需要订阅 JoVE 才能查看此. 登录或开始免费试用。
Method Article
基于刀豆球蛋白A的沉降测定法,用于测量葡聚糖磷酸酶的底物结合
摘要
该方法描述了一种基于凝集素的 体外 沉降测定,以量化葡聚糖磷酸酶和支链淀粉的结合亲和力。该共沉降测定法对于测量葡聚糖磷酸酶底物结合是可靠的,可应用于各种可溶的葡聚糖底物。
摘要
葡聚糖磷酸酶属于更大的双重特异性磷酸酶(DSP)家族,可去磷酸化葡聚糖底物,例如动物中的糖原和植物中的淀粉。具有模型葡聚糖底物的葡聚糖磷酸酶的晶体结构揭示了由DSP和碳水化合物结合结构域组成的独特的葡聚糖结合界面。然而,定量测量葡聚糖-葡聚糖磷酸酶与生理相关底物的相互作用对于葡聚糖磷酸酶家族酶的生物学理解和能量代谢的调节至关重要。本手稿报告了一种基于刀豆球蛋白 A (ConA) 的体外沉降测定,旨在检测葡聚糖磷酸酶对不同葡聚糖底物的底物结合亲和力。作为概念验证,测定了葡聚糖磷酸酶拟南芥淀粉过量4(SEX4)和支链淀粉的解离常数(KD)。SEX4突变体和葡聚糖磷酸酶家族的其他成员的表征进一步证明了该测定法在评估蛋白质 - 碳水化合物相互作用的差异结合方面的实用性。这些数据证明了该测定法适用于表征各种淀粉和糖原相互作用的蛋白质。
引言
葡聚糖磷酸酶是蛋白酪氨酸磷酸酶 (PTP) 超家族1 中功能多样化的双重特异性磷酸酶 (DSP) 亚家族的成员。它们存在于大多数生命形式中,包括广泛不同的光合生物、人类、脊椎动物以及一些无脊椎动物和原生生物2,3,4。植物含有三种已知的葡聚糖磷酸酶:淀粉过量4(SEX4),Like Four1(LSF1)和Like Four2(LSF2)5,6,7。缺乏葡聚糖磷酸酶的植物显示出暂时淀粉降解和淀粉在叶片中积累的速率降低8,9。拉福林是葡聚糖磷酸酶家族的创始成员,该家族使脊椎动物和人类的糖原去磷酸化3,10。拉福林的突变导致神经退行性拉福拉病,这是一种致命的常染色体隐性癫痫形式11。葡聚糖磷酸酶是糖原和淀粉代谢所必需的,并且已成为调节植物淀粉含量和治疗神经退行性Lafora病的重要酶12,13。最近对葡聚糖磷酸酶的X射线晶体学研究与模型葡聚糖底物揭示了底物结合和葡聚糖去磷酸化的催化机制14,15,16,17。然而,目前对葡聚糖磷酸酶如何与其生理底物结合的理解是不完整的。
淀粉是由80%-90%支链淀粉和10%-20%直链淀粉18制成的葡萄糖不溶性聚合物。植物葡聚糖磷酸酶的底物是磷酸化的碳水化合物分子,如糖原和淀粉颗粒。磷酸化的葡萄糖基残基以1:600的磷酸:葡萄糖基残基比例存在。有趣的是,磷酸盐仅存在于支链淀粉分子19上。主要植物葡聚糖磷酸酶SEX4作用于淀粉颗粒以去磷酸化支链淀粉分子。SEX4的X射线晶体结构与结构引导的诱变研究相结合,证明了SEX4对葡聚糖结构内不同位置的独特底物特异性15。我们最近表明,SEX4的生物学相关活性只能在作用于其可溶的支链淀粉底物20时观察到。然而,由于底物的结构复杂性、更广泛的结合特异性以及蛋白质与其底物之间的低结合亲和力,理解葡聚糖-SEX4 相互作用已被证明是困难的。这些问题阻碍了利用蛋白质-配体相互作用中常用的方法的能力,例如等温滴定量热法(ITC),核磁共振(NMR)波谱和基于酶联免疫吸附测定(ELISA)的测定。
有趣的是,我们对碳水化合物 - 蛋白质相互作用的大部分理解都来自研究凝集素。刀豆球蛋白A(ConA)是一种豆科凝集素家族,最初是从杰克豆中提取的蛋白质。ConA以高特异性结合碳水化合物,这有利于其在药物靶向和递送应用中的使用。ConA与含有非还原性α-D-甘露糖基和α-D-葡萄糖基的各种底物的结合已被广泛研究19,20。市售ConA结合琼脂糖珠通常用于纯化糖蛋白和糖脂21。ConA通过葡萄糖残基的C3,C4和C6羟基与这些葡聚糖结合。ConA-琼脂糖珠也已成功用于测量糖原-蛋白质和淀粉-蛋白质相互作用的结合22,23。在这项研究中,我们使用ConA-琼脂糖珠开发了一种结合测定法,以测量葡聚糖磷酸酶-支链淀粉相互作用的结合特异性。
以前,使用基于ConA的沉降测定来评估葡聚糖磷酸酶底物结合能力14,20,24。在这项研究中,使用相同的策略开发了一种确定葡聚糖-葡聚糖磷酸酶和碳水化合物相互作用的结合亲和力的新方法。该方法在研究各种可溶的碳水化合物-蛋白质相互作用方面也具有优势。
研究方案
1. ConA-琼脂糖珠的制备
- 制备 250 mL 含有 67 mM HEPES (pH 7.5)、10 mM MgCl 2 和 0.2 mM CaCl2 的结合缓冲液。使用1M NaOH溶液调节pH值。
- 将 250 μL ConA-琼脂糖珠悬浮液移液到 1.5 mL 微量离心管中。将内容物在4°C下以10,000× g 离心30秒。 弃去上清液。
注意:用于测定的每个支链淀粉浓度需要 250 μL 1.5 mL 微量离心管中的 ConA-琼脂糖珠。 - 向含有 250 μL ConA-琼脂糖珠的每个试管中加入 750 μL 结合缓冲液。在4°C下以10,000× g 离心管1分钟。 除去上清液。重复此步骤2x,以确保用结合缓冲液适当洗涤和平衡磁珠。
2.支链淀粉溶液的制备
- 制作 10 mg/mL 马铃薯支链淀粉的储备溶液。支链淀粉不溶于水,受热溶解。为了溶解,将 0.1 g 马铃薯支链淀粉加入 10 mL 蒸馏水中。在80°C的水浴中加热悬浮液1小时或直到溶液不再浑浊。
- 让溶液恢复到室温(RT),反复涡旋以避免结块。
- 醇碱处理是溶解支链淀粉底物的替代方法。要使用此方法溶解,请按照以下步骤操作。
- 将 0.5 g 支链淀粉底物悬浮在 5 mL 20% 乙醇和 5 mL 2 M NaOH 中。在室温下剧烈搅拌内容物15-20分钟。
- 接下来,加入 10 mL 水,并通过加入 2 M HCl 将溶液的 pH 值调节至 6.5。 用蒸馏水将所得溶液的体积提高到 50 mL,制成 10 mg/mL 支链淀粉溶液。
- 稀释 10 mg/mL 可溶的支链淀粉溶液,制成一系列 2 mL 稀释的支链淀粉溶液。例如,进行 10 mg/mL 的半稀释以制备一系列支链淀粉浓度(5 mg/mL、2.5 mg/mL、1.25 mg/mL、0.625 mg/mL、0.3125 mg/mL、0.156 mg/mL、0.078 mg/mL、0.039 mg/mL、0.019 mg/mL 和 0 mg/mL)。
3.ConA-琼脂糖的制备:支链淀粉珠
- 将 250 μL 每种稀释的支链淀粉溶液加入 1.5 mL 微量离心管中,该离心管含有在结合缓冲液中预先平衡的 250 μL ConA-琼脂糖微球。将内容物充分混合。用相应的支链淀粉浓度标记试管。
- 将内容物在4°C的旋转轮上孵育30分钟。
注意:20分钟后,ConA-琼脂糖:支链淀粉结合复合物随着时间的推移没有变化。通过从10分钟到1小时的变化孵育时间来选择30分钟的孵育时间,以确保达到平衡。 - 将试管以10,000× g 离心1分钟。将上清液收集在新标记的 1.5 mL 微量离心管中。保存这些上清液级分以进行D-葡萄糖测定12 (支链淀粉的酸水解,然后通过酶测定 法测定 葡萄糖的UV)。此步骤对于确保所有支链淀粉都与珠子结合是必要的。
- 将 750 μL 结合缓冲液加入 ConA-琼脂糖:支链淀粉珠中。将试管以10,000× g 离心1分钟。弃去上清液以去除任何未结合的支链淀粉分子。
- 重复步骤3.4以确保充分洗涤。现在,每个试管都含有与不同数量的支链淀粉底物结合的ConA-琼脂糖珠。
4. 用ConA-琼脂糖孵育SEX4:支链淀粉珠
- 将 250 μL ConA-琼脂糖:支链淀粉珠与 100 μL 结合缓冲液混合,其中包括 10 μg SEX4 蛋白、10 mM 二硫苏糖醇 (DTT) 和 10 μM 蛋白酶抑制剂混合物 (PIC)。请注意,每个试管中的总体积为 350 μL。
注意:添加蛋白酶抑制剂混合物作为预防措施,以避免任何不必要的SEX4降解。这是一个可选步骤。在该测定中,使用重组蛋白拟南芥SEX4(AtSEX4)。纯化的蛋白质含有通过化学发光检测蛋白质所必需的N末端组氨酸标签。有关葡聚糖磷酸酶纯化的详细信息在先前的出版物14,20,24中描述。 - 将蛋白质和ConA-琼脂糖:支链淀粉珠悬浮液在4°C下孵育45分钟,轻轻旋转。
注意:选择45分钟的孵育时间以确保复合物达到平衡。 - 将试管以10,000× g 离心1分钟。使用凝胶上样吸头将 50 μL 上清液小心移液到新的 1.5 mL 微量离心管中。向含有 50 μL 收集的上清液级分的每个试管中加入 20 μL 4x SDS-PAGE 染料和 10 μL 水。将样品在95°C下加热10分钟。保存这些样品以运行SDS-PAGE凝胶。确保标有"上清液(S)"的10个新管具有相应的底物浓度。
- 将 750 μL 结合缓冲液加入 ConA-琼脂糖:支链淀粉:SEX4 磁珠中,以去除磁珠中的任何未结合蛋白。将试管以10,000× g 离心1分钟。再次重复此步骤以确保正确洗涤。弃去上清液。
- 将 20 μL 4x SDS-PAGE 染料和 80 μL 蒸馏水加入含有洗涤过的 ConA-琼脂糖:支链淀粉:SEX4 珠子的管中。将样品在95°C下加热10分钟,并以10,000× g 离心1分钟。
- 弃去沉淀并保存上清液以运行SDS-PAGE凝胶。将 80 μL 上清液移液到新管中,并将其标记为"沉淀 (P)"。
5. 运行SDS-PAGE凝胶
- 将 40 μL 未结合的蛋白质样品(在步骤 2.3 中制成,标记为 S)从最低底物浓度到最高底物浓度加载到 4%-12% 预制聚丙烯酰胺凝胶孔中,但保持第一泳道自由以加载蛋白质分子量标记物。使用第二块凝胶上样步骤2.5中制备的10个结合蛋白样品(标记为P)。
- 将新鲜制备的1x SDS-PAGE电泳缓冲液添加到设备的两个腔室中。在150 V下运行凝胶35分钟或直到染料前沿到达凝胶底部。
- 从设备中取出电泳凝胶并去除垫片和玻璃板。使用分离的凝胶进行蛋白质印迹分析。
6. 用于化学发光检测的蛋白质印迹14,15
注意:根据用户实验室中的蛋白质印迹设备,可以轻松修改/调整此方法。
- 制作 1 L 转膜缓冲液,其中含有 5.8 g Tris 碱、2.9 g 甘氨酸、0.37 g SDS 和 200 mL 甲醇。
- 将大小分离的蛋白质从聚丙烯酰胺凝胶转移到硝酸纤维素膜上。根据蛋白质转印方案14,15简要组装海绵,滤纸,凝胶和硝酸纤维素膜。在 70 V 下运行 1 小时。
- 为防止非特异性蛋白质结合,将含有1%-5%牛血清白蛋白(BSA)或乳蛋白的硝酸纤维素膜在50mL TBST缓冲液(20mM Tris [pH 7.5],150mM NaCl,0.1%吐温20)中孵育1小时。使用TBST缓冲液洗涤膜3x,以除去任何未结合的封闭溶液。
- 将膜与辣根过氧化物酶(HRP)连接的特异性His标记蛋白抗体孵育1小时。在TBST缓冲液中洗涤膜3次,以去除任何未结合的抗体。使用1:2,000稀释的TBST抗体,以获得最佳重现性和灵敏度。
- HRP 酶联抗体与 SEX4 蛋白的组氨酸标签特异性结合,在化学发光试剂存在下产生条带。对于数字成像,在 1.5 mL 管中制备等份化学发光底物溶液(每个 750 μL)的溶液。将膜在溶液中孵育至少5分钟。
- 将膜蛋白面朝下放在印迹扫描仪上,并运行采集软件以定量沉淀和上清液级分中的蛋白质。
7. 数据分析
- 使用采集软件和印迹扫描仪进行定量信号测量。将上清液和沉淀级分中的所有定量测量标准化为上样的总蛋白质。
注意:该软件允许量化上清液和沉淀级分中每个蛋白质条带的强度。 - 在饱和结合实验中,绘制蛋白质结合与支链淀粉浓度的百分比。将数据拟合为 Y = Bmax x X/(K D + X),使用数据分析软件计算 KD。
注意:Bmax是最大特异性结合,Y轴是蛋白质结合的百分比,X轴是支链淀粉浓度。

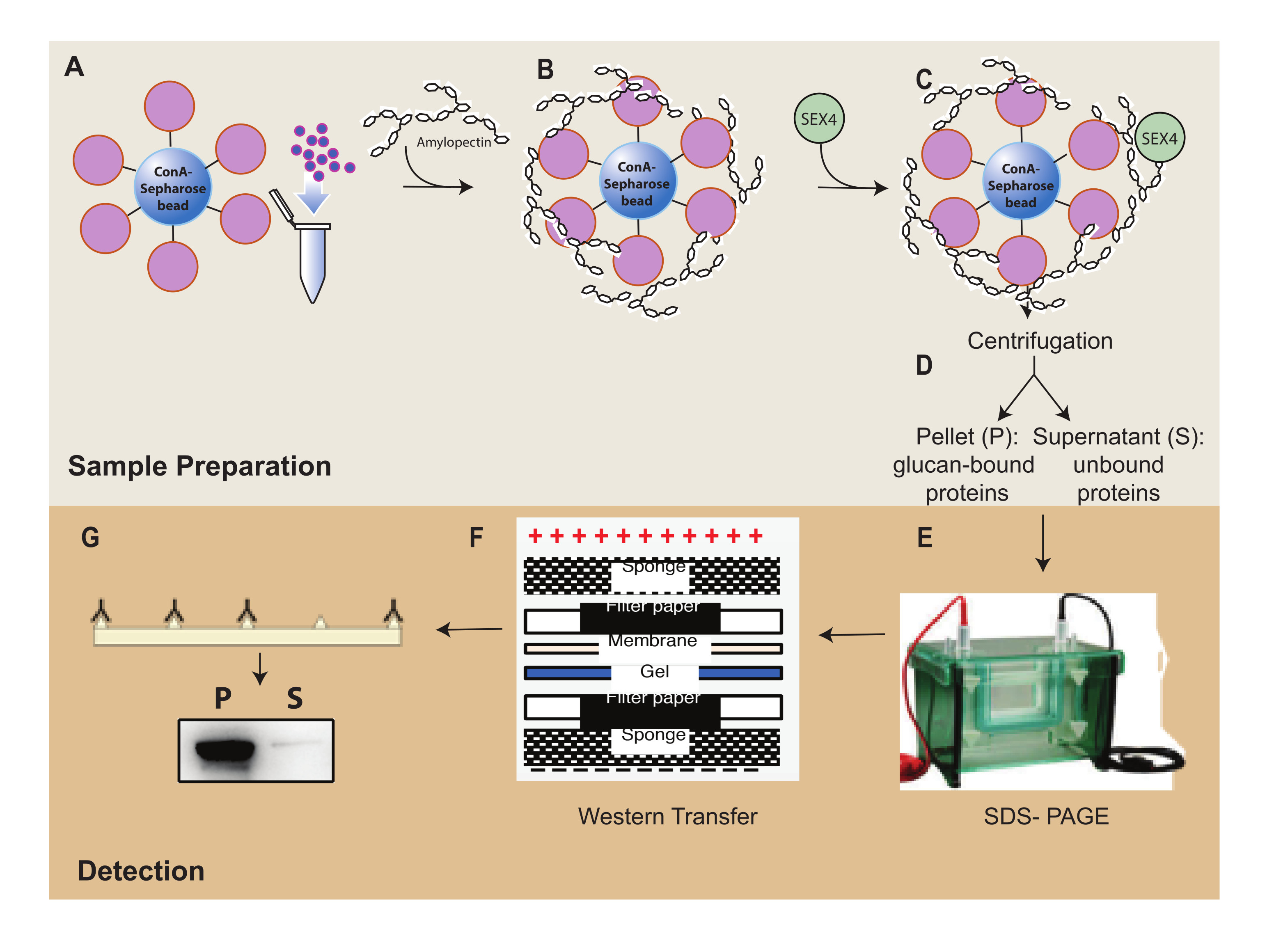
图 1:ConA-琼脂糖沉降测定工作流程概述。 (A) ConA-琼脂糖微球的制备。(B)与支链淀粉底物孵育。(C)与SEX4蛋白孵育。(D)通过离心分离结合和未结合的蛋白质级分。(E) 通过SDS-PAGE分离蛋白质。(F) 蛋白质印迹分析。(G)His标记的SEX4蛋白的化学发光检测。请点击此处查看此图的大图。
{kind=link}
结果
葡聚糖磷酸酶家族蛋白质的关键特征之一是它们与葡聚糖底物结合的能力。首先,使用SDS-PAGE分析SEX4与ConA-琼脂糖:支链淀粉珠的结合能力(图2A)。牛血清白蛋白(BSA)作为阴性对照,用于检测蛋白质与ConA-琼脂糖:支链淀粉珠的任何非特异性结合。蛋白质的SDS-PAGE分析显示沉淀级分中存在SEX4蛋白,上清液级分中存在BSA。W278A是一种已知的SEX4突变体,具有显着降低的葡聚糖结?...
讨论
这项研究证明了一种新的 体外 沉降测定法的成功开发,该测定法可以测定葡聚糖-葡聚糖磷酸酶相互作用的结合亲和力。该测定设计利用凝集素ConA通过葡萄糖的羟基残基 与 葡聚糖的特异性结合,间接捕获溶解的碳水化合物底物到琼脂糖珠上。这允许 通过离心分离 结合和未结合的蛋白质级分,并确定可溶的葡聚糖底物和葡聚糖磷酸酶的结合亲和力。发现所有测试的葡聚糖磷?...
披露声明
作者声明不存在利益冲突。
致谢
这项研究得到了美国国家科学基金会奖MCB-2012074的支持。作者感谢佛罗里达大学生物化学与分子生物学系的Craig W. Vander Kooi博士的宝贵讨论和支持。作者还感谢佛罗里达大学生物化学和分子生物学系的Matthew S. Gentry博士的支持。我们要感谢斯基德莫尔学院神经科学项目主席Sara Lagalwar博士允许我们使用LICOR C数字印迹扫描仪进行蛋白质印迹成像。
材料
| Name | Company | Catalog Number | Comments |
| 6x-His Tag monoclonal antibody (HIS.H8), HRP | Therm Fisher Scientific | MA1-21315-HRP | |
| Biorad gel electrophoresis and Western blot kit | Biorad | 1703930 | |
| Calcium chloride | Sigma-Aldrich | 208291 | |
| C-Digit blot scanner | LICOR | 3600-00 | Blot scanner |
| Complete protease inhibitor cocktail | Sigma-Aldrich | 11836170001 | |
| Concanavalin A-sepharose beads | Sigma-Aldrich | C9017 | This product contains in 0.1 M acetate buffer, pH 6, containing 1 M NaCl, 1 mM CaCl2, 1 mM MnCl2, and 1 mM MgCl2 in 20% ethanol |
| Centrifuge | Eppendorf | 5425R | |
| Glycine | Fisher Scientific | BP381-5 | |
| GraphPad Prism 8.0 software | GraphPad | Version 8.0 | Data analysis software |
| HEPES | Sigma-Aldrich | H8651 | |
| Image Studio | LICOR | 3600-501 | Acquisition Software |
| Magnesium chloride | Sigma-Aldrich | M2670 | |
| Methanol | Fisher Scientific | A452SK-4 | |
| Sodium dodecyl sulfate | Fisher Scientific | PI28312 | |
| Potato amylopectin | Sigma-Aldrich | A8515 | |
| Precast SDSPAGE Gels | Genscript | M00653S | |
| Tris base | Fisher Scientific | BP154-1 | |
| Tween 20 | Fisher Scientific | MP1TWEEN201 | |
| Westernsure premium chemiluminescence substrate | LI-COR | 926-95000 |
参考文献
- Meekins, D. A., Vander Kooi, C. W., Gentry, M. S. Structural mechanisms of plant glucan phosphatases in starch metabolism. The FEBS Journal. 283 (13), 2427-2447 (2016).
- Gentry, M. S., et al. The phosphatase laforin crosses evolutionary boundaries and links carbohydrate metabolism to neuronal disease. The Journal of Cell Biology. 178 (3), 477-488 (2007).
- Worby, C. A., Gentry, M. S., Dixon, J. E. Laforin, a dual specificity phosphatase that dephosphorylates complex carbohydrates. The Journal of Biological Chemistry. 281 (41), 30412-30418 (2006).
- Gentry, M. S., Pace, R. M. Conservation of the glucan phosphatase laforin is linked to rates of molecular evolution and the glucan metabolism of the organism. BMC Evolutionary Biology. 9, 138 (2009).
- Niittyla, T., et al. Similar protein phosphatases control starch metabolism in plants and glycogen metabolism in mammals. The Journal of Biological Chemistry. 281 (17), 11815-11818 (2006).
- Kotting, O., et al. STARCH-EXCESS4 is a laforin-like Phosphoglucan phosphatase required for starch degradation in Arabidopsis thaliana. The Plant Cell. 21 (1), 334-346 (2009).
- Comparot-Moss, S., et al. A putative phosphatase, LSF1, is required for normal starch turnover in Arabidopsis leaves. Plant Physiology. 152 (2), 685-697 (2010).
- Zeeman, S. C., Northrop, F., Smith, A. M., Rees, T. A starch-accumulating mutant of Arabidopsis thaliana deficient in a chloroplastic starch-hydrolysing enzyme. The Plant Journal: For Cell and Molecular Biology. 15 (3), 357-365 (1998).
- Kotting, O., et al. Identification of a novel enzyme required for starch metabolism in Arabidopsis leaves. The phosphoglucan, water dikinase. Plant Physiology. 137 (1), 242-252 (2005).
- Tagliabracci, V. S., et al. Laforin is a glycogen phosphatase, deficiency of which leads to elevated phosphorylation of glycogen in vivo. Proceedings of the National Academy of Sciences. 104 (49), 19262-19266 (2007).
- Gentry, M. S., Guinovart, J. J., Minassian, B. A., Roach, P. J., Serratosa, J. M. Lafora disease offers a unique window into neuronal glycogen metabolism. The Journal of Biological Chemistry. 293 (19), 7117-7125 (2018).
- Brewer, M. K., et al. Targeting pathogenic lafora bodies in lafora disease using an antibody-enzyme fusion. Cell Metabolism. 30 (4), 689-705 (2019).
- Santelia, D., Zeeman, S. C. Progress in Arabidopsis starch research and potential biotechnological applications. Current Opinion in Biotechnology. 22 (2), 271-280 (2011).
- Raththagala, M., et al. Structural mechanism of laforin function in glycogen dephosphorylation and lafora disease. Molecular Cell. 57 (2), 261-272 (2015).
- Meekins, D. A., et al. Phosphoglucan-bound structure of starch phosphatase Starch Excess4 reveals the mechanism for C6 specificity. Proceedings of the National Academy of Sciences. 111 (20), 7272-7277 (2014).
- Vander Kooi, C. W., et al. Structural basis for the glucan phosphatase activity of Starch Excess4. Proceedings of the National Academy of Sciences. 107 (35), 15379-15384 (2010).
- Meekins, D. A., et al. Structure of the Arabidopsis glucan phosphatase like sex four2 reveals a unique mechanism for starch dephosphorylation. The Plant Cell. 25 (6), 2302-2314 (2013).
- Smith, A. M., Zeeman, S. C. Starch: A flexible, adaptable carbon store coupled to plant growth. Annual Review of Plant Biology. 71, 217-245 (2020).
- Jane, J., Kasemuwan, T., Chen, J. F., Juliano, B. O. Phosphorus in rice and other starches. Cereal Foods World. 41 (11), 827-832 (1996).
- Mak, C. A., et al. Cooperative kinetics of the glucan phosphatase starch excess4. Biochemistry. 60 (31), 2425-2435 (2021).
- Campbell, K. P., MacLennan, D. H. Purification and characterization of the 53,000-dalton glycoprotein from the sarcoplasmic reticulum. The Journal of Biological Chemistry. 256 (9), 4626-4632 (1981).
- Campbell, K. P., MacLennan, D. H., Jorgensen, A. O., Mintzer, M. C. Purification and characterization of calsequestrin from canine cardiac sarcoplasmic reticulum and identification of the 53,000 dalton glycoprotein. The Journal of Biological Chemistry. 258 (2), 1197-1204 (1983).
- Davey, M. W., Sulkowski, E., Carter, W. A. Binding of human fibroblast interferon to concanavalin A-agarose. Involvement of carbohydrate recognition and hydrophobic interaction. Biochemistry. 15 (3), 704-713 (1976).
- Meekins, D. A., et al. Mechanistic insights into glucan phosphatase activity against polyglucan substrates. The Journal of Biological Chemistry. 290 (38), 23361-23370 (2015).
- Wilkens, C., et al. Plant α-glucan phosphatases SEX4 and LSF2 display different affinity for amylopectin and amylose. FEBS Letters. 590 (1), 118-128 (2016).
- Atanasova, M., Bagdonas, H., Agirre, J. Structural glycobiology in the age of electron cryo-microscopy. Current Opinion in Structural Biology. 62, 70-78 (2020).
- Doyle, M. L. Characterization of binding interactions by isothermal titration calorimetry. Current Opinion in Biotechnology. 8 (1), 31-35 (1997).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。