
A subscription to JoVE is required to view this content. Sign in or start your free trial.
Method Article
קביעת פני התא יחסית והביטוי סה"כ של תעלות יונים רקומביננטי שימוש cytometry זרימה
In This Article
Summary
הפרעות בקצב לב בירושה לעתים קרובות נגרמים על ידי מוטציות המשנות את מסירת השטח של תעלות יונים אחד או יותר. הנה, אנחנו מתאימים זרימת cytometry מבחני לספק כימות של ביטוי החלבון הכולל ותא יחסית משטח של תעלות יונים רקומביננטי לידי ביטוי בתאים TSA-201.
Abstract
Inherited or de novo mutations in cation-selective channels may lead to sudden cardiac death. Alteration in the plasma membrane trafficking of these multi-spanning transmembrane proteins, with or without change in channel gating, is often postulated to contribute significantly in this process. It has thus become critical to develop a method to quantify the change of the relative cell surface expression of cardiac ion channels on a large scale. Herein, a detailed protocol is provided to determine the relative total and cell surface expression of cardiac L-type calcium channels CaV1.2 and membrane-associated subunits in tsA-201 cells using two-color fluorescent cytometry assays. Compared with other microscopy-based or immunoblotting-based qualitative methods, flow cytometry experiments are fast, reproducible, and large-volume assays that deliver quantifiable end-points on large samples of live cells (ranging from 104 to 106 cells) with similar cellular characteristics in a single flow. Constructs were designed to constitutively express mCherry at the intracellular C-terminus (thus allowing a rapid assessment of the total protein expression) and express an extracellular-facing hemagglutinin (HA) epitope to estimate the cell surface expression of membrane proteins using an anti-HA fluorescence conjugated antibody. To avoid false negative, experiments were also conducted in permeabilized cells to confirm the accessibility and proper expression of the HA epitope. The detailed procedure provides: (1) design of tagged DNA (deoxyribonucleic acid) constructs, (2) lipid-mediated transfection of constructs in tsA-201 cells, (3) culture, harvest, and staining of non-permeabilized and permeabilized cells, and (4) acquisition and analysis of fluorescent signals. Additionally, the basic principles of flow cytometry are explained and the experimental design, including the choice of fluorophores, titration of the HA antibody and control experiments, is thoroughly discussed. This specific approach offers objective relative quantification of the total and cell surface expression of ion channels that can be extended to study ion pumps and plasma membrane transporters.
Introduction
מאמר זה מספק assay אמין לדווח הביטוי שטח פני התא היחסי של חלבונים בממברנה כגון תעלות יונים לידי ביטוי בתאים רקומביננטי באמצעות טכנולוגית cytometry הזרימה הקיימת. תעלות יונים הן חלבונים בממברנה נקבוביים יוצרים שאחראים לשליטת אותות חשמליים על ידי gating את זרימת יונים על פני קרום התא. הם מסווגים על ידי מנגנון הפעלה, טבע, סלקטיביות של מינים יון העוברים דרך הנקבוביות שבו הם נקודתיים. ברמה התאית ורקמות, והנתיבים יון מקרוסקופית דרך תעלות יונים הם תוצר של biophysical (gating ו חלחול), ביוכימיות (זירחון), ו biogenesis (סינתזה, glycosylation, סחר, השפלה) נכסים 1. כל התהליכים הללו הוא ייחודי לכל סוג של תעלות יונים, והוא מותאם למלא את התפקיד הפיזיולוגי של ערוץ יון. כתוצאה מכך, שינויים בכל תהליכים המכוילים אלה באמצעותבירושה או הנדסה גנטית, המכונה לעתים קרובות "channelopathy", יכול להיות מזיק הומאוסטזיס תא. חשוב להדגיש, כי מסירת הסכום "הנכון" של תעלות יונים על פני התא היא קריטית הומאוסטזיס תא. גם עליות קטנות (רווח של פונקציה) וירידה קטנה (הפסד של פונקציה) בפעילות ערוץ יון יש פוטנציאל לגרום פתולוגיה רצינית לאורך החיים. פגמי המשלוח פני התא של תעלות יונים בוגרות הוא הקובע חשוב channelopathies רב, כגון סיסטיק פיברוזיס (ערוץ יון CFTR) 2 ו הפרעות בקצב לב של טופס תסמונת QT הארוך (תעלות אשלגן לב) 3.
Channelopathies המשויכים מוות פתאומי של הלב 4. שהשכיחות העולמית הנוכחית של כל channelopathies הלב נחשב לפחות 1: 2,000-1: 3,000 לנפש 5 ואחראים כמחצית ca למוות קרדיאלי פתאומי בקצב לא סדירses 6. תפקוד לקוי של נתרן מתח מגודר לב, potassium-, ו בסידן תעלות יונים סלקטיבי ידועות לשחק תפקיד מרכזי בתהליך זה. The L-סוג Ca V 1.2 תעלות סידן מתח מגודרת נדרש ליזום התכווצות שריר הלב מסונכרן. הלב L- סוג Ca V 1.2 ערוץ הינו קומפלקס חלבון רב-למקטע מורכב למקטע α1 הראשי יוצרי הנקבוביות Ca V ו- CA V SS ו- CA V α2δ1 יחידות משנה עזר 7-12. ראוי לציין, כי ההשלמה המלאה של יחידות משנת עזר נדרשת לייצר ערוצי Ca V 1.2 פעילות לפי קרום הפלזמה ואינטראקציות דינמיות בין יחידות המשנה אלה חיוניות כדי לתמוך בפונקציה החשמלית הרגילה של הלב 13. Ca V SS מקדם את הביטוי על פני קרום התא של Ca V 1.2 ערוצים באמצעות אינטראקציה הידרופובי nanomolar הלא קוולנטיים 14. שיתוף ביטוי של wi למקטע Ca V α2δ1ה Ca V SS הנכנס Ca V α1 מגרה ביטוי השיא הנוכחי (5 עד פי 10) ומקדם ההפעלה ערוץ במתח שלילי יותר. קבל של פונקצית מוטציות של למקטע יוצרי הנקבוביות Ca V 1.2 נקשר עם צורה של הפרעות קצב חדריות שנקראת תסמונת QT הארוך 15 בעוד שורה של מוטציות נקודתיות בשלוש היחידות משנה הראשיים להרכיב ערוץ L- סוג Ca V 1.2 זוהה במקצועות הסובלים הפרעות קצב של טופס תסמונת QT הקצר 16,17. תעלות יונים הן חלבוני בממברנה שניתן לחקור מנקודת מבט ביוכימית (כימית חלבון) או באמצעות כלי אלקטרו (מכונה ייצור הנוכחי) ולעתים קרובות תוך שימוש בגישות משלימות אלה. אלקטרופיזיולוגיה, ב תיקון- clamping כל תא מסוים, היא פתרון ראוי להבהיר את הפונקציה של תעלות יוני 15 אבל לא יכולה לפתור שינויים בסחר חלבון משינויי biophysical שלהםנכסים. כימית חלבון יש, עם זאת, לעתים קרובות שימוש אפשרי מוגבלת בשל הביטוי הנמוך היחסי של חלבונים בממברנה גדולים ביחס חלבונים מסיסים קטנים. שיטות תפוקה גבוהה חזקות באמצעות הודעת קרינה צריכות להיות מפותחות על מנת לתת מענה ליקויים דווקא biogenesis חלבון גורמים לשינויי הביטוי פני התא של תעלות יונים.
Cytometry זרימה היא טכנולוגיה biophysical מועסק ספירת תאים, מיון, איתור סמן ביולוגי, והנדסת חלבון 18. כאשר פתרון מדגם של תאי חיים או חלקיקים מוזרק לתוך cytometer זרימה, התאים מסודרים לזרם יחיד שיכול להיות נחקר על ידי מערכת זיהוי של המכונה (איור 1). התזרים הראשון cytometer המכשיר המיוצר ב -1956 19 זוהה רק פרמטר אחד אבל cytometers הזרימה המודרני יש לייזרים מרובים גלאי קרינה המאפשרים זיהוי של יותר מ -30 פרמטרי ניאון 20,21.מסננים ומראות (אופטיקה פליטה) לכוון את פיזור האור או אור הניאון של תאי רשת אלקטרונית (פוטודיודה וגלאים) הממירה את אור יחסי עוצמתו. נתונים דיגיטליים מנותחים באמצעות תוכנות מיוחדות הפלט העיקרי מוצג כמזימת נקודה 21.

איור 1:. עקרונות Biophysical של זרימת cytometry מיון תאים בודדים נדחפים דרך נחיר בלחץ גבוה בתוך זרם של נוזל נדן אשר נע אותם על אחד או יותר נקודות החקירה לייזר. אלומת האור היא מוסחת על ידי התאים עוברים והאור שנאסף בכיוון קדימה (Forward הפיזור, FCS) נשלח פוטודיודה הממיר האור לאות ביחס ישירה לגודל של התא. האור נאסף גם בזווית של 90 ° לנתיב לייזר ונשלח גלאי (המכונה גם photomultipliers (PMT)).אור זה מנותב דרך מראות dichroic המאפשרות זיהוי של אות הפיזור בצד (SSC), המשקפת את הגרעיניות בתוך התאים, ואת פליטת פלורסנט אם fluorochromes הנרגש נוכחים בתא. שלושה גלאים (ירוק, צהוב, אדום) מיוצגים עם מסננים bandpass אורך גל שונה, מה שמאפשר זיהוי סימולטני של fluorochromes שונים. האותות השונים דיגיטציה ידי למחשב חיצוני והוסבו נתונים שינותחו לכמת את המאפיינים של התאים. אנא לחצו כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
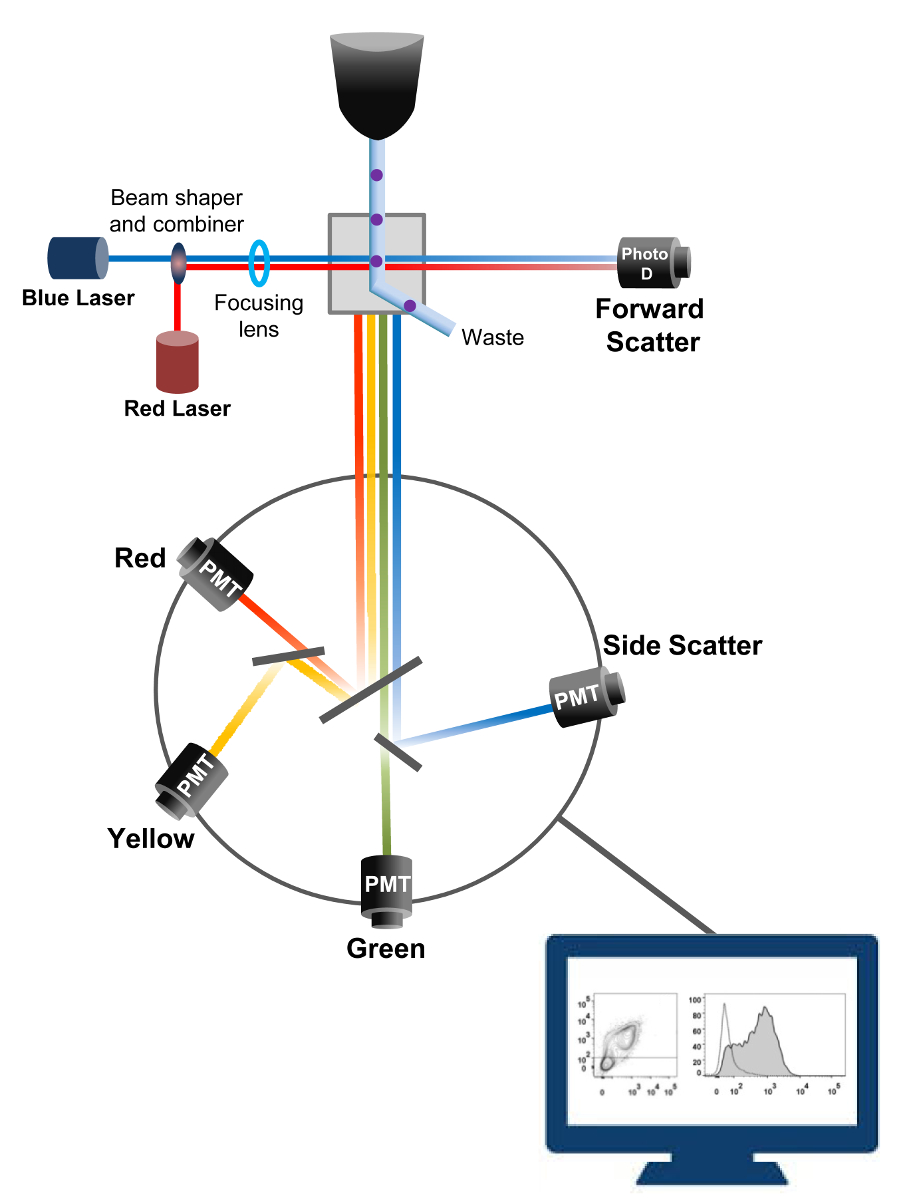{kind=link}
קיבולת התפוקה הגבוהה של cytometers הזרימה נוצלה לכמת את ביטוי הקרום היחסי של סוג-בר רקומביננטי מתח מגודר סחר מחסר L- סוג Ca V 1.2 ערוצים יחידים משנה הקשורים בתאים חיים. בונת cDNA שיתוףדינג עבור החלבונים תויגו כפליים לבצע אפיטופ ללא ניאון תאי זמנית שיכול להיות מזוהה על ידי נוגדן מצומדות ניאון בלתי חדיר ו fluorophore תאי כי הוא פלורסנט constitutively. הן epitope התאית, הוכנסה בלופ תאי של החלבון, ואת fluorophore התאי, מוכנסת לאחר C- הסופית, מתורגם עם החלבון. בסדרה זו של ניסויים, חלבון Ca V α2δ1 תוכנן להביע hemagglutinin התאי epitope (HA) (YPYDVPDYA) זוהה על ידי (isothiocyanate והעמסה) FITC בלתי חדירה, מצומדות אנטי-HA ו mCherry כמו fluorophore התאי המהותי. כדי לקבוע את רמת הביטוי שטח פני התא היחסי של α2δ1 V mCherry-Ca HA-tagged חