
A subscription to JoVE is required to view this content. Sign in or start your free trial.
Method Article
מתודולוגיות לימוד
In This Article
Summary
This study presents the development of reproducible methodologies to study biofilm inhibitors and their effects on Bacillus subtilis multicellularity.
Abstract
This work assesses different methodologies to study the impact of small molecule biofilm inhibitors, such as D-amino acids, on the development and resilience of Bacillus subtilis biofilms. First, methods are presented that select for small molecule inhibitors with biofilm-specific targets in order to separate the effect of the small molecule inhibitors on planktonic growth from their effect on biofilm formation. Next, we focus on how inoculation conditions affect the sensitivity of multicellular, floating B. subtilis cultures to small molecule inhibitors. The results suggest that discrepancies in the reported effects of such inhibitors such as D-amino acids are due to inconsistent pre-culture conditions. Furthermore, a recently developed protocol is described for evaluating the contribution of small molecule treatments towards biofilm resistance to antibacterial substances. Lastly, scanning electron microscopy (SEM) techniques are presented to analyze the three-dimensional spatial arrangement of cells and their surrounding extracellular matrix in a B. subtilis biofilm. SEM facilitates insight into the three-dimensional biofilm architecture and the matrix texture. A combination of the methods described here can greatly assist the study of biofilm development in the presence and absence of biofilm inhibitors, and shed light on the mechanism of action of these inhibitors.
Introduction
קהילות חיידקים רבות תאי לשחק תפקידים משמעותיים הסביבה טבעית אנתרופוגניים, והוא יכול להיות מועיל או מזיק ביותר. מושבות רבות התאיות אלה ידועים כמו biofilms, שבה התאים הבודדים המוטבעים מטריצת חומרים פולימריים תאיים בהפקה עצמית (EPS). EPS מאוד לדבוק התאים אל פני השטח הוא ליישב. הם לשמש כמגן נגד כוחות מכניים וכימיים וליצור קשר הדוק בין תאים שכנים, בהקלת התקשורת הסלולרית 1. Biofilm ניתן לצפות כקהילת בדיל, שבו משתמשי תאי פיקוח הדוק, מתוזמר תהליכים כדי לתאם את פעילותם בתוך הקהילה, כמו גם על פני מיני 2-5. המעבר ממצב planktonic ונטול חיים של צמיחה למצב ביופילם קשור לעתים קרובות עם תהליכים התפתחותיים. דוגמה טובה לכך היא subtilis Bacillus חיידק אדמה חיובי-גראם, ולפיכך undomeזן sticated משמש כאורגניזם מודל חזק כדי ללמוד את שלבי ההתפתחות מובילים להיווצרות ביופילם. חיידק זה, תאי ניע להתארגן מבנים רבים-תאיים בולטים מבצעות משימות מיוחדות 4. קבוצה אחת של תאים, המטריצה מפיק להפריש exopolysaccharides 6, חלבון עמילואיד טס 7,8, ואת חלבון הידרופוביות משטח BslA 9,10; שכולן להשתתף באסיפה של EPS 11-13.
בהתחשב השפע של biofilms בנישות טבעיות אנתרופוגניים והניזק הקטלני המשוערת הם יכולים לגרום, יש צורך דוחק למצוא דרכים למנוע היווצרותם. מעכבי מולקולה קטנים יכולים לסייע לגילוי מסלולי רגולטוריות חדשים, אנזימים וחלבונים מבניים המעורבים ביצירת ביופילם, ובכך לקדם תובנות התהליכים המורכבים של הרכבת קהילה רבה-תאית. כב ' subtilis הוא מודל למד היטב עבור ביוהיווצרות סרט 14,15, זה יכול לשמש כדי להעריך את ההשפעות של מעכבי ביופילם שונים. מחקר זה מתמודד עם ארבע שיטות יסוד שהן המפתח להערכת התגובה של biofilms למעכבי מולקולה קטנה. ראשית, על מנת להבטיח כי מעכבים אלה יש מטרת biofilm הספציפי, הפרדת ההשפעה על צמיחת planktonic מהשפעת על היווצרות ביופילם היא קריטית. רוב סוכני אנטיבקטריאלי למקד תאים בשלב הצמיחה planktonic שלהם, אבל מולקולות המשפיעות על אורח החיים ביופילם הם נדירים. בנוסף, כמו מולקולות שאינו משפיעות צמיחת planktonic אינן רעילות, הם יכולים להפחית את לחץ הסלקטיבי העדפת מוטציות אנטיביוטיות עמידות 16. לדוגמה, כאשר biofilms מטופלים עם חומצות אמינו D-או מולקולות מקיר מפריעים תאים אחרים מסוימים, הם גם מוטרדים או מפורקים, אבל מעכבי אלה רק במתינות להשפיע 12,17 צמיחה planktonic. לעומת זאת, אנטיביוטיקה רבה לפגוע בצמיחת planktonic דרמטית, עם little או אין השפעה על היווצרות ביופילם 17.
שנית, הקמת מסגרת ניסיונית עקבית ויציב כדי לחקור את ההשפעה של המולקולות הקטנות חיונית. צפינו כי טווח הריכוז הפעיל של מעכבי מולקולה קטנים היה רגיש לתנאים מראש התרבות להגדרות הניסוי נעשו שימוש כדי לחקור את ההשפעה של מעכבי מולקולה הקטנים אלה. דיווחים שונים, במיוחד הלומדים ב subtilis, חשף וריאציות בטווח הריכוז שבו D-אמינו חומצות לדכא את היווצרות pellicles - את biofilms חיידקי צף 12,17-19. התוצאות המוצגות כאן עולות כי הגורמים הבאים להסביר את ההבדלים בטווח הריכוז הפעיל: התנאים מראש התרבות (12,17 לוגריתמים לעומת שלב מאוחר-נייח 20 צמיחה), את מדיום הגידול בשימוש המצב טרום התרבות (עשירות, undefined [מרק לוריא, LB] לעומת מוגדר [מונוסודיום גלוטמט-glycerol, MSgg]), יחס החיסון ובמיוחד הסרת המדיום מראש התרבות לפני החיסון. הטמפרטורה של צמיחה קרומית סטטי הראה תפקיד חשוב פחות בטווח הפעילות של D-לאוצין מעכב מולקולה קטנה, חומצת אמינו D-נציג השתמשו במחקר זה.
לבסוף, לאחר biofilms מטופלים עם מעכבי biofilm ספציפיים, שיטות חזקות ואינפורמטיבי נדרשות לאפיין את ההשפעות של מעכבים אלה על כושר ביופילם. הנה, שתי שיטות כדי לאפיין את ההשפעה עצמאית של מעכבי מולקולה קטנים מתוארות בפירוט: (1) ההשפעה על תאים בודדים בתוך מושבה ביופילם ואת התנגדותם סוכני מיקרוביאלית. תאי biofilms הם בדרך כלל יותר עמיד לאנטיביוטיקה בהשוואת חיידקים חינם-חי 21-23. בעוד תופעה זו תלויה בכמה גורמים, היכולת של EPS כדי לצמצם את חדירת אנטיביוטיקה לעתים קרובות נחשב כהסבר מושך 24 . שיטה זו מעריכה את הישרדותם של תאי biofilm שנקבע מראש לאחר חשיפה לחומרים אנטי בקטריאליים. (2) ההשפעה על אדריכלות המושבה ביופילם, מן הגדולים אל בקנה מידה קטן. מושבות Biofilm מאופיינים המבנה התלת-ממדי שלהם ואת הנוכחות של EPS. ניצול במיקרוסקופ אלקטרונים סורק, שינויים במורפולוגיה התא, מבנה המושבה ביופילם והארכיטקטורה ושפע של EPS ניתן דמיינו מן גדול (מ"מ) אל בקנה מידה קטן (מיקרומטר).
Protocol
1. הערכת השפעת מולקולה קטנה מעכבי על קרומית היווצרות המושבה Biofilm
- כן פתרון 2x של מדיום MSgg וישכנע ביופילם מוגדר 25 ללא כלוריד סידן וברזל hexahydrate כלוריד (III). לאחר העיקור מסנן, מוסיפים את כלוריד סידן. המדיום הוא מוכן לשימוש ישירות או שזה יכול להיות מאוחסן על 4 ° C בחושך.
- הכן את דילול 1x MSgg ביום הניסוי.
- לדלל את המדיום 2x MSgg ל X 1 עם מים מזוקקים סטריליים (pellicles) או חם 3% סטרילי (80 ° C) אגר (biofilms) ולהוסיף ברזל (III) כלוריד hexahydrate לריכוז סופי של 50 מיקרומטר (pellicles) או 250 מיקרומטר ( biofilms). להוסיף אנטיביוטיקה או מעכבי מולקולה קטנה לריכוז הרצוי ומערבבים היטב. לדוגמה, כדי לקבל ריכוז סופי של 0.5 מ"מ D-לאוצין ב 30 מ"ל להקים pellicles או biofilms, להוסיף 196.6 μl של 76.3 מ"מ (10 מ"ג / מ"ל) פתרון המניות D-לאוצין.
לאTE:. הרכב 1x MSgg הסופי מתואר בטבלת 1 לעומת המתכון המקורי 25, המדיום הכיל 50 מיקרוגרם / מיליליטר תראונין ואת ריכוז הברזל לגדול מושבות ביופילם על מדיום MSgg מוצק הוגדלו 2.5x כדי לייעל את מורפולוגיה המושבה המקומטת .
- לדלל את המדיום 2x MSgg ל X 1 עם מים מזוקקים סטריליים (pellicles) או חם 3% סטרילי (80 ° C) אגר (biofilms) ולהוסיף ברזל (III) כלוריד hexahydrate לריכוז סופי של 50 מיקרומטר (pellicles) או 250 מיקרומטר ( biofilms). להוסיף אנטיביוטיקה או מעכבי מולקולה קטנה לריכוז הרצוי ומערבבים היטב. לדוגמה, כדי לקבל ריכוז סופי של 0.5 מ"מ D-לאוצין ב 30 מ"ל להקים pellicles או biofilms, להוסיף 196.6 μl של 76.3 מ"מ (10 מ"ג / מ"ל) פתרון המניות D-לאוצין.
- לאחר ההתמצקות של אגר, לייבש את צלחות MSgg המוצקות במנדף ביולוגי 30-45 דקות לפני החיסון.
- כדי לבחור מעכבים ספציפיים שמפריעים מנגנוני ההיווצרות קרומית (איור 1), פוסל כי ריכוזי בשימוש להשפיע planktonic וצמיחה סטטי.
- לקבוע צמיחת planktonic (גידול הצפיפות אופטית לאורך זמן בתרבות נוזלית) בתוך עקומת צמיחה פשוט על ידי מדידת הצפיפות האופטית ב 600 ננומטר כל שעה עד שלב הצמיחה הנייח.
- כדי לאשר כי עכירות תרבות נמדדת מייצג ספירת תא חי, לקבוע את מספרי יחידות המושבה להרכיב (CFU)של תאים בשלב הצמיחה planktonic מתרבות רועד לאחר מועדים לפי שעון מספר.
- כדי להעריך את ההשפעה של מעכבי מולקולה קטנה על צמיחה קרומית סטטי, תאים הקציר בסוף הדגירה 3 ימים ב 23 ° C מתרבות תא-24 גם כן, מחוסן באותם תנאים כמפורט בסעיפים 1.7-1.9 ולקבוע את CFU. עבור שליטה זו, להשתמש זן קרומי מחסרת כי חסר את operons קידוד עבור הרכיבים התאים מטריקס (כלומר, ב subtilis Δ epsH, Δ טסה).
הערה: זן זה הוא מסוגל לגדול בתנאים סטטיים, אך בניגוד סוג בר יוצרים קרומי, זה הוא חסר את היכולת לצוף לממשק הנוזל האוויר שצמח היא מועדף בשל רמות חמצן מוגברים 26. לפיכך, מטריצה תאית זו ומתח קרומי מחסרת הוא זן התייחסות מומלץ להעריך צמיחה בתנאים סטטיים.
הערה: לקבלת לשעבר המבוקשבשפע של D-לאוצין חומצת קאנונית אמינו-D כמתואר להלן, השפעה על planktonic וצמיחה סטטי בריכוזים שהפריעו היווצרות קרומית נפסלה 12,17. השיטות לקביעת planktonic וצמיחה סטטי מתוארות בפירוט 17.

באיור 1. סקירה מושגית לזיהוי של התקנה ניסיונית חזקה כדי להעריך את העיכוב הספציפי של היווצרות ביופילם. קריטריונים לבחירה עבור מעכבי מולקולה קטנים המציינים הפרעה ספציפית עם היווצרות ביופילם ללא השפעה מובהקת על צמיחת planktonic. אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של נתון זה.
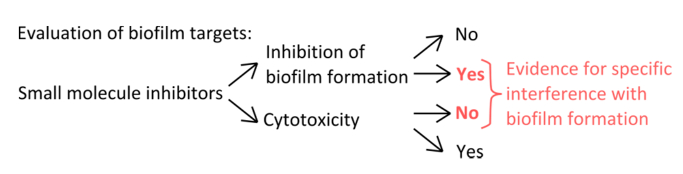{kind=link}
- Streak את B. subtilis ממלאי -80 מעלות צלזיוס (LB cultuמחדש של 10 9 תאים / מ"ל קפוא 20% גליצרול) לבודד מושבות אחת על צלחת אגר LB-1.5% במקל טיפ או המוליך סטרילית.
- לגדול בין לילה ב 30 מעלות צלזיוס.
- שלב קריטי: עבור עיכוב קרומי חזק ידי חומצות קאנונית D-אמינו כגון-לאוצין D, לגדל מושבה אחת הרים מצלחת LB-1.5% אגר ב 3 מיליליטר מרק LB ב 37 מעלות צלזיוס למשך 4 שעות בתוך רועד חממה (רועד סל"ד מהיר 200). החזר את מרק LB עם המדיום biofilm וישכנע MSgg לפני חיסון ידי צנטריפוגה 1.5 מ"ל תרבות המתנע עבור 4 דקות ב 6000 XG, בזהירות הסרת supernatant מחדש השעיית גלולה ב 1.5 מ"ל בינוני MSgg. שאר התרבות יכולה להיות מושלך.
חשוב: כדי להבטיח את איתנות המערכת, את הצפיפות האופטית ב 600 ננומטר (OD 600) של תרבות המתנע שטף צריכה להיות בין 0.6 ל -1. - במהלך הצמיחה של התרבות המתנע, להכין containin צלחת multidish 12 גם תא-תרבותמ"ל גרם 3 של המדיום MSgg בלי או עם טווח הריכוז של מעכבי מולקולה קטנה (למשל, 0.3, 0.5, 1 מ"מ D-לאוצין 17). כדי לשלול השפעות קצה, להפיץ את המיקום של הריכוזים השונים על פני צלחת multidish. לחלופין, השתמש 24 גם צלחות תרבית תאים multidish המכיל 1.5 מ"ל של מדיום MSgg.
- לחסן את בארות הצלחת 12 גם תא-תרבות multidish עם 3 μl של תרבות המתנע שטף (1: 1,000 דילול).
הערה: יחס דילול נמוך, כלומר, 1: 500 ניתן להשתמש. זה מקטין את זמן הפיתוח של pellicles. - לגדל את pellicles ב 23 ° C בתנאים סטטי במשך שלושה ימים. אל תזיז את pellicles במהלך הזמן הזה, כפי שהוא יכול להשפיע על מורפולוגיה השטח הסופית של קרומי.
- רוכש תמונות עם חשיפה משקפת הומוגנית של ברק. לחלופין, לצלם תמונה של pellicles עם מצלמה ברזולוציה גבוהה. כדי למנוע חפצים הנגרמים על ידי inconsisזוויות וצללי אוהל אור, לצלם מלמעלה למטה עם המצלמה הקבועה על חצובה ולהשתמש מקור אור רך גדול ב 45 מעלות משני הצדדים.
הערה: שיטה חלופית כדי ללמוד B. subtilis רב-תאיות הוא assay המושבה ביופילם על המדיום MSgg מוצק, וישכנע ביופילם. כמו pellicles, assay זה מאפשר לימוד של תהליכים spatiotemporal. ברגע בטווח הפעיל של מעכבי מולקולה קטנים נקבע, והשפיעו על היווצרות מושבת biofilm ניתן ללמוד. - לגדול מושבות ביופילם, לזהות באופן סימטרי 1.5 μl של תרבות מראש רחוץ (שלב 1.7) על צלחת אגר MSgg 1.5% יבשים בעזרת תבנית - 4 טיפות לכל צלחת פטרי בקוטר 8.5 ס"מ. תנו טיפות לספוג לצלחת לפני שעבר עליהם.
הערה: התבנית עוזרת לקבל חלוקה שווה של מושבות ביופילם בתוך האזור שבו התאים גדלים. כדי להכין את התבנית, לצייר את השטח הכולל של צמיחה בסקלות מקורית, לחלק אותו הכת שווהORS ולסמן במרכז. עבור צלחת פטרי עגול בקוטר 8.5 ס"מ, זה מקצה 14 ס"מ 2 למושבה ביופילם אחד. - דגירת הצלחות ב 30 מעלות צלזיוס למשך שלושה ימים. במהלך תקופה זו, מושבות ביופילם לפתח ליצור מבנה תלת-ממדי, מקומט.
- צלם תמונות כמו בשלב 1.11.
2. Assay ההתנגדות אתנול
- לגדול biofilms כמתואר בשלבים 1.1-1.7 ו 1.12-1.14.
- לאחר 68 שעות של צמיחה ב 30 ° C, לחתוך את המושבות ביופילם לשני חלקים שווים בעזרת סכין גילוח ואת התבנית.

דוגמא איור 2. עיצוב ניסיוני כדי להעריך את ההתנגדות של תאי מושבה ביופילם לסוכני חיטוי. (א) תבנית המשמשת לחלוקה השוויונית של מושבות ביופילם על פני צלחת פטרי לחיתוך. (ב)תמונות מלמעלה למטה של ביופילם wild-type מטופל גדלו במשך 68 שעות על מדיום MSgg וישכנע ביופילם מוצק, מוגדר על 30 מעלות צלזיוס. ההגדלה מראה כיצד מושבה biofilm ניתן לחתוך לשני חצאים שווים. (ג) שני חצאים ביופילם שווה זוכים ליחס שווה (שליטה, PBS) או עם או PBS או עיקור סוכן ומעובד כמתואר. סרגל קנה מידה:. 1 ס"מ אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
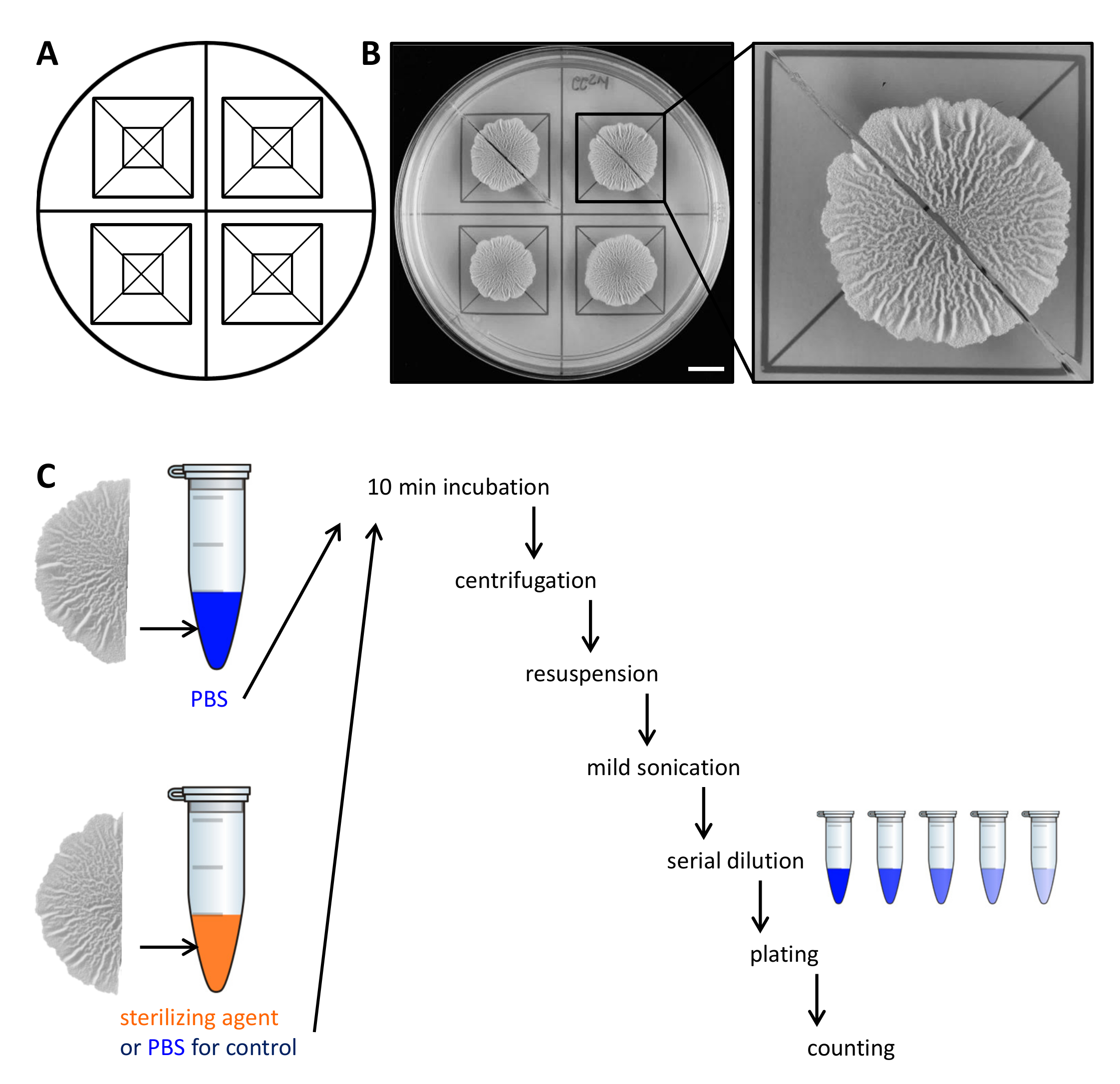{kind=link}
- הרם בזהירות כל מחצית של המושבה ביופילם מהצלחת אגר עם מרית קטנה ולהעביר אותו צינור 1.5 מ"ל microcentrifuge המכיל 500 μl של תמיסת מלח שנאגרו פוספט (PBS). במידת הצורך, לגרד את התאים הנותרים מהצלחת ולהעבירם צינור microcentrifuge כמו גם.
הערה: המחצית השנייה של המושבה ביופילם מטופל באופן דיפרנציאלי, תלוי אם זה הוא שליטה או כדי לבדוק את עמידות Steriסוכני lizing. - על הבקרה, דגירה במחצית השנייה של המושבה biofilm ב 500 μl PBS כמו בשלב 2.3. כדי להעריך התנגדות לסוכנים עיקור, להעביר את החצי השני של המושבה ביופילם 500 μl 50% (V / V) אתנול.
הערה: סוכני חיטוי אלטרנטיבי כגון חומצת נתרן יכולים לשמש. עבור כל הסוכנים לסטריליזציה בשימוש, לקבוע את זמן ריכוז דגירה הפעיל ניסוי ראשוני. - דגירה המושבות ביופילם במשך 10 דקות על העליונה הספסל בטמפרטורת החדר.
- צנטריפוגה מושבות ביופילם במשך 5 דקות ב 18,000 XG ובזהירות להסיר את supernatant עם טפטפת. הוספת 300 μl של PBS.
- Sonicate התאים מעט (sec 10%, דופק 5 משרעת) עם microtip של sonicator.
הערה: אנרגית sonication חייבת להיות מספיק כדי אגרגטים ביופילם נפרדים. עם זאת, sonication קשה מדי עלול lyse התאים. אשר מראש על ידי מיקרוסקופ אור שהאנרגיה sonicationבשימוש לא lyse התאים ושכל אגרגטים הם מומסים. - הוסף 700 μl של PBS לנפח סופי של 1 מ"ל. בצע דילול סדרתי (10 -7) ב PBS ולהתפשט 100 μl של 3 דילולים על צלחת אגר LB-1.5% באמצעות חרוזי זכוכית סטרילית.
הערה: הדילולים האופטימליים להיות מצופות צריכה להיקבע בניסוי ראשוני, כמו זה תלוי בכמות של תאים במושבה ביופילם הריבית לבין שיעור ההישרדות של התאים בתגובת סוכן החיטוי. - דגירה צלחות לילה ב 30 מעלות צלזיוס, לספור את CFU ולקבוע CFU / ml. מתוך CFU / ml הסופי של כל המושבות ביופילם וחצי, לחשב את אחוז הניצולים.
הערה: כאשר מבוצע ונותח כמתואר, שני חצי המושבה המלאה ביופילם ואת המטופל מול החצי שטופל אתנול של המושבה ביופילם המטופל אמור להניב הבדלים מתחת ל -10% בספירת תא קיימא, אימות הסימטריה או ההתנגדות של המושבה , respectively. לחלופין, התוצאות יכולות להיות מיוצגות הכולל CFU. ספירת התאים של מלאת המושבה ביופילם מטופל צריכה להישאר באותו סדר הגודל. לעומת זאת, ספירת תאי המחצית שטופל אתנול של מושבה ביופילם מולקולה שטופלה קטנה צפויות לרדת ידי מינימום של שני סדרי הגודל לתבוע רגישות מוגברת סוכן החיטוי.
3. הכנת דוגמאות מושבת Biofilm עבור במיקרוסקופ אלקטרונים סורק
- לגדול מושבות ביופילם כמתואר בשלבים 1.1-1.7 ו 1.12-1.14.
- הכן קבוצה חדשה של 2% (v / v) glutaraldehyde, 3% (v / v) פתרון paraformaldehyde ב cacodylate נתרן 100 מ"מ, 5 מ"מ חיץ סידן כלורי, pH 7.3. הכן 5 מ"ל של מקבע לכל צלחת פטרי בקוטר 8.5 ס"מ.
זהירות: Glutaraldehyde ו paraformaldehyde הם מסוכנים. לטפל בהם עם ציוד בטיחות בתוך במנדף כימי. מחק את הפתרונות ואת החומרים מזוהמים אל האזפסולת ardous. - בזהירות להוסיף מקבע למושבות ביופילם, ללא מחלק ישירות על גבי biofilms.
הערה: עקב האופי ההידרופובי של המושבה ביופילם, המושבות לאט להתנתק אגר ולהתחיל לצוף. - בזהירות לאטום את הצלחות עם פס של Parafilm. דגירה על שייקר רוטרי 2 שעות בטמפרטורת החדר ולאחר מכן להעביר את הצלחות עד 4 מעלות צלזיוס למשך לילה.
- למחרת, להסיר בזהירות את הנוזל עם טפטפת זכוכית פסטר מחובר משאבת ואקום.
- בזהירות להוסיף 10 מ"ל 100 מ"מ cacodylate נתרן, 5 מ"מ חיץ סידן כלורי לשטוף את ביופילם דגירה במשך 5 דקות. הוצא בעדינות את הנוזל עם טפטפת זכוכית פסטר מפינת הצלחת כדי למנוע נזק ביופילם ולהוסיף פתרון הכביסה טריים ידי pipetting עדין. חזור על פעולה זו פעם.
- עבור ההתייבשות של המושבות ביופילם, המשך בביצוע השלבים הבאים: 2x 5 דקות DDH 2 O; 2x 20 דקות 30% אתנול; 2x 20 דקות ב 50% אתנול; 2x 20 דקות ב 70% אתנול; 2x 20 דקות ב -96% אתנול; 2x 30 דקות ב 100% אתנול.
- הוסף 15 מ"ל של נוזל על כל צלחת פטרי בקוטר 8.5 ס"מ כל צעד ולהסיר את הנוזל בזהירות אחרי כל הדגירה.
- השתמש באחת משתי שיטות שונות לייבוש דגימות אתנול.
- עבור אוויר-ייבוש מן אתנול:
- חותכים נייר סינון תאית (בקוטר של 9 ס"מ) ברבעונים. בקצרה לצלול רבע ב 100% אתנול, ולאחר מכן בזהירות להעביר את אחד מושבה ביופילם צף על גבי זה. מכניס את נייר הסינון הרטוב בצלחת פטרי מרופדת נייר סינון. מכסים את צלחת פטרי ולתת מושבות ביופילם לילה יבש במנדף כימי.
- עבור נקודה קריטית (CP) -drying באמצעות פחמן דו-חמצני (CO 2) כמו נוזל המעבר:
- מלאו 75% של חדר מכונת ייבוש נקודה הקריטי עם אתנול 100%. העברת דגימות לתוך מחזיק, כל דגימה לתוך ג משלהhamber. במידת הצורך, לחתוך את biofilm עם מספריים לחתיכות קטנות. השאר את הדגימות שקועות אתנול במהלך כל הטיפול. ואז, להעביר את בעל ההחדרה ולסגור את החדר בחוזקה.
- מצננים את החדר כדי C 7 ° ולהתחיל ערבוב. ממלא את החדר לחלוטין עם נוזל CO 2. במהלך זמן דגירה 7 דקות, לתת לתערובת אתנול עם CO 2. ואז, פרוק 25% מהפתרון.
הערה: אין לרוקן את התא מתחת לרמה של המדגם. - חזור על שלב 3.8.2.2 ארבע פעמים.
- חזור על שלב 3.8.2.2 חמש פעמים עם זמן דגירה של 5 דקות בלבד. לבסוף, אתנול צריך להיות מוחלף לחלוטין על ידי ה- CO 2.
- בסבב האחרון, ריק רק 5% של החדר. כבה את הבחישה ואת הקירור. התחל חימום בתא עד 42 מעלות צלזיוס. בטמפרטורה של 31.1 מעלות צלזיוס, בלחץ של 73.9 בר, CO 2 הנוזלים מגיעים לנקודה הקריטית שלה, המדינה שבה ph הגזיםASE יש אותו הצפיפות כשלב הנוזלי של 27 הממס. ברגע שהטמפרטורה מגיעה 42 ° C, דגירה של 10 דקות. בגיל 42 ° C, CO 2 בתא קיים כגז סופר קריטי.
הערה: כל זמן לבדוק את הלחץ של החדר. הלחץ לא יעלה על 120 בר ב 42 מעלות צלזיוס. - התחל כדי לשחרר את הגז לאט עם חימום רציף. פעולה זו שומרת על דגימות בשלב CO 2 -gas ומונע את העיוות של מורפולוגיה המדגם דרך מתח פני הנוזל. הגדר את flowmeter עד 5 L / hr ידי כוונון עדין על שסתום ויסות שליטה על מד הזרימה. המתן עד כל הלחץ בתא הוא שוחרר. עכשיו לפתוח את התא ולהסיר את הדגימות בזהירות מבעל.
- עבור אוויר-ייבוש מן אתנול:
- מעיל בדל במיקרוסקופ אלקטרונים עם קלטת פחמן. בעזרת פינצטה, בזהירות להעביר את המושבות ביופילם על הבדל. חבר כל מושבה אל הבדל ידי הוספת גשר דק מן המכוניתbon קלטת, שהוא חיוני עבור חיסול תשלום תחת אלומת אלקטרונים. בשלב זה, להתמודד עם המושבות ביופילם בזהירות כפי שהם מאוד שבירים. אחסן את דגימות ייבוש לפחות 24 שעות או עד בדיקה.
- ביום הבדיקה עם מיקרוסקופ אלקטרונים סורק, גמגום מעיל המושבות ביופילם עבור 2 דקות בתוך זווית 60 ° בתוך coater-גמגום זהב-פלדיום. חזור על פעולה זו פעמיים וסובב את הדגימות על ידי 120 ° שביניהם. בסוף, גמגום מעיל דגימות פעם במשך 3 דקות מלמעלה. 20 ננומטר שכבה דקה של פלדיום זהב משפר את המוליכות ומשפר את הניגודיות של המדגם עבור הדמיה של SEM.
- אחסן את דגימות ייבוש להימנע ההתייבשות של המדגם 28 עד הדמיה באמצעות מיקרוסקופ אלקטרונים סורק 29,30.
תוצאות
את assay הקרומית היא אחת שיטות כדי לחקור את תהליכי פיקוח הדוק והדינמיים של B. subtilis רב-תאיות. מלבד זאת, assay הקרומית מתאימה לבדוק מגוון של תנאים או טרום פתיחה או ריכוזי מולקולה קטנים בצלחת multidish תא-תרבות אחת בניסוי אחד. עם זאת, ב ' ההיווצרות קרומית subtilis רגישה ?...
Discussion
Subtilis Bacillus צורות חזקה מובנה מאוד biofilms הוא נוזל (pellicles) ועל בינוני מוצק (מושבות). לפיכך, היא משמשת אורגניזם מודל אידיאלי לאפיין את מצב הפעולה של מעכבי biofilm ספציפיים. על תקשורת מוצקה, תאי טופס מבנים רבים-תאי עם מאפייני בולטים שאינם ניכרים pellicles, כמו קמטים הקורנים מהמרכ?...
Disclosures
The authors have nothing to disclose.
Acknowledgements
Electron microscope imaging was conducted at the Electron Microscopy Unit of the Weizmann Institute of Science, supported in part by the Irving and Cherna Moskowitz Center for Nano and Bio-Nano Imaging. This research was also supported by the ISF I-CORE grant 152/1, Mr. and Mrs. Dan Kane, Ms. Lois Rosen, by a Yeda-Sela research grant, by the Larson Charitable Foundation, by Ruth and Herman Albert Scholars Program for New Scientists, by the Ilse Katz Institute for Materials Sciences and Magnetic Resonance Research grant, by the Ministry of Health grant for alternative research methods, and by the France-Israel Cooperation - Maimonide-Israel Research Program. IKG is a recipient of the Rowland and Sylvia Career Development Chair.
Materials
| Name | Company | Catalog Number | Comments |
| Luria Broth, Lennox | Difco | 240230 | |
| Bacto Agar | Difco | 214010 | |
| potassium phosphate monobasic | Sigma, 136.09 g/mol | P0662-500G | |
| potassium phosphate dibasic | Fisher Scientific, 174.18 g/mol | BP363-1 | |
| 3-(N-morpholino)propanesulfonic acid | Fisher Scientific, 209.27 g/mol | BP308-500 | |
| magnesium chloride hexahydrate | Merck, 203.30 g/mol | 1.05833.0250 | |
| calcium chloride anhydrous | J.T. Baker, 110.98 g/mol | 1311-01 | |
| manganese(II) chloride tetrahydrate | Sigma, 197.91 g/mol | 31422-250G-R | |
| iron(III) chloride hexahydrate | Sigma, 270.30 g/mo) | F2877-500G | |
| zinc chloride anhydrous | Acros Organics, 136.29 g/mol | 424592500 | |
| thiamine hydrochloride | Sigma, 337.27 g/mol | T1270-100G | |
| L-tryptophan | Fisher Scientific, 204.1 g/mol | BP395-100 | |
| L-phenylalanine | Sigma, 165.19 g/mol | P5482-100G | |
| L-threonine | Sigma, 119.12 g/mol | T8625-100G | |
| glycerol anhydrous | Bio-Lab Itd | 712022300 | |
| L-glutamic acid monosodium salts hydrate | Sigma, 169.11 g/mol | G1626-1KG | |
| D-leucine | Sigma, 169.11 g/mol | 855448-10G | |
| ethanol anhydrous | Gadot | 830000054 | |
| razor blade | Eddison | NA | |
| circular cellulose filter papers | Whatman, 90 mm | 1001-090 | |
| glutaraldehyde | EMS (Electron Micoscopy Science), 25% in water | 16220 | |
| paraformaldehyde | EMS, 16% in water | 15710 | |
| sodium cacodylate | Merck, 214.05 g/mol | 8.2067 | |
| calcium chloride 2-hydrate | Merck, 147.02 g/mol | 1172113 | |
| stub-aluminium mount | EMS, sloted head | 75230 | |
| carbon adhesive tape | EMS | 77825-12 | |
| Shaker 37 °C | New Brunswick Scientific Innowa42 | NA | |
| Centrifuge | Eppendorf table top centrifuge 5424 | NA | |
| Digital Sonifier, Model 250, used with Double Step Microtip | Branson | NA | |
| Incubator 30 °C | Binder | NA | |
| Incubator 23 °C | Binder | NA | |
| Filter System, 500 ml, polystyrene | Cornig Incorporated | NA | |
| Rotary Shaker - Orbitron Rotatory II | Boekel | NA | |
| S150 Sputter Coater | Edwards | NA | |
| CPD 030 Critical Point Dryer | BAL-TEC | NA | |
| Environmental Scanning Electron Microscope | XL30 ESEM FEG Philips (FEI) | NA |
References
- Branda, S. S., Vik, S., Friedman, L., Kolter, R. Biofilms: the matrix revisited. Trends Microbiol. 13, 20-26 (2005).
- Stoodley, P., Sauer, K., Davies, D. G., Costerton, J. W. Biofilms as complex differentiated communities. Annu Rev Microbiol. 56, 187-209 (2002).
- Miller, M. B., Bassler, B. L. Quorum sensing in bacteria. Annu Rev Microbiol. 55, 165-199 (2001).
- Aguilar, C., Vlamakis, H., Losick, R., Kolter, R. Thinking about Bacillus subtilis as a multicellular organism. Curr Opin Microbiol. 10, 638-643 (2007).
- Kolter, R., Greenberg, E. P. Microbial sciences: the superficial life of microbes. Nature. 441, 300-302 (2006).
- Kearns, D. B., Chu, F., Branda, S. S., Kolter, R., Losick, R. A master regulator for biofilm formation by Bacillus subtilis. Mol Microbiol. 55, 739-749 (2005).
- Branda, S. S., Chu, F., Kearns, D. B., Losick, R., Kolter, R. A major protein component of the Bacillus subtilis biofilm matrix. Mol Microbiol. 59, 1229-1238 (2006).
- Romero, D., Aguilar, C., Losick, R., Kolter, R. Amyloid fibers provide structural integrity to Bacillus subtilis biofilms. Proc Natl Acad Sci USA. 107, 2230-2234 (2010).
- Kobayashi, K., Iwano, M. BslA(YuaB) forms a hydrophobic layer on the surface of Bacillus subtilis biofilms. Mol Microbiol. 85, 51-66 (2012).
- Hobley, L., et al. BslA is a self-assembling bacterial hydrophobin that coats the Bacillus subtilis biofilm. Proc Natl Acad Sci USA. 110, 13600-13605 (2013).
- Romero, D., Vlamakis, H., Losick, R., Kolter, R. An accessory protein required for anchoring and assembly of amyloid fibres in B. subtilis biofilms. Mol Microbiol. 80, 1155-1168 (2011).
- Kolodkin-Gal, I., et al. D-amino acids trigger biofilm disassembly. Science. 328, 627-629 (2010).
- Chan, Y. G., Kim, H. K., Schneewind, O., Missiakas, D. The capsular polysaccharide of Staphylococcus aureus is attached to peptidoglycan by the LytR-CpsA-Psr (LCP) family of enzymes. J Biol Chem. 289, 15680-15690 (2014).
- Mielich-Suss, B., Lopez, D. Molecular mechanisms involved in Bacillus subtilis biofilm formation. Environ Microbiol. 17, 555-565 (2014).
- Cairns, L. S., Hobley, L., Stanley-Wall, N. R. Biofilm formation by Bacillus subtilis: new insights into regulatory strategies and assembly mechanisms. Mol Microbiol. 93, 587-598 (2014).
- Chen, M., Yu, Q., Sun, H. Novel strategies for the prevention and treatment of biofilm related infections. Int J Mol Sci. 14, 18488-18501 (2013).
- Bucher, T., Oppenheimer-Shaanan, Y., Savidor, A., Bloom-Ackermann, Z., Kolodkin-Gal, I. Disturbance of the bacterial cell wall specifically interferes with biofilm formation. Environ Microbiol Rep. 7, 990-1004 (2015).
- Sarkar, S., Pires, M. M. D-Amino acids do not inhibit biofilm formation in Staphylococcus aureus. PLoS One. 10, e0117613 (2015).
- Wei, W., Bing, W., Ren, J., Qu, X. Near infrared-caged D-amino acids multifunctional assembly for simultaneously eradicating biofilms and bacteria. Chem Commun (Camb). 51, 12677-12679 (2015).
- Leiman, S. A., et al. D-amino acids indirectly inhibit biofilm formation in Bacillus subtilis by interfering with protein synthesis. J Bacteriol. 195, 5391-5395 (2013).
- Costerton, J. W., Stewart, P. S., Greenberg, E. P. Bacterial biofilms: a common cause of persistent infections. Science. 284, 1318-1322 (1999).
- Davies, D. Understanding biofilm resistance to antibacterial agents. Nat Rev Drug Discov. 2, 114-122 (2003).
- Olsen, I. Biofilm-specific antibiotic tolerance and resistance. Eur J Clin Microbiol Infect Dis. 34, 877-886 (2015).
- Tseng, B. S., et al. The extracellular matrix protects Pseudomonas aeruginosa biofilms by limiting the penetration of tobramycin. Environ Microbiol. 15, 2865-2878 (2013).
- Branda, S. S., Gonzalez-Pastor, J. E., Ben-Yehuda, S., Losick, R., Kolter, R. Fruiting body formation by Bacillus subtilis. Proc Natl Acad Sci USA. 98, 11621-11626 (2001).
- Holscher, T., et al. Motility, Chemotaxis and Aerotaxis Contribute to Competitiveness during Bacterial Pellicle Biofilm Development. J Mol Biol. 427, 3695-3708 (2015).
- Bray, D. . Methods in Biotechnology. 13, 235-243 (2000).
- Ensikat, H. J., Ditsche-Kuru, P., Barthlott, W. . Scanning electron microscopy of plant surfaces: simple but sophisticated methods for preparation and examination. 1, 248-255 (2010).
- Hayat, M. A. . Principles and techniques of scanning electron microscopy: Biological applications. 2, (1976).
- Schatten, H. . Scanning Electron Microscopy for the Life Sciences. , (2013).
- Bridier, A., Meylheuc, T., Briandet, R. Realistic representation of Bacillus subtilis biofilms architecture using combined microscopy (CLSM, ESEM and FESEM). Micron. 48, 65-69 (2013).
- Boyde, A., MacOnnachie, E. Volume changes during preparation of mouse embryonic tissue for scanning electron microscopy. SCANNING. 2, 149-163 (1979).
- Yao, Z., Kahne, D., Kishony, R. Distinct single-cell morphological dynamics under beta-lactam antibiotics. Mol Cell. 48, 705-712 (2012).
- Epstein, A. K., Pokroy, B., Seminara, A., Aizenberg, J. Bacterial biofilm shows persistent resistance to liquid wetting and gas penetration. Proc Natl Acad Sci USA. 108, 995-1000 (2011).
- Vlamakis, H., Chai, Y., Beauregard, P., Losick, R., Kolter, R. Sticking together: building a biofilm the Bacillus subtilis way. Nat Rev Microbiol. 11, 157-168 (2013).
- Shemesh, M., Chai, Y. A combination of glycerol and manganese promotes biofilm formation in Bacillus subtilis via histidine kinase KinD signaling. J Bacteriol. 195, 2747-2754 (2013).
- Kolodkin-Gal, I., et al. Respiration control of multicellularity in Bacillus subtilis by a complex of the cytochrome chain with a membrane-embedded histidine kinase. Genes Dev. 27, 887-899 (2013).
- Oppenheimer-Shaanan, Y., et al. Spatio-temporal assembly of functional mineral scaffolds within microbial biofilms. npj Biofilms and Microbiomes. 2, 15031 (2016).
- Garcia-Betancur, J. C., Yepes, A., Schneider, J., Lopez, D. Single-cell analysis of Bacillus subtilis biofilms using fluorescence microscopy and flow cytometry. J Vis Exp. , e3796 (2012).
- Bogino, P. C., Oliva Mde, L., Sorroche, F. G., Giordano, W. The role of bacterial biofilms and surface components in plant-bacterial associations. Int J Mol Sci. 14, 15838-15859 (2013).
- Fratamico, P. M., Annous, B. A., Guenther, N. W. . Biofilms in the Food and Beverage Industires. 1, (2009).
- Gao, G., et al. Effect of biocontrol agent Pseudomonas fluorescens 2P24 on soil fungal community in cucumber rhizosphere using T-RFLP and DGGE. PLoS One. 7, e31806 (2012).
- Chen, Y., et al. Biocontrol of tomato wilt disease by Bacillus subtilis isolates from natural environments depends on conserved genes mediating biofilm formation. Environ Microbiol. 15, 848-864 (2013).
- Bryers, J. D. Medical biofilms. Biotechnol Bioeng. 100, 1-18 (2008).
- Logan, B. E. Exoelectrogenic bacteria that power microbial fuel cells. Nat Rev Microbiol. 7, 375-381 (2009).
- Nevin, K. P., Woodard, T. L., Franks, A. E., Summers, Z. M., Lovley, D. R. Microbial electrosynthesis: feeding microbes electricity to convert carbon dioxide and water to multicarbon extracellular organic compounds. MBio. 1, (2010).
- Torres, C. I., et al. A kinetic perspective on extracellular electron transfer by anode-respiring bacteria. FEMS Microbiol Rev. 34, 3-17 (2010).
- Li, J., Wang, N. Foliar application of biofilm formation-inhibiting compounds enhances control of citrus canker caused by Xanthomonas citri subsp. citri. Phytopathology. 104, 134-142 (2014).
- Okegbe, C., Price-Whelan, A., Dietrich, L. E. Redox-driven regulation of microbial community morphogenesis. Curr Opin Microbiol. 18, 39-45 (2014).
- Mann, E. E., Wozniak, D. J. Pseudomonas biofilm matrix composition and niche biology. FEMS Microbiol Rev. 36, 893-916 (2012).
- Bouffartigues, E., et al. Sucrose favors Pseudomonas aeruginosa pellicle production through the extracytoplasmic function sigma factor SigX. FEMS Microbiol Lett. 356, 193-200 (2014).
- Wu, C., Lim, J. Y., Fuller, G. G., Cegelski, L. Quantitative analysis of amyloid-integrated biofilms formed by uropathogenic Escherichia coli at the air-liquid interface. Biophys J. 103, 464-471 (2012).
- Serra, D. O., Richter, A. M., Hengge, R. Cellulose as an Architectural Element in Spatially Structured Escherichia coli Biofilms. J Bacteriol. 195, 5540-5554 (2013).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved