
Method Article
Culture d’organoïdes 3D intestinaux de porcelet à partir de cryptes épithéliales cryoconservées et établissement de monocouches cellulaires
Dans cet article
Résumé
Ici, nous décrivons un protocole de culture d’organoïdes 3D intestinaux de porc à partir de cryptes épithéliales cryoconservées. Nous décrivons également une méthode pour établir des monocouches cellulaires dérivées d’organoïdes 3D, permettant l’accès à la face apicale des cellules épithéliales.
Résumé
Les organoïdes intestinaux sont de plus en plus utilisés pour étudier l’épithélium intestinal pour la modélisation des maladies digestives, ou pour étudier les interactions avec les médicaments, les nutriments, les métabolites, les agents pathogènes et le microbiote. Des méthodes de culture d’organoïdes intestinaux sont maintenant disponibles pour de multiples espèces, y compris les porcs, qui est une espèce d’intérêt majeur à la fois comme animal de ferme et comme modèle translationnel pour les humains, par exemple, pour étudier les maladies zoonotiques. Ici, nous donnons une description approfondie d’une procédure utilisée pour cultiver des organoïdes 3D intestinaux de porc à partir de cryptes épithéliales gelées. Le protocole décrit comment cryoconserver les cryptes épithéliales de l’intestin du porc et les procédures ultérieures pour cultiver des organoïdes intestinaux 3D. Les principaux avantages de cette méthode sont (i) la dissociation temporelle de l’isolement des cryptes de la culture d’organoïdes 3D, (ii) la préparation de stocks importants de cryptes cryoconservées provenant de plusieurs segments intestinaux et de plusieurs animaux à la fois, et donc (iii) la réduction du besoin d’échantillonner des tissus frais d’animaux vivants. Nous détaillons également un protocole pour établir des monocouches cellulaires dérivées d’organoïdes 3D pour permettre l’accès à la face apicale des cellules épithéliales, qui est le site d’interactions avec les nutriments, les microbes ou les médicaments. Dans l’ensemble, les protocoles décrits ici constituent une ressource utile pour étudier l’épithélium intestinal du porc dans la recherche vétérinaire et biomédicale.
Introduction
L’épithélium intestinal est formé d’une monocouche de cellules recouvrant la muqueuse digestive à l’interface avec l’environnement luminal. Cette position est associée à diverses fonctions, telles que l’absorption des nutriments et la fonction de barrière, qui sont soutenues par la présence de cellules souches et de multiples types de cellules épithéliales différenciées (cellules absorbantes, entéroendocrines, Paneth et caliciformes)1. Les lignées cellulaires immortalisées traditionnellement utilisées pour étudier les cellules épithéliales présentent des limitations majeures, car elles ne reflètent pas la complexité cellulaire de l’épithélium intestinal et présentent des anomalies génomiques2. Le développement d’organoïdes tridimensionnels (3D) par Sato et al.3 a fourni un nouveau modèle pour étudier l’épithélium intestinal avec une pertinence physiologique améliorée. En effet, les organoïdes intestinaux sont dérivés de cellules souches non transformées, sont composés de multiples types de cellules, et récapitulent la fonctionnalité de l’épithélium intestinal. Les organoïdes intestinaux sont de plus en plus utilisés pour comprendre le développement et les fonctions de l’épithélium intestinal et ses interactions avec les agents pathogènes, les nutriments, les toxines, les médicaments, le microbiote et ses métabolites2.
Initialement développées pour l’homme et la souris, les méthodes utilisées pour la culture des organoïdes intestinaux ont récemment été adaptées à d’autres espèces, dont les porcs4. Gonzales et al.5 ont été les premiers à cultiver des organoïdes porcins du jéjunum ; Depuis lors, des organoïdes porcins ont été décrits pour d’autres segments intestinaux (duodénum, iléon et côlon)6,7,8, et il a été démontré qu’ils conservent un phénotype spécifique à l’emplacement 9,10,11. Les organoïdes 3D intestinaux de porc sont maintenant couramment utilisés pour étudier l’effet des nutriments 12,13 ou des infections entériques 6,8,14.
La plupart des études ont décrit la culture d’organoïdes intestinaux à partir de cryptes épithéliales fraîchement isolées. Cependant, cela n’est pas toujours possible pour des raisons logistiques, notamment lorsque l’on travaille avec de gros animaux tels que les porcs. En effet, les animaleries pour porcs peuvent être situées loin du laboratoire où les organoïdes sont cultivés, ce qui complique l’organisation du travail. De plus, la culture organoïde prend beaucoup de temps; Ainsi, il n’est pas pratique de cultiver simultanément plusieurs lignées organoïdes, par exemple, à partir de différents segments intestinaux ou de plusieurs animaux. Pour contourner ces problèmes, quelques études chez l’homme, les chevaux et les porcs ont décrit des méthodes pour cultiver des organoïdes à partir de tissus intestinaux congelés (ou biopsies) ou de cryptes épithéliales isolées 4,15,16,17. Ces méthodes permettent la cryoconservation de cellules souches épithéliales intestinales provenant de plusieurs segments intestinaux d’un seul animal, qui peuvent ensuite être utilisées pour cultiver des organoïdes en cas de besoin. De plus, cela permet une forte réduction du nombre d’animaux vivants utilisés comme donneurs de cellules souches, car d’importants stocks de cryptes cryoconservées peuvent être créés (principes des 3R). Un autre avantage de cette méthode est la croissance des organoïdes intestinaux uniquement à partir d’animaux d’intérêt après avoir obtenu des résultats phénotypiques ou génotypiques, ce qui est très rentable.
In vivo, les cellules épithéliales intestinales sont polarisées, le côté apical étant dirigé vers la lumière. In vitro, dans les organoïdes 3D, la face apicale des cellules épithéliales est également face à la lumière (c’est-à-dire à l’intérieur des organoïdes)4. Cette organisation empêche l’accès au côté apical, ce qui est un problème lors de l’étude des effets des composants luminaux (p. ex. nutriments, microbes, métabolites) sur les cellules épithéliales. Pour contourner cet inconvénient, plusieurs méthodes ont été développées, telles que la culture de cellules organoïdes en monocouches 2D, la micro-injection et l’inversion de polarité (« organoïdes apical-out »)18,19. La culture de monocouches de cellules organoïdes est en train de devenir le système le plus efficace et le plus traitable. Le principe est de dissocier les organoïdes 3D en cellules uniques et de les ensemencer sur un récipient de culture cellulaire préalablement recouvert d’une fine couche de matrice extracellulaire (MEC)20. Dans ces conditions de culture, la face apicale des cellules épithéliales est orientée vers le haut, et est donc accessible à des traitements expérimentaux20. La culture de monocouches de cellules organoïdes a été récemment adaptée pour l’intestin de porc21,22; Les monocouches cellulaires dérivées d’organoïdes 3D porcins ont été utilisées pour de multiples applications, notamment l’étude des infections entériques 6,23,24,25, le transport des nutriments9 et la modélisation des maladies digestives 26.
Ici, cette étude présente d’abord un protocole détaillé pour la culture et le maintien d’organoïdes 3D intestinaux de porc dérivés de cryptes épithéliales cryoconservées (Figure 1). Ensuite, un protocole est décrit pour établir des monocouches cellulaires à partir d’organoïdes 3D intestinaux de porc. Les méthodes décrites ici fournissent des outils expérimentaux qui peuvent être utilisés pour étudier l’épithélium intestinal du porc pour le transport des nutriments, la fonction de barrière et les interactions hôte-microorganisme.
Protocole
Ce protocole a été approuvé par le comité local d’éthique (N°TOXCOM/0136/PP) conformément à la directive européenne sur la protection des animaux utilisés à des fins scientifiques (2010/63/UE). Ce protocole est décrit pour le jéjunum à titre d’exemple, mais il peut être utilisé pour chaque segment de l’intestin grêle et du gros intestin (duodénum, jéjunum, iléon, côlon).
1. Isolement des cryptes épithéliales de l’intestin du porcelet
REMARQUE : Préparer un stock complet de milieu aigle modifié de Dulbecco (DMEMc) avec du DMEM complété par 10 % de sérum fœtal bovin (FBS) et 1 % de pénicilline-streptomycine (P/S). Préparer des aliquotes de 50 mL et les conserver à 4 °C pendant 1 mois.
- Préparation des solutions (à faire le jour de l’isolement de la crypte)
- Préparer la solution de dissociation contenant une solution saline tamponnée au phosphate (PBS), 3 mM de dithiothréitol (DTT), 9 mM d’acide éthylènediaminetétraacétique (EDTA), 10 μM Y27632 inhibiteur de ROCK et 1 % de pénicilline-streptomycine (P/S) et conserver sur de la glace.
- Préparer la solution de congélation contenant du DMEMc, 10 % de FBS, 10 % de diméthylsulfoxyde (DMSO) et 10 μM d’inhibiteur de ROCK Y27632 et conserver sur de la glace (la concentration finale de FBS est de 18 %).
- Préparez la solution de transport contenant du PBS froid complété par 1% P/S et stockez-la sur de la glace.
- Isolement des cryptes épithéliales
- Abattage d’un porcelet par électronarcose suivie d’une exsanguination.
- Immédiatement après l’abattage, ouvrez l’abdomen du porcelet avec un scalpel et retirez tout l’intestin.
- Prélever environ 2 cm d’un segment intestinal et le conserver dans une solution de transport frigorifique. Gardez le segment sur la glace jusqu’à l’isolement de la crypte (jusqu’à 2 h).
- Placez le mouchoir dans une boîte de Pétri. Ouvrez le segment intestinal longitudinalement et lavez soigneusement le tissu dans du PBS froid complété par 1% P / S pour éliminer le contenu intestinal.
- Transférer le tissu dans une nouvelle boîte de Petri remplie de 10 ml de PBS froid complété par 1% P/S.
- Tenez le tissu avec une pince à épiler et retirez les villosités et le mucus restant en grattant avec une lame de microscope.
NOTE: L’ablation des villosités (la structure en forme de langue) peut être vérifiée par observation microscopique du surnageant. - Transférer le tissu dans un tube conique de 15 mL contenant 5 mL de solution de dissociation glacée et incuber pendant 30 minutes à température ambiante (RT) sur un agitateur rotatif (15 rpm).
- Transférer le tissu dans une nouvelle boîte de Petri et ajouter 10 ml de PBS froid complété par 1% P/S.
- Isolez les cryptes mécaniquement en grattant fermement la muqueuse avec une lame de microscope.
REMARQUE : Au microscope, vérifier la présence de cryptes épithéliales dans le PBS (Figure 2A). - Aspirer la solution cryptique avec une pipette sérologique et filtrer à travers une crépine cellulaire de 100 μm dans un tube conique de 50 mL.
- Piper 10 μL de la solution et vérifier la présence de cryptes au microscope. Centrifuger à 300 x g pendant 5 min à 4 °C.
- Sous une enceinte de biosécurité stérile, jeter le surnageant et remettre en suspension la pastille de cryptes dans 10 mL de DMEMc froid additionné d’inhibiteur de ROCK Y27632 de 10 μM.
- Piper 10 μL de la solution cryptique dans une plaque de 48 puits. Comptez manuellement le nombre de cryptes au microscope avec un grossissement de 10x et calculez la concentration de cryptes par mL de solution.
NOTE: Les cryptes isolées peuvent être utilisées directement pour la culture d’organoïdes intestinaux. Cependant, il est souvent plus pratique de cryoconserver un grand lot de cryptes de chaque porcelet et de les utiliser plus tard pour la culture organoïde.
- Congélation des cryptes épithéliales
- Transférer un volume correspondant à 900 cryptes dans un tube conique de 15 mL. Centrifuger à 300 x g pendant 5 min à 4 °C.
- Jeter le surnageant et remettre en suspension la pastille de cryptes dans 1 mL de la solution de congélation. Transférer dans un cryotube et placer le flacon dans un récipient de congélation cellulaire.
- Conservez le récipient de congélation cellulaire à -80 °C pendant 24 h, puis transférez les flacons dans de l’azote liquide pour un stockage à long terme.
2. Mise en place d’organoïdes 3D intestinaux de porcelet à partir de cryptes épithéliales congelées
REMARQUE : Les organoïdes 3D intestinaux des porcelets sont cultivés dans un milieu de culture commercial formulé pour la croissance des organoïdes humains, complétés par 1 % P/S et 100 μg/mL d’un agent antimicrobien pour les cellules primaires, et stockés à 4 °C pendant 1 semaine. Une matrice extracellulaire dérivée d’une tumeur (ECM) est utilisée pour la culture d’organoïdes 3D. Toutes les références de produits commerciaux sont présentées dans le tableau des matériaux.
- Préparation des matériaux
- Placez les embouts de pipettes à -20 °C (au moins pendant la nuit).
- Placer les aliquotes congelées de l’ECM (500 μL) à 4 °C au moins 1 h à l’avance.
- Préchauffer une plaque de 48 puits dans un incubateur à 37 °C, 5 % de CO2 .
- Placez le milieu de culture à RT.
- Préchauffer un bain-marie à 37 °C.
- Placez un petit seau à glace sous le capot dans des conditions aseptiques.
- Décongélation des cryptes épithéliales gelées
- Décongeler rapidement un flacon contenant 900 cryptes gelées au bain-marie à 37 °C (moins de 5 min).
- Transférer la solution cryptique dans un tube conique de 15 mL.
- Centrifuger à 300 x g pendant 5 min à TA. Retirer le surnageant
- Ajouter 150 μL de l’ECM avec des pointes refroidies pour obtenir une concentration finale de 150 cryptes par 25 μL d’ECM. Pipet de haut en bas 10 fois pour obtenir une suspension homogène des cryptes dans l’ECM.
REMARQUE: Gardez toujours l’ECM sur la glace pour éviter la polymérisation. Utilisez toujours des embouts de pipette pré-refroidis à -20 °C pour manipuler l’ECM. Pipet lentement pour éviter de faire des bulles d’air dans l’ECM. Un ECM non dilué est utilisé à cette étape pour empêcher les petites gouttes de s’effondrer. - Ensemencer six puits avec une goutte de 25 μL par puits avec des pointes refroidies dans une plaque préchauffée de 48 puits.
REMARQUE: Gardez la pointe verticale, au centre du puits, et pipetez lentement sans introduire d’air pour obtenir un dôme. Ici, des plaques à 48 puits sont utilisées, car le nombre d’organoïdes est généralement faible lorsque l’on commence à partir de cryptes gelées. - Incuber pendant 30 min dans un incubateur à 37 °C, 5% CO2 pour la polymérisation de l’ECM.
- Ajouter 250 μL par puits de milieu de culture à TA. Incuber dans un incubateur à 37 °C, 5% de CO 2 et changer le milieu de culture tous les2-3 jours
REMARQUE: Les cryptes ne sont généralement pas visibles après la procédure de décongélation et la plupart des cellules sont dissociées dans l’ECM (Figure 2B).
3. Passage d’organoïdes 3D intestinaux de porcelets dérivés de cryptes congelées
REMARQUE: Le temps pour obtenir des organoïdes à partir de cryptes gelées est généralement plus long que lorsque vous commencez à partir de cryptes fraîches. Les organoïdes sont généralement prêts à se fendre 10 jours après la décongélation (figure 2B).
- Préparation des matériaux
- Placer les aliquotes congelées de l’ECM (500 μL) à 4 °C pendant au moins 1 h.
- Préchauffer les plaques de 24 puits à 37 °C.
- Préchauffer le PBS et le réactif de dissociation enzymatique supplémenté en inhibiteur de ROCK Y27632 10 μM dans un bain-marie à 37 °C.
- Placez le milieu de culture à RT.
- Placez un petit seau à glace sous le capot dans des conditions aseptiques.
- Passage d’organoïdes 3D dérivés de cryptes gelées
- Retirer le milieu de culture et laver avec 250 μL de PBS préchauffé à 37 °C.
- Ajouter 250 μL de réactif de dissociation enzymatique préchauffé complété par un inhibiteur de ROCK Y27632 de 10 μM à 37 °C dans chaque puits.
NOTE: En raison du faible nombre d’organoïdes, la dissociation des organoïdes est effectuée directement dans chaque puits. - Détacher les organoïdes dans l’ECM en grattant avec une pipette P1000 et homogénéiser soigneusement en pipetant cinq fois.
- Incuber pendant 5 min dans un incubateur à 37 °C, 5% CO2 . Dissocier les cellules en pipetant de haut en bas 10 fois à l’aide d’une pipette P1000.
NOTE: L’objectif est d’obtenir des cellules isolées ou des amas de petites cellules (<10 cellules). Vérifiez la dissociation au microscope. Si de gros fragments d’organoïdes sont encore observés, répéter l’étape 3.2.4. - Ajouter 500 μL de DMEMc dans chaque puits contenant des cellules dissociées et regrouper jusqu’à 12 puits dans un tube conique de 15 mL contenant 3 mL de DMEMc froid.
- Centrifuger à 500 x g pendant 5 min à 4 °C. Jeter le surnageant et remettre en suspension la pastille dans 1 mL de DMEMc froid.
- Compter les cellules avec une dilution de 1:2 dans du bleu de trypan avec un compteur de cellules.
Remarque : Le compteur de cellules automatisé peut compter les cellules dans de petits clusters, le cas échéant. - Centrifuger le volume nécessaire de la solution cellulaire pour avoir 3 000 cellules vivantes par dôme (un dôme par puits de la plaque de 24 puits) à 500 x g pendant 5 min à 4 °C.
- Remettez les cellules en suspension avec 17 μL de DMEMc froid par 3 000 cellules vivantes sur de la glace. Ajustez le volume au nombre de puits requis.
- Ajouter lentement 33 μL d’ECM froid avec des pointes refroidies pour 3 000 cellules vivantes et homogénéiser sur de la glace sans faire de bulles. Ajustez le volume au nombre de puits requis.
REMARQUE: Les cellules sont remises en suspension dans une solution contenant 1/3 DMEMc et 2/3 ECM. Pour chaque dôme, 50 μL de cette solution sont nécessaires. L’ECM dilué est moins cher et plus facile à pipeter. - Ensemencer les puits avec 50 μL de suspension de cellules ECM par puits avec des pointes refroidies dans une plaque préchauffée de 24 puits.
- Incuber pendant 30 min dans un incubateur à 37 °C, 5% CO2 pour la polymérisation de l’ECM.
- Ajouter 500 μL du milieu de culture par puits. Incuber dans un incubateur à 37 °C, 5% de CO 2 et changer le milieu de culture tous les2-3 jours
NOTE: Les organoïdes peuvent être utilisés directement pour i) des expériences, ii) le maintien de la culture organoïde 3D, iii) la congélation ou iv) l’ensemencement des monocouches de cellules organoïdes (Figure 1). Vérifiez la croissance des organoïdes avec un microscope tous les jours pour choisir le moment optimal pour diviser les organoïdes. Les organoïdes doivent avoir une lumière claire et vide et des bords bien définis. Les organoïdes matures avec des débris noirs dans la lumière ne doivent pas être utilisés pour la division (Figure 3).
4. Maintenance de la culture organoïde en 3D
REMARQUE: Pour le passage, les organoïdes doivent apparaître clairs avec une lumière vide. Des débris noirs apparaissent dans la lumière environ 5 jours après la division et indiquent la présence de cellules mortes. Limiter le nombre de cellules mortes au moment du passage est préférable pour un maintien optimal de la culture. Le calendrier devrait donc être adapté pour éviter d’atteindre ce stade de maturité.
- Préparation du matériel
- Placer les aliquotes de l’ECM (500 μL) à 4 °C pour les décongeler pendant au moins 1 h.
- Préchauffer une plaque de 24 puits à 37 °C.
- Préchauffer le réactif de dissociation enzymatique complété par un inhibiteur de ROCK Y27632 de 10 μM dans un bain-marie à 37 °C.
- Placez le milieu de culture à RT.
- Placez un petit seau à glace sous le capot dans des conditions aseptiques.
- Passage des organoïdes 3D intestinaux
- Détacher les organoïdes avec l’ECM en grattant avec une pipette P1000. Homogénéiser soigneusement en pipetant dans le milieu de culture et transférer dans un tube conique de 15 mL contenant 5 mL de DMEMc froid sur de la glace.
REMARQUE : Un tube conique de 15 mL est requis pour une piscine comportant 12 puits d’organoïdes cultivés dans des dômes de 50 μL dans des plaques de 24 puits. - Centrifuger les organoïdes collectés à 500 x g pendant 5 min à 4 °C.
NOTE: Les organoïdes forment une pastille blanche au fond du tube. Si les organoïdes sont toujours dans la suspension dans l’ECM après centrifugation, aspirer soigneusement le surnageant supérieur (sans toucher la couche ECM), homogénéiser en pipetant 10 fois avec une pipette P1000 et répéter l’étape de centrifugation. La couche ECM ne doit pas être visible après cette procédure. - Aspirer soigneusement le surnageant et remettre en suspension la pastille cellulaire dans 1 mL de réactif de dissociation enzymatique préchauffé complété par un inhibiteur de ROCK Y27632 de 10 μM. Pipeter de haut en bas 10 fois pour initier la dissociation des organoïdes.
- Incuber pendant 5 min dans un bain-marie à 37 °C pour la digestion enzymatique.
- Perturber mécaniquement les organoïdes en pipetant 10 fois avec une pipette P1000. Vérifiez la suspension cellulaire au microscope.
NOTE: L’objectif est d’obtenir des cellules isolées ou des amas de petites cellules. Si de gros fragments organoïdes sont encore observés, répéter l’incubation (étape 4.2.4) et la perturbation mécanique (étape 4.2.5). - Ajouter 4 mL de DMEMc glacé. Centrifugeuse à 500 x g pendant 5 min à 4 °C
- Jeter le surnageant et remettre en suspension la pastille de cellule organoïde dans 1 mL de DMEMc.
- Procéder comme décrit ci-dessus (étapes 3.2.7 à 3.2.13) pour compter les cellules et ensemencer les cellules organoïdes dans 50 μL de dômes ECM contenant 3000 cellules vivantes dans des plaques préchauffées à 24 puits.
- Détacher les organoïdes avec l’ECM en grattant avec une pipette P1000. Homogénéiser soigneusement en pipetant dans le milieu de culture et transférer dans un tube conique de 15 mL contenant 5 mL de DMEMc froid sur de la glace.
5. Congélation d’organoïdes 3D
- Préparer le volume nécessaire (1 mL pour une piscine de deux dômes) de solution de congélation contenant du DMEMc additionné de 10 % de FBS, de 10 % de DMSO et d’inhibiteur de ROCK Y27632 à 10 μM et conserver sur de la glace (la concentration finale de FBS est de 18 %).
- Retirez le milieu de culture des puits à congeler.
- Ajouter 1 mL de la solution de congélation au premier puits à congeler, détacher l’ECM par grattage et homogénéiser avec l’embout de la pipette.
- Transférer la suspension organoïde du premier puits au second puits à congeler.
- Transférer la piscine des deux puits dans un cryotube et placer le flacon dans un récipient de congélation cellulaire.
- Conservez le récipient de congélation cellulaire à -80 °C pendant 24 h, puis transférez les flacons dans de l’azote liquide pour un stockage à long terme.
6. Culture de monocouches cellulaires dérivées d’organoïdes 3D
NOTE: Les monocouches de cellules organoïdes porcines sont cultivées dans un milieu 2D composé du milieu de culture utilisé pour les organoïdes 3D complété par 20% FBS.
- Préparation des matériaux
- Préparez le support 2D et conservez-le à RT.
- Préchauffer le réactif de dissociation enzymatique complété par un inhibiteur de ROCK Y27632 de 10 μM dans un bain-marie à 37 °C.
- Revêtement de l’insert de culture
- Stérilisez une paire de pinces à épiler et transférez-les dans l’enceinte de biosécurité.
- Placez les inserts de culture cellulaire (0,33 cm2) dans une plaque de 24 puits avec la pince à épiler.
- Préparer la solution d’enrobage contenant du collagène IV dilué à 50 μg/mL dans du PBS froid. Tuyler de haut en bas pour mélanger
- Ajouter 150 μL de la solution IV de collagène dilué à chaque insert de culture cellulaire, ce qui correspond à 22,7 μg/cm2.
REMARQUE: Orienter soigneusement la pipette verticalement au centre de la membrane perméable et vérifier que la solution de collagène couvre toute la membrane. - Placer la plaque dans un incubateur à 37 °C, 5% de CO2 et laisser reposer toute la nuit (ou pendant un minimum de 3 h).
- Ensemencement de cellules organoïdes 3D dans des inserts de culture cellulaire
- Préparez une suspension cellulaire à partir des organoïdes 3D, comme décrit aux étapes 4.2.1 à 4.2.7.
- Compter les cellules avec une dilution de 1:2 dans du bleu de trypan avec un compteur de cellules, et calculer le volume nécessaire pour ensemencer 2,5 x 10 5 cellules par insert de culture, correspondant à 7,6 x 105 cellules/cm2.
- Centrifuger le volume nécessaire de la suspension cellulaire à 500 x g pendant 5 min à 4 °C.
- Pendant la centrifugation, aspirer soigneusement la solution de revêtement des inserts de culture et laisser sécher à TA sous le capot sans couvercle pendant 5 min.
- Après centrifugation, jeter le surnageant et remettre en suspension la pastille cellulaire dans le volume nécessaire de milieu 2D complété par un inhibiteur de ROCK Y27632 de 10 μM. Un volume de 200 μL de milieu 2D contenant les cellules est nécessaire pour chaque insert.
- Ensemencer 200 μL de la suspension cellulaire (2,5 x 105 cellules) sur la membrane perméable enrobée (face apicale) (figure 4A).
REMARQUE: Pipet lentement au centre de la membrane et garder l’embout vertical. - Ajouter 500 μL du milieu 2D supplémenté en inhibiteur de ROCK Y27632 10 μM dans le compartiment inférieur (face basale). Incuber dans un incubateur à 37 °C, 5% CO2 .
- Un jour après l’ensemencement, remplacez les milieux apical et basal par un milieu 2D frais sans l’inhibiteur Y27632 ROCK.
- Changez le support 2D chaque jour. La monocouche devient confluente 1 jour après l’ensemencement et peut ensuite être utilisée pour des expériences.
REMARQUE : Une valeur de résistance électrique transépithéliale (TEER) au-dessus du blanc (insérer sans cellules) confirme que la confluence est atteinte (Figure 4B).
7. Immunomarquage des monocouches de cellules organoïdes
- Préparation des solutions
NOTE: Ajuster le volume des solutions en fonction du nombre de puits à colorer; 200 μL de solution sont nécessaires à chaque étape pour un puits. Les références à tous les produits commerciaux sont fournies dans le tableau des matériaux.- Préparer une solution de paraformaldéhyde (PFA) à 4 % sous une hotte chimique en ajoutant 5 mL de PFA à 32 % à 35 mL de PBS. Préparer des aliquotes de 10 ml et conserver à -20 °C.
ATTENTION : Manipulez toujours le PFA sous le capot chimique, tout en portant des gants en nitrile. - Préparer une solution de Triton X100-PBS à 0,2 %, en ajoutant 2 μL de Triton X100 à 1 mL de PBS immédiatement avant utilisation. Garder à RT.
- Préparer une solution d’albumine sérique bovine (BSA)-PBS à 10 % en ajoutant 100 mg de BSA à 1 mL de PBS. Garder à RT.
- Préparer une solution de BSA-PBS à 1% en ajoutant 10 mg de BSA à 1 mL de PBS. Garder à RT.
- Préparer une solution d’anticorps primaire occludine diluée à 1:200 en ajoutant 5 μL d’anticorps primaire à 995 μL de PBS BSA à 1%. Gardez la solution sur la glace.
- Préparer une solution de l’anticorps secondaire dilué à 1:1 000 en ajoutant 1 μL d’anticorps secondaire à 999 μL de BSA-PBS à 1%. Gardez la solution sur la glace, à l’abri de la lumière.
- Préparer une solution de phalloïdine TRITC à 10 μg/mL en ajoutant 10 μL de TRITC à 1 mg/mL à 990 μL de PBS. Gardez sur la glace, à l’abri de la lumière.
- Préparer une solution de paraformaldéhyde (PFA) à 4 % sous une hotte chimique en ajoutant 5 mL de PFA à 32 % à 35 mL de PBS. Préparer des aliquotes de 10 ml et conserver à -20 °C.
- Immunomarquage
NOTE: Sauf indication contraire, toutes les incubations sont effectuées à RT, sous agitation lente sur une plate-forme à bascule (30 tr/min). L’immunomarquage est effectué directement dans l’insert de culture cellulaire.- Retirez le milieu basal et apical. Placez la plaque sous le capot chimique.
- Laver les monocouches deux fois avec 200 μL de PBS à TA et incuber pendant 5 min.
- Fixer les monocouches cellulaires avec 200 μL de PFA à 4% à TA et incuber pendant 20 min à TA.
- Laver les monocouches deux fois avec 200 μL de PBS à TA et incuber pendant 5 min.
REMARQUE: Après les étapes de fixation et de lavage, la monocouche peut être maintenue à 4 ° C dans PBS pendant 1 semaine. - Retirer le PBS, perméabiliser avec 200 μL de Triton X100-PBS à 0,2% et incuber pendant 20 min.
- Laver les monocouches deux fois avec 200 μL de PBS à TA et incuber pendant 5 min.
- Ajouter 200 μL de solution d’anticorps primaires dans 1% BSA-PBS et incuber pendant une nuit à 4 °C, sous agitation lente sur une plate-forme à bascule. Inclure un puits témoin négatif en ajoutant seulement 1% BSA-PBS sans l’anticorps primaire.
- Laver les monocouches trois fois avec 200 μL de PBS à TA et incuber pendant 5 min.
- Ajouter 200 μL d’anticorps secondaire dans 1% BSA-PBS et incuber à TA pendant 2 h, à l’abri de la lumière.
- Laver les monocouches trois fois avec 200 μL de PBS à TA et incuber pendant 5 min.
- Ajouter 200 μL de phalloïdine TRITC à 10 μg/mL et incuber pendant 10 min.
- Laver les monocouches deux fois avec 200 μL de PBS à TA et incuber pendant 5 min.
- Retirez le PBS et coupez la membrane avec un scalpel.
- Récupérez la membrane avec une pince à épiler et placez-la sur une lame de microscope, avec la face apicale tournée vers le haut.
- Ajouter 15 μL du support de montage complété par DAPI à 1:1 000 directement sur la membrane. Placez un bordereau de couverture et sceller.
- Conserver à 4 °C, à l’abri de la lumière, jusqu’à l’imagerie.
Résultats
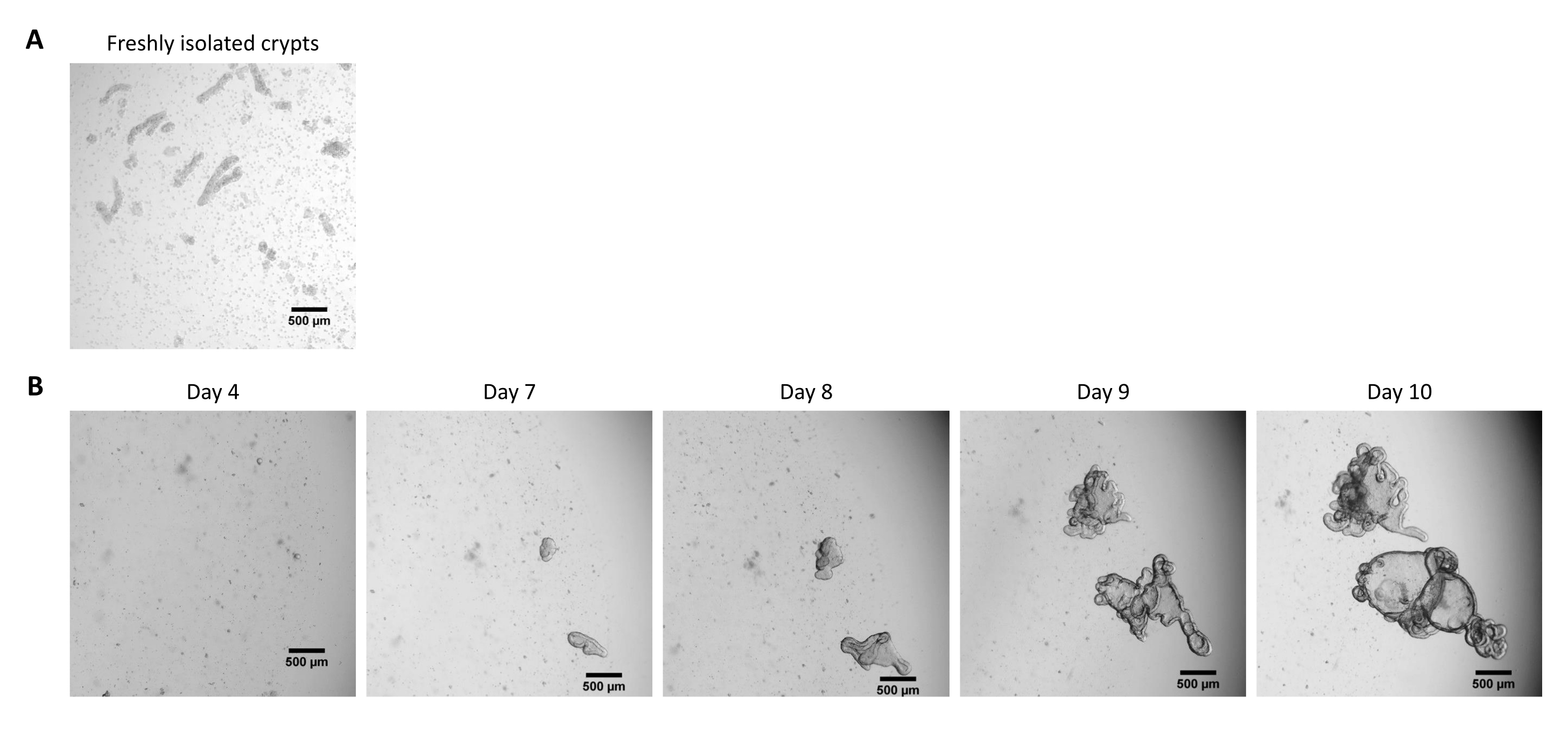
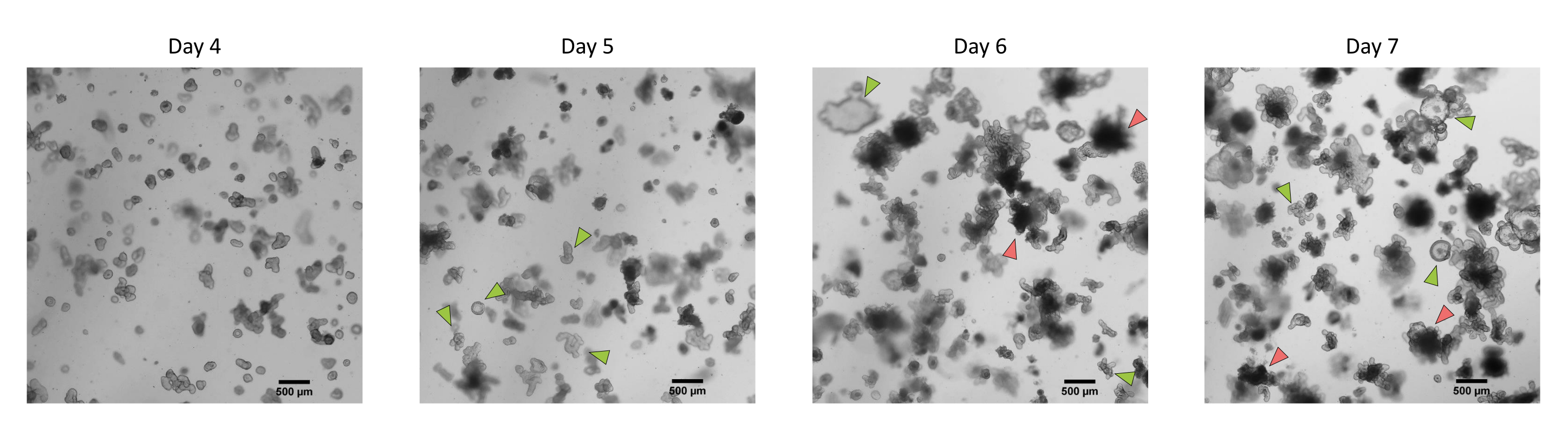
Selon le protocole décrit ci-dessus, les cryptes épithéliales sont obtenues à partir de l’intestin du porc et cryoconservées pour un stockage à long terme dans de l’azote liquide (Figure 1 et Figure 2A). Après décongélation, les cellules souches cryptiques sont ensemencées dans l’ECM (Figure 2B). La structure de la crypte est généralement perdue après cette étape, en raison de la désintégration de la structure de la crypte dans l’ECM. Les organoïdes peuvent être observés en 3-4 jours, puis se développent rapidement et développent des structures bourgeonnantes (Figure 2B). Nous avons réussi à obtenir des organoïdes après décongélation de cryptes gelées dans >80% des tentatives. Environ 10 jours après la décongélation (selon le taux de croissance des organoïdes), un passage des organoïdes est effectué pour élargir la culture (Figure 3). Les organoïdes se développent plus rapidement après la division, et ils présentent diverses morphologies, avec quelques organoïdes kystiques et une majorité d’organoïdes bourgeonnants. Pour un maintien optimal de la culture, les organoïdes utilisés pour le passage doivent présenter une lumière claire et vide et des bords bien définis (exemples indiqués par des flèches vertes) sans débris noirs dans la lumière (indiqués par des flèches rouges), comme observé chez les organoïdes matures (Figure 3). Nous avons constaté que les débris de globules noirs commencent à s’accumuler vers le jour 6 après le décès. Ainsi, il est recommandé de diviser ou de congeler les organoïdes au jour 4-5 après le passage.
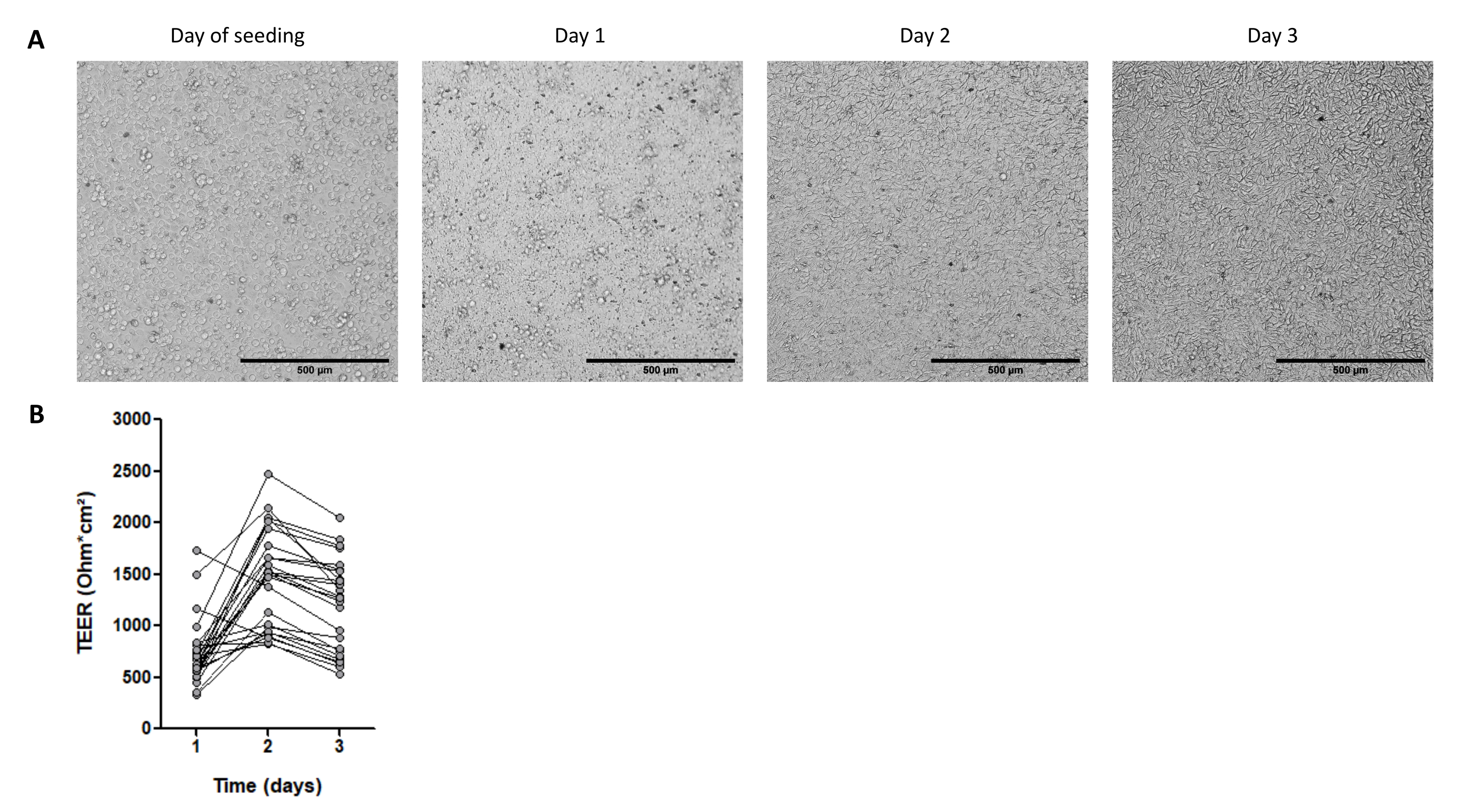
Le stade de maturité des organoïdes est également un point important dans l’obtention des monocouches cellulaires. Les organoïdes à maturité avancée (indiquée par la présence de débris de cellules noires dans la lumière) ne sont pas optimaux pour ensemencer des monocouches. Nous dissocions généralement les organoïdes 3D 4 jours après le passage pour collecter des cellules pour la culture 2D. Environ un à trois puits d’organoïdes 3D cultivés à 3 000 cellules par dôme de 50 μL de l’ECM sont nécessaires pour ensemencer un insert de culture de 2,5 x 105 cellules. Les cellules se fixent et forment une monocouche entièrement confluente en 1 jour (Figure 4A), ce qui est confirmé par le TEER élevé d’environ 700 Ω·cm2 (Figure 4B). Cependant, les bords cellulaires sont difficiles à visualiser par microscopie à fond clair à ce stade précoce, probablement en raison d’un faible niveau de différenciation. Le TEER reste élevé pendant 3 jours (Figure 4B).
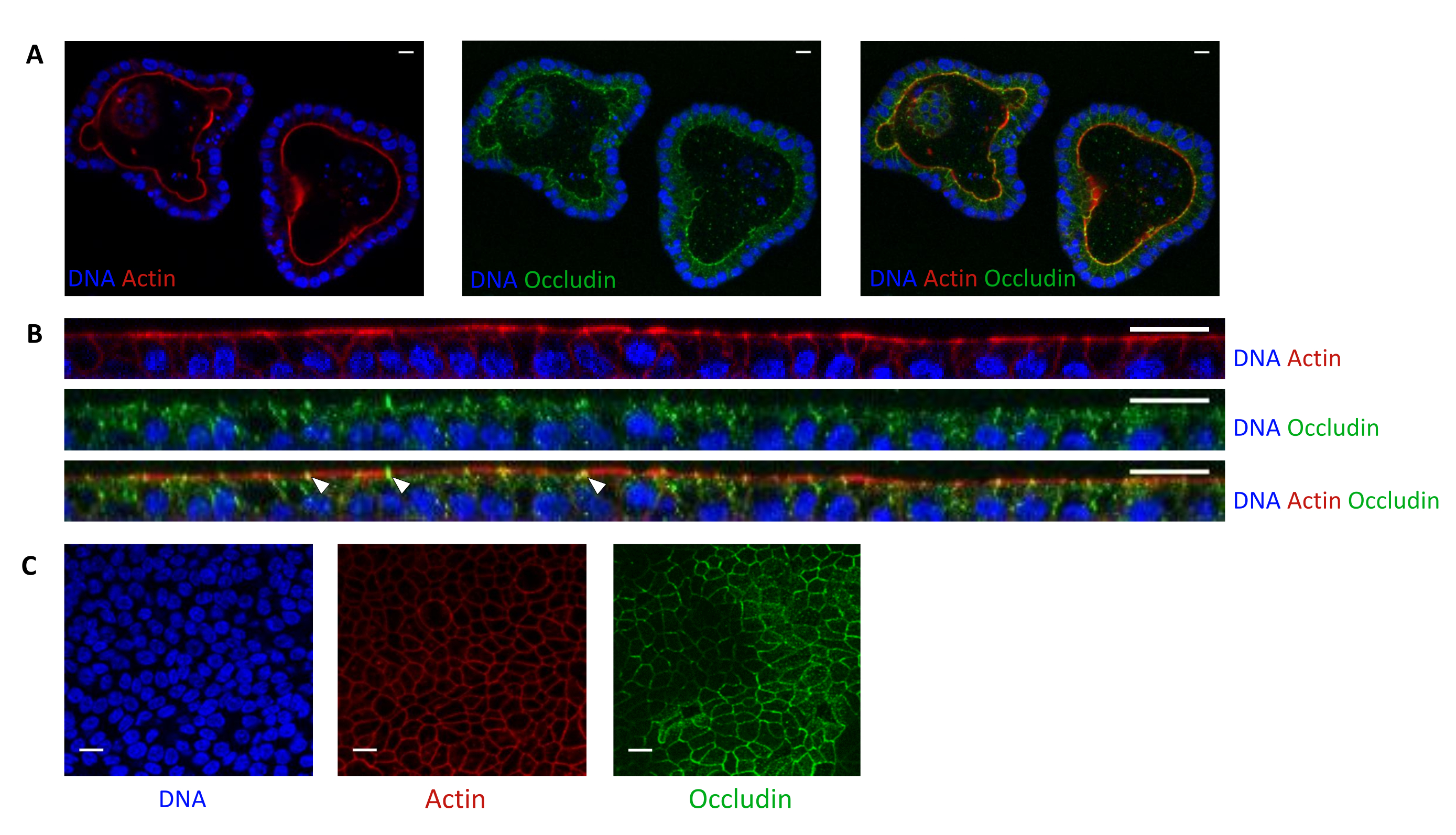
La coloration à l’actine indique que la face apicale des cellules épithéliales est orientée vers la lumière dans les organoïdes 3D (Figure 5A). Les cellules organoïdes ensemencées en culture forment une couche unique confluente de cellules épithéliales, la face apicale étant orientée vers le compartiment supérieur (Figure 4B,C). La coloration à l’occludine révèle la présence de jonctions serrées du côté apical des cellules épithéliales dans les organoïdes 3D et dans les monocouches cellulaires (Figure 5A-C).

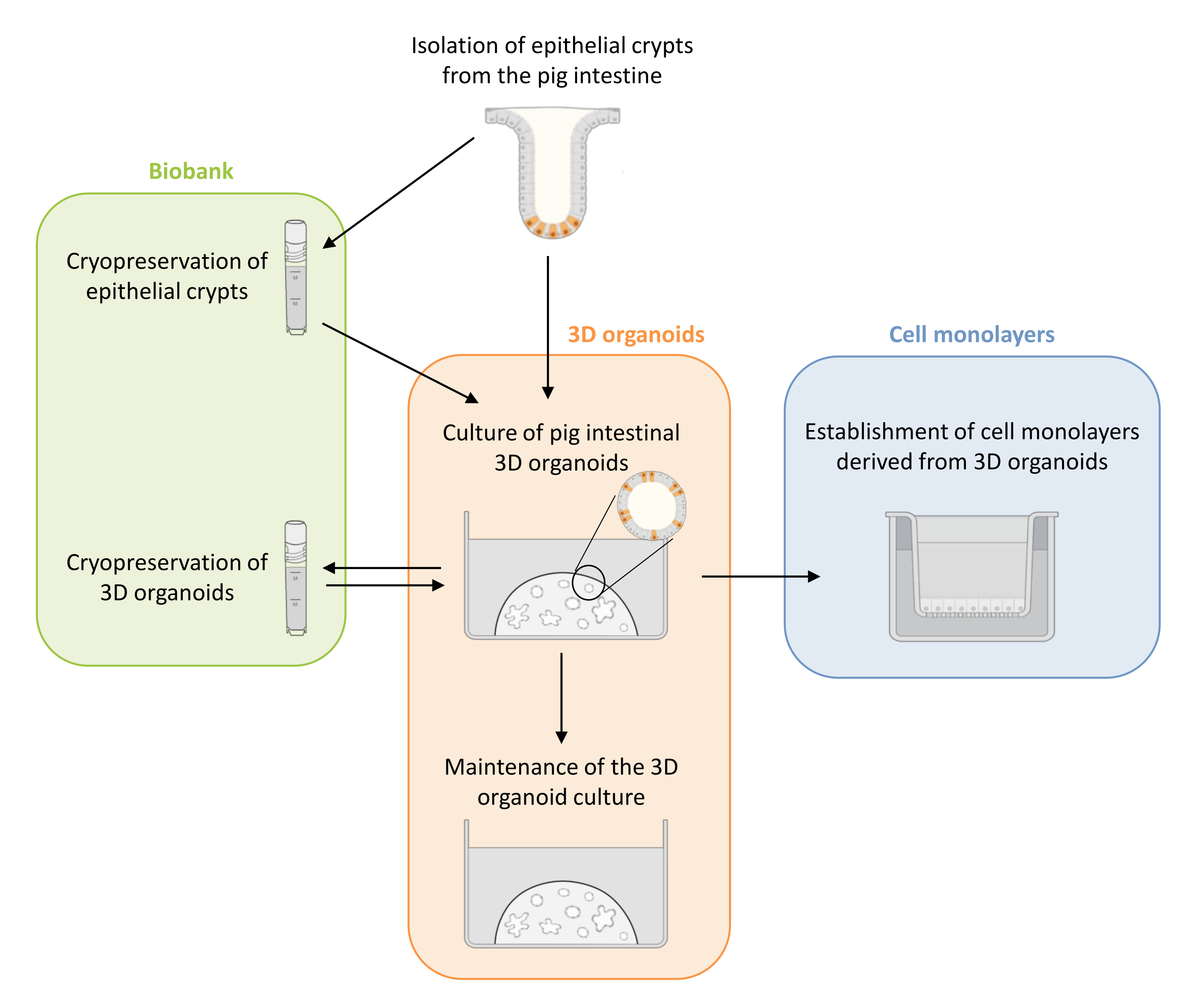
Figure 1 : Représentation schématique des méthodes utilisées pour cultiver des organoïdes 3D et des monocouches cellulaires dérivées d’organoïdes. Les cryptes épithéliales sont isolées de l’intestin du porcelet. Ces cryptes peuvent i) être utilisées immédiatement pour la culture d’organoïdes 3D ou ii) être congelées et stockées dans une biobanque dans de l’azote liquide. Les cryptes cryoconservées peuvent être décongelées et utilisées pour cultiver des organoïdes 3D. La culture organoïde 3D peut être maintenue par fractionnement successif, ou congelée et stockée dans la biobanque. Des monocouches cellulaires peuvent être obtenues à partir de la culture organoïde 3D pour permettre l’accès à la face apicale des cellules et étudier la fonction de barrière épithéliale. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 2 : Culture d’organoïdes 3D intestinaux de porc provenant de cryptes épithéliales cryoconservées. (A) Image microscopique représentative de cryptes jéjunales fraîchement isolées. (B) Images microscopiques représentatives d’organoïdes 3D obtenues après décongélation des cryptes jéjunales. Les organoïdes ont été cultivés dans un dôme d’ECM de 25 μL dans une plaque de 48 puits. La figure montre des images des organoïdes 3D 4, 7, 8, 9 et 10 jours après l’ensemencement. La barre d’échelle représente 500 μm. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 3 : Morphologie des organoïdes 3D intestinaux de porc dérivés de cryptes épithéliales cryoconservées après le premier passage. Images microscopiques représentatives montrant le développement d’organoïdes dérivés de cryptes de jéjunum cryoconservées après le premier passage du jour 4 au jour 7. Les flèches vertes indiquent des organoïdes clairs appropriés pour le passage ou l’ensemencement des monocouches. Les flèches rouges indiquent des organoïdes matures qui ne conviennent pas à la division ou à l’ensemencement des monocouches; Ainsi, les puits doivent être utilisés avant l’apparition de cette morphologie. La barre d’échelle représente 500 μm. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 4 : Caractéristiques des monocouches cellulaires dérivées d’organoïdes 3D intestinaux de porc. (A) Images microscopiques représentatives de la morphologie de la monocouche sur 3 jours. Les cellules organoïdes de jéjunum ont été ensemencées à 2,5 x 105 cellules dans des inserts de culture cellulaire de 0,33 cm 2 recouvertsde collagène IV à 50 ng / mL. La barre d’échelle représente 500 μm. (B) Résistance électrique transépithéliale (TEER) des monocouches de cellules organoïdes sur 3 jours. Les points reliés par une ligne correspondent au même puits à des moments différents. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 5 : Imagerie d’organoïdes de jéjunum de porc cultivés en 3D ou 2D. Imagerie par microscopie confocale de (A) organoïdes 3D 5 jours après le fractionnement et de monocouches de cellules organoïdes (B,C) 3 jours après l’ensemencement (B: section XZ; C : section XY). L’ADN (bleu) a été coloré avec du DAPI. L’actine (rouge) était colorée à la phalloïdine. L’occludine (vert) a été colorée avec un anticorps polyclonal. Les flèches blanches indiquent l’occludine localisée à la jonction serrée. La barre d’échelle représente 20 μm. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Discussion
Ce protocole décrit une méthode utilisée pour cryoconserver les cryptes épithéliales de l’intestin du porcelet pour le stockage à long terme et la culture ultérieure d’organoïdes 3D. Ce protocole utilise une solution de congélation contenant du DMSO, du FBS, l’inhibiteur Y27632 ROCK, du DMEM et des antibiotiques. Une autre étude chez le porc a obtenu des organoïdes provenant de cryptes cryoconservées dans une solution de congélation similaire mais sans l’inhibiteur ROCK15. L’inhibiteur Y27632 ROCK a été inclus pour prévenir l’apoptose et maintenir le pool de cellules souches puisque, après décongélation, les cellules cryptiques épithéliales sont dissociées, ce qui peut entraîner la mort cellulaire (« anoikis »)27,28. Il est intéressant de noter que des entéroïdes équins ont été obtenus à partir de cryptes épithéliales congelées dans un milieu de culture contenant uniquement du DMEM et du DMSO16; Cette méthode simple n’a pas encore été testée pour les cryptes épithéliales du porc. D’autres méthodes ont été publiées pour cultiver des organoïdes humains et porcins à partir de tissus congelés ou de biopsies au lieu de cryptes épithéliales 4,17. L’avantage de cette méthode est la possibilité de cryoconserver directement les tissus intestinaux sans effectuer la procédure d’isolement de la crypte, qui nécessite du temps et du matériel de laboratoire. Cela peut être pratique lorsque les tissus doivent être prélevés loin du laboratoire. Cependant, lors de l’isolement des cryptes immédiatement après l’abattage, de grands segments de l’intestin peuvent être traités pour obtenir un très grand nombre de cryptes, ce qui n’est pas le cas à partir de petits fragments de tissus congelés. Après le dégel des cryptes épithéliales, des organoïdes ont été observés 3-4 jours après l’ensemencement et se sont divisés après 10 jours. Il s’agit d’un taux de croissance plus lent que lors du démarrage de la culture à partir de cryptes épithéliales fraîches, pour lesquelles les organoïdes ont été obtenus dès le jour 1 après l’ensemencement et peuvent généralement être divisés vers le jour 5et 11. Khalil et al. ont également signalé un retard de croissance des entéroïdes porcins à partir de cryptes congelées15, ce qui suggère que les cellules souches pourraient avoir besoin de temps pour récupérer leur capacité proliférative. Nous avons également obtenu un nombre inférieur d’organoïdes en partant de cryptes congelées par rapport aux cryptes fraîches, ce qui pourrait être dû à la mort des cellules souches pendant le processus de congélation. Dans certaines tentatives de décongélation des cryptes (<20%), nous n’avons pas obtenu d’organoïdes à partir de cryptes gelées, probablement en raison d’une procédure de cryoconservation sous-optimale (par exemple, congélation retardée après isolement de la crypte de probablement plus de 1 h). Ainsi, nous recommandons de garder les cryptes sur la glace jusqu’à ce qu’elles soient comptées et de les congeler le plus rapidement possible.
Pour les organoïdes 3D, nous avons choisi d’utiliser un milieu de culture organoïde commercial formulé pour les humains. En effet, des rapports antérieurs ont montré que les organoïdes intestinaux de porc se développent efficacement avec ce milieu 8,11,14,19,25,26,29,30. Il est intéressant que ce milieu de culture soit prêt à l’emploi et présente une concentration standardisée de facteurs de croissance au sein d’un lot. Cependant, ce milieu de culture est coûteux, sa composition n’est pas divulguée et il n’est donc pas possible de moduler sa composition. En revanche, d’autres études ont mis en culture des organoïdes intestinaux de porc dans des milieux personnalisés contenant des inhibiteurs pharmacologiques, un facteur de croissance recombinant et/ou des milieux conditionnés 5,6,7,21. Bien que très flexible et moins chère, cette méthode prend beaucoup de temps pour la production de milieux conditionnés et pourrait manquer de reproductibilité en raison de la variabilité potentielle de la concentration des facteurs de croissance dans les milieux conditionnés. Ainsi, la qualité de chaque lot de milieux conditionnés doit être validée en mesurant la croissance organoïde ou l’expression géniquemarqueur 31.
Une étude a montré que les organoïdes jéjunaux de porc cultivés dans le même milieu de culture organoïde commercial utilisé ici se développaient plus rapidement et semblaient moins différenciés que les entéroïdes cultivés avec des milieux contenant un facteur de croissance recombinant et/ou des milieux conditionnés23. Un état prolifératif élevé facilite la culture d’organoïdes 3D, mais peut nécessiter d’induire une différenciation pour être plus représentatif des caractéristiques physiologiques intestinales. Dans ce protocole, pour le passage des organoïdes 3D, les cellules sont entièrement dissociées pour le comptage, ce qui permet de contrôler le nombre de cellules ensemencées en ECM. Cela augmente la reproductibilité du phénotype des organoïdes, qui est fortement influencé par leur densité. De plus, compter les cellules évite d’obtenir une culture trop faible ou surpeuplée, ce qui nécessite d’adapter le calendrier de culture. La plupart des autres études ont préparé des fragments organoïdes non complètement dissociés en cellules individuelles et ont utilisé un rapport de dilution pour le passage. Cette méthode est plus simple, mais pourrait induire une variabilité en fonction de la densité organoïde de la culture.
Pour la culture en monocouches, les cellules organoïdes sont ensemencées dans des inserts de culture préenduits d’une fine couche d’ECM, qui permet la fixation des cellules mais évite la croissance des organoïdes en 3D. Ce protocole utilisait du collagène de type IV comme protéine ECM, comme décrit précédemment chez les porcs23. D’autres études avec des monocouches organoïdes de porc ont utilisé la même ECM dérivée de la tumeur utilisée ici pour cultiver des organoïdes 3D 6,8,9,21,25,30. L’avantage de l’utilisation du collagène est la possibilité de normaliser la concentration en protéines avec une composition entièrement définie, ce qui n’est pas le cas dans l’ECM dérivée de la tumeur. Une étape critique pour le succès de la culture des monocouches cellulaires est de prêter attention à l’apparence visuelle des organoïdes 3D précurseurs, qui devraient avoir des bords bien définis et une lumière vide sans débris noirs. En effet, les organoïdes avec un niveau de maturation élevé et un faible taux de prolifération ne sont pas une source appropriée de cellules pour la culture 2D. Ainsi, le moment de la dissociation des organoïdes 3D en cellules uniques est crucial pour le succès de cette étape.
La culture de monocouches 2D à partir de cellules individuelles permet de standardiser le nombre de cellules ensemencées, ce qui est plus difficile à partir de fragments organoïdes, comme cela est effectué dans d’autres méthodes. Nous avons ensemencé 7,6 x 10 5 cellules par cm 2, ce qui est élevé par rapport à la plupart des autres études 21,22,23 chez des porcs qui utilisaient une densité cellulaire inférieure, allant de0,25 x 10 5 cellules par cm 2 à 1,78 x 10 5 cellules par cm2. L’exigence d’un nombre élevé de cellules organoïdes constitue une limitation de ce protocole, mais elle nous a permis d’obtenir rapidement une monocouche confluente, recouvrant entièrement l’insert de culture après 1 jour. En revanche, Vermeire et al.23 ont obtenu une confluence après 4-7 jours avec une densité plus faible de cellules ensemencées (de 0,25 x 10 5 cellules/cm 2 à 0,4 x 105 cellules/cm2). Certaines études ont également utilisé des monocouches de cellules organoïdes de porc qui ne couvraient pas entièrement la surface de culture pour les infections par les virus 8,30. Dans ces conditions, la face apicale des cellules épithéliales est accessible pour les traitements, mais des monocouches entièrement confluentes sont nécessaires si l’objectif est d’étudier l’absorption des nutriments ou la perméabilité épithéliale.
Pour les monocouches de cellules organoïdes, un milieu de culture organoïde commercial complété par 20 % de FBS a été utilisé, sur la base d’une étude récente sur les monocouches dérivées d’entéroïdesbovins 32. Dans nos tests, la supplémentation avec 20% FBS était nécessaire pour obtenir des monocouches entièrement confluentes, probablement en raison d’une exigence élevée en facteur de croissance. Au contraire, d’autres études utilisant le même milieu commercial ont établi des monocouches sans FBS 8,25,30 supplémentaire, mais sans atteindre la confluence complète. D’autres études ont également utilisé une supplémentation avec 20% FBS dans un milieu personnalisé pour la culture de monocouches de cellules organoïdesporcines 21,22. Dans nos expériences, TEER est élevé 1 jour après l’ensemencement (environ 700 Ω·cm2), et reste élevé jusqu’au jour 3 (environ 1 500 Ω·cm2 ; ceci est cohérent avec la formation de jonctions serrées, comme l’indique l’expression de l’occludine. Van der Hee et al. ont obtenu des valeurs TEER similaires sur 72 h pour les monocouches organoïdes de jéjunum21. Ils ont également démontré que les monocouches peuvent être maintenues jusqu’au jour 12-15 avec des changements quotidiens de support. En revanche, d’autres études ont rapporté des valeurs TEER beaucoup plus faibles (environ 200 Ω·cm2) pour les monocouches de cellules organoïdesporcines 6,22. Ces différences entre les études peuvent être liées au segment intestinal étudié ou aux milieux utilisés qui influencent la différenciation épithéliale.
En conclusion, le protocole ci-dessus pour cultiver des organoïdes 3D intestinaux de porc à partir de cryptes épithéliales congelées facilite l’organisation du travail de culture. Il réduit le besoin d’obtenir des tissus frais à partir d’animaux vivants. Nous expliquons également comment établir des monocouches cellulaires entièrement confluentes dérivées d’organoïdes porcins en moins de 3 jours. Ainsi, nos protocoles pourraient être des ressources utiles pour les scientifiques qui étudient l’épithélium intestinal du porc pour la recherche vétérinaire ou biomédicale.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
Ce travail a été soutenu par l’Institut Carnot France Futur Elevage (projet OrganoPig) et par INRAE HOLOFLUX (projet Holopig). Les auteurs sont reconnaissants envers les installations centrales de Genotoul (TRI). Nous remercions Christelle Knudsen (GenPhySE, INRAE, Toulouse) pour sa relecture attentive.
matériels
| Name | Company | Catalog Number | Comments |
| 0.5 M EDTA, pH 8.0 | Thermo Fisher Scientific | AM9260G | Store at room temperature. |
| 15 mL conical tube | Sarstedt | 62.554.502 | |
| 24-well cell culture plate | Corning | 003526 | |
| 48-well cell culture plate | Corning | 003548 | |
| 50 mL polypropylene conical tube | Falcon | 352070 | |
| Bovine Serum Albumine (BSA) | Euromedex | A6003 | Store at 4 °C. |
| Centrifuge Universal 320 R | Hettich | 1406 | |
| Collagene type IV from human placenta | Sigma | C5533 | Prepare the stock solution at 1 mg/mL in acetic acid according to the manufacturer's recommendation. Aliquot (500 µL) and store at -20 °C. |
| CoolCell LX Cell Freezing Container | Corning | 432003 | Used to cryopreserve crypts and organoids. |
| Countess 3 Automated Cell Counter | Thermo Fisher Scientific | 16842556 | |
| Coverslips, 22 mm x 50 mm | VWR | 630-1845 | |
| Cryotube ClearLine 1 mL | Clear line | 390706 | Used to cryopreserve crypts and organoids. |
| DAPI | Invitrogen | D1306 | Prepare the stock solution at 5 mg/mL in water according to the manufacturer’s recommendation. Aliquot (20 µL) and store at -20 °C |
| Dithiothreitol (DTT) | Merck | 10197777001 | Store at 4 °C. |
| DMEM, high glucose, GlutaMAX Supplement, pyruvate | Thermo Fisher Scientific | 31966047 | Store at 4 °C. |
| DMSO (Dimethyl Sulfoxide) | Corning | 25-950-CQC | Store at room temperature. |
| Epredia Superfrost Plus Adhesion microscopic slide | VWR | 631-9483 | |
| Fetal Bovine Serum (FBS) | Thermo Fisher Scientific | 10270-106 | Store 5 mL aliquots at -20 °C. |
| Fisherbrand Sterile Cell Strainers | Thermo Fisher Scientific | 22363549 | Used for crypt isolation. |
| Fixed Tilt 3D Platform Rotator | VWR | 97025-564 | Used for incubations in the immunostaining protocol. |
| Gibco PBS, pH 7.4 | Thermo Fisher Scientific | 10010015 | Store at 4 °C. |
| Gibco TrypLE Express Enzyme (1x), phenol red | Thermo Fisher Scientific | 12605-010 | Enzyme dissociation reagent. Store at room temperature. |
| Goat anti-rabbit IgG, Alexa fluor 488 | Thermo Fisher Scientific | A-11008 | Secondary antibody. Store at 4 °C. Working dilution 1:1000. |
| IntestiCult Organoid Growth Medium (Human) | Stem Cell Technology | 6010 | Organoid culture medium. Store at -20 °C. Thaw the basal medium and organoid supplement at room temperature and mix (1:1). Store the mix at 4 °C for up to 1 week. |
| Insert with PET membrane transparent Falcon for plate 24 wells | Corning | 353095 | |
| Inverted microscope | Nikon | Eclipse TS2 | |
| Matrigel Basement Membrane Matrix | Corning | 354234 | Tumor-derived extracellular matrix used for the 3D culture of organoids. Matrigel polymerizes at room temperature. Use cooled tips to pipet the Matrigel. Prepare 500 µL aliquots and store at -20 °C. |
| Mounting medium for fluorescence with DAPI | Vectashield | H1250 | Store at 4 °C. |
| Occludin polyclonal antibody | Thermo Fisher Scientific | 71-1500 | Primary antibody. Store at -20 °C. Working dilution 1:200. |
| Paraformaldehyde 32% | Electron microscopy science | 15714 | Prepare 4% paraformaldehyde (PFA) solution under chemical hood by adding 5 mL of 32% PFA to 35 mL of PBS. Aliquot by 10 mL and store at -20 °C. |
| Penicillin-Streptomycin | Sigma | P4333 | Antibacterial. Store 5 mL aliquots at -20 °C. |
| Phalloidin TRITC | Sigma | P1951 | Probe for actin staining. 1 mg/mL stock solution. Store at 4 °C. |
| Primocin | InvivoGen | ant-pm-05 | Antimicrobial agent for primary cells acting on bacteria, mycoplasma and fungi. Store at -20 °C. |
| ROCK Inhibitor (Y27632) | ATCC | ACS-3030 | Used to maintain the stem cells. Prepare the stock solution at 10 mM in sterile water according to the manufacturer's recommendation and store aliquots (50 µL) at -20 °C. |
| Rotating shaker mix XL | Clear line | 062646CL | Used for crypt isolation. |
| Stripette Serological Pipets 10 mL | Corning | 4488 | |
| Tissue Culture Dish | TPP | 93100 | |
| Triton X100 | Sigma | 8787 | Store at room temperature. |
| Trypan Blue stain 0.4% | Thermo Fisher Scientific | T10282 | Store at room temperature. |
| Vacuum system Vacusip | Integra | 159000 | Used to remove the medium of organoid wells. |
Références
- Peterson, L. W., Artis, D. Intestinal epithelial cells: regulators of barrier function and immune homeostasis. Nature Reviews Immunology. 14 (3), 141-153 (2014).
- In, J. G., et al. Human mini-guts: new insights into intestinal physiology and host-pathogen interactions. Nature Reviews Gastroenterology & Hepatology. 13 (11), 633-642 (2016).
- Sato, T., et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature. 459 (7244), 262-265 (2009).
- Beaumont, M., et al. Intestinal organoids in farm animals. Veterinary Research. 52 (1), 33 (2021).
- Gonzalez, L. M., Williamson, I., Piedrahita, J. A., Blikslager, A. T., Magness, S. T. Cell lineage identification and stem cell culture in a porcine model for the study of intestinal epithelial regeneration. PLoS One. 8 (6), 66465 (2013).
- Holthaus, D., Delgado-Betancourt, E., Aebischer, T., Seeber, F., Klotz, C. Harmonization of protocols for multi-species organoid platforms to study the intestinal biology of toxoplasma gondii and other protozoan infections. Frontiers in Cellular and Infection Microbiology. 10, 610368 (2021).
- Powell, R. H., Behnke, M. S. WRN conditioned media is sufficient for in vitro propagation of intestinal organoids from large farm and small companion animals. Biology Open. 6 (5), 698-705 (2017).
- Li, L., et al. Porcine intestinal enteroids: a new model for studying enteric coronavirus porcine epidemic diarrhea virus infection and the host innate response. Journal of Virology. 93 (5), 01682 (2019).
- vander Hee, B., Madsen, O., Vervoort, J., Smidt, H., Wells, J. M. Congruence of transcription programs in adult stem cell-derived jejunum organoids and original tissue during long-term culture. Frontiers in Cell and Developmental Biology. 8, 375 (2020).
- Barnett, A. M., et al. Porcine colonoids and enteroids keep the memory of their origin during regeneration. American Journal of Physiology. Cell Physiology. 320 (5), 794-805 (2021).
- Mussard, E., et al. The phenotype of the gut region is more stably retained than developmental stage in piglet intestinal organoids. Frontiers in Cell and Developmental Biology. 10, 983031 (2022).
- Zhu, M., Qin, Y. -. C., Gao, C. -. Q., Yan, H. -. C., Wang, X. -. Q. l-Glutamate drives porcine intestinal epithelial renewal by increasing stem cell activity via upregulation of the EGFR-ERK-mTORC1 pathway. Food & Function. 11 (3), 2714-2724 (2020).
- Wang, Z., et al. Dietary vitamin A affects growth performance, intestinal development, and functions in weaned piglets by affecting intestinal stem cells. Journal of Animal Science. 98 (2), (2020).
- Derricott, H., et al. Developing a 3D intestinal epithelium model for livestock species. Cell and Tissue Research. 375 (2), 409-424 (2019).
- Khalil, H. A., et al. A novel culture system for adult porcine intestinal crypts. Cell and Tissue Research. 365 (1), 123-134 (2016).
- Stewart, A. S., Freund, J. M., Gonzalez, L. M. Advanced three-dimensional culture of equine intestinal epithelial stem cells. Equine Veterinary Journal. 50 (2), 241-248 (2018).
- Tsai, Y. -. H., et al. A method for cryogenic preservation of human biopsy specimens and subsequent organoid culture. Cellular and Molecular Gastroenterology and Hepatology. 6 (2), 218-222 (2018).
- Wilson, S. S., Tocchi, A., Holly, M. K., Parks, W. C., Smith, J. G. A small intestinal organoid model of non-invasive enteric pathogen-epithelial cell interactions. Mucosal Immunology. 8 (2), 352-361 (2015).
- Li, Y., et al. Next-generation porcine intestinal organoids: an apical-out organoid model for swine enteric virus infection and immune response investigations. Journal of Virology. 94 (21), 01006-01020 (2020).
- Moon, C., VanDussen, K. L., Miyoshi, H., Stappenbeck, T. S. Development of a primary mouse intestinal epithelial cell monolayer culture system to evaluate factors that modulate IgA transcytosis. Mucosal Immunology. 7 (4), 818-828 (2014).
- vander Hee, B., et al. Optimized procedures for generating an enhanced, near physiological 2D culture system from porcine intestinal organoids. Stem Cell Research. 28, 165-171 (2018).
- Hoffmann, P., et al. Intestinal organoid-based 2D monolayers mimic physiological and pathophysiological properties of the pig intestine. PLoS One. 16 (8), 0256143 (2021).
- Vermeire, B., Gonzalez, L. M., Jansens, R. J. J., Cox, E., Devriendt, B. Porcine small intestinal organoids as a model to explore ETEC-host interactions in the gut. Veterinary Research. 52 (1), 94 (2021).
- Luo, H., et al. Utility evaluation of porcine enteroids as PDCoV infection model in vitro. Frontiers in Microbiology. 11, 821 (2020).
- Resende, T. P., Medida, R. L., Vannucci, F. A., Saqui-Salces, M., Gebhart, C. Evaluation of swine enteroids as in vitro models for Lawsonia intracellularis infection1,2. Journal of Animal Science. 98 (2), 011 (2020).
- Engevik, A. C., et al. Editing myosin VB gene to create porcine model of microvillus inclusion disease, with microvillus-lined inclusions and alterations in sodium transporters. Gastroenterology. 158 (8), 2236-2249 (2020).
- Watanabe, K., et al. A ROCK inhibitor permits survival of dissociated human embryonic stem cells. Nature Biotechnology. 25 (6), 681-686 (2007).
- Gracz, A. D., Puthoff, B. J., Magness, S. T. Identification, isolation, and culture of intestinal epithelial stem cells from murine intestine. Somatic Stem Cells: Methods and Protocols. 879, 89-107 (2012).
- Ferrandis Vila, M., et al. Dietary fiber sources and non-starch polysaccharide-degrading enzymes modify mucin expression and the immune profile of the swine ileum. PloS One. 13 (11), 0207196 (2018).
- Li, L., et al. IFN-lambda 3 mediates antiviral protection against porcine epidemic diarrhea virus by inducing a distinct antiviral transcript profile in porcine intestinal epithelia. Frontiers in Immunology. 10, 2394 (2019).
- VanDussen, K. L., Sonnek, N. M., Stappenbeck, T. S. L-WRN conditioned medium for gastrointestinal epithelial stem cell culture shows replicable batch-to-batch activity levels across multiple research teams. Stem Cell Research. 37, 101430 (2019).
- Sutton, K. M., Orr, B., Hope, J., Jensen, S. R., Vervelde, L. Establishment of bovine 3D enteroid-derived 2D monolayers. Veterinary Research. 53 (1), 15 (2022).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationExplorer plus d’articles
This article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.