
Method Article
Количественные подходы для скоринга in vivo Neuronal Aggregate и Organelle Extrusion в больших экзоферных пузырьках в C. elegans
* Эти авторы внесли равный вклад
В этой статье
Резюме
В этом протоколе описаны подходы к обнаружению и количественной оценке крупных агрегатных и/или органелловых экструзий, производимых клетками C. elegans в виде мембранных экзоферов. Мы описываем штаммы, условия роста, критерии скоринга, сроки и соображения микроскопии, необходимые для облегчения вскрытия этого механизма высылки мусора.
Аннотация
Токсичность неправильно сфоленные белки и митохондриальной дисфункции являются ключевыми факторами, которые способствуют возрастных функциональных нейронов снижение и нейродегенеративных заболеваний между видами. Хотя эти нейротоксические проблемы уже давно считается клеточной внутренней, значительные доказательства в настоящее время подтверждает, что неправильное использование белков болезни человека, происходящих в одном нейроне может появиться в соседних клетках, явление, предлагаемое для содействия патологии распространения в человеческих нейродегенеративных заболеваний.
C. elegans взрослых нейронов, которые выражают агрегации белков может выдавить большие (4 мкм) мембраны окруженные пузырьки, которые могут включать в себя агрегированный белок, митохондрии, и лизосомы. Эти большие пузырьки называются "экзоферами" и отличаются от экзосом (которые примерно в 100 раз меньше и имеют различный биогенез). Выбрасывать клеточный мусор в экзоферах может происходить с помощью консервативного механизма, который представляет собой фундаментальную, но ранее непризнанную ветвь протеостаза нейронов и митохондриального контроля качества, относящуюся к процессам, с помощью которых агрегаты распространяются при нейродегенеративных заболеваниях человека.
В то время как экзоферы были в основном изучены у животных, которые выражают высокую копию трансгенных mCherry в пределах сенсорных нейронов, эти протоколы одинаково полезны в изучении экзофергенеза с использованием флуоресцентно помеченных органелл или других белков, представляющих интерес в различных классах нейронов.
Здесь описаны физические особенности экзоферов C. elegans, стратегии их обнаружения, критерии идентификации, оптимальные сроки количественной оценки и протоколы роста животных, которые контролируют стрессы, которые могут модулировать уровни производства экзоферов. Вместе детали протоколов, изложенных здесь, должны служить для установления стандарта количественного анализа экзоферов в лабораториях. Этот документ призван служить ресурсом в этой области для лабораторий, стремящихся разработать молекулярные механизмы, с помощью которых производятся экзоферы и на которые экзоферы реагируют соседние и отдаленные клетки.
Введение
Нейротоксические проблемы агрегатов и дисфункциональных митохондрий уже давно считаются клеточными, но в последнее время стало ясно, что неправильно сфоленные белки болезни человека, происходящие в одном нейроне, также могут распространиться на соседние клетки, способствуяпатологии 1. Аналогичным образом, млекопитающие митохондрии могут быть отправлены из клетки их первоначального производства для трансклеточнойдеградации 2 или для спасения митохондриальных популяций в проблемных соседнихклетках 3. Vesicles различных размеров, как правило, наблюдается для передачи клеточных материалов в соседние клетки или жидкости окрестности4. Некоторые экструдированы пузырьки приближаются к размеру средней нейрональной сомы (средняя сенсорная нейронная сома 6 мкм) и могут вместить большие агрегаты и органеллы.
Яркий пример большого экструзии пузырьков, которые могут нести белковые агрегаты и органеллы происходит в C. elegans сенсорных рецепторов нейронов, которые выражают высокое число копий репортер построить кодирование вредных агрегации подверженных, деградации устойчивостью mCherry5. Экструзии от сенсорных нейронов, называемые экзоферами, имеют средний диаметр 4 мкм, избирательно включают mCherry или другие агрегаты, и поставляются непосредственно в соседний гиподермис, который обычно окружает нейроны сенсорного рецептора. Гиподермис попытки лизосомы основе деградации, но некоторые неудобоваримые содержимое, такие как mCherry агрегаты могут быть повторно экструдированы гиподермиса в заполненные жидкостью псевдокоэком животного, из которых mCherry может быть принято удаленных клеток мусорщика называется coelomocytes длядолгосрочного хранения ( Рисунок 1, Рисунок 2)5.
Большие экструдированы экзоферные пузырьки покидают клетку, окруженную плазменной мембраной сенсорного рецептора, и могут содержать агрегированные белки болезни человека, митохондрии и лизосомы. Процесс производства экзофера, как представляется, включает в себя сортировку потенциально токсичных видов (например, агрегация подверженных выраженной mCherry отделена от растворимых, безобидных белков, как GFP, который остается в основном в нейрональной сомы). Таким образом, направленное изгнание угрожающих сущностей осуществляется нейроном5. Протеостаз вызов, такие как стресс, вызванный аутофагией нокдаун, MG132-опосредованное протеасомное ингибирование, или трансгенное выражение белков болезни человека, таких как болезнь Генттона связанных расширенный полиглутамин No 128 или болезни Альцгеймера причастных фрагментА 1-42, может увеличить число нейронов, которыепроизводят экзосферы 5.
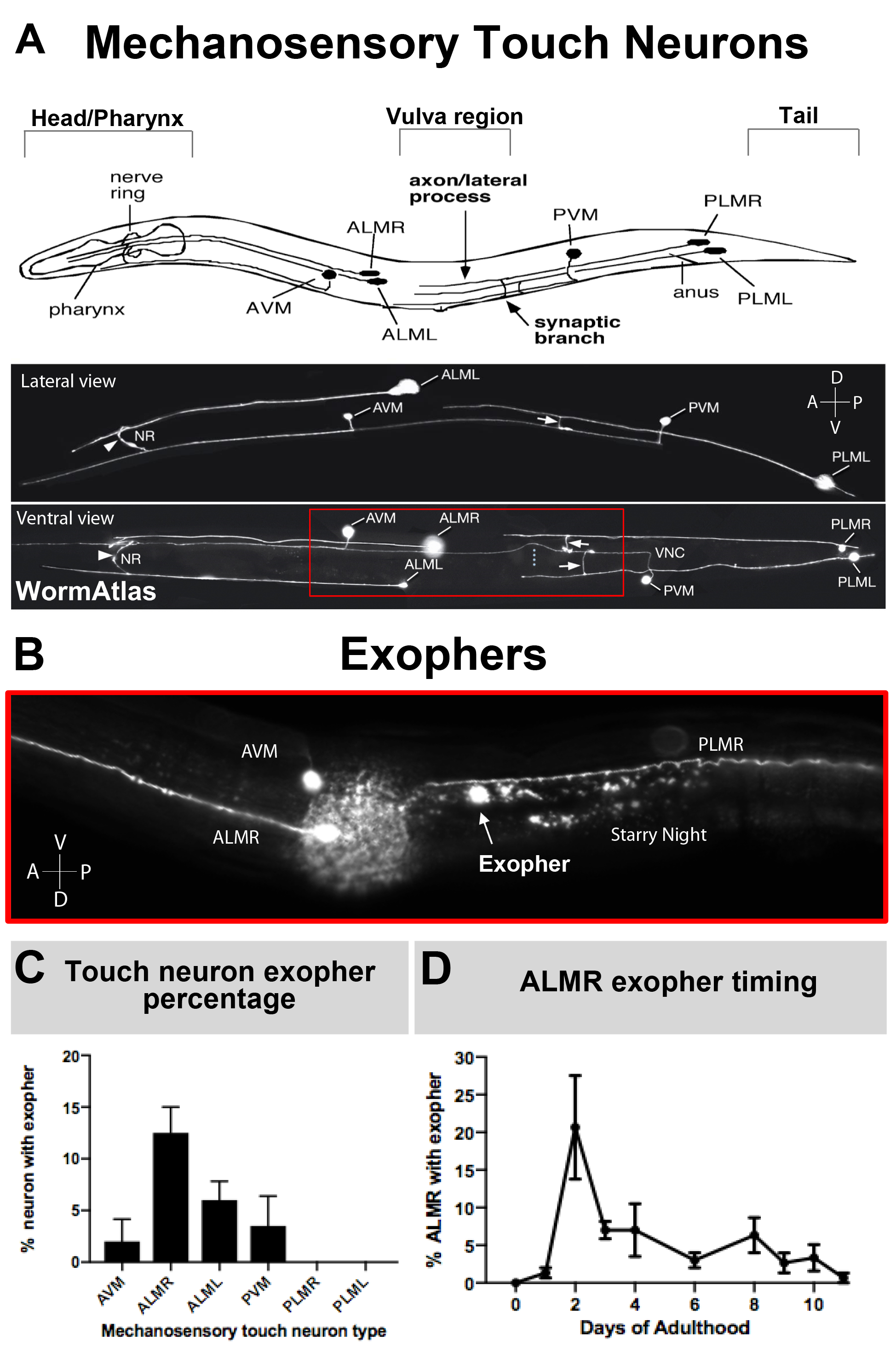
Поскольку экзоферы были задокументированы совсем недавно, то, что известно об их биологии, заслуживает описания. Экзоферы были обнаружены в, и являются наиболее хорошо изучены в, C. elegans сенсорных рецепторов нейронов. Есть шесть C. elegans механосенсорные сенсорные нейроны, которые имеют клеточные тела, распределенные по всему телу(рисунок 3A) и называются микротрубочные клетки, потому что их ультраструктурные особенности отличительные 15 микротрубочек протофиламента. Нейроны сенсорного рецептора являются передней AVM (передний брюшной микротрубообразной нейрон), ALMR, и ALML (передние боковой микротрубообразной нейронов справа и слева), более центральный PVM (задний брюшной микротрубочек нейрон), и задний PLMR и PLML (задний боковой микротрубочек нейронов вправо и влево) в хвосте. Интересно, что шесть нейронов сенсорных рецепторов производят экзоферы с разной скоростью, несмотря на выражение одного и того же наступательного трансгена(рисунок 3C). Из шести механосенсорных сенсорных нейронов, нейрон ALMR подвергается экзофергенезу чаще, чем другие сенсорные нейроны. Квантиция числа экзоферов от сенсорных нейронов, таким образом, как правило, устанавливается путем сосредоточения внимания на ALMR.
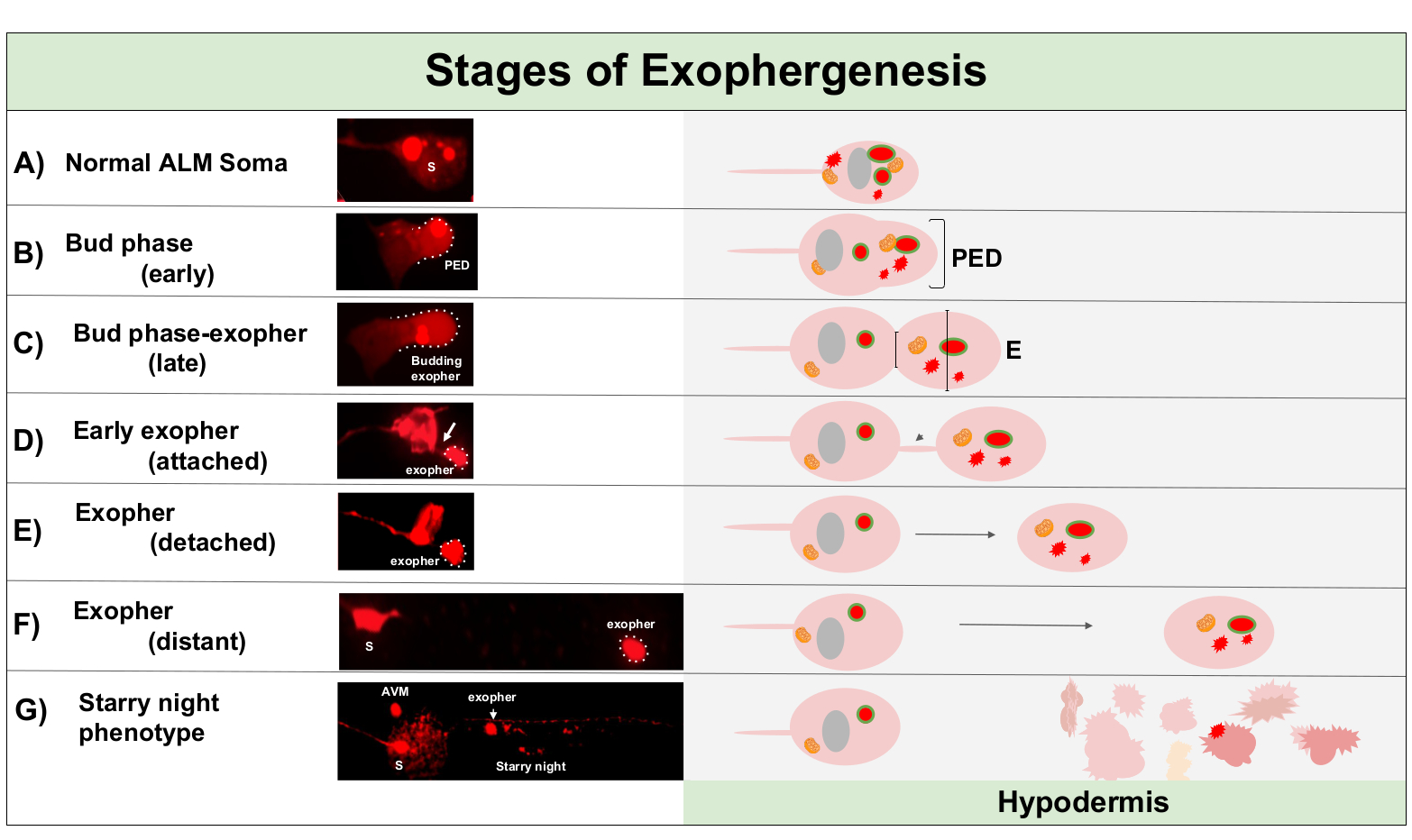
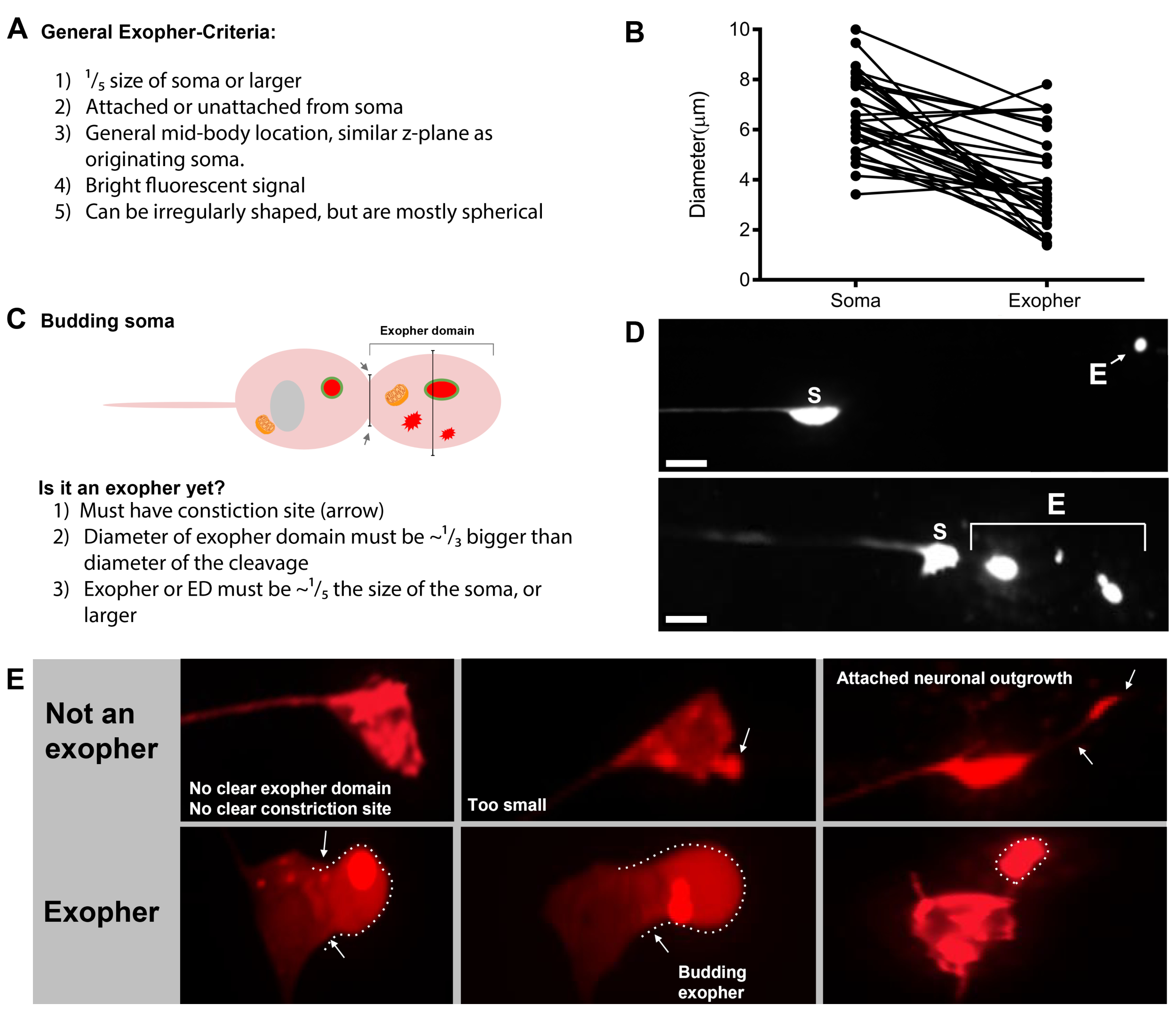
Экзофергенез является динамичным процессом, который обычно начинается с отеком цитоплазмы нейронов(рисунок 1A-B). Клеточное содержимое, органеллы или белковые агрегаты собираются в одну сторону нейрональной сомы, чаще всего к задней части нейрона ALMR (вдали от проецирования нейрита), образуя домен до экзофера (PED) (Рисунок 1B). Раннее выступ наблюдается, как PED начинает проецировать наружу, образуя узнаваемый выступающий бутон. Поздний бутон определяется, когда самый широкий диаметр домена до экзофера примерно на 1/3 больше диаметра сужения шеи сома-экзофера(рисунок 1С). Экзоферы могут быть выброшены практически в любом направлении от сомы, но большинство экзоферов выходят из тела клетки и остаются примерно в той же фокусной плоскости, что и возникаюющая сома.
Экзофер может отойти от возникают сомы, как шея бутон сужается в тонкую нить. Экзоферы могут оставаться прикрепленными к соме через эту нить(рисунок 1D, стрелка),а затем могут стать отделенными. Содержимое клеток, таких как кальций, агрегаты и митохондрии могут быть переданы через эту нить в прикрепленныйэкзофер 5, хотя основная часть экструдированного материала положена в отсек экзофера массивным начинающим событием. Экзоферы считаются зрелыми, когда нет видимой соединительной трубки или тонкой нити и экзофер полностью отделен от отправки сомы(рисунок 1E).
Экзоферы производства C. elegans коснуться нейронов сразу же сталкиваются с гиподермисом, ткань, которая окружает сенсорный нейрон. Чаще всего, экзофер везикулы, как представляется, путешествия в пределах гиподермиса задней к хвосту, и может быть довольно далеко от сомы, прежде чем содержимое экзофора появляются целевые для деградации (например, расстояние может быть 100 мкм от сомы (Рисунок 1F)). Флуоресцентный экзофер vesicle распадается на многие мелкие пузырьки в гиподермиса, принимая на вид называют "звездная ночь"(рисунок 1G и рисунок 2). В "звездной ночи" этапе, прокалывают флуоресцентный материал можно наблюдать разбросаны по гиподермальной синцитии во многих небольших точках флуоресценции по сравнению с оригинальным одиночным экзофером. Звездная ночь может выглядеть пунктуальной при низком увеличении и с более высоким увеличением, может выглядеть пунктуальной и / или сетевой в гиподермиса. Флуоресцентный сигнал звездной ночи, как правило, тусклым, чем экзофер и нейронно выраженной флуоресценции (Рисунок 2B-C). Разгон mCherry во многие пунктуальные пузырьки, как полагают, связаны фагосом созревания и слияния с эндосомной / лизосомальной сети гиподермальной клетки. Некоторые экзосферные материалы, вероятно, деградируют в гиподермальной лизосомальной сети, но остаточные виды, устойчивые к деградации (такие как агрегаты mCherry), выбрасываются из гиподермиса в псевдоколом, жидкий отсек, который может содержать клеточный мусор. Флуоресцентный материал позже принимаются в удаленных клеток мусорщика называется coelomocytes (Рисунок 2C), который может сосредоточиться, хранить, и снова попытка деградации mCherry.
Явление агрегированного экструзии и переноса, как представляется, сохраняется через фила, будучи зарегистрированы в генетических моделях, таких как C. elegans5,,6,7 и D. melanogaster8,9, а также в нескольких моделях млекопитающих., Экзофер-как экструзии были зарегистрированы для клетокмлекопитающих 10, наблюдение, предполагающее, что сохраненные механизмы могут лежать в основе совокупности и изгнания органеллы. Таким образом, производство экзофера может быть сохраненным механизмом управления клеточным мусором, который представляет собой фундаментальную, но ранее непризнанную ветвь протеостаза нейронов и митохондриального контроля качества, которые при несбалансированности могут активно способствовать нейродегенеративному заболеванию. Идентификация молекул, участвующих в дискриминации мусора и сортировки, транспортировки в различные субклеточные локали, экструзии, формирования / схия трубчатого соединения, связывающего сомы и позднего экзофера, и признание большого экструдированного пузырька для удаленной деградации соседней клетки остаются для будущей работы. Исследования в моделях нематод и мух будут иметь решающее значение для определения механизмов агрегированного и органельного сбора и переноса, используя объективные генетические подходы и мощные клеточные биологические инструменты, предлагаемые этими моделями для выявления участвующих молекул в физиологическом контексте.
Критические первые шаги в расшифровке механизмов, задействованных в биологии экзоферов, включают определение протоколов воспроизводимой количественной оценки экзоферов in vivo. Модель C. elegans предлагает особое преимущество для таких усилий, так как тело прозрачно и экзоферы можно легко наблюдать, когда они содержат флуоресцентно помеченные белки или органеллы. Экзоферы, как сообщается, генерируются C. elegans дофаминергических нейронов PDE и CEP, ASE и ASER сенсорных нейронов, и красителя заполнения амфид нейронов5. Поскольку экзоферы, вырабатываемые нейронами сенсорных рецепторов, лучше всего характеризуются, основное внимание здесь уделяется использованию сенсорных нейронов для анализа экзоферов. Однако основной подход может быть применен для измерения производства экзофера из любой ячейки. Протоколы для обнаружения и количественного экзоферов производства C. elegans коснуться рецепторов нейронов, которые трансгенически выразить mCherry белка изложены, с акцентом на грузы, которые могут контролироваться и временные ограничения в скоринга. В этой статье определяются подходы к идентификации экзоферов in vivo, а также количественная оценка экологических и генетических условий, модулирующие производство экзоферов. Протоколы подчеркивают критическое внимание к постоянным неспокойных условиях для определения базового производства экзоферов и для сопоставлений по генотипам.
протокол
1. Штаммы, полезные для обнаружения экзоферов
- Выберите штамм, который выражает флуоресцентные грузы в нейронах C. elegans, чтобы легко визуализировать экзоферы.
ПРИМЕЧАНИЕ: Таблица 1 перечисляет штаммы, которые были использованы для визуализации экзоферов, вырабатываемых всенсорных нейронах рецепторов 5,,11,,12. В принципе, любая клетка или нейрональный тип может быть проверена на производство экзофера с помощью клетки или ткани конкретных промоутер диск выражение флуоресцентного белка, который агрегируется или иным образом выбран для экструзии. - Кроме того, используйте красителя заполнения анализ для визуализации экзоферов в амфидных нейронов головы, которые открыты для окружающей среды и подложкидля заполнения 5,13.
2. Средства массовой информации роста
- Подготовка стандартных средств роста нематод (NGM) к культуре штаммов в соответствии со стандартнымиметодами 14,15.
ПРИМЕЧАНИЕ: Недостаток пищи, или фтор-дезиуридин (FuDR), часто используется для блокирования производства потомства, и может резко повлиять на производство экзоферона. Держите население постоянно кормили (избежать даже коротких периодов бактериального истощения пищи) и поддерживать животных при постоянной температуре.
3. Животноводство имеет решающее значение для последовательного производства экзоферов
- Поднимите животных на последовательных средствах массовой информации и с последовательными бактериальными источниками пищи. У животных не должно быть бактериальной пищи, даже в течение коротких периодов времени, так как ограничение пищевых продуктов может резко изменить уровень производства экзоферов.
- Храните средства массовой информации рецепты и подготовки равномерной всей исследования.
ПРИМЕЧАНИЕ: Изменение мультимедиа может повлиять на базальные уровни производства экзоферов. Партии Agar могут влиять на базовые уровни экзофера, поэтому при изменении лотов поставок сделайте заметку о дате. Выбрось фондовые тарелки через две недели, чтобы обеспечить здоровую бактериальную пищу и предотвратить сушеный агар, который вызывает изменения в осмолярности агара, которые влияют на уровень экзофера. - Для базальных условий держите животных при постоянной температуре 20 градусов по Цельсию. Воспитание животных при переменных температурах (даже временные изменения температуры) может вызвать изменения в сроках производства максимальных экзоферов.
ПРИМЕЧАНИЕ: Температурная изменчивость не ограничивается культурными условиями. Колебания температуры во время экспериментов или на лабораторной скамейке могут быть ударными. Например, температура в комнате микроскопа не должна резко отличаться от культурного инкубатора или лабораторной скамейки. - Не используйте фармакологические антиф рождаемость вмешательства, потому что оплодотворенные яйца имеют решающее значение для раннего взрослого производства экзоферов.
ПРИМЕЧАНИЕ: Использование фтор-дезиуридина (FuDR)16 или C2217, следует избегать. При выполнении жизненных или старостарых экспериментов на животных, возрастных синхронизированных популяций должны поддерживаться путем физического удаления взрослых из их меньшего потомства, выбирая их на свежие пластины распространяются с бактериями, а не с помощью общих фармакологических анти-фертильности мероприятий. - Не используйте загрязненные культуры; повторное участие в экспериментах в случае биологического компромисса населения или пластины. Бактериальное или грибковое загрязнение может вызвать стрессы и метаболические изменения у животных и должно отсутствовать в экспериментальных популяциях.
- Чтобы максимизировать воспроизводимые результаты, поддерживайте культуры, по крайней мере, для двух здоровых, сытых, свободных от загрязнения поколений при 20 градусах Цельсия, прежде чем экспериментировать, чтобы избежать потенциальных экологически индуцированных эпигенетических изменений.
4. Возраст синхронизации для экзофера скоринга путем отбеливания, флотации сахарозы, или сбор личинок L4
- Держите экспериментальных популяций того же биологического возраста, как экзофер обнаружения моделей варьируются в зависимости от возраста взрослого и сравнение животных смешанных возрастных популяций может сбить с толку результаты. Всегда обеспечить успешную синхронизацию экспериментальных популяций животных, проверяя на "белый полумесяц" вульвы морфологии на этапе L4.
ПРИМЕЧАНИЕ: Как правило, пик производства экзофера для C. elegans механосенсорных нейронов ALMR происходит на взрослый день 2-3(рисунок 3D), как измеряется от дней после L4 этапе. Взрослый день 1 составляет 24 часа после L4 личинок этапе, который отличается "белый полумесяц" вульвы морфологии (Рисунок 5E). - Подготовка синхронизированных популяций яиц путем отбеливания gravid взрослых.
- Соберите gravid взрослых, наполненных яйцами, мыть животных, растущих на пластине NGM. Для мытья, затопить пластину с 1 мл буфера M9, трубчатые вверх и вниз, чтобы собрать жидкость с приостановлено животных и трубчатых в 1,5 мл микроцентрифуг трубки. Пеллет животных путем гравитационного заселения или мягкой центрифугации с помощью мини-центрифуги и удалить супернатант.
- Добавьте 150 МКЛ из 5M NaOH и 150 МКЛ 6% гипохлорита натрия (отбеливатель) в 1 мл в H2O и смешайте инверсией в течение примерно 5 минут.
ПРИМЕЧАНИЕ: Свежий раствор отбеливания гарантирует, что кутикула животного может быть нарушена для сбора яиц. Прогресс в нарушении кутикулы можно контролировать под рассеченным микроскопом; взрослые должны разбить и выпустить яйца в том смысле, что отбеливание должно быть остановлено. - Аккуратно центрифуга с миницентрифугой трубки на 20 с и удалить супернатант. Добавьте 1 мл буфера M9 и центрифуги снова, оставляя около 100 йл на верхней части гранулы.
- Повторите шаги 4.2.3 дважды, чтобы удалить следы раствора отбеливателя.
- Переусейте яйца в оставшемся объеме и перенесите на свежую семенную пластину NGM. Взрослые будут lysed, но многие жизнеспособные яйца должны быть в подготовке.
- Подготовка синхронизированных популяций по приуготовимому яйцекладу.
- Выберите 20 gravid взрослых к семенной пластине NGM, используя стандартные протоколыпередачи 14.
- Разрешить животным свободно ползать и откладывать яйца на 1,5 ч (мутант штаммов с низкими размерами выводка может потребовать введения более взрослых животных).
- Удалите всех взрослых животных из тарелки, собирая, оставляя синхронизированную популяцию яиц позади. Проверьте пластины несколько часов спустя, чтобы убедиться, что нет жизнеспособных взрослых были пропущены во время удаления взрослых.
- Подготовка синхронизированных популяций яиц путем выбора флотации сахарозы яиц.
- Собирайте животных и яйца из пяти пластин NGM, на которых гравидные животные откладывают яйца в течение по крайней мере 24 часов, заливая пластины раствором M9 с моющим средством 0,1% (например, Tween 20 или Triton X-100) и собирая в трубку 15 мл. Пеллет взрослых при мягкой центрифуге при комнатной температуре (2000 х г на 30 с).
- Удалить супернатант и мыть животных в 15 мл свежего M9 три раза, отбрасывая супернатант после каждой стирки, будучи уверенным, чтобы сохранить гранулы обогащены в животных и яиц.
- Сохранить 2 мл супернатанта и повторно помыть гранулы. Добавьте 2 мл 60% веса по объему сахарозы.
- Центрифуга при 2000 x г в течение 5 мин. Раствор теперь будет отображать верхнюю фазу, сильно обогащенную яйцами.
- Перенесите около 2,5 мл верхней фазы в новую трубку 15 мл и добавьте 10 мл M9.
- Смешайте инверсией в течение 1 мин, а затем центрифугу 2000 х г в течение 1 мин.
- Удалите супернатант и вымойте обогащенную яйцом гранулу в M9. 10-15 МКЛ яичной гранулы могут быть распределены по свежей OP50 семенами NGM пластины.
ПРИМЕЧАНИЕ: Этот метод готовит большое количество яиц; не позволяют собранным животным выбежать из OP50 E. coli пищи.
- Подготовка синхронизированных популяций путем сбора животных на этапе разработки L4.
- Выращивать животных на семенных пластинах NGM, как описано выше.
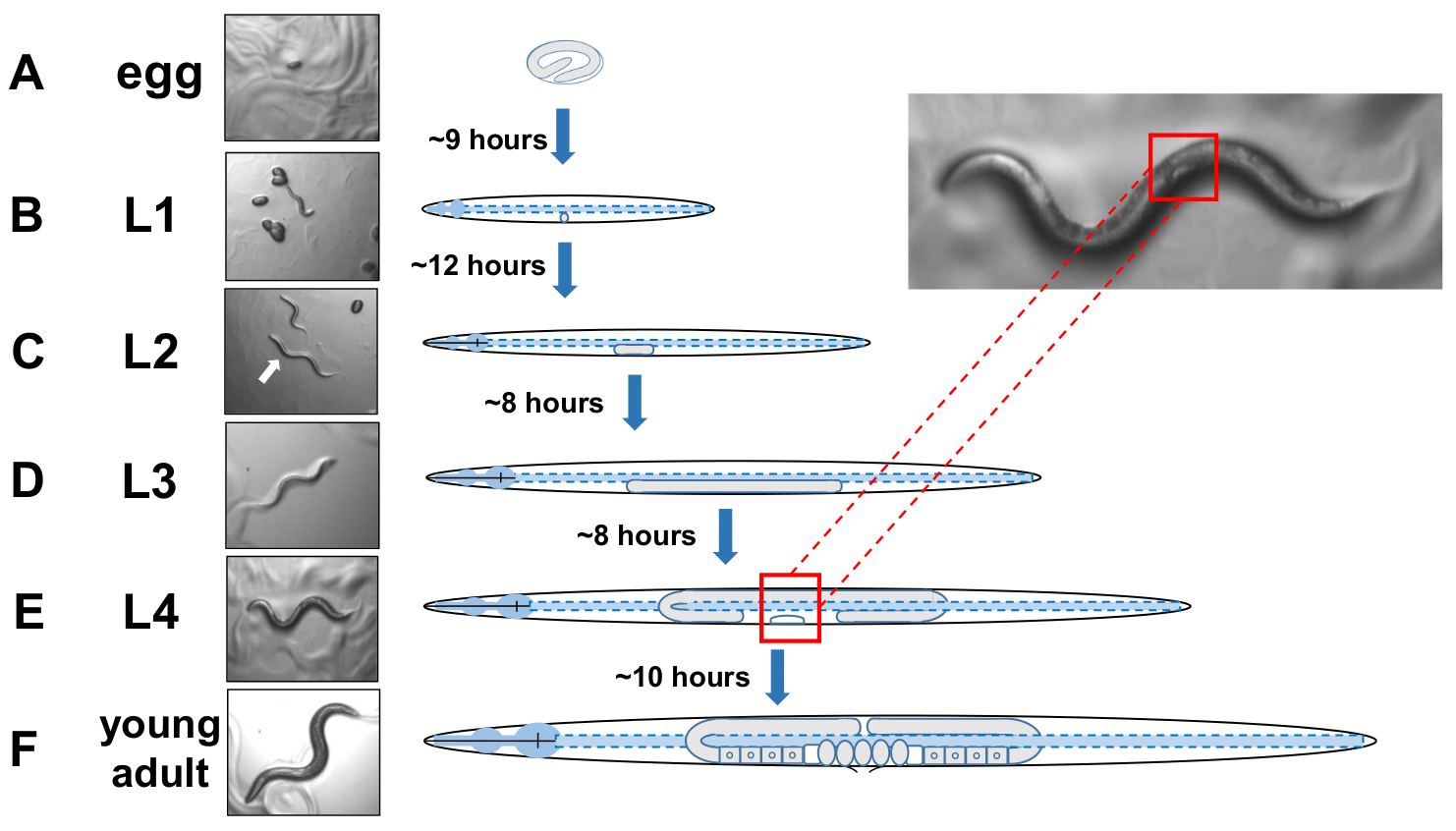
ПРИМЕЧАНИЕ: C. elegans развиваются в четырех дискретных стадиях. При 20 градусов по Цельсию, недавно заложенное яйцо занимает около 9 часов, чтобы люк (Рисунок 5A). Почтовый люк, животное проходит через личинку этапе 1 (L1) личинок этапе 4 (L4), с каждой стадии продолжительностью 8-12 часов между каждой линьки (Рисунок 5A-F). Таким образом, пластина, подготовленная путем инокуляции с яйцами должны иметь много L4 животных, чтобы забрать около 40 часов после яйца вводятся. - Определите L4-постановочных животных, найдя белый полумесяц полумесяца форму развивающихся вульвы (Рисунок 5E).
ПРИМЕЧАНИЕ: животные на стадии L4 однородны по размеру и пигментации тела. Выберите животных с белым полумесяцем к свежей пластине роста для изучения постановочных животных. На следующий день (24 часа спустя) следует считать взрослым день 1. - Оценка популяции животных ежедневно на взрослый день 2.
ПРИМЕЧАНИЕ: Экзоферы, как правило, забил в день 2 взрослой жизни, которая является пик производства экзофера в базальных условиях. Однако, поскольку пик экзофергенеза и сроки могут быть сдвинуты экологическими или генетическими изменениями, которые изучаются, рекомендуется забивать популяцию взрослых животных ежедневно в течение четырех дней, чтобы создать наиболее полную картину(рисунок 3D).
- Выращивать животных на семенных пластинах NGM, как описано выше.
5. Обнаружение экзоферов с помощью флуоресцентного микроскопа
- Наблюдайте за экзоферами с помощью микроскопа с высоким увеличением псевдо-стерео, который оснащен микроскопией флуоресценции.
- Обездвижить животных на пластинах NGM путем пипетки 100-200 МЛ 10-100 мМ левамизол / теттрамизол раствор на поверхности пластины NGM агара. Через 2-4 минуты животные парализованы и могут наблюдаться непосредственно на агарной пластине.
ПРИМЕЧАНИЕ: Иммобилизация лечения не являются абсолютно необходимыми, так что с обученным глазом, нейронной идентификации и присутствия экзофера может быть забил визуально после сканирования животных под микроскопом на пластине при определении того, является ли экзофер был произведен. - Наблюдайте за флуоресцентными нейронами, используя общее увеличение в 100 раз, чтобы выполнить обнаружение экзоферов вскрытом микроскопе.
ПРИМЕЧАНИЕ: Оценка событий экзофера с помощью микроскопии вскрытия позволяет осуществлять наблюдение за большим количеством животных с относительной легкостью непосредственно на агарных пластинах, на которых они воспитаны. - Live изображения и монтажа репортер штаммов для экзофер исследований с использованием конфокальные микроскопии
- Используйте конфокальный микроскоп для живого изображения внутриклеточной динамики и характеристик экзофергенеза.
ПРИМЕЧАНИЕ: Live Imaging является выгодным подходом для наблюдения тонкие детали производства экзофера, потому что производство экзофера является динамичным процессом. - Ограничьте движение животных для живой визуализации высокого разрешения с помощью удобных методов, включая использование левамизола или теттрамизола на 10-100 мМ или применение гидрогелевых полистироловых микробусов (диаметром 15 мкм, 30 мкм или 40мкм) 18.
- Используйте конфокальный микроскоп для живого изображения внутриклеточной динамики и характеристик экзофергенеза.
- Подготовка слайда для соединения и конфокальные микроскопии
- Гора 20-50 животных в иммобилизирующий агент на слайде микроскопа. Многоразовые кольчатые цитологические слайды, окрашенные поднятыми кольцами диаметром 13 мм, полезны для монтажа.
- Выберите живых животных в 5-20 йл паралитического, таких как 10-100 мМ левамизола или теттрамизола в окрашенном круге или на агар-площадке.
- Подождите 4 минуты до паралича, а затем накройте слайд крышкой (рекомендуем No 11/2 (0,16 - 0,19 мм) или No 2 (0,17 - 0,25 мм).
- Монтаж небольшого количества животных
- Не раздавить смонтированных животных; при наблюдении только несколько (менее 20) животных на слайде, есть риск дробления некоторых животных из-за неравного давления крышки. Этот риск может быть сведен к минимуму с помощью низкого процента агарозы площадку для монтажа.
- Сделайте 2-4% агарозы площадку слайд, а затем добавить 2-15 йл паралитического раствора на площадку. Имейте в виду, левамизол и теттрамизол диффузных в площадку, уменьшая их эффективную концентрацию.
- Гора, собирая животных в 2-15 йл капли паралитического раствора или микробусы, опираясь на агар площадку. Поместите крышку сверху и убедитесь, что животные нетронутыми18.
- Подготовка Агар площадки
- Чтобы подготовить 2% агар колодки, тепло 2% агарозы в растворе M9 и микроволновой печи, пока агароза находится в однородном и расплавленном состоянии.
- Для достижения агар площадку достаточного качества, альтернативные смешивания и микроволновой воды при низкой мощности менее чем за 20 секунд. Избегайте включения пузырьков воздуха в площадку, поместив кипящий агар на нагревательный блок и позволяя пузырькам подняться на поверхность.
- Используйте пипетки Pasteur, чтобы нарисовать агар из глубины расплавленного раствора ниже воскресшего пузыря.
- Подготовьте два лентой слайды и поместите по обе стороны от чистого стеклянного микроскопа слайд на плоской поверхности. Для того, чтобы записанные на пленку слайды поместите две 5-сантиметровые полоски лабораторной ленты на каждыйслайд (рисунок 6A).
- Используя трубу Pasteur, поместите одну каплю агара на чистый слайд микроскопа, зажатый между записанными на пленку слайдами(рисунок 6B).
- Тщательно и быстро, покрыть падение расплавленного агара с четвертой чистой слайд, поместив в через лентой слайды (Рисунок 6Cc).
ПРИМЕЧАНИЕ: Слайд должен осторожно нажмите расплавленный агар в сплющенный круг толщиной около 0,4 мм (толщина ленты) (Рисунок 6D). Агар должен быстро остыть. - Удалите верхний слайд, сдвинув его с(рисунок 6E). Агар колодки высыхают быстро и лучше всего использовать в течение нескольких минут. После того, как верхний слайд удаляется, используйте гель площадку немедленно для монтажа животных. Избегайте использования прокладок с пузырьками воздуха.
- Храните агарные прокладки до 30 минут, заключенные между двумя стеклянными горками. Сушеный агар заставляет животных слипаться и высыхать. Гора животных в пределах 2-15 мкл паралитического раствора или микробусов и крышка с крышкой; экран слайда в течение 20 минут после паралича и монтажа(рисунок 6).
ПРИМЕЧАНИЕ: Поскольку стрессовые условия могут изменить скорость экзофера, избегайте паралитики, которая может вызвать окислительный стресс (например: азид натрия) при скрининге на экзоферы.
- Обнаружение экзоферов с помощью конфокального микроскопа с вращающимся диском
- Наблюдайте за биологическими особенностями клеток, такими как органеллы и другое содержимое с 1,4 численными целями диафрагмы в 63x и 100x.
- Используйте программное обеспечение, способное управлять этапом и получением изображений, используя многомерное приобретение. Микроскопы и программное обеспечение для обработки изображений также должны подходить для визуализации и сбора данных, поскольку эти шаги включают стандартные подходы к визуализации.
6. Определение сенсорных нейронов и скоринга для экзоферов с установленными животными
- Гора парализована взрослого животного(рисунок 6).
- Определите желаемую плоскость. Используйте низкое увеличение яркого поля (10-40x) для определения подходящего З-плоскости животного, принимая к сведению позиционирование животного, ориентацию головы и хвоста, и расположение вульвы - которые являются ориентирами для более поздней нейронной и экзофернойидентификации (рисунок 3A и рисунок 5E).
- Сосредоточьтесь на сигнале флуоресценции выбранного репортера. Оставаясь в том же самолете, переключитесь на широкоугольный просмотр флуоресценции в 10-40x для выбранного цитозоляного репортера.
ПРИМЕЧАНИЕ: В этом примере флуоресцентное выражение определяется mec-4 механосенсорное прикосновение нейрона конкретных промоутер. Высокие массивы копий и различные флюорофоры имеют изменчивость выражения и, следовательно, переменную флуоресцентную интенсивность. При необходимости отрегулируйте. - Прокрутите в пределах оси Кью, чтобы наблюдать глубину животного и флуоресцентное выражение в фокусной плоскости. При этом подтвердите ориентацию головы и хвоста; голова/глотка будет иметь флуоресцентное нервное кольцо, и в этом случае хвост будет содержать 1-2 видимых PLM сома(рисунок 3A).
- Определить сенсорные нейроны
- Определите, установлено ли животное слева или справа(рисунок 3A).
ПРИМЕЧАНИЕ: Учитывая трехмерность животного, лучшее разрешение изображения осуществляется на стороне ближе всего к оптике. - Определите сому (ALM, ALMR, AVM) путем наблюдения - начните с головы, чтобы определить нервное кольцо и боковой нейрональных процессов.
- При увеличении в 10-40 раз медленно прокручивайте ось, чтобы определить прилагаемый процесс.
- После того, как процесс определен, следуйте его боковой в задней направлении к вульве, где сома будет очевидно, отмечен круглым телом клетки в конце процесса. После того, как наиболее в фокусе нейрональной сомы найдено, он может быть идентифицирован с помощью других нейронных ориентиров следующим образом:
- Используйте AVM, близлежащий брюшной нейрон, чтобы помочь назначить ориентацию животных. Если avM нейрон находится в той же плоскости, как ALM, то животное опирается на его стороне и нейрон снаружи, что плоскость ALMR . Если нейрон AVM находится не в той же плоскости, что и ALM, ближайший сенсорный нейрон к фокусной плоскости ALML.
- Определите нейрон PVM, другой брюшной сенсорный нейрон, расположенный рядом с хвостом, чтобы указать, находится ли передний сенсорный нейрон в той же плоскости. Если это так, сенсорный нейрон наблюдается ALML.
- Получите представление о положении других тел сомы, вблизи области интереса (флуоресцентные нейроны, расположенные по обе стороны от сомы), и во всех плоскостях, даже если это не возможно, чтобы получить глубокий нейрон установлен в четком фокусе.
ПРИМЕЧАНИЕ: Идентификация всех сенсорных нейронов сома имеет важное значение, потому что вне фокуса сома может быть ошибочно принято за экзоферы.
- Определите, установлено ли животное слева или справа(рисунок 3A).
7. Выявление и скоринг для экзоферов
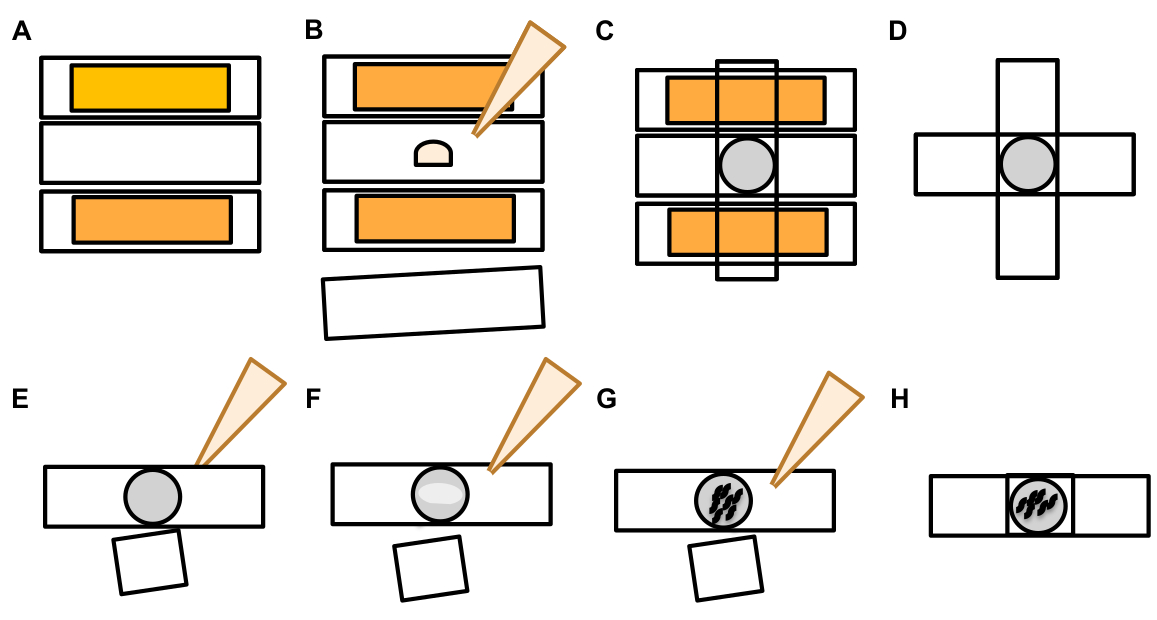
- После того, как сенсорный нейрон найден, проверить его на большие выступы (экзофер доменов) достаточно большой, чтобы считаться бутон экзофер, (достижение покрайней мере 1/5 й размер происходящих сома) (Рисунок 1C).
ПРИМЕЧАНИЕ: Средний экзофер измеряет около 2-8 мкм в диаметре, в то время как средняя сома a (B4065 bzIs166(Pmec-4::mCherry) животных измеряет 6-10 мкм в день 2 взрослых(рисунок 7B). - Если не наблюдается бутон или область экзофера, проинспектировать нейрональной сомы для прилагается тонкая нить, исходящей от сомы. Прикрепленные экзоферы, как правило, расположены ближе к исходной соме и в аналогичной плоскости.
ПРИМЕЧАНИЕ: Экзоферы не всегда остаются прикрепленными к соме. Обнаружение прикрепленной нити является окончательным признаком того, что объект является экзофером. - Чтобы определить незакрепленный экзофер, ищите содержимое экзофера. Экзоферы могут концентрировать исключенные флуоресцентные белки и поэтому часто ярче, чем сома.
ПРИМЕЧАНИЕ: Содержимое экзоферов неоднородно и изменчиво. Клеточные органеллы, такие как лизосомы и митохондрии, также могут быть экструдированы в экзоферах(рисунок 4C-E). - Ищите незакрепленные экзоферы в различных координационных плоскостях, чем самолет, в котором была найдена возникаюющая сома. Хотя было замечено, что экзоферы выступают из Сомы ALM в любом направлении, это типично, что экзоферы выступают от сомы, в заднем направлении от нейронального процесса.
- Проверьте наличие крупных сферических объектов, которые не расположены и идентифицируются как нейрональные сомы. Экзоферы могут быть неправильной формы, но, как правило, сферические структуры. Экзоферы деградируют с течением времени, поэтому старые экзоферы, как правило, имеют более нерегулярную форму.
ПРИМЕЧАНИЕ: Зрелые или старые экзоферы отличаются от рассеянной стадии «звездной ночи» более яркой интенсивностью флуоресценции экзоферов и их сферической формой. - Исследуйте фенотип «звездной ночи» как свидетельство более раннего экзофергенеза. Экзоферы прогресса в "звезднойночи" этапе, как экзофер распадается на меньшие пузырьки и окружающие гиподермиса попытки ухудшить содержание экзофера (Рисунок 1G, 2B, 3B и 7A).
ПРИМЕЧАНИЕ: Звездная ночная сцена отмечена фрагментированными и рассеянными (иногда сетевыми) флуоресцентными образованиями, которые потеряли структурную целостность и отображают тусклую флуоресценцию по сравнению с сенсорными нейронами и структурами экзоферов. - Ищите экземпляры "несколько событий экзофера". Экзоферы обычно производятся как особое явление (1 экзофер, исходящий из 1 сомы), но при некоторых обстоятельствах более одного экзофера может быть освобожден от одной сомы (Рисунок 7D).
ПРИМЕЧАНИЕ: Зрелые экзоферы могут деградировать в несколько пузырьков, поскольку они деградируют в гиподермии. Различие в том, был ли каждый экзофер создан независимым событием экзофергенеза, или же один исходный экзофер раскололся, чтобы создать дополнительную пузырьку, можно определить только путем наблюдения замедленного действия. - Имейте в виду, что не все морфологические аномалии созревают в экзоферы.
- Не забивай разтянутую сому в качестве экзофера. Расширенная или заостренная сома может наблюдаться иногда (особенно с возрастом или в состоянии стресса), но расширение без четкого места сужения не засовывано в качестве экзофера.
- Отклонить небольшие решенные почки, которые не достигают1/5th размером сомы в количестве событий экзофера.
- Не считайте нейритные нарои экзоферами. Зрелые невриты могут резко расширяться с возрастом (обычно в противоположном направлении нейронального процесса) и флуоресцентный белок может мигрировать в дистальный конецтаких структур 19.
ПРИМЕЧАНИЕ: Эти neurite нарои не экзоферы, поскольку они имеют различные модели развития в течение нескольких дней и недель, не образуют почки, и не отсоединяются(рисунок 7E).
- Определите флуоресцентные сущности, которые не являются экзоферами.
ПРИМЕЧАНИЕ: Важно, чтобы получить представление о фоновой флуоресценции для обеспечения правильной идентификации экструдированного флуоресцентного лица против аутофторесценции.-
Трансгенное флуоресцентное выражение против аутофторесценции. Не ошибитесь в аутофторесценции для трансгенного выражения. Истинный сигнал экзоферона не будет в кишечнике или кишечнике (подтверждение DIC может быть использовано для идентификации этих тканей), и сигнал экзофора будет значительно ярче, чем фоновое аутофторесценция.
ПРИМЕЧАНИЕ: Аутофторесценция вызвана кишечной гранулы кишечной флуоресцентной пигментации и накапливается с возрастом. Это неоднородно, особенно если рассматривать с различными длинами волн. - Сигнал от эмбрионов. Не прими сигнал эмбриона для экзофергенеза. Подтвердите подозрения на сигнал эмбриона, переключившись с флуоресценции на яркое освещение и проверяя на наличие ассоциаций сигнала с яйцеклетками в матке.
-
Из самолета или близлежащих тел сомы. Избегайте принятия из самолета сомы для экзофера, идентифицяя все близлежащие тела сомы, даже вне фокуса сомы в начале наблюдения.
ПРИМЕЧАНИЕ: Если забил для экзоферов из ALMR, определить и учитывать местоположение AVM и ALMR сомы. Более подробная информация об идентификации тела сомы описана на рисунке 3A.
-
Трансгенное флуоресцентное выражение против аутофторесценции. Не ошибитесь в аутофторесценции для трансгенного выражения. Истинный сигнал экзоферона не будет в кишечнике или кишечнике (подтверждение DIC может быть использовано для идентификации этих тканей), и сигнал экзофора будет значительно ярче, чем фоновое аутофторесценция.
8. Скоринг и статистика
- Оценка экзоферов как двоичный (да, есть экзофер / нет, нет экзофера).
- Рассмотрите обнаружение экзоферов как «экзофер-событие» для данного нейрона. Событие экзофера может представлять собой наблюдение одного экзофера вблизи сомы или нескольких экзоферов.
ПРИМЕЧАНИЕ: Для количественной оценки числа отдельных событий экзофергенеза используются наблюдения замедленного действия. - Подсчитайте события экзофера на ту или иную идентифицированную ячейку, поскольку различные клетки не производят экзоферы с одинаковой скоростью (см., например, рисунок 3C). Нейроны ALMR производят наиболее базовые экзоферы в штаммах, описанных в настоящем, и поэтому часто это клетка, выбранная для количественной оценки экзофера из нейронов сенсорных рецепторов.
- Для статистики, в целом, провести по крайней мере 3 биологических испытаний, по крайней мере 30 животных забил за испытание с соответствующим количеством наблюдений, необходимых для анализа нарушения.
- Для нескольких испытаний с участием одного или двух мутантов / лечения по сравнению с контролем, тест Кокран-Мантель-Генцель подходит для определения значений р.
- Для испытаний с участием более двух мутантов лечения по сравнению с контролем, также целесообразно использовать двоичный анализ логистической регрессии для оценки значимости для любого числа категорических предикторов.
Результаты
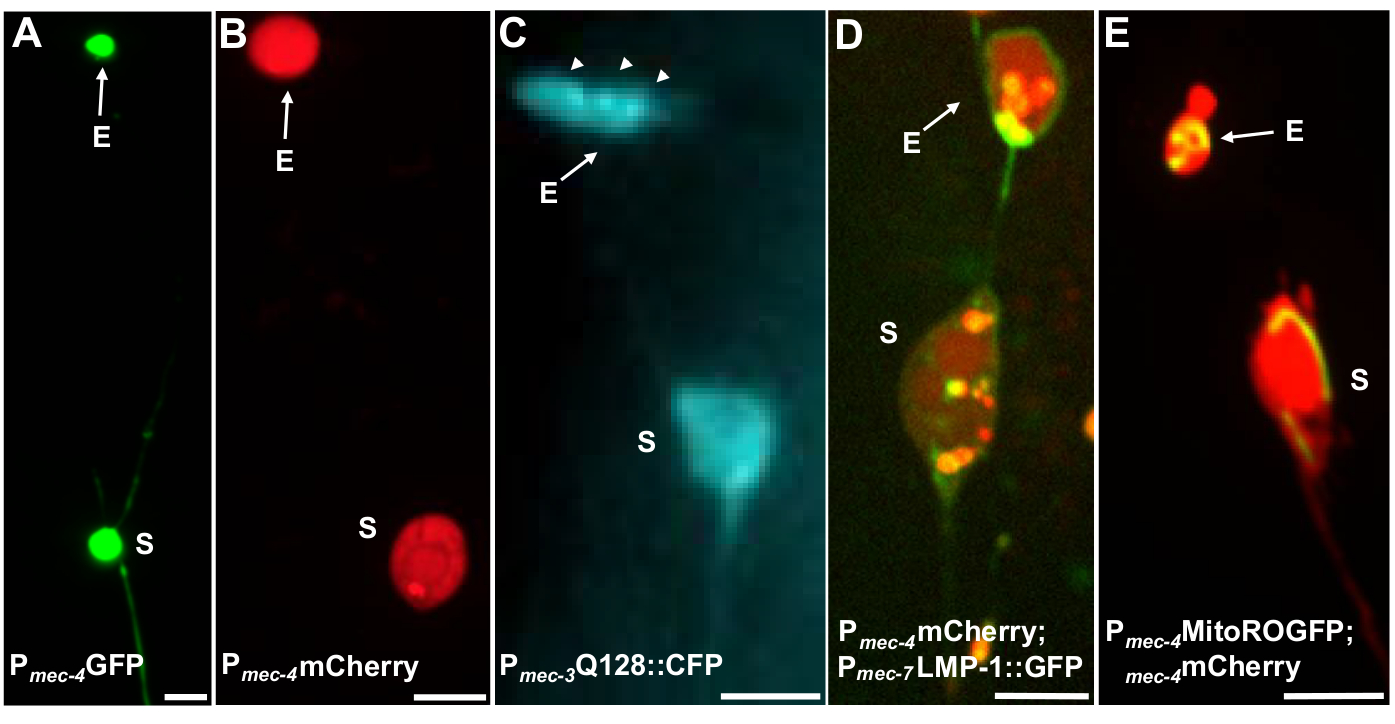
Несколько флуоресцентных репортеров могут быть использованы для измерения экзоферов. Сенсорные нейронные экзоферы легко визуализированы in vivo с помощью флуоресцентных меток белков, которые могут быть выбраны для экструзии, путем маркировки органелл, которые могут быть экструдированы, или путем пометки клеточных мембран. Таблица 1 определяет сенсорный нейрон, выраженный флуоресцентными репортерами, которые были использованы для мониторинга экзоферов, с репрезентативными примерами, включенными в рисунок 4. Грузы, которые, как известно, экструдированы в экзоферах включают слияние N-терминальной области человеческого ханттина для расширения полиглутамина (No128) (Рисунок 4B), лизосомы, которые GFP-тегами с лизосомальным ассоциированным мембранным белком (LMP-1) (Рисунок 4C), и митохондриями, помеченными матрицей-локализованной GFP (Рисунок 4D). Цитоплазмический GFP не сильно исключен и преимущественно сохраняется всоме 5, хотя GFP может слабо визуализировать экзоферы(рисунок 4A). Когда GFP сливается с белками, которые изгоняются, этот тег может быть использован для визуализации экзоферов. Важным моментом является то, что, помечая различные белки, большой спектр вопросов о высылке конкретных грузов и органелл, а также на белки и мембраны, которые составляют экзоферы, могут быть решены.
Псевдо-стереомикроскоп установки является эффективным инструментом для просмотра экзоферов у животных на агар пластин. Эта установка является гибридом соединения и стереоскопической технологии, которая включает в себя высокое числовое диафрагмы оптики на каждом увеличении, псевдо-стерео технологии (дискретные цели над стереоскопической базы), и зум операционного переключателя для просмотра на увеличения промежуточных установленных целей. Микроскоп, как это должно быть оснащено 10x окуляры и цели достаточно мощным для наблюдения нейрональной морфологии и экзофер производства для высокой пропускной способности скоринга (2x цель, используемая для сканирования / выбора, 10x цель, используемая для выявления и скоринга).
В то время как возможности увеличения стандартных стереомикроскопов обычно имеют достаточно высокое разрешение, чтобы увидеть сеть сенсорных нейронов, выражаюющих флуоресцентные белки, стандартных рассеченных микроскопов недостаточно для наблюдения за подклеточными деталями экзоферов, такими как трубчатые соединения сомы с экзофером. Такие наблюдения требуют конфокальные микроскопии (см. таблицу материалов для деталей оборудования).
Исследования количественной оценки экзофера требуют строгого контроля для устранения экспериментальных стрессов. Для воспроизводимого производства экзоферов требуется внимательное поддержание последовательных условий роста. В частности, производство экзоферов реагирует на стресс, что постоянное кормление, постоянная температура и рост без загрязнения на протяжении поколений имеют решающее значение для воспроизводимости. В базальных условиях роста с высокой нейронной экспрессией mCherry, производство экзофера является относительно низким (5-25% ALMRs производят экзоферы), но некоторые стрессы, в том числе осмотического и окислительного стресса, может увеличить скорость экзофера. В то время как выражение mCherry можно рассматривать как стресс, следствием стрессочувствительность уровней экзофера является то, что, при правильном контроле, экспериментальное введение стресса может быть стратегией для более легкого индуцирования и наблюдения экзоферогенеза.
Сроки и ожидаемые уровни производства экзоферов. Экзоферы практически отсутствуют при разработке личинок. Период пикового производства экзофера в молодой взрослой жизни, как представляется, весьма ограничено во время взрослых дней 1-4, чаще всего проявляется во взрослом день 2 или 3. Поскольку пик может немного сдвинуться вперед или назад, наиболее полная оценка профиля производства экзоферов заключается в том, чтобы забивать несколько испытаний ежедневно в течение взрослых дней 1-4. В целом, ALMR производит один крупный экзофер, с пузырьком сохраняется, по крайней мере 24 часов. Экзофер может быть произведен довольно быстро (в порядке минут в кратчайшиесроки). Чаще всего, только один крупный экзофер производится на нейрон в ранней взрослой жизни, но производство нескольких экзоферов возможно.
В целом производство экзоферов АЛМР, выражающих mCherry в базальных условиях, колеблется от 5-25% АЛМР, исследованных в оптимальные сроки взрослого дня 2-3(рисунок 3D). Протеостаз кризисов5, а также воздействие других стрессов может модулировать уровень экзофера. Стресс или генетические возмущения могут увеличить производство экзоферов до уровня обнаружения до 90% нейронов ALMR, производящих экструзии экзоферов.
РНК на основе кормления для тестирования ролей конкретных генов в экзофергенезе. Нематод C. elegans обычно подвергается РНК сбить путем кормления животных преобразованы штамм кишечной палочки HT115, которые выражают двойной мель РНК (dsRNA) ориентации генаинтереса 20. HT115 бактерии могут быть использованы при забил для экзоферов в кормлении РНК5. Хотя стенограммы в большинстве тканей могут быть направлены на РНК с помощью этой техники, нейроны являются более огнеупорным. Чувствительность к РНК может быть откалибрована с помощью животных, которые выражают трансгенный транспортер dsRNA SID-1 под нейрон-специфическим промоутером. Таким образом, нейрональной ткани могут быть сенсибилизированы до РНК21.
Ткань конкретных нокдаун гена интереса может быть достигнуто путем выражения компонента эндогенного метаболизма РНК в мутант, который не хватает в этом компоненте. Например: белок Argonaute RDE-1 может быть выражен конкретно в нейронах животных-мутантов rde-1 для достижения нокдауна гена, представляющих интерес только в нейронах, когда животные подвергаются вмешательству РНК, направленному против этого гена.
Использование стандартных нематод РНКпротоколов 20,22, воздействие родителей на этапе L4 в РНК и позволяет их потомство развивать потребление преобразованных бактерий HT115 до взрослой жизни генерирует сильный генетический нокдаун, но быть внимательным к потенциальным задержкам развития, вызванных РНК, как экспериментальные животные могут расти иначе, чем пустой вектор контроля. Важно всегда включать в себя пустой векторный контроль для отрицательного сравнения контроля. HT115 бактерии могут быть использованы при забил для экзоферов в кормлении РНК. Однако, обратите внимание, что некоторые гены эффективны при изменении показателей экзофергенеза даже в более короткие периоды воздействия РНК5. Если таргетинг на определенные гены приводит к нарушению развития, избегая подвергать животных пожизненному нокдауну, животных можно просто подбирать на этапе L4 на пластинах РНК для воздействия от L4 до взрослых D2 или D3.
| Процедить имя | Генотип | Описание | Процент экзофера | Ссылки |
| SK4005 | zdIs5(Pmec-4GFP) | Цитозолическое выражение GFP в сенсорных нейронах. | 1-8% ALM | Рисунок 4A, Мелентиевич 2017 |
| ЗБ4065 | bzIs166-Pмек-4::mCherry | Переэкспрессия mCherry (bzIs166) в сенсорных нейронах, производит как цитозолический сигнал, так и агрегаты mCherry. bzIs166 является индуктором экзофера. агрегаты mCherry являются предикторами экзофергенеза и преимущественно экструдируются в экзоферах. | 3-20% ALM (нормальные условия). 20-80% ALM (условия поста). | Рисунок 4B, Мелентиевич 2017 |
| ЗБ4067 | bzIs167-Pmec-4mitogfp Pmec-4mCherry4; igIs1'Pmec-7YFP Pmec-3htt57-128::cfp lin-15;; | YFP цитосолически маркирует сенсорные нейроны MEC-7. Совместно выраженный No128::CFP агрегирует и индуцирует экзоферы. CFP преимущественно замалчивается. | 25% | Рисунок 4C, Мелетиевич 2017 |
| ЗБ4509 | bzIs166-Pmec-4mCherry; bzIs168-Pmec-7LMP-1::GFP | bzIs168 LMP-1::GFP маркирует плазменные мембраны и лизосомальные мембраны. bzIs168 может быть использован для идентификации нейронных мембран, экзоферов (так как они связаны мембраной) и лизосомальных мембранных структур. | 3-20% ALM | Рисунок 4D, Мелентиевич 2017 |
| ЗБ4528 | bzIs166-Pmec-4mCherry; zhsEx17 (Pmec-4mitoLS::ROGFP) | Allele zhsEx17 является митохондриализированным репортером, который изменяет свою пиковую длину волны возбуждения с 405 нм (окисленный) до 476 нм (уменьшенный) в зависимости от местной окислительной среды. Он выражается в сенсорных нейронах и может быть использован сам по себе для идентификации митохондрий в сенсорных нейронах и в мито-экзоферах. | 3-20% протео-экзофер ALM. % ALM мито-экзофер количественной оценки в прогрессе. | Рисунок 4E, Мелентиевич 2017, Кэннон 2008, Ghose 2013 |
Таблица 1. Штаммы, которые были использованы для визуализации сенсорных нейронов, сенсорных нейронов-экзоферов и содержимого экзофера.

Рисунок 1: Этапы экзофергенеза. Процесс изготовления и выброса экзофера называется «экзофер-генезис». Динамический процесс формирования экзофера может занять от нескольких минут до нескольких часов. Изображены примеры морфологии сомы и экзофера на определенных этапах в процессе динамического экзофергенеза в штамме с высоким экзофером, производящем штамм, «B4065 bzIs166»Pmec-4mCherry». Все изображения дня 2 взрослых нейронов ALM приняты с целью 100x. (A)Нормальная сома. Взрослый механосенсорный сенсорный нейрон ALM трансгенно выражает Pmec-4mCherry. Морфология сомы изображена типична для молодых взрослых нейронов в этом штамме, с концентрациями mCherry в цитоплазме. (B)Ранняя фаза бутона. Первый наблюдаемый этап экзофергенеза предполагает поляризацию выбранного цитоплазмического материала к краю мембраны сомы. Этот шаг часто сопровождается расширением или отеком сомы. В случае сенсорных нейронов, до экзофер домена (PED) распространяется на окружающий гиподермис (не видно здесь). Обратите внимание на большую концентрацию материала mCherry в области раннего бутона. (C)Поздняя фаза бутона. После дальнейшей клеточной поляризации и расширения домена до экзофера становится очевидным сужение между сомой и экзофером (стрелкой). Это событие сигнализирует о переходе к поздней фазе бутона. Хотя на поздней стадии бутон клетки экспонатов четкое место сужения и отдельных сома и экзофер доменов, он еще не ущипнул полностью от сомы; начинающий экзофер может быть прикреплен толстым стеблем (стрелка). Начинающий домен считается ранним экзофером, когда диаметр домена экзофера, о котором идет речь, примерно на 1/3 больше диаметра строительной площадки/стебля. (D)Фаза раннего экзофера. Ранние экзоферы могут быть прикреплены стеблем от уходящей сомы – диаметр этого соединения может истончаться по мере того, как экзофер отходит от сомы. Цитоплазмический материал может быть передан из сомы в экзофер через эту трубку, хотя большая часть материала загружается в процессе создания. Экзоферы могут отделяться от сомы, как поизображено в( E ), разделенные экзоферы считаются зрелыми экзоферами (F). Зрелый экзофер может проходить через окружающие гиподермальные ткани, отходя от уходящей сомы. (G)Разбивка мчерри-маркированных экзоферов на более мелкие пузырьки в гиподермисах приводит к рассеянному пунктуату появления материала mCherry, скорее всего, когда он входит в гиподермальной эндолисосомной сети. Рассеянный пунктуатный сигнал называется фазой «звездной ночи». Деградация некоторых содержание экзоферов, вероятно, достигается гиподермальной лизосомой, но некоторые материалы не полностью деградировали и часто повторно экструдируются гиподермисом в псевдоколом. Пост-экзофергенез mCherry транзита описывается более подробно на рисунке 2. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 2: mCherry экструдированы из сенсорных нейронов в экзоферов занимается окружающих гиподермальной лизосомальной сети, но позже может быть экструдирован в псевдокоелом, где coelomocytes может хранить / ухудшать mCherry. (A) Мультфильм резюме о том, как mCherry экструдированы в экзоферы транзитов тела после изгнания нейронами. Во время экзоферогенеза выбранное клеточное содержимое, такое как mCherry, становится локализованным и отключается от отправки нейрональной сомы в независимой везикуле, окруженной нейрональными и гиподермальными плазменными мембранами. Так как сенсорные нейроны встроены в гиподермальной ткани, как экзофер домена почки наружу он движется дальше в гиподермис. Экзофер может транзит гиподермиса, и через несколько часов до нескольких дней, содержимое экзофора может фрагментировать в эндолисосомной сети гиподермиса. MCherry может появиться как рассеянный puncta всей hypodermis, этап называется "звездная ночь". Через несколько дней, некоторые из mCherry может выйти из гиподермиса в окружающих псевдоколом, где клетки мусорщика называется coelomocytes может получить доступ к, и занять, mCherry, которые могут быть сохранены. (B)Пример появления звездных ночных пузырьков mCherry. Изображение ALM сомы помечены mCherry с большими фрагментами экзофера и звездные ночные пузырьки. Штамм : «B4065 bzIs166»Pmec-4mCherry». (C)Пример концентрации мчерри в далеких келомоцитах. Sideview взрослого животного день 10 штамма »B4065 bzIs166»Pmec-4mCherry» показаны mCherry сосредоточены в coelomocytes (стрелки). Некоторые звездные ночные пузырьки также очевидны. В целом концентрация коломоцитов становится очевидной примерно после взрослого дня 5 жизни. (Bдно) Мультфильм воспроизведения (B), с сенсорными нейронами и процессами, изложенными в красном, как и яркие фрагменты экзофера; рассеянные маленькие пузырьки различных глубин показаны в светло-розовом цвете. (C дно) Мультфильм версия изображения ( C ),показываянейронный процесс в красном, звездная ночь в розовом и coelomocytes в зеленом цвете. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 3: Механосенсорные сенсорные нейроны производят экзоферы на разных уровнях с точным временным профилем. (A) (Вверху) Мультфильм изображение механосенсорных сенсорных нейронов в пространственном отношении к ключевым анатомическим ориентирам C. elegans включая насосную глотку и нейронно-плотное нервное кольцо во главе животного, вульву в середине тела и конические хвост. (Внизу) Нет, нет, нет. Флуоресцентно помечены сенсорные нейроны, выражаюющие GFP, как рассматривать с верхней и левой стороны (изображения, адаптированные из WormAtlas). Красная коробка изображает область, где обычно расположены экзоферы ALM. (B)Высокое увеличение зрения средней области тела, в которой ALM полученных экзоферов производятся в штамм, выражаюющий "Pmec-4mCherry". AVM и ALMR нейрон изображены, и показано, является экзофер ALMR вместе с mCherry звездной ночи. Нейроны ALMR наиболее легко производят экзоферы. (C) ALMR механосенсорные сенсорные нейроны с большей готовностью производят экзоферы по сравнению с другими сенсорными нейронами в гермафродитах в базальных условиях. Механосенсорное прикосновение нейрона экзофер производства на взрослый день 2, как забил для отдельных нейронов сенсорных рецепторов указывается. Штамм: «B4065 bzIs166»Pmec-4mCherry», N'gt;150, бары ошибок SEM.(D) сенсорныенейроны ALMR производят больше экзоферов в течение дней 2 и 3 взрослой жизни по сравнению с подростковой стадией L4 или с животными в преклонных возрастах. Штамм: «B4065 bzIs166»Pmec-4mCherry», N'gt;150, бары ошибок SEM. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 4: Примеры некоторых флуоресцентных репортеров, которые помечает содержимое экзофера. Простой способ наблюдать экзоферы путем создания трансгенных животных, которые выражают флюорофоры от нейрональных промоутеров. Фторфоры позволяют визуализировать экзофер и трансгенное выражение вызывает агрегацию и/или протеостресс, что увеличивает экзофергенез. Экзоферы, производимые амфидными нейронами, также могут наблюдаться в родных условиях, используя пломбы красителя для визуализации. Показаны примеры общих штаммов, которые могут быть использованы для наблюдения экзоферов, (E) экзофер, (S) сома. (A) Сома и экзофер из ALM взрослого штамма SK4005 zdIs5«Pmec-4GFP»,100x цель, используемая для фотографии, масштаб бар 3 м. В этом штамме измеряются экзоферы, включаемые растворимые GFP, но производство экзоферов происходит нечасто. Сплавливание GFP на белки, которые могут быть преимущественно экструдированы в экзоферах в других исследованиях, подтверждает, что синтезы GFP могут быть обнаружены в зрелых экзоферах. (B) ALM сома и экзофер взрослого штамма »B4065 bzIs166»Pmec-4mCherry», который выражает mCherry и вызывает производство сенсорного нейрона экзофера. 100x цель, используемая для фотографии, масштаб бар 5 мкм. (C) ALM сома и экзофер взрослого штамма «B4067 bzIs167»Pmec-4mitogfp Pmec-4mCherry4»; igIs1(Pmec-7YFP Pmec-3htt57-128::cfp lin-15); селективный синий канал, используемый для изображения htt57-128::CFP. Экзофер содержит агрегаты htt57-128::CFP (стрелки), которые кажутся более концентрированными в экзофере, чем в соме. 40x цель, используемая для фотографии, масштаб бар 5 м. (D-E) Экзоферы могут содержать органеллы и органеллы-специфические пометки флуоресцентными белками, что позволяет контролировать экструзию органелл. (D) Лизосомальная мембрана тег LMP-1::GFP излагается сома и экзофер мембраны и теги плазменных мембран слабо (плазменная мембрана локализации является шагом торговли на пути к лизосомальной ориентации) и этикетки лизосомальных органелл сильно. Показан взрослый ALM сома совместно выражая Pmec-4mCherry и PMEC-7LMP-1::GFP, который локализует мембраны и лизосомы. Сома имеет прикрепленный экзофер с другими меньшими экструзиями, которые могут быть фрагментами экзофера (стрелками). Положительные структуры GFP включены в сому и присутствуют в большомэкзофере, штамме: «B4509 bzIs166»Pmec-4mCherry;bzIs168«Pmec-7LMP-1::GFP». 100x цель, используемая для фотографии, масштаб бар 5 мкм. E)Митохондриальный маркер GFP может быть использован для идентификации митохондрий в соме и экзоферах. Показано взрослое ALM сома, выражающ Pmec-4mCherry и mito::ROGFP, который локализуется в митохондриальной матрице. мито::ROGFP выражается в одиночку, без mCherry, также может быть легко использован для выявления нейронов и оценка для экзоферов, которые включают митохондрии. Штамм: «B4528 bzIs166»Pmec-4mCherry»; zhsEx17 (Pmec-4mitoLS::ROGFP). 100x цель, используемая для фотографии; шкала бар 5 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 5: Цикл развития C. elegans и идентификации L4. (A) При 20 градусов по Цельсию яйцо занимает около 9 часов, чтобы люк один раз положил мать. (B)Недавно вылупившихся животных находится в личинок стадии 1 (L1) и линьки в личинку L2 после 12 часов. (C) животные остаются в L2 и ( D )L3личиночинок этапов около 8 часов каждый. (E) Подростковые животные считаются четвертой личиночковой стадии (L4) и отмечены заметным развивающихся вульвы, которая появляется как белый полумесяц вблизи среднего тела. Наличие этого в то время как полумесяц позволяет легко идентификации и сбора L4 поставил животных для создания синхронизированных культур, которые позже облегчить забил для экзоферов. Звери остаются в стадии L4 в течение примерно 10 часов, прежде чем их окончательный линьки в gravid взрослых, F) определены путем разработки яйцеклеток, видимых сперматозоидов, и начало яйцекладки. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 6: Подготовка микроскопа слайд агар площадку. (A)Подготовь два слайда с одной полосой лабораторной ленты, расположенной вдоль верхней части. Поместите не-записанный слайд микроскопа между ними, как по фото. B) Поместите каплю расплавленной агарозы на вершине слайда. (C) Поместите чистый слайд мягко на верхней части капли, нажав агарозы в спущенный круг площадку. (D) Удалите записанные на пленку слайды, которые действуют для достижения равномерного уплощения агара, который необходим для создания ровной колодки. (E)Удалите верхнюю горку после того, как агарозная колодка высохнет. (F)Pipette паралитический раствор (левамизол или теттрамизол) на верхней части агар площадку. (G)Выберите надлежащим образом поставил животных в паралитической. (H)Аккуратно накройте животных крышкой и убедитесь, что животные живы. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 7: Символы экзоферов и критерии идентификации экзоферов. (A)Общие критерии, которые определяют экзофер. (B)Диаметр сравнения между отправкой сомы и экструдированного экзофера, измеряется в мкм. Взрослые ALM сомы, N-35, штамм: «B4065 bzIs166»Pmec-4mCherry» - 6,53 мкм средний размер сомы и 3,83 мкм средний размер экзофера. (C)Определение критериев дифференциации между областью экзофера и начинающим экзофером. (D) Чаще всего, отдельные нейроны делают один большой экзофер, который позже распадается или фрагменты, как гиподермис пытается ухудшить его содержание. Тем не менее, несколько экзоферов могут наблюдаться рядом с одним сенсорным нейроном, которые могут вытекать из нескольких событий экзофера от одного нейрона или в качестве альтернативы, экзоферы могут также бутон или фрагмент себя. Происхождение нескольких экзофер-похожих сущностей может быть определено только с помощью микроскопии промежуток времени. Сверху изображена сенсорная нейронная сома ALMR с одним далеким экзофером. Внизу изображена сенсорная нейронная сома ALMR с несколькими экструзиями, похожими на экзоферы. (E)Общие морфологические особенности во взрослых ALM сенсорных нейронов сома, которые могут быть ошибочно приняты за экзофер событий. Вверху слева - растянутая ALM сома, без четкого домена экзофера или сужения сайта. Верхняя середина - Нейроны могут иметь небольшие внеклеточные выступы, которые могут быть аналогичны экзоферам, но не отвечают критериям требования размера, которые должны рассматриваться как экзофер. Верхний правый - С возрастом, сенсорные нейроны могут развиваться нарои вдоль их незначительные нейрита. Часто материал mCherry можно собрать на кончике нарости neurite. Это не забил в качестве экзофера, если собранный mCherry не отвечает требованиям размера экзофера до сомы. Внизу изображены взрослые нейроны ALM, которые имеют определяющие критерии для домена экзофера или экзофера. Botom left - ALM soma, которая имеет известный домен экзофера, который выборочно включает цитозол mCherry и помеченные mCherry агрегаты. Сайт сужения домена экзофера отмечен стрелками и соответствует критериям размера (по крайней мере1/5 th размером сомы). Самый большой диаметр домена экзофера почти на 1/3 больше диаметра участка сужения, что соответствует критериям события экзофера. Нижняя середина - ALM сома, которая имеет видный начинающий экзофер, который отвечает критериям размера. Существует четкий сайт сужения. Внизу справа - ALM сома, которая имеет прикрепленный mCherry заполненный экзофер, который отвечает требованиям размера экзофера. Экзофер крепится тонкой соединительной нитью. Все изображения из штамма «B4065 bzIs166»(Pmec-4mCherry). Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}
Обсуждение
Характеристика молекулярных механизмов in vivo агрегата и ликвидации органеллы в виде крупных экзоферов находится в зачаточном состоянии. Вопросы о назначении грузов для высылки, поляризованном сборе этих грузов внутри ячейки, регулировании решения о генерации экзоферов, механизме, который является посредником в экструзиях, и взаимодействии экзоферов с деградативным механизмом в соседней камере - все это еще не решено. Кроме того, визуализация виво трубчатых соединений, которые могут передавать биологические материалы, которые включают кальций, агрегаты и митохондрии, интересна и недостаточно изучена биологией в своем собственном праве. Вопросы о том, почему некоторые клетки более склонны к выработке экзоферов, чем другие, также остаются нерешенными, но могут начать генетически рассекаться с помощью подходов, изложенных в этом протоколе.
Подробно описаны в этом протоколе подходы к достижению воспроизводимых скоринга производства экзоферов, с вниманием к различему экзоферов от близлежащих сом клеток, сроки проведения анализов для захвата пика производства экзофера, а также строгий контроль условий роста для устранения непреднамеренных напряжений, которые могут модулировать уровни экзофера. Оба различия большого раннего экзофера, или "звездная ночь" дисперсии в окружающем гиподермис может быть количественно в качестве доказательства производства экзофера. При этом нейроны, выражаюющие mCherry в базальных условиях, чаще всего ассоциируются с 5-25% нейронов определенного типа, производящих экзофер. Контролируемое введение стрессовых условий может быть применено для увеличения производства экзоферов для обнаружения до 90% нейронов, производящих экструзии, особенно полезные для генетических или фармакологических экранов для модификаторов.
При нейродегенеративных заболеваниях человека большие агрегаты могут передаваться из больных нейронов в соседние клетки для содействия распространению патологии. Механизм экзоферона может происходить через сохраненный механизм, используемый для агрегирования экструзии через филу. Определение молекул in vivo, которые либо повышают эффективность этого процесса (считается более эффективным контролем протеостаза), либо блокируют его можно использовать для влияния на разработку новых стратегий борьбы с многочисленными нейродегенеративными заболеваниями. Таким образом, описанный здесь протокол может быть использован для классических генетических экранов мутагенеза, геномных РНК-экранов, которые систематически сбивают гены для выявления усилителей и супрессоров, или для исследований лекарственного вмешательства, которые определяют кандидат фармакологические модификаторы этого процесса. Подход прост, хотя и несколько трудоемкий. Экзоферы настолько велики, что их можно рассматривать с помощью микроскопа с высоким увеличением. Тем не менее, C. elegans нейроны относительно малы и, глядя на их органеллы или их мембраны требуют более высокой мощности конфокальных изображений и медленный процесс. Варианты более высокой пропускной способности могут включать подходы к визуализации с высоким содержанием в формате нескольких хорошо пластин.
Применение стандартизированного подхода к забил экзофер должен лежать в основе согласованного генетического вскрытия процесса, с помощью которого нейроны могут организовать и устранить клеточного мусора.
Раскрытие информации
Ни один
Благодарности
Мы признаем следующие гранты NIH: R01AG047101 и R37AG56510. Сотрудники лабораторий Driscoll и Grant внесли значительный вклад в разработку и тонкую настройку описанных протоколов, с помощью строгих экспериментов и сильной коммуникации.
Материалы
| Name | Company | Catalog Number | Comments |
| 95B Scientific CMOS camera | Photometrics Prime | ||
| 1,000 μL low retention tips | Sarstedt | ||
| 10 mL serological pipette | Appleton Woods | CC214 | |
| 10 μL low retention tips | Sarstedt | 70.1130.105 | |
| 13% sodium hypochlorite | Acros Organics | AC219255000 | |
| 15 mL centrifuge tubes | Fisher Scientific | 05-539-12 | |
| 2 L erlenmeyer flasks | Scientific Laboratory Supplies | FLA4036 | |
| 25 mL serological pipette | Appleton Woods | CC216 | |
| 300 μL low retention tips | Sarstedt | 70.765.105 | |
| 50 mL serological pipette | Appleton Woods | CC117 | |
| 5-Fluoro-2'-deoxyuridine 98% | Alfa Aesar | L16497.ME | |
| 9 cm sterile Petri dishes | Fisher Scientific | 11309283 | |
| absolute ethanol | Vwr | 20821.33 | |
| Agar | Sigma Aldrich | A1296 | |
| C. elegans strain wild type | Supplied by CGC | N2 | C. elegans strain |
| calcium chloride dihydrate | Sigma Aldrich | C3881 | |
| cholesterol | Acros | 110190250 | |
| dibasic sodium phosphate | Sigma Aldrich | S3264 | |
| E. coli strain OP50 | Supplied by CGC | Op50 | E coli strain |
| FBS10 Standard microscope | Meyer Instruments | KSC 410-1-100-1 | FBS10 Standard with Plate Base, 100/100 Trinocular Head and Flip zoom |
| glass pipette 270 mm | Fisherbrand | FB50255 | |
| Heraeus Multifuge X3R | Thermofisher scientific | 75004515 | |
| Inoculating Spreaders | Fisher Scientific | 11821741 | |
| LB medium capsules | MP biomedicals | 3002-031 | |
| LDI – Laser Diode Illuminator | 89 North | ||
| levamisole | Sigma Aldrich | 16595-80-5 | |
| M4 multipette | Eppendorf | 4982000012 | |
| magnesium sulphate | Sigma Aldrich | M7506 | |
| monobasic potassium phosphate | Sigma Aldrich | P0662 | |
| Multitron Standard shaking incubator | Infors HT | INFO28573 | |
| Nalgene 1 L Centrifuge pots | Fisher Scientific | 3120-1000 | |
| P10 pipette | Eppendorf Research Plus | 3123000020 | |
| P1000 pipette | Eppendorf Research Plus | ||
| P200 pipette | Eppendorf Research Plus | 3123000055 | |
| pipeteboy 2 | VWR | 612-0927 | |
| Polystyrene microbeads | Sigma Aldrich | MFCD00131491 | |
| RC5C plus floor mounted centrifuge | Sorvall | 9900884 | |
| Reusable ringed cytology slides | ThermoFisher Scientific | 22037242 | |
| SK4005 zdIs5[Pmec-4GFP] | contract Driscoll lab | GFP expressed in touch neurons | |
| sodium chloride | Sigma Aldrich | 13422 | |
| Sodium hydroxide | Fisher Chemical | S/4880/53 | |
| Tactrol 2 Autoclave | Priorclave | ||
| Triton-X | Thermofisher scientific | 28313 | |
| Tween 20 | Sigma Aldrich | 9005-64-5 | |
| X-Light V2 Spinning Disk Confocal Unit | CrestOptics | ||
| ZB4065 bzIs166[Pmec-4mCherry] | contract Driscoll lab | mCherry expressed in touch neurons | |
| ZB4067 bzIs167[Pmec-4mitogfp Pmec-4mCherry4]; igIs1[Pmec-7YFP Pmec-3htt57Q128::cfp lin-15+] | contract Driscoll lab | Q128 expressed in touch neurons | |
| ZB4509 bzIs166[Pmec-4mCherry]; bzIs168[Pmec-7LMP-1::GFP] | contract Driscoll lab | mitoROGFP expressed in touch neurons | |
| ZB4528 bzIs166[Pmec-4mCherry]; zhsEx17 [Pmec-4mitoLS::ROGFP] | contract Driscoll lab | autophagy marker expressed in touch neurons | |
| ZEISS Axio Vert.A1 | Zeiss |
Ссылки
- Davis, A. A., Leyns, C. E. G., Holtzman, D. M. Intercellular Spread of Protein Aggregates in Neurodegenerative Disease. Annual Review of Cell and Developmental Biology. 34, 545-568 (2018).
- Davis, C. H., et al. Transcellular degradation of axonal mitochondria. Proceedings of the National Academy of Sciences of the United States of America. 111 (26), 9633-9638 (2014).
- Torralba, D., Baixauli, F., Sanchez-Madrid, F. Mitochondria Know No Boundaries: Mechanisms and Functions of Intercellular Mitochondrial Transfer. Frontiers in Cell and Developmental Biology. 4, 107 (2016).
- Stahl, P. D., Raposo, G. Extracellular Vesicles: Exosomes and Microvesicles Integrators of Homeostasis. Physiology (Bethesda, Md.). 34 (3), 169-177 (2019).
- Melentijevic, I., et al. C-elegans neurons jettison protein aggregates and mitochondria under neurotoxic stress. Nature. 542 (7641), 367 (2017).
- Nussbaum-Krammer, C. I., Park, K. W., Li, L., Melki, R., Morimoto, R. I. Spreading of a prion domain from cell-to-cell by vesicular transport in Caenorhabditis elegans. PLoS Genetics. 9 (3), 1003351 (2013).
- Tyson, T., et al. Novel animal model defines genetic contributions for neuron-to-neuron transfer of alpha-synuclein. Scientific Reports. 7, (2017).
- Babcock, D. T., Ganetzky, B. Transcellular spreading of huntingtin aggregates in the Drosophila brain. Proceedings of the National Academy of Sciences of the United States of America. 112 (39), 5427-5433 (2015).
- Pearce, M. M. P., Spartz, E. J., Hong, W., Luo, L., Kopito, R. R. Prion-like transmission of neuronal huntingtin aggregates to phagocytic glia in the Drosophila brain. Nature Communications. 6, 6768 (2015).
- Fu, H., Li, J., Du, P., Jin, W., Cui, D. Metabolic wastes are extracellularly disposed by excretosomes, nanotubes and exophers in mouse HT22 cells through an autophagic vesicle clustering mechanism. bioRxiv. 10 (1), (2019).
- Ghose, P., Park, E. C., Tabakin, A., Salazar-Vasquez, N., Rongo, C. Anoxia-reoxygenation regulates mitochondrial dynamics through the hypoxia response pathway, SKN-1/Nrf, and stomatin-like protein STL-1/SLP-2. PLoS Genetics. 9 (12), 1004063 (2013).
- Cannon, M. B., Remington, S. J. Redox-sensitive green fluorescent protein: probes for dynamic intracellular redox responses. A review. Methods in Molecular Biology. 476, 51-65 (2008).
- Perkins, L. A., Hedgecock, E. M., Thomson, J. N., Culotti, J. G. Mutant sensory cilia in the nematode Caenorhabditis elegans. Developmental Biology. 117 (2), 456-487 (1986).
- Stiernagle, T. Maintenance of C. elegans. WormBook: The Online Review of C. Elegans Biology. , 1-11 (2006).
- Sutphin, G. L., Kaeberlein, M. Measuring Caenorhabditis elegans life span on solid media. Journal of Visualized Experiments. (27), (2009).
- Mitchell, D. H., Stiles, J. W., Santelli, J., Sanadi, D. R. Synchronous growth and aging of Caenorhabditis elegans in the presence of fluorodeoxyuridine. Journal of Gerontology. 34 (1), 28-36 (1979).
- Weicksel, S. E., et al. A novel small molecule that disrupts a key event during the oocyte-to-embryo transition in C. elegans. Development. 143 (19), 3540-3548 (2016).
- Dong, L., et al. Reversible and long-term immobilization in a hydrogel-microbead matrix for high-resolution imaging of Caenorhabditis elegans and other small organisms. PloS One. 13 (3), 0193989 (2018).
- Toth, M. L., et al. Neurite sprouting and synapse deterioration in the aging Caenorhabditis elegans nervous system. Journal of Neuroscience. 32 (26), 8778-8790 (2012).
- Conte, D., MacNeil, L. T., Walhout, A. J. M., Mello, C. C. RNA Interference in Caenorhabditis elegans. Current Protocols in Molecular Biology. 109, (2015).
- Calixto, A., Chelur, D., Topalidou, I., Chen, X., Chalfie, M. Enhanced neuronal RNAi in C. elegans using SID-1. Nature Methods. 7 (7), 554-559 (2010).
- Maher, K. N., Catanese, M., Chase, D. L. Large-scale gene knockdown in C. elegans using dsRNA feeding libraries to generate robust loss-of-function phenotypes. Journal of Visualized Experiments. (79), e50693 (2013).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеThis article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены