
Method Article
Injektionen von AAV-Vektoren für die Optogenetik in anästhesiertem und wach verhaltendem nicht-menschlichem Primatengehirn
In diesem Artikel
Zusammenfassung
Wie derzeit implementiert, erfordert die Optogenetik bei nichtmenschlichen Primaten die Injektion viraler Vektoren in das Gehirn. Eine optimale Injektionsmethode sollte zuverlässig sein und für viele Anwendungen in der Lage sein, einzelne Stellen beliebiger Tiefe anzuvisieren, die in der postmortalen Histologie leicht und eindeutig identifiziert werden können. Ein Injektionsverfahren mit diesen Eigenschaften wird vorgestellt.
Zusammenfassung
Optogenetische Techniken haben die neurowissenschaftliche Forschung revolutioniert und sind bereit, dasselbe für die neurologische Gentherapie zu tun. Der klinische Einsatz der Optogenetik erfordert jedoch, dass Sicherheit und Wirksamkeit in Tiermodellen nachgewiesen werden, idealerweise in nicht-menschlichen Primaten (NHPs), aufgrund ihrer neurologischen Ähnlichkeit mit dem Menschen. Die Anzahl der Kandidatenvektoren, die potenziell für die Neurowissenschaften und die Medizin nützlich sind, ist enorm, und es gibt noch keine Hochdurchsatzmittel, um diese Vektoren zu testen. Daher besteht ein Bedarf an Techniken, um mehrere räumlich und volumetrisch genaue Injektionen von viralen Vektoren in das NHP-Gehirn durchzuführen, die durch postmortale Histologie eindeutig identifiziert werden können. Hierin wird ein solches Verfahren beschrieben. Injektionskanülen bestehen aus gekoppelten Polytetrafluorethylen- und Edelstahlrohren. Diese Kanülen sind autoklavierbar, Einwegkanülen und haben ein geringes Minimalbelastungsvolumen, was sie ideal für die Injektion teurer, hochkonzentrierter viraler Vektorlösungen macht. Ein inertes, rot gefärbtes Mineralöl füllt den Totraum und bildet mit der Vektorlösung einen sichtbaren Meniskus, der eine sofortige und genaue Messung von Injektionsraten und -volumina ermöglicht. Das Öl wird in die Rückseite der Kanüle geladen, wodurch das Risiko einer Co-Injektion mit dem Vektor verringert wird. Kanülen können in 10 Minuten geladen werden, und Injektionen können in 20 Minuten durchgeführt werden. Dieses Verfahren eignet sich gut für Injektionen in wache oder betäubte Tiere. Wenn es verwendet wird, um qualitativ hochwertige virale Vektoren zu liefern, kann dieses Verfahren eine robuste Expression optogenetischer Proteine erzeugen, was eine optische Kontrolle der neuronalen Aktivität und des Verhaltens in NHPs ermöglicht.
Einleitung
Die Optogenetik bei nicht-menschlichen Primaten (NHPs) beinhaltet typischerweise die Injektion von viralen Vektoren direkt in das Gehirn. Eine Klasse von Vektoren, die für diese Anwendung gut geeignet ist, basiert auf Adeno-assoziierten Viren (AAV). Diese Vektoren bestehen aus einem Proteinkapsid, das ein einzelsträngiges DNA-Genom umgibt, das wiederum aus einem Promotor, einem Opsin-Gen und gegebenenfalls anderen proteinkodierenden und genregulatorischen Elementen besteht. Fortschritte beim molekularen Klonen haben die Manipulation und Kombination dieser Komponenten für die Entwicklung neuer Vektoren erleichtert. Folglich ist die Sammlung von AAV-Vektoren, die für die NHP-Optogenetik potenziell nützlich ist, groß und wächst schnell.
Derzeit erfordert der Nutzen eines AAV-Vektors für die NHP-Optogenetik Tests in vivo. Diese Tatsache ist ein wesentliches Hindernis für den Fortschritt. Tiere müssen sparsam verwendet werden, und das Testen mehrerer Vektoren in einem einzigen Tier erfordert, dass die Injektionsstellen in Bezug auf die neuronale Architektur vernünftig positioniert und in Bezug auf die Virusausbreitung gut getrennt werden. Eine genaue histologische Beurteilung erfordert, dass Injektionen räumlich und volumetrisch genau sind. Eine Injektionstechnik, die zuvor für die fokale Verabreichung von pharmakologischen Wirkstoffen 1,2,3,4 verwendet wurde, wurde angepasst und vereinfacht, um solche Injektionen durchzuführen. Diese Injektionstechnik ist kostengünstig, verwendet sterilisierbare Einwegkomponenten, kann bei betäubten oder wach verhaltenden Affen verwendet werden und kann verwendet werden, um verschiedene Gehirnbereiche jeder Tiefe anzusprechen. Das folgende Protokoll beschreibt Schritt-für-Schritt-Verfahren zur Herstellung der Einwegkomponenten und zur Durchführung von Injektionen in das NHP-Gehirn. Die Vor- und Nachteile der Technik werden diskutiert.
Protokoll
Alle Experimente wurden in Übereinstimmung mit dem Leitfaden für die Pflege und Verwendung von Labortieren durchgeführt und übertrafen die vom Institute of Laboratory Animal Resources und der Association for Assessment and Accreditation of Laboratory Animal Care International empfohlenen Mindestanforderungen. Alle Verfahren wurden vom Animal Care and Use Committee der University of Washington bewertet und genehmigt (UW IACUC Protokoll #4167-01). Fünf gesunde Makaken (2 Macaca mulatta, 3 Macaca nemestrina; männlich, 4-11 Jahre alt) nahmen an dieser Studie teil. Bei allen chirurgischen Eingriffen wurden sterile Instrumente und Techniken verwendet.
1. Herstellung einer Kanüle (Abbildung 1A)
- Vorbereitung jedes Teils
- Stumpfen Sie die Spitze einer Injektionsnadel (30 G, 13 mm Länge) mit einer Scheibenschleifmaschine.
- Schneiden Sie ein Edelstahlrohr (30 G, Innendurchmesser = 0,16 mm, Außendurchmesser = 0,31 mm) auf eine Länge, die auf die Tiefe des Zielhirnbereichs abgestimmt ist (25 mm ist gut geeignet, um die dorsale Oberfläche der Großhirnrinde zu injizieren). Mit einer Scheibenschleifmaschine ein Ende des geschnittenen Rohrs abschrägen und das andere glatt streichen. Entgraten Sie das Innere des Rohres mit einer Räumung.
- Schneiden Sie den Schlauch aus Polytetrafluorethylen (PFTE) (Innendurchmesser = 0,23 mm ± 0,02 mm, Wand = 0,23 mm ± 0,02 mm, 1 mm entspricht 42 nL ± 7 nL Flüssigkeit) auf eine Länge, die der Menge der zu ladenden Vektorlösung entspricht (1 μL Vektorlösung belegt 24 mm Schlauch). Flackern Sie beide Enden des PTFE-Röhrchens durch Einführen der abgestumpften Injektionsnadel.
- Führen Sie die abgestumpfte Injektionsnadel ca. 5 mm in ein Ende des PTFE-Röhrchens ein. Führen Sie das abgeschrägte Ende des Edelstahlrohrs ca. 5 mm in das andere Ende ein (Abbildung 1A).
- Führen Sie Tests vor der Injektion durch. Injizieren Sie gefiltertes Wasser durch die Injektionsnadel der Kanüle. Vergewissern Sie sich, dass Wasser aus dem abgeschrägten Edelstahlrohr von der Spitze austritt und dass aus keiner der beiden Verbindungen Wasser austritt.
2. Injektionsverfahren für betäubte Tiere
- Chirurgische Vorbereitung
- Sterilisieren Sie chirurgische Instrumente und Verbrauchsmaterialien mit den Verfahren in der Materialtabelle.
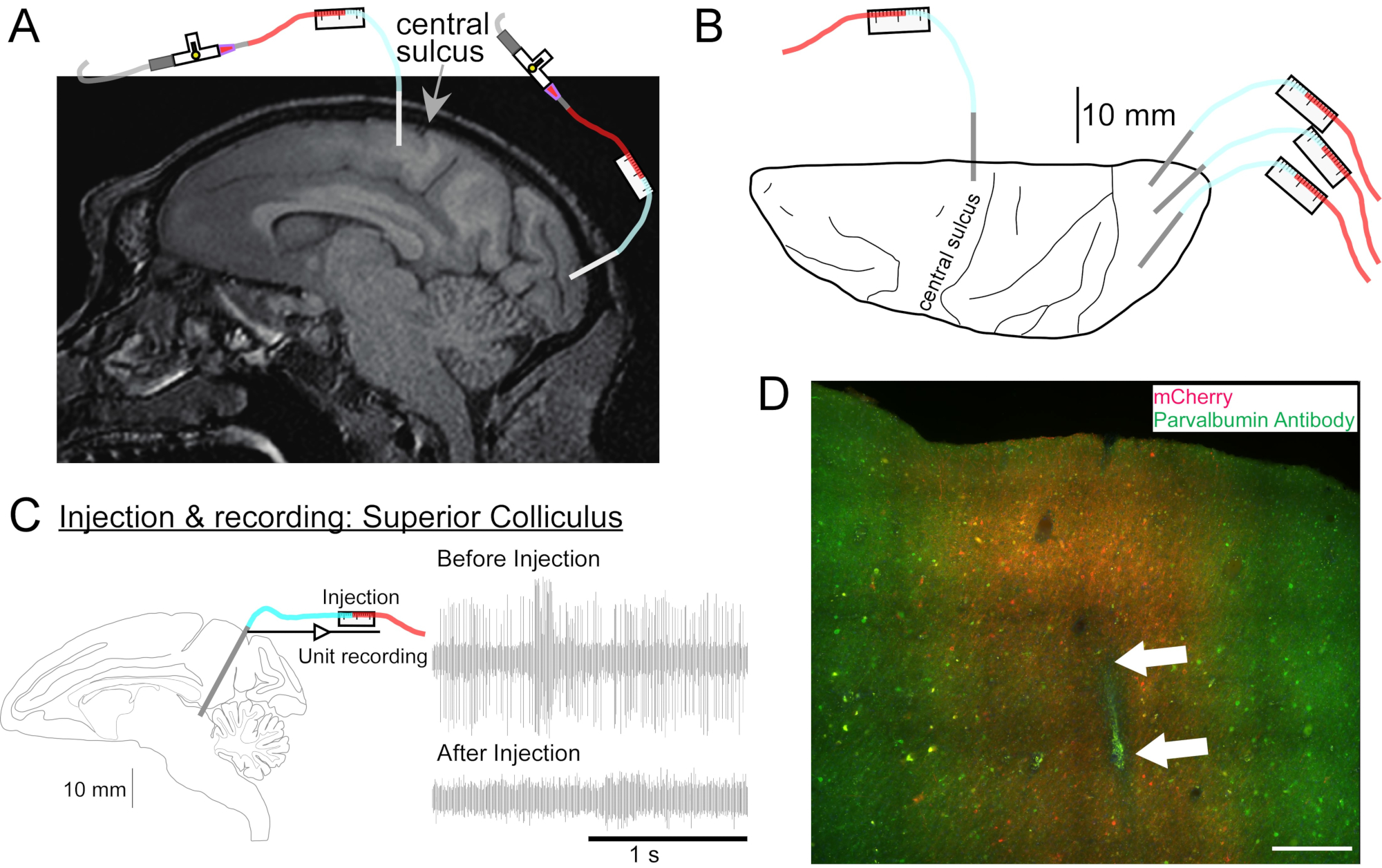
- Führen Sie bei Bedarf eine Kopf-MRT durch, um tiefe Hirnstrukturen anzuvisieren (Abbildung 2A).
- Unmittelbar vor der Operation die Tiere mit Ketamin (10-15 mg/kg) sedieren und Antibiotika (Cefazolin) und Analgetika (Retard-Buprenorphin und Meloxicam) intramuskulär verabreichen. Dann verabreichen Sie Propofol über einen intravenösen (IV) Katheter in die Venen saphena oder cephalic.
- Intubieren Sie das Tier und wandeln Sie es in Isoflurangas um. Bestätigen Sie die richtige Anästhesie durch stabile Herzfrequenz, Blutdruck, Atemfrequenz, entspannte Skelettmuskulatur und das Fehlen von Lid- oder Entzugsreflexen.
- Rasieren Sie den Kopf des Tieres. Tragen Sie künstliche Tränensalbe auf die Hornhaut auf, um ein Austrocknen zu verhindern.
- Vorbereitung des Injektionsbereichs
- Legen Sie den Kopf des Tieres in den stereotaktischen Rahmen. Tragen Sie die chirurgische Peelinglösung mit Mullschwämmen auf die rasierte Haut auf, üben Sie leichten Druck aus, um Ablagerungen zu lösen, und spülen Sie sie mit Isopropylalkohol ab. Wiederholen Sie diesen Vorgang dreimal. Bedecken Sie das Tier mit einem sterilen fenestrierten Vorhang. Schneiden Sie die Haut ein und reflektieren Sie den Muskel. Platzieren Sie den Manipulator auf dem stereotaktischen Arm, positionieren Sie ihn so, dass er auf den Zielhirnbereich zielt, und markieren Sie die Kraniotomieposition auf dem Schädel mit einem sterilisierten Stift.
- Entfernen Sie den stereotaktischen Manipulator und führen Sie die Kraniotomie durch. Schneiden Sie die Dura auf Wunsch ein (z. B. um sulkale Wahrzeichen zu visualisieren). Bringen Sie den Manipulator wieder in den stereotaktischen Arm zurück.
- Laden der Vektorlösung (Abbildung 1C)
- Übertragen Sie die Vektorlösung vorsichtig in ein sterilisiertes PCR-Röhrchen mit einem P20-Pipettor, um Blasen zu vermeiden.
- Befestigen Sie eine Kanüle mit der abgeschrägten Spitze nach unten an einem vertikal ausgerichteten stereotaktischen Halter. Verbinden Sie eine 1-ml-Luer-lock-Spritze mit der Injektionsnadelnadel der Kanüle.
- Tauchen Sie die abgeschrägte Spitze der Kanüle in die Vektorlösung.
HINWEIS: Die Spritze sollte bereits angebracht sein; Das Anhängen an dieser Stelle würde Blasen in die Vektorlösung einführen. - Laden Sie die Lösung in die Kanüle, indem Sie mit der 1-ml-Spritze leichten Unterdruck anlegen. Verfolgen Sie den Meniskus visuell zwischen der Lösung und der Luft.
- Sobald die Vektorlösung geladen wurde, setzen Sie den leichten Unterdruck fort, bis die Lösung die Nadelnabe erreicht. Entfernen Sie die 1-ml-Spritze und injizieren Sie das farbige Mineralöl in die Nadelnadel.
HINWEIS: Das Öl sollte langsam entlang der Innenwand der Nadelnabe injiziert werden, um mit der Vektorlösung einen klar geschnittenen Meniskus zu bilden und Luftblasen zu vermeiden. - Befestigen Sie die Injektionsnadelnabe an einem der beiden offenen Anschlüsse eines 3-Wege-Luer-Lock-Absperrhahns.
HINWEIS: Wenn Sie die Injektionsnadel am geschlossenen Anschluss befestigen, entsteht unerwünschter Luftdruck hinter dem Öl. - Schließen Sie den Anschluss, der mit der Nadelspitze verbunden ist, füllen Sie eine 1-ml-Spritze mit Luft und befestigen Sie sie an einem der beiden anderen Anschlüsse. Schließen Sie schließlich die verbleibende Spitze des Absperrhahns, um die Spritze mit der Kanüle zu verbinden.
- Drücken Sie langsam Luft in die Kanüle. Sobald das farbige Öl an der Spitze der stumpfen Nadel im PTFE-Schlauch erscheint, prüfen Sie, ob Luft zwischen der Lösung und dem farbigen Öl vorhanden ist.
- Wenn Luft vorhanden ist, üben Sie Unterdruck auf die Spritze aus, um das farbige Öl in die Nadelnabe zurückzuführen. Entfernen Sie die Blase und üben Sie Überdruck aus, bis ein Tropfen Vektorlösung an der abgeschrägten Kanülenspitze sichtbar ist.
- Entfernen Sie die 1-ml-Spritze, um unerwünschten Luftdruck hinter dem Öl freizusetzen, und schließen Sie den Absperrhahn, um zu verhindern, dass der Vektor durch die Schwerkraft aus der Kanüle austritt.
- Kleben Sie ein Kunststofflineal an den PTFE-Schlauch, um die Bewegung des Meniskus während der Injektion zu messen (Abbildung 1B,D,F).
- Einführen von Kanülen in den Zielhirnbereich (Abbildung 1B)
- Befestigen Sie die Kanüle am stereotaktischen Manipulator.
- Übertragen Sie den Pumpenschlauch (der in einem Luer-Lock-Anschluss endet) manuell vom nicht sterilen Assistenten zum Chirurgen. Der Chirurg sollte den Luer-Lock-Konnektor durch die Wand einer sterilen Hülse greifen, einen zweiten sterilen Absperrhahn am Konnektor befestigen, die Hülse fest darum kleben und dann den Kragen der Hülse fallen lassen, damit er sich durch die Schwerkraft entlang des Röhrchens erstrecken kann.
- Verbinden Sie den am Kanülenschlauch befestigten Absperrhahn mit dem an der Luftpumpe befestigten Absperrhahn. Stellen Sie die Luftpumpe auf niedrigen Druck, schalten Sie sie ein und erhöhen Sie den Druck, bis das Öl durch die Kanüle vordringt und ein Tropfen Vektorlösung an der Kanülenspitze sichtbar ist.
- Stellen Sie die Position des Kunststofflineals am PTFE-Schlauch ein, um die Bewegung des Meniskus während der Injektion zu messen.
- Fahren Sie die Kanüle mit dem stereotaktischen Manipulator nach unten und notieren Sie die Tiefe, in der die Spitze an die Oberfläche reicht (Dura oder Pia mater).
- Fahren Sie die Kanüle zur tiefsten Stelle, um entlang der Spur injiziert zu werden. Die Oberfläche wird grübchen. Wenn Sie Oberflächenkortex injizieren, bestätigen Sie visuell, dass die Kanüle in die Oberfläche eingedrungen ist, mit einem Operationsmikroskop oder Lupenbrillen, falls verfügbar.
- Um Fehlzielen aufgrund von Gewebekompression zu minimieren, fahren Sie die Kanüle langsam (1 mm/min), schnell (0,5 mm/s) mit 1-5 Minuten Wartezeit am Boden ein oder schießen Sie die tiefste Injektionsstelle um 500 μm über und ziehen Sie sie dann zurück.
- Injektion
- Injizieren Sie 0,5 μL der Vektorlösung mit der elektrischen Luftpumpe über 10-30 s. Bestätigen Sie den Injektionsfluss, indem Sie den Meniskus zwischen dem farbigen Öl und der Vektorlösung im PTFE-Röhrchen verfolgen.
- Warten Sie 1 min und ziehen Sie die Kanüle zur nächsten Injektionsstelle entlang der Spur zurück.
- Lassen Sie die Kanüle nach der letzten Injektion 10 Minuten an Ort und Stelle, um einen Vektorausfluss zu vermeiden.
- Ziehen Sie die Kanüle zurück und entsorgen Sie sie in einem Behälter für scharfe Biogefahren.
- Optional ist eine kleine Menge (≤1 μL) fluoreszierende Mikroperlen in der Nähe der Vektorinjektionsstelle zu injizieren, um die Identifizierung der Injektionsstelle post mortem zu erleichtern.
- Wiederholen Sie diesen Vorgang wie gewünscht für die anderen Vektorlösungen an anderen Stellen (Abbildung 2B).
- Schließung der Operation
- Nähen Sie die Dura, den Muskel und die Haut.
- Entfernen Sie den Affen aus dem stereotaktischen Rahmen, und entfernen Sie alle Monitorkabel.
- Entfernen Sie den Affen aus der Isoflurananästhesie und extubieren Sie nach der Rückkehr des Schluckreflexes.
- Führen Sie eine postoperative Behandlung durch (3-5 Tage Meloxicam und 7-10 Tage Cephalexin). Überwachen Sie das Tier mindestens einmal alle 10 Minuten, bis es in der Lage ist, eine stabile aufrechte Sitzposition beizubehalten.
3. Chirurgie und AAV-Vektorinjektion für wach verhaltende Tiere (Abbildung 1D)
HINWEIS: Eine Variante der Technik kann verwendet werden, um Injektionen in das Gehirn von wachen, sich verhaltenden Affen zu machen, wie unten beschrieben.
- Simultane Injektion mit Aufzeichnung
- Um die elektrische Aktivität an der Injektionsstelle aufzuzeichnen, beschichten Sie die Außenseite der Kanüle mit Epoxidharz (Boden ~15 mm) und Polyimidschlauch (Restlänge). Legen Sie das Metall an der Spitze frei, indem Sie das Epoxidharz davon abkratzen (Injectrode, Abbildung 1F)2. Alternativ können Sie die Kanüle und eine separate extrazelluläre Elektrode nebeneinander in ein doppelläufiges Führungsrohr einführen (Doppelzylinder-Führungsrohr, Abbildung 1E).
- Kanüleneinführung in den Zielhirnbereich.
- Setzen Sie den Affen in die Versuchskabine, schränken Sie die Bewegung des Kopfes ein und reinigen Sie die am Schädel montierte Aufnahmekammer mit Standardtechniken.
- Befestigen Sie ein Führungsrohr am Mikrolaufwerk. Führen Sie die Injektionskanüle in das Führungsrohr ein.
- Schieben Sie die Kanüle vor, bis die Spitze aus dem Führungsrohr herausragt.
- Schließen Sie den Absperrhahn an die elektrische Luftpumpe an. Um die ordnungsgemäße Systemfunktion zu bestätigen, drücken Sie einen Tropfen Vektorlösung mit der Luftpumpe von der Spitze und bestätigen Sie die Bewegung des Öl-Vektor-Lösungsmeniskus.
- Ziehen Sie die Kanüle ~5 mm in das Führungsrohr, um sie vor Beschädigungen während des Einführens des Röhrchens in das Gehirn zu schützen. Führen Sie das Führungsrohr in das Gehirn ein.
- Fahren Sie die Kanüle mit dem Mikrolaufwerk zur zu injizierenden Stelle. Identifizieren Sie die Zielstelle entweder durch elektrische Aufzeichnung (Abbildung 2C) oder Stimulation.

Abbildung 1: Aufbau von Chirurgie und Apparatur . (A) Injektionskanüle. Jeder Teil der Kanüle ist angegeben. Unten rechts eingefügt: vergrößertes Bild der Kanülenspitze, Maßstabsleiste: 500 μm. (B) Operationsaufbau für anästhesierte Affen. Der Affe wird in einem stereotaktischen Rahmen unter einem chirurgischen Vorhang platziert. Beatmungsgerät (V), intravenöse Leitung (IV), Blutdruckmessgerät (BP) und Sauerstoffsättigungsmonitor (O2) sind mit dem Affen verbunden. Die Injektionskanüle wird mit einem stereotaktischen Mikromanipulator in den Zielbereich eingeführt. Die Vektorlösung wird von einer elektrischen Luftpumpe (unten links eingesetzt, braun) eingespritzt, die an einen Luftkompressor gekoppelt ist (unten links eingesetzt, blau). Ein Kunststofflineal (oberer Einschub) wird auf den PTFE-Schlauch geklebt, um die Bewegung des Meniskus zwischen farbigem Öl (oberer Einschub, rot) und Vektorlösung (oberer Einschub, klar) während der Injektion zu messen. (C) Einrichtung zum Laden der Vektorlösung in die Kanüle. (D) Ein Affe während einer Injektion von Vektorlösung in einer Versuchskabine. Der Kopf des Tieres wird von drei Stabilisierungspfosten gehalten, und die Augenposition wird durch ein Skleralsuchspulensystem aufgezeichnet. Die Injektionskanüle wird gehalten und mit einem Mikroelektrodenhalter/-treiber in die Zieltiefe getrieben. Die Injektion wird gesteuert, indem der Meniskus zwischen dem farbigen Öl und der Vektorlösung über eine USB-Kamera überwacht wird (Einschubbild). (E) Doppelrohreinspritzung. Ein doppelläufiger Führungsrohrhalter/-treiber hält eine Injektionskanüle und eine Mikroelektrode (siehe Einschub). (F) Injizieren. Das Metall an der Spitze der Kanüle, das durch Abkratzen der Epoxidschicht freigelegt wird, ermöglicht den elektrischen Zugang zu Neuronen (Einschub, Maßstabsbalken: 500 μm). (G) Einrichtung der Laserstimulation. Ein doppelläufiger Führungsrohrhalter/-treiber hält sowohl eine optische Faser als auch eine Mikroelektrode (siehe Einschub). Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
{kind=link}

Abbildung 2: Diagramm der AAV-Injektionsstellen . (A) Sagittaler Schnitt des Gehirns MR-Bild mit Injektionsstellen im primären motorischen Kortex und primären visuellen Kortex einer Macaca-Nemestrina. (B) Ansicht von der dorsalen Oberfläche auf der entsprechenden Atlasplatte, die die Kanülenplatzierung relativ zum zentralen Sulcus (primärer motorischer Kortex) und zum primären visuellen Kortex zeigt. (C) Einheitenaufzeichnung durch Injektrode im Colliculus superior. Eine Einheit, die vor der Injektion isoliert wurde (rechts oben), verschwand nach der Injektion (rechts unten). (D) Einspritzspur (weiße Pfeile). Maßstabsleiste: 500 μm. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
{kind=link}
4. Hirngewebeaufbereitung für die Histologie
- Warten Sie 6-8 Wochen nach der Injektion, um die Transgenexpression zu maximieren.
HINWEIS: Die optimale Dauer hängt vom genauen viralen Vektor ab, der im Experiment verwendet wird. - Verarbeiten Sie das Gehirn mit herkömmlichen histologischen Techniken, um die Transduktionseffizienz und Selektivität zu beurteilen 5,6,7.
Ergebnisse
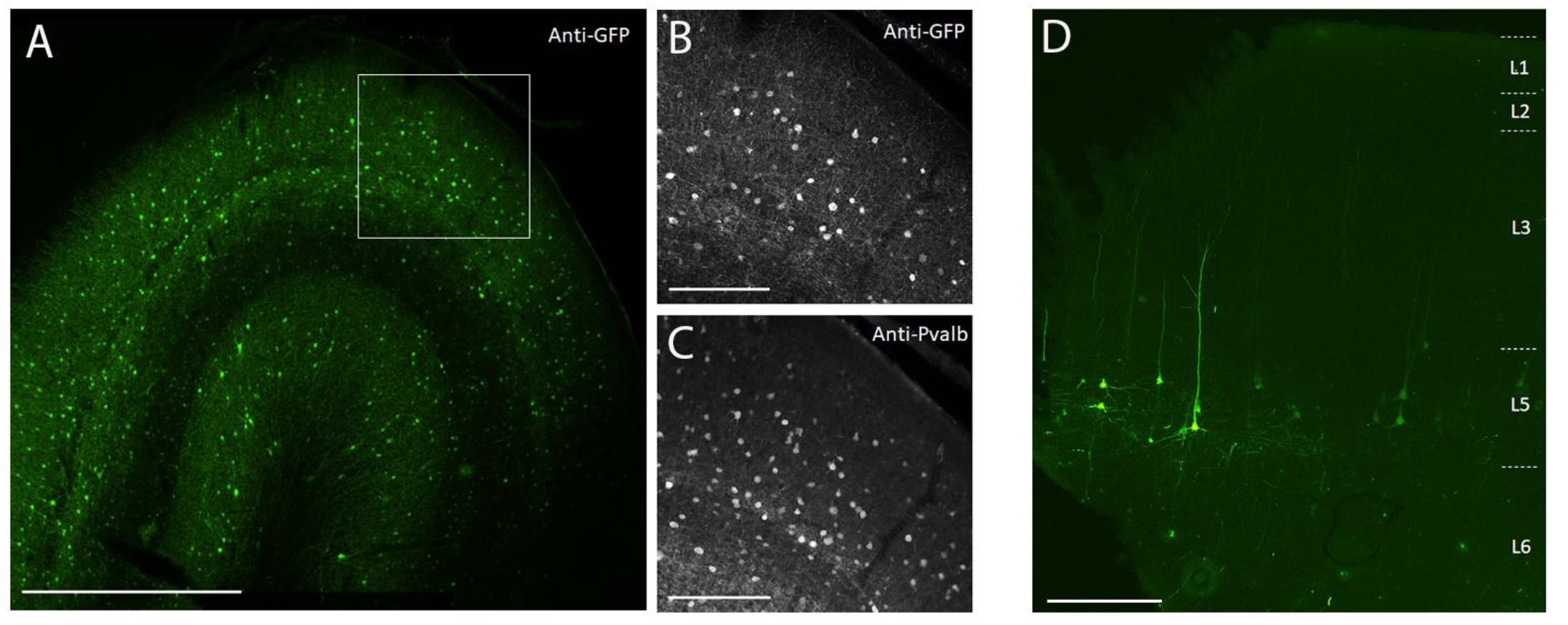
Um die Transgenexpression durch in vivo stereotaktische Injektion in das NHP-Gehirn unter Verwendung der hier beschriebenen chirurgischen Injektionsmethode zu demonstrieren, wurden zwei Vektoren ausgewählt, die Enhancer enthielten, die die Expression des supergelb fluoreszierenden Protein-2 (SYFP2) in verschiedenen neuronalen Typen 8,9 antreiben. Virale Vektoren wurden in das PHP.eB-Kapsid10 verpackt, durch Iodixanol-Zentrifugation gereinigt und dann auf hohen Titer (>1E13 virale Genome/ml) konzentriert, gemessen mittels qPCR. Ein Volumen von 0,5 μL wurde in jeder der zehn Tiefen entlang von zehn Spuren durch den Kortex injiziert, was ein Gesamtinjektionsvolumen von 5 μL / Spur ergibt. Abbildung 3A-C zeigt die SYFP2-Expression durch Anti-GFP-Immunfärbung 113 Tage nach Injektion des PVALB-subklassenspezifischen AAV-Vektors CN2045 in den primären visuellen Kortex einer erwachsenen männlichen Macaca-Nemestrina. Das SYFP2-Transgen wird robust in zahlreichen nicht-pyramidalen Neuronen nachgewiesen, die über die kortikale Tiefe verstreut sind, und die meisten SYFP2-exprimierenden Neuronen waren auch immunreaktiv für PVALB7. Abbildung 3D zeigt die native SYFP2-Expression im primären motorischen Kortex 64 Tage nach der Injektion des L5-Neuronen-Subklassen-spezifischen AAV-Vektors CN2251. Die SYFP2-markierten Neuronen haben alle eine klare pyramidale Morphologie mit Somata, die auf Schicht 5 beschränkt sind, und charakteristischen dicken apikalen Dendriten. Diese Daten zeigen eindeutig die präzise Kontrolle der Transgenexpression in ausgewählten Populationen neokortikaler Neuronen im NHP-Gehirn durch stereotaktische Injektion von zellartigen AAV-Vektoren.

Abbildung 3: Beispiel einer zelltypspezifischen SYFP2-Expression, die durch AAV-Vektoren vermittelt wird, die in das NHP-Gehirn injiziert wurden . (A) Epifluoreszenz-Mikroaufnahme eines festen Schnitts aus dem primären visuellen Kortex der Makaken 113 Tage nach Injektion eines PVALB-Subklassen-spezifischen AAV-Vektors. Maßstabsstab: 1 mm. (B,C) Bild mit höherer Vergrößerung des eingerahmten Bereichs in A. (B) Anti-GFP-Signal. (C) Anti-PVALB-Signal. Maßstabsbalken: 250 μm. (D) Epifluoreszenz-Mikroaufnahme der nativen SYFP2-Fluoreszenz in einem festen Schnitt aus dem primärmotorischen Kortex der Makaken 64 Tage nach Injektion eines extratelenzephalen subklassenspezifischen AAV-Vektors der Schicht 5. Maßstabsleiste: 500 μm. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
{kind=link}
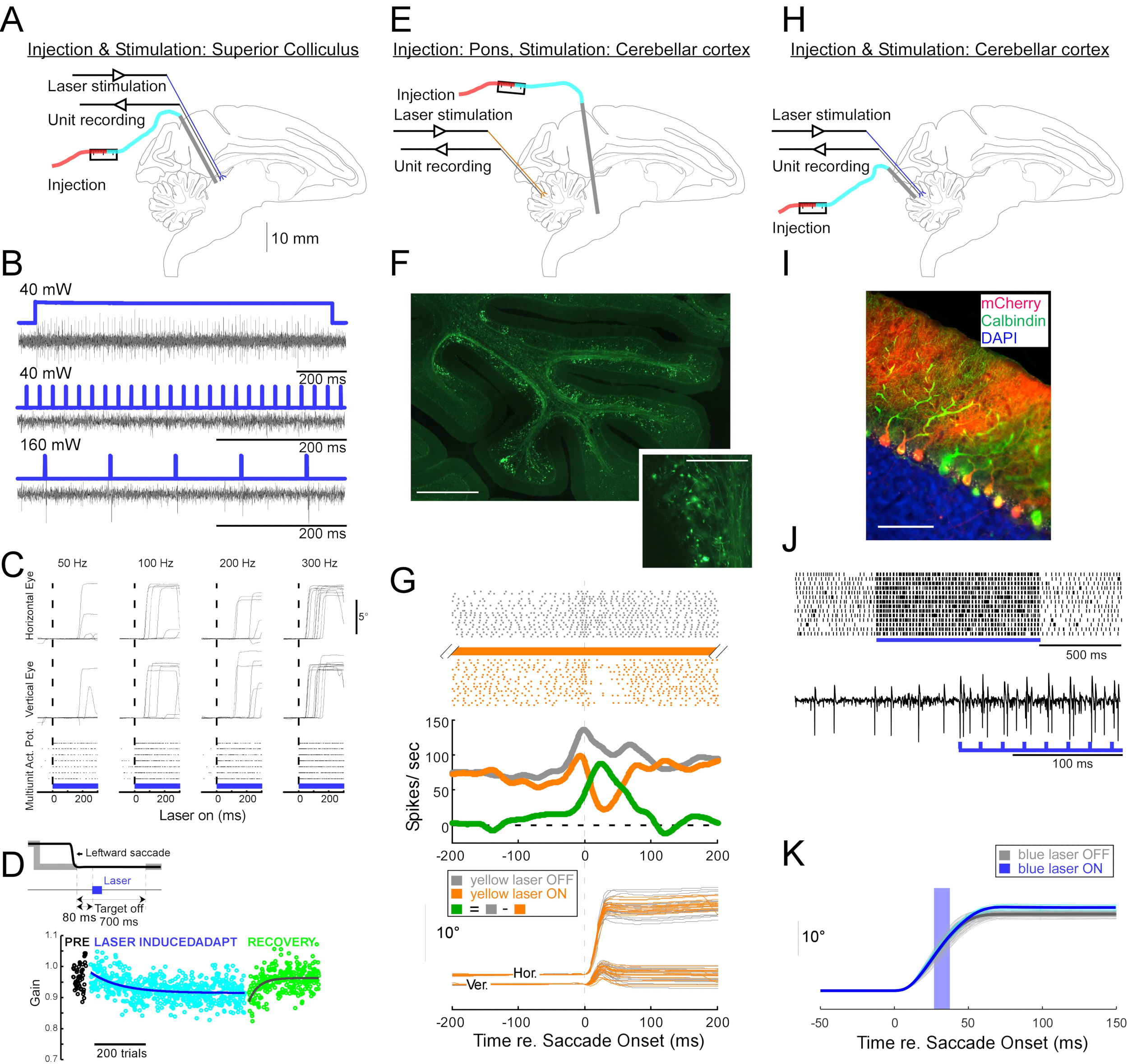
Um den Nutzen dieser Injektionstechnik für neurophysiologische und verhaltensbezogene optogenetische Manipulationen zu demonstrieren, wurden drei Experimente an jeweils einem anderen Affen (Macaca mulatta) durchgeführt. Im ersten Experiment (Abbildung 4A-D) wurden AAV-Vektoren, die das Kanalrhodopsin-2-Transgen (AAV1-hSyn1-ChR2-mCherry) tragen, in den linken Colliculus superior (SC) injiziert. Der Vektor wurde alle 250 μm in 19 Tiefen für insgesamt 12 μL injiziert. Im zweiten Experiment (Abbildung 4E-G) wurden 3 μL AAV1-hSyn-ArchT-EYFP-Lösung in den Nucleus reticularis tegmenti pontis (NRTP) injiziert. Im dritten Experiment (Abbildung 4H-K) wurden 24 μL AAV9-L7-ChR2-mCherry-Lösung in die Kleinhirnrindeinjiziert 6. Sechs bis acht Wochen nach jeder Injektion wurden eine optische Faser und eine Wolframelektrode über ein doppelläufiges Führungsrohr in das Gehirn eingeführt (Abbildung 1G).
Abbildung 4B zeigt die Reaktion eines SC-Neurons auf blaues Licht (450 nm). Dauerlicht (1,2 s) bei 40 mW erzeugte eine Reihe aufeinanderfolgender Aktionspotentiale (Abbildung 4B, oben). Lichtimpulse von 1 ms Dauer konnten bei 40 mW keine Aktionspotentiale hervorrufen (Abbildung 4B, Mitte), aber zuverlässig Aktionspotentiale bei 160 mW, dem einzigen anderen getesteten Leistungspegel, mit einer Latenz von 2,7 ± 0,6 ms (Abbildung 4B, unten). Eine Impulsfolge (160 mW, Frequenz: 300 Hz, Tastverhältnis: 15%, Dauer: 300 ms) evozierte Sakkaden konsistent mit einer durchschnittlichen Latenz von 97 ± 32 ms, einer mittleren Amplitude von 10,4° und einem mittleren Winkel von 47° (nach oben und rechts; Abbildung 4C).
In Übereinstimmung mit Studien, die die Sakkadenverstärkung durch subschwellige elektrische Stimulation des SC11,12 modifizierten, reduzierte die optische Stimulation des SC nach 15°, 18° und 20° links- und abwärts (225°) Sakkaden allmählich den Sakkadengewinn (Abbildung 4D). Diese Abnahme der Verstärkung erforderte ~250 Versuche (grüne Kreise), um zum Voranpassungsgewinn (schwarze Kreise) zurückzukehren, was seine Grundlage in der langfristigen Plastizität bestätigt.
Im zweiten Experiment (Abbildung 4E) wurde die moosige Faserprojektion vom NRTP zum okulomotorischen Vermis (OMV) der Kleinhirnrinde (Läppchen VIc und VII) optisch unterdrückt. Abbildung 4F zeigt fluoreszierend markierte moosartige Fasern und Rosetten in der OMV (Einschub). Gelbes Laserlicht (589 nm) wurde über optische Fasern an die OMV abgegeben, und eine nahe gelegene Wolframelektrode wurde verwendet, um die Aktivität der Purkinje-Zellen aufzuzeichnen. Abbildung 4G zeigt die einfache Spike-Aktivität vor (grau) und nach (orange) optogenetischer Inaktivierung von NRTP-Projektionen (Abbildung 4G, oben). Vor der Inaktivierung zeigte die Purkinje-Zelle ein Doppelburstmuster für 12° rechtsgerichtete Sakkaden (Abbildung 4G, Mitte, grau). Während der Inaktivierung nahm die Feuerrate ab und wechselte zu einem Burst-Pause-Muster (Abbildung 4G, Mitte, orange). Der Vergleich dieser beiden Antwortmuster deutet darauf hin, dass der moosige Fasereintrag in die Purkinje-Zellen die Verzögerungsphase der Sakkade beeinflusst, indem er den zweiten Ausbruch antreibt (Abbildung 4G, Mitte, grün). Die Variabilität der rechtsgerichteten Sakkaden wurde während der optogenetischen Inaktivierung reduziert, was mit der Idee übereinstimmt, dass ein Teil der Variabilität von Versuch zu Versuch in Sakkadenmetriken auf die Variabilität der Signale zurückzuführen ist, die von moosigen Fasern getragen werden (Abbildung 4G, unten, orange).
Im dritten Experiment (Abbildung 4H) wurden Purkinje-Zellen der OMV optogenetisch stimuliert (Abbildung 4I). Ein Zug kurzer Lichtpulse (1,5-ms-Impulse, 65 mW, 50 Hz) erhöhte die einfache Spike-Aktivität einer isolierten Purkinje-Zelle (Abbildung 4J, oben). Einzelne 1,5-ms-Lichtimpulse rufen häufig >1 einfache Spitze hervor (Abbildung 4J, unten). Optogenetische einfache Spike-Aktivierung, zeitlich abgestimmt während einer Sakkade (10-ms-Lichtpuls, 60 mW), erhöhte Sakkadenamplitude (Abbildung 4K), bestätigt die disinhibitorische Rolle von Purkinje-Zellen auf dem okulomotorischen Burst-Generator.

Abbildung 4: Zusammenfassung von drei optogenetischen Experimenten, die an wachen Affen durchgeführt wurden. (A-D) Experiment 1, panneuronale Erregung: virale Injektion, Laserstimulation und Einheitenaufzeichnung wurden im Colliculus superior (A) durchgeführt. (B) Repräsentative Einheitsaktivität, hervorgerufen durch Laserstimulation. (C), horizontale (oben) und vertikale (mittlere) Komponenten von Augenbewegungen und Rasterdiagramm der Einheitsaktivität (unten), hervorgerufen durch Laserstimulation. (D) Eine repräsentative Sitzung der Sakkadenanpassung, die durch Laserstimulation induziert wird. Die Stimulation (100 0,5-ms-Laserpulse) wurde 80 ms nach jeder Sakkade (Einschub) abgegeben. Der Sakkadengewinn (Sakkadenamplitude / Zielamplitude) nahm in den Studien allmählich ab. (E-G) Experiment 2, Signalweg-spezifische Hemmung: Ein viraler Vektor wurde in den Nucleus reticularis tegmenti pontis injiziert, und Laserstimulation und Einheitenaufzeichnung wurden im okulomotorischen Vermis (E) durchgeführt. (F) Histologischer Schnitt des okulomotorischen Wurms mit markierten moosigen Fasern (Maßstabsbalken: 1 mm) und ihren Rosetten (Einschub, Maßstabsbalken: 100 μm). (G) Purkinje-Zellaktivität (oben: Raster, Mitte: durchschnittliche Feuerrate) und Flugbahnen visuell geführter Sakkaden (unten) mit und ohne Laserstimulation. Grau: Laser-Off-Versuche, Orange: Laser auf Versuchen, Grün: Unterschied zwischen Grau und Orange. (H-K) Experiment 3, zelltypspezifische Aktivierung: Virusinjektion, Laserstimulation und Einheitenaufzeichnung wurden im okulomotorischen Vermis (H) durchgeführt. (I) Histologischer Schnitt des okulomotorischen Vermis mit markierten Purkinje-Zellen. Maßstabsbalken: 100 μm. (J) Einfache Spike-Aktivität einer Purkinje-Zelle, hervorgerufen durch Laserstimulation. Oben: Rasterplot aus 14 Versuchen. Unten: Spannungsleiter aus einem einzigen repräsentativen Versuch. (K) Trajektorien visuell geführter Sakkaden mit und ohne Laserstimulation. Ein 10-ms-Lichtimpuls während der Sakkaden erhöhte die Sakkadenamplituden. Individuelle Sakkadentrajektorien (Cyan) und ihr Durchschnitt (blau) bei Laser-On-Versuchen. Individuelle Sakkadentrajektorien (hellgrau) und ihr Durchschnitt (dunkelgrau) bei Laser-Off-Versuchen. Die Lichtwellenlänge betrug 450 nm in den Experimenten 1 und 3 und 589 nm in Experiment 2. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
{kind=link}
Diskussion
Fortschritte in der NHP-Optogenetik haben einen Bedarf an genauen, zuverlässigen intrakraniellen Injektionsmethoden geschaffen. Vorteile der in diesem Bericht beschriebenen Methode sind, dass sie kostengünstig ist, sterilisierbare und Einwegkomponenten verwendet und die Fähigkeit hat, verschiedene Gehirnbereiche jeder Tiefe anzusprechen. Es ermöglicht auch die Steuerung der Einspritzgeschwindigkeit und des Volumens aufgrund der Geschwindigkeit, mit der das Luftventil gesteuert werden kann. Der Luftdruck kann vorübergehend erhöht werden, um eine Verstopfung zu lösen, und dann schnell reduziert werden, um eine nachfolgende Überinjektion zu vermeiden, die durch anhaltenden Druck erzeugt würde. Einwegkomponenten reduzieren das Risiko einer Kreuzkontamination zwischen den Injektionsstellen.
Zu den kritischen Schritten in diesem Injektionsprotokoll gehören die Konstruktion hochwertiger Kanülen, deren Beladung ohne Einführung von Blasen und die Auswahl von Injektionsstellen, die nicht zu nahe beieinander liegen. Injektionen ≥ Abstand von 1 cm transduzieren normalerweise nicht überlappende Regionen, aber diese Heuristik ist abhängig von viralem Serotyp, Titer, Promotor, Volumen, Ziel und Nachweismethode. Die Auswahl von Injektionsstellen, die nicht direkt miteinander verbunden sind, vermeidet potenzielle Störfaktoren, die durch den Opsintransport entlang von Axonen und die Neigung einiger AAV-Serotypen zur retrograden Transduktion verursacht werden.
Die Methode kann verwendet werden, um NHPs zu injizieren, während sie betäubt und in einem stereotaktischen Rahmen (Abbildung 3) oder alarmiert und kopffixiert (Abbildung 4) sind. Ersteres hat den Vorteil, dass Injektionen gezielt in stereotaktischen Koordinaten durchgeführt werden können, und es ermöglicht eine visuelle Bestätigung der Kanülenpenetration durch eine akute Durotomie (das Einschneiden der Dura bei einem wachen Affen durch eine chronische Kraniotomie erhöht das Infektionsrisiko). Der letztere Ansatz hat den Vorteil, dass er die Anzahl der Überlebensoperationen und damit den Stress für das Tier reduziert, mit elektrophysiologischen Aufzeichnungen während des Verhaltens kompatibel ist und den gleichen Koordinatenrahmen und die gleichen Instrumente verwendet, die zum Einfügen optischer Fasern für Experimente nach der Injektion verwendet werden. Die Injektionstechnik bei wachen Affen könnte durch Injektionen durch künstliche Dura13,14,15 weiter verbessert werden. Dies würde die zusätzlichen Vorteile der direkten Visualisierung von Injektionsstellen und der Gewebefluoreszenz, die eine erfolgreiche Transduktion anzeigt, verleihen.
Mehrere andere AAV-Injektionstechniken wurden in NHPs verwendet. Vor kurzem wurde eine Mehrkanal-Injektionsvorrichtung entwickelt, um AAV-Vektoren gleichmäßig an große kortikale NHP-Regionen zu liefern16. Ähnliche Ergebnisse können mit konvektionsverstärkter Abgabe17,18 erzielt werden. Diese Methoden zielen darauf ab, die Transduktionsausbreitung zu maximieren, was ein wichtiges Ziel ist, das sich jedoch von der räumlichen Präzision unterscheidet, die unsere Methode erreichen soll.
Eine weitere alternative Methode besteht darin, AAV-Vektoren durch Borosilikatschläuche zu injizieren, die an einem Ende an einer scharfen Spitze abgeschrägt undam anderen Ende 5,6 an einer Hamiltonspritze befestigt sind. Diese Methode hat viel mit der in diesem Artikel beschriebenen Methode gemeinsam. Der virale Vektor wird in einer Länge von Schläuchen gehalten, der Raum im Schlauch hinter dem Virus ist mit gefärbtem Öl gefüllt, und der Fluss des Vektors wird über die Bewegung des Ölvektormeniskus überwacht. Diese alternative Technik erfordert weniger Ausrüstung und Vorbereitung, erfordert jedoch das Ziehen von Öl in das Borosilikatrohr durch die abgeschrägte Spitze durch Unterdruck und das anschließende Laden des Vektors über den gleichen Weg. Dies führt unweigerlich zu Spuren von Öl, das an das Gehirn abgegeben wird. Darüber hinaus muss der Borosilikatschlauch unserer Erfahrung nach einen Durchmesser von ~350 μm haben, um Dura auch bei abgeschrägter Abschrägung zu durchdringen und verursacht daher größere mechanische Schäden als die in diesem Artikel beschriebene dünnere Metallkanüle (Abbildung 2D). 30 G-Schläuche wurden verwendet, weil ihre kritische Knicklast hoch genug ist, um trotz einer Länge von 1-10 cm eine Dura zu vermitteln, weil sie fest in den PTFE-Schlauch passen und weil sie selten verstopft werden. 33 G Schläuche verstopft und biegt sich leichter und ist schwieriger mit dem PTFE-Schlauch zu verbinden. 36 G Schläuche sind nicht steif genug, um NHP dura mater zu durchdringen.
Eine weitere alternative Injektionstechnik besteht darin, den Ausgang der Luftpumpe mit einer Rückseite einer vektorgeladenen, gezogenen Glaspipette19 zu verbinden. Der Vektor wird durch direkten, intermittierenden Luftdruck von der Pumpe aus der Pipettenspitze gedrückt, wodurch Öl überflüssig wird. Ähnlich wie bei der oben erläuterten Einschlauchmethode verringert das Fehlen von Materialverbindungen zwischen Meniskus und Kanülenspitze das Risiko von Leckagen. Die scharfe Verjüngung und die empfindlichen Spitzen von Glaspipetten verhindern jedoch, dass sie NHP dura durchdringen oder auf tiefe Strukturen abzielen.
Offenlegungen
Die Autoren haben nichts offenzulegen.
Danksagungen
Diese Studie wurde von WaNPRC / ITHS P51OD010425 (JTT), National Institute of Health (NIH) Grants EY023277 (R01 für YK), EY030441 (R01 für GH), MH114126 (RF1 to JTT, Boaz Levi, Ed Lein), MH120095 (UG3 für JTT und GH), EY028902 (R01 für RS) unterstützt und durch NIH-Zuschüsse OD010425 (P51 für WaNPRC) und University of Washington Royalty Research Fund A148416 ermöglicht. Die Autoren danken Yasmine El-Shamayleh und Victoria Omstead für die Histologie, Refugio Martinez für das Klonen viraler Vektoren und John Mich für die Unterstützung bei der Verarbeitung von NHP-Hirngewebe.
Materialien
| Name | Company | Catalog Number | Comments |
| Equipment: Stereotaxic set | |||
| Item | |||
| Allen keys | BONDHUS | 10936 | STERRAD |
| Cannula holder | KOPF | 1770 | STERRAD |
| Carrier (manipulator) | KOPF | 1404 | STERRAD |
| Carrier platform | KOPF | 1430 | NA |
| Carrier stand | KOPF | 1449 | STERRAD |
| Eye, ear, mouth bars | KOPF | 1430 | NA |
| Stereotaxic base | KOPF | 1210 | NA |
| Equipment: Cannula | |||
| Item | |||
| 1 mL Luer-lock syringes | BD | 309628 | NA (sterilized package) |
| Cannulas* | (homemade - see below) | NA | steam (autoclave) |
| Colored oil** | (homemade - see below) | NA | NA |
| Elevator (for tube rack) | Cole-Parmer | UX-08057-10 | STERRAD |
| Filter tip | Genesee Scientific | 23-404 | NA (sterilized package) |
| Fluorescent microbeads | Lumafluor | R170 | NA |
| P20 pipetman | Gilson | FA10003M | NA |
| PCR tubes | Olympus Plastics | 22-161 | STERRAD |
| Stopcock | Cole-Parmer | 3060004 | STERRAD |
| Tube rack | homemade | NA | STERRAD |
| Vector solution | (homemade) | NA | NA |
| Equipment: Electric air pump set | |||
| Item | |||
| Electric air pump | World Precision Instruments | PV830 | NA |
| Foot pedal | World Precision Instruments | 3260 | NA |
| Tube cover | EZ Drape | A400-1000 | NA (sterilized package) |
| Equipment: General surgery tools | |||
| Item | |||
| Beaker | MEDLINE | azlon | STERRAD |
| Burrs | STRYKER | 277-10-235 | STERRAD |
| Double pronged tissue pick | Fine Science Tools | 18067-11 | STERRAD |
| Drapes | MEDLINE | DYNJP3004 | NA (sterilized package) |
| Dressing forceps | Miltex | 6-118 | STERRAD |
| Drill | STRYKER | Q9R-5400 | NA |
| Drill bits | STRYKER | 277-82-87 | STERRAD |
| Gauze | MEDLINE | NON26334 | NA (sterilized package) |
| Hemostatic mosquito forceps | Miltex | 7-2, 7-4 | STERRAD |
| Light handles | SKYTRON | Stellar XL | STERRAD |
| Needle hodler | Miltex | 8-2 | STERRAD |
| Periosteal elevator | Miltex | 18-1968 | STERRAD |
| Rongeurs | Miltex | 17-4800 | STERRAD |
| Saline | BAXTER | 2F7122 | STERRAD |
| Scalpel | Bard-Parker | 372610 | STERRAD |
| Scissors | Miltex | 5-12, 5-114 | STERRAD |
| Senn retractors | Miltex | 28065 | STERRAD |
| Sterile gloves | MEDLINE | Triumph Micro | NA (sterilized package) |
| Suction | medela | 200-4869 | NA |
| Suction tip | MEDLINE | DYNDFR12S | NA (sterilized package) |
| Suction tube | COVIDEN | 8888301614 | NA (sterilized package) |
| Surgical gowns | MEDLINE | DYNJP2002S | NA (sterilized package) |
| Surgical pens & ruler | MEDLINE | DYNJSM03 | NA (sterilized package) |
| Suture | COVIDEN | SL-635 | NA (sterilized package) |
| Tissue forceps | Miltex | 6-114 | STERRAD |
| Towel clamps | Miltex | 7-504 | STERRAD |
| Wood swabs | MEDLINE | MDS202095 | NA (sterilized package) |
| Equipment: *cannulas | |||
| Item | |||
| Hypodermic needle | EXELINT INTERNATIONAL | 26437 | NA (sterilized package) |
| Stainless steel tube | K-TUBE | K30R | NA |
| PTFE tube | ZEUS | 216200 | NA |
| Equipment: **colored oil | |||
| Item | |||
| Liquid Candle Dye Concentrate | PremiumCraft | Red/Pink | NA |
| Mineral oil | Vi-Jon | S0883 | NA |
| STERRAD: low-temperature hydrogen peroxide gas plasma |
Referenzen
- Kojima, Y., Robinson, F. R., Soetedjo, R. Cerebellar fastigial nucleus influence on ipsilateral abducens activity during saccades. Journal of Neurophysiology. 111 (8), 1553-1563 (2014).
- Kojima, Y., Soetedjo, R. Elimination of the error signal in the superior colliculus impairs saccade motor learning. Proceedings of the National Academy of Sciences of the United States of America. 115 (38), 8987-8995 (2018).
- Kojima, Y., Soetedjo, R., Fuchs, A. F. Effects of GABA agonist and antagonist injections into the oculomotor vermis on horizontal saccades. Brain Research. 1366, 93-100 (2010).
- Kojima, Y., Soetedjo, R., Fuchs, A. F. Effect of inactivation and disinhibition of the oculomotor vermis on saccade adaptation. Brain Research. 1401, 30-39 (2011).
- De, A., El-Shamayleh, Y., Horwitz, G. D. Fast and reversible neural inactivation in macaque cortex by optogenetic stimulation of GABAergic neurons. eLife. 9, 52658 (2020).
- El-Shamayleh, Y., Kojima, Y., Soetedjo, R., Horwitz, G. D. Selective optogenetic control of Purkinje cells in monkey cerebellum. Neuron. 95 (1), 51-62 (2017).
- Mich, J. K., et al. Functional enhancer elements drive subclass-selective expression from mouse to primate neocortex. Cell Reports. 34 (13), 108754 (2021).
- Graybuck, L. T., et al. Enhancer viruses for combinatorial cell subclass-specific labeling. Neuron. (21), 00159 (2020).
- Michel, J., Benninger, D., Dietz, V., van Hedel, H. J. Obstacle stepping in patients with Parkinson's disease. Complexity does influence performance. Journal of Neurology. 256 (3), 457-463 (2009).
- Chan, K. Y., et al. Engineered AAVs for efficient noninvasive gene delivery to the central and peripheral nervous systems. Nature Neuroscience. 20 (8), 1172-1179 (2017).
- Kaku, Y., Yoshida, K., Iwamoto, Y. Learning signals from the superior colliculus for adaptation of saccadic eye movements in the monkey. Journal of Neuroscience. 29 (16), 5266-5275 (2009).
- Soetedjo, R., Fuchs, A. F., Kojima, Y. Subthreshold activation of the superior colliculus drives saccade motor learning. Journal of Neuroscience. 29 (48), 15213-15222 (2009).
- Kleinbart, J. E., et al. A modular implant system for multimodal recording and manipulation of the primate brain. Annual International Conference of the IEEE Engineering in Medicine and Biology Society. IEEE Engineering in Medicine and Biology Society. Annual International Conference. 2018, 3362-3365 (2018).
- Arieli, A., Grinvald, A., Slovin, H. Dural substitute for long-term imaging of cortical activity in behaving monkeys and its clinical implications. Journal of Neuroscience Methods. 114 (2), 119-133 (2002).
- Ruiz, O., et al. Optogenetics through windows on the brain in the nonhuman primate. Journal of Neurophysiology. 110 (6), 1455-1467 (2013).
- Fredericks, J. M., et al. Methods for mechanical delivery of viral vectors into rhesus monkey brain. Journal of Neuroscience Methods. 339, 108730 (2020).
- Bankiewicz, K. S., et al. Convection-enhanced delivery of AAV vector in parkinsonian monkeys; in vivo detection of gene expression and restoration of dopaminergic function using pro-drug approach. Experimental Neurology. 164 (1), 2-14 (2000).
- Yazdan-Shahmorad, A., et al. Widespread optogenetic expression in macaque cortex obtained with MR-guided, convection enhanced delivery (CED) of AAV vector to the thalamus. Journal of Neuroscience Methods. 293, 347-358 (2018).
- Nathanson, J. L., Yanagawa, Y., Obata, K., Callaway, E. M. Preferential labeling of inhibitory and excitatory cortical neurons by endogenous tropism of adeno-associated virus and lentivirus vectors. Neuroscience. 161 (2), 441-450 (2009).
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenWeitere Artikel entdecken
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten