanal::CFP. Eksopher htt57Q128 içerir::CFP agregalar (oklar), daha soma daha eksopher konsantre görünür. Fotoğrafçılık için kullanılan 40x objektif, ölçek çubuğu 5μm. (D-E) Ekophers organeller içerebilir ve floresan proteinler ile organel özgü etiketleme organel ekstrüzyon izleme sağlar. (D) Lysozomal membran etiketi LMP-1::GFP soma ve eksoher membranı özetler ve plazma membranlarını zayıf etiketler (plazma membran lokalizasyonu lisozomal hedefleme yolunda bir kaçakçılık adımıdır) ve lysosomal organelleri güçlü bir şekilde etiketler. Gösterilen bir yetişkin ALM soma co-ifade Pmec-4mCherry ve Pmec-7LMP-1::GFP membranlar ve lyzozomlar lokalize olduğunu. Soma eksopher parçaları (oklar) olması muhtemel diğer küçük ekstrüzyonlar ile ekli bir eksopher vardır. GFP pozitif yapılar soma dahil ve büyük eksopher mevcuttur, gerginlik: ZB4509 bzIs166[Pmec-4mCherry]; bzIs168[Pmec-7LMP-1::GFP]. Fotoğrafçılık için kullanılan 100x objektif, ölçek çubuğu 5 μm. E) Soma ve ekopherlerde mitokondriyi tanımlamak için mitokondriyal GFP belirteci kullanılabilir. Gösterilen pmec-4mCherry ve mito ifade yetişkin bir ALM soma::ROGFP, mitokondriyal matris lokalize. mito::ROGFP tek başına ifade, mCherry olmadan, aynı zamanda kolayca nöronlar ve mitokondri içeren eksophers için puan tanımlamak için kullanılabilir. Suşu: ZB4528 bzIs166[Pmec-4mCherry]; zhsEx17 [Pmec-4mitoLS::ROGFP]. Fotoğrafçılıkiçin kullanılan 100x objektif; ölçek çubuğu 5μm. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 5: C. elegans ve L4 tanımlamasının gelişim döngüsü. (A) 20 °C'de bir yumurtanın anne tarafından döşendikten sonra yumurtadan çıkması yaklaşık 9 saat sürer. (B) Yeni yumurtadan çıkan bir hayvan larva evre1 (L1) ve 12 saat sonra L2 larvaiçine molts olduğunu. (C) Hayvanlar L2 ve (D) L3 larva evrelerinde yaklaşık 8 saat ertelerinde kalır. (E) Ergen hayvanlar dördüncü larva evresi (L4) olarak kabul edilir ve orta gövdeye yakın beyaz bir hilal olarak görünen dikkat çekici gelişen vulva ile işaretlenirler. Hilal daha sonra eksophers için puanlama kolaylaştırmak senkronize kültürler oluşturmak için L4 sahnelenen hayvanların kolay belirlenmesi ve toplanması sağlar iken bu varlığı. Hayvanlar gravid yetişkinler içine son molt önce yaklaşık 10 saat Boyunca L4 aşamasında kalır, F) yumurta geliştirme ile tanımlanan, görünür spermatheca, ve yumurta yumurtlama başlatılması. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 6: Mikroskop slayt agar ped hazırlanması. (A) Laboratuvar bandı nın tek bir şeridi ile iki slayt hazırlayın. Resimde olduğu gibi arasına bantsız bir mikroskop slaytı yerleştirin. B) Slaytın üzerine erimiş agarose damlası yerleştirin. (C) Temiz bir slaydı yavaşça damlanın üzerine yerleştirin, sönmüş bir daire pedi içine agarose tuşuna basarak. (D) Bantlı slaytları çıkarın, hangi bir çift ped oluşturmak için gerekli olan agar eşit bir düzleme gerçekleştirmek için hareket. (E) Agarose ped kuruduktan sonra üst teki kaydırağı çıkarın. (F) Pipet agar ped üstüne felçedici bir çözelti (levamisole veya tetramisole). (G) Uygun şekilde sahnelenmiş hayvanları felç haline getirin. (H) Hayvanları hafifçe örtün ve hayvanların hayatta olduğundan emin olun. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 7: Eksophers ve exopher tanımlama kriterleri karakterleri. (A) Bir eksopher tanımlayan genel kriterler. (B) Gönderen soma ile ekstrüzyon eksopher arasındaki çap karşılaştırmaları, μm cinsinden ölçülür. Yetişkin ALM somas, N = 35, zorlanma: ZB4065 bzIs166[Pmec-4mCherry] - soma 6.53 μm ortalama boyutu ve 3.83 μm dışa dönük ortalama boyutu. (C) Bir eksopher etki alanı ile tomurcuklanan bir eksopher arasında ayrım için ölçütler tanımlama. (D) En yaygın olarak, bireysel nöronlar bir büyük eksopher yapmak, daha sonra bölünür veya hipodermis içeriğini aşağılamak için çalışır gibi parçalar. Yine de, birden fazla eksophers bir nöron ya birden fazla exopher olaylar dan türetilmiş olabilir bir dokunmatik nöron yanında görülebilir ya da alternatif olarak, eksophers da tomurcuk veya parça kendilerini olabilir. Birden fazla eksopher benzeri varlığın kökeni sadece zaman atlamalı mikroskopi kullanılarak belirlenebilir. Top tek bir uzak eksopher ile bir ALMR dokunmatik nöron soma tasvir. Alt birden fazla eksopher benzeri ekstrüzyonları ile bir ALMR dokunmatik nöron soma tasvir. (E) Erişkin ALM dokunma nöron somas yaygın morfolojik özellikleri eksosher olaylar için yanlış olabilir. Sol üstte - Açık bir exopher etki alanı veya daralma sitesi ile bir distended ALM soma. Üst orta - Nöronlar eksophers benzer olabilir küçük hücre dışı çıkıntılar olabilir, ancak bir ekofher olarak kabul edilecek boyut gereksinimi kriterlerini karşılamaz. Sağ üstte – Yaşla birlikte, dokunma nöronları küçük neurite boyunca büyümeler gelişebilir. Genellikle mCherry malzeme neurite outgrowth ucunda toplanabilir. Toplanan mCherry exopher-to-soma boyut gereksinimlerini karşılamazsa bu bir exopher olarak puanlandırılamaz. Alt bir eksopher etki alanı veya eksopher için tanımlayıcı kriterlere sahip yetişkin ALM nöronlar göstermektedir. Botom sol - ALM soma seçici mCherry sitosol ve mCherry etiketli agregalar içeren önemli bir eksopher etki alanı vardır. Eksopher etki alanı daralma sitesi oklarla işaretlenir ve boyut ölçütlerini (soma boyutunun en az1/5'i) karşılar. Eksopher etki alanının en büyük çapı, bir eksopher olayı için kriterleri karşılayan, daralma alanının çapından neredeyse 1/3 daha büyüktür. Alt orta - ALM soma boyut ölçütlerini karşılayan önemli bir tomurcuklanma exopher vardır. Açık bir daralma sitesi vardır. Sağ alt - Eksopher boyut gereksinimlerini karşılayan bağlı bir mCherry dolu eksopher olan ALM soma. Eksopher ince bir bağlantı filamenti ile bağlanır. Tüm görüntüler gerginlik ZB4065 bzIs166[Pmec-4mCherry] vardır. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
Tartışmalar
Agrega ve organel eliminasyonunun in vivo moleküler mekanizmalarının büyük eksophers şeklinde karakterizasyonu emekleme dönemindedir. Sınırdışı için yüklerin atanması, hücre içindeki bu yüklerin polarize toplaması, eksopher oluşturma kararının düzenlenmesi, ekstrüzyonlara aracılık eden makinelerin düzenlenmesi ve ekstrüzyonların komşu hücredeki bozunan makinelerle etkileşimi ile ilgili sorular ele alınmaya devam etmektedir. Ayrıca, kalsiyum, agrega ve mitokondri içeren biyolojik maddeleri geçirebilen boru bağlantılarının in vivo görselleştirilmesi ilginçtir ve kendi başına yeterince incelenmiştir. Bazı hücrelerin neden eksopher üretimine diğerlerinden daha yatkın olduğu sorusu da çözülmese de, bu protokolde belirtilen yaklaşımlarla genetik olarak incelenmeye başlanabilir.
Bu protokolde ayrıntılı olarak açıklanan eksopher üretiminin tekrarlanabilir skorlama elde etmek için yaklaşımlar, yakındaki hücre somas gelen eksophers ayırt dikkat ile, eksopher üretiminin doruk yakalamak için analizzamanlaması, ve ekofher düzeylerini modüle etmek istenmeyen stresleri ortadan kaldırmak için büyüme koşullarının sıkı kontrolü. Büyük erken dışa dönük lük ya da çevredeki hipodermisin "yıldızlı gece" dağılımı, eksopher üretiminin kanıtı olarak ölçülebilir. Bu söyleniyor, bazal koşullar altında mCherry ifade nöronlar en sık bir eksopher üreten belirli bir tip nöronların% 5-25 ile ilişkilidir. Ekstrüzyon üreten nöronların %90'ı kadar yüksek olan ve özellikle modifiye ediciler için genetik veya farmakolojik ekranlar için yararlı olan eksopher üretimini artırmak için stres koşullarının kontrollü olarak getirilmesi uygulanabilir.
İnsan nörodejeneratif hastalık, büyük agregalar patoloji yayılmasını teşvik etmek için komşu hücrelere hastalıklı nöronlar aktarabilirsiniz. Eksopher mekanizması phyla boyunca agrega ekstrüzyon için kullanılan korunmuş bir mekanizma ile transpire olabilir. Ya bu sürecin verimliliğini artırmak in vivo molekülleri tanımlanması (daha etkili proteostaz kontrolü olarak kabul edilir) ya da birden fazla nörodejeneratif hastalıklarla mücadele için yeni stratejilerin tasarımı nı etkilemek için harnessed olabilir blok. Bu nedenle, burada açıklanan protokol klasik genetik mutagenez ekranları, genom çapındaki RNAi ekranları, arttırıcıları ve bastırıcıları tanımlamak için genleri sistematik olarak deviren ekranlar veya bu sürecin aday farmakolojik değiştiricilerini tanımlayan ilaç müdahale çalışmaları için kullanılabilir. Yaklaşım, biraz zahmetli olmasına rağmen, basittir. Eksophers onlar yüksek büyütme diseksiyon mikroskop ile görülebilir büyüktür. Yine de, C. elegans nöronlar nispeten küçük ve organelleri veya membranlar bakarak daha yüksek güç konfokal görüntüleri gerektirir ve yavaş bir süreçtir. Daha yüksek iş elde etme seçenekleri, çok iyi plaka biçiminde yüksek içerikli görüntüleme yaklaşımlarını içerebilir.
Eksopher puanlama için standart bir yaklaşım uygulaması hangi nöronlar organize ve hücresel enkaz ortadan kaldırmak sürecin uyumlu bir genetik diseksiyon altında yatan olmalıdır.
Açıklamalar
Hiçbiri
Teşekkürler
Aşağıdaki NIH hibelerini kabul ediyoruz: R01AG047101 ve R37AG56510. Driscoll ve Grant laboratuvarlarının üyeleri, sıkı deneyler ve güçlü iletişim le tanımlanan protokollerin geliştirilmesine ve ince ayarlanmasına büyük katkıda bulunmuştur.
