
Method Article
Approcci quantitativi per il punteggio in vivo Neuronal Aggregate e Organelle Extrusion in Grandi Vesicles esopher in C. elegans
* Questi autori hanno contribuito in egual misura
In questo articolo
Riepilogo
Questo protocollo descrive gli approcci per il rilevamento e la quantificazione di grandi estrusioni aggregate e/o organelle (4 m) prodotte dalle cellule C. elegans sotto forma di esopher legati alla membrana. Descriviamo i ceppi, le condizioni di crescita, i criteri di punteggio, i tempi e le considerazioni di microscopia necessarie per facilitare la dissezione di questo meccanismo di espulsione dei detriti.
Abstract
La tossicità delle proteine misfolded e la disfunzione mitocondriale sono fattori cardine che promuovono il declino neuronale funzionale associato all'età e la malattia neurodegenerativa tra le specie. Anche se queste sfide neurotossiche sono state a lungo considerate intrinseche alle cellule, prove considerevoli ora supportano che le proteine della malattia umana misfolded originate in un neurone possono apparire nelle cellule vicine, un fenomeno proposto per promuovere la diffusione della patologia nella malattia neurodegenerativa umana.
C. elegans neuroni adulti che esprimono proteine aggreganti possono estrudere grandi vesicelle di membrana-circondato da membrana che possono includere la proteina aggregata, mitocondri, e lisosomi. Queste grandi vesicles sono chiamate "esophers" e sono distinte dagli esosomi (che sono circa 100 volte più piccoli e hanno una biogenesi diversa). Gettare detriti cellulari negli esopher può verificarsi da un meccanismo conservato che costituisce un ramo fondamentale, ma precedentemente non riconosciuto, della proteostasi neuronale e del controllo della qualità mitocondriale, rilevante per i processi con cui gli aggregati si diffondono nelle malattie neurodegenerative umane.
Mentre gli esopher sono stati studiati per lo più in animali che esprimono mCherry transgenici ad alta copia all'interno dei neuroni touch, questi protocolli sono ugualmente utili nello studio dell'esoferegenesi utilizzando organelli con tag fluorescente o altre proteine di interesse in varie classi di neuroni.
Qui sono descritte le caratteristiche fisiche degli esopher C. elegans, le strategie per il loro rilevamento, i criteri di identificazione, i tempi ottimali per la quantificazione e i protocolli di crescita animale che controllano le sollecitazioni che possono modulare i livelli di produzione esopher. Insieme, i dettagli dei protocolli qui descritti dovrebbero servire a stabilire uno standard per l'analisi quantitativa degli esopher tra i laboratori. Questo documento cerca di fungere da risorsa nel campo per i laboratori che cercano di elaborare meccanismi molecolari attraverso i quali vengono prodotti gli esopher e con i quali gli esopher vengono reagiti da cellule vicine e lontane.
Introduzione
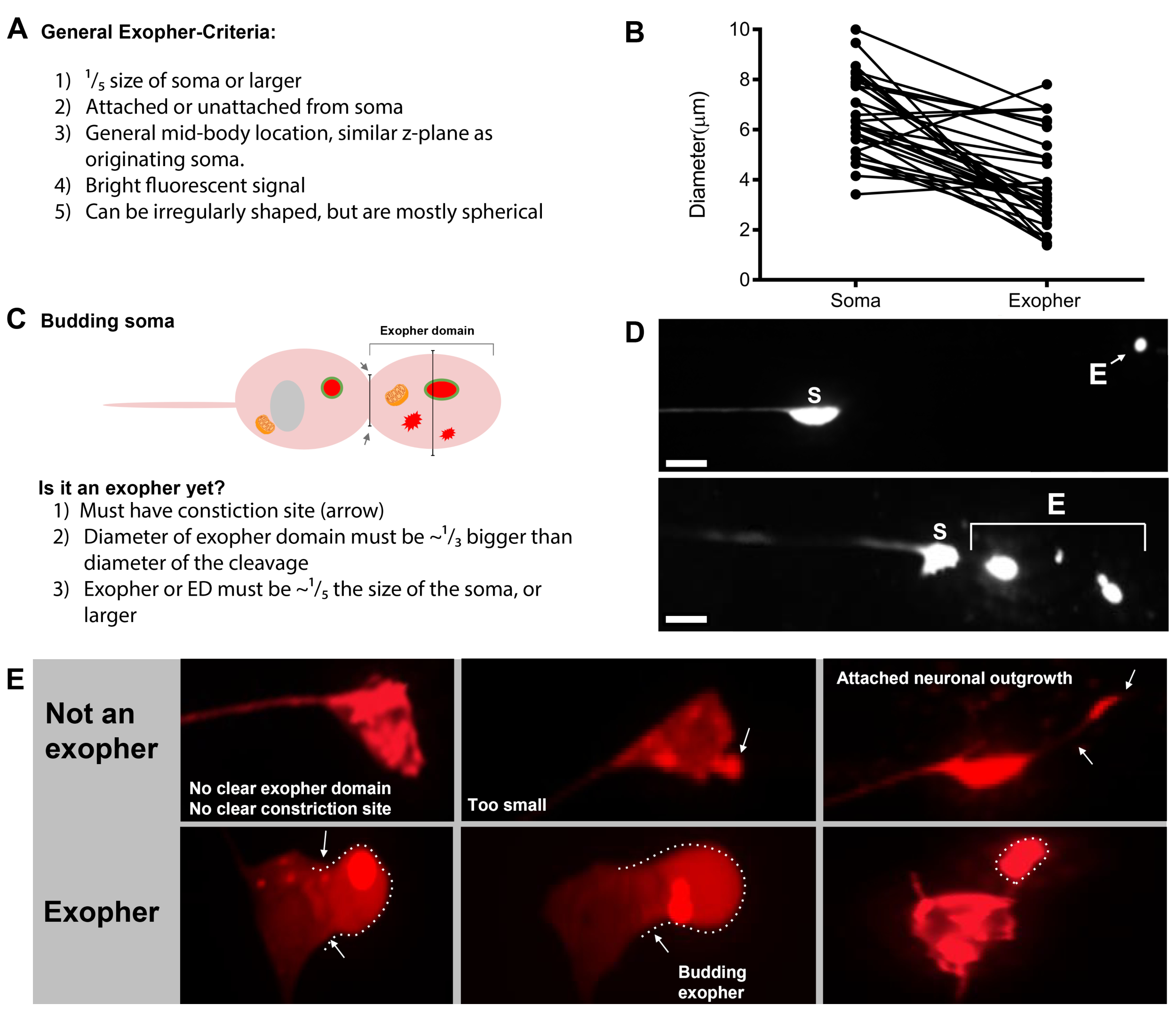
Le sfide neurotossiche degli aggregati e dei mitocondri disfunzionali sono state a lungo considerate intrinseche alle cellule, ma più recentemente è diventato chiaro che le proteine della malattia umana mal ben posizionate provenienti da un neurone possono diffondersi anche nelle cellule vicine, promuovendo la patologia1. Allo stesso modo, i mitocondri dei mammiferi possono essere inviati fuori dalla cellula della loro produzione originale per la degradazione transcellulare2 o per il salvataggio di popolazioni mitocondriali nelle cellule vicine contestate3. Vescicle di varie dimensioni sono stati generalmente osservati per trasferire materiali cellulari alle cellule vicine o all'ambiente fluido4. Alcune vesciche estruse si avvicinano alle dimensioni del soma neuronale medio (soma neurone di tocco medio - 6 m) e possono ospitare grandi aggregati e organelli.
Un esempio lampante di grande estrusione vescicola che può trasportare aggregati proteici e organelli si verifica nei neuroni del recettore del tocco C. elegans che esprimono un alto numero di copia reporter costruire codificando un mCherry 5 soggetto di aggregazione nocivo e resistente alladegradazione. Le estrusioni dai neuroni tattili, chiamati esopher, sono di diametro medio di 4 m, includono selettivamente mCherry o altri aggregati, e vengono consegnate direttamente nell'ipodermide vicino, che normalmente circonda i neuroni del recettore tat toccato. L'ipodermide tenta la degradazione a base di lisosomi, ma alcuni contenuti non digeribili come gli aggregati mCherry possono essere riesaciti dagli ipodermi nello pseudocoelom pieno di liquido dell'animale, da cui il mCherry può essere ripreso da celle di scavenger remote chiamate coelomociti per l'archiviazione a lungo termine (Figura 1, Figura 2)5.
Le grandi vesciche esodiche estruse lasciano la cellula circondata da membrana plasmatica del recettore tat toccato e possono contenere proteine aggregate della malattia umana, mitocondri e lisosomi. Il processo di produzione di esopher sembra comportare lo smistamento di specie potenzialmente tossiche (ad esempio, un mCherry espresso soggetto ad aggregazione è separato da proteine solubili e inoffensive come il GFP che rimane per lo più nel soma neuronale). In questo modo, l'espulsione diretta delle entità minacciose viene eseguita dal neurone5. Una sfida di proteostasi, come lo stress indotto dal knockdown autofagia, l'inibizione proteasome mediata da MG132 o l'espressione transgenica delle proteine della malattia umana come la poliglutamina espansa Q128 associata alla malattia di Huntington o il frammento implicato dalla malattia di Alzheimer Aβ1-42, può aumentare il numero di neuroni che producono esophers5.
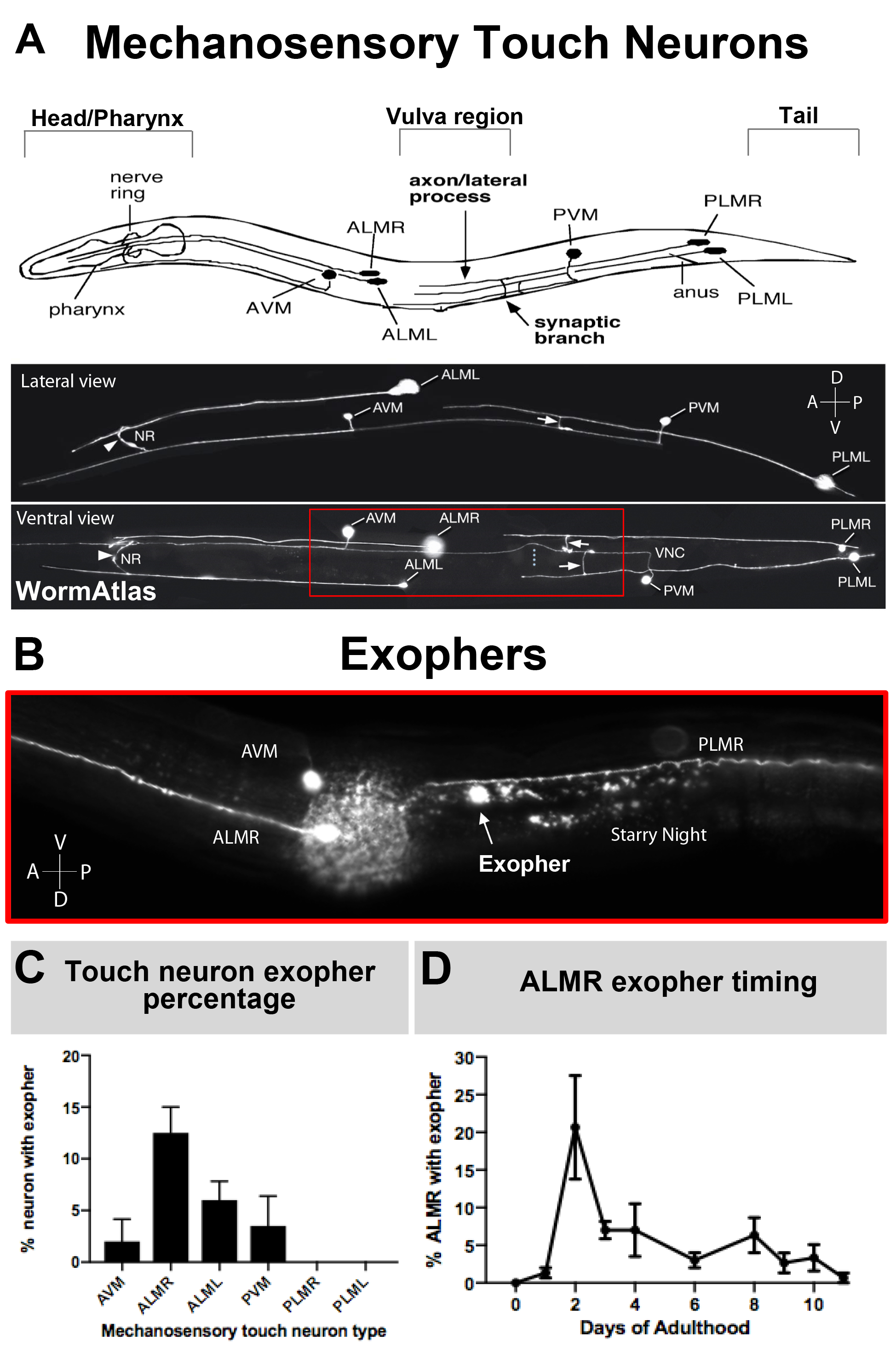
Come gli esopher sono stati documentati solo di recente, ciò che è noto della loro biologia merita la descrizione. Gli esopher sono stati scoperti in, e sono i più studiati in, C. elegans toccare i neuroni recettori. Ci sono sei neuroni del tocco meccanossensoriale C. elegans che hanno corpi cellulari distribuiti intorno al corpo (Figura 3A) e sono chiamati cellule microtubuli perché la loro ultrastruttura presenta distintivi 15 microtubuli protofilament. I neuroni del recettore tatuatore sono l'AVM anteriore (neurone ventrale anteriore), ALMR e ALML (neuroni microtubuli intermedi a destra e sinistra), il PVM più centrale (neurone ventrale posteriore), e il PLMR posteriore e PLML (neuroni microtubuli successivi posteri a destra ea sinistra) nella coda. È interessante notare che, i sei neuroni del recettore tattile producono esopher a velocità diverse, pur esprimendo lo stesso transgene offensivo (Figura 3C). Dei sei neuroni del recettore del tocco meccanossenoriale, il neurone ALMR subisce l'esophergenesi più spesso degli altri neuroni touch. La quantitazione dei numeri esopher dai neuroni touch è quindi di solito stabilita concentrandosi sull'ALMR.
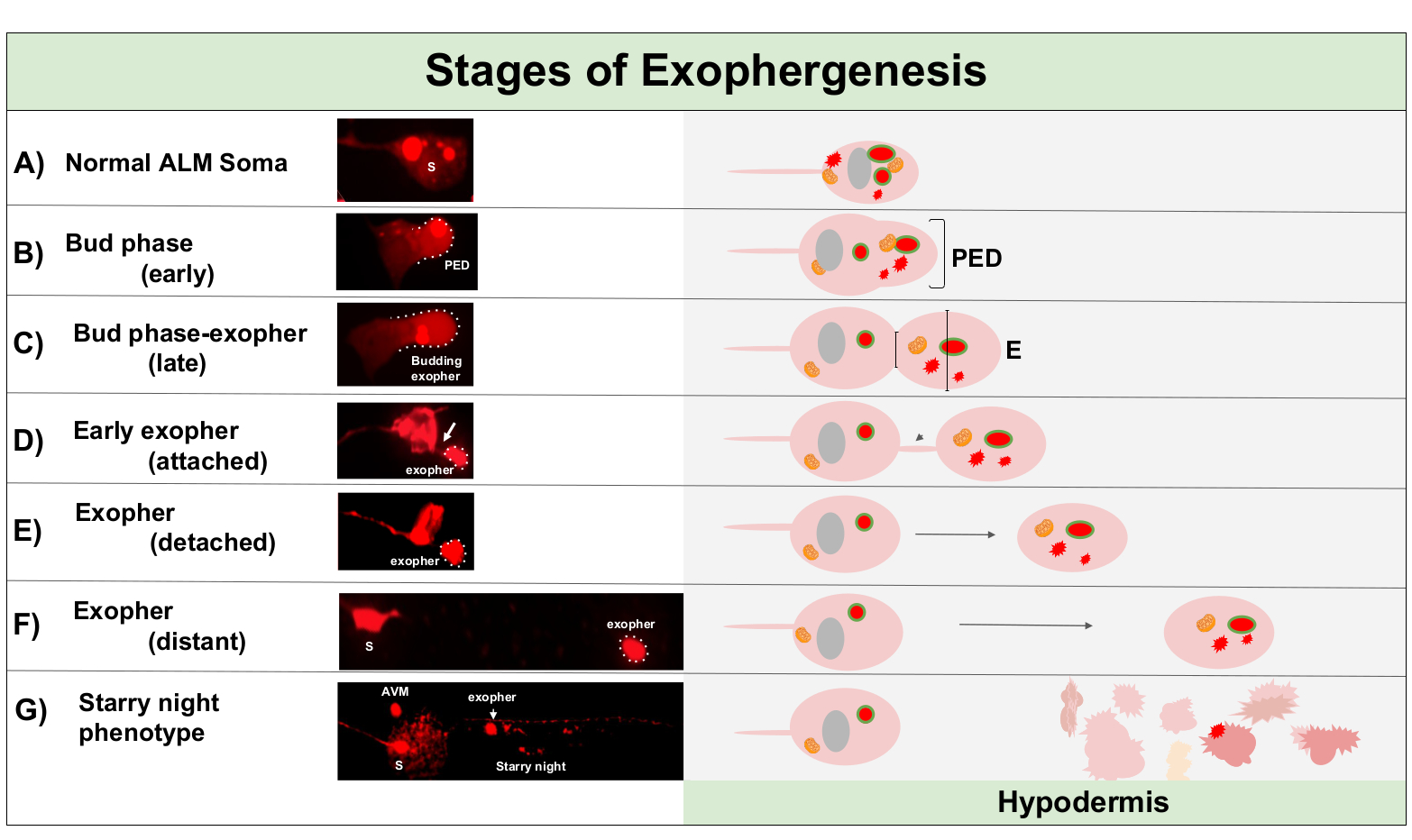
L'esophergenesi è un processo dinamico che in genere inizia con il gonfiore del citoplasma neuronale (Figura 1A-B). I contenuti cellulari, gli organelli o gli aggregati proteici vengono raccolti su un lato del soma neuronale, più comunemente verso l'estremità posteriore del neurone ALMR (lontano dal neurite di progetto), formando un dominio pre-esopher (PED) (Figura 1B). La sporgenze precoce si osserva quando il PED inizia a proiettarsi verso l'esterno, formando un bocciolo sporgente riconoscibile. Il bocciolo tardivo è definito quando il diametro più largo del dominio pre-esopher è di circa 1/3 più grande del diametro della costrizione del collo soma-exopher (Figura 1C). Gli esopher possono essere espulsi in quasi tutte le direzioni dal soma, ma la maggior parte degli esopher escono posteriormente dal corpo della cellula e rimangono approssimativamente nello stesso piano focale del soma originario.
L'esopher può allontanarsi dal soma originario mentre il collo del germoglio si restringe in un filamento sottile. Gli esopher possono rimanere attaccati al soma tramite questo filamento (Figura 1D, freccia) e possono successivamente staccarsi. Il contenuto cellulare come calcio, aggregati e mitocondri può essere trasferito tramite questo filamento nell'esopherallegato 5, anche se la maggior parte del materiale estruso viene messo nel compartimento esodario dal massiccio evento in erba. Gli esopher sono considerati maturi quando non vi è alcun tubo di collegamento visibile o filamento sottile e l'esopher è completamente separato dal soma di invio (Figura 1E).
Gli esopher prodotti da C. elegans toccano i neuroni incontrano immediatamente l'ipodermide, il tessuto che circonda il neurone touch. Più comunemente, il vescicolo esopher sembra viaggiare all'interno dell'ipodermio posteriormente verso la coda, e può essere abbastanza distante dal soma prima che i contenuti esopher appaiano mirati alla degradazione (ad esempio, la distanza può essere di 100 m di distanza dal soma (Figura 1F)). La vescica esopher fluorescente si rompe in molte vescicole più piccole all'interno dell'ipodermide, assumendo un aspetto chiamato "notte stellata" (Figura 1G e Figura 2). Nella fase della "notte stellata", il materiale fluorescente puntuale può essere osservato sparso attraverso il sincizio ipodermico in molti punti più piccoli di fluorescenza rispetto all'esopher solitario originale. Notte stellata può sembrare puntuale sotto basso ingrandimento e con ingrandimento maggiore, può sembrare puntuale e / o in rete all'interno dell'ipodermide. Il segnale fluorescente della notte stellata è tipicamente dimmer rispetto all'esopher e alla fluorescenza neuronalmente espressa (Figura 2B-C). La dispersione di mCherry in molte vesciche puntuali è pensata per coinvolgere la maturazione fagosomica e la fusione con la rete endosomal/lisosomica della cellula ipodermica. Alcuni materiali esopher sono probabilmente degradati nella rete lisosomica ipodermica, ma le specie residue resistenti alla degradazione (come gli aggregati mCherry) vengono gettate dall'ipodermide nello pseudocoelom, un compartimento fluido che può contenere detriti cellulari. Il materiale fluorescente viene successivamente assunto da celle di scavenger remote chiamate coelomociti (Figura 2C), che possono concentrarsi, immagazzinare e tentare nuovamente la degradazione di mCherry.
Il fenomeno dell'estrusione aggregata e del trasferimento appare conservato attraverso la fita, essendo stato riportato in modelli genetici come C. elegans5,6,7 e D. melanogaster8,9 così come in più modelli di mammiferi. Sono state segnalate estrusioni simili a esopher per le cellule dei mammiferi10, un'osservazione che suggerisce che i meccanismi conservati potrebbero essere alla base dell'espulsione aggregata e degli organelli. La produzione di esopher può quindi essere un meccanismo conservato di gestione dei detriti cellulari che costituisce un ramo fondamentale, ma precedentemente non riconosciuto, della proteostasi neuronale e del controllo della qualità mitocondriale, che, quando squilibrato, potrebbe contribuire attivamente alla malattia neurodegenerativa. L'identificazione delle molecole coinvolte nella discriminazione e nello smistamento dei detriti, il trasporto in un locale subcellulare distinto, l'estrusione, la formazione/scissione della connessione tubolare che collega il soma e l'esopher tardivo, e il riconoscimento della grande vescicola estrusa per la degradazione remota da parte di una cellula vicina rimangono per il lavoro futuro. Gli studi sui modelli di nematode e mosca saranno di fondamentale importanza per definire i meccanismi di raccolta e trasferimento di aggregati e organelli, utilizzando approcci genetici imparziali e potenti strumenti biologici cellulari offerti da questi modelli per identificare le molecole partecipanti in un contesto fisiologico.
I primi passi critici nella decifrazione dei meccanismi operativi nella biologia esopher comportano la definizione di protocolli per la quantificazione dell'esopher in vivo riproducibili. Il modello C. elegans offre un vantaggio particolare per tali sforzi poiché il corpo è trasparente e gli esopher possono essere facilmente osservati quando contengono proteine o organelli con etichetta fluorescente. Exophers sono stati segnalati per essere generato da C. elegans neuroni dopaminergici PDE e CEP, neuroni sensoriali ASE e ASER, e neuroni anfami di riempimentocolorante 5. Poiché gli esopher prodotti dai neuroni del recettore tat toccato sono meglio caratterizzati, l'attenzione qui è sull'uso di neuroni tattili per l'analisi esopher. Tuttavia l'approccio di base può essere applicato per misurare la produzione di esopher da qualsiasi cellula. Vengono delineati protocolli per rilevare e quantificare gli eopher prodotti da C. elegans che toccano i neuroni recettori che esprimono transgenicamente la proteina mCherry, con un'enfasi sui carichi che possono essere monitorati e sui vincoli temporali nel punteggio. Questo articolo definisce gli approcci verso l'identificazione dell'esopher in vivo e la quantificazione delle condizioni ambientali e genetiche che modulano la produzione di esopher. I protocolli sottolineano l'attenzione critica alle condizioni costanti non di stress per la determinazione della produzione di esopher di base e per i confronti tra i genotipi.
Protocollo
1. Ceppi utili per il rilevamento esopher
- Selezionare un ceppo che esprime carichi fluorescenti all'interno dei neuroni di C. elegans per visualizzare facilmente gli esopher.
NOTA: nella tabella 1 sono elencati i ceppi che sono stati utilizzati per visualizzare gli esopher prodotti nei neuroni del recettoretat toccato 5,11,12. In linea di principio, qualsiasi cellula o tipo neuronale può essere testato per la produzione di esodiche utilizzando un promotore specifico di cellule o tessuti per guidare l'espressione di una proteina fluorescente che aggrega o è altrimenti selezionata per l'estrusione. - In alternativa, utilizzare un saggio di riempimento del colorante per visualizzare gli esopher nei neuroni della testa amphide, che sono aperti all'ambiente e sussalibili al riempimento5,13.
2. Media di crescita
- Preparare i supporti di crescita nematode standard (NGM) ai ceppi di coltura secondo i metodi standard14,15.
NOTA: La mancanza di cibo, o fluoro-deossiuridina (FuDR), usata comunemente per bloccare la produzione di progenie, e può influenzare drammaticamente la produzione di esopher. Mantenere la popolazione continuamente nutrita (evitare anche brevi periodi di esaurimento degli alimenti batterici) e mantenere gli animali a una temperatura costante.
3. Allevamento di animali critico per una produzione coerente di esopher
- Allevare animali su mezzi di comunicazione coerenti e con fonti di cibo batterico coerenti. Gli animali non devono esersi di cibo batterico, anche per brevi periodi di tempo, poiché la limitazione degli alimenti può cambiare drasticamente i livelli di produzione di esopher.
- Mantenere le ricette multimediali e la preparazione uniforme durante tutto uno studio.
NOTA: la modifica dei supporti può influire sui livelli basali di produzione di esopher. I lotti di agar possono influenzare i livelli di esopher di base, quindi quando i lotti di fornitura cambiano, prendere nota della data. Gettare le piastre di brodo dopo due settimane per garantire cibo batterico sano e per prevenire l'agar essiccato, che provoca cambiamenti nell'osmolarità dell'agar che influenzano i livelli di esodato. - Per le condizioni basali, tenere gli animali ad una temperatura costante di 20 gradi centigradi. L'allevamento di animali a temperature variabili (anche variazioni temporanee di temperatura) può causare variazioni nella tempistica della produzione massima di esopher.
NOTA: la variabilità della temperatura non è limitata alle condizioni di coltura. Le variazioni delle temperature durante gli esperimenti o al banco di laboratorio possono avere un impatto. Ad esempio, le temperature all'interno di una stanza al microscopio non dovrebbero differire drammaticamente dall'incubatore di coltura o dal banco di laboratorio. - Non utilizzare interventi farmacologici anti-fertilità perché le uova fecondate sono fondamentali per la produzione adulta precoce di esopher.
NOTA: L'uso di Fluoro-deoxyuridine (FuDR)16 o C2217, deve essere evitato. Quando si eseguono esperimenti sugli animali di vecchiaia o di vecchiaia, le popolazioni sincronizzate con l'età dovrebbero essere mantenute rimuovendo fisicamente gli adulti dalla loro progenie più piccola raccogliendoli su piastre fresche diffuse con batteri piuttosto che utilizzando comuni interventi farmacologici anti-fertilità. - Non utilizzare colture contaminate; reinizializzare gli esperimenti in caso di compromesso biologico della popolazione o della piastra. La contaminazione batterica o fungina può indurre stress e cambiamenti metabolici negli animali e deve essere assente dalle popolazioni sperimentali.
- Per massimizzare i risultati riproducibili, mantenere le colture per almeno due generazioni sane, ben nutrite e senza contaminazione a 20 gradi centigradi prima della sperimentazione per evitare potenziali cambiamenti epigenetici indotti dall'ambiente.
4. Sincronizzazione dell'età per il punteggio esopher mediante sbiancamento, galleggiamento di saccarosio o raccolta di larve L4
- Mantenere le popolazioni sperimentali della stessa età biologica, poiché i modelli di rilevamento degli esopher variano con l'età adulta e il confronto degli animali con popolazioni miste di età mista può confondere i risultati. Garantire sempre la corretta sincronizzazione delle popolazioni animali sperimentali controllando la morfologia della vulva bianca nella fase L4.
NOTA: Generalmente, la produzione di esopher di picco per i neuroni ALMR meccanosi C. elegans si verifica nel giorno adulto 2-3 (Figura 3D), come misurato dai giorni dopo lo stadio L4. Adulto giorno 1 è 24 ore dopo la fase larvale L4 che si distingue per morfologia vulva "mezzaluna bianca" (Figura 5E). - Preparare popolazioni di uova sincronizzate sbiancando gli adulti gravid.
- Raccogliere adulti gravid riempiti di uova lavando gli animali che crescono su un piatto NGM. Per lavare, inondare la piastra con 1 mL di tampone M9, pipetta su e giù per raccogliere il liquido con animali sospesi e pipetta in un tubo microcentrifuge da 1,5 mL. Pellet animali da assestamento gravitazionale o centrifugazione dolce con una mini centrifuga e rimuovere il supernatant.
- Aggiungere 150 L di 5M NaOH e 150 L del 6% di ipoclorito di sodio (candeggina) in 1 mL in H2O e mescolare per inversione per circa 5 minuti.
NOTA: La soluzione di sbiancamento fresca assicura che la cuticola animale possa essere interrotta per la raccolta delle uova. I progressi nell'interruzione della cuticola possono essere monitorati al microscopio sezionante; gli adulti devono rompere e rilasciare le uova nel punto in cui lo sbiancamento deve essere interrotto. - Centrifugare delicatamente con un tubo minicentrifuge per 20 s e rimuovere il supernante. Aggiungere di nuovo 1 mL di buffer M9 e centrifugare, lasciando circa 100 l sulla parte superiore del pellet.
- Ripetere i passaggi 4.2.3 due volte per rimuovere le tracce della soluzione di candeggina.
- Risundere le uova nel volume rimanente e trasferirlo in una piastra NGM fresca. Gli adulti saranno lised, ma molte uova vitali dovrebbero essere nella preparazione.
- Preparare popolazioni sincronizzate a tempo.
- Raccogliere 20 adulti gravid a una piastra NGM semita utilizzando i protocolli di trasferimento standard14.
- Consentire agli animali di strisciare liberamente e deporre le uova per 1,5 h (i ceppi mutanti con basse dimensioni della covata possono richiedere l'introduzione di più animali adulti).
- Rimuovere tutti gli animali adulti dalla piastra raccogliendo, lasciando la popolazione di uova sincronizzate alle spalle. Controllare le piastre poche ore dopo per verificare che nessun adulto vitale sia stato perso durante la rimozione degli adulti.
- Preparare popolazioni di uova sincronizzate per selezione di uova di galleggiamento di saccarosio.
- Raccogliere animali e uova da cinque piastre NGM su cui gli animali gravide hanno deposto le uova per almeno 24 ore allagando le piastre con soluzione M9 con detergente 0,1% (come Tween 20 o Triton X-100) e raccogliendo in un tubo da 15 mL. Pellet adulti da centrifugazione delicata a temperatura ambiente (2.000 x g per 30 s).
- Rimuovere tre volte gli animali supernanti e lavarli in 15 mL di M9 fresco, scartando il supernante dopo ogni lavaggio, assicurandosi di mantenere il pellet arricchito in animali e uova.
- Conservare 2 mL di supernante e rissuspendere il pellet. Aggiungere 2 mL di peso del 60% per saccarosio volume.
- Centrifuga a 2000 x g per 5 min. La soluzione mostrerà ora una fase superiore altamente arricchita nelle uova.
- Trasferire circa 2,5 mL della fase superiore in un nuovo tubo da 15 mL e aggiungere 10 mL di M9.
- Mescolare per inversione per 1 min, quindi centrifugare 2000 x g per 1 min.
- Togliere il supernante e lavare il pellet arricchito di uova in M9. 10-15 L del pellet d'uovo possono essere distribuiti a una piastra NGM fresca con semi OP50.
NOTA: Questo metodo prepara un gran numero di uova; non consentono agli animali raccolti di esarsi dal cibo OP50 E. coli.
- Preparare popolazioni sincronizzate raccogliendo gli animali nella fase di sviluppo L4.
- Coltivare animali su piastre NGM semi come descritto sopra.
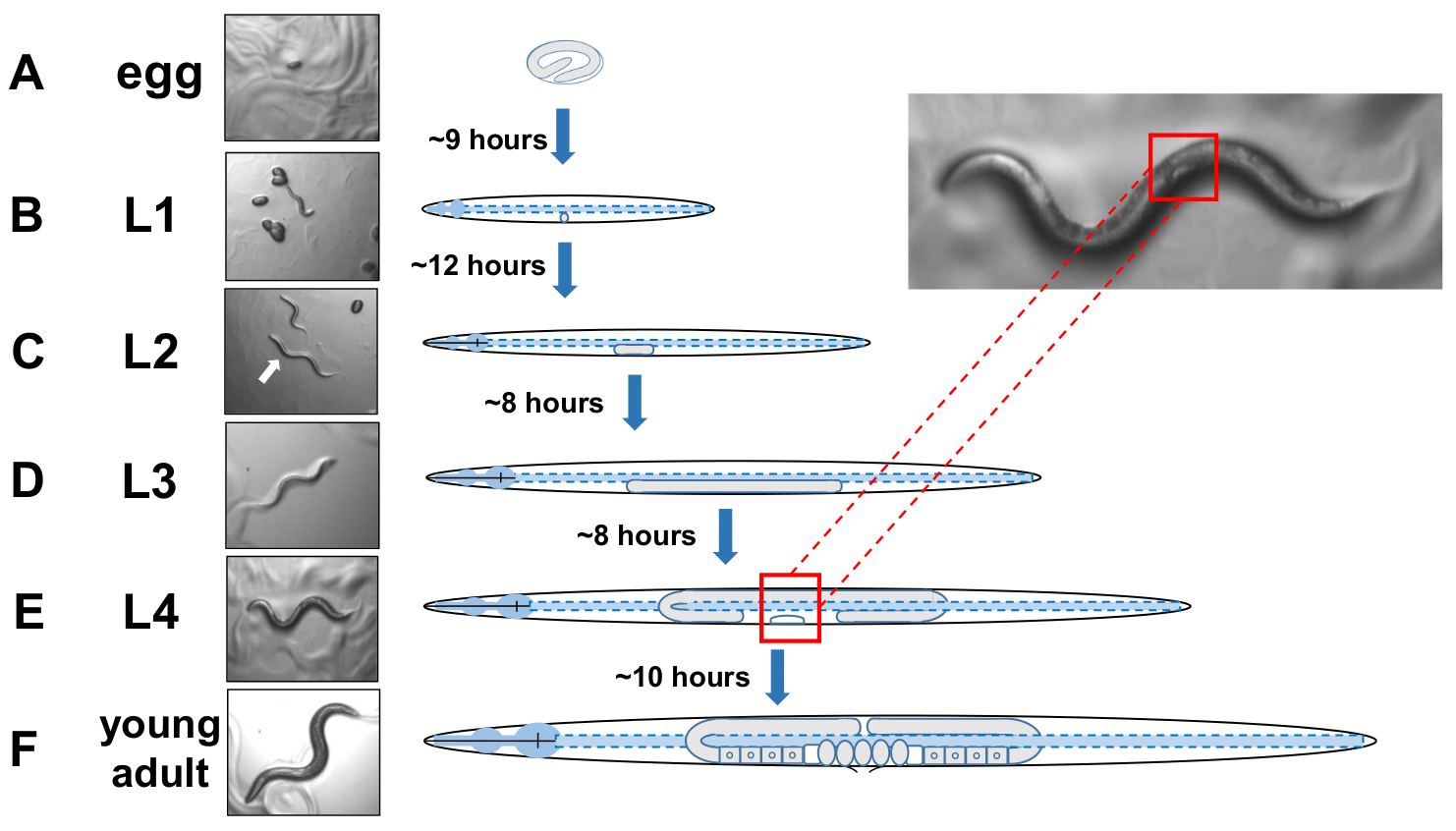
NOTA: I C. elegans si sviluppano in quattro fasi distinte. A 20 gradi centigradi, un uovo appena deposto impiega circa 9 ore per schiudersi (Figura 5A). Post hatch, un animale passa attraverso larvale fase 1 (L1) alla fase larvale 4 (L4), con ogni fase che dura 8-12 ore tra ogni muta(Figura 5A-F). Pertanto, un piatto preparato da inoculazione con uova dovrebbe avere molti animali L4 per raccogliere circa 40 ore dopo l'introduzione delle uova. - Identificare gli animali in fasi L4 individuando la forma a mezza luna bianca della vulva in via di sviluppo (Figura 5E).
NOTA: Gli animali allo stadio L4 sono uniformi per dimensioni e pigmentazione del corpo. Raccogliere gli animali con la mezzaluna bianca in un piatto di crescita fresco per l'esame degli animali in scena. Il giorno successivo (24 ore dopo) deve essere conteggiato come adulto giorno 1. - Segnare una popolazione di animali ogni giorno il giorno adulto 2.
NOTA: Gli esopher sono in genere segnati al secondo giorno dell'età adulta, che è il picco della produzione di esopher in condizioni basali. Tuttavia, poiché il picco e la tempistica dell'esophergenesi può essere spostato da cambiamenti ambientali o genetici che vengono studiati, si consiglia di segnare una popolazione di animali adulti ogni giorno per quattro giorni per generare il quadro più completo (Figura 3D).
- Coltivare animali su piastre NGM semi come descritto sopra.
5. Rilevamento di esopher utilizzando un microscopio fluorescente
- Osservare gli esopher utilizzando un microscopio pseudo-stereo sezionante ad alto ingrandimento dotato di microscopia a fluorescenza.
- Immobilizzare gli animali su piastre NGM mediante pipettamento 100-200 L di soluzione levamisole/tetramisole da 10-100 mM sulla superficie della piastra di agar NGM. Dopo 2-4 minuti, gli animali diventano paralizzati e possono essere osservati direttamente sulla piastra dell'agar.
NOTA: I trattamenti di immobilizzazione non sono assolutamente necessari, in modo tale che con un occhio addestrato, l'identificazione neuronale e la presenza esopher possano essere segnati seguendo visivamente gli animali che strisciano al microscopio sulla piastra quando si determina se è stato prodotto o meno un esopher. - Osservare i neuroni fluorescenti utilizzando un ingrandimento totale di 100x per eseguire il rilevamento del microscopio sezionante degli esopher.
NOTA: il punteggio di eventi esopher utilizzando la microscopia di dissezione consente l'osservazione di un gran numero di animali con relativa facilità direttamente sulle piastre di agar su cui sono allevati. - Imaging dal vivo e montaggio ceppi reporter per studi esopher utilizzando microscopia confocale
- Utilizzare un microscopio confocale per l'immagine intracellulare dinamica e le caratteristiche dell'esophergenesi.
NOTA: L'imaging dal vivo è un approccio vantaggioso per osservare dettagli sottili della produzione di esopher perché la produzione di esopher è un processo dinamico. - Limitare il movimento degli animali per l'imaging dal vivo ad alta risoluzione utilizzando metodi convenienti, tra cui l'utilizzo di levamisole o tetramisole a 10-100 mM o l'applicazione di microsfere idrogel (con diametri di 15 m, 30 m o 40 m)18.
- Utilizzare un microscopio confocale per l'immagine intracellulare dinamica e le caratteristiche dell'esophergenesi.
- Preparazione del scorrimento per microscopia composta e confocale
- Montare 20-50 animali in un agente immobilizzante su uno scivolo al microscopio. I vetrini di citologia ad anello riutilizzabile verniciati con anelli rialzati di 13 mm di diametro sono utili per il montaggio.
- Raccogliere gli animali vivi in 5-20 L di un paralitico come levamisole da 10-100 mM o tetramisole all'interno del cerchio dipinto o sul cuscinetto di agar.
- Attendere 4 minuti per la paralisi, quindi coprire il scorremento con un coverslip (consigliato n. 11/2 (0,16 – 0,19 mm) o n. 2 (0,17 - 0,25 mm).
- Montaggio di un piccolo numero di animali
- Non schiacciare gli animali montati; quando si osservano solo pochi (meno di 20) animali per scivolo, vi è il rischio di schiacciare alcuni degli animali a causa della pressione disuguale del costo. Questo rischio può essere ridotto al minimo utilizzando un pad di agarose a bassa percentuale per il montaggio.
- Fare un 2-4% agarose pad scivolo, e quindi aggiungere 2-15 L di soluzione paralitica al pad. Tenete a mente levamisole e tetramisole diffuse nel pad, diminuendo la loro concentrazione effettiva.
- Montare raccogliendo gli animali in una goccia di soluzione paralitica di 2-15 L o microsfere appoggiate sul cuscinetto di agar. Posizionare il coperture sulla parte superiore e verificare che gli animali siano intatti18.
- Preparazione del pad Agar
- Per preparare 2% agar pad, calore 2% agarose nella soluzione M9 e microonde fino a quando l'agarose è in uno stato omogeneo e fuso.
- Per ottenere un cuscinetto di agar di qualità sufficiente, miscelazione alternativa e microwaving a bassa potenza per meno di 20 secondi. Evitare l'inclusione di bolle d'aria all'interno del pad mettendo agar bollente su un blocco di riscaldamento e permettendo alle bolle di salire in superficie.
- Utilizzare una pipetta Pasteur per disegnare l'agar dal profondo all'interno della soluzione fusa sotto le bolle alzate.
- Preparare due vetrini con spina e posizionare su entrambi i lati di un vetrerà al microscopio di vetro pulito su una superficie piana. Per fare in modo che le diapositive con nastro adesivo possentino due strisce di nastro da laboratorio da 5 cm su ogni vetrini (Figura 6A).
- Utilizzando un pipetto Pasteur, posizionare una singola goccia di agar sul vetrini al microscopio pulito inserito tra i vetrini spinati (Figura 6B).
- Con attenzione e rapidità, coprire la goccia di agar fuso con una quarta diapositiva pulita posizionando attraverso le diapositive toccate (Figura 6Cc).
NOTA: La diapositiva deve premere delicatamente l'agar fuso in un cerchio appiattito di circa 0,4 mm di spessore (lo spessore del nastro) (Figura 6D). L'agar dovrebbe raffreddarsi rapidamente. - Rimuovere la diapositiva superiore facendola scorrere ( Figura6E) . Le pastiglie di agar si asciugano rapidamente e sono meglio utilizzate in pochi minuti. Una volta rimosso lo scivolo superiore, utilizzare immediatamente il gel pad per il montaggio degli animali. Evitare di utilizzare pastiglie con bolle d'aria.
- Conservare pastiglie di agar fino a 30 minuti racchiusi tra i due scivoli di vetro. L'agar essiccato fa sì che gli animali si agglomerino e dislocateno. Montare gli animali entro 2-15 L di soluzione paralitica o microsfere e coprire con coperture; schermo la diapositiva entro 20 minuti di paralisi e montaggio (Figura 6).
NOTA: Poiché le condizioni di stress possono alterare i tassi di esopher, evitare i paralitici che possono indurre stress ossidativo (adesivi di sodio) durante lo screening per esophers.
- Rilevamento di esopher utilizzando un microscopio confocale a disco rotante
- Osservare le caratteristiche biologiche delle cellule come organelli e altri contenuti con obiettivi di apertura numerica 1.4 a 63x e 100x.
- Utilizzare un software in grado di controllare le stage e l'acquisizione di immagini utilizzando l'acquisizione multidimensionale. I microscopi e il software di elaborazione delle immagini dovrebbero anche essere adatti per l'imaging e la raccolta dei dati, poiché questi passaggi prevedono approcci di imaging standard.
6. Identificare i neuroni tattili e il punteggio per gli esopher con animali montati
- Monte animale adulto paralizzato (Figura 6).
- Identificare il piano z desiderato. Utilizzare basso ingrandimento campo luminoso (10-40x) per identificare il piano z adatto dell'animale, prendendo atto del posizionamento dell'animale, orientamento testa-coda, e la posizione della vulva - che sono punti di riferimento per l'identificazione neuronale ed esopher successiva (Figura 3A & Figura 5E).
- Concentrati sul segnale di fluorescenza del reporter prescelto. Rimanendo nello stesso piano z, passa alla visualizzazione della fluorescenza widefield a 10-40x per il reporter citosolico scelto.
NOTA: In questo esempio l'espressione fluorescente è guidata dal promotore mec-4 mechanosensory touch neurone specifico. Alti array di copia e fluorofori diversi hanno variabilità nell'espressione e quindi intensità fluorescente variabile. Regolare se necessario. - Scorrere all'interno dell'asse z per osservare la profondità dell'animale e l'espressione fluorescente nel piano focale. In questo modo, confermare l'orientamento testa-coda; la testa/pharynx avrà l'anello del nervo fluorescente e in questo caso, la coda conterrà 1-2 somi PLM visibili (Figura 3A).
- Identificare i neuroni tattiti
- Identificare se l'animale è montato sul lato sinistro o destro (Figura 3A).
NOTA: Considerando la tridimensionalità dell'animale, la migliore risoluzione di imaging viene eseguita sul lato più vicino all'ottica. - Identificare il soma (ALM, ALMR, AVM) osservando - iniziare dalla testa per identificare l'anello nervoso e processi neuronali laterali.
- Con un ingrandimento di 10-40x, scorrere lentamente l'asse z per identificare il processo associato.
- Una volta identificato il processo, seguirlo lateralmente nella direzione posteriore verso la vulva, dove il soma sarà evidente, segnato da un corpo cellulare rotondo alla fine del processo. Una volta che il soma neuronale più in-focus si trova, può essere identificato utilizzando altri punti di riferimento neuronali come segue:
- Utilizzare l'AVM, un neurone ventrale nelle vicinanze, per aiutare ad assegnare l'orientamento animale. Se il neurone AVM è nello stesso piano dell'ALM, l'animale sta poggiando su un fianco e il neurone all'esterno di quell'aereo è l'ALMR . Se il neurone AVM non si trova nello stesso piano dell'ALM in questione, il neurone touch più vicino al piano focale è ALML.
- Identificare il neurone PVM, un altro neurone ventrale touch situato vicino alla coda, per indicare se il neurone di tocco anteriore è nello stesso piano. Se è così, il neurone tatto osservato è ALML.
- Ottenere un senso della posizione di altri corpi soma, vicino all'area di interesse (neuroni fluorescenti situati su entrambi i lati del soma), e in tutti i piani z, anche se non è possibile ottenere il neurone più profondo impostato in chiaro fuoco.
NOTA: L'identificazione di tutti i somi neuronali touch è importante perché il soma fuori fuoco può essere scambiato per esophers.
- Identificare se l'animale è montato sul lato sinistro o destro (Figura 3A).
7. Identificazione e punteggio per gli esopher
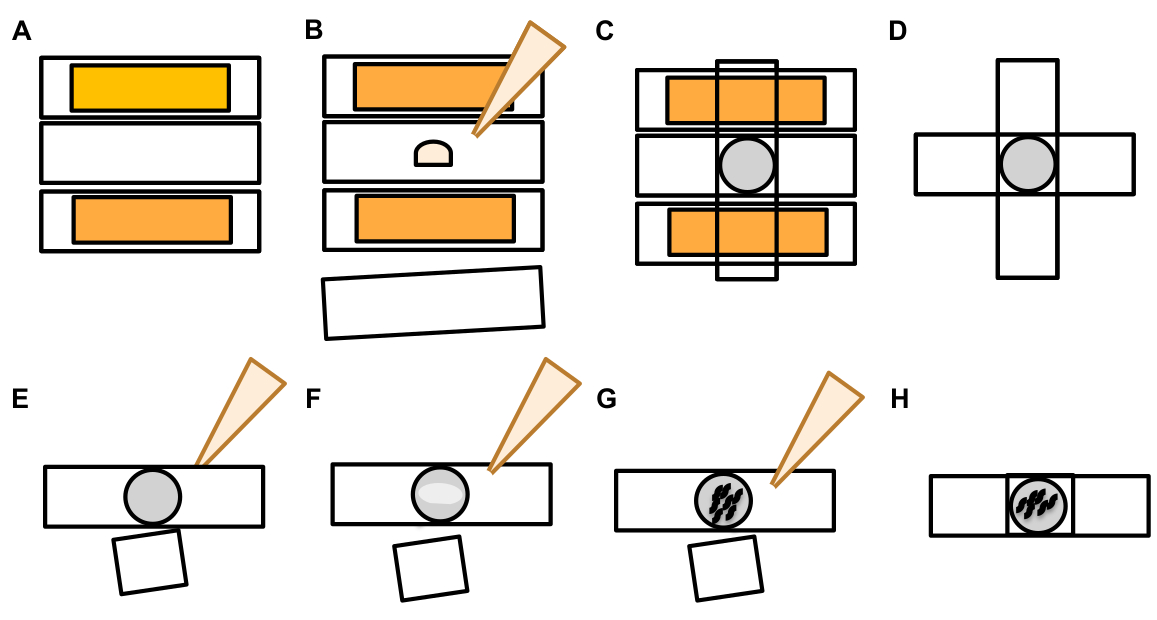
- Una volta trovato un neurone touch, ispezionarlo per grandi sporgenze (domini esopher) abbastanza grandi da essere considerato un esopher gemma, (raggiungendo almeno 1/5th la dimensione del soma di origine) ( Figura1C).
NOTA: l'esopher medio misura circa 2-8 m di diametro, mentre il soma medio di un animale (B4065 bzIs166[Pmec-4::mCherry]) misura 6-10 m nel secondo giorno degli adulti(Figura 7B). - Se non si osserva alcun germoglio o dominio esopher, ispezionare il soma neuronale per un filamento sottile attaccato emanato dal soma. Gli esopher attaccati tendono ad essere situati più vicino al soma originario e in un piano z simile.
NOTA: gli esopher non sempre rimangono attaccati al soma. Il rilevamento di un filamento collegato è un'indicazione definitiva che l'oggetto è un esopher. - Per identificare un esopher non collegato, cercare il contenuto di un esopher. Gli esopher possono concentrare le proteine fluorescenti espulse e sono quindi spesso più luminose del soma.
NOTA: il contenuto degli esopher è eterogeneo e variabile. Gli organelli cellulari come i lisosomi e i mitocondri possono anche essere estrusi all'interno di esodi (Figura 4C-E). - Cercare esodi non collegati in piani focali diversi rispetto al piano in cui è stato trovato il soma originario. Anche se gli esopher sono stati visti sporgere dal soma ALM in qualsiasi direzione, è tipico che gli esopher sporgono lontano dal soma, in una direzione posteriore dal processo neuronale.
- Verificare la presenza di oggetti sferici di grandi dimensioni che non sono posizionati e identificati come somi neuronali. Gli esopher possono essere di forma irregolare, ma sono tipicamente strutture sferiche. Gli esopher si degradano nel tempo, quindi gli esopher più vecchi tendono ad avere una forma più irregolare.
NOTA: Gli esopher maturi o più vecchi si distinguono dal palco disperso della "notte stellata" attraverso l'intensità di fluorescenza più luminosa degli esopher e la loro forma sferica. - Indaga sul fenotipo della "notte stellata" come prova di una precedente esofradnesi. Gli esopher progrediscono in una fase di "nottestellata" mentre l'esopher si rompe in vesciche più piccole e l'ipodermide circostante tenta di degradare il contenuto dell'esopher ( Figura1G, 2B, 3B & 7A).
NOTA: Lo stadio notturno stellato è caratterizzato da entità fluorescenti frammentate e sparse (a volte in rete) che hanno perso l'integrità strutturale e mostra una dem fluorescenza rispetto ai neuroni touch e alle strutture esopher. - Cercare le istanze di "eventi esopher multipli". Gli esopher sono di solito prodotti come un evento singolare (1 esopher emanato da 1 soma), ma in alcune circostanze più di un esopher può essere rilasciato da un singolo soma (Figura 7D).
NOTA: Gli esopher maturi possono degradarsi in vesicelle multiple in quanto sono degradati nell'ipodermide. Distinguendo se ogni esopher è stato generato da un evento esophergenesi indipendente o se una divisione esopher originale per creare una vescica aggiuntiva può essere determinata solo dall'osservazione time-lapse. - Tenete a mente che non tutte le anomalie morfologiche maturano in esophers.
- Non segnare un soma disassodato come un esopher. Un soma esteso o appuntito può essere osservato a volte (soprattutto con l'età o sotto stress), ma un'estensione senza un sito di costrizione chiara non è segnata come un esopher.
- Rifiutare piccole gemme risolte che non raggiungono 1/5th la dimensione del soma nella quantificazione dell'evento exopher.
- Non contare le escrescenze neurite come esophers. I neuriti maturi possono estendersi drammaticamente con l'età (di solito nella direzione opposta del processo neuronale) e la proteina fluorescente può migrare verso l'estremità distale di talistrutture 19.
NOTA: Queste escrescenze neurite non sono esopher in quanto hanno un modello di sviluppo distinto per giorni e settimane, non formano gemme e non si staccano (Figura 7E).
- Identificare le entità fluorescenti che non sono esopher.
NOTA: è importante avere un'idea della fluorescenza di fondo per garantire la corretta identificazione dell'entità fluorescente estrusa rispetto all'autofluorescenza.-
Espressione fluorescente transgenica vs autofluorescenza. Non confondere l'autofluorescenza per l'espressione transgenica. Il vero segnale esopher non sarà nell'intestino o nell'intestino (la conferma DIC può essere utilizzata per identificare questi tessuti) e il segnale esopher sarà significativamente più luminoso dell'autofluorescenza di fondo.
NOTA: L'autofluorescenza è causata dalla pigmentazione fluorescente intestinale del granulo intestinale e si accumula con l'età. È eterogeneo, soprattutto se visto con diverse lunghezze d'onda. - Segnale dagli embrioni. Non confondere il segnale dell'embrione per l'esoferegenesi. Confermare i sospetti di segnale embrionale passando dalla fluorescenza all'illuminazione del campo luminoso e controllando le associazioni di segnale con le uova nell'utero.
-
Fuori dall'aereo o corpi di Soma nelle vicinanze. Evitare di confondere un soma fuori dall'aereo per un esopher identificando tutti i corpi soma vicini, anche somi fuori fuoco all'inizio dell'osservazione.
NOTA: se il punteggio per esophers da ALMR, identificare e tenere conto della posizione di AVM e ALMR somas. Ulteriori dettagli sull'identificazione del corpo soma sono descritti nella Figura 3A.
-
Espressione fluorescente transgenica vs autofluorescenza. Non confondere l'autofluorescenza per l'espressione transgenica. Il vero segnale esopher non sarà nell'intestino o nell'intestino (la conferma DIC può essere utilizzata per identificare questi tessuti) e il segnale esopher sarà significativamente più luminoso dell'autofluorescenza di fondo.
8. Punteggio e statistiche
- Punteggio esophers come binario (sì, c'è un esopher / no, non c'è un esopher).
- Considerare il rilevamento esopher come un "exopher-event" per un determinato neurone. Un evento esopher può costituire l'osservazione di un singolo esopher vicino a un soma o a più esopher.
NOTA: per quantificare il numero di singoli eventi di esophergenesi, utilizzare l'osservazione time-lapse. - Contare gli eventi esopher per una particolare cella identificata perché celle diverse non producono esopher alla stessa velocità (vedere ad esempio Figura 3C). I neuroni ALMR producono gli esopher più basali nei ceppi descritti nel presente documento e quindi spesso questa è la cellula selezionata per la quantificazione esopher dai neuroni del recettore del tocco.
- Per le statistiche, in generale, condurre almeno 3 prove biologiche, di almeno 30 animali segnati per prova con il corrispondente numero di osservazioni necessarie per l'analisi della perturbazione.
- Per più prove che coinvolgono uno o due mutanti/trattamenti rispetto al controllo, il test Cochran-Mantel-Haenszel è appropriato per determinare i valori p.
- Per le sperimentazioni che coinvolgono più di due mutanti di trattamenti rispetto al controllo, è anche opportuno utilizzare un'analisi binaria di regressione logistica per valutare la significatività per qualsiasi numero di predittori categori.
Risultati
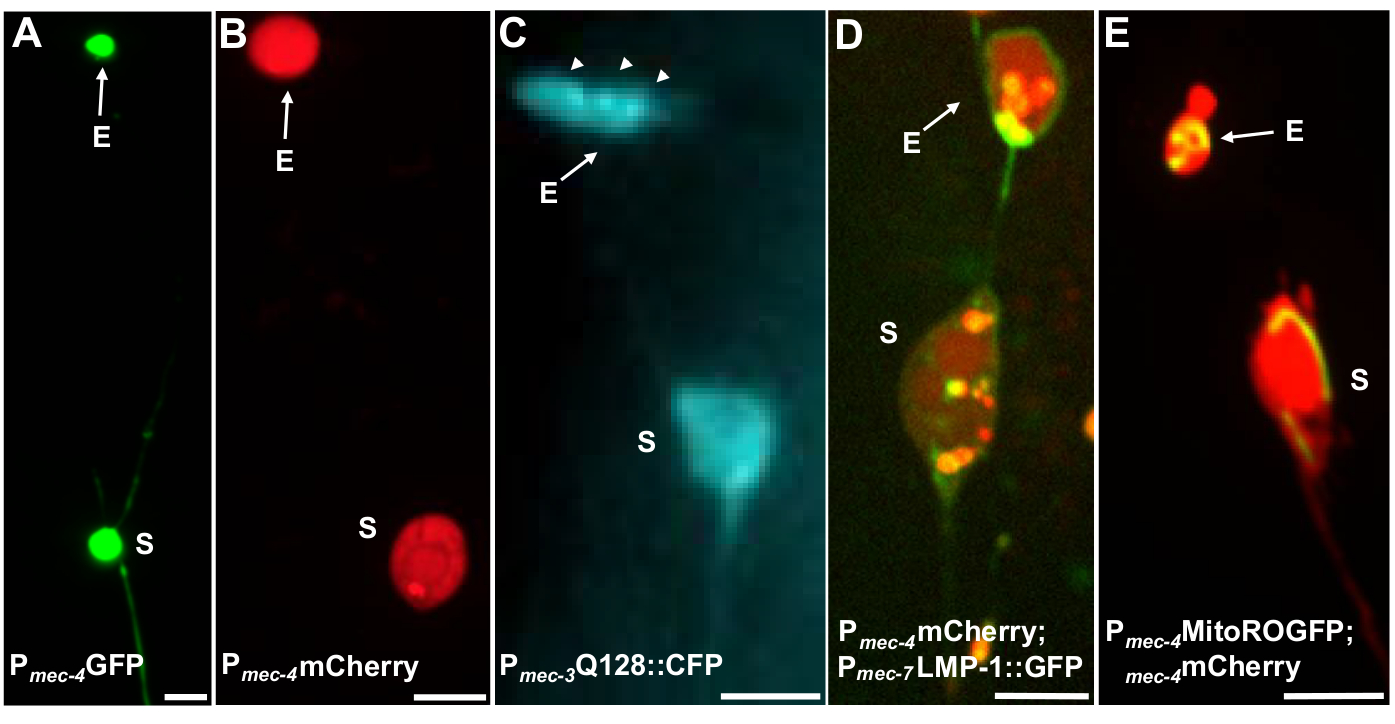
Più reporter fluorescenti possono essere utilizzati per misurare gli esopher. Gli esodati di neuroni tattili sono facilmente visualizzati in vivo attraverso l'etichettatura fluorescente delle proteine che possono essere selezionate per l'estrusione, etichettando organelli che possono essere estrusi o etichettando le membrane cellulari. La tabella 1 identifica i reporter fluorescenti che sono stati utilizzati per monitorare gli esopher, con esempi rappresentativi inclusi nella Figura 4. I carichi che sono noti per essere estrusi negli esopher includono una fusione del dominio N-terminale della huntingtina umana alla poliglutamina espansa (Q128) (Figura 4B), lysosomi che sono GFP contrassegnati con proteina di membrana associata al lisosomiale (LMP-1) (Figura 4C) e mitocondri con GFP con codificata a matrice (Figura4D). Cytoplasmic GFP non è fortemente espulso ed è preferibilmente mantenuto nel soma5, anche se GFP può debolmente visualizzare esosti(Figura 4A). Quando GFP è fusa con proteine che vengono espulse, questo tag può essere utilizzato per visualizzare gli esopher. Un punto importante è che etichettando diverse proteine, una vasta gamma di domande sull'espulsione di specifici carichi e organelli, così come sulle proteine e le membrane che costituiscono gli esopher, possono essere affrontate.
Una configurazione pseudo-stereomicroscope è uno strumento efficace per la visualizzazione di esopher negli animali su piastre di agar. Questa configurazione è un ibrido di tecnologia composta e stereoscopica che include alte ottiche ad apertura numerica su ogni ingrandimento, tecnologia pseudo-stereo (obiettivi discreti su una base stereoscopica) e un interruttore operativo zoom per la visualizzazione a ingraziati intermedi agli obiettivi installati. Un microscopio come questo dovrebbe essere dotato di 10x oculurti e obiettivi abbastanza potenti per osservare la morfologia neuronale e la produzione di esopher per il punteggio ad alta velocità effettiva (2x obiettivo utilizzato per la scansione / raccolta, obiettivo 10x utilizzato per l'identificazione e il punteggio).
Mentre le capacità di ingrandimento degli stereomicrossi standard in genere hanno una risoluzione sufficientemente elevata per vedere la rete di neuroni touch che esprimono proteine fluorescenti, i microscopi standard di sezionazione non sono sufficienti per osservare i dettagli subcellulari degli esopher come le connessioni tubolari del soma da esopher. Tali osservazioni richiedono la microscopia confocale (vedere la Tabella dei Materiali per i dettagli delle attrezzature).
Gli studi di quantificazione dell'esopher richiedono controlli rigorosi per eliminare le sollecitazioni sperimentali. Per la produzione di esopher riproducibile è necessario un attento mantenimento di condizioni di crescita costanti. Più specificamente, la produzione di esopher è sensibile allo stress, in modo tale che l'alimentazione costante, la temperatura costante e la crescita senza contaminazione tra le generazioni sono fondamentali per la riproducibilità. In condizioni di crescita basale con elevata espressione neuronale di mCherry, la produzione di esopher è relativamente bassa (5-25% degli ALMR producono esopher) ma alcune sollecitazioni, tra cui lo stress osmotico e ossidativo, possono aumentare i tassi di esopher. Mentre l'espressione mCherry può essere pensata come stress, un corollario della sensibilità allo stress dei livelli di esopher è che, se adeguatamente controllata, l'introduzione dello stress sperimentale può essere una strategia per indurre e osservare più facilmente l'esoferegenesi.
Tempismo e livelli di produzione esopher previsti. Gli esopher sono praticamente assenti durante lo sviluppo larvale. Il periodo di picco della produzione di esopher nella vita dei giovani adulti sembra essere molto limitato durante i giorni adulti 1-4, più comunemente evidente al giorno adulto 2 o 3. Poiché il picco può spostarsi un po' avanti o indietro, la valutazione più completa di un profilo di produzione esopher consiste nel segnare più prove al giorno nei giorni adulti 1-4. In generale, un ALMR produce un importante esopher, con la vescica persistente per almeno 24 ore. L'esopher può essere prodotto abbastanza rapidamente (nell'ordine di minuti al suo più veloce). Più comunemente, solo un exopher principale è prodotto per neurone nella prima vita adulta, ma la produzione di esopher multipli è possibile.
In generale, la produzione di esopher da parte degli ALMR che esprimono mCherry in condizioni basali varia dal 5-25% degli ALMR esaminati entro il periodo di tempo ottimale del giorno adulto 2-3 (Figura 3D). Crisi di proteostasi5, così come l'esposizione ad altre sollecitazioni possono modulare il livello di esopher. Lo stress o le perturbazioni genetiche possono aumentare la produzione di esodiche a tassi di rilevamento fino al 90% dei neuroni ALMR che producono estrusioni esopher.
RNAi a base di alimentazione per testare ruoli di geni specifici nell'esofrafere. Il nematode C. elegans è comunemente sottoposto a RNAi abbattere alimentando gli animali trasformati E. coli CEPPO HT115 che esprimono un doppio RNA spiaggiato (dsRNA) mirando un gene di interesse20. I batteri HT115 possono essere utilizzati quando si segnano per esopher nell'alimentazione RNAi5. Mentre le trascrizioni nella maggior parte dei tessuti possono essere prese di mira da RNAi utilizzando questa tecnica, i neuroni sono più refrattari. La sensibilità all'RNAi può essere calibrata utilizzando animali che esprimono il trasportatore di dsRNA transgenico SID-1 sotto un promotore specifico del neurone. In questo modo il tessuto neuronale può essere sensibilizzato a RNAi21.
Il knockdown specifico del tessuto di un gene di interesse può essere realizzato esprimendo una componente del metabolismo endogeno dell'RNAi all'interno di un mutante che è carente in quel componente. Ad esempio: la proteina Argonauta RDE-1 può essere espressa specificamente nei neuroni degli animali mutanti rde-1 per ottenere il knockdown di un gene di interesse solo nei neuroni quando gli animali sono esposti a un intervento RNAi mirato a quel gene.
Utilizzando i protocolli RNAi nematodi standard20,22, l'esposizione dei genitori allo stadio L4 all'RNAi e permettendo alla loro progenie di sviluppare batteri HT115 trasformati consumanti fino all'età adulta genera il forte knock-down genetico, ma essere attenti a potenziali ritardi di sviluppo indotti da RNAi come animali sperimentali possono crescere in modo diverso rispetto a un controllo vettoriale vuoto. È importante includere sempre il controllo vettore vuoto per il confronto dei controlli negativi. I batteri HT115 possono essere utilizzati quando si segna per gli esopher nell'alimentazione dell'RNAi. Tuttavia, si noti che alcuni geni sono efficaci nel cambiare i tassi di esophergenesi anche durante periodi più brevi di esposizione RNAi5. Se il targeting di alcuni geni porta a un fallimento dello sviluppo, evitare di esporre gli animali a un knockdown permanente, gli animali possono semplicemente essere raccolti allo stadio L4 su piastre RNAi per l'esposizione da L4 a D2 o D3 adulti.
| Nome deformazione | Genotipo | Descrizione | Percentuale esopher | Riferimento |
| SK4005 | zdIs5[Pmec-4GFP] | Espressione citosolica di GFP nei neuroni touch. | 1-8% ALM | Figura 4A, Melentijevic 2017 |
| B4065 | bzIs166[Pmec-4::mCherry] | La sovraespressione di mCherry (bzIs166) nei neuroni touch produce sia segnali citosolici che aggregati mCherry. bzIs166 è un induttore esore. Gli aggregati mCherry sono predittori di esophergenesi e sono preferenzialmente estrusi negli esodati. | 3-20% ALM (condizioni normali). 20-80% ALM (condizioni di digiuno). | Figura 4B, Melentijevic 2017 |
| B4067 | bzIs167[Pmec-4mitogfp Pmec-4mCherry4]; igIs1[Pmec-7YFP Pmec-3htt57Q128::cfp lin-15']; | YFP etichetta citosolmente i neuroni touch mec-7. Co-espresso Q128::CFP aggrega e induce esophers. CFP silenzia preferibilmente. | 25% | Figura 4C, Meletijevic 2017 |
| N. 4509 | bzIs166[Pmec-4mCherry]; bzIs168[Pmec-7LMP-1::GFP] | bzIs168 LMP-1::GFP etichetta le membrane plasmatiche e le membrane lisosomiche. bzIs168 può essere utilizzato per identificare le membrane neuronali, gli esopher (in quanto sono legati alla membrana) e le strutture a membrana lisosomica. | 3-20% ALM | Figura 4D, Melentijevic 2017 |
| N. 4528 | bzIs166[Pmec-4mCherry]; zhsEx17 [Pmec-4mitoLS::ROGFP] | Allele zhsEx17 è un reporter localizzato mitoconndrialmente che cambia la sua lunghezza d'onda di eccitazione di picco da 405nm (ossidato) a 476nm (ridotto) secondo l'ambiente ossidativo locale. Si esprime nei neuroni touch e può essere utilizzato da solo per identificare i mitocondri nei neuroni touch e nei mito-esopher. | 3-20% PROteo-esopher ALM. % quantitazione MITO-esopher ALM in corso. | Figura 4E, Melentijevic 2017, Cannone 2008, Ghose 2013 |
Tabella 1. Ceppi che sono stati utilizzati per la visualizzazione di neuroni tattili, toccare neurone-esophers, e contenuti esopher.

Figura 1: Fasi dell'esophergenesi. Il processo di creazione ed espulsione di un esopher è chiamato "exopher-genesis". Il processo dinamico di formazione degli esopher può richiedere da diversi minuti a diverse ore. Sono raffigurati esempi di soma ed esopher morfologia a passi specifici durante il processo di esophergenesi dinamica in un ceppo di produzione ad alta esopher, .B4065 bzIs166[Pmec-4mCherry]. Tutte le immagini sono del giorno 2 neuroni ALM adulti presi con un obiettivo 100x. (A) Normale soma. Adulto tocco meccanosensoriale neurone ALM transgenicamente esprimendo Pmec-4mCherry. La morfologia soma raffigurata è tipica dei neuroni giovani adulti in questo ceppo, con concentrazioni di mCherry nel citoplasma. (B) Fase iniziale del germoglio. Il primo passo osservabile di esophergenesi consiste nella polarizzazione di materiale citoplasmico selezionato al bordo della membrana soma. Questo passaggio è spesso accompagnato da un'espansione o gonfiore del soma. Nel caso dei neuroni touch, il dominio pre-esopher (PED) si estende nell'ipodermide circostante (non visibile qui). Si noti la maggiore concentrazione di materiale mCherry nel dominio delle prime gemme. (C) Fase di bocciolo tardivo. Dopo un'ulteriore polarizzazione cellulare e un'espansione del dominio pre-esopher, una costrizione tra il soma e l'esopher (freccia) diventa evidente. Questo evento segnala la transizione alla fase di bocciolo in ritardo. Anche se nella fase di tarda bocciolo la cellula mostra un chiaro sito di costrizione e domini soma ed esopher separati, non è ancora pizzicato completamente dal soma; l'esopher in erba può essere attaccato da un gambo spesso (freccia). Il dominio in erba è considerato un esopher precoce quando il diametro del dominio esopher in questione è circa 1/3 più grande del diametro del cantiere / gambo. (D) Fase di esopher precoce. I primi esopher possono essere attaccati da un gambo dal soma in partenza: il diametro di questa connessione può assottigliarsi man mano che l'esopher si allontana dal soma. Il materiale citoplasmico può essere trasferito dal soma all'esopher attraverso questo tubo, anche se la maggior parte del materiale viene caricato durante il processo di germogliazione. Gli esopher possono staccarsi dal soma come raffigurato in (E), gli esopher separati sono considerati esopher maturi (F). L'esopher maturo può transitare attraverso il tessuto ipodermico circostante, allontanandosi dal soma in partenza. (G) La ripartizione dell'esopher etichettata mCherry in vescicle più piccole all'interno dell'ipodermide si traduce in un aspetto puntuale sparso del materiale mCherry, molto probabilmente quando entra nella rete endolisosomica ipodermica. Il segnale di foratura disperso è chiamato la fase "notte stellata". La degradazione di alcuni contenuti esopher è probabilmente compiuta da lisosomi ipodermici, ma alcuni materiali non sono completamente degradati ed è spesso riesstruito dagli ipodermi nello pseudocoelom. Il transito mCherry post-exophergenesis è descritto in modo più dettagliato nella Figura 2. Si prega di fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 2: mCherry estruso dai neuroni tattili negli esopher coinvolge la rete lisosomica ipodermica circostante, ma può essere successivamente estruso nello pseudocoelom dove i coelomociti possono immagazzinare/degradare l'mCherry. (A) Riassunto del cartone animato di come mCherry estruso negli esomi transita nel corpo dopo l'espulsione da parte dei neuroni. Durante l'esophergenesi selezionato contenuto cellulare come mCherry diventano localizzati e germogliare fuori dal soma neuronale di invio in una vescicola indipendente circondato dalle membrane plasmatiche neuronali e ipodermiche. Poiché i neuroni touch sono incorporati nel tessuto ipodermico, come il dominio esopher germogli verso l'esterno si muove ulteriormente nell'ipodermide. L'esopher può transitare l'ipodermide, e dopo ore a giorni, il contenuto di esopher può frammentarsi all'interno della rete endolisomica dell'ipodermide. L'mCherry può apparire come puncta sparsi in tutto l'ipodermide, un palco chiamato "notte stellata". Dopo alcuni giorni, alcuni dei mCherry possono passare dall'ipodermide nello pseudocoelom circostante, dove le cellule scavenger chiamate coelomociti possono accedere e prendere mCherry che possono essere conservate. (B) Esempio dell'aspetto delle vescie mCherry notte stellate. Immagine di un soma ALM etichettato con mCherry con grandi frammenti di esopher e vesciche notturne stellate. La deformazione è B4065 bzIs166[Pmec-4mCherry]. (C) Esempio di concentrazione mCherry in coelomociti distanti. Vista laterale di un animale adulto giorno 10 di ceppo B4065 bzIs166[Pmec-4mCherry] che mostra mCherry concentrato in coelomociti (frecce). Alcune vesicles notturne stellate sono anche evidenti. In generale la concentrazione di coelomociti diventa evidente dopo circa il giorno 5 della vita. (B in basso) Riproduzione a cartoni animati di (B), con neuroni touch e processi delineati in rosso, così come i frammenti esopher più luminosi; piccole vesicles sparse di diverse profondità di z sono mostrate in rosa più chiaro. (C in basso) Versione cartoon dell'immagine di (C), che mostra il processo neuronale in rosso, notte stellata in rosa e coelomociti in verde. Si prega di fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 3: I neuroni del tocco meccanosensale producono esopher a diversi livelli con un profilo temporale preciso. (A) (Top) rappresentazione a cartoni animati dei neuroni del tocco meccanosensoriale in relazione spaziale ai principali punti di riferimento anatomici di C. elegans, tra cui il pharynx di pompaggio e l'anello nervoso ad alta densità neuronale alla testa dell'animale, la vulva nel corpo medio e la coda affusolata. (In basso) Neuroni touch fluorescenti etichettati che esprimono GFP come visti dal lato superiore e sinistro (immagini adattate da WormAtlas). La scatola rossa raffigura l'area in cui si trovano in genere gli esopher ALM. (B) Alta vista di ingrandimento della regione del corpo medio in cui gli esopher derivati da ALM sono prodotti in un ceppo che esprime [Pmec-4mCherry]. AVM e ALMR neurone sono raffigurati, e mostrato è un esopher ALMR insieme a mCherry notte stellata. I neuroni ALMR producono più facilmente esopher. (C) I neuroni del tocco meccanossensoriale ALMR producono più facilmente esopher rispetto ad altri neuroni tattili in erefroditi in condizioni basali. Mechanosensory touch neurone esopher produzione il giorno adulto 2, come segnato per singoli neuroni recettori tocco è indicato. Ceppo: B4065 bzIs166[Pmec-4mCherry], N>150, barre di errore sono SEM. (D) I neuroni tattili ALMR producono più esopher durante i giorni 2 e 3 dell'età adulta rispetto allo stadio L4 adolescente o con gli animali in età avanzata. Ceppo: B4065 bzIs166[Pmec-4mCherry], N>150, le barre di errore sono SEM. Fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 4: Esempi di alcuni reporter fluorescenti che taggano contenuti esopher. Un modo semplice per osservare gli esopher è la creazione di animali transgenici che esprimono fluorofori dai promotori neuronali. I fluorofori consentono la visualizzazione dell'espressione esopher e transgenica induce l'aggregazione e/o la proteostress che aumenta l'esophergenesi. Gli esopher prodotti dai neuroni anfiidi possono anche essere osservati in condizioni native, utilizzando il riempimento dei coloranti per la visualizzazione. Sono mostrati esempi di ceppi comuni che possono essere utilizzati per osservare esophers, (E) exopher, (S) soma. (A) Soma ed esopher da un ALM di un adulto di ceppo SK4005 zdIs5[Pmec-4GFP],100x obiettivo utilizzato per la fotografia, scala bar 3m. In questo ceppo, vengono misurati gli esopher che includono GFP solubile, ma la produzione di esopher si verifica raramente. La fusione di GFP a proteine che possono essere estruse preferibilmente negli esopher in altri studi conferma che le fusioni GFP possono essere rilevate negli esophermaturi. (B) ALM soma ed esopher di un adulto di ceppo B4065 bzIs166[Pmec-4mCherry], che esprime mCherry e induce la produzione di esodiche neuronali touch. Obiettivo 100x utilizzato per la fotografia, scala barra 5 m. (C) ALM soma ed esopher di un adulto di ceppo - B4067 bzIs167[Pmec-4mitogfp Pmec-4mCherry4]; igIs1[Pmec-7YFP Pmec-3htt57Q128::cfp lin-15]; canale blu selettivo utilizzato per l'immagine di htt57Q128::CFP. L'esopher contiene aggregati htt57Q128::CFP (frecce), che appaiono più concentrati nell'esopher che nel soma. Obiettivo 40x utilizzato per la fotografia, scala barra 5m. (D-E) Gli esopher possono contenere organelli e tag specifici per organelli con proteine fluorescenti che consentono il monitoraggio dell'estrusione degli organelli. (D) Tag membrana lisosomica LMP-1::GFP delinea il soma e la membrana esopher e tag membrane plasmatiche debolmente (localizzazione della membrana plasmatica è un passo di traffico sulla strada per il targeting lisosomico) ed etichetta fortemente organelli lisosomili. Mostrato è un soma ALM adulto che co-esprime Pmec-4mCherry e il Pmec-7LMP-1::GFP che si localizza a membrane e lisosomi. Il soma ha un esopher collegato con altre estrusioni più piccole che potrebbero essere frammenti esopher (frecce). Le strutture positive GFP sono incluse nel soma e sono presenti nel grande esopher, ceppo: B4509 bzIs166[Pmec-4mCherry]; bzIs168[Pmec-7LMP-1::GFP]. Obiettivo 100x utilizzato per la fotografia, scala barra 5 m. E) Un marcatore GFP mitocondriale può essere utilizzato per identificare i mitocondri in soma ed esopher. Mostrato è un soma ALM adulto che esprime Pmec-4mCherry e mito::ROGFP, che si localizza alla matrice mitocondriale. mito::ROGFP espresso da solo, senza il mCherry, può anche essere facilmente utilizzato per identificare i neuroni e punteggio per esophers che includono mitocondri. Ceppo: B4528 bzIs166[Pmec-4mCherry]; zhsEx17 [Pmec-4mitoLS::ROGFP]. Obiettivo 100x utilizzato per la fotografia; barra della scala 5m. Si prega di fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 5: Ciclo di sviluppo di C. elegans e identificazione L4. (A) A 20 gradi centigradi un uovo impiega circa 9 ore per schiudersi una volta deposto dalla madre. (B) Un animale appena schiuso è in fase larvale 1 (L1) e muta in una larva L2 dopo 12 ore. (C) Gli animali rimangono nelle fasi larva L2 eD) L3 per circa 8 ore ciascuna. (E) Gli animali adolescenti sono considerati il quarto stadio larvale (L4) e sono contrassegnati da una cospicua vulva in via di sviluppo che appare come una mezzaluna bianca vicino al corpo medio. La presenza di questo mentre mezzaluna consente una facile identificazione e raccolta di L4 animali in scena per stabilire colture sincronizzate che in seguito facilitano il punteggio per gli esopher. Gli animali rimangono nella fase L4 per circa 10 ore prima della loro muta finale in adulti gravide, F) identificati dallo sviluppo di uova, spermatheca visibile, e l'inizio della deposizione delle uova. Si prega di fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 6: Preparazione del microscopio slide agar pad. (A) Preparare due diapositive con una singola striscia di nastro da laboratorio posizionato lungo la parte superiore. Posizionare una diapositiva al microscopio non registrato in mezzo come illustrato. B) Posizionare una goccia di agarose fuso sulla parte superiore della diapositiva. (C) Posizionare delicatamente una diapositiva pulita sulla parte superiore della goccia, premendo l'agarose in un cerchio sgonfio. (D) Rimuovere le diapositive toccate, che agiscono per realizzare un appiattimento uniforme dell'agar che è necessario per creare un pad uniforme. (E) Rimuovere la diapositiva superiore una volta che il pad agarose si è asciugato. (F) Pipetta una soluzione paralitica (levamisole o tetramisole) sulla parte superiore del pad agar. (G) Scegli gli animali opportunamente messi in scena nel paralitico. (H) Coprire delicatamente gli animali con un coperture e garantire che gli animali siano vivi. Si prega di fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 7: Caratteri di esopher e criteri di identificazione esopher. (A) Criteri generali che identificano un esopher. (B) Confronto di diametro tra il soma di invio e l'esopher estruso, misurato in m. Somi ALM adulti, N-35, ceppo: B4065 bzIs166[Pmec-4mCherry] - 6,53 m dimensione media di soma e 3,83 m dimensione media di esopher. (C) Definizione dei criteri per la differenziazione tra un dominio esopher e un esopher in erba. (D) Più comunemente, i singoli neuroni fanno un grande esopher, che in seguito si divide o frammenti come l'ipodermide tenta di degradare il suo contenuto. Ancora, più esophers possono essere osservati accanto a un neurone tocco che potrebbe derivare da più eventi esopher da un neurone o in alternativa, esophers possono anche germogliare o frammentarsi. L'origine di più entità simili a esopher può essere determinata solo utilizzando la microscopia time lapse. Top raffigura un soma neurone al tocco ALMR con un singolo esopher distante. In basso raffigura un soma neurone al tocco ALMR con più estrusioni esopher-like. (E) Caratteristiche morfologiche comuni nei somi del neurone ALM per adulti che possono essere scambiati per eventi esopher. In alto a sinistra - Un soma ALM disastento, senza un dominio esopher chiaro o un sito di costrizione. Mezzo superiore - I neuroni possono avere piccole protrusioni extracellulari che possono essere analoghe agli esopher, ma non soddisfano i criteri di requisito di dimensione per essere considerati un esopher. In alto a destra – Con l'età, i neuroni tattili possono sviluppare le crescita lungo il loro neurite minore. Spesso il materiale mCherry può essere raccolto sulla punta della crescita neurita. Questo non è segnato come un esopher se il mCherry raccolto non soddisfa i requisiti di dimensione exopher-to-soma. In basso raffigura i neuroni ALM adulti che hanno criteri di definizione per un dominio esopher o un esopher. Botom left - ALM soma che ha un dominio exopher prominente che include selettivamente mCherry cytosol e mCherry aggregati con tag. Il sito di costrizione del dominio esopher è contrassegnato da frecce e soddisfa i criteri di dimensione (almeno 1/5th la dimensione del soma). Il diametro più grande del dominio esopher è quasi 1/3 più grande del diametro del sito di costrizione, che soddisfano i criteri per un evento esopher. In basso al centro - SOma ALM che ha un prominente esopher in erba che soddisfa i criteri di dimensione. C'è un chiaro sito di costrizione. In basso a destra - ALM soma che ha un exopher riempito mCherry allegato che soddisfa i requisiti di dimensione esopher. L'esopher è attaccato da un sottile filamento di collegamento. Tutte le immagini provengono da ceppo B4065 bzIs166[Pmec-4mCherry]. Si prega di fare clic qui per visualizzare una versione più grande di questa figura.
{kind=link}
Discussione
La caratterizzazione dei meccanismi molecolari in vivo di eliminazione aggregata e organello sotto forma di grandi esopher è agli inizi. Le domande sulla designazione dei carichi per l'espulsione, la raccolta polarizzata di questi carichi all'interno della cellula, la regolazione della decisione di generare esodiche, i macchinari che mediano le estrusioni e l'interazione degli esopher con i macchinari degradativi in una cella vicina rimangono tutti da affrontare. Inoltre, la visualizzazione in vivo delle connessioni tubolari che possono passare materiali biologici che includono calcio, aggregati e mitocondri è interessante e la biologia poco studiata a sé stante. Anche le domande sul perché alcune cellule sono più inclini alla produzione di esopher rispetto ad altre sono irrisolte, ma possono iniziare a essere geneticamente sezionate con gli approcci delineati in questo protocollo.
Descritti in dettaglio in questo protocollo sono gli approcci per ottenere il punteggio riproducibile della produzione di esopher, con attenzione a distinguere gli esopher dai somi cellulari vicini, la tempistica delle analisi per catturare il picco di produzione di esopher e il rigoroso controllo delle condizioni di crescita per eliminare le sollecitazioni indesiderate che possono modulare i livelli di esodiche. Sia la distinzione del grande esopher precoce, sia la dispersione della "notte stellata" nell'ipodermide circostante possono essere quantificate come prova della produzione di esopher. Detto questo, neuroni che esprimono mCherry in condizioni basali sono più spesso associati con 5-25% dei neuroni di un tipo specifico che producono un esopher. L'introduzione controllata delle condizioni di stress potrebbe essere applicata per aumentare la produzione di esopher al rilevamento fino al 90% dei neuroni che producono estrusioni, particolarmente utile per gli schermi genetici o farmacologici per i modificatori.
Nella malattia neurodegenerativa umana, grandi aggregati possono trasferire dai neuroni matidi nelle cellule vicine per promuovere la diffusione della patologia. Il meccanismo di esodatoro potrebbe trasparire tramite un meccanismo conservato utilizzato per l'estrusione aggregata attraverso la fita. Definire le molecole in vivo che migliorano l'efficienza di questo processo (considerato un controllo della proteostasi più efficace) o bloccano potrebbe essere sfruttato per influenzare la progettazione di nuove strategie per combattere più malattie neurodegenerative. Come tale, il protocollo qui descritto potrebbe essere utilizzato per schermi di mutagenesi genetica classica, schermi RNAi a livello di genoma che abbatteno sistematicamente i geni per identificare potenziatori e soppressori, o per studi di intervento farmacologico che identificano i modificatori farmacologici candidati di questo processo. L'approccio è semplice, anche se un po 'faticoso. Gli esopher sono così grandi che possono essere visualizzati con un microscopio di sezionazione ad alto ingrandimento. Ancora, I neuroni C. elegans sono relativamente piccoli e guardando i loro organelli o le loro membrane richiedono immagini confocali di maggiore potenza ed è un processo lento. Le opzioni per una maggiore velocità effettiva potrebbero comportare approcci di imaging ad alto contenuto in formato multi-well plate.
L'applicazione di un approccio standardizzato al punteggio esopher dovrebbe essere alla base di una dissezione genetica concertata del processo mediante il quale i neuroni possono organizzare ed eliminare i detriti cellulari.
Divulgazioni
Nessuno
Riconoscimenti
Riconosciamo le seguenti sovvenzioni NIH: R01AG047101 e R37AG56510. I membri dei laboratori Driscoll e Grant hanno contribuito ampiamente allo sviluppo e alla messa a punto dei protocolli descritti, con esperimenti rigorosi e una forte comunicazione.
Materiali
| Name | Company | Catalog Number | Comments |
| 95B Scientific CMOS camera | Photometrics Prime | ||
| 1,000 μL low retention tips | Sarstedt | ||
| 10 mL serological pipette | Appleton Woods | CC214 | |
| 10 μL low retention tips | Sarstedt | 70.1130.105 | |
| 13% sodium hypochlorite | Acros Organics | AC219255000 | |
| 15 mL centrifuge tubes | Fisher Scientific | 05-539-12 | |
| 2 L erlenmeyer flasks | Scientific Laboratory Supplies | FLA4036 | |
| 25 mL serological pipette | Appleton Woods | CC216 | |
| 300 μL low retention tips | Sarstedt | 70.765.105 | |
| 50 mL serological pipette | Appleton Woods | CC117 | |
| 5-Fluoro-2'-deoxyuridine 98% | Alfa Aesar | L16497.ME | |
| 9 cm sterile Petri dishes | Fisher Scientific | 11309283 | |
| absolute ethanol | Vwr | 20821.33 | |
| Agar | Sigma Aldrich | A1296 | |
| C. elegans strain wild type | Supplied by CGC | N2 | C. elegans strain |
| calcium chloride dihydrate | Sigma Aldrich | C3881 | |
| cholesterol | Acros | 110190250 | |
| dibasic sodium phosphate | Sigma Aldrich | S3264 | |
| E. coli strain OP50 | Supplied by CGC | Op50 | E coli strain |
| FBS10 Standard microscope | Meyer Instruments | KSC 410-1-100-1 | FBS10 Standard with Plate Base, 100/100 Trinocular Head and Flip zoom |
| glass pipette 270 mm | Fisherbrand | FB50255 | |
| Heraeus Multifuge X3R | Thermofisher scientific | 75004515 | |
| Inoculating Spreaders | Fisher Scientific | 11821741 | |
| LB medium capsules | MP biomedicals | 3002-031 | |
| LDI – Laser Diode Illuminator | 89 North | ||
| levamisole | Sigma Aldrich | 16595-80-5 | |
| M4 multipette | Eppendorf | 4982000012 | |
| magnesium sulphate | Sigma Aldrich | M7506 | |
| monobasic potassium phosphate | Sigma Aldrich | P0662 | |
| Multitron Standard shaking incubator | Infors HT | INFO28573 | |
| Nalgene 1 L Centrifuge pots | Fisher Scientific | 3120-1000 | |
| P10 pipette | Eppendorf Research Plus | 3123000020 | |
| P1000 pipette | Eppendorf Research Plus | ||
| P200 pipette | Eppendorf Research Plus | 3123000055 | |
| pipeteboy 2 | VWR | 612-0927 | |
| Polystyrene microbeads | Sigma Aldrich | MFCD00131491 | |
| RC5C plus floor mounted centrifuge | Sorvall | 9900884 | |
| Reusable ringed cytology slides | ThermoFisher Scientific | 22037242 | |
| SK4005 zdIs5[Pmec-4GFP] | contract Driscoll lab | GFP expressed in touch neurons | |
| sodium chloride | Sigma Aldrich | 13422 | |
| Sodium hydroxide | Fisher Chemical | S/4880/53 | |
| Tactrol 2 Autoclave | Priorclave | ||
| Triton-X | Thermofisher scientific | 28313 | |
| Tween 20 | Sigma Aldrich | 9005-64-5 | |
| X-Light V2 Spinning Disk Confocal Unit | CrestOptics | ||
| ZB4065 bzIs166[Pmec-4mCherry] | contract Driscoll lab | mCherry expressed in touch neurons | |
| ZB4067 bzIs167[Pmec-4mitogfp Pmec-4mCherry4]; igIs1[Pmec-7YFP Pmec-3htt57Q128::cfp lin-15+] | contract Driscoll lab | Q128 expressed in touch neurons | |
| ZB4509 bzIs166[Pmec-4mCherry]; bzIs168[Pmec-7LMP-1::GFP] | contract Driscoll lab | mitoROGFP expressed in touch neurons | |
| ZB4528 bzIs166[Pmec-4mCherry]; zhsEx17 [Pmec-4mitoLS::ROGFP] | contract Driscoll lab | autophagy marker expressed in touch neurons | |
| ZEISS Axio Vert.A1 | Zeiss |
Riferimenti
- Davis, A. A., Leyns, C. E. G., Holtzman, D. M. Intercellular Spread of Protein Aggregates in Neurodegenerative Disease. Annual Review of Cell and Developmental Biology. 34, 545-568 (2018).
- Davis, C. H., et al. Transcellular degradation of axonal mitochondria. Proceedings of the National Academy of Sciences of the United States of America. 111 (26), 9633-9638 (2014).
- Torralba, D., Baixauli, F., Sanchez-Madrid, F. Mitochondria Know No Boundaries: Mechanisms and Functions of Intercellular Mitochondrial Transfer. Frontiers in Cell and Developmental Biology. 4, 107 (2016).
- Stahl, P. D., Raposo, G. Extracellular Vesicles: Exosomes and Microvesicles Integrators of Homeostasis. Physiology (Bethesda, Md.). 34 (3), 169-177 (2019).
- Melentijevic, I., et al. C-elegans neurons jettison protein aggregates and mitochondria under neurotoxic stress. Nature. 542 (7641), 367 (2017).
- Nussbaum-Krammer, C. I., Park, K. W., Li, L., Melki, R., Morimoto, R. I. Spreading of a prion domain from cell-to-cell by vesicular transport in Caenorhabditis elegans. PLoS Genetics. 9 (3), 1003351 (2013).
- Tyson, T., et al. Novel animal model defines genetic contributions for neuron-to-neuron transfer of alpha-synuclein. Scientific Reports. 7, (2017).
- Babcock, D. T., Ganetzky, B. Transcellular spreading of huntingtin aggregates in the Drosophila brain. Proceedings of the National Academy of Sciences of the United States of America. 112 (39), 5427-5433 (2015).
- Pearce, M. M. P., Spartz, E. J., Hong, W., Luo, L., Kopito, R. R. Prion-like transmission of neuronal huntingtin aggregates to phagocytic glia in the Drosophila brain. Nature Communications. 6, 6768 (2015).
- Fu, H., Li, J., Du, P., Jin, W., Cui, D. Metabolic wastes are extracellularly disposed by excretosomes, nanotubes and exophers in mouse HT22 cells through an autophagic vesicle clustering mechanism. bioRxiv. 10 (1), (2019).
- Ghose, P., Park, E. C., Tabakin, A., Salazar-Vasquez, N., Rongo, C. Anoxia-reoxygenation regulates mitochondrial dynamics through the hypoxia response pathway, SKN-1/Nrf, and stomatin-like protein STL-1/SLP-2. PLoS Genetics. 9 (12), 1004063 (2013).
- Cannon, M. B., Remington, S. J. Redox-sensitive green fluorescent protein: probes for dynamic intracellular redox responses. A review. Methods in Molecular Biology. 476, 51-65 (2008).
- Perkins, L. A., Hedgecock, E. M., Thomson, J. N., Culotti, J. G. Mutant sensory cilia in the nematode Caenorhabditis elegans. Developmental Biology. 117 (2), 456-487 (1986).
- Stiernagle, T. Maintenance of C. elegans. WormBook: The Online Review of C. Elegans Biology. , 1-11 (2006).
- Sutphin, G. L., Kaeberlein, M. Measuring Caenorhabditis elegans life span on solid media. Journal of Visualized Experiments. (27), (2009).
- Mitchell, D. H., Stiles, J. W., Santelli, J., Sanadi, D. R. Synchronous growth and aging of Caenorhabditis elegans in the presence of fluorodeoxyuridine. Journal of Gerontology. 34 (1), 28-36 (1979).
- Weicksel, S. E., et al. A novel small molecule that disrupts a key event during the oocyte-to-embryo transition in C. elegans. Development. 143 (19), 3540-3548 (2016).
- Dong, L., et al. Reversible and long-term immobilization in a hydrogel-microbead matrix for high-resolution imaging of Caenorhabditis elegans and other small organisms. PloS One. 13 (3), 0193989 (2018).
- Toth, M. L., et al. Neurite sprouting and synapse deterioration in the aging Caenorhabditis elegans nervous system. Journal of Neuroscience. 32 (26), 8778-8790 (2012).
- Conte, D., MacNeil, L. T., Walhout, A. J. M., Mello, C. C. RNA Interference in Caenorhabditis elegans. Current Protocols in Molecular Biology. 109, (2015).
- Calixto, A., Chelur, D., Topalidou, I., Chen, X., Chalfie, M. Enhanced neuronal RNAi in C. elegans using SID-1. Nature Methods. 7 (7), 554-559 (2010).
- Maher, K. N., Catanese, M., Chase, D. L. Large-scale gene knockdown in C. elegans using dsRNA feeding libraries to generate robust loss-of-function phenotypes. Journal of Visualized Experiments. (79), e50693 (2013).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati