
Method Article
Enfoques cuantitativos para la puntuación in vivo de la extrusión de agregados neuronales y organelle en vesículas de exoféro grandes en C. elegans
* Estos autores han contribuido por igual
En este artículo
Resumen
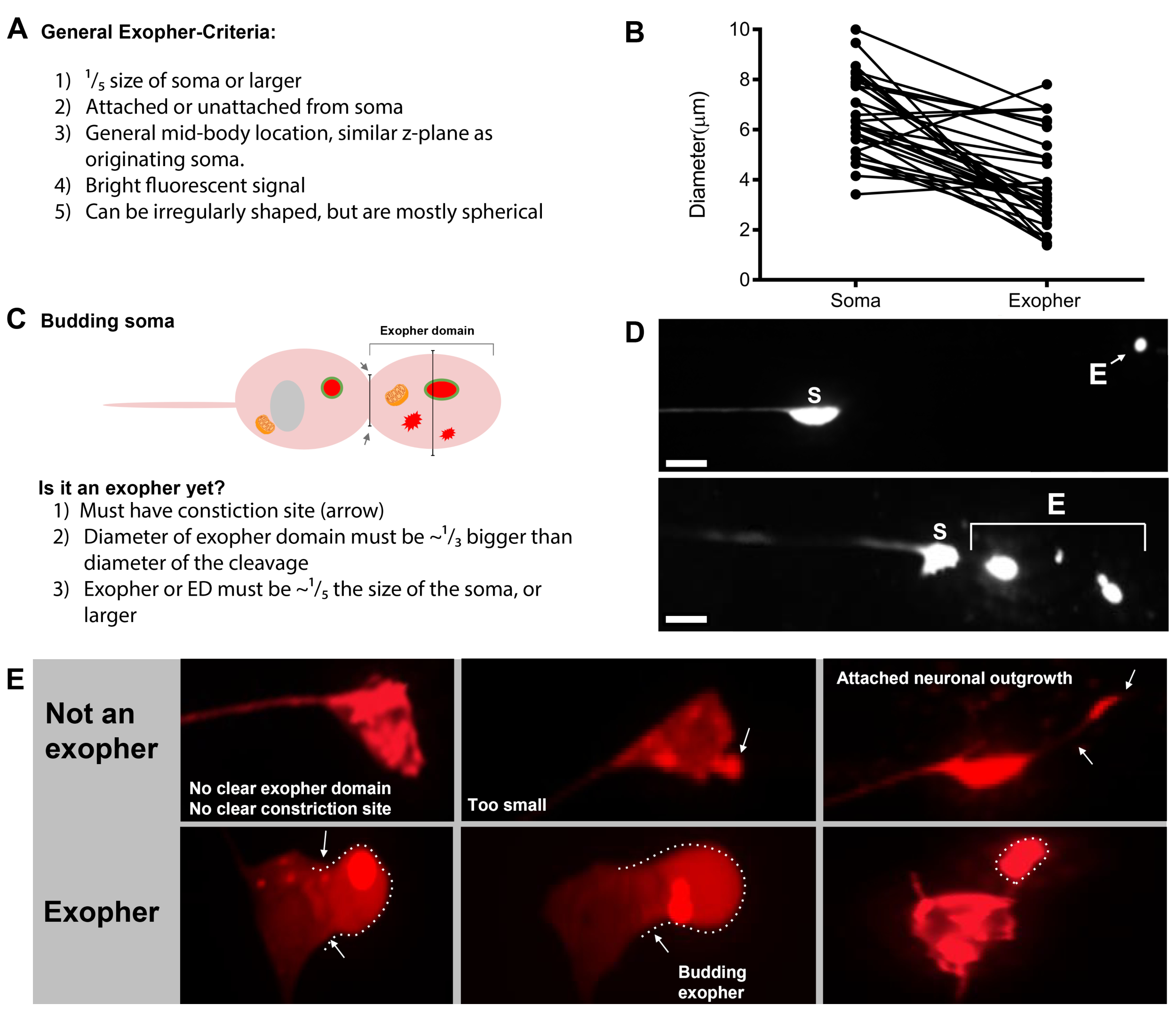
Este protocolo describe los enfoques para la detección y cuantificación de grandes extrusiones de agregados y/o orgánulos (4 m) producidos por células C. elegans en forma de exoferos unidos a la membrana. Describimos las tensiones, las condiciones de crecimiento, los criterios de puntuación, el momento y las consideraciones de microscopía necesarias para facilitar la disección de este mecanismo de expulsión de escombros.
Resumen
La toxicidad de las proteínas mal pleidas y la disfunción mitocondrial son factores fundamentales que promueven el deterioro neuronal funcional asociado a la edad y la enfermedad neurodegenerativa entre especies. Aunque estos desafíos neurotóxicos se han considerado durante mucho tiempo como intrínseco de las células, la evidencia considerable ahora apoya que las proteínas de enfermedad humana mal pleidas que se originan en una neurona pueden aparecer en las células vecinas, un fenómeno propuesto para promover la propagación de la patología en la enfermedad neurodegenerativa humana.
C. elegans neuronas adultas que expresan proteínas agregantes pueden extruir grandes vesículas rodeadas por membrana (4 m) que pueden incluir la proteína agregada, las mitocondrias y los lisosomas. Estas vesículas grandes se llaman "exoferas" y son distintas de los exosomas (que son aproximadamente 100 veces más pequeñas y tienen diferente biogénesis). La eliminación de desechos celulares en exophers puede ocurrir por un mecanismo conservado que constituye una rama fundamental, pero antes no reconocida, de la proteostasis neuronal y el control de calidad mitocondrial, relevante para los procesos por los cuales los agregados se propagan en enfermedades neurodegenerativas humanas.
Mientras que los exoferores han sido estudiados principalmente en animales que expresan mCherry transgénico de alta copia dentro de las neuronas táctiles, estos protocolos son igualmente útiles en el estudio de la exofergénesis utilizando orgánulos etiquetados fluorescentemente u otras proteínas de interés en varias clases de neuronas.
Aquí se describen las características físicas de los exophers de C. elegans, las estrategias para su detección, los criterios de identificación, el momento óptimo para la cuantificación y los protocolos de crecimiento animal que controlan las tensiones que pueden modular los niveles de producción de exoféros. Juntos, los detalles de los protocolos descritos aquí deben servir para establecer un estándar para el análisis cuantitativo de los exophers en todos los laboratorios. Este documento busca servir como un recurso en el campo para laboratorios que buscan elaborar mecanismos moleculares mediante los cuales se producen los exophers y por los cuales los exopheros son reaccionados por células vecinas y distantes.
Introducción
Los desafíos neurotóxicos de los agregados y las mitocondrias disfuncionales se han considerado durante mucho tiempo intrínsecos a las células, pero más recientemente se ha puesto de manifiesto que las proteínas de enfermedad humana mal pleidas que se originan en una neurona también pueden propagarse a las células vecinas, promoviendo la patología1. Del mismo modo, las mitocondrias de mamíferos pueden enviarse fuera de la célula de su producción original para la degradación transcelular2 o para el rescate de poblaciones mitocondriales en células vecinas desafiadas3. Generalmente se han observado vesículas de varios tamaños para transferir materiales celulares a células vecinas o a entornos de fluidos4. Algunas vesículas extruidas se acercan al tamaño del soma neuronal promedio (soma de neurona táctil promedio a 6 m) y pueden acomodar grandes agregados y orgánulos.
Un ejemplo llamativo de gran extrusión de vesículas que puede transportar agregados proteicos y orgánulos se produce en las neuronas receptoras táctiles de C. elegans que expresan una construcción de reportero de número alto de copia que codifica un mCherry5nociva propenso a la agregación y resistente a la degradación. Las extrusiones de las neuronas táctiles, llamadas exoferas, tienen un diámetro medio de 4 m, incluyen selectivamente mCherry u otros agregados, y se entregan directamente en la hipodermis vecina, que normalmente rodea las neuronas de los receptores táctiles. La hipodermis intenta la degradación basada en lisosomas, pero algunos contenidos no digeribles como los agregados mCherry pueden ser reextruidos por la hipodermis en el pseudocoelom lleno de fluidos del animal, del cual el mCherry puede ser tomado por células carroñeras remotas llamadas comoelocitos para almacenamiento a largo plazo (Figura 1, Figura 2)5.
Las grandes vesículas exoféreas extruidas salen de la célula rodeadas por la membrana plasmática del receptor táctil y pueden contener proteínas de enfermedad humana agregadas, mitocondrias y lisosomas. El proceso de producción de exoferas parece implicar la clasificación de especies potencialmente tóxicas (por ejemplo, un mCherry expresado propenso a la agregación se separa de proteínas solubles e inofensivas como la GFP que permanece principalmente en el soma neuronal). De esta manera, la expulsión dirigida de las entidades amenazantes se lleva a cabo por la neurona5. Un desafío de proteostasis, como el estrés inducido por la despojeación por la autofagia, la inhibición proteosoma mediada por MG132, o la expresión transgénica de proteínas de la enfermedad humana como la poliglutamina expandida asociada a la enfermedad de Huntington Q128 o el fragmento implicado por la enfermedad de AlzheimerA-1-42, puede aumentar el número de neuronas que producen que producen5.
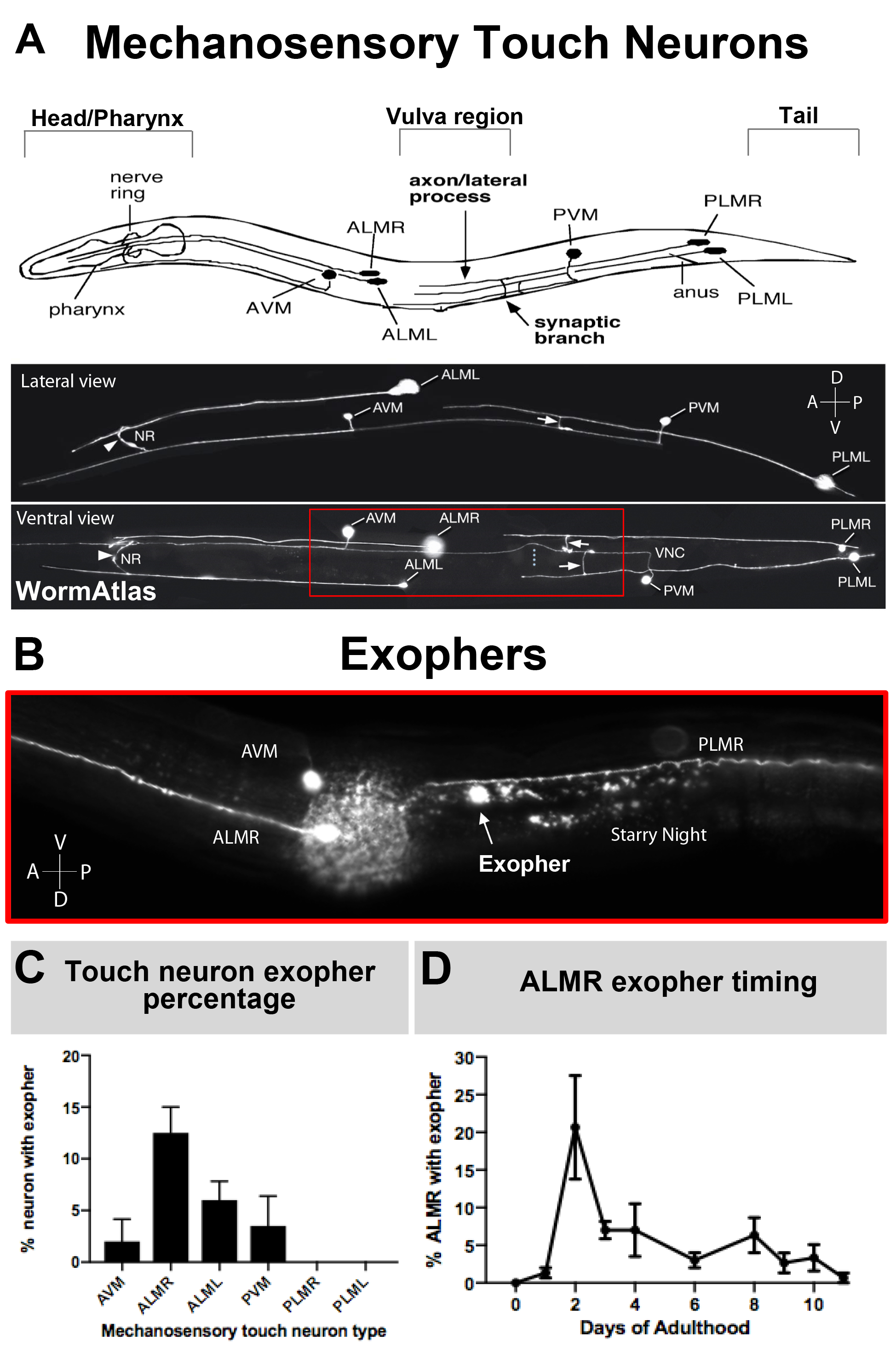
Como los exoferos han sido documentados recientemente, lo que se conoce de su biología merece una descripción. Los exoferos fueron descubiertos en, y son los más bien estudiados en, C. elegans tacto neuronas receptoras. Hay seis neuronas táctiles mecanosensoriales C. elegans que tienen cuerpos celulares distribuidos por todo el cuerpo (Figura 3A) y se llaman células de microtúbulos porque su ultraestructura cuenta con 15 microtúbulos de protofilamento distintivos. Las neuronas del receptor táctil son la malformación arteriovenosa anterior (neurona de microtúbulo ventral anterior), ALMR y ALML (neuronas de microtúbulo laterales anteriores derecha e izquierda), la PVM más central (neurona de microtúbulo ventral posterior) y la PLMR posterior y PLML (neuronas de microtúbulos laterales posteriores derecha e izquierda) en la cola. Curiosamente, las seis neuronas receptoras táctiles producen exophers a diferentes velocidades, a pesar de expresar el mismo transgén ofensivo (Figura 3C). De las seis neuronas receptoras táctiles mecanosensoriales, la neurona ALMR sufre exofergénesis más a menudo que las otras neuronas táctiles. Por lo tanto, la cuantificación de los números de exoferes de las neuronas táctiles se establece generalmente centrándose en el ALMR.
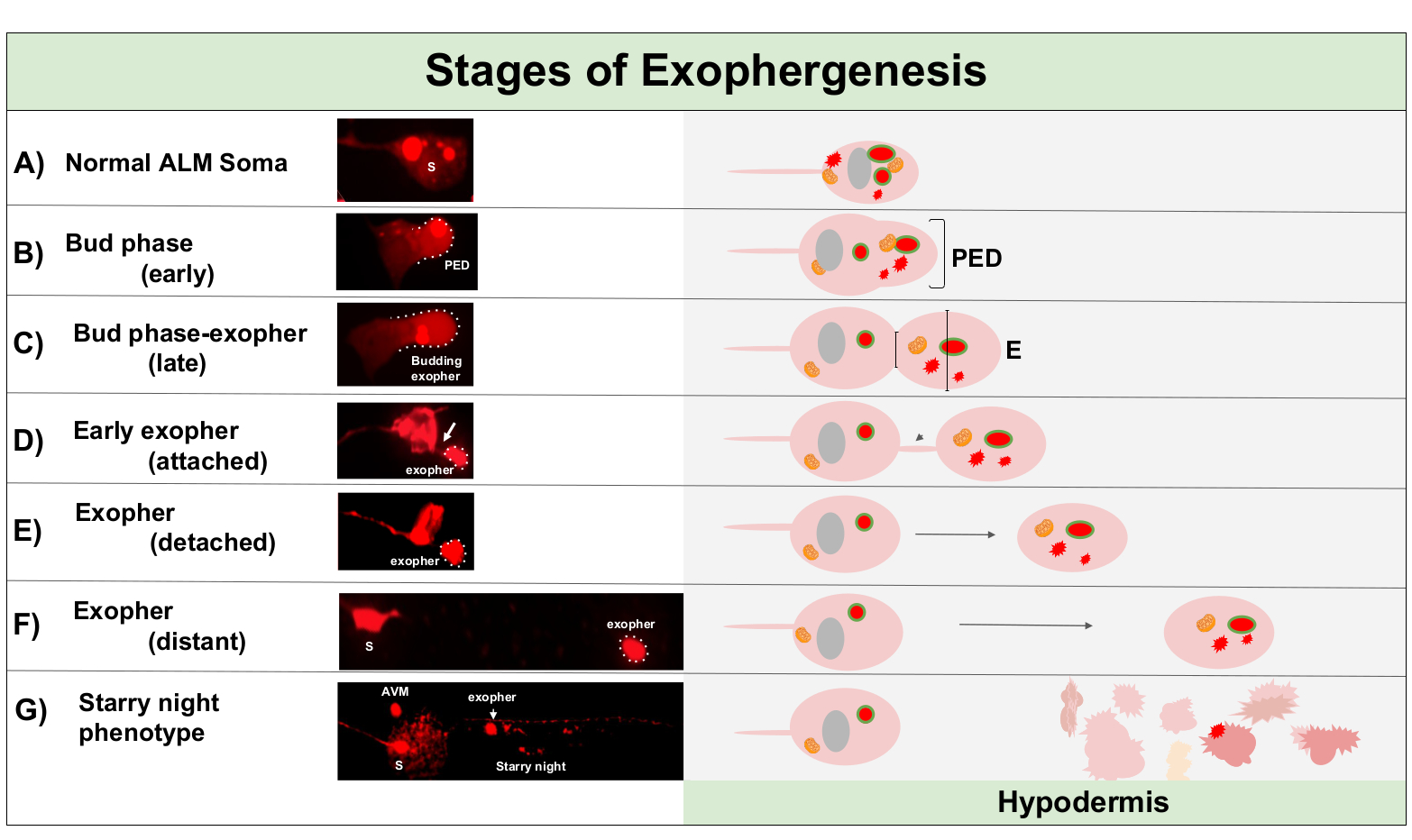
La exofergénesis es un proceso dinámico que normalmente comienza con la hinchazón del citoplasma neuronal(Figura 1A-B). El contenido celular, los orgánulos o los agregados proteicos se recogen a un lado del soma neuronal, más comúnmente hacia el extremo posterior de la neurona ALMR (lejos de la neurita que se proyecta), formando un dominio de pre-exofero (PED) (Figura 1B). La protuberancia temprana se observa a medida que el PED comienza a proyectarse hacia afuera, formando un cogollo sobresaltado reconocible. El brote tardío se define cuando el diámetro más ancho del dominio pre-exoférico es aproximadamente 1/3 más grande que el diámetro de la constricción del cuello soma-exopher (Figura 1C). Los exoferos pueden ser expulsados en casi cualquier dirección del soma, pero la mayoría de los exophers salen posteriormente del cuerpo celular y permanecen aproximadamente en el mismo plano focal que el soma de origen.
El exofer puede alejarse del soma de origen a medida que el cuello del cogollo se estrecha en un filamento delgado. Los exoferos pueden permanecer unidos al soma a través de este filamento(Figura 1D, flecha)y más tarde pueden separarse. El contenido celular como calcio, áridos y mitocondrias se puede transferir a través de este filamento al exopher5adjunto, aunque la mayor parte del material extruido se coloca en el compartimiento del exofero por el evento en ciernes masivo. Los exoferos se consideran maduros cuando no hay tubo de conexión visible o filamento delgado y el exofero está completamente separado del soma de envío (Figura 1E).
Los exoferos producidos por C. elegans tocan las neuronas inmediatamente encuentran la hipodermis, el tejido que rodea la neurona táctil. Más comúnmente, la vesícula exoférica parece viajar dentro de la hipodermis posteriormente hacia la cola, y puede estar bastante distante del soma antes de que el contenido del exoféro aparezca dirigido a la degradación (por ejemplo, la distancia puede estar a 100 m del soma(Figura 1F)). La vesícula de exoférico fluorescente se divide en muchas vesículas más pequeñas dentro de la hipodermis, tomando una apariencia conocida como "noche estrellada"(Figura 1G y Figura 2). En la etapa de la "noche estrellada", se puede observar material fluorescente punctato esparcido por el sinítcio hipodérmico en muchos puntos más pequeños de fluorescencia en comparación con el exofero solitario original. La noche estrellada puede verse punctata bajo bajo aumento y con mayor aumento, puede parecer punctato y / o en red dentro de la hipodermis. La señal fluorescente de la noche estrellada es típicamente más tenue que el exofer y la fluorescencia expresada neuronalmente (Figura 2B-C). Se cree que la dispersión de mCherry en muchas vesículas punctatas implica la maduración y fusión de fagosos con la red endosomal/lisosomal de la célula hipodérmica. Es probable que algunos materiales de exoférico se degraden en la red lisosomal hipodérmica, pero las especies residuales que son resistentes a la degradación (como los agregados mCherry) se echan de la hipodermis en el pseudocoelom, un compartimento de fluidos que puede contener desechos celulares. El material fluorescente es tomado más tarde por células carroñeras remotas llamadas coelomocícitos (Figura 2C), que pueden concentrarse, almacenarse e intentar de nuevo la degradación de mCherry.
El fenómeno de la extrusión y transferencia de agregados parece conservado a través de phyla, habiendo sido reportado en modelos genéticos como C. elegans5,6,7 y D. melanogaster8,9 así como en múltiples modelos de mamíferos. Se han notificado extrusiones similares a exoféros para las células de mamíferos10,una observación que sugiere que los mecanismos conservados podrían subyacen a la expulsión de agregados y orgánulos. Por lo tanto, la producción de exoféros puede ser un mecanismo conservado de manejo de desechos celulares que constituye una rama fundamental, pero antes no reconocida, de la proteostasis neuronal y el control de calidad mitocondrial, que, cuando está desequilibrado, podría contribuir activamente a la enfermedad neurodegenerativa. La identificación de las moléculas implicadas en la discriminación y clasificación de escombros, el transporte a una localidad subcelular distinta, la extrusión, la formación/ciencia de la conexión tubular que une el soma y el exofero tardío, y el reconocimiento de la gran vesícula extruida para la degradación remota por una célula vecina permanecen para el trabajo futuro. Los estudios en modelos de nematodos y moscas serán de vital importancia para definir mecanismos de recolección y transferencia de agregados y orgánulos, utilizando enfoques genéticos imparciales y potentes herramientas biológicas celulares ofrecidas por estos modelos para identificar moléculas participantes en contexto fisiológico.
Los primeros pasos críticos para descifrar los mecanismos operativos en biología de exoférfos implican la definición de protocolos para la cuantificación de exoferos in vivo reproducible. El modelo C. elegans ofrece una ventaja particular para tales esfuerzos, ya que el cuerpo es transparente y los exoferores se pueden observar fácilmente cuando contienen proteínas o orgánulos etiquetados fluorescentemente. Los exoferos han sido reportados para ser generados por C. elegans neuronas dopaminérgicas PDE y CEP, ASE y ASER neuronas sensoriales, y las neuronas ampancias de relleno de tinte5. Debido a que los exoferores producidos por las neuronas receptoras táctiles se caracterizan mejor, el enfoque aquí es en el uso de neuronas táctiles para el análisis de exoferos. Sin embargo, el enfoque básico se puede aplicar para medir la producción de exoféros desde cualquier célula. Se describen los protocolos para detectar y cuantificar los exophers producidos por C. elegans que expresan transgénicamente la proteína mCherry, con énfasis en las cargas que pueden ser monitoreadas y las restricciones temporales en la puntuación. Este artículo define los enfoques hacia la identificación de exoféricos in vivo y la cuantificación de las condiciones ambientales y genéticas que modulan la producción de exoferos. Los protocolos hacen hincapié en la atención crítica a las condiciones constantes de no estrés para la determinación de la producción de exoferos basales y para las comparaciones entre genotipos.
Protocolo
1. Tensiones útiles para la detección de exoferos
- Seleccione una cepa que exprese cargas fluorescentes dentro de las neuronas de C. elegans para visualizar fácilmente los exophers.
NOTA: Tabla 1 enumera las cepas que se han utilizado para visualizar los exoferos producidos en las neuronas de receptores táctiles5,11,12. En principio, cualquier célula o tipo neuronal se puede probar para la producción de exoferas mediante el uso de una célula o tejido promotor específico para impulsar la expresión de una proteína fluorescente que se agrega o se selecciona de otro modo para la extrusión. - Alternativamente, utilice un ensayo de relleno de tinte para visualizar los exophers en las neuronas de la cabeza amptidas, que están abiertas al medio ambiente y susceptibles de rellenar5,13.
2. Medios de crecimiento
- Preparar medios estándar de crecimiento de nematodos (NGM) para cultivos de cepas de acuerdo con los métodos estándar14,,15.
NOTA: La falta de alimentos, o fluoro-desoxiuridina (FuDR), utilizado comúnmente para bloquear la producción de progenie, y puede afectar dramáticamente la producción de exoferos. Mantener a la población alimentada continuamente (evitar incluso períodos cortos de agotamiento de alimentos bacterianos) y mantener a los animales a una temperatura constante.
3. La cría de animales es fundamental para una producción constante de exoferos
- Criar animales en medios consistentes y con fuentes de alimentos bacterianos consistentes. Los animales no deben quedado sin alimentos bacterianos, incluso durante cortos períodos de tiempo ya que la limitación de los alimentos puede cambiar drásticamente los niveles de producción de exoféros.
- Mantenga las recetas de los medios de comunicación y la preparación uniforme durante todo el estudio.
NOTA: El cambio de medios puede afectar a los niveles basales de producción de exoferos. Los lotes de agar pueden influir en los niveles de exoféro basal, por lo que cuando cambien los lotes de suministro, tome nota de la fecha. Tire los platos de stock después de dos semanas para asegurar alimentos bacterianos saludables y para prevenir el agar seco, que causa cambios en la osmolaridad del agar que influyen en los niveles de exoféro. - Para condiciones basales, mantenga a los animales a una temperatura constante de 20 oC. La cría de animales a temperaturas variables (incluso cambios temporales de temperatura) puede causar variaciones en el momento de la producción máxima de exoferos.
NOTA: La variabilidad de la temperatura no se limita a las condiciones de cultivo. Las variaciones de temperatura durante los experimentos o en el banco del laboratorio pueden ser impactantes. Por ejemplo, las temperaturas dentro de una sala de microscopio no deben diferir drásticamente de la incubadora de cultivo o banco de laboratorio. - No utilice intervenciones farmacológicas antialic fertilidad porque los óvulos fertilizados son críticos para la producción temprana de exoferos en adultos.
NOTA: Debe evitarse el uso de Fluoro-desoxiuridina (FuDR)16 o C2217. Al realizar experimentos con animales de por vida o de edad avanzada, las poblaciones sincronizadas por la edad deben mantenerse eliminando físicamente a los adultos de su progenie más pequeña recogiéndolos en placas frescas que se proponían con bacterias en lugar de utilizar intervenciones farmacológicas antialtilidad farmacológicas comunes. - No utilice cultivos contaminados; reiniciar experimentos en caso de compromiso biológico de la población o la placa. La contaminación bacteriana o fúngica puede inducir tensiones y cambios metabólicos en los animales y debe estar ausente de las poblaciones experimentales.
- Para maximizar los resultados reproducibles, mantenga los cultivos para al menos dos generaciones sanas, bien alimentadas y libres de contaminación a 20 oC antes de la experimentación para evitar posibles cambios epigenéticos inducidos por el medio ambiente.
4. Sincronización de edad para la puntuación de exoféroes por blanqueo, flotación de sacarosa o recolección de larvas L4
- Mantener las poblaciones experimentales en la misma edad biológica, ya que los patrones de detección de exoferos varían según la edad adulta y la comparación de animales de poblaciones mixtas en edad puede confundir resultados. Asegurar siempre la sincronización exitosa de las poblaciones animales experimentales mediante la comprobación de la morfología de la vulva "media luna blanca" en la etapa L4.
NOTA: Generalmente, la producción máxima de exoferos para las neuronas ALMR mecanosensoriales de C. elegans se produce en el día adulto 2-3(Figura 3D),medido a partir de los días después de la etapa L4. El día 1 adulto es 24 horas después de la etapa larval L4 que se distingue por la morfología vulva "media luna blanca"(Figura 5E). - Preparar las poblaciones sincronizadas de óvulos blanqueando adultos gravídidos.
- Recoger adultos gravid llenos de huevos lavando animales que crecen en un plato NGM. Para lavar, inunde la placa con 1 ml de tampón M9, pipetee hacia arriba y hacia abajo para recoger líquido con animales suspendidos y pipetear en un tubo de microcentrífuga de 1,5 ml. Pellet animales por sedimentación gravitacional o centrifugación suave con una mini centrífuga y quitar el sobrenadante.
- Añadir 150 l de NaOH de 5M y 150 l de hipoclorito sódico al 6% (blanqueador) en 1 ml en H2O y mezclar por inversión durante aproximadamente 5 minutos.
NOTA: La solución de blanqueo fresco garantiza que la cutícula animal pueda ser interrumpida para la cosecha de óvulos. El progreso en la interrupción de la cutícula se puede monitorear bajo un microscopio diseccionado; adultos deben romper y liberar óvulos en el punto en que el blanqueo debe detenerse. - Centrifugar suavemente con un tubo de minicentrífuga durante 20 s y retire el sobrenadante. Añadir 1 ml de tampón M9 y centrífuga de nuevo, dejando alrededor de 100 l en la parte superior del pellet.
- Repita los pasos 4.2.3 dos veces para eliminar los restos de solución de lejía.
- Resuspender los huevos en el volumen restante y transferirlos a una placa NGM de semilla fresca. Los adultos serán lezados, pero muchos huevos viables deben estar en la preparación.
- Preparar las poblaciones sincronizadas por huevos cronomebados.
- Elija 20 adultos gravid a una placa NGM sin semillas utilizando los protocolos de transferencia estándar14.
- Permita que los animales se arrastren libremente y pongan huevos durante 1,5 h (las cepas mutantes con tamaños bajos de cría pueden requerir la introducción de más animales adultos).
- Retire todos los animales adultos del plato recogiendo, dejando atrás la población de huevos sincronizados. Revise las placas unas horas más tarde para verificar que no se han perdido adultos viables durante la extracción de adultos.
- Preparar las poblaciones sincronizadas de huevos mediante la selección de flotación de sacarosa de los huevos.
- Recoger animales y huevos de cinco placas NGM sobre las que los animales gravid han estado poniendo huevos durante al menos 24 horas inundando platos con solución M9 con 0.1% detergente (como Tween 20 o Triton X-100) y recogiendo en un tubo de 15 ml. Pellet adultos por centrifugación suave a temperatura ambiente (2.000 x g para 30 s).
- Retirar el sobrenadante y lavar a los animales en 15 ml de M9 fresco tres veces, descartando el sobrenadante después de cada lavado, asegurándose de mantener el pellet enriquecido en animales y huevos.
- Conservar 2 ml de sobrenadante y resuspender el pellet. Añadir 2 ml de 60% de peso por sacarosa volumen.
- Centrífuga a 2000 x g durante 5 min. La solución ahora mostrará una fase superior altamente enriquecida en huevos.
- Transfiera unos 2,5 ml de la fase superior a un nuevo tubo de 15 ml y añada 10 ml de M9.
- Mezclar por inversión durante 1 min, y luego centrifugar 2000 x g durante 1 min.
- Retire el sobrenadante y lave el pellet enriquecido con huevos en M9. 10-15 l del huevo-pellet se puede distribuir a una placa NGM fresca de semilla de OP50.
NOTA: Este método prepara un gran número de huevos; no permita que los animales recolectados se quedas sin alimentos OP50 E. coli.
- Preparar poblaciones sincronizadas mediante la recogida de animales en la etapa L4 de desarrollo.
- Cultivar animales en placas NGM de semillas como se describió anteriormente.
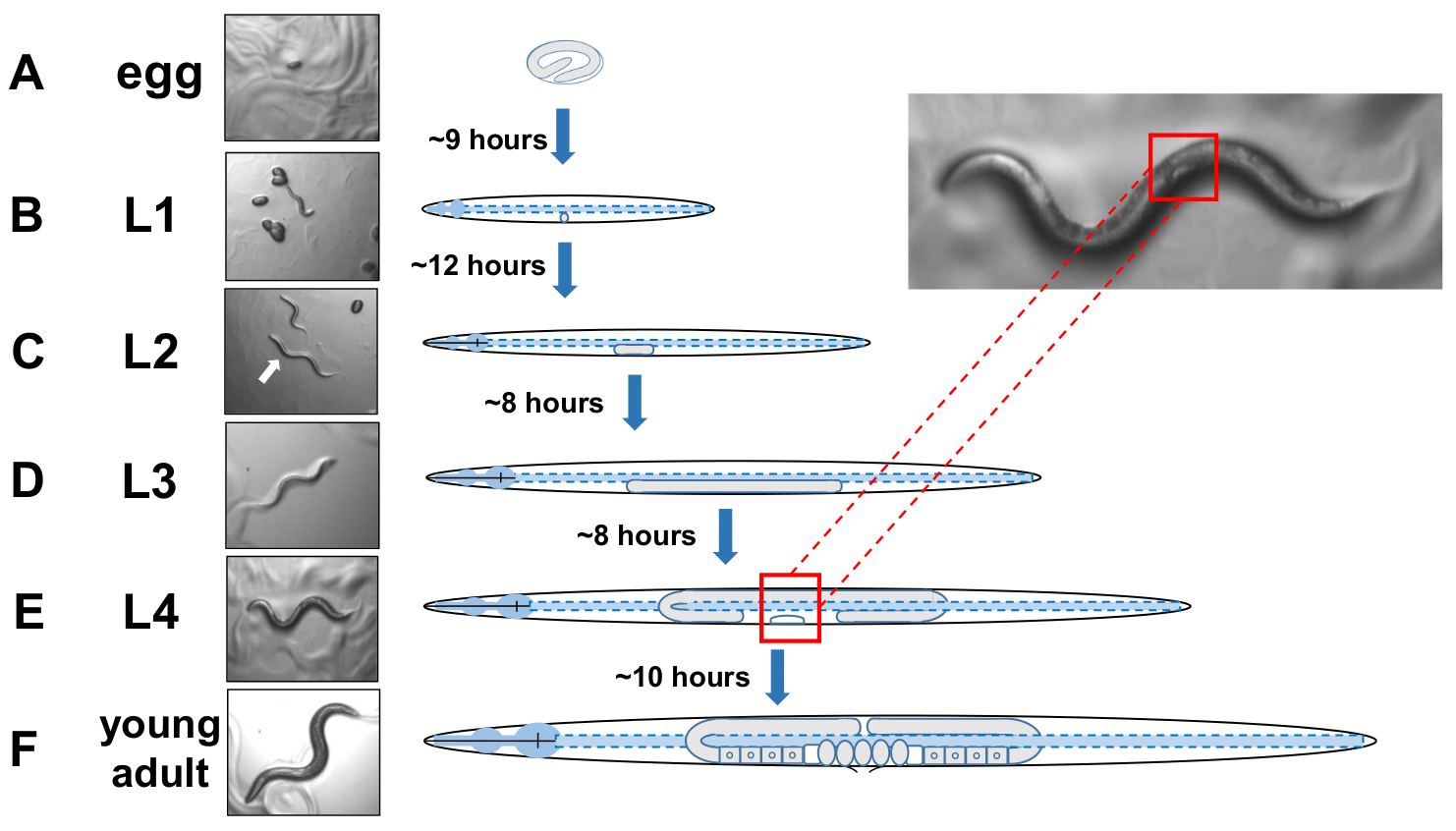
NOTA: C. elegans se desarrollan en cuatro etapas discretas. A 20 oC, un huevo recién puesto tarda unas 9 horas en eclosionar(Figura 5A). Después de la escotilla, un animal pasa a través de la etapa larval 1 (L1) a la etapa larvaria 4 (L4), con cada etapa que dura 8-12 horas entre cada molt (Figura 5A-F). Por lo tanto, un plato preparado por inoculación con huevos debe tener muchos animales L4 para recoger alrededor de 40 horas después de que se introducen los huevos. - Identificar animales de etapa L4 mediante la localización de la forma media luna blanca de la vulva en desarrollo (Figura 5E).
NOTA: Los animales en la etapa L4 son uniformes en tamaño y en pigmentación corporal. Recoger animales con la media luna blanca a una placa de crecimiento fresco para el examen de los animales escenificados. Al día siguiente (24 horas más tarde) se debe contar como adulto día 1. - Puntuar una población de animales diariamente en el adulto día 2.
NOTA: Los exophers suelen puntuar en el día 2 de la edad adulta, que es la producción máxima de exoferes en condiciones basales. Sin embargo, debido a que el pico y el momento de la exofergénesis pueden ser cambiados por los cambios ambientales o genéticos que se están estudiando, se recomienda puntuar una población de animales adultos diariamente durante cuatro días para generar el cuadro más completo(Figura 3D).
- Cultivar animales en placas NGM de semillas como se describió anteriormente.
5. Detección de exopheros usando un microscopio fluorescente
- Observe los exoferos usando un microscopio de disección pseudo-estéreo de alto aumento que está equipado para la microscopía de fluorescencia.
- Inmovilizar a los animales en placas NGM pipeteando 100-200 l de solución de 10-100 mM de levamisole/tetramisole sobre la superficie de la placa de agar NGM. Después de 2-4 minutos, los animales quedan paralizados y se pueden observar directamente en el plato de agar.
NOTA: Los tratamientos de inmovilización no son absolutamente necesarios, de tal manera que con un ojo entrenado, la identificación neuronal y la presencia de exoferas pueden ser puntuados siguiendo visualmente animales rastreados bajo el microscopio en la placa al determinar si se ha producido o no un exofero. - Observe las neuronas fluorescentes usando un aumento total de 100x para lograr la detección diseccionante del microscopio de los exophers.
NOTA: La puntuación de eventos de exofero mediante microscopía de disección permite la observación de un gran número de animales con relativa facilidad directamente en las placas de agar en las que se crían. - Imágenes en vivo y tensiones de reportero de montaje para estudios de exoféro utilizando microscopía confocal
- Utilice un microscopio confocal para la dinámica intracelular de imagen en vivo y las características de la exofergénesis.
NOTA: Las imágenes en vivo son un enfoque ventajoso para observar detalles sutiles de la producción de exoferes porque la producción de exoféros es un proceso dinámico. - Restringir el movimiento de los animales para obtener imágenes en vivo de alta resolución utilizando métodos convenientes, incluida la utilización de levamisol o tetramisol a 10-100 mM o la aplicación de microperlas de poliestireno de hidrogel (con diámetros de 15 m, 30 o 40 m)18.
- Utilice un microscopio confocal para la dinámica intracelular de imagen en vivo y las características de la exofergénesis.
- Preparación de diapositivas para microscopía compuesta y confocal
- Monte 20-50 animales en un agente inmovilizador en un portaobjetos. Las guías citologías anilladas reutilizables pintadas con anillos elevados de 13 mm de diámetro son útiles para el montaje.
- Escoge animales vivos en 5-20 l de un paralítico como 10-100 mM de levamisol o tetramisole dentro del círculo pintado o en la almohadilla de agar.
- Espere 4 minutos para la parálisis y luego cubra el portaobjetos con un cubreobjetos (recomendar No. 11/2 (0.16 – 0.19 mm) o No 2 (0.17 - 0.25 mm).
- Montaje de un pequeño número de animales
- No aplastar animales montados; cuando se observan sólo unos pocos (menos de 20) animales por portaobjetos, existe el riesgo de aplastar a algunos de los animales debido a la presión desigual del cubreobjetos. Este riesgo se puede minimizar mediante el uso de una almohadilla de agarosa de bajo porcentaje para el montaje.
- Haga una diapositiva de almohadilla de agarosa del 2-4%, y luego agregue 2-15 l de solución paralítica a la almohadilla. Tenga en cuenta que el levamisol y el tetramisol se difunden en la almohadilla, disminuyendo su concentración efectiva.
- Montar recogiendo animales en una gota de 2-15 l de solución paralítica o microperlas que descansan sobre la almohadilla de agar. Coloque el cubreobjetos en la parte superior y compruebe que los animales están intactos18.
- Preparación de la almohadilla de agar
- Para preparar almohadillas de agar al 2%, calienta el 2% de agarosa en solución M9 y microondas hasta que la agarosa esté en un estado homogéneo y fundido.
- Para lograr una almohadilla de agar de calidad suficiente, mezcla alternativa y microwaving a baja potencia durante menos de 20 segundos. Evite la inclusión de burbujas de aire dentro de la almohadilla colocando el agar hirviendo en un bloque de calentamiento y permitiendo que las burbujas se eleven a la superficie.
- Utilice una pipeta Pasteur para extraer el agar desde lo más profundo de la solución fundida por debajo de las burbujas levantadas.
- Prepare dos diapositivas grabadas y colóquelos a cada lado de un portaobjetos de microscopio de vidrio limpio sobre una superficie plana. Para hacer las correderas grabadas coloque dos tiras de 5 cm de cinta de laboratorio en cada diapositiva (Figura 6A).
- Usando una pipeta Pasteur, coloque una sola gota de agar sobre la corredera limpia del microscopio entre las diapositivas grabadas (Figura 6B).
- Con cuidado y rapidez, cubra la gota de agar fundido con una cuarta diapositiva limpia colocándola a través de las diapositivas grabadas (Figura 6Cc).
NOTA: La diapositiva debe presionar suavemente el agar fundido en un círculo aplanado de aproximadamente 0,4 mm de espesor (el grosor de la cinta) (Figura 6D). El agar debe enfriarse rápidamente. - Retire la diapositiva superior deslizándola (Figura 6E) . Las almohadillas de agar se secan rápidamente y se utilizan mejor en cuestión de minutos. Una vez que se retire la corredera superior, utilice la almohadilla de gel inmediatamente para montar animales. Evite el uso de almohadillas con burbujas de aire.
- Almacene las almohadillas de agar hasta 30 minutos entre los dos portaobjetos de vidrio. El agar seco hace que los animales se agrupen y se desecúen. Montar animales dentro de 2-15 l de solución paralítica o microperas y cubrir con cubreobjetos; pantalla de la diapositiva dentro de los 20 minutos de parálisis y montaje (Figura 6).
NOTA: Debido a que las condiciones de estrés pueden alterar las tasas de exoféro, evite los paralíticos que pueden inducir estrés oxidativo (por ejemplo: azida sódica) al realizar pruebas de detección de exoferos.
- Detección de exopheros mediante un microscopio confocal de disco giratorio
- Observe las características biológicas celulares como los orgánulos y otros contenidos con 1,4 objetivos de apertura numérica a 63x y 100x.
- Utilice un software capaz de controlar la etapa y la adquisición de imágenes utilizando la adquisición multidimensional. Los microscopios y el software de procesamiento de imágenes también deben ser adecuados para la recopilación de imágenes y datos, ya que estos pasos implican enfoques de imagen estándar.
6. Identificación de neuronas táctiles y puntuación para los exophers con animales montados
- Monte animal adulto paralizado (Figura 6).
- Identifique el plano Z deseado. Utilice campo brillante de bajo aumento (10-40x) para identificar el plano Z adecuado del animal, tomando nota del posicionamiento del animal, la orientación de la cola de la cabeza y la ubicación de la vulva , que son puntos de referencia para la identificación neuronal y del exofero posterior(Figura 3A y Figura 5E).
- Concéntrese en la señal de fluorescencia del reportero elegido. Permaneciendo en el mismo plano Z, cambie a la visualización de fluorescencia de campo ancho a 10-40x para el reportero citosólico elegido.
NOTA: En este ejemplo, la expresión fluorescente es impulsada por el promotor específico de la neurona mec-4 mecanosensorial. Las matrices de copias altas y los diferentes fluoróforos tienen variabilidad en la expresión y, por lo tanto, intensidad fluorescente variable. Ajuste si es necesario. - Desplácese dentro del eje Z para observar la profundidad del animal y la expresión fluorescente en el plano focal. Al hacerlo, confirme la orientación de la cabeza-cola; la cabeza/faringe tendrá el anillo nervioso fluorescente y en este caso, la cola contendrá 1-2 somas PLM visibles(Figura 3A).
- Identificar neuronas táctiles
- Identifique si el animal está montado en el lado izquierdo o derecho(Figura 3A).
NOTA: Teniendo en cuenta la 3 dimensiones del animal, la mejor resolución de imagen se logra en el lado más cercano a la óptica. - Identifique el soma (ALM, ALMR, AVM) observando - comience en la cabeza para identificar el anillo nervioso y los procesos neuronales laterales.
- En aumento de 10-40x, desplácese lentamente por el eje Z para identificar el proceso adjunto.
- Una vez identificado el proceso, síguelo lateralmente en la dirección posterior hacia la vulva, donde el soma será evidente, marcado por un cuerpo celular redondo al final del proceso. Una vez que se encuentra el soma neuronal más enfocado, se puede identificar mediante el uso de otros puntos de referencia neuronales de la siguiente manera:
- Utilice la malformación arteriovenosa, una neurona ventral cercana, para ayudar a asignar la orientación animal. Si la neurona AVM está en el mismo plano que la ALM, entonces el animal está descansando sobre su lado y la neurona fuera de ese plano es el ALMR. Si la neurona AVM no está en el mismo plano que la ALM en cuestión, la neurona táctil más cercana al plano focal es ALML.
- Identifique la neurona PVM, otra neurona táctil ventral situada cerca de la cola, para indicar si la neurona del tacto anterior está en el mismo plano. Si es así, la neurona táctil observada es ALML.
- Obtener una idea de la posición de otros cuerpos soma, cerca del área de interés (neuronas fluorescentes situadas a ambos lados del soma), y en todos los planos Z, incluso si no es posible conseguir la neurona más profunda establecida en un enfoque claro.
NOTA: La identificación de todos los somas de neuronas táctiles es importante porque el soma fuera de foco se puede confundir con los exophers.
- Identifique si el animal está montado en el lado izquierdo o derecho(Figura 3A).
7. Identificación y puntuación de los exophers
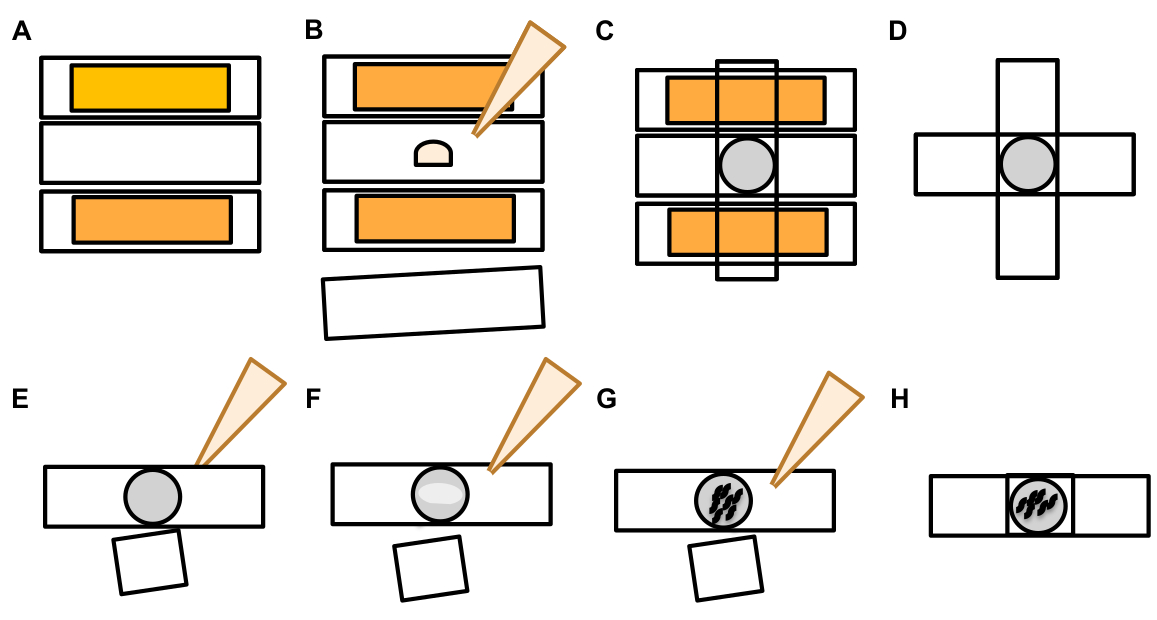
- Una vez que se encuentra una neurona táctil, inspeccione las protuberancias grandes (dominios de exoféro) lo suficientemente grandes como para ser consideradas un exofero de brotes, (alcanzando al menos 1/5o del tamaño del soma originario) ( Figurath 1C).
NOTA: El exofer medio mide alrededor de 2-8 m de diámetro, mientras que el soma promedio de un animal (ZB4065 bzIs166[Pmec-4::mCherry]) mide 6-10 m en el día 2 adultos (Figura 7B). - Si no se observa ningún dominio de brote o exoférico, inspeccione el soma neuronal en busca de un filamento delgado unido que emana del soma. Los exoferos adjuntos tienden a estar situados más cerca del soma de origen y en un plano Z similar.
NOTA: Los exoferos no siempre permanecen unidos al soma. La detección de un filamento adjunto es una indicación definitiva de que el objeto es un exofero. - Para identificar un exofero no conectado, busque el contenido de un exofer. Los exoferores pueden concentrar proteínas fluorescentes expulsadas y, por lo tanto, a menudo son más brillantes que el soma.
NOTA: El contenido de los exopheros es heterogéneo y variable. Los orgánulos celulares como los lisosomas y las mitocondrias también se pueden extruir dentro de los exoferos (Figura 4C-E). - Busque exophers no conectados en planos focales diferentes al plano en el que se encontró el soma de origen. Aunque se ha visto que los exoferos sobresalen del soma de ALM en cualquier dirección, es típico que los exophers prosime lejos del soma, en una dirección posterior del proceso neuronal.
- Compruebe si hay objetos esféricos grandes que no estén posicionados e identificados como somas neuronales. Los exoferos pueden tener forma irregular, pero suelen ser estructuras esféricas. Los los eores se degradan con el tiempo, por lo que los exoferos más viejos tienden a tener una forma más irregular.
NOTA: Los exoferes maduros o más antiguos se distinguen de la etapa de "noche estrellada" dispersa a través de la intensidad de fluorescencia más brillante de los exoferos y su forma esférica. - Investigue el fenotipo de la "noche estrellada" como evidencia de exofergénesis anterior. Los exoferos progresan en una etapa de "nocheestrellada" mientras el exofero se rompe en vesículas más pequeñas y la hipodermis circundante intenta degradar el contenido del exofer(Figura 1G,2B, 3B y 7A).
NOTA: La etapa nocturna estrellada está marcada por entidades fluorescentes fragmentadas y dispersas (a veces en red) que han perdido integridad estructural y muestra una fluorescencia tenue en comparación con las neuronas táctiles y las estructuras de exoféro. - Busque ejemplos de "múltiples eventos de exofero". Los exoferos se producen generalmente como una ocurrencia singular (1 exofum que emana de 1 soma) pero en algunas circunstancias más de un exofer puede ser liberado de un solo soma (Figura 7D).
NOTA: Los exoféricos maduros pueden degradarse en varias vesículas a medida que se degradan en la hipodermis. Distinguir si cada exoferero fue generado por un evento de exofergénesis independiente o si una división de exoféro original para crear una vesícula adicional sólo puede determinarse mediante la observación de lapso de tiempo. - Tenga en cuenta que no todas las anomalías morfológicas maduran en exofers.
- No anote un soma distendido como un exofero. Un soma extendido o puntiagudo se puede observar en ocasiones (especialmente con la edad o bajo estrés), pero una extensión sin un sitio de constricción clara no se puntuó como un exofero.
- Rechazar pequeños cogollos resueltos que no alcanzan 1/5del tamaño del soma en la cuantificación de eventos de exofer.
- No cuente los crecimientos de neurita como exoferos. Las neuritas maduras pueden extenderse dramáticamente con la edad (generalmente en la dirección opuesta del proceso neuronal) y la proteína fluorescente puede migrar al extremo distal de dichas estructuras19.
NOTA: Estos crecimientos de neurita no son exoferas, ya que tienen un patrón de desarrollo distinto a lo largo de días y semanas, no forman cogollos y no se separan (Figura 7E).
- Identifique entidades fluorescentes que no sean exophers.
NOTA: Es importante hacerse una idea de la fluorescencia de fondo para asegurar la correcta identificación de la entidad fluorescente extruida frente a la autofluorescencia.-
Expresión fluorescente transgénica vs autofluorescencia. No confunda la autofluorescencia con la expresión transgénica. La verdadera señal de exoférico no estará en el intestino o el intestino (la confirmación diciembre se puede utilizar para identificar estos tejidos) y la señal de exoférico será significativamente más brillante que la autofluorescencia de fondo.
NOTA: La autofluorescencia es causada por la pigmentación fluorescente intestinal del gránulo intestinal intestinal y se acumula con la edad. Es heterogéneo, especialmente visto con diferentes longitudes de onda. - Señal de embriones. No confunda la señal embrionaria con la exofergénesis. Confirme las sospechas de señal embrionaria cambiando de la fluorescencia a la iluminación de campo brillante y comprobando las asociaciones de señal con los huevos en el útero.
-
Cuerpos de soma fuera de avión o cercanos. Evite confundir un soma fuera del avión con un exofero identificando todos los cuerpos de soma cercanos, incluso somas fuera de foco al comienzo de la observación.
NOTA: Si califica para los exophers de ALMR, identifique y tenga en cuenta la ubicación de los somas AVM y ALMR. Más detalles sobre la identificación del cuerpo soma se describen en la Figura 3A.
-
Expresión fluorescente transgénica vs autofluorescencia. No confunda la autofluorescencia con la expresión transgénica. La verdadera señal de exoférico no estará en el intestino o el intestino (la confirmación diciembre se puede utilizar para identificar estos tejidos) y la señal de exoférico será significativamente más brillante que la autofluorescencia de fondo.
8. Puntuación y estadísticas
- Puntuación de los exophers como binario (sí, hay un exofero/no, no hay un exofer).
- Considere la detección de exoferos como un "evento de exofero" para una neurona dada. Un evento de exofer puede constituir la observación de un solo exopher cerca de un soma o de múltiples exophers.
NOTA: Para cuantificar el número de eventos individuales de exofergénesis, utilice la observación de lapso de tiempo. - Recuento de eventos de exofero por una célula identificada en particular porque diferentes células no producen exoferos a la misma velocidad (véase, por ejemplo, la Figura 3C). Las neuronas ALMR producen los exoferes más basales en las cepas descritas en este documento y, por lo tanto, a menudo esta es la célula seleccionada para la cuantificación de exofericos a partir de neuronas receptoras táctiles.
- Para las estadísticas, en general, realizar al menos 3 ensayos biológicos, de al menos 30 animales puntuados por ensayo con el número correspondiente de observaciones necesarias para el análisis de la interrupción.
- Para múltiples ensayos con uno o dos mutantes/tratamientos en comparación con el control, la prueba Cochran-Mantel-Haenszel es apropiada para determinar los valores p.
- Para los ensayos que involucran más de dos mutantes de tratamientos en comparación con el control, también es apropiado utilizar un análisis de regresión logística binaria para evaluar la significancia para cualquier número de predictores categóricos.
Resultados
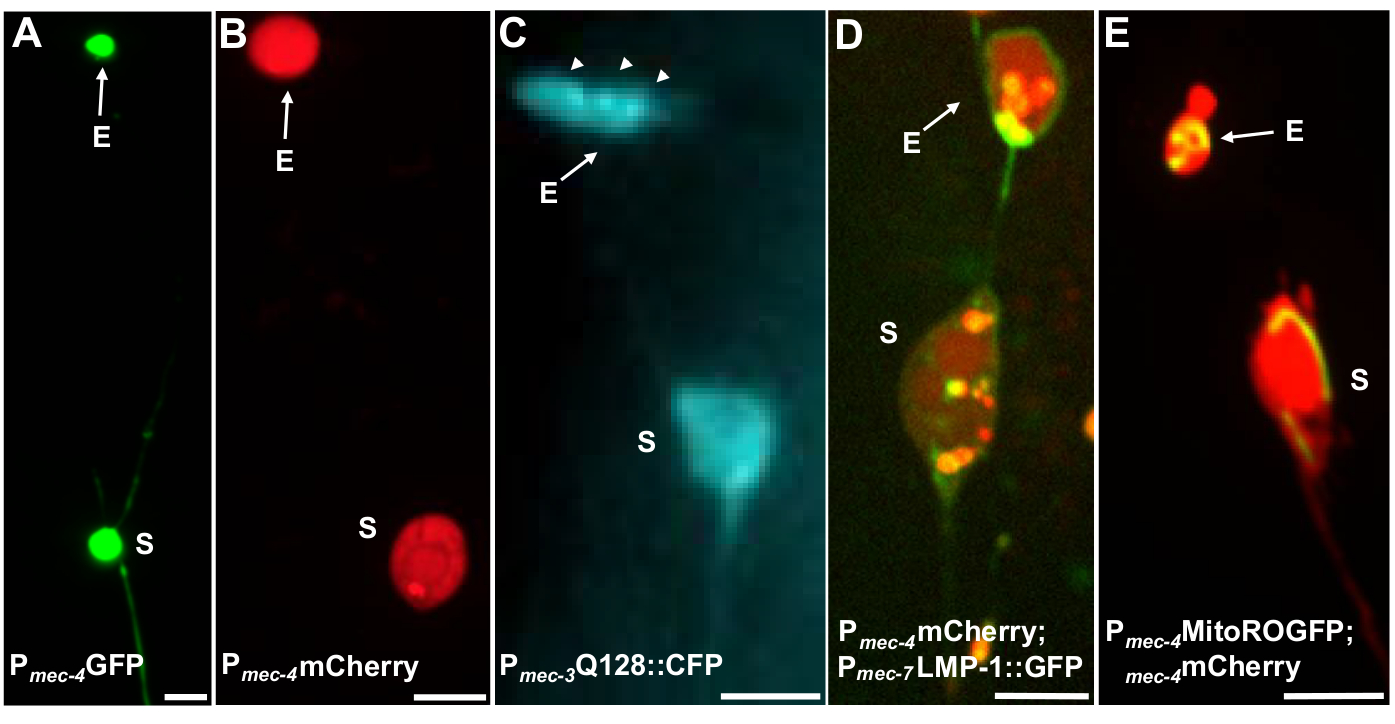
Se pueden utilizar varios reporteros fluorescentes para medir los exophers. Los exoferes de neuronas táctiles se visualizan fácilmente in vivo a través del etiquetado fluorescente de proteínas que se pueden seleccionar para la extrusión, mediante el etiquetado de orgánulos que se pueden extruir o mediante el etiquetado de membranas celulares. La Tabla 1 identifica los reporteros fluorescentes expresados en neuronas táctiles que se han utilizado para monitorear los exophers, con ejemplos representativos incluidos en la Figura 4. Las cargas que se sabe que se extruyen en los exoféros incluyen una fusión del dominio N-terminal de la huntingtina humana a la poliglutamina expandida (Q128) (Figura 4B), los lisosomas que están etiquetados con proteína de membrana asociada a lisosomal (LMP-1) (Figura 4C), y mitocondrias etiquetadas con GFP localizado en matriz (Figura 4D). La GFP citoplasmática no se expulsa enérgicamente y se retiene preferentemente en el soma5,aunque la GFP puede visualizar débilmente los exferores(Figura 4A). Cuando la GFP se fusiona con proteínas que se expulsan, esta etiqueta se puede utilizar para visualizar los exophers. Un punto importante es que al etiquetar diferentes proteínas, se puede abordar una amplia gama de preguntas sobre la expulsión de cargas y orgánulos específicos, así como sobre las proteínas y membranas que componen los exophers.
Una configuración de pseudo-estereomicroscopio es una herramienta eficaz para ver los exopheros en animales sobre placas de agar. Esta configuración es un híbrido de tecnología compuesta y estereoscópica que incluye óptica de alta apertura numérica en cada aumento, tecnología pseudo-estéreo (objetivos discretos sobre una base estereoscópica) y un interruptor de operación de zoom para ver los aumentos intermedios a los objetivos instalados. Un microscopio como este debe estar equipado con 10 oculares y objetivos lo suficientemente potentes como para observar la morfología neuronal y la producción de exoféricos para la puntuación de alto rendimiento (objetivo 2x utilizado para escanear / recoger, objetivo 10x utilizado para la identificación y puntuación).
Mientras que las capacidades de aumento de los estereomicroscopios estándar suelen tener una resolución lo suficientemente alta como para ver la red de neuronas táctiles que expresan proteínas fluorescentes, los microscopios diseccionantes estándar no son suficientes para observar los detalles subcelulares de los exferores como las conexiones tubulares de soma a la exofería. Estas observaciones requieren una microscopía confocal (véase la Tabla de Materiales para más detalles del equipo).
Los estudios de cuantificación de exoféros requieren controles estrictos para eliminar las tensiones experimentales. Se requiere un mantenimiento atento de las condiciones de crecimiento constantes para la producción de exoferas reproducibles. Más específicamente, la producción de exoferes responde al estrés de tal manera que la alimentación constante, la temperatura constante y el crecimiento libre de contaminación a través de generaciones son críticos para la reproducibilidad. En condiciones de crecimiento basal con alta expresión neuronal de mCherry, la producción de exoféricos es relativamente baja (5-25% de los ALMR producen exophers), pero algunas tensiones, incluyendo el estrés osmótico y oxidativo, pueden aumentar las tasas de exofero. Mientras que la expresión mCherry se puede considerar como estrés, un corolario de la sensibilidad al estrés de los niveles de exoféro es que, si se controla adecuadamente, la introducción experimental de estrés puede ser una estrategia para inducir y observar más fácilmente la exofergénesis.
Tiempo y niveles de producción de exoferes anticipados. Los exoferores están prácticamente ausentes durante el desarrollo larvario. El período de producción máxima de exoferos en la vida adulta joven parece estar muy restringido durante los días adultos 1-4, siendo más comúnmente evidente en el día 2 o 3 de adultos. Debido a que el pico puede desplazarse un poco hacia adelante o hacia atrás, la evaluación más completa de un perfil de producción de exoféro es puntuar múltiples ensayos diarios durante los días adultos 1-4. En general, un ALMR produce un exofero importante, con la vesícula persistente durante al menos 24 horas. El exofer se puede producir con bastante rapidez (en el orden de minutos en su másrápido). Más comúnmente, sólo un exofero importante se produce por neurona en la vida adulta temprana, pero la producción de múltiples exophers es posible.
En general, la producción de exoféro por ALMRs que expresa mCherry en condiciones basales oscila entre el 5-25% de los ALMR examinados dentro del marco óptimo del día 2-3 del adulto(Figura 3D). Las crisis de proteostasis5,así como la exposición a otras tensiones pueden modular el nivel de exoféro. El estrés o las perturbaciones genéticas pueden aumentar la producción de exoferas a tasas de detección de hasta el 90% de las neuronas ALMR que producen extrusiones de exofer.
ARNi basado en la alimentación para probar roles de genes específicos en la exofergénesis. El nematodo C. elegans es comúnmente sometido a ARNi derribado por la alimentación de animales transformados E. coli cepa HT115 que expresan un ARN de doble cadena (dsRNA) dirigido a un gen de interés20. Las bacterias HT115 se pueden utilizar al puntuar para los exoferos en la alimentación de RNAi5. Mientras que las transcripciones en la mayoría de los tejidos pueden ser objetivo de ARNi utilizando esta técnica, las neuronas son más refractarias. La sensibilidad al ARNi se puede calibrar utilizando animales que expresan el transportador transgénico de ARN-1 bajo un promotor específico de la neurona. De esta manera el tejido neuronal se puede sensibilizar a RNAi21.
El derribo específico del tejido de un gen de interés se puede lograr expresando un componente del metabolismo endógeno del ARNi dentro de un mutante que es deficiente en ese componente. Por ejemplo: la proteína Argonaute RDE-1 puede expresarse específicamente en las neuronas de los animales mutantes rde-1 para lograr la eliminación de un gen de interés sólo en las neuronas cuando los animales están expuestos a una intervención de ARNi dirigida a ese gen.
Utilizando los protocolos estándar de NEMAtodo RNAi20,22, la exposición de los padres en la etapa L4 al ARNi y permitiendo que su progenie desarrolle bacterias HT115 transformadas consumadas hasta la edad adulta genera el fuerte derribo genético, pero estar atento a posibles retrasos en el desarrollo inducidos por el ARNi, ya que los animales experimentales pueden crecer de manera diferente que un control vectorial vacío. Es importante incluir siempre el control vectorial vacío para la comparación de controles negativos. Las bacterias HT115 se pueden utilizar al puntuar para los exophers en la alimentación de ARNi. Sin embargo, tenga en cuenta que algunos genes son eficaces para cambiar las tasas de exofergénesis incluso durante períodos más cortos de exposición a ARNi5. Si la focalización de ciertos genes conduce a un fracaso del desarrollo, evite exponer a los animales a un derribo de por vida, los animales simplemente pueden ser recogidos en la etapa L4 en placas de ARNi para la exposición de L4 a D2 o D3 adulto.
| Nombre de la cepa | Genotipo | Descripción | Porcentaje de exoferos | Referencia |
| SK4005 | zdIs5[Pmec-4GFP] | Expresión citosólica de GFP en neuronas táctiles. | 1-8% ALM | Figura 4A, Melentijevic 2017 |
| ZB4065 | bzIs166[Pmec-4::mCherry] | La sobreexpresión de mCherry (bzIs166) en las neuronas táctiles, produce tanto señal citosólica como agregados mCherry. bzIs166 es un inductor de exofer. Los agregados mCherry son predictores de exofergénesis y se extruyen preferentemente en exophers. | 3-20% ALM (condiciones normales). 20-80% ALM (condiciones de ayuno). | Figura 4B, Melentijevic 2017 |
| ZB4067 | bzIs167[Pmec-4mitogfp Pmec-4mCherry4]; igIs1[Pmec-7YFP Pmec-3htt57Q128::cfp lin-15+]; | YFP etiquetas citosómicamente las neuronas táctiles mec-7. Co-expresado Q128::CFP agrega e induce los exophers. La PPC silencia preferentemente. | 25% | Figura 4C, Meletijevic 2017 |
| ZB4509 | bzIs166[Pmec-4mCherry]; bzIs168[Pmec-7LMP-1::GFP] | bzIs168 LMP-1::GFP etiqueta membranas plasmáticas y membranas lisosomales. bzIs168 se puede utilizar para identificar membranas neuronales, exoferas (ya que están unidas a membranas) y estructuras de membrana lisosomal. | 3-20% ALM | Figura 4D, Melentijevic 2017 |
| ZB4528 | bzIs166[Pmec-4mCherry]; zhsEx17 [Pmec-4mitoLS::ROGFP] | Allele zhsEx17 es un reportero mitocondrialmente localizado que cambia su longitud de onda de excitación máxima de 405 nm (oxidado) a 476 nm (reducido) según el entorno oxidativo local. Se expresa en las neuronas táctiles y se puede utilizar por sí mismo para identificar mitocondrias en neuronas táctiles y en mito-exophers. | 3-20% PROteo-exofer ALM. % alm mito-exopher cantidad en curso. | Figura 4E, Melentijevic 2017, Cannon 2008, Ghose 2013 |
Tabla 1. Cepas que se han utilizado para la visualización de neuronas táctiles, neuro-exophers táctiles, y contenido de exopher.

Figura 1: Etapas de la exofergénesis. El proceso de hacer y expulsar un exofero se llama "exofer-génesis". El proceso dinámico de formación de exoféros puede tardar de varios minutos a varias horas. Se muestran ejemplos de morfología soma y exoférico en pasos específicos durante el proceso de exofergénesis dinámica en una cepa de producción de alto exofero, ZB4065 bzIs166[Pmec-4mCherry]. Todas las imágenes son del día 2 neuronas alM adultas tomadas con un objetivo 100x. (A) Soma normal. Neurona táctil mecanosensorial adulta ALM que expresa transgénicamente Pmec-4mCherry. La morfología soma representada es típica de las neuronas adultas jóvenes en esta cepa, con concentraciones de mCherry en el citoplasma. (B) Fase temprana de los cogollos. El primer paso observable de exofergénesis implica la polarización del material citoplasmático seleccionado hasta el borde de la membrana soma. Este paso a menudo va acompañado de una expansión o hinchazón del soma. En el caso de las neuronas táctiles, el dominio pre-exoféro (PED) se extiende a la hipodermis circundante (no visible aquí). Tenga en cuenta la mayor concentración de material mCherry en el dominio de brotes tempranos. (C) Fase tardía del brote. Tras una mayor polarización celular y una expansión del dominio pre-exopher, una constricción entre el soma y el exofer (flecha) se hace evidente. Este evento indica la transición a la fase de brotes tardíos. Aunque en la etapa de brote tardío la célula exhibe un sitio de constricción clara y dominios soma y exopher separados, todavía no se pellizca completamente del soma; el exofer en ciernes puede ser unido por un tallo grueso (flecha). El dominio en ciernes se considera un exópher temprano cuando el diámetro del dominio del exofero en cuestión es aproximadamente 1/3 mayor que el diámetro del sitio de construcción / tallo. (D) Fase de exofero temprano. Los primeros exoféros se pueden conectar mediante un tallo del soma que sale, el diámetro de esta conexión puede adelgazar a medida que el exofer se aleja del soma. El material citoplasmático se puede transferir del soma al exofero a través de este tubo, aunque la mayoría del material se carga durante el proceso de entrada. Los exoferos pueden separarse del soma como se representa en (E), los exoferos separados se consideran exoferos maduros (F). El exofer maduro puede transitar a través del tejido hipodérmico circundante, alejándose del soma que sale. (G) El desglose del exofer etiquetado con mCherry en vesículas más pequeñas dentro de la hipodermis da como resultado una apariencia punctata dispersa del material mCherry, muy probablemente al entrar en la red endolysosomal hipodérmica. La señal punctata dispersa se llama la fase de "noche estrellada". La degradación de algunos contenidos de exoférceos es probablemente lograda por lisosomas hipodérmicos, pero algunos materiales no están completamente degradados y a menudo son reextruidos por la hipodermis en el pseudocoelom. El tránsito post-exofergénesis mCherry se describe con más detalle en la Figura 2. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 2: mCherry extruido de neuronas táctiles en exophers involucra la red lisomal hipodérmica circundante, pero más tarde se puede extruir en el pseudocoelom donde los coelomocícitos pueden almacenar / degradar el mCherry. (A) Resumen de dibujos animados de cómo mCherry extruido en exophers transita el cuerpo después de la expulsión por las neuronas. Durante la exofergénesis, los contenidos celulares seleccionados como mCherry se localizan y se alejan del soma neuronal de envío en una vesícula independiente rodeada por las membranas plasmáticas neuronales e hipodérmicas. Dado que las neuronas táctiles están incrustadas en el tejido hipodérmico, a medida que el dominio exoférico brota hacia afuera se mueve más hacia la hipodermis. El exofer puede transitar la hipodermis, y después de horas a días, el contenido del exoféro puede fragmentarse dentro de la red endolysosomal de la hipodermis. El mCherry puede aparecer como puncta disperso a lo largo de la hipodermis, un escenario llamado "noche estrellada". Después de unos días, algunos de los mCherry pueden pasar de la hipodermis hacia el pseudocoelom circundante, donde las células carroñeras llamadas coelomocícitos pueden tener acceso a, y tomar, mCherry que se pueden almacenar. (B) Ejemplo de la aparición de la noche estrellada mCherry vesicles. Imagen de un soma alM etiquetado con mCherry con grandes fragmentos de exoféro y vesículas nocturnas estrelladas. La cepa es ZB4065 bzIs166[Pmec-4mCherry]. (C) Ejemplo de concentración de mCherry en coelomocitos distantes. Vista lateral de un animal adulto día 10 de cepa ZB4065 bzIs166[Pmec-4mCherry] mostrando mCherry concentrado en coelomocitos (flechas). Algunas vesículas nocturnas estrelladas también son evidentes. En general, la concentración de coelomocícito se hace evidente después de aproximadamente el día adulto 5 de la vida. (B inferior) Reproducción de dibujos animados de (B), con neuronas táctiles y procesos descritos en rojo, al igual que los fragmentos de exofer más brillantes; las vesículas pequeñas dispersas de diferentes profundidades Z se muestran en rosa más claro. (C inferior) Versión de dibujos animados de la imagen de (C), mostrando el proceso neuronal en rojo, noche estrellada en rosa y coelomocitos en verde. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 3: Las neuronas táctiles mecanosensoriales producen exferores a diferentes niveles con un perfil temporal preciso. (A) (Arriba) Representación de dibujos animados de neuronas táctiles mecanosensoriales en relación espacial con puntos de referencia anatómicos clave de C. elegans incluyendo la faringencia de bombeo y el anillo nervioso densa de neuronas en la cabeza del animal, la vulva en el cuerpo medio, y la cola cónica. (Abajo) Neuronas táctiles con etiqueta fluorescente que expresan GFP como se ve desde la parte superior e izquierda (imágenes adaptadas de WormAtlas). El cuadro rojo representa el área donde se encuentran normalmente los exoferos de ALM. (B) Vista de gran aumento de la región del cuerpo medio en la que se producen los exoferos derivados de ALM en una cepa que expresa [Pmec-4mCherry]. Se representan las neuronas AVM y ALMR, y se muestra que es un exofero ALMR junto con la noche estrellada mCherry. Las neuronas ALMR producen más fácilmente exophers. (C) Las neuronas táctiles mecanosensoriales ALMR producen más fácilmente exophers en comparación con otras neuronas táctiles en hermafroditas en condiciones basales. Se indica la producción de exofer de neuronas táctiles mecanosensoriales en el día 2 adulto, como se puntó para las neuronas receptoras táctiles individuales está indicada. Tensión: ZB4065 bzIs166[Pmec-4mCherry], N>150, las barras de error son sem. (D) LAS neuronas táctiles ALMR producen más exophers durante los días 2 y 3 de la edad adulta en comparación con la etapa adolescente L4 o con animales en edad avanzada. Strain: ZB4065 bzIs166[Pmec-4mCherry], N>150, barras de error son SEM. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 4: Ejemplos de algunos reporteros fluorescentes que etiquetan el contenido de exopher. Una manera sencilla de observar los exopheros es creando animales transgénicos que expresan fluoróforos de promotores neuronales. Los fluoróforos permiten la visualización del exofero y la expresión transgénica induce la agregación y/o proteostresa que aumenta la exofergénesis. Los exoferos producidos por las neuronas ampaquinas también se pueden observar en condiciones nativas, utilizando el relleno de tinte para la visualización. Se muestran ejemplos de cepas comunes que se pueden utilizar para observar exoferos, (E) exopher, (S) soma. (A) Soma y exopher de un ALM de un adulto de cepa SK4005 zdIs5[Pmec-4GFP],100x objetivo utilizado para la fotografía, barra de escala 3m. En esta cepa, se miden los exophers que incluyen GFP soluble, pero la producción de exoféros ocurre con poca frecuencia. La fusión de la GFP a proteínas que se pueden extruir preferentemente en los exoféros en otros estudios confirma que las fusiones GFP se pueden detectar en los exoferosmaduros. (B) ALM soma y exopher de un adulto de cepa ZB4065 bzIs166[Pmec-4mCherry], que expresa mCherry e induce la producción de exofero de neuronas táctiles. Objetivo 100x utilizado para la fotografía, barra de escala de 5 m. (C) ALM soma y exopher de un adulto de cepa ZB4067 bzIs167[Pmec-4mitogfp Pmec-4mCherry4]; igIs1[Pmec-7YFP Pmec-3htt57Q128::cfp lin-15+]; canal azul selectivo utilizado para la imagen de htt57Q128::CFP. El exofer contiene htt57Q128::CFP agregados (flechas), que aparecen más concentrados en el exofer que en el soma. Objetivo 40x utilizado para la fotografía, barra de escala de 5 m. (D-E) Los exoferos pueden contener orgánulos y etiquetado específico de orgánulos con proteínas fluorescentes que permite la monitorización de la extrusión de organela. (D) Etiqueta de membrana lisosomal LMP-1::GFP describe la membrana soma y exoférico y etiqueta las membranas plasmáticas débilmente (la localización de la membrana plasmática es un paso de tráfico en el camino hacia la orientación lisosomal) y etiqueta fuertemente los orgánulos lisosomales. Se muestra un soma ALM adulto co-expresando Pmec-4mCherry y el Pmec-7LMP-1::GFP que se localiza en membranas y lisosomas. El soma tiene un exofero unido con otras extrusiones más pequeñas que probablemente sean fragmentos de exoféro (flechas). Las estructuras positivas de GFP se incluyen en el soma y están presentes en el gran exoféro, cepa: ZB4509 bzIs166[Pmec-4mCherry]; bzIs168[Pmec-7LMP-1::GFP]. Objetivo 100x utilizado para la fotografía, barra de escala de 5 m. E) Se puede utilizar un marcador de GFP mitocondrial para identificar mitocondrias en soma y exophers. Se muestra un soma ALM adulto que expresa Pmec-4mCherry y mito::ROGFP, que se localiza en la matriz mitocondrial. mito::ROGFP expresado solo, sin el mCherry, también se puede utilizar fácilmente para identificar las neuronas y la puntuación de los exopheros que incluyen mitocondrias. Tensión: ZB4528 bzIs166[Pmec-4mCherry]; zhsEx17 [Pmec-4mitoLS::ROGFP]. Objetivo 100x utilizado para la fotografía; barra de escala de 5 m. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 5: Ciclo de desarrollo de C. elegans e identificación L4. (A) A 20 oC un huevo tarda aproximadamente 9 horas en eclosionar una vez puesto por la madre. (B) Un animal recién nacido se encuentra en la etapa larval 1 (L1) y se muda en una larva L2 después de 12 horas. (C) Los animales permanecen en las etapas larvales L2 y L3(D)durante unas 8 horas cada una. (E) Los animales adolescentes se consideran la cuarta etapa larval (L4) y están marcados por una es vulva en desarrollo visible que aparece como una media luna blanca cerca del cuerpo medio. La presencia de esto, mientras que la media luna permite una fácil identificación y recolección de animales escenificados L4 para establecer cultivos sincronizados que más tarde facilitan la puntuación de los exophers. Los animales permanecen en la etapa L4 durante unas 10 horas antes de su muda final en adultos gravid, F) identificada por el desarrollo de óvulos, espermatozoides visibles, y el inicio de la puesta de óvulos. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 6: Preparación de la almohadilla de agar deslizante del microscopio. (A) Prepare dos diapositivas con una sola tira de cinta de laboratorio colocada longitudinalmente en la parte superior. Coloque una diapositiva de microscopio sin cintas en el medio como se muestra en la imagen. B) Coloque una gota de agarosa fundida en la parte superior de la diapositiva. (C) Coloque una diapositiva limpia suavemente en la parte superior de la gota, presionando la agarosa en una almohadilla circular desinflada. (D) Retire las diapositivas grabadas, que actúan para lograr un aplanamiento uniforme del agar que se necesita para crear una almohadilla uniforme. (E) Retire la corredera superior una vez que la almohadilla de agarosa se haya secado. (F) Pipetear una solución paralítica (levamisole o tetramisole) encima de la almohadilla de agar. (G) Escoja animales adecuadamente escenificados en el paralítico. (H) Cubrir suavemente los animales con un cubreobjetos y asegurar que los animales estén vivos. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 7: Caracteres de los criterios de identificación de los exophers y exopher. (A) Criterios generales que identifican un exofero. (B) Comparaciones de diámetros entre el soma emisor y el exofero extruido, medidos en m. Somas DE ALM adultos, N-35, cepa: ZB4065 bzIs166[Pmec-4mCherry] - 6,53 m de tamaño medio de soma y 3,83 m de tamaño medio de exofer. (C) Definición de criterios para diferenciar entre un dominio de exoféro y un exofero en ciernes. (D) Más comúnmente, las neuronas individuales hacen un gran exofer, que más tarde se divide o fragmentos como la hipodermis intenta degradar su contenido. Aún así, se pueden observar múltiples exophers junto a una neurona táctil que podrían derivar de múltiples eventos de exoferos de una neurona o alternativamente, los exophers también pueden brotar o fragmentarse a sí mismos. El origen de varias entidades similares a exoferas solo se puede determinar mediante microscopía de lapso de tiempo. La parte superior representa un soma de neurona táctil ALMR con un solo exofero distante. La parte inferior representa un soma de neurona táctil ALMR con múltiples extrusiones similares a exoferas. (E) Características morfológicas comunes en somas de neuronas táctiles ALM adultos que pueden confundirse con eventos de exofer. Arriba a la izquierda - Un soma ALM distendido, sin dominio de exoférico claro o sitio de constricción. Media superior - Las neuronas pueden tener pequeñas protuberancias extracelulares que pueden ser análogas a los exóphers, pero no cumplen con los criterios de requisito de tamaño para ser considerado un exofero. Arriba a la derecha – Con la edad, las neuronas táctiles pueden desarrollar crecimientos a lo largo de su neurita menor. A menudo, el material de mCherry se puede recolectar en la punta del crecimiento de la neurita. Esto no se anota como un exofero si el mCherry recogido no cumple con los requisitos de tamaño de exopher-to-soma. El fondo representa las neuronas alM adultas que tienen criterios definitorios para un dominio de exoféro o un exofer. Botom izquierda - ALM soma que tiene un dominio exopher prominente que incluye selectivamente citosol mCherry y mCherry etiquetado agregados. El sitio de constricción de dominio exopher está marcado por flechas y cumple con los criterios de tamaño (al menos 1/5del tamaño del soma). El diámetro más grande del dominio del exofer es casi 1/3 más grande que el diámetro del sitio de constricción, cumpliendo con los criterios para un evento de exofer. Medio inferior - ALM soma que tiene un exofero en ciernes prominente que cumple con los criterios de tamaño. Hay un sitio de constricción clara. Abajo a la derecha - ALM soma que tiene un exofer relleno de mCherry adjunto que cumple con los requisitos de tamaño de exopher. El exofer está unido por un filamento de conexión delgado. Todas las imágenes son de tensión ZB4065 bzIs166[Pmec-4mCherry]. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Discusión
La caracterización de los mecanismos moleculares in vivo de eliminación de agregados y orgánulos en forma de grandes exoferos está en su infancia. Quedan por abordar las cuestiones relativas a la designación de cargas para la expulsión, la recogida polarizada de estas cargas dentro de la célula, la regulación de la decisión de generar exophers, la maquinaria que media las extrusiones y la interacción de los exoferos con la maquinaria degradante en una célula vecina. Además, la visualización in vivo de conexiones tubulares que pueden pasar materiales biológicos que incluyen calcio, agregados y mitocondrias es interesante y su poco estudiada biología por derecho propio. Las preguntas sobre por qué ciertas células son más propensas a la producción de exoferos que otras también están sin resolverse, pero pueden comenzar a ser diseccionadas genéticamente con los enfoques descritos en este protocolo.
En este protocolo se describen en detalle los enfoques para lograr la puntuación reproducible de la producción de exoferas, con atención a distinguir los exophers de los somas celulares cercanos, el momento de los análisis para capturar el pico de producción de exoferos, y el estricto control de las condiciones de crecimiento para eliminar las tensiones no deseadas que pueden modular los niveles de exofero. Tanto la distinción del gran exopher temprano, o la dispersión de la "noche estrellada" en la hipodermis circundante puede ser cuantificada como evidencia de la producción de exoferos. Dicho esto, las neuronas que expresan mCherry en condiciones basales se asocian más a menudo con 5-25% de las neuronas de un tipo específico que produce un exofero. La introducción controlada de condiciones de estrés podría aplicarse para aumentar la producción de exoferos hasta la detección hasta el 90% de las neuronas que producen extrusiones, particularmente útiles para pantallas genéticas o farmacológicas para modificadores.
En la enfermedad neurodegenerativa humana, grandes agregados pueden transferirse de las neuronas enfermas a las células vecinas para promover la propagación de la patología. El mecanismo de exoférico podría transpirar a través de un mecanismo conservado utilizado para la extrusión agregada a través de phyla. Definir las moléculas in vivo que mejoran la eficiencia de este proceso (considerado control de proteostasis más eficaz) o bloquearlo podría ser aprovechado para influir en el diseño de nuevas estrategias para combatir múltiples enfermedades neurodegenerativas. Como tal, el protocolo descrito aquí podría utilizarse para pantallas de mutagénesis genética clásica, pantallas de ARNi en todo el genoma que derriban sistemáticamente genes para identificar potenciadores y supresores, o para estudios de intervención de fármacos que identifican modificadores farmacológicos candidatos de este proceso. El enfoque es sencillo, aunque algo laborioso. Los exoferores son tan grandes que se pueden ver con un microscopio diseccionado de alto aumento. Aún así, las neuronas C. elegans son relativamente pequeñas y mirando sus orgánulos o sus membranas requieren imágenes confocales de mayor potencia y es un proceso lento. Las opciones para un mayor rendimiento podrían incluir enfoques de imágenes de alto contenido en formato de placa de varios pozos.
La aplicación de un enfoque estandarizado para la puntuación de exofericos debe subyacen a una disección genética concertada del proceso mediante el cual las neuronas pueden organizar y eliminar los desechos celulares.
Divulgaciones
Ninguno
Agradecimientos
Reconocemos las siguientes subvenciones NIH: R01AG047101 y R37AG56510. Los miembros de los laboratorios Driscoll y Grant han contribuido ampliamente al desarrollo y ajuste fino de los protocolos descritos, con experimentos rigurosos y una fuerte comunicación.
Materiales
| Name | Company | Catalog Number | Comments |
| 95B Scientific CMOS camera | Photometrics Prime | ||
| 1,000 μL low retention tips | Sarstedt | ||
| 10 mL serological pipette | Appleton Woods | CC214 | |
| 10 μL low retention tips | Sarstedt | 70.1130.105 | |
| 13% sodium hypochlorite | Acros Organics | AC219255000 | |
| 15 mL centrifuge tubes | Fisher Scientific | 05-539-12 | |
| 2 L erlenmeyer flasks | Scientific Laboratory Supplies | FLA4036 | |
| 25 mL serological pipette | Appleton Woods | CC216 | |
| 300 μL low retention tips | Sarstedt | 70.765.105 | |
| 50 mL serological pipette | Appleton Woods | CC117 | |
| 5-Fluoro-2'-deoxyuridine 98% | Alfa Aesar | L16497.ME | |
| 9 cm sterile Petri dishes | Fisher Scientific | 11309283 | |
| absolute ethanol | Vwr | 20821.33 | |
| Agar | Sigma Aldrich | A1296 | |
| C. elegans strain wild type | Supplied by CGC | N2 | C. elegans strain |
| calcium chloride dihydrate | Sigma Aldrich | C3881 | |
| cholesterol | Acros | 110190250 | |
| dibasic sodium phosphate | Sigma Aldrich | S3264 | |
| E. coli strain OP50 | Supplied by CGC | Op50 | E coli strain |
| FBS10 Standard microscope | Meyer Instruments | KSC 410-1-100-1 | FBS10 Standard with Plate Base, 100/100 Trinocular Head and Flip zoom |
| glass pipette 270 mm | Fisherbrand | FB50255 | |
| Heraeus Multifuge X3R | Thermofisher scientific | 75004515 | |
| Inoculating Spreaders | Fisher Scientific | 11821741 | |
| LB medium capsules | MP biomedicals | 3002-031 | |
| LDI – Laser Diode Illuminator | 89 North | ||
| levamisole | Sigma Aldrich | 16595-80-5 | |
| M4 multipette | Eppendorf | 4982000012 | |
| magnesium sulphate | Sigma Aldrich | M7506 | |
| monobasic potassium phosphate | Sigma Aldrich | P0662 | |
| Multitron Standard shaking incubator | Infors HT | INFO28573 | |
| Nalgene 1 L Centrifuge pots | Fisher Scientific | 3120-1000 | |
| P10 pipette | Eppendorf Research Plus | 3123000020 | |
| P1000 pipette | Eppendorf Research Plus | ||
| P200 pipette | Eppendorf Research Plus | 3123000055 | |
| pipeteboy 2 | VWR | 612-0927 | |
| Polystyrene microbeads | Sigma Aldrich | MFCD00131491 | |
| RC5C plus floor mounted centrifuge | Sorvall | 9900884 | |
| Reusable ringed cytology slides | ThermoFisher Scientific | 22037242 | |
| SK4005 zdIs5[Pmec-4GFP] | contract Driscoll lab | GFP expressed in touch neurons | |
| sodium chloride | Sigma Aldrich | 13422 | |
| Sodium hydroxide | Fisher Chemical | S/4880/53 | |
| Tactrol 2 Autoclave | Priorclave | ||
| Triton-X | Thermofisher scientific | 28313 | |
| Tween 20 | Sigma Aldrich | 9005-64-5 | |
| X-Light V2 Spinning Disk Confocal Unit | CrestOptics | ||
| ZB4065 bzIs166[Pmec-4mCherry] | contract Driscoll lab | mCherry expressed in touch neurons | |
| ZB4067 bzIs167[Pmec-4mitogfp Pmec-4mCherry4]; igIs1[Pmec-7YFP Pmec-3htt57Q128::cfp lin-15+] | contract Driscoll lab | Q128 expressed in touch neurons | |
| ZB4509 bzIs166[Pmec-4mCherry]; bzIs168[Pmec-7LMP-1::GFP] | contract Driscoll lab | mitoROGFP expressed in touch neurons | |
| ZB4528 bzIs166[Pmec-4mCherry]; zhsEx17 [Pmec-4mitoLS::ROGFP] | contract Driscoll lab | autophagy marker expressed in touch neurons | |
| ZEISS Axio Vert.A1 | Zeiss |
Referencias
- Davis, A. A., Leyns, C. E. G., Holtzman, D. M. Intercellular Spread of Protein Aggregates in Neurodegenerative Disease. Annual Review of Cell and Developmental Biology. 34, 545-568 (2018).
- Davis, C. H., et al. Transcellular degradation of axonal mitochondria. Proceedings of the National Academy of Sciences of the United States of America. 111 (26), 9633-9638 (2014).
- Torralba, D., Baixauli, F., Sanchez-Madrid, F. Mitochondria Know No Boundaries: Mechanisms and Functions of Intercellular Mitochondrial Transfer. Frontiers in Cell and Developmental Biology. 4, 107 (2016).
- Stahl, P. D., Raposo, G. Extracellular Vesicles: Exosomes and Microvesicles Integrators of Homeostasis. Physiology (Bethesda, Md.). 34 (3), 169-177 (2019).
- Melentijevic, I., et al. C-elegans neurons jettison protein aggregates and mitochondria under neurotoxic stress. Nature. 542 (7641), 367 (2017).
- Nussbaum-Krammer, C. I., Park, K. W., Li, L., Melki, R., Morimoto, R. I. Spreading of a prion domain from cell-to-cell by vesicular transport in Caenorhabditis elegans. PLoS Genetics. 9 (3), 1003351 (2013).
- Tyson, T., et al. Novel animal model defines genetic contributions for neuron-to-neuron transfer of alpha-synuclein. Scientific Reports. 7, (2017).
- Babcock, D. T., Ganetzky, B. Transcellular spreading of huntingtin aggregates in the Drosophila brain. Proceedings of the National Academy of Sciences of the United States of America. 112 (39), 5427-5433 (2015).
- Pearce, M. M. P., Spartz, E. J., Hong, W., Luo, L., Kopito, R. R. Prion-like transmission of neuronal huntingtin aggregates to phagocytic glia in the Drosophila brain. Nature Communications. 6, 6768 (2015).
- Fu, H., Li, J., Du, P., Jin, W., Cui, D. Metabolic wastes are extracellularly disposed by excretosomes, nanotubes and exophers in mouse HT22 cells through an autophagic vesicle clustering mechanism. bioRxiv. 10 (1), (2019).
- Ghose, P., Park, E. C., Tabakin, A., Salazar-Vasquez, N., Rongo, C. Anoxia-reoxygenation regulates mitochondrial dynamics through the hypoxia response pathway, SKN-1/Nrf, and stomatin-like protein STL-1/SLP-2. PLoS Genetics. 9 (12), 1004063 (2013).
- Cannon, M. B., Remington, S. J. Redox-sensitive green fluorescent protein: probes for dynamic intracellular redox responses. A review. Methods in Molecular Biology. 476, 51-65 (2008).
- Perkins, L. A., Hedgecock, E. M., Thomson, J. N., Culotti, J. G. Mutant sensory cilia in the nematode Caenorhabditis elegans. Developmental Biology. 117 (2), 456-487 (1986).
- Stiernagle, T. Maintenance of C. elegans. WormBook: The Online Review of C. Elegans Biology. , 1-11 (2006).
- Sutphin, G. L., Kaeberlein, M. Measuring Caenorhabditis elegans life span on solid media. Journal of Visualized Experiments. (27), (2009).
- Mitchell, D. H., Stiles, J. W., Santelli, J., Sanadi, D. R. Synchronous growth and aging of Caenorhabditis elegans in the presence of fluorodeoxyuridine. Journal of Gerontology. 34 (1), 28-36 (1979).
- Weicksel, S. E., et al. A novel small molecule that disrupts a key event during the oocyte-to-embryo transition in C. elegans. Development. 143 (19), 3540-3548 (2016).
- Dong, L., et al. Reversible and long-term immobilization in a hydrogel-microbead matrix for high-resolution imaging of Caenorhabditis elegans and other small organisms. PloS One. 13 (3), 0193989 (2018).
- Toth, M. L., et al. Neurite sprouting and synapse deterioration in the aging Caenorhabditis elegans nervous system. Journal of Neuroscience. 32 (26), 8778-8790 (2012).
- Conte, D., MacNeil, L. T., Walhout, A. J. M., Mello, C. C. RNA Interference in Caenorhabditis elegans. Current Protocols in Molecular Biology. 109, (2015).
- Calixto, A., Chelur, D., Topalidou, I., Chen, X., Chalfie, M. Enhanced neuronal RNAi in C. elegans using SID-1. Nature Methods. 7 (7), 554-559 (2010).
- Maher, K. N., Catanese, M., Chase, D. L. Large-scale gene knockdown in C. elegans using dsRNA feeding libraries to generate robust loss-of-function phenotypes. Journal of Visualized Experiments. (79), e50693 (2013).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados