
Method Article
Approches quantitatives pour la notation in vivo Neuronal Aggregate and Organelle Extrusion in Large Exopher Vesicles in C. elegans
* Ces auteurs ont contribué à parts égales
Dans cet article
Résumé
Ce protocole décrit les approches de détection et de quantitation des grandes extrusions d’agrégats et/ou d’orgelles (~4 μm) produites par les cellules de C. elegans sous forme d’exophers à membrane. Nous décrivons les souches, les conditions de croissance, les critères de notation, le moment et les considérations de microscopie nécessaires pour faciliter la dissection de ce mécanisme d’expulsion des débris.
Résumé
La toxicité des protéines mal repliées et du dysfonctionnement mitochondrial sont des facteurs essentiels qui favorisent le déclin neuronal fonctionnel associé à l’âge et les maladies neurodégénératives entre les espèces. Bien que ces défis neurotoxiques aient longtemps été considérés comme intrinsèques aux cellules, des preuves considérables soutiennent maintenant que les protéines mal repliées sur les maladies humaines provenant d’un neurone peuvent apparaître dans les cellules voisines, un phénomène proposé pour favoriser la propagation de la pathologie dans les maladies neurodégénératives humaines.
Les neurones adultes C. elegans qui expriment les protéines agrégéssantes peuvent extruder de grandes vésicules entourées de membrane (~4 μm) qui peuvent inclure les protéines agrégées, les mitochondries et les lysosomes. Ces grands vésicules sont appelées « xopher » et sont distinctes des exosomes (qui sont environ 100x plus petits et ont une biogenèse différente). Jeter les débris cellulaires dans les exophers peut se produire par un mécanisme conservé qui constitue une branche fondamentale, mais autrefois non reconnue, de la protéostase neuronale et le contrôle de la qualité mitochondriale, pertinente aux processus par lesquels les agrégats se propagent dans les maladies neurodégénératives humaines.
Alors que les exophers ont été principalement étudiés chez les animaux qui expriment une copie élevée transgénique mCherry dans les neurones tactiles, ces protocoles sont également utiles dans l’étude de l’exophergenesis en utilisant des organites fluorescents marqués ou d’autres protéines d’intérêt dans diverses classes de neurones.
Décrit ici sont les caractéristiques physiques des exophers C. elegans, les stratégies pour leur détection, les critères d’identification, le moment optimal pour la quantitation, et les protocoles de croissance animale qui contrôlent les contraintes qui peuvent moduler les niveaux de production d’exophe. Ensemble, les détails des protocoles décrits ici devraient servir à établir une norme pour l’analyse quantitative des exophérs dans les laboratoires. Ce document vise à servir de ressource sur le terrain pour les laboratoires qui cherchent à élaborer des mécanismes moléculaires par lesquels les exopheurs sont produits et par lesquels les exopheurs sont réagis par les cellules voisines et éloignées.
Introduction
Les défis neurotoxiques des agrégats et des mitochondries dysfonctionnelles ont longtemps été considérés comme intrinsèques aux cellules, mais plus récemment, il est devenu clair que les protéines mal repliées sur la maladie humaine provenant d’un neurone peuvent également se propager aux cellules voisines, favorisant la pathologie1. De même, les mitochondries de mammifères peuvent être envoyées hors de la cellule de leur production originale pour la dégradation transcellulaire2 ou pour le sauvetage des populations mitochondriales dans les cellules voisines contestées3. Des vésicules de différentes tailles ont généralement été observées pour transférer des matériaux cellulaires vers les cellules voisines ou vers l’environnement fluide4. Certains vésicules extrudées approchent de la taille du soma neuronal moyen (soma moyen de neurone tactile ~ 6 μm) et peuvent accueillir de grands agrégats et organites.
Un exemple frappant de grande extrusion de vésicule qui peut transporter des agrégats de protéines et des organites se produit dans les neurones récepteurs tactiles de C. elegans qui expriment un nombre élevé de reporter de copie construct encodage d’une agrégation nocive-sujette, mCherry-résistant à la dégradation5. Les extrusions des neurones tactiles, appelées exophers, ont un diamètre moyen d’environ 4 μm, comprennent sélectivement mCherry ou d’autres agrégats, et sont livrées directement dans l’hypoderme voisin, qui entoure normalement les neurones récepteurs tactiles. L’hypoderme tente une dégradation à base de lysosome, mais certains contenus non digestibles tels que les agrégats de mCherry peuvent être ré-extrudés par l’hypoderme dans le pseudocoelom rempli de liquide de l’animal, à partir de laquelle le mCherry peut être absorbé par des cellules de charognard à distance appelées coélomocytes pour le stockage à long terme (Figure 1, Figure 2)5.
Les grandes vésicules extrudées exophaires quittent la cellule entourée d’une membrane plasmatique à récepteur tactile et peuvent contenir des protéines, des mitochondries et des lysosomes de maladies humaines agrégées. Le processus de production d’exopher semble impliquer le tri d’espèces potentiellement toxiques (par exemple, un mCherry exprimé sujet à l’agrégation est séparé des protéines solubles et inoffensives comme le GFP qui reste principalement dans le soma neuronal). De cette façon, l’expulsion dirigée des entités menaçantes est accomplie par le neurone5. Un défi de protéostase, tel que le stress induit par l’abattissement d’autophagie, l’inhibition protéasome médiée par MG132, ou l’expression transgénique des protéines de la maladie humaine telles que la polyglutamine étendue Q128 associée à la maladie de Huntington ou le fragment Aβ1-42impliquant la maladie d’Alzheimer, peut augmenter le nombre de neurones qui produisent des exophers5.
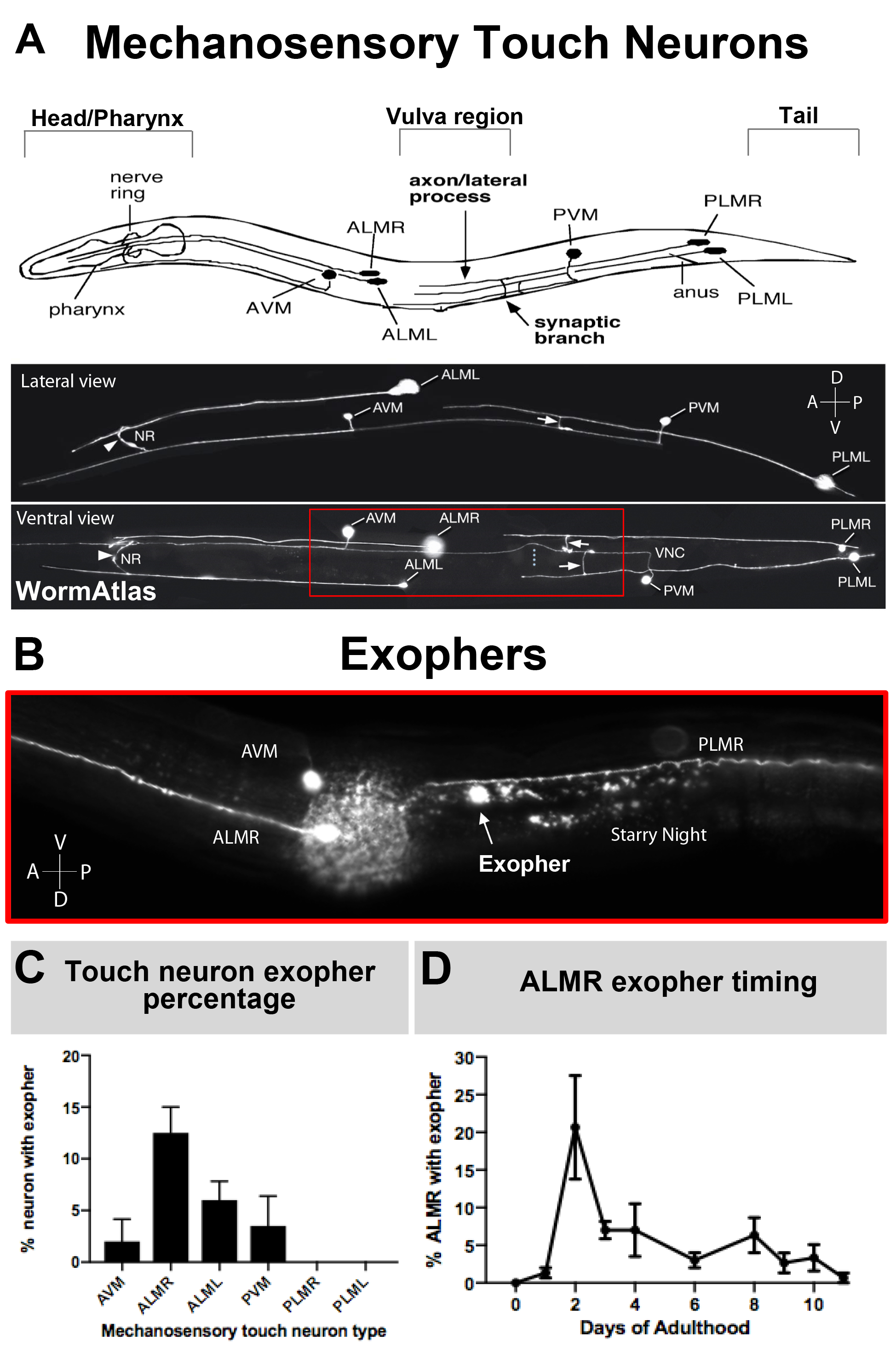
Comme les exophers n’ont été documentés que récemment, ce que l’on sait de leur biologie mérite la description. Des exophers ont été découverts dans les neurones récepteurs tactiles C. elegans les plus étudiés. Il y a six neurones tactiles mécanosensoriels C. elegans qui ont des corps cellulaires répartis autour du corps (Figure 3A) et sont appelés cellules microtubules parce que leur ultrastructure comporte 15 microtubules protofilament distinctifs. Les neurones récepteurs tactiles sont l’AVM antérieur (neurone microtubule ventral antérieur), l’ALMR et l’ALML (neurones latéraux antérieurs microtubules à droite et à gauche), le PVM plus central (neurone microtubule ventral postérieur), et le PLMR postérieur et PLML (neurones latéraux postérieurs microtubules à droite et à gauche) dans la queue. Fait intéressant, les six neurones récepteurs tactiles produisent des exophers à des taux différents, en dépit d’exprimer le même transgène offensant (Figure 3C). Des six neurones sensibles mécanosensoriels, le neurone ALMR subit l’exophergenesis plus souvent que les autres neurones tactiles. La quantitation des nombres d’exopher des neurones tactiles est donc généralement établie en se concentrant sur l’ALMR.
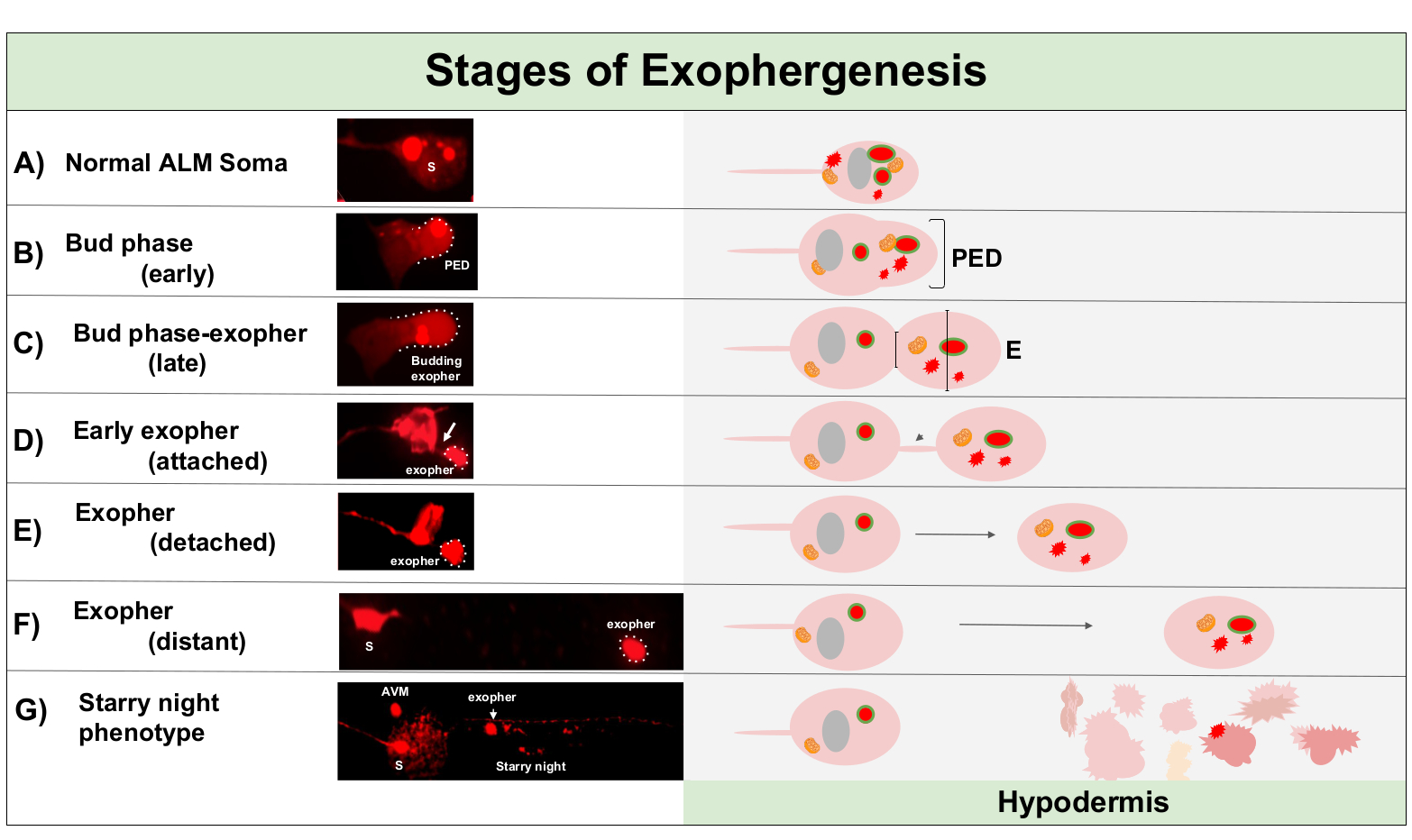
L’exophégenèse est un processus dynamique qui commence généralement par un gonflement du cytoplasme neuronal (figure 1A-B). Le contenu cellulaire, les organites ou les agrégats protéiques sont recueillis d’un côté du soma neuronal, le plus souvent vers l’extrémité postérieure du neurone ALMR (loin de la neurite de projection), formant un domaine pré-exopher (PED) (Figure 1B). La saillie précoce est observée lorsque le PED commence à se projeter vers l’extérieur, formant un bourgeon saillant reconnaissable. Le bourgeon tardif est défini lorsque le diamètre le plus large du domaine pré-exopher est environ 1/3 plus grand que le diamètre de la constriction du cou soma-exopher (figure 1C). Les exophers peuvent être éjectés dans presque n’importe quelle direction du soma, mais la plupart des exophers sortent postérieurement du corps cellulaire et restent dans approximativement le même plan focal que le soma originaire.
L’exopher peut s’éloigner du soma d’origine lorsque le cou du bourgeon se rétrécit en un mince filament. Les exophers peuvent rester attachés au soma par ce filament (figure 1D, flèche)et peuvent plus tard se détacher. Les contenus cellulaires tels que le calcium, les agrégats et les mitochondries peuvent être transférés par l’intermédiaire de ce filament dans l’exopherattaché 5, bien que la majeure partie du matériel extrudé soit mise dans le compartiment exopher par l’événement massif en herbe. Les exophers sont considérés comme matures lorsqu’il n’y a pas de tube de connexion visible ou de filament mince et que l’exopher est complètement séparé du soma d’envoi (figure 1E).
Les exomères produits par C. elegans touchent immédiatement l’hypoderme, le tissu qui entoure le neurone tactile. Le plus souvent, le vésicule exopher semble voyager dans l’hypoderme postérieurement vers la queue, et peut être assez éloigné de la soma avant que le contenu exopher apparaissent ciblés pour la dégradation (par exemple, la distance peut être ~100 μm loin de la soma (Figure 1F)). La vésicule exophaire fluorescente se décompose en de nombreux vésicules plus petites dans l’hypoderme, prenant une apparence appelée « nuit étoilée » (figure 1G et figure 2). Au stade de la « nuit étoilée », on peut observer des matériaux fluorescents punctate dispersés à travers le syncytium hypodermique en de nombreux petits points de fluorescence par rapport à l’exopher solitaire d’origine. Nuit étoilée peut sembler punctate sous le grossissement bas et avec le grossissement plus élevé, peut sembler punctate et/ou réseauté dans l’hypoderme. Le signal fluorescent de la nuit étoilée est généralement plus faible que l’exopher et la fluorescence exprimée par neurone (Figure 2B-C). La dispersion de mCherry en beaucoup de vesicules punctate est pensé pour impliquer la maturation phagosome et la fusion avec le réseau endosome/lysosomal de la cellule hypodermique. Certains matériaux exophaires sont probablement dégradés dans le réseau lysosomal hypodermique, mais les espèces résiduelles qui sont résistantes à la dégradation (comme les agrégats de mCherry) sont jetées hors de l’hypoderme dans le pseudocoelom, un compartiment fluide qui peut contenir des débris cellulaires. Le matériau fluorescent est plus tard pris en charge par les cellules charognardes éloignées appelées coélomocytes (Figure 2C), qui peuvent se concentrer, stocker, et à nouveau tenter la dégradation de mCherry.
Le phénomène de l’extrusion et du transfert d’agrégats semble conservé à travers le phyla, ayant été rapporté dans des modèles génétiques tels que C. elegans5,6,7 et D. melanogaster8,9 ainsi que dans plusieurs modèles de mammifères. Des extrusions exophéro-semblables ont été rapportées pour les cellules de mammifères10, une observation suggérant que les mécanismes conservés pourraient sous-être l’expulsion d’agrégat et d’organelle. La production d’exopher peut donc être un mécanisme conservé de la gestion des débris cellulaires qui constitue une branche fondamentale, mais autrefois non reconnue, de la protéostase neuronale et du contrôle de la qualité mitochondriale, qui, lorsqu’elle est déséquilibrée, pourrait contribuer activement à la maladie neurodégénérative. L’identification des molécules impliquées dans la discrimination et le tri des débris, le transport vers un lieu subcellulaire distinct, l’extrusion, la formation/scission de la connexion tubulaire reliant le soma et l’exopher tardif, et la reconnaissance de la grande vésicule extrudée pour la dégradation à distance par une cellule voisine restent pour des travaux futurs. Les études sur les modèles de nématodes et de mouches seront d’une importance cruciale pour définir les mécanismes de collecte et de transfert d’agrégats et d’orgales, en utilisant des approches génétiques impartiales et des outils biologiques cellulaires puissants offerts par ces modèles pour identifier les molécules participantes dans le contexte physiologique.
Les premières étapes critiques dans le déchiffrement des mécanismes opératoires dans la biologie exopher impliquent la définition de protocoles pour la quantification in vivo exopher in vivo. Le modèle C. elegans offre un avantage particulier pour de tels efforts puisque le corps est transparent et les exophers peuvent être facilement observés lorsqu’ils contiennent des protéines ou des organites fluorescents. Des exomères ont été générés par les neurones dopaminergiques C. elegans PDE et CEP, les neurones sensoriels ASE et ASER, et les neurones amphid deteinture-remplissage 5. Parce que les exophes produits par les neurones récepteurs tactiles sont mieux caractérisés, l’accent est mis ici sur l’utilisation des neurones tactiles pour l’analyse exopher. Toutefois, l’approche de base peut être appliquée pour mesurer la production d’exophér à partir de n’importe quelle cellule. Les protocoles de détection et de quantitation des exophers produits par les neurones récepteurs tactiles C. elegans qui expriment transgénétiquement la protéine mCherry sont décrits, en mettant l’accent sur les cargaisons qui peuvent être surveillées et les contraintes temporelles dans la notation. Cet article définit les approches vers l’identification in vivo exopher, et la quantitation des conditions environnementales et génétiques qui modulent la production d’exopher. Les protocoles mettent l’accent sur l’attention critique portée aux conditions constantes de non-stress pour la détermination de la production d’exopher de base et pour les comparaisons entre les génotypes.
Protocole
1. Souches utiles pour la détection d’exopher
- Sélectionnez une souche qui exprime des cargaisons fluorescentes dans les neurones de C. elegans pour visualiser facilement les exophes.
REMARQUE : Le tableau 1 répertorie les souches qui ont été utilisées pour visualiser les exopers produits dans les neurones récepteurs tactiles5,11,12. En principe, n’importe quel type de cellule ou de neurone peut être testé pour la production d’exopher en utilisant une cellule ou un promoteur spécifique de tissu pour conduire l’expression d’une protéine fluorescente qui s’agrége ou est autrement sélectionnée pour l’extrusion. - Alternativement, utilisez un test de remplissage de colorant pour visualiser les exophers dans les neurones de tête amphid, qui sont ouverts à l’environnement et pouvant se remplir5,13.
2. Médias de croissance
- Préparer les supports de croissance standard des nématodes (NGM) aux souches de culture selon les méthodes standard14,15.
REMARQUE : Le manque de nourriture, ou fluoro-désoxyuridine (FuDR), utilisé couramment pour bloquer la production de descendance, et peut affecter considérablement la production d’exopher. Gardez la population nourrie en permanence (éviter même de courtes périodes d’épuisement des aliments bactériens) et maintenir les animaux à une température constante.
3. Élevage critique pour une production constante d’exopher
- Élever des animaux sur des supports cohérents et avec des sources de nourriture bactériennes cohérentes. Les animaux ne doivent pas manquer de nourriture bactérienne, même pendant de courtes périodes de temps puisque la limitation des aliments peut changer considérablement les niveaux de production d’exopher.
- Gardez les recettes médiatiques et la préparation uniformes tout au long d’une étude.
REMARQUE : L’évolution des médias peut affecter les niveaux basaux de production d’exopher. Les lots d’agar peuvent influencer les niveaux d’exopher de base, de sorte que lorsque les lots d’approvisionnement changent, notez la date. Jetez les plaques de bouillon après deux semaines pour assurer la nourriture bactérienne saine et pour empêcher l’agar séché, qui provoque des changements dans l’osmolarité d’agar qui influencent les niveaux d’exopher. - Pour les conditions basales, garder les animaux à une température constante de 20 °C. L’élevage des animaux à des températures variables (même des changements temporaires de température) peut entraîner des variations dans le moment de la production maximale d’exophér.
REMARQUE : La variabilité de la température ne se limite pas aux conditions de culture. Les variations de température pendant les expériences ou sur le banc de laboratoire peuvent avoir un impact. Par exemple, les températures dans une salle de microscope ne devraient pas différer considérablement de l’incubateur de culture ou du banc de laboratoire. - N’utilisez pas d’interventions pharmacologiques anti-fertilité parce que les ovules fécondés sont essentiels pour la production précoce d’exophers chez les adultes.
REMARQUE : L’utilisation de fluoro-désoxyuridine (FuDR)16 ou C2217, doit être évitée. Lors de l’exécution d’expériences sur des animaux âgés ou âgés, les populations synchronisées par âge devraient être maintenues en retirant physiquement les adultes de leur plus petite progéniture en les cueillant sur des plaques fraîches propagées par des bactéries plutôt que d’utiliser des interventions pharmacologiques anti-fertilité courantes. - N’utilisez pas de cultures contaminées; relancer les expériences en cas de compromis biologique de la population ou de la plaque. La contamination bactérienne ou fongique peut induire des stress et des changements métaboliques chez les animaux et doit être absente des populations expérimentales.
- Pour maximiser les résultats reproductibles, maintenez les cultures pendant au moins deux générations saines, bien alimentées et sans contamination à 20 °C avant l’expérimentation afin d’éviter d’éventuels changements épigénétiques induits par l’environnement.
4. Synchronisation d’âge pour la notation d’exopher par blanchiment, flottation de saccharose, ou la cueillette de larves de L4
- Garder les populations expérimentales au même âge biologique, car les modèles de détection des exopher varient selon l’âge adulte et la comparaison des animaux de populations d’âges mixtes peut confondre les résultats. Assurez-vous toujours une synchronisation réussie des populations animales expérimentales en vérifiant la morphologie de la vulve « croissant blanc » au stade L4.
REMARQUE : En général, la production d’exopher de pointe pour les neurones ALMR mécanosensoriels de C. elegans se produit le jour adulte 2-3 (figure 3D), tel que mesuré à partir des jours après le stade L4. Le jour adulte 1 est 24 heures après le stade larvaire L4 qui se distingue par la morphologie de la vulve « croissant blanc » (Figure 5E). - Préparer les populations d’œufs synchronisés en blanchissant les adultes gravid.
- Recueillir les adultes gravid remplis d’œufs en lavant les animaux qui poussent sur une plaque NGM. Pour laver, inonder la plaque d’un tampon M9 de 1 ml, de monter et de descendre pour recueillir le liquide avec les animaux en suspension et le pipet dans un tube de microcentrifuge de 1,5 m L. Pelleter les animaux par décantation gravitationnelle ou centrifugeuse douce avec une mini centrifugeuse et enlever le supernatant.
- Ajouter 150 μL de 5M NaOH et 150 μL d’hypochlorite de sodium (eau de Javel) en 1 mL en H2O et mélanger par inversion pendant environ 5 minutes.
REMARQUE : La solution de blanchiment frais garantit que la cuticule animale peut être perturbée pour la récolte d’œufs. Les progrès dans la perturbation de la cuticule peuvent être surveillés sous un microscope de dissection ; les adultes devraient casser et libérer des œufs au point d’arrêter le blanchiment. - Centrifugeuse doucement avec un tube minicentrifuge pour 20 s et enlever le supernatant. Ajouter 1 mL de tampon M9 et la centrifugeuse à nouveau, en laissant environ 100 μL sur le dessus de la pastille.
- Répétez les étapes 4.2.3 deux fois pour enlever les traces de solution d’eau de Javel.
- Resuspendez les œufs dans le volume restant et transférez-les dans une assiette ngm fraîche ensemencée. Les adultes seront lysés, mais de nombreux œufs viables devraient être dans la préparation.
- Préparer les populations synchronisées en pondant les œufs chronométrés.
- Choisissez 20 adultes gravid à une plaque NGM ensemencée en utilisant les protocoles de transfert standard14.
- Permettre aux animaux de ramper librement et pondre des œufs pendant 1,5 h (des souches mutantes de faible taille de couvée peuvent nécessiter l’introduction d’un plus grand nombre d’animaux adultes).
- Retirez tous les animaux adultes de l’assiette en cueillant, laissant derrière vous la population d’œufs synchronisés. Vérifiez les plaques quelques heures plus tard pour vérifier qu’aucun adulte viable n’a été manqué lors de l’enlèvement des adultes.
- Préparer les populations d’œufs synchronisés par la sélection de flottaison de saccharose des œufs.
- Recueillir les animaux et les œufs de cinq plaques NGM sur lesquelles les animaux gravid pondent des œufs depuis au moins 24 heures en inondant les plaques avec la solution M9 avec 0,1% de détergent (comme Tween 20 ou Triton X-100) et la collecte dans un tube de 15 mL. Pellet adultes par centrifugation douce à température ambiante (2 000 x g pour 30 s).
- Retirer les supernatants et laver les animaux dans 15 mL de M9 frais trois fois, en jetant le supernatant après chaque lavage, en étant sûr de garder la pastille enrichie en animaux et œufs.
- Conserver 2 mL de supernatant et resuspendez le granulé. Ajouter 2 mL de 60% de poids en volume de saccharose.
- Centrifugeuse à 2000 x g pendant 5 min. La solution affichera désormais une phase supérieure très enrichie en œufs.
- Transférer environ 2,5 ml de la phase supérieure sur un nouveau tube de 15 mL et ajouter 10 ml de M9.
- Mélanger par inversion pendant 1 min, puis centrifuge 2000 x g pendant 1 min.
- Retirer le supernatant et laver la boulette enrichie d’œufs en M9. 10-15 μL de l’œuf-granulé peut être distribué dans une assiette fraîche épépinée de NGM.
REMARQUE : Cette méthode prépare un grand nombre d’œufs; ne permettent pas aux animaux recueillis de manquer d’aliments op50 E. coli.
- Préparer les populations synchronisées en cueillant des animaux au stade de développement L4.
- Cultiver des animaux sur des plaques ngm ensemencées comme décrit ci-dessus.
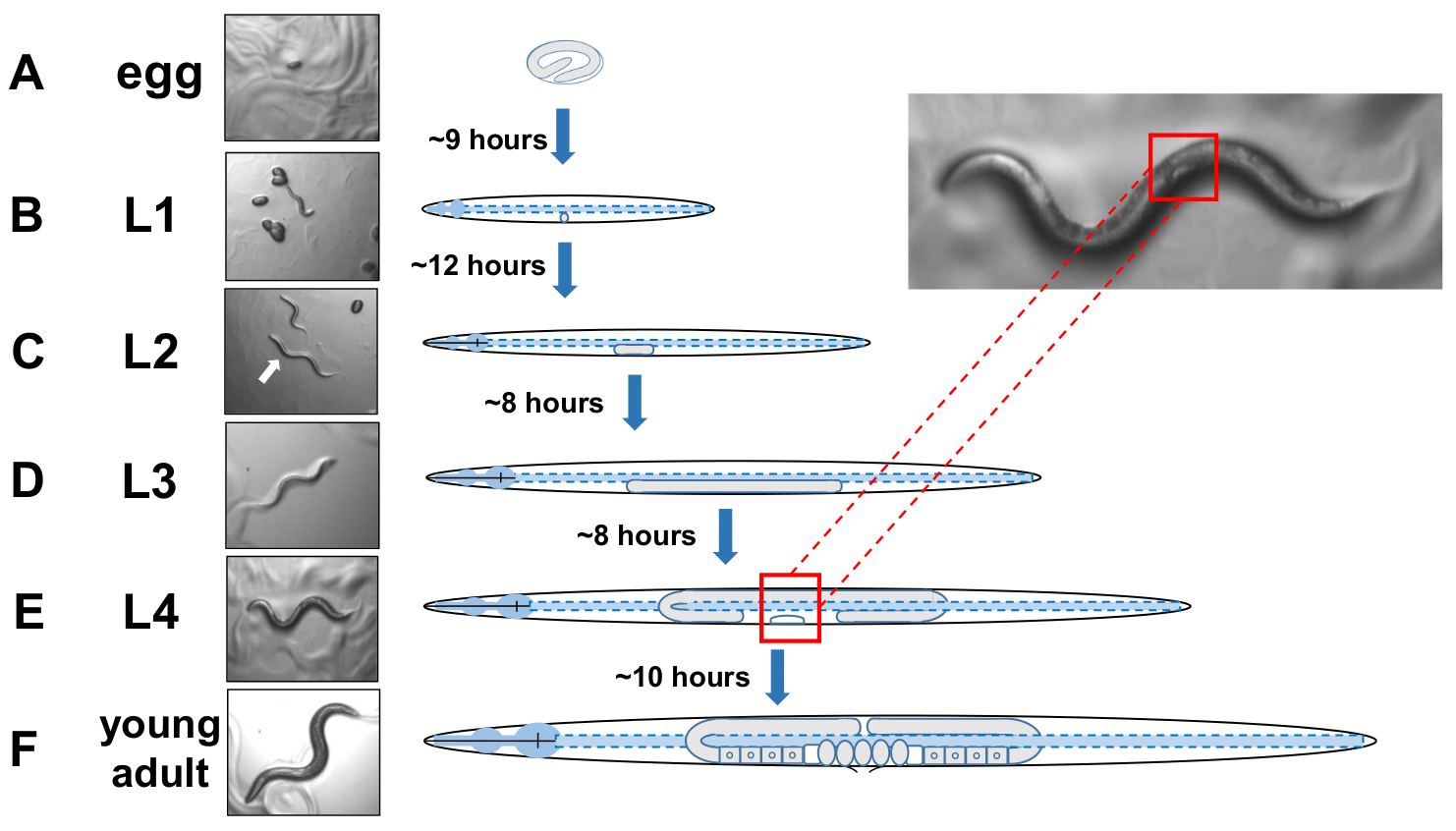
REMARQUE : C. elegans se développe en quatre étapes distinctes. À 20 °C, un œuf nouvellement pondu prend environ 9 heures à éclore (figure 5A). Après l’éclosion, un animal passe par le stade larvaire 1 (L1) jusqu’au stade larvaire 4 (L4), chaque étape d’une durée de 8 à 12 heures entre chaque mue (figure 5A-F). Par conséquent, une assiette préparée en inoculant avec des œufs devrait avoir beaucoup d’animaux L4 à cueillir environ 40 heures après l’introduction des œufs. - Identifier les animaux mis en scène L4 en localisant la forme blanche du croissant de demi-lune de la vulve en développement (figure 5E).
REMARQUE : Les animaux à l’étape L4 sont uniformes en taille et en pigmentation corporelle. Choisissez des animaux avec le croissant blanc à une plaque de croissance fraîche pour l’examen des animaux mis en scène. Le lendemain (~ 24 heures plus tard) doit être compté comme jour adulte 1. - Marquer une population d’animaux par jour le jour adulte 2.
NOTE : Les exophers sont généralement notés au jour 2 de l’âge adulte, qui est la production d’exopher de pointe dans des conditions basales. Toutefois, étant donné que le pic et le moment de l’exophergenese peuvent être décalés par des changements environnementaux ou génétiques à l’étude, il est conseillé de marquer une population d’animaux adultes par jour sur quatre jours pour générer l’image la plus complète (Figure 3D).
- Cultiver des animaux sur des plaques ngm ensemencées comme décrit ci-dessus.
5. Détection d’exophers à l’aide d’un microscope fluorescent
- Observez les exophers à l’aide d’un microscope de dissection pseudo-stéréo à grossissement élevé qui est équipé pour la microscopie de fluorescence.
- Immobiliser les animaux sur les plaques NGM en canalisant 100-200 μL de 10-100 mM de levamisole/tetramisole sur la surface de la plaque d’agar NGM. Après 2-4 minutes, les animaux deviennent paralysés et peuvent être observés directement sur la plaque d’agar.
REMARQUE : Les traitements d’immobilisation ne sont pas absolument nécessaires, de sorte qu’avec un œil formé, l’identification neuronale et la présence d’exopher peuvent être notées en suivant visuellement les animaux rampants sous le microscope sur la plaque pour déterminer si oui ou non un exopher a été produit. - Observez les neurones fluorescents en utilisant un grossissement total de 100x pour effectuer la détection au microscope de disséquer des exophers.
REMARQUE : La notation des événements exopher à l’aide de la microscopie de dissection permet l’observation d’un grand nombre d’animaux avec une relative facilité directement sur les plaques d’agar sur lesquelles ils sont élevés. - L’imagerie en direct et le montage des souches de journaliste pour les études d’exophe à l’aide de la microscopie confocale
- Utilisez un microscope confocal pour la dynamique intracellulaire d’image en direct et les caractéristiques de l’exophergenesis.
REMARQUE : L’imagerie en direct est une approche avantageuse pour observer des détails subtils de la production d’exopher parce que la production d’exopher est un processus dynamique. - Restreindre le mouvement des animaux pour l’imagerie vivante à haute résolution en utilisant des méthodes pratiques, y compris l’utilisation de la levamisole ou de la tétramisole à 10-100 mM ou l’application de microbilles de polystyrène hydrogel (avec des diamètres de 15 μm, 30 μm ou 40 μm)18.
- Utilisez un microscope confocal pour la dynamique intracellulaire d’image en direct et les caractéristiques de l’exophergenesis.
- Préparation des diapositives pour la microscopie composée et confocale
- Montez 20-50 animaux en un agent immobilisant sur une lame de microscope. Des lames de cytologie à anneaux réutilisables peintes avec des anneaux surélevés de 13 mm de diamètre sont utiles pour le montage.
- Choisissez des animaux vivants dans 5-20 μL d’un paralytique tel que la levamisole de 10 à 100 mM ou la tétramisole dans le cercle peint ou sur le tampon d’agar.
- Attendez 4 minutes pour la paralysie, puis couvrez la diapositive d’un couvercle (recommander le numéro 11/2 (0,16 – 0,19 mm) ou le numéro 2 (0,17 - 0,25 mm).
- Montage d’un petit nombre d’animaux
- N’écrasez pas les animaux montés; lorsque vous observez seulement quelques animaux (moins de 20) par glissement, il y a un risque d’écrasement de certains animaux en raison de la pression inégale du couvercle. Ce risque peut être minimisé en utilisant un faible pourcentage de pad agarose pour le montage.
- Faites une lame de pad agarose de 2 à 4 %, puis ajoutez 2 à 15 μL de solution paralytique au tampon. Gardez à l’esprit le levamisole et tétramisole diffusent dans le pad, diminuant leur concentration efficace.
- Montez en cueillant les animaux dans une goutte de 2 à 15 μL de solution paralytique ou de microbilles reposant sur le tampon d’agar. Placez le couvercle sur le dessus et vérifiez que les animaux sont intacts18.
- Préparation de pad d’Agar
- Pour préparer 2% de tampons agar, chauffer 2% d’agarose dans la solution M9 et micro-ondes jusqu’à ce que l’agarose soit dans un état homogène et fondu.
- Pour obtenir un tampon agar de qualité suffisante, alternez le mélange et le micro-ondes à faible puissance pendant moins de 20 secondes. Évitez l’inclusion de bulles d’air dans le tampon en plaçant l’agar bouillant sur un bloc de chauffage et en permettant aux bulles de remonter à la surface.
- Utilisez une pipette Pasteur pour dessiner l’agar des profondeurs de la solution en fusion sous les bulles levées.
- Préparer deux lames scotchées et placer de chaque côté d’une lame de microscope en verre propre sur une surface plane. Pour faire les diapositives scotchées placer deux bandes de 5 cm de ruban de laboratoire sur chaque diapositive (figure 6A).
- À l’aide d’un pipet Pasteur, placez une seule goutte d’agar sur la lame de microscope propre coincée entre les lames scotchées (Figure 6B).
- Soigneusement et rapidement, couvrir la goutte d’agar fondu avec une quatrième diapositive propre en plaçant à travers les diapositives enregistrées (Figure 6Cc).
REMARQUE : La glissière doit appuyer doucement sur l’agar fondu dans un cercle aplati d’environ 0,4 mm d’épaisseur (l’épaisseur du ruban) (figure 6D). L’agar doit rapidement refroidir. - Retirez la diapositive supérieure en la faisant glisser (figure 6E). Les tampons agar sèchent rapidement et sont mieux utilisés en quelques minutes. Une fois la lame supérieure enlevée, utilisez immédiatement le tampon de gel pour le montage des animaux. Évitez d’utiliser des coussinets avec des bulles d’air.
- Conserver les tampons d’agar jusqu’à 30 minutes encastrés entre les deux glissières en verre. L’agar séché fait en sorte que les animaux s’agglutinent et se dessésent. Monter les animaux dans un rayon de 2 à 15 μL de solution paralytique ou de microbilles et couvrir de couvercles; filtrer la diapositive dans les 20 minutes suivant la paralysie et le montage (figure 6).
REMARQUE : Parce que les conditions de stress peuvent modifier les taux d’exopher, évitez les paralytiques qui peuvent induire un stress oxydatif (par exemple : azide de sodium) lors du dépistage des exophers.
- Détection d’exophers à l’aide d’un microscope confocal à disque tournant
- Observez les caractéristiques biologiques cellulaires telles que les organites et d’autres contenus avec des objectifs d’ouverture numérique de 1,4 à 63x et 100x.
- Utilisez un logiciel capable de contrôler la scène et d’acquérir des images en utilisant l’acquisition multidimensionnelle. Les microscopes et les logiciels de traitement d’images devraient également convenir à l’imagerie et à la collecte de données, car ces étapes impliquent des approches d’imagerie standard.
6. Identifier les neurones tactiles et marquer pour les exophers avec des animaux montés
- Mont animaux adultes paralysés (Figure 6).
- Identifiez le plan Z souhaité. Utiliser un champ lumineux à faible grossissement (10-40x) pour identifier le plan Z approprié de l’animal, en prenant note du positionnement de l’animal, de l’orientation de la queue de tête et de l’emplacement de la vulve - qui sont des repères pour l’identification neuronale et exopher ultérieure (figure 3A et figure 5E).
- Concentrez-vous sur le signal de fluorescence du journaliste choisi. En restant dans le même Z-plane, passez à l’observation de fluorescence widefield à 10-40x pour le journaliste cytosolique choisi.
NOTE : Dans cet exemple, l’expression fluorescente est pilotée par le promoteur mécanosensoriel de neurone de contact mec-4. Les tableaux de copie élevés et les fluorophores différents ont une variabilité d’expression et donc une intensité fluorescente variable. Ajuster si nécessaire. - Faites défiler dans l’axe Z pour observer la profondeur de l’expression animale et fluorescente dans le plan focal. Ce faisant, confirmez l’orientation tête-queue; la tête/pharynx aura l’anneau de nerf fluorescent et dans ce cas, la queue contiendra 1-2 somes visibles plm (figure 3A).
- Identifier les neurones tactiles
- Déterminer si l’animal est monté sur le côté gauche ou le côté droit (figure 3A).
REMARQUE : Compte tenu de la 3-dimensionnalité de l’animal, la meilleure résolution d’imagerie est accomplie sur le côté le plus proche de l’optique. - Identifiez le soma (ALM, ALMR, AVM) en observant - commencez à la tête pour identifier l’anneau nerveux et les processus neuronaux latéraux.
- Au grossissement 10-40x, faites défiler lentement l’axe Z pour identifier le processus ci-joint.
- Une fois que le processus est identifié, suivez-le ultérieurement dans la direction postérieure vers la vulve, où le soma sera apparent, marqué par un corps cellulaire rond à la fin du processus. Une fois que le soma neuronal le plus concentré est trouvé, il peut être identifié en utilisant d’autres repères neuronaux comme suit:
- Utilisez l’AVM, un neurone ventral à proximité, pour aider à assigner l’orientation animale. Si le neurone AVM est dans le même plan que l’ALM, alors l’animal se repose sur son côté et le neurone à l’extérieur de ce plan est l’ALMR . Si le neurone AVM n’est pas dans le même plan que l’ALM en question, le neurone tactile le plus proche du plan focal est ALML.
- Identifier le neurone PVM, un autre neurone tactile ventral situé près de la queue, pour indiquer si le neurone tactile antérieur se trouve dans le même plan. Si c’est le cas, le neurone tactile observé est ALML.
- Obtenez une idée de la position des autres corps de soma, près de la zone d’intérêt (neurones fluorescents situés de chaque côté de la soma), et dans tous les Z-planes, même s’il n’est pas possible d’obtenir le neurone le plus profond mis en évidence.
NOTE : L’identification de tous les somas de neurone tactile est importante parce que le soma hors foyer peut être confondu avec des exophers.
- Déterminer si l’animal est monté sur le côté gauche ou le côté droit (figure 3A).
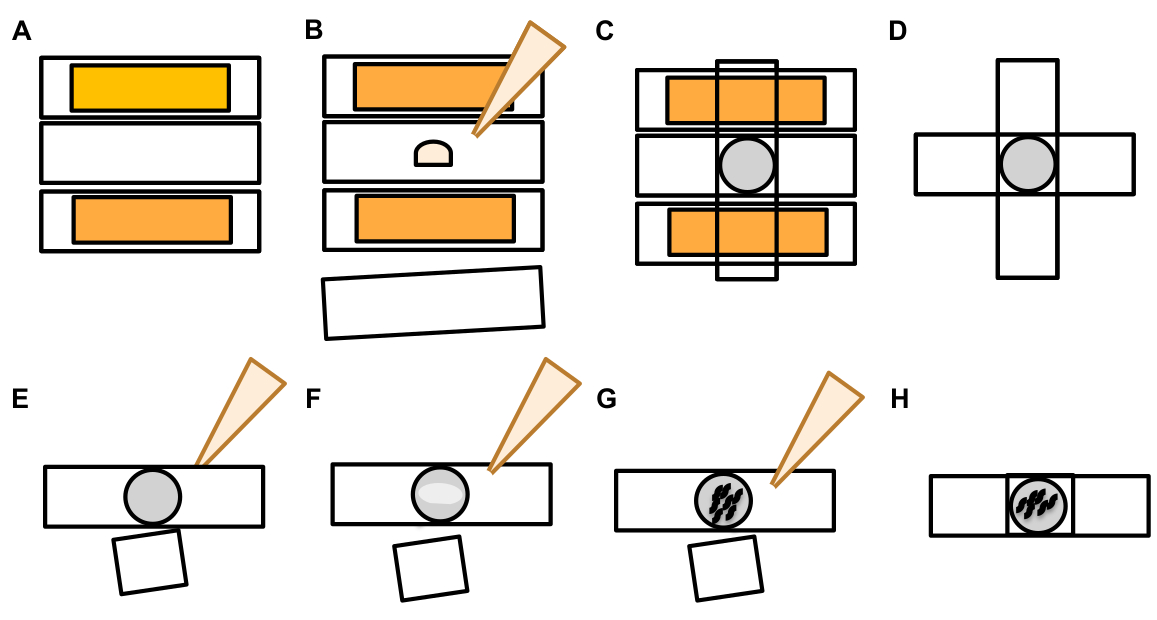
7. Identification et notation des exophers
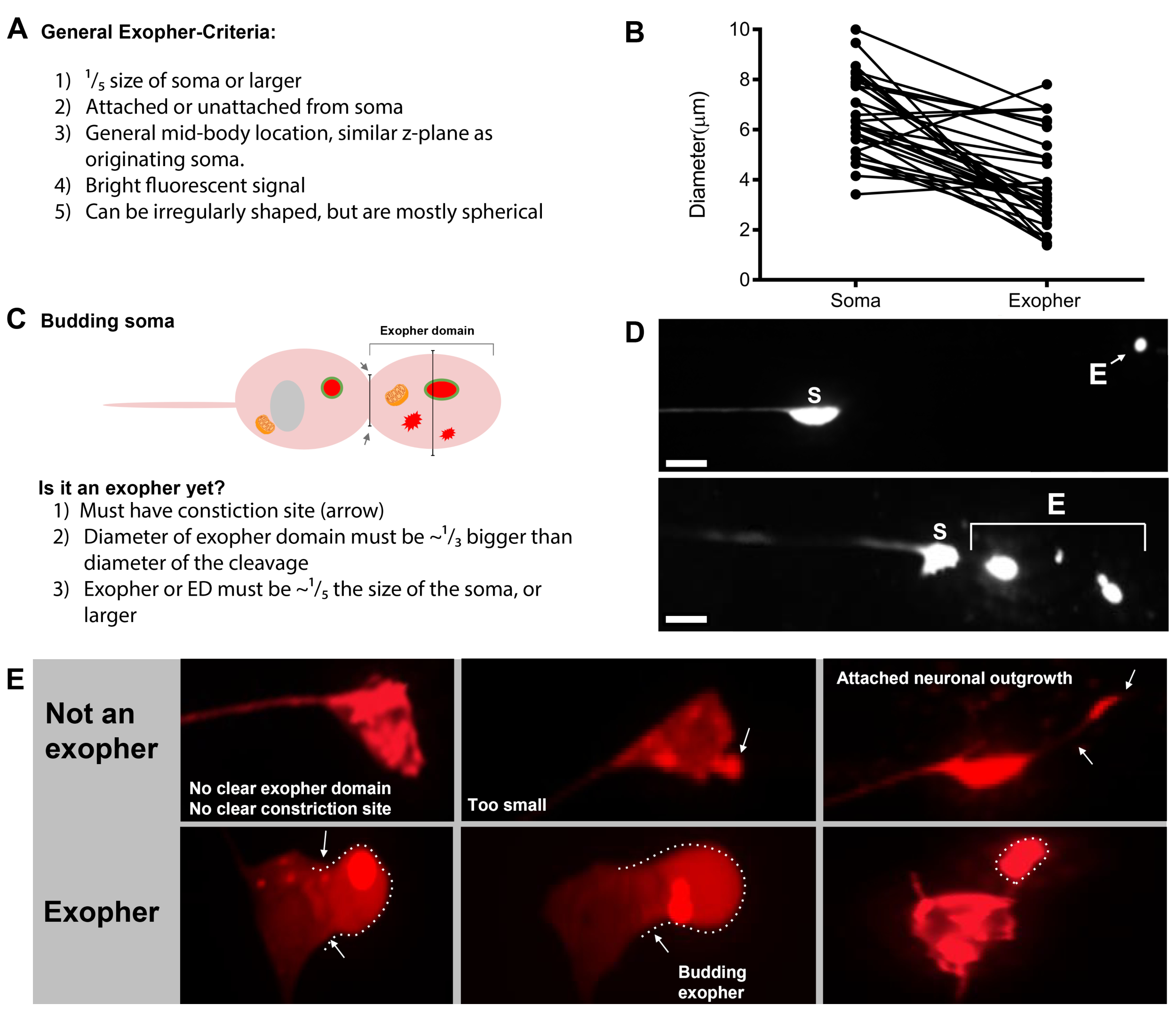
- Une fois qu’un neurone tactile est trouvé, inspectez-le pour les grandes saillies (domaines exopher) assez grandes pour être considérées comme un exopher de bourgeons , (atteignant au moins 1/5e de la taille du soma d’origine) (Figure 1C).
REMARQUE : L’exopher moyen mesure environ 2-8 μm de diamètre, tandis que le soma moyen d’un (ZB4065 bzIs166[Pmec-4::mCherry]) l’animal mesure 6-10 μm dans le jour 2 adultes (Figure 7B). - Si aucun bourgeon ou domaine exopher n’est observé, inspectez le soma neuronal pour un filament mince attaché émanant du soma. Les exophers attachés ont tendance à être situés plus près du soma d’origine et dans un plan Z similaire.
REMARQUE : Les exophers ne restent pas toujours attachés au soma. La détection d’un filament attaché est une indication définitive que l’objet est un exopher. - Pour identifier un exopher non attaché, recherchez le contenu d’un exopher. Les exophers peuvent concentrer les protéines fluorescentes expulsées et sont donc souvent plus brillants que le soma.
REMARQUE : Le contenu des exophers est hétérogène et variable. Les organites cellulaires tels que les lysosomes et les mitochondries peuvent également être extrudés dans les exophers (Figure 4C-E). - Recherchez des exopheurs non attachés dans différents plans focaux que l’avion dans lequel le soma d’origine a été trouvé. Bien que les exophers aient été vus pour faire saillie de la soma D’ALM dans n’importe quelle direction, il est typique que les exophers dépassent loin du soma, dans une direction postérieure du processus neuronal.
- Vérifiez les gros objets sphériques qui ne sont pas positionnés et identifiés comme des somas neuronaux. Les exophers peuvent avoir une forme irrégulière, mais sont généralement des structures sphériques. Les exophers se dégradent avec le temps, de sorte que les exophers plus âgés ont tendance à avoir une forme plus irrégulière.
REMARQUE : Les exophers matures ou plus âgés se distinguent de l’étape dispersée de la « nuit étoilée » par l’intensité de fluorescence plus brillante des exophers et de leur forme sphérique. - Étudier le phénotype de la « nuit étoilée » comme preuve de l’exophergenesis antérieure. Les exophers progressent dans une étape de «nuit étoilée » pendant que l’exopher se brise en vésicules plus petites et que l’hypoderme environnant tente de dégrader le contenu d’exopher (figure 1G, 2B, 3B et 7A).
REMARQUE : L’étape nocturne étoilée est marquée par des entités fluorescentes fragmentées et dispersées (parfois en réseau) qui ont perdu leur intégrité structurelle et affichent une fluorescence faible par rapport aux neurones tactiles et aux structures exopher. - Recherchez des instances d'« événements exopher multiples ». Les exophers sont généralement produits comme une occurrence singulière (1 exopher émanant de 1 soma) mais dans certaines circonstances plus d’un exopher peut être libéré d’un seul soma (Figure 7D).
REMARQUE : Les exophers matures peuvent se dégrader en vésicules multiples car elles sont dégradées dans l’hypoderme. La distinction entre chaque exopher a été générée par un événement d’exophégenèse indépendant ou si une scission exopher originale pour créer une vésicule supplémentaire ne peut être déterminée que par l’observation en time-lapse. - Gardez à l’esprit que toutes les anomalies morphologiques ne mûrissent pas en exophers.
- Ne marquez pas un soma distendé comme un exopher. Un soma étendu ou pointu peut être observé à l’occasion (surtout avec l’âge ou sous le stress), mais une extension sans un site de constriction claire n’est pas marqué comme un exopher.
- Rejeter les petits bourgeons résolus qui n’atteignent pas1/5 e de la taille du soma dans la quantification de l’événement exopher.
- Ne comptez pas les excroissances neurite comme exophers. Les neurites matures peuvent s’étendre de façon spectaculaire avec l’âge (généralement dans la direction opposée du processus neuronal) et les protéines fluorescentes peuvent migrer dans l’extrémité distale de ces structures19.
REMARQUE : Ces excroissances de neurite ne sont pas des exophes car elles ont un modèle de développement distinct au cours des jours et des semaines, ne forment pas les bourgeons et ne se détachent pas (figure 7E).
- Identifiez les entités fluorescentes qui ne sont pas des exophers.
REMARQUE : Il est important d’avoir une idée de la fluorescence de fond pour assurer une identification correcte de l’entité fluorescente extrudée par rapport à l’autofluorescence.-
Expression fluorescente transgénique vs autofluorescence. Ne confondez pas l’autofluorescence avec l’expression transgénique. Le vrai signal exopher ne sera pas dans l’intestin ou l’intestin (la confirmation de DIC peut être employée pour identifier ces tissus) et le signal d’exopher sera sensiblement plus lumineux que l’autofluorescence de fond.
REMARQUE : L’autofluorescence est causée par la pigmentation fluorescente intestinale des granules intestinaux et s’accumule avec l’âge. Il est hétérogène, surtout vu avec différentes longueurs d’onde. - Signal d’embryons. Ne confondez pas le signal embryonnaire avec l’exophégenèse. Confirmer les soupçons de signal embryonnaire en passant de la fluorescence à l’éclairage brightfield et en vérifiant les associations de signal avec les œufs dans l’utérus.
-
Hors de l’avion ou à proximité des corps de soma. Évitez de confondre un soma hors de l’avion pour un exopher en identifiant tous les corps de soma à proximité, même hors de la mise au point somas au début de l’observation.
REMARQUE : Si vous marquez pour les exophers d’ALMR, identifiez et expliquez l’emplacement des somas AVM et ALMR. Plus de détails sur l’identification du corps soma sont décrits à la figure 3A.
-
Expression fluorescente transgénique vs autofluorescence. Ne confondez pas l’autofluorescence avec l’expression transgénique. Le vrai signal exopher ne sera pas dans l’intestin ou l’intestin (la confirmation de DIC peut être employée pour identifier ces tissus) et le signal d’exopher sera sensiblement plus lumineux que l’autofluorescence de fond.
8. Notation et statistiques
- Score exophers comme binaire (oui, il ya un exopher / non, il n’y a pas un exopher).
- Considérez la détection d’exopher comme un « événement d’exopher » pour un neurone donné. Un événement exopher peut constituer l’observation d’un seul exopher près d’un soma ou d’un exophers multiples.
REMARQUE : Pour quantifier le nombre d’événements individuels d’exophégenèse, utilisez l’observation en time-lapse. - Compter les événements exopher par cellule identifiée particulière parce que différentes cellules ne produisent pas d’exophers au même rythme (voir par exemple la figure 3C). Les neurones ALMR produisent le plus d’exophers de base dans les souches décrites ici et donc souvent c’est la cellule sélectionnée pour la quantification exopher des neurones récepteurs tactiles.
- Pour les statistiques, en général, mener au moins 3 essais biologiques, sur au moins 30 animaux notés par essai, avec le nombre correspondant d’observations nécessaires à l’analyse de la perturbation.
- Pour les essais multiples impliquant un ou deux mutants/traitements par rapport au contrôle, le test Cochran-Mantel-Haenszel est approprié pour déterminer les valeurs p.
- Pour les essais portant sur plus de deux mutants de traitements par rapport au contrôle, il convient également d’utiliser une analyse de régression logistique binaire pour évaluer l’importance d’un certain nombre de prédicteurs catégoriques.
Résultats
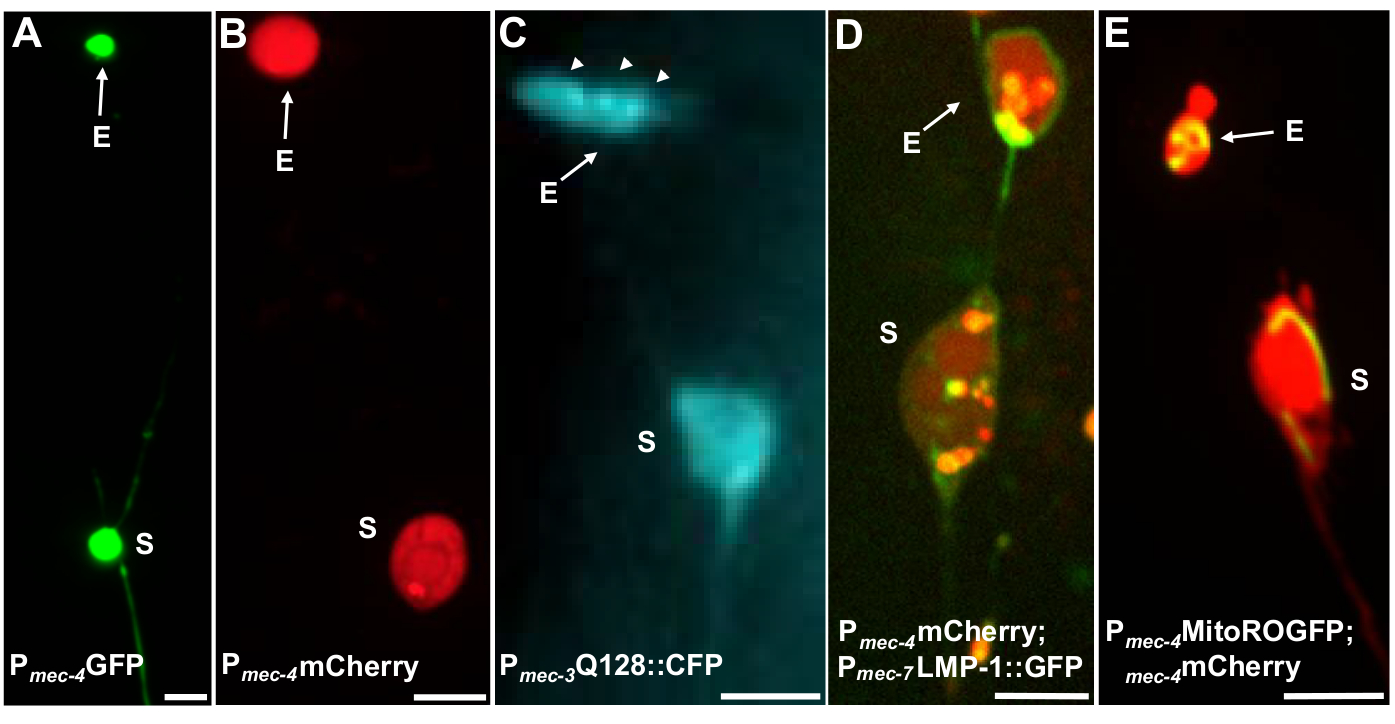
Plusieurs reporters fluorescents peuvent être utilisés pour mesurer les exopheurs. Les exopherons des neurones tactiles sont facilement visualisés in vivo par le marquage fluorescent des protéines qui peuvent être sélectionnées pour l’extrusion, par l’étiquetage des organites qui peuvent être extrudés, ou par le marquage des membranes cellulaires. Le tableau 1 identifie les neurones tactiles exprimés par des journalistes fluorescents qui ont été utilisés pour surveiller les exophers, avec des exemples représentatifs inclus à la figure 4. Les cargaisons qui sont connues pour être extrudées dans les exophers comprennent une fusion du domaine n-terminal de huntingtine humaine à la polyglutamine étendue (Q128) (Figure 4B), lysosomes qui sont GFP-marqués avec la protéine de membrane associée au lysosomal (LMP-1) (Figure 4C), et les mitochondries marquées avec GFP(figure 4D)localisée par matrice ( Figure 4D ). Cytoplasmique GFP n’est pas fortement expulsé et est préférentiellement conservé dans le soma5, bien que GFP peut faiblement visualiser exophers (Figure 4A). Lorsque GFP est fusionné aux protéines qui sont expulsées, cette balise peut être utilisée pour visualiser les exophers. Un point important est qu’en taguant différentes protéines, un large éventail de questions sur l’expulsion de cargaisons et d’organites spécifiques, ainsi que sur les protéines et les membranes qui composent les exophers, peuvent être abordées.
Une configuration pseudo-stéréomicroscope est un outil efficace pour visualiser les exopers chez les animaux sur des plaques d’agar. Cette configuration est un hybride de la technologie composée et stéréoscopique qui comprend des optiques à ouverture numérique élevée sur chaque grossissement, la technologie pseudo-stéréo (objectifs discrets sur une base stéréoscopique), et un commutateur d’exploitation de zoom pour la visualisation à grossissements intermédiaires à des objectifs installés. Un microscope comme celui-ci devrait être équipé d’oculaires 10x et d’objectifs suffisamment puissants pour observer la morphologie neuronale et la production d’exophér pour la notation à haut débit (objectif 2x utilisé pour la numérisation/cueillette, objectif 10x utilisé pour l’identification et la notation).
Alors que les capacités de grossissement des stéréomicroscopes standard ont généralement une résolution assez élevée pour voir le réseau de neurones tactiles exprimant des protéines fluorescentes, les microscopes de dissection standard ne sont pas suffisants pour observer les détails subcellulaires des exophers comme les connexions tubulaires de soma à exopher. De telles observations nécessitent une microscopie confocale (voir le Tableau des matériaux pour plus de détails sur l’équipement).
Les études de quantitation d’Exopher exigent des contrôles stricts pour éliminer les contraintes expérimentales. Le maintien attentif de conditions de croissance cohérentes est nécessaire pour la production exopher reproductible. Plus précisément, la production d’exopher est sensible au stress, de sorte qu’une alimentation constante, une température constante et une croissance sans contamination d’une génération à l’autre sont essentielles à la reproductibilité. Dans des conditions de croissance basale avec une expression neuronale élevée de mCherry, la production d’exopher est relativement faible (5-25% des ALMR produisent des exophers) mais certains stress, y compris le stress osmotique et oxydatif, peuvent augmenter les taux d’exophér. Alors que l’expression mCherry peut être considérée comme un stress, un corollaire de la sensibilité au stress des niveaux d’exopher est que, si correctement contrôlé, l’introduction expérimentale de stress peut être une stratégie pour induire et observer plus facilement l’exophégénèse.
Calendrier et niveaux de production d’exopher prévus. Les exophers sont pratiquement absents pendant le développement larvaire. La période de production maximale d’exopher dans la vie des jeunes adultes semble être fortement limitée à pendant les jours adultes 1-4, le plus souvent étant évident au jour adulte 2 ou 3. Parce que le pic peut se déplacer en avant ou en arrière un peu, l’évaluation la plus complète d’un profil de production exopher est de marquer plusieurs essais par jour sur les jours adultes 1-4. En général, un ALMR produit un exopher majeur, la vésicule persistant pendant au moins 24 heures. L’exopher peut être produit assez rapidement (de l’ordre des minutes à son plus rapide). Le plus souvent, un seul exopher majeur est produit par neurone au début de la vie adulte, mais la production de plusieurs exophers est possible.
En général, la production d’exopher par almrs exprimant mCherry dans des conditions basales varie de 5-25% des ALMR examinés dans le délai optimal du jour adulte 2-3 (Figure 3D). Les crises de protéostasie5, ainsi que l’exposition à d’autres contraintes peuvent moduler le niveau d’exopher. Le stress ou les perturbations génétiques peuvent augmenter la production d’exopher à des taux de détection aussi élevés que 90% des neurones ALMR produisant des extrusions exopher.
RNAi à base d’alimentation pour tester les rôles de gènes spécifiques dans l’exophégenèse. Le nématode C. elegans est généralement soumis à l’RNAi abattre par l’alimentation des animaux transformés E. coli souche HT115 qui expriment un ARN double brin (dsRNA) ciblant un gène d’intérêt20. Les bactéries HT115 peuvent être utilisées lors de la notation pour les exophers dans l’alimentation RNAi5. Alors que les transcriptions dans la plupart des tissus peuvent être ciblées par l’ARNI en utilisant cette technique, les neurones sont plus réfractaires. La sensibilité à l’ARN peut être calibrée à l’aide d’animaux qui expriment le transporteur transgénique dsRNA SID-1 sous un promoteur spécifique aux neurones. De cette façon, le tissu neuronal peut être sensibilisé à RNAi21.
L’effondrement tissulaire d’un gène d’intérêt peut être accompli en exprimant une composante du métabolisme endogène de l’RNAi au sein d’un mutant qui est déficient dans ce composant. Par exemple: la protéine argonaute RDE-1 peut être exprimée spécifiquement dans les neurones des animaux mutants rde-1 pour atteindre le renversement d’un gène d’intérêt que dans les neurones lorsque les animaux sont exposés à une intervention RNAi ciblant ce gène.
En utilisant les protocoles standard de l’ARNI20,22, l’exposition des parents au stade L4 à l’ARNI et permettant à leur progéniture de développer la consommation transformée HT115 bactéries jusqu’à l’âge adulte génère la forte perte génétique, mais être attentif aux retards de développement potentiels induits par l’ARNI que les animaux expérimentaux peuvent croître différemment d’un vecteur de contrôle vide. Il est important d’inclure toujours la lutte antivectorielle vide pour la comparaison de contrôle négatif. Les bactéries HT115 peuvent être utilisées lors de la notation pour les exophers dans l’alimentation RNAi. Cependant, notez que certains gènes sont efficaces pour modifier les taux d’exophégenèse, même pendant les périodes plus courtes de l’exposition àl’ARN 5. Si le ciblage de certains gènes conduit à une défaillance du développement, évitez d’exposer les animaux à l’effondrement à vie, les animaux peuvent simplement être cueillis au stade L4 sur des plaques RNAi pour être exposés de L4 à l’adulte D2 ou D3.
| Nom de la souche | Génotype | Description | Pourcentage d’exopher | Référence |
| SK4005 | zdIs5[Pmec-4GFP] | Expression cytosolique de GFP dans les neurones tactiles. | 1-8% ALM | Figure 4A, Melentijevic 2017 |
| ZB4065 | bzIs166[Pmec-4::mCherry] | La surexpression de mCherry (bzIs166) dans les neurones tactiles produit à la fois le signal cytosolique et les agrégats mCherry. bzIs166 est un inducteur d’exopher. Les agrégats mCherry sont des prédicteurs de l’exophergenesis et sont extrudés de préférence chez les exophaires. | 3-20% ALM (conditions normales). 20-80% ALM (conditions de jeûne). | Figure 4B, Melentijevic 2017 |
| ZB4067 | bzIs167[Pmec-4mitogfp Pmec-4mCherry4]; igIs1[Pmec-7YFP Pmec-3htt57Q128::cfp lin-15+]; | YFP étiquettes cytosolétiquement mec-7 neurones tactiles. Co-exprimé Q128::CFP agrége et induit exophers. La PCP fait un silence préférentiel. | ~25% | Figure 4C, Meletijevic 2017 |
| ZB4509 | bzIs166[Pmec-4mCherry]; bzIs168[Pmec-7LMP-1::GFP] | bzIs168 LMP-1::GFP étiquette les membranes plasmatiques et les membranes lysosomales. bzIs168 peut être utilisé pour identifier les membranes neuronales, les exophers (car ils sont liés à la membrane), et les structures lysosomal-membrane. | 3-20% ALM | Figure 4D, Melentijevic 2017 |
| ZB4528 | bzIs166[Pmec-4mCherry]; zhsEx17 [Pmec-4mitoLS::ROGFP] | Allele zhsEx17 est un journaliste mitochondrial localisé qui change sa longueur d’onde de pointe d’excitation de 405nm (oxydé) à 476nm (réduit) selon l’environnement oxydatif local. Il est exprimé dans les neurones tactiles et peut être utilisé seul pour identifier les mitochondries dans les neurones tactiles et dans les mito-exophers. | 3-20% ALM protéo-exopher. % quantitation mito-exopher ALM en cours. | Figure 4E, Melentijevic 2017, Cannon 2008, Ghose 2013 |
Tableau 1. Souches qui ont été utilisées pour la visualisation des neurones tactiles, toucher les neurones-exophers, et le contenu exopher.

Figure 1 : Étapes de l’exophégenèse. Le processus de fabrication et d’éjection d’un exopher est appelé « xopher-genesis ». Le processus dynamique de formation d’exopher peut prendre plusieurs minutes à plusieurs heures. Sont représentés des exemples de soma et de morphologie d’exopher à des étapes spécifiques pendant le processus dynamique d’exophergenesis dans une souche productrice d’exopher élevé, ZB4065 bzIs166[Pmec-4mCherry]. Toutes les images sont de jour 2 neurones adultes ALM prises avec un objectif 100x. (A) Soma normal. Neurone tactile mécanosensoriel adulte ALM exprimant transgéniquement Pmec-4mCherry. La morphologie soma représentée est typique des neurones jeunes adultes dans cette souche, avec des concentrations de mCherry dans le cytoplasme. (B) Phase précoce des bourgeons. La première étape observable de l’exophergenesis implique la polarisation du matériau cytoplasmique sélectionné jusqu’au bord de la membrane soma. Cette étape est souvent accompagnée d’une expansion ou d’un gonflement du soma. Dans le cas des neurones tactiles, le domaine pré-exopher (PED) s’étend dans l’hypoderme environnant (non visible ici). Notez la plus grande concentration de matière mCherry dans le domaine des bourgeons précoces. (C) Phase tardive des bourgeons. Dès la polarisation cellulaire et l’expansion du domaine pré-exopher, une constriction entre le soma et l’exopher (flèche) devient évidente. Cet événement signale la transition vers la phase de fin de bourgeon. Bien qu’à l’étape tardive de la bourgeon la cellule présente un site de constriction claire et des domaines séparés de soma et d’exopher, elle n’est pas encore complètement pincée du soma ; l’exopher en herbe peut être fixé par une tige épaisse (flèche). Le domaine en herbe est considéré comme un exopher précoce lorsque le diamètre du domaine exopher en question est d’environ 1/3 plus grand que le diamètre du chantier de construction/tige. (D) Phase précoce de l’exopher. Les exophers précoces peuvent être attachés par une tige du soma au départ — le diamètre de cette connexion peut s’amincir au fur et à mesure que l’exophe s’éloigne du soma. Le matériel cytoplasmique peut être transféré du soma à l’exopher par l’intermédiaire de ce tube, bien que la plupart du matériel soit chargé pendant le processus de bourgeonnement. Les exophers peuvent se détacher de la soma telle qu’elle est représentée dans (E), les exophers séparés sont considérés comme des exophers matures (F). L’exopher mature peut transiter par le tissu hypodermique environnant, s’éloignant du soma au départ. (G) La dégradation de l’exopher mCherry-étiqueté dans de plus petites vésicules dans l’hypoderme se traduit par une apparence punctate dispersée du matériau mCherry, très probablement comme il entre dans le réseau endolysosomal hypodermique. Le signal punctate dispersé est appelé la phase de la « nuit étoilée ». La dégradation de certains contenus exopher est probablement réalisée par des lysosomes hypodermiques, mais certains matériaux ne sont pas entièrement dégradés et sont souvent ré-extrudés par l’hypoderme dans le pseudocoelom. Le transit post-exophergesis mCherry est décrit plus en détail dans la figure 2. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 2 : mCherry extrudé des neurones tactiles dans les exophers engage le réseau lysosomal hypodermique environnant mais peut plus tard être extrudé dans le pseudocoelom où les coelomocytes peuvent stocker/dégrader le mCherry. (A) Résumé de dessin animé de la façon dont mCherry extrudé dans les exophers transite le corps après l’expulsion par les neurones. Pendant l’exophergenesis, certains contenus cellulaires tels que le mCherry deviennent localisés et s’éloignent du soma neuronal d’envoi dans une vésicule indépendante entourée des membranes plasmatiques neuronales et hypodermiques. Puisque les neurones tactiles sont incorporés dans le tissu hypodermique, comme les bourgeons de domaine exopher vers l’extérieur, il se déplace plus loin dans l’hypoderme. L’exopher peut transiter l’hypoderme, et après des heures à des jours, le contenu exopher peut se fragmenter dans le réseau endolysosomal de l’hypoderme. Le mCherry peut apparaître comme puncta dispersés tout au long de l’hypoderme, une scène appelée « nuit étoilée ». Après quelques jours, une partie de la mCherry peut s’évanouir de l’hypoderme dans le pseudocoelom environnant, où les cellules charognards appelées coélomocytes peuvent accéder à, et prendre, mCherry qui peuvent être stockés. (B) Exemple de l’apparition des vésicules étoilées de nuit mCherry. Image d’un soma ALM marqué avec mCherry avec de grands fragments d’exopher et des vésicules nocturnes étoilées. Souche est ZB4065 bzIs166[Pmec-4mCherry]. (C) Exemple de concentration de mCherry dans les coélomocytes éloignés. Sideview d’un animal adulte jour 10 de souche ZB4065 bzIs166[Pmec-4mCherry] montrant mCherry concentré dans les coélomocytes (flèches). Certaines vésicules étoilées de nuit sont également évidentes. En général, la concentration de coélomocytes devient évidente après environ le jour adulte 5 de la vie. (En bas B) Reproduction de dessin animé de (B), avec des neurones tactiles et des processus décrits en rouge, comme le sont les fragments d’exopher les plus brillants; de petites vésicules éparses de différentes profondeurs Z sont montrées en rose plus clair. (C en bas) Version dessin animé de l’image de (C), montrant le processus neuronal en rouge, nuit étoilée en rose et coélomocytes en vert. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 3 : Les neurones tactiles mécanosensoriels produisent des exophes à différents niveaux avec un profil temporel précis. (A) (Haut) Représentation de dessin animé des neurones tactiles mécanosensoriels en relation spatiale avec les principaux repères anatomiques de C. elegans, y compris le pharynx de pompage et l’anneau nerveux neuronal-dense à la tête de l’animal, la vulve au milieu du corps, et la queue effilée. (En bas) Neurones tactiles fluorescents exprimant GFP vu du côté supérieur et gauche (images adaptées de WormAtlas). La boîte rouge représente la zone où les exophers ALM sont généralement situés. (B) Vue de grossissement élevée de la région du corps moyen à laquelle les exopheurs dérivés de l’ALM sont produits dans une souche exprimant [Pmec-4mCherry]. AVM et neurone ALMR sont représentés, et montré est un exopher ALMR avec mCherry nuit étoilée. Les neurones ALMR produisent le plus facilement des exophers. (C) Neurones tactiles mécanosensoriels ALMR produisent plus facilement des exopers que d’autres neurones tactiles dans les hermaphrodites dans des conditions basales. La production mécanosensorielle d’exopher de neurone de contact le jour adulte 2, comme indiqué pour les neurones individuels de récepteur de contact est indiquée. Souche : ZB4065 bzIs166[Pmec-4mCherry], N>150, barres d’erreur sont SEM. (D) Les neurones tactiles ALMR produisent plus d’exophers pendant les jours 2 et 3 de l’âge adulte par rapport au stade L4 adolescent ou avec des animaux à l’âge avancé. Souche: ZB4065 bzIs166[Pmec-4mCherry], N>150, barres d’erreur sont SEM. Veuillez cliquer ici pour voir une version plus grande de cette figure.
{kind=link}

Figure 4 : Exemples de certains reporters fluorescents qui identifient le contenu de l’exopher. Une façon simple d’observer les exophers est de créer des animaux transgéniques qui expriment les fluorophores des promoteurs neuronaux. Les fluorophores permettent la visualisation de l’expression exopher et transgénique induit l’agrégation et/ou la protéostress qui augmente l’exophergenesis. Les exopers produits par les neurones amphid peuvent également être observés dans des conditions indigènes, en utilisant le remplissage de colorant pour la visualisation. Sont montrés des exemples de souches communes qui peuvent être utilisées pour observer exophers, (E) exopher, (S) soma. (A) Soma et exopher d’un ALM d’un adulte de souche SK4005 zdIs5[Pmec-4GFP], 100x objectif utilisé pour la photographie, barre d’échelle 3μm. Dans cette souche, les exopheurs qui incluent le GFP soluble sont mesurés, mais la production d’exopher se produit rarement. La fusion de GFP à des protéines qui peuvent être extrudées de préférence dans les exophers dans d’autres études confirme que les fusions de GFP peuvent être détectées dans les exophersmatures. (B) ALM soma et exopher d’un adulte de la souche ZB4065 bzIs166[Pmec-4mCherry], qui exprime mCherry et induit la production d’exopher de neurone tactile. Objectif 100x utilisé pour la photographie, barre d’échelle 5 μm. (C) ALM soma et exopher d’un adulte de souche ZB4067 bzIs167[Pmec-4mitogfp Pmec-4mCherry4]; igIs1[Pmec-7YFP Pmec-3htt57Q128::cfp lin-15+]; canal bleu sélectif utilisé pour l’image de htt57Q128::CFP. L’exopher contient des agrégats htt57Q128::CFP (flèches), qui apparaissent plus concentrés dans l’exopher que dans le soma. Objectif 40x utilisé pour la photographie, barre d’échelle 5μm. (D-E) Les exophers peuvent contenir des organites et un marquage spécifique à l’orgelle avec des protéines fluorescentes permet de surveiller l’extrusion organelle. (D) Étiquette de membrane lysosomal LMP-1:::GFP décrit la membrane soma et exopher et identifie les membranes plasmatiques faiblement (la localisation de membrane plasmatique est une étape de trafic sur le chemin du ciblage lysosomal) et étiquette fortement les organites lysosomal. Montré est un adulte ALM soma co-exprimant Pmec-4mCherry et le Pmec-7LMP-1::GFP qui se localise aux membranes et lysosomes. Le soma a un exopher attaché avec d’autres extrusions plus petites susceptibles d’être des fragments d’exopher (flèches). GFP structures positives sont inclus dans le soma et sont présents dans le grand exopher, souche: ZB4509 bzIs166[Pmec-4mCherry]; bzIs168[Pmec-7LMP-1::GFP]. Objectif 100x utilisé pour la photographie, barre d’échelle 5 μm. E) Un marqueur Mitochondrial GFP peut être utilisé pour identifier les mitochondries dans le soma et les exophers. Montré est un adulte ALM soma exprimant Pmec-4mCherry et mito::ROGFP, qui localise à la matrice mitochondriale. mito::ROGFP exprimé seul, sans le mCherry, peut également être facilement utilisé pour identifier les neurones et le score pour les exophers qui comprennent les mitochondries. Souche : ZB4528 bzIs166[Pmec-4mCherry]; zhsEx17 [Pmec-4mitoLS::ROGFP]. Objectif 100x utilisé pour la photographie; barre d’échelle 5μm. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 5 : Cycle de développement de l’identification de C. elegans et L4. (A) À 20 °C, un œuf prend environ 9 heures à éclore une fois pondus par la mère. (B) Un animal nouvellement éclos est au stade larvaire 1 (L1) et mue dans une larve L2 après 12 heures. (C) Les animaux restent dans les stades larvaires L2 etDL3 pendant environ 8 heures chacun. (E) Les animaux adolescents sont considérés comme le quatrième stade larvaire (L4) et sont marqués par une vulve en développement remarquable qui apparaît comme un croissant blanc près du corps moyen. La présence de ce croissant permet d’identifier et de cueillir facilement les animaux mis en scène L4 afin d’établir des cultures synchronisées qui facilitent plus tard la notation des exophers. Les animaux restent au stade L4 pendant environ 10 heures avant leur mue finale chez les adultes gravid, F) identifiés par le développement d’ovules, le spermatheca visible, et l’initiation de la ponte. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 6 : Préparation d’un tampon d’agar à glissière de lame de microscope. (A) Préparer deux diapositives avec une seule bande de ruban de laboratoire placée dans le sens de la longueur sur le dessus. Placez une lame de microscope non scotchée entre les deux comme sur la photo. B) Placez une goutte d’agarose en fusion sur le dessus de la glissière. (C) Placez une lame propre doucement sur le dessus de la goutte, en appuyant sur l’agarose dans un tapis de cercle dégonflé. (D) Retirez les diapositives enregistrées, qui agissent pour accomplir un aplatissement uniforme de l’agar qui est nécessaire pour créer un tampon uniforme. (E) Retirez la lame supérieure une fois que le tampon d’agarose a séché. (F) Pipette une solution paralytique (levamisole ou tétramisole) sur le dessus de l’agar pad. (G) Choisissez les animaux mis en scène de manière appropriée dans le paralytique. (H) Couvrez doucement les animaux d’un couvercle et assurez-vous que les animaux sont vivants. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}

Figure 7 : Caractères d’exophers et critères d’identification de l’exophé. (A) Critères généraux qui identifient un exopher. (B) Comparaisons de diamètre entre le soma d’envoi et l’exopher extrudé, mesurée en μm. Somas adultes ALM, N=35, souche : ZB4065 bzIs166[Pmec-4mCherry] - 6,53 μm taille moyenne de soma et 3,83 μm taille moyenne d’exopher. (C) Définition des critères de différenciation entre un domaine exopher et un exopher en herbe. (D) Le plus souvent, les neurones individuels font un grand exopher, qui se divise plus tard ou des fragments que l’hypoderme tente de dégrader son contenu. Pourtant, plusieurs exophers peuvent être observés à côté d’un neurone tactile qui pourrait dériver soit de multiples événements exopher d’un neurone ou alternativement, exophers peuvent également bourgeonner ou se fragmenter. L’origine de plusieurs entités de l’exopher ne peut être déterminée qu’à l’aide de la microscopie time lapse. Le dessus représente un soma de neurone tactile ALMR avec un seul exopher lointain. Le bas représente un soma de neurone tactile ALMR avec de multiples extrusions exopher-like. (E) Caractéristiques morphologiques communes dans les somas adultes de neurone tactile d’ALM qui peuvent être confondus avec des événements d’exopher. En haut à gauche - Un soma ALM distendé, sans domaine exopher clair ou site de constriction. Milieu supérieur - Les neurones peuvent avoir de petites protubérances extracellulaires qui peuvent être analogues aux exophers, mais ne répondent pas aux critères d’exigence de taille pour être considérés comme un exopher. En haut à droite – Avec l’âge, les neurones tactiles peuvent développer des excroissances le long de leur neurite mineure. Souvent, le matériel mCherry peut être recueilli à la pointe de l’excroissance de la neurite. Ce n’est pas marqué comme un exopher si le mCherry recueilli ne répond pas aux exigences de taille exopher-à-soma. En bas représente les neurones ALM adultes qui ont des critères de définition pour un domaine exopher ou un exopher. Botom gauche - ALM soma qui a un domaine exopher proéminent qui inclut sélectivement cytosol mCherry et mCherry marqué agrégats. Le site de constriction de domaine exopher est marqué par des flèches et répond aux critères de taille (au moins1/5 e de la taille du soma). Le plus grand diamètre du domaine exopher est presque 1/3 plus grand que le diamètre du site de constriction, répondant aux critères d’un événement exopher. Milieu inférieur - ALM soma qui a un exopher en herbe en avant qui répond aux critères de taille. Il y a un site de constriction clair. En bas à droite - ALM soma qui a un exopher attaché rempli de mCherry qui répond aux exigences de taille exopher. L’exopher est attaché par un mince filament de raccordement. Toutes les images proviennent de la souche ZB4065 bzIs166[Pmec-4mCherry]. Veuillez cliquer ici pour voir une version plus grande de ce chiffre.
{kind=link}
Discussion
La caractérisation des mécanismes moléculaires in vivo de l’élimination des agrégats et des organelles sous la forme de grands exophes n’en est qu’à ses balbutiements. Il reste à régler les questions relatives à la désignation des cargaisons pour expulsion, à la collecte polarisée de ces cargaisons dans la cellule, à la réglementation de la décision de générer des exophers, à la machinerie qui sert de médiateur aux extrusions et à l’interaction des exopherons avec la machinerie dégradatrice d’une cellule voisine. En outre, la visualisation in vivo des connexions tubulaires qui peuvent passer des matériaux biologiques qui incluent le calcium, les agrégats, et les mitochondries est intéressante et la biologie sous-étudiée dans son propre droit. Les questions de savoir pourquoi certaines cellules sont plus sujettes à la production d’exopher que d’autres sont également non résolues, mais peuvent commencer à être génétiquement disséquées avec les approches décrites dans ce protocole.
Décrit en détail dans ce protocole sont les approches pour atteindre la notation reproductible de la production d’exopher, avec l’attention à distinguer les exopheurs des somas cellulaires voisins, le moment des analyses pour capturer le pic de la production d’exopher, et le contrôle strict des conditions de croissance pour éliminer les contraintes involontaires qui peuvent moduler les niveaux d’exopher. La distinction du grand exopher précoce, ou la dispersion de la « nuit étoilée » dans l’hypoderme environnant peut être quantité comme preuve de la production d’exopher. Cela étant dit, les neurones exprimant mCherry dans des conditions basales sont le plus souvent associés à 5-25% des neurones d’un type spécifique produisant un exopher. L’introduction contrôlée de conditions de stress pourrait être appliquée pour augmenter la production d’exopher à la détection aussi élevé que 90% des neurones produisant des extrusions, particulièrement utile pour les écrans génétiques ou pharmacologiques pour les modificateurs.
Dans les maladies neurodégénératives humaines, de grands agrégats peuvent se transférer des neurones malades dans les cellules voisines pour favoriser la propagation de la pathologie. Le mécanisme d’exopher pourrait se produire par l’intermédiaire d’un mécanisme conservé utilisé pour l’extrusion globale à travers phyla. Définir les molécules in vivo qui améliorent l’efficacité de ce processus (considéré comme un contrôle plus efficace de la protéostase) ou bloquent- ils pourraient être exploités pour influencer la conception de nouvelles stratégies de lutte contre les maladies neurodégénératives multiples. En tant que tel, le protocole décrit ici pourrait être utilisé pour les écrans de mutagenèse génétique classique, les écrans RNAi à l’échelle du génome qui assomment systématiquement les gènes pour identifier les exhausteurs et les suppresseurs, ou pour des études d’intervention médicamenteuse qui identifient les modificateurs pharmacologiques candidats de ce processus. L’approche est simple, bien que quelque peu laborieuse. Les exophers sont si grands qu’ils peuvent être vus avec un microscope à dissection à grossissement élevé. Pourtant, les neurones C. elegans sont relativement petits et regarder leurs organites ou leurs membranes nécessitent des images confocales de puissance supérieure et est un processus lent. Les options pour un débit plus élevé pourraient impliquer des approches d’imagerie à haute teneur en format de plaque multi-puits.
L’application d’une approche normalisée à la notation exopher devrait sous-tendre une dissection génétique concertée du processus par lequel les neurones peuvent organiser et éliminer les débris cellulaires.
Déclarations de divulgation
Aucun
Remerciements
Nous reconnaissons les subventions suivantes aux NIH : R01AG047101 et R37AG56510. Les membres des laboratoires Driscoll et Grant ont largement contribué à l’élaboration et à l’ajustement des protocoles décrits, avec des expériences rigoureuses et une communication solide.
matériels
| Name | Company | Catalog Number | Comments |
| 95B Scientific CMOS camera | Photometrics Prime | ||
| 1,000 μL low retention tips | Sarstedt | ||
| 10 mL serological pipette | Appleton Woods | CC214 | |
| 10 μL low retention tips | Sarstedt | 70.1130.105 | |
| 13% sodium hypochlorite | Acros Organics | AC219255000 | |
| 15 mL centrifuge tubes | Fisher Scientific | 05-539-12 | |
| 2 L erlenmeyer flasks | Scientific Laboratory Supplies | FLA4036 | |
| 25 mL serological pipette | Appleton Woods | CC216 | |
| 300 μL low retention tips | Sarstedt | 70.765.105 | |
| 50 mL serological pipette | Appleton Woods | CC117 | |
| 5-Fluoro-2'-deoxyuridine 98% | Alfa Aesar | L16497.ME | |
| 9 cm sterile Petri dishes | Fisher Scientific | 11309283 | |
| absolute ethanol | Vwr | 20821.33 | |
| Agar | Sigma Aldrich | A1296 | |
| C. elegans strain wild type | Supplied by CGC | N2 | C. elegans strain |
| calcium chloride dihydrate | Sigma Aldrich | C3881 | |
| cholesterol | Acros | 110190250 | |
| dibasic sodium phosphate | Sigma Aldrich | S3264 | |
| E. coli strain OP50 | Supplied by CGC | Op50 | E coli strain |
| FBS10 Standard microscope | Meyer Instruments | KSC 410-1-100-1 | FBS10 Standard with Plate Base, 100/100 Trinocular Head and Flip zoom |
| glass pipette 270 mm | Fisherbrand | FB50255 | |
| Heraeus Multifuge X3R | Thermofisher scientific | 75004515 | |
| Inoculating Spreaders | Fisher Scientific | 11821741 | |
| LB medium capsules | MP biomedicals | 3002-031 | |
| LDI – Laser Diode Illuminator | 89 North | ||
| levamisole | Sigma Aldrich | 16595-80-5 | |
| M4 multipette | Eppendorf | 4982000012 | |
| magnesium sulphate | Sigma Aldrich | M7506 | |
| monobasic potassium phosphate | Sigma Aldrich | P0662 | |
| Multitron Standard shaking incubator | Infors HT | INFO28573 | |
| Nalgene 1 L Centrifuge pots | Fisher Scientific | 3120-1000 | |
| P10 pipette | Eppendorf Research Plus | 3123000020 | |
| P1000 pipette | Eppendorf Research Plus | ||
| P200 pipette | Eppendorf Research Plus | 3123000055 | |
| pipeteboy 2 | VWR | 612-0927 | |
| Polystyrene microbeads | Sigma Aldrich | MFCD00131491 | |
| RC5C plus floor mounted centrifuge | Sorvall | 9900884 | |
| Reusable ringed cytology slides | ThermoFisher Scientific | 22037242 | |
| SK4005 zdIs5[Pmec-4GFP] | contract Driscoll lab | GFP expressed in touch neurons | |
| sodium chloride | Sigma Aldrich | 13422 | |
| Sodium hydroxide | Fisher Chemical | S/4880/53 | |
| Tactrol 2 Autoclave | Priorclave | ||
| Triton-X | Thermofisher scientific | 28313 | |
| Tween 20 | Sigma Aldrich | 9005-64-5 | |
| X-Light V2 Spinning Disk Confocal Unit | CrestOptics | ||
| ZB4065 bzIs166[Pmec-4mCherry] | contract Driscoll lab | mCherry expressed in touch neurons | |
| ZB4067 bzIs167[Pmec-4mitogfp Pmec-4mCherry4]; igIs1[Pmec-7YFP Pmec-3htt57Q128::cfp lin-15+] | contract Driscoll lab | Q128 expressed in touch neurons | |
| ZB4509 bzIs166[Pmec-4mCherry]; bzIs168[Pmec-7LMP-1::GFP] | contract Driscoll lab | mitoROGFP expressed in touch neurons | |
| ZB4528 bzIs166[Pmec-4mCherry]; zhsEx17 [Pmec-4mitoLS::ROGFP] | contract Driscoll lab | autophagy marker expressed in touch neurons | |
| ZEISS Axio Vert.A1 | Zeiss |
Références
- Davis, A. A., Leyns, C. E. G., Holtzman, D. M. Intercellular Spread of Protein Aggregates in Neurodegenerative Disease. Annual Review of Cell and Developmental Biology. 34, 545-568 (2018).
- Davis, C. H., et al. Transcellular degradation of axonal mitochondria. Proceedings of the National Academy of Sciences of the United States of America. 111 (26), 9633-9638 (2014).
- Torralba, D., Baixauli, F., Sanchez-Madrid, F. Mitochondria Know No Boundaries: Mechanisms and Functions of Intercellular Mitochondrial Transfer. Frontiers in Cell and Developmental Biology. 4, 107 (2016).
- Stahl, P. D., Raposo, G. Extracellular Vesicles: Exosomes and Microvesicles Integrators of Homeostasis. Physiology (Bethesda, Md.). 34 (3), 169-177 (2019).
- Melentijevic, I., et al. C-elegans neurons jettison protein aggregates and mitochondria under neurotoxic stress. Nature. 542 (7641), 367 (2017).
- Nussbaum-Krammer, C. I., Park, K. W., Li, L., Melki, R., Morimoto, R. I. Spreading of a prion domain from cell-to-cell by vesicular transport in Caenorhabditis elegans. PLoS Genetics. 9 (3), 1003351 (2013).
- Tyson, T., et al. Novel animal model defines genetic contributions for neuron-to-neuron transfer of alpha-synuclein. Scientific Reports. 7, (2017).
- Babcock, D. T., Ganetzky, B. Transcellular spreading of huntingtin aggregates in the Drosophila brain. Proceedings of the National Academy of Sciences of the United States of America. 112 (39), 5427-5433 (2015).
- Pearce, M. M. P., Spartz, E. J., Hong, W., Luo, L., Kopito, R. R. Prion-like transmission of neuronal huntingtin aggregates to phagocytic glia in the Drosophila brain. Nature Communications. 6, 6768 (2015).
- Fu, H., Li, J., Du, P., Jin, W., Cui, D. Metabolic wastes are extracellularly disposed by excretosomes, nanotubes and exophers in mouse HT22 cells through an autophagic vesicle clustering mechanism. bioRxiv. 10 (1), (2019).
- Ghose, P., Park, E. C., Tabakin, A., Salazar-Vasquez, N., Rongo, C. Anoxia-reoxygenation regulates mitochondrial dynamics through the hypoxia response pathway, SKN-1/Nrf, and stomatin-like protein STL-1/SLP-2. PLoS Genetics. 9 (12), 1004063 (2013).
- Cannon, M. B., Remington, S. J. Redox-sensitive green fluorescent protein: probes for dynamic intracellular redox responses. A review. Methods in Molecular Biology. 476, 51-65 (2008).
- Perkins, L. A., Hedgecock, E. M., Thomson, J. N., Culotti, J. G. Mutant sensory cilia in the nematode Caenorhabditis elegans. Developmental Biology. 117 (2), 456-487 (1986).
- Stiernagle, T. Maintenance of C. elegans. WormBook: The Online Review of C. Elegans Biology. , 1-11 (2006).
- Sutphin, G. L., Kaeberlein, M. Measuring Caenorhabditis elegans life span on solid media. Journal of Visualized Experiments. (27), (2009).
- Mitchell, D. H., Stiles, J. W., Santelli, J., Sanadi, D. R. Synchronous growth and aging of Caenorhabditis elegans in the presence of fluorodeoxyuridine. Journal of Gerontology. 34 (1), 28-36 (1979).
- Weicksel, S. E., et al. A novel small molecule that disrupts a key event during the oocyte-to-embryo transition in C. elegans. Development. 143 (19), 3540-3548 (2016).
- Dong, L., et al. Reversible and long-term immobilization in a hydrogel-microbead matrix for high-resolution imaging of Caenorhabditis elegans and other small organisms. PloS One. 13 (3), 0193989 (2018).
- Toth, M. L., et al. Neurite sprouting and synapse deterioration in the aging Caenorhabditis elegans nervous system. Journal of Neuroscience. 32 (26), 8778-8790 (2012).
- Conte, D., MacNeil, L. T., Walhout, A. J. M., Mello, C. C. RNA Interference in Caenorhabditis elegans. Current Protocols in Molecular Biology. 109, (2015).
- Calixto, A., Chelur, D., Topalidou, I., Chen, X., Chalfie, M. Enhanced neuronal RNAi in C. elegans using SID-1. Nature Methods. 7 (7), 554-559 (2010).
- Maher, K. N., Catanese, M., Chase, D. L. Large-scale gene knockdown in C. elegans using dsRNA feeding libraries to generate robust loss-of-function phenotypes. Journal of Visualized Experiments. (79), e50693 (2013).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.