Malzemeler
| Name | Company | Catalog Number | Comments |
| 95B Scientific CMOS camera | Photometrics Prime | ||
| 1,000 μL low retention tips | Sarstedt | ||
| 10 mL serological pipette | Appleton Woods | CC214 | |
| 10 μL low retention tips | Sarstedt | 70.1130.105 | |
| 13% sodium hypochlorite | Acros Organics | AC219255000 | |
| 15 mL centrifuge tubes | Fisher Scientific | 05-539-12 | |
| 2 L erlenmeyer flasks | Scientific Laboratory Supplies | FLA4036 | |
| 25 mL serological pipette | Appleton Woods | CC216 | |
| 300 μL low retention tips | Sarstedt | 70.765.105 | |
| 50 mL serological pipette | Appleton Woods | CC117 | |
| 5-Fluoro-2'-deoxyuridine 98% | Alfa Aesar | L16497.ME | |
| 9 cm sterile Petri dishes | Fisher Scientific | 11309283 | |
| absolute ethanol | Vwr | 20821.33 | |
| Agar | Sigma Aldrich | A1296 | |
| C. elegans strain wild type | Supplied by CGC | N2 | C. elegans strain |
| calcium chloride dihydrate | Sigma Aldrich | C3881 | |
| cholesterol | Acros | 110190250 | |
| dibasic sodium phosphate | Sigma Aldrich | S3264 | |
| E. coli strain OP50 | Supplied by CGC | Op50 | E coli strain |
| FBS10 Standard microscope | Meyer Instruments | KSC 410-1-100-1 | FBS10 Standard with Plate Base, 100/100 Trinocular Head and Flip zoom |
| glass pipette 270 mm | Fisherbrand | FB50255 | |
| Heraeus Multifuge X3R | Thermofisher scientific | 75004515 | |
| Inoculating Spreaders | Fisher Scientific | 11821741 | |
| LB medium capsules | MP biomedicals | 3002-031 | |
| LDI – Laser Diode Illuminator | 89 North | ||
| levamisole | Sigma Aldrich | 16595-80-5 | |
| M4 multipette | Eppendorf | 4982000012 | |
| magnesium sulphate | Sigma Aldrich | M7506 | |
| monobasic potassium phosphate | Sigma Aldrich | P0662 | |
| Multitron Standard shaking incubator | Infors HT | INFO28573 | |
| Nalgene 1 L Centrifuge pots | Fisher Scientific | 3120-1000 | |
| P10 pipette | Eppendorf Research Plus | 3123000020 | |
| P1000 pipette | Eppendorf Research Plus | ||
| P200 pipette | Eppendorf Research Plus | 3123000055 | |
| pipeteboy 2 | VWR | 612-0927 | |
| Polystyrene microbeads | Sigma Aldrich | MFCD00131491 | |
| RC5C plus floor mounted centrifuge | Sorvall | 9900884 | |
| Reusable ringed cytology slides | ThermoFisher Scientific | 22037242 | |
| SK4005 zdIs5[Pmec-4GFP] | contract Driscoll lab | GFP expressed in touch neurons | |
| sodium chloride | Sigma Aldrich | 13422 | |
| Sodium hydroxide | Fisher Chemical | S/4880/53 | |
| Tactrol 2 Autoclave | Priorclave | ||
| Triton-X | Thermofisher scientific | 28313 | |
| Tween 20 | Sigma Aldrich | 9005-64-5 | |
| X-Light V2 Spinning Disk Confocal Unit | CrestOptics | ||
| ZB4065 bzIs166[Pmec-4mCherry] | contract Driscoll lab | mCherry expressed in touch neurons | |
| ZB4067 bzIs167[Pmec-4mitogfp Pmec-4mCherry4]; igIs1[Pmec-7YFP Pmec-3htt57Q128::cfp lin-15+] | contract Driscoll lab | Q128 expressed in touch neurons | |
| ZB4509 bzIs166[Pmec-4mCherry]; bzIs168[Pmec-7LMP-1::GFP] | contract Driscoll lab | mitoROGFP expressed in touch neurons | |
| ZB4528 bzIs166[Pmec-4mCherry]; zhsEx17 [Pmec-4mitoLS::ROGFP] | contract Driscoll lab | autophagy marker expressed in touch neurons | |
| ZEISS Axio Vert.A1 | Zeiss |
Referanslar
- Davis, A. A., Leyns, C. E. G., Holtzman, D. M. Intercellular Spread of Protein Aggregates in Neurodegenerative Disease. Annual Review of Cell and Developmental Biology. 34, 545-568 (2018).
- Davis, C. H., et al. Transcellular degradation of axonal mitochondria. Proceedings of the National Academy of Sciences of the United States of America. 111 (26), 9633-9638 (2014).
- Torralba, D., Baixauli, F., Sanchez-Madrid, F. Mitochondria Know No Boundaries: Mechanisms and Functions of Intercellular Mitochondrial Transfer. Frontiers in Cell and Developmental Biology. 4, 107 (2016).
- Stahl, P. D., Raposo, G. Extracellular Vesicles: Exosomes and Microvesicles Integrators of Homeostasis. Physiology (Bethesda, Md.). 34 (3), 169-177 (2019).
- Melentijevic, I., et al. C-elegans neurons jettison protein aggregates and mitochondria under neurotoxic stress. Nature. 542 (7641), 367 (2017).
- Nussbaum-Krammer, C. I., Park, K. W., Li, L., Melki, R., Morimoto, R. I. Spreading of a prion domain from cell-to-cell by vesicular transport in Caenorhabditis elegans. PLoS Genetics. 9 (3), 1003351 (2013).
- Tyson, T., et al. Novel animal model defines genetic contributions for neuron-to-neuron transfer of alpha-synuclein. Scientific Reports. 7, (2017).
- Babcock, D. T., Ganetzky, B. Transcellular spreading of huntingtin aggregates in the Drosophila brain. Proceedings of the National Academy of Sciences of the United States of America. 112 (39), 5427-5433 (2015).
- Pearce, M. M. P., Spartz, E. J., Hong, W., Luo, L., Kopito, R. R. Prion-like transmission of neuronal huntingtin aggregates to phagocytic glia in the Drosophila brain. Nature Communications. 6, 6768 (2015).
- Fu, H., Li, J., Du, P., Jin, W., Cui, D. Metabolic wastes are extracellularly disposed by excretosomes, nanotubes and exophers in mouse HT22 cells through an autophagic vesicle clustering mechanism. bioRxiv. 10 (1), (2019).
- Ghose, P., Park, E. C., Tabakin, A., Salazar-Vasquez, N., Rongo, C. Anoxia-reoxygenation regulates mitochondrial dynamics through the hypoxia response pathway, SKN-1/Nrf, and stomatin-like protein STL-1/SLP-2. PLoS Genetics. 9 (12), 1004063 (2013).
- Cannon, M. B., Remington, S. J. Redox-sensitive green fluorescent protein: probes for dynamic intracellular redox responses. A review. Methods in Molecular Biology. 476, 51-65 (2008).
- Perkins, L. A., Hedgecock, E. M., Thomson, J. N., Culotti, J. G. Mutant sensory cilia in the nematode Caenorhabditis elegans. Developmental Biology. 117 (2), 456-487 (1986).
- Stiernagle, T. Maintenance of C. elegans. WormBook: The Online Review of C. Elegans Biology. , 1-11 (2006).
- Sutphin, G. L., Kaeberlein, M. Measuring Caenorhabditis elegans life span on solid media. Journal of Visualized Experiments. (27), (2009).
- Mitchell, D. H., Stiles, J. W., Santelli, J., Sanadi, D. R. Synchronous growth and aging of Caenorhabditis elegans in the presence of fluorodeoxyuridine. Journal of Gerontology. 34 (1), 28-36 (1979).
- Weicksel, S. E., et al. A novel small molecule that disrupts a key event during the oocyte-to-embryo transition in C. elegans. Development. 143 (19), 3540-3548 (2016).
- Dong, L., et al. Reversible and long-term immobilization in a hydrogel-microbead matrix for high-resolution imaging of Caenorhabditis elegans and other small organisms. PloS One. 13 (3), 0193989 (2018).
- Toth, M. L., et al. Neurite sprouting and synapse deterioration in the aging Caenorhabditis elegans nervous system. Journal of Neuroscience. 32 (26), 8778-8790 (2012).
- Conte, D., MacNeil, L. T., Walhout, A. J. M., Mello, C. C. RNA Interference in Caenorhabditis elegans. Current Protocols in Molecular Biology. 109, (2015).
- Calixto, A., Chelur, D., Topalidou, I., Chen, X., Chalfie, M. Enhanced neuronal RNAi in C. elegans using SID-1. Nature Methods. 7 (7), 554-559 (2010).
- Maher, K. N., Catanese, M., Chase, D. L. Large-scale gene knockdown in C. elegans using dsRNA feeding libraries to generate robust loss-of-function phenotypes. Journal of Visualized Experiments. (79), e50693 (2013).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır