
需要订阅 JoVE 才能查看此. 登录或开始免费试用。
Method Article
在实验室条件下收集和长期维护切叶蚁(Atta)
摘要
在这里,描述了一种在实验室条件下成功收集和维持健康的 Atta (膜翅目:Formicidae)蚁群的方案。此外,还详细介绍了不同的巢穴类型和配置以及可能的实验程序。
摘要
蚂蚁是地球上生物多样性最丰富的动物群体之一,栖息在不同的环境中。在受控环境中维持蚁群可以丰富对其生物学的理解,从而有助于应用研究。这种做法通常用于造成经济损失的物种(如 阿塔 蚂蚁)的种群控制研究。为了培养它们的互利真菌,这些切叶蚁收集叶子,因此被认为是广泛分布在整个美洲大陆的农业害虫。它们是高度社会化的,栖息在由各种房间组成的精心制作的地下巢穴中。它们在受控环境中的维护取决于此处描述的几种程序和频繁护理的日常工作。它从在繁殖季节(即婚礼飞行)收集蜂王开始,然后将其单独转移到塑料容器中。由于蜂王的死亡率很高,第二次收集可以在婚礼飞行后约6个月进行,届时挖掘出具有发达真菌团的早期巢穴,手工采摘并放置在塑料容器中。在实验室中,每天将叶子提供给已建立的菌落,每周将蚂蚁产生的废物与剩余的干燥植物材料一起清除。随着真菌园的不断生长,菌落根据实验目的转移到不同类型的容器中。切叶蚁群被放置在相互连接的容器中,代表组织系统,其功能室由自然界中的这些昆虫建造。此设置非常适合监控废物量、真菌花园健康以及工人和蜂王的行为等因素。促进数据收集和更详细的观察被认为是将蚁群保持在受控条件下的最大优势。
引言
蚂蚁由对大多数陆地环境产生影响的多样化个体组成1.它们充当高效的分散者2,3,4,捕食者5和生态系统工程师6,7,8,9,10,突出了它们对自然生态系统的重要性和生态成功。所有蚂蚁物种都被归类为真社会昆虫;然而,它们的社会组织在不同物种群体之间差异很大,即分工系统、功能群体、个体之间的交流、牧草组织、菌落基础和繁殖过程11。作为一个高度多元化的群体,他们求助于多种食物资源和专门的喂养行为。事实上,农业不仅是人类文明的一大步,也是蚂蚁物种的一大步。大约 55 至 65 Ma 前12,蚂蚁开始培养真菌并将它们纳入几乎排他性的饮食中。他们变得如此专业化,以至于他们发展出严格、依赖和强制性的互动,被归类为共生,其中一个个体没有另一个个体就无法生存。
低等真菌生长的蚂蚁收集和处理死亡的有机物,例如腐烂的叶子碎片,以培养它们的互利真菌;而高等真菌生长的蚂蚁收获新鲜的植物材料,构成了最成功的共生自然系统之一13。这种高度专业化的农业技术使他们能够抓住一个新的利基市场。高等蚂蚁包括切叶蚁,这是一个单系群,在19Ma(15-24Ma)和18Ma(14-22Ma)之间引起14,15,16由四个有效属组成:Atta Fabricius,Acromyrmex Mayr,Amoimyrmex Cristiano和Pseudoatta Gallardo。由切叶蚁执行的切叶农业系统是从衍生的农业系统演变而来的17.这些物种中的大多数专门利用互惠真菌物种Leucoagaricus gongylophorus Singer 18(也称为Leucocoprinus gongylophorus Heim 19),标志着一个显着的进化转变11。真菌品种垂直传播,从原始巢到后代,表明它们是克隆繁殖的20。
值得注意的是, 阿塔 社会发展了一个复杂的组织结构,其环境非常重要,并且引起了myrmecologists的极大兴趣。他们的人口可以由数百万人组成,其中大多数是不育的女工,表现出明显的多态性,即不同的大小和解剖形态。种群根据年龄、生理状态、形态类型、行为和殖民地的专门活动按种姓区分21.工人可能被区分为园丁和护士、巢内通才、觅食者和挖掘者以及捍卫者或士兵21.这种组织允许合作执行任务,以及可以产生高度结构化的集体行为的自组织系统,使他们能够有效地对环境干扰做出反应22.
人口更新的作用由单个女王(即一夫一妻制)扮演,只要她活着,就构成了永久的生殖种姓22.众所周知,阿塔女王的寿命超过20年,在其一生中产卵23。由于女王是不可替代的,它的耐力对于殖民地13,20,23,24的生存至关重要。然而,在繁殖季节,巢中可以找到数千只有翅膀的繁殖雌性和雄性,但没有一只留在原来的巢中,形成了一个临时种姓22。在Atta sexdens殖民地,产生了近3,000只生殖雌性和14,000只生殖雄性25。它发生在一个菌落达到性成熟时,大约在实施后38个月,并且每年重复一次,直到它被消灭23,25。新的阿塔殖民地是通过单倍体建立的,其中一个女王开始一个新的巢穴。
当环境条件有利时,繁殖者离开地下巢穴开始婚礼飞行。其发生时间因地区而异,根据物种的不同,整个巴西领土全年都在变化。然而,该事件之前似乎有降雨和湿度升高26,这可能与土壤湿度22导致的挖掘便利有关。通常,在婚前1-5周,巢穴入口和通道被拓宽,以方便生殖个体离开。在离开它们的母群之前,有翅膀的雌性在颊下腔中收集并储存一部分互助真菌20,27。在飞行途中进行多次交配,并计算出在某些物种中,一个女王可以被三到八只雄性(即一妻多夫制)授精28,确保遗传变异性29。之后,蜂王进入土壤,优先考虑没有或很少植被的地方25,在那里它们移除翅膀并挖掘它们的第一个巢室。这是唯一可以在巢穴外看到女王的时期。虽然在人工巢中看到了临时种姓的个体,但尚不清楚在实验室条件下是否进行了任何成功的交配(即婚礼飞行)24。
最初的筑巢对应于殖民地最关键的时期,可以持续6小时到8小时23,25。这时,女王在最初的房间里隐居,几天后,产卵开始了。第一批卵被喂给蜂王反刍的菌丝体,标志着殖民地真菌花园的开始。第一只幼虫大约在 25 天22 出现,几乎在第一个月末,菌落由一个增殖真菌垫组成,未成熟的真菌(卵、幼虫和蛹)嵌套在那里,以及女王,她孤立地抚养她最初的后代23。卵也是第一批幼虫的食物资源,被蜂王高度消耗13.此外,女王通过脂肪身体储备和分解代谢不再使用的翅膀肌肉来维持自己13.最初的真菌培养物不会被消耗,因为菌落的生存取决于其发育,在此期间,蜂王用粪液使其受精13。出巢几天后,第一批工人打开巢穴入口,开始在巢穴附近区域进行觅食活动13.他们将收集的材料作为真菌花园的基质,现在作为工人的食物13,22。在加入真菌培养物之前,将工人携带的植物材料切成小块并用粪便液体润湿13。蚂蚁操纵真菌接种物来增加和控制其生长,这将用于分隔大土壤挖掘室,专门用于调理花园13,22,25。
婚后约6个月, A. sexdens 巢包含一个真菌室和几个通道。切叶蚁巢的建设具有很大的专业化作用,可以作为抵御天敌和不利环境因素的防御机制22.众所周知,切叶蚁会在真菌花园中破碎,并在房间开始变干时将其转移到高湿度的房间中13。因此,尽管挖掘巢穴具有相当大的能源成本,但投入的能量在殖民地本身的利益中被逆转了22。除了少数例外, 阿塔 物种还为殖民地的废物制作专门的房间,主要由耗尽的真菌基质和死蚂蚁的尸体制成,将其与巢穴的其余部分隔离开来,并建立了重要的社会免疫策略30。此外,一组不同的工人直接操纵垃圾,以避免污染其他人。工人们不断觅食以培育真菌,这是菌落的主要营养资源。然而,它们也可以在切割碎片时以植物汁液为食。植物材料经过精心挑选用于真菌园维护,并受到许多因素的影响,例如叶片性状和生态系统的特性13。
切叶蚁获取新鲜材料的觅食策略非常复杂,再加上已建立的殖民地的高收获需求,给农业生产者造成了相当大的经济损失,并危及森林恢复区22,31。因此,这些蚂蚁在可能遇到的大多数地区都可以归类为害虫,范围从美国南部到阿根廷东北部11,13,22,32。由于这些昆虫生物学中固有的一系列适应性(即社会组织、觅食、真菌栽培、卫生和复杂的巢穴结构)33,因此消灭有问题的菌落具有挑战性33。因此,种群控制策略不同于通常应用于其他害虫的策略,主要诉诸有吸引力的受污染诱饵产品33,34。然而,由于这些蚂蚁可以拒绝对真菌和群体个体的有害物质,并损害耕地33,因此不断测试新的天然化合物和控制替代品33,35,36。由于很难在经过现场测试的菌落上监测实验结果,因此初步论文是在受控环境中进行的。
因此,考虑到蚂蚁的异质生活方式,支持物种水平的研究,并将群体视为操作单位,其中一只蚂蚁是复杂超有机体的元素,实验方案必须适应感兴趣的群体11。到目前为止收集的有关 Atta 属的报告使得在实验室条件下成功收集和维持菌落并承认其基本需求和一般功能成为可能。基于它们的繁殖、群体建立和摄食行为等自然过程,已经开发出一种常规的做法,允许在不同类型的巢穴中长期建立群体。在这里,描述了在实验室中维持切叶蚁的程序协议,并强调了可能的一般研究,具有不同的实验目的和科学推广。
研究方案
1. 皇后收藏
- 在文献中搜索感兴趣地区的阿塔繁殖季节。繁殖季节的发生、频率和日间时间因地区气候条件而异(表1)。虽然它通常在春季进行,但必须针对打算进行收集的位置收集此信息37,38,39,40,41,42,43,44,45,46。
- 确定并标记将 阿塔 巢视为收集蜂王和年轻殖民地的可能区域的位置。在婚礼飞行期间,蜂王分散在巢穴周围;因此,拥有更多殖民地的地区更有可能拥有女王着陆点,在那里它们开始新的巢穴挖掘。
- 检查先前选择的区域,在 阿塔 蚂蚁的繁殖季节是否有结婚飞行的迹象。跟踪婚礼飞行日的环境状况,例如炎热和降雨天气。
- 在先前选择的区域识别切叶蚁巢,并寻找表明有翼繁殖蚁即将离开的外部特征。巢穴的特征包括隧道入口变宽(图1),增加的工人流量对可能的捕食者表现出更具攻击性的行为,以及出现在隧道入口处的有翼繁殖蚁(图1)。当心降雨后的高湿度天,因为它们通常在婚礼飞行之前。
- 准备带有底部石膏层的塑料盖容器,以单独保留皇后。确保容器体积约为 200 mL,底部的石膏层高度约为 1 cm,并且具有高吸水性以控制湿度。
注意:要准备石膏底座,请按照制造商的说明进行操作。 - 准备一个温度恒定为23±1°C的环境,相对湿度约为70%±10%。选择一个没有激烈活动和人流量大的地方,以避免振动和干扰。使用中性香味的清洁产品,以防止对蚂蚁行为的任何干扰。
注意:特定环境条件的波动会导致水凝结或水分流失,并损害真菌园。 - 婚礼飞行后,收集已经开始筑巢挖掘的无翼女王,并将它们小心地单独放入用石膏层准备的塑料容器中。避免徒手触摸蜂王,并使用乳胶手套或昆虫镊子。
注意:翅膀移除和土壤挖掘行为表明已经交配的生殖雌性,因此能够开始一个新的殖民地。皇后区收藏也被视为这件作品中的第一个收藏。 - 将装有蜂王的容器移动到先前选择的受控环境的位置。执行蜂王运输时要格外小心,避免过多的干扰并保持最低温度恒定。
- 收集后约3天内不要操纵或移动蜂王,以避免压力。

图1.巢穴入口扩大,有翅膀的蚂蚁繁殖和工人。加宽的隧道入口是指示阿塔婚礼飞行发生的巢穴特征之一。 请点击此处查看此图的大图。
{kind=link}
2. 皇后区的维护
- 最初,在针头注射器的帮助下,每2天向受体的石膏层添加2.5毫升水。
- 不要打开收件人,而是小心地用针刺穿容器盖,以避免由于操作而受到干扰。在此期间可以使用相同的孔。确保添加的水不会浸泡石膏层。避免直接浇水女王、最初的真菌海绵和任何未成熟的。只要真菌花园表现出干燥的一面,缺水,请灌溉石膏层。
- 收集两周后,检查真菌是否被蜂王反刍。如果没有真菌,转移从已建立的菌落获得的约2g真菌。此外,如果真菌没有发育,请执行此步骤。
注意:对于真菌转移,有必要从已建立的菌落中收集健康的真菌并清除其上可能出现的所有蚂蚁。使用汤匙、昆虫学镊子和乳胶手套来操纵真菌。 - 在第一批工人出现后,根据菌落的切割活动,开始定期提供年轻和薄叶的碎片。确保提供的叶子是健康的,并且植物没有经过杀虫剂或其他化学物质的处理。在早期阶段,确保叶子碎片不超过4厘米。
注意:由于第一批工人开始觅食,因此必须在它们出现后提供植物材料。产品频率取决于工人将植物材料加入真菌的敏捷性,但可以每周 2-3 天。也可以提供燕麦片和玉米片,但应与叶子交替使用,以避免真菌干燥。 - 提供新叶时,清除菌落废物和干燥的叶子碎片。操纵皇后时,避免使用香水、保湿霜、面霜或任何有强烈气味的物质。此外,在所有过程中使用乳胶手套。
- 跟随菌落的发育,当真菌园达到容器体积的至少一半时,将菌落转移到人工持久巢中。
注意:由于每个菌落的发育速度是固有的,因此没有估计的菌落转移时间。通常,由于真菌花园小,来自第一个收集的菌落被转移到具有最大体积为1L的真菌花园室的巢穴中。
3. 幼菌群的收集
- 获取大约 500 mL 体积的塑料容器。
- 婚后约6个月,在先前标记的切叶蚁位置上识别带有早期Atta巢穴的颗粒状土壤颗粒(图1)(图2)的指示性塔形土丘。
注意:婚礼飞行六个月后,幼蜂群的巢穴估计在土壤中深达1米。在此期间指出了一个新的收藏品,以实现大量成功和持久菌落的更高机会。 - 用花园锄头挖掘巢穴入口,直到到达容纳年轻殖民地的房间。收集蜂王、真菌园、未成熟和年轻工人,并将它们放入塑料容器中。尽可能轻柔地执行收集过程。
注意:当然,也会收集大量的土壤,应该在实验室未来的维护程序中逐渐清除。 - 将装有菌落的塑料盖容器移动到指定的受控环境中。运输幼菌时要格外小心,避免过多干扰并保持最低温度恒定性。避免操纵或移动菌落约 3 天以避免压力。如果房间有活跃的例行程序,可以在菌落上一块深色布。

图2.塔形土丘。 特征性的塔形土丘表明存在Atta sexdens和Atta laevigata的早期殖民地。请点击此处查看此图的大图。
{kind=link}
4. 维持幼菌落
- 每周提供3次薄嫩叶。
- 确保提供的叶子是健康的,并且植物没有经过杀虫剂或其他化学物质的处理。在此阶段,确保叶子碎片长度至少为 7 厘米。
注意:也可以提供燕麦片和玉米片,但应与叶子交替使用,以避免真菌干燥。 - 供应频率取决于工人将植物材料加入真菌的敏捷性。随着切割活动的激烈,每周三次或每周 5 天每天增加两次供应。
- 确保提供的叶子是健康的,并且植物没有经过杀虫剂或其他化学物质的处理。在此阶段,确保叶子碎片长度至少为 7 厘米。
- 提供新叶子时,请借助勺子清除菌落废物,包括土壤残留物。在所有过程中使用乳胶手套。在操纵年轻菌落时,避免使用香水,保湿剂,面霜或任何具有强烈气味的物质。
注意:工人自己将从真菌中分离土壤和废物。 - 跟随菌落的发育,当真菌园达到容器体积的至少一半时,将菌落转移到人工持久巢中。
注意:由于每个菌落的发育速度是固有的,因此没有估计的菌落转移时间。

图3:容纳Atta sexdens 和 Atta laevigata 菌落的人工巢的类型。 切叶蚁的持久人造巢的插图:隐蔽的垂直巢设置,隐蔽的水平巢设置和开放的竞技场巢设置。 请点击此处查看此图的大图。
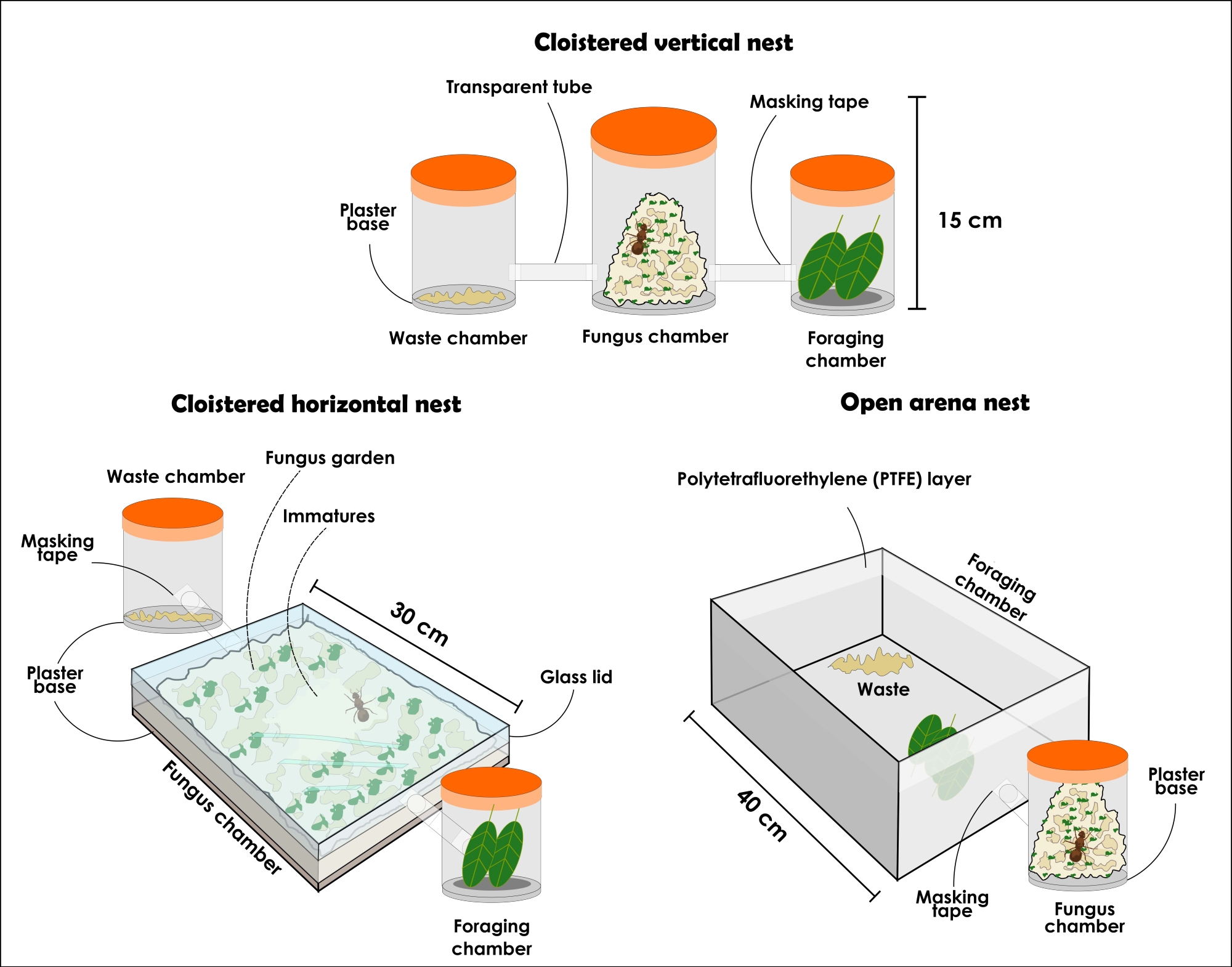{kind=link}
5. 耐用的人工巢
- 如下所述准备一个封闭的垂直巢穴设置(图3和 图4)。
注意:与世隔绝的巢穴配置应始终具有不同的接收者,分别摆出(1)真菌花园室,(2)废物处理室和(3)觅食室。最初,它应该从三个容器开始,但可以添加更多的收件人来增加真菌花园。巢的大小和材料可能根据其实验目的而有所不同,尽管这里描述并建议使用透明材料。容器必须没有开口,否则蚂蚁会逃脱。下面描述的巢穴类型可用于一般研究,但由于取下盖子时会引起干扰,因此不建议用于自信的蚂蚁收集,从而导致个体之间的极大骚动。尽管如此,由于材料透明,即使真菌园填满了整个容器,也可以找到女王和不同的蚂蚁种姓。由于这是一种趋势,因此始终假设未成熟的真菌位于真菌花园的中心,因为它占据了垂直容器中的大部分可用空间。- 选择一个大约 1 L 的透明有盖容器,并添加 1 厘米的高吸水性石膏底座。这将是真菌花园室。选择两个带盖的透明容器,每个容器约 500 mL,作为废物处理和觅食室。蚂蚁将选择每个房间,然后确保它们不互换。
注意:要准备石膏底座,请按照制造商的说明进行操作。 - 穿孔并用透明管或软管连接三个容器。如有必要,在管子边缘贴上遮蔽胶带,以确保大腿与容器的连接并避免蚂蚁逃逸。将石膏基础容器放在中间,其他容器放在相对的两侧。
- 小心地将选定菌落的真菌海绵(参见步骤2.5和步骤4.3)与蜂王,工人和未成熟菌一起转移到石膏基础容器中。在转移之前,请确保石膏底座已浇水。使用乳胶手套。
- 选择一个大约 1 L 的透明有盖容器,并添加 1 厘米的高吸水性石膏底座。这将是真菌花园室。选择两个带盖的透明容器,每个容器约 500 mL,作为废物处理和觅食室。蚂蚁将选择每个房间,然后确保它们不互换。
- 如下所述准备一个隐蔽的水平巢穴设置(图3和 图4)。
注意:水平配置的巢可以密切观察真菌花园和工人朝向它的活动。由于真菌园的年轻部分位于顶部,因此可以观察到工人最近提供的基质正在合并。真菌的新部分可以通过其颜色发现,这将类似于最后提供的资源的颜色,而较旧的部分通常带有米色。后代和蜂王也可以很容易地找到,因为在水平容器中,它们通常位于真菌花园的顶部,即使它占据了大部分空间。这种配置可用于以行为为中心的研究、焦点采样和科学推广目的,因为它给出了巢穴内部组织的感觉。- 获取一个尺寸约为 31 厘米 x 21 厘米 x 4.5 厘米的透明带盖容器,并添加一层 1 厘米的高吸水性石膏底座。这将是真菌花园室。选择两个带盖的透明容器,每个容器约 500 mL,作为废物处理和觅食室。蚂蚁将选择每个房间,然后确保它们不互换。
注意:要准备石膏底座,请按照制造商的说明进行操作。如有必要,用遮蔽胶带关闭盖子和容器之间的小空间,以防止蚂蚁逃脱。 - 穿孔并用透明管或软管连接容器。如有必要,在管子边缘贴上遮蔽胶带,以确保大腿与容器的连接并避免蚂蚁逃逸。将石膏基础容器放在中间,其他容器放在相对的两侧。
- 小心地将选定菌落的真菌海绵(参见步骤2.5和步骤4.3)与蜂王,工人和未成熟菌一起转移到石膏基础容器中。在转移之前,请确保石膏底座已浇水。使用乳胶手套。
- 获取一个尺寸约为 31 厘米 x 21 厘米 x 4.5 厘米的透明带盖容器,并添加一层 1 厘米的高吸水性石膏底座。这将是真菌花园室。选择两个带盖的透明容器,每个容器约 500 mL,作为废物处理和觅食室。蚂蚁将选择每个房间,然后确保它们不互换。
- 如下所述准备一个开放的竞技场巢穴设置(图 3 和 图 5)。
注意:开放的竞技场巢穴允许收集蚂蚁而不会受到太大干扰和觅食行为分析。它们还可以为科学推广目的提供自然界中发现的殖民地的可靠表示。- 选择一个大约 1 L 的透明盖容器,并添加 1 厘米的高吸水性石膏层底座。这将是真菌花园室。
注意:建议从 1 L 容器开始,然后逐渐传递到容量较大的容器中,以获得更大的真菌花园。但是,容器不应超过5升的容积。根据需要添加任意数量的容器。 - 选择一个开放的竞技场。竞技场的大小可以根据学习目的而有所不同。如果选择大型竞技场,请将装有真菌园的容器放在其内部(图5)。如果是小竞技场,请使用透明软管或管将其连接到真菌花园容器(图3)。竞技场将用作觅食和废物处理室,因此请确保它不会太小。
- 将一层聚四氟乙烯液体单次涂抹到竞技场边界以容纳蚂蚁。使用浸有液体的棉花和丁腈手套。
注意:使用聚四氟乙烯液体时避免吸入和皮肤接触。 - 小心地将选定菌落的真菌海绵(参见步骤2.5和步骤4.3)与蜂王,工人和未成熟菌一起转移到石膏基础容器中。在转移之前,请确保石膏底座已浇水。使用乳胶手套。
- 选择一个大约 1 L 的透明盖容器,并添加 1 厘米的高吸水性石膏层底座。这将是真菌花园室。

图4:切叶蚁Atta sexdens 和 Atta laevigata的人工隐居巢穴。 与世隔绝的垂直巢设置顶部(A)和侧视图(B);回廊水平巢设置顶部(C)和侧视图(D)。 请点击此处查看此图的大图。
{kind=link}

图5:切叶蚁Atta sexdens 和 Atta laevigata的人工开放竞技场巢穴。 阿 塔六分球顶部(A)和侧视图(B)的开放式竞技场巢穴设置。1)真菌花园室;2)浪费;3)橙片;4)带有聚四氟乙烯(PTFE)层的玻璃。 请点击此处查看此图的大图。
6. 发达菌落的维护
- 每天在每个菌落中至少提供一片大叶子进入觅食室,并附有 1 L 真菌园。如果蚂蚁的切割活动很强烈,请增加叶子的数量。如果真菌干燥,请预先润湿叶子以提供额外的湿度。在与世隔绝的巢穴中,迅速进行祭品,以避免蚂蚁逃离觅食室。
注意:在这里,叶子是从桑树(桑葚),芒果(芒果),桉树(桉 树属),jambolan(Syzygium cumini),芙蓉(木槿 属),金合欢(Acalypha wilkesiana)和Ligustrum(Ligustrum lucidum)等植物物种中收集的。- 提供橙子和苹果等水果,以及燕麦和玉米片,以多样化和补充饮食。对于觅食活动频繁的大型殖民地,每天提供燕麦片和玉米片,每周提供一次水果。如果不是这种情况,请与叶子交替提供薄片,但每周不超过三次,每月一次或两次水果。根据蚂蚁的觅食活动调整每种食物的数量和频率。
- 如果上述选项不可用,请确定蚂蚁在区域树木和灌木的叶子、花朵和果实中的觅食偏好,甚至是商业化的蔬菜、谷物和其他薄片。避免提供含有防御性化合物和杀虫剂的资源。
- 每2周从所有菌落中取出废物室的所有内容物。出于人口控制目的,也移除工人。如果工人将健康的真菌转移到废物室,请确保蜂王不在上面并将其移除。如果处理的废物量很大或太潮湿,请每周清除一次。
- 每当提供新的蚂蚁时,请从蚂蚁室中取出未带走的材料,并确保它始终干净。
- 如果工人将健康的真菌转移到觅食室,请打扰它,打开容器盖并将中性滑石粉涂抹在觅食室边缘表面。仅当真菌室上仍有一些空间时才执行此过程,这样工人才会将真菌转移回容器而不会丢失它或任何未成熟物。
- 如果需要更多的真菌花园,请添加另一个抹灰容器,并将一部分真菌海绵移入其中。直到真菌到达容器的一半,在真菌室中加入叶子。真菌园的生长应该逐渐发生,以免损害菌落平衡。如果需要更大的容器,请确保在转移之前让真菌占据最小容器的整个空间。不应让菌落废物和干叶积聚到真菌花园室中。
- 检查容器中的石膏底座,随着时间的推移,由于蚂蚁排泄、废物运输和高湿度浓度,它可能会变成深棕色并变得无效。此外,一些菌落可能会切割该层并处理掉它。在这些情况下,将真菌园转移到新的抹灰容器中。
结果
描述蚂蚁收集过程的流程图如图 6 所示。这里显示了使用上述收集、维护和巢穴设置协议获得的一些结果。

图6:采摘切叶蚁群的流程图。按照协议,第一次收集在婚礼飞行后立即进行。取下翅膀的蜂王被收集起来,放在一?...
讨论
这里描述的维持切叶蚁群的协议已经以自信和可复制的方式开发和应用了三十多年。它允许开展受现场条件限制的研究。因此,健康的蚂蚁和蚁群可用于多个领域的研究,例如个体和群体水平的比较形态学,毒理学51,52,组织学53和微生物学54,55,56。它们在实验?...
披露声明
作者没有利益冲突需要披露。
致谢
献给马里奥·奥托里(纪念) 和沃尔特·雨果·德·安德拉德·库尼亚,他们对切叶蚂蚁研究做出了巨大贡献。我们感谢圣保罗州立大学和生物科学研究所的支持。这项研究的部分资金来自巴西高级国家财政代码001、国家公民和技术发展委员会、圣保罗国家宪法权利保障基金和联合国国家发展基金会。
材料
| Name | Company | Catalog Number | Comments |
| Entomologic forceps | N/A | N/A | N/A |
| Glass tank | N/A | N/A | Tempered glass, custom made |
| Hose | N/A | N/A | Transparent, PVC 1/2 Inch x 2,0 mm |
| Latex gloves | Descarpack | 550301 | N/A |
| Nitrile gloves | Descarpack | 433301 | N/A |
| Open arena | N/A | N/A | Polypropylene crate |
| Plaster pouder | N/A | N/A | Plaster pouder used in construction, must be absorbant |
| Plastic Containers for collection | Prafesta | Natural Cód.: 8231/Natural Cód.: 8262 | Lidded, transparent , polypropylene |
| Plastic containers for nests | Prafesta | Discontinued | Polystyrene, hermetic |
| Teflon | Dupont | N/A | Polytetrafluoroethylene liquid (PTFE Dispertion 30) |
参考文献
- Wilson, E. O. . The Insect Societies. , (1971).
- Ortiz, D. P., Elizalde, L., Pirk, G. I. Role of ants as dispersers of native and exotic seeds in an understudied dryland. Ecological Entomology. 46 (3), 626-636 (2021).
- Christianini, A. V., Oliveira, P. S. Birds and ants provide complementary seed dispersal in a neotropical savanna. Journal of Ecology. 98 (3), 573-582 (2010).
- Camargo, P. H. S. A., Martins, M. M., Feitosa, R. M., Christianini, A. V. Bird and ant synergy increases the seed dispersal effectiveness of an ornithochoric shrub. Oecologia. 181 (2), 507-518 (2016).
- Sanders, D., van Veen, F. J. F. Ecosystem engineering and predation: the multi-trophic impact of two ant species. Journal of Animal Ecology. 80 (3), 569-576 (2011).
- Swanson, A. C., et al. Welcome to the Atta world: A framework for understanding the effects of leaf-cutter ants on ecosystem functions. Functional Ecology. 33 (8), 1386-1399 (2019).
- Meyer, S. T., et al. Leaf-cutting ants as ecosystem engineers: topsoil and perturbations around Atta cephalotes nests reduce nutrient availability. Ecological Entomology. 38 (5), 497-504 (2013).
- Sosa, B., Brazeiro, A. Positive ecosystem engineering effects of the ant Atta vollenweideri on the shrub Grabowskia duplicata. Journal of Vegetation Science. 21 (3), 597-605 (2010).
- De Almeida, T., et al. Above- and below-ground effects of an ecosystem engineer ant in Mediterranean dry grasslands. Proceedings of the Royal Society B: Biological Sciences. 287 (1935), 20201840 (2020).
- Folgarait, P. J. Ant biodiversity and its relationship to ecosystem functioning: a review. Biodiversity & Conservation. 7 (9), 1221-1244 (1998).
- Hölldobler, B., Wilson, E. O. . The Ants. , (1990).
- Barrera, C. A., Sosa-Calvo, J., Schultz, T. R., Rabeling, C., Bacci, M. Phylogenomic reconstruction reveals new insights into the evolution and biogeography of Atta leaf-cutting ants (Hymenoptera: Formicidae). Systematic Entomology. 47 (1), 13-35 (2021).
- Hölldobler, B., Wilson, E. O. . The Leafcutter Ants: Civilization By Instinct. , (2011).
- Branstetter, M. G., et al. Dry habitats were crucibles of domestication in the evolution of agriculture in ants. Proceedings of the Royal Society B: Biological Sciences. 284 (1852), 20170095 (2017).
- Solomon, S. E., et al. The molecular phylogenetics of Trachymyrmex Forel ants and their fungal cultivars provide insights into the origin and coevolutionary history of ‘higher-attine’ ant agriculture. Systematic Entomology. 44 (4), 939-956 (2019).
- Cristiano, M. P., Cardoso, D. C., Sandoval-Gómez, V. E., Simões-Gomes, F. C. Amoimyrmex Cristiano, Cardoso & Sandoval, gen. nov. (Hymenoptera: Formicidae): a new genus of leaf-cutting ants revealed by multilocus molecular phylogenetic and morphological analyses. Austral Entomology. 59 (4), 643-676 (2020).
- Schultz, T. R., Brady, S. G. Major evolutionary transitions in ant agriculture. Proceedings of the National Academy of Sciences of the United States of America. 105 (14), 5435-5440 (2008).
- Mueller, U. G., et al. Phylogenetic patterns of ant–fungus associations indicate that farming strategies, not only a superior fungal cultivar, explain the ecological success of leafcutter ants. Molecular Ecology. 27 (10), 2414-2434 (2018).
- Mueller, U. G., et al. Biogeography of mutualistic fungi cultivated by leafcutter ants. Molecular Ecology. 26 (24), 6921-6937 (2017).
- Weber, N. A. The fungus-culturing behavior of ants. American Zoologist. 12 (3), 577-587 (1972).
- Wilson, E. O. Caste and division of labor in leaf-cutter ants (Hymenoptera: Formicidae: Atta). Behavioral Ecology and Sociobiology. 7 (2), 157-165 (1980).
- Della Lucia, T. M. C. Formigas cortadeiras: da bioecologia ao manejo. Viçosa Editora UFV. 421, (2011).
- Autuori, M. Contribuição para o conhecimento da saúva (Atta spp). I. Evolução do sauveiro (Atta sex dens rubolpilosa Forel, 1908). Arquivos do Instituto Biologico Saul Paulo. 12, 197-228 (1941).
- Bueno, O. C., Hebling, M. J. A., Schneider, M. O., Pagnocca, F. C. Ocorrência de formas aladas de Atta sexdens rubropilosa Forel (Hymenoptera: Formicidae) em colônias de laboratório. Neotropical Entomology. 31 (3), 469-473 (2002).
- Mariconi, F. A. M. Biologo. As Saúvas. , (2021).
- Bento, J. M. S. . Condições climáticas para o vôo nupcial e reconhecimento dos indivíduos em Atta sexdens rubropilosa (Hymenoptera: Formicidae). , (1993).
- Little, A. E. F., Murakami, T., Mueller, U. G., Currie, C. R. The infrabuccal pellet piles of fungus-growing ants. Naturwissenschaften. 90 (12), 558-562 (2003).
- Kerr, W. E. Acasalamento de rainhas com vários machos em duas espécies da tribo Attini. Revista Brasileira de Biologia. 21, 45-48 (1961).
- Kerr, W. E. Tendências evolutivas na reprodução dos himenópteros sociais. Arquivos do Museu Nacional. 52, (1962).
- Cremer, S., Armitage, S. A. O., Schmid-Hempel, P. Social immunity. Current Biology. 17 (16), 693-702 (2007).
- Hernández, J. V., Jaffé, K. Dano econômico causado por populações de formigas Atta laevigata (F. Smith) em plantações de Pinus caribaea (Mor.) e elementos para o manejo da praga. Anais da Sociedade Entomológica do Brasil. 24 (2), 287-298 (1995).
- Kempf, W. W. Catálogo abreviado das formigas da Região Neotropical Studia Entomologica. antbase.org. 15, 3 (1972).
- Della Lucia, T. M. C., Gandra, L. C., Guedes, R. N. C. Managing leaf-cutting ants: peculiarities, trends and challenges. Pest Management Science. 70 (1), 14-23 (2013).
- Boaretto, M. A. C., Forti, L. C. Perspectivas no controle de formigas-cortadeiras. Série Técnica IPEF. 11 (30), 31-46 (1997).
- Folgarait, P. J., Goffré, D. Conidiobolus lunulus, a newly discovered entomophthoralean species, pathogenic and specific to leaf-cutter ants. Journal of Invertebrate Pathology. 186, 107685 (2021).
- Cardoso, S. R. S., Rodrigues, A., Forti, L. C., Nagamoto, N. S. Pathogenicity of filamentous fungi towards Atta sexdens rubropilosa (Hymenoptera: Formicidae). International Journal of Tropical Insect Science. 42 (2), 1215-1223 (2022).
- Ichinose, K., Rinaldi, I., Forti, L. C. Winged leaf-cutting ants on nuptial flights used as transport by Attacobius spiders for dispersal. Ecological Entomology. 29 (5), 628-631 (2004).
- Pagnocca, F. C., Rodrigues, A., Nagamoto, N. S., Bacci, M. Yeasts and filamentous fungi carried by the gynes of leaf-cutting ants. Antonie Van Leeuwenhoek. 94 (4), 517-526 (2008).
- Attili-Angelis, D., et al. Novel Phialophora species from leaf-cutting ants (tribe Attini). Fungal Diversity. 65 (1), 65-75 (2014).
- Delabie, J. H. C., do Nascimento, I. C., Mariano, C. S. F. Estratégias de reprodução e dispersão em formigas attines, com exemplos do sul da Bahia. XIX Congresso Brasileiro de Entomologia. , 16-21 (2002).
- Fjerdingstad, E. J., Boomsma, J. J. Variation in size and sperm content of sexuals in the leafcutter ant Atta colombica. Insectes Sociaux. 44 (3), 209-218 (1997).
- Currie, C. R., Mueller, U. G., Malloch, D. The agricultural pathology of ant fungus gardens. Proceedings of the National Academy of Sciences of the United States of America. 96 (14), 7998-8002 (1999).
- Moser, J. C., et al. Eye size and behaviour of day-and night-flying leafcutting ant alates. Journal of Zoology. 264 (1), 69-75 (2004).
- Moreira, S. M., Rodrigues, A., Forti, L. C., Nagamoto, N. S. Absence of the parasite Escovopsis in fungus garden pellets carried by gynes of Atta sexdens. Sociobiology. 62 (1), 34-38 (2015).
- Arcuri, S. L., et al. Yeasts found on an ephemeral reproductive caste of the leaf-cutting ant Atta sexdens rubropilosa. Antonie Van Leeuwenhoek. 106 (3), 475-487 (2014).
- Staab, M., Kleineidam, C. J. Initiation of swarming behavior and synchronization of mating flights in the leaf-cutting ant Atta vollenweideri FOREL, 1893 (Hymenoptera: Formicidae). Myrmecol. News. 19, 93-102 (2014).
- Gálvez, D., Chapuisat, M. Immune priming and pathogen resistance in ant queens. Ecology and Evolution. 4 (10), 1761-1767 (2014).
- Baer, B., Armitage, S. A. O., Boomsma, J. J. Sperm storage induces an immunity cost in ants. Nature. 441 (7095), 872-875 (2006).
- Carlos, A. A. . Semioquímicos e comunicação sonora em formigas cortadeiras (Hymenoptera: Formicidae). , (2013).
- Veja um FORMIGUEIRO por DENTRO. Boravê Available from: https://youtu.be/sN99x_Rjf90 (2021)
- Ortiz, G., Vieira, A. S., Bueno, O. C. Toxicological and morphological comparative studies of insecticides action in leaf-cutting ants. International Journal of Agriculture Innovations and Research. 6 (3), 516-522 (2017).
- Decio, P., Silva-Zacarin, E. C. M., Bueno, F. C., Bueno, O. C. Toxicological and histopathological effects of hydramethylnon on Atta sexdens rubropilosa (Hymenoptera: Formicidae) workers. Micron. 45, 22-31 (2013).
- Vieira, A. S., Morgan, E. D., Drijfhout, F. P., Camargo-Mathias, M. I. Chemical composition of metapleural gland secretions of fungus-growing and non-fungus-growing ants. Journal of Chemical Ecology. 38 (10), 1289-1297 (2012).
- Vieira, A. S., Ramalho, M. O., Martins, C., Martins, V. G., Bueno, O. C. Microbial communities in different tissues of Atta sexdens rubropilosa leaf-cutting ants. Current Microbiology. 74 (10), 1216-1225 (2017).
- Ramalho, M. d. e. O., Martins, C., Morini, M. S. C., Bueno, O. C. What can the bacterial community of Atta sexdens (Linnaeus, 1758) tell us about the habitats in which this ant species evolves. Insects. 11 (6), 332 (2020).
- Machado, L. M., et al. Attractivity or repellence: relation between the endophytic fungi of Acalypha, Colocasia and the leaf-cutting ants—Atta sexdens. Advances in Entomology. 9 (2), 85-99 (2021).
- Moreira, A., Forti, L. C., Andrade, A. P., Boaretto, M. A., Lopes, J. Nest architecture of Atta laevigata (F. Smith, 1858) (Hymenoptera: Formicidae). Studies on Neotropical Fauna and Environment. 39 (2), 109-116 (2004).
- Della Lucia, T. M. C., Moreira, D. D. O., Oliveira, M. A., Araújo, M. S. Perda de peso de rainhas de Atta durante a fundação e o estabelecimento das colônias. Revista Brasileira de Biologia. 55 (4), 533-536 (1995).
- Fujihara, R. T., Camargo, R. d. a. S., Forti, L. C. Lipid and energy contents in the bodies of queens of Atta sexdens rubropilosa Forel (Hymenoptera, Formicidae): pre-and post-nuptial flight. Revista Brasileira de Entomologia. 56 (1), 73-75 (2012).
- da Silva Camargo, R., Forti, L. C. Queen lipid content and nest growth in the leaf cutting ant (Atta sexdens rubropilosa) (Hymenoptera: Formicidae). Journal of Natural History. 47, 65-73 (2013).
- Camargo, R. S., Forti, L. C., Fujihara, R. T., Roces, F. Digging effort in leaf-cutting ant queens (Atta sexdens rubropilosa) and its effects on survival and colony growth during the claustral phase. Insectes Sociaux. 58 (1), 17-22 (2011).
- Mota Filho, T. M. M., Garcia, R. D. M., Camargo, R. S., Stefanelli, L. E. P., Forti, L. C. Observations about founding queens (Atta sexdens) and their unusual behavior. International Journal of Agriculture Innovations and Research. 9, 352-357 (2021).
- Barcoto, M. O., Pedrosa, F., Bueno, O. C., Rodrigues, A. Pathogenic nature of Syncephalastrum in Atta sexdens rubropilosa fungus gardens. Pest Management Science. 73 (5), 999-1009 (2017).
- Silva, A., Bacci, M., Pagnocca, F. C., Bueno, O. C., Hebling, M. J. A. Production of polysaccharidases in different carbon sources by Leucoagaricus gongylophorus Möller (Singer), the symbiotic fungus of the leaf-cutting ant Atta sexdens Linnaeus. Curr. Microbiology. 53 (1), 68-71 (2006).
- Majoe, M., Libbrecht, R., Foitzik, S., Nehring, V. Queen loss increases worker survival in leaf-cutting ants under paraquat-induced oxidative stress. Philosophical Transactions of the Royal Society B. 376 (1823), 20190735 (2021).
- Della Lucia, T. M. C., Peternelli, E. F. O., Lacerda, F. G., Peternelli, L. A., Moreira, D. D. O. Colony behavior of Atta sexdens rubropilosa (Hymenoptera: Formicidae) in the absence of the queen under laboratory conditions. Behavioural Processes. 64 (1), 49-55 (2003).
- Sales, T. A., Toledo, A. M. O., Zimerer, A., Lopes, J. F. S. Foraging for the fungus: why do Acromyrmex subterraneus (Formicidae) queens need to forage during the nest foundation phase. Ecological Entomology. 46 (6), 1364-1372 (2021).
- Forti, L. C., et al. Do workers from subspecies Acromyrmex subterraneus prepare leaves and toxic baits in similar ways for their fungus garden. Neotropical Entomology. 49 (1), 12-23 (2020).
- Dorigo, A. S., et al. Projeto Primeiros Passos na Ciência: rompendo barreiras sociais e estreitando laços entre a comunidade acadêmica e o ensino médio público. Revista Brasileira de Extensão Universitária. 11 (1), 47-59 (2020).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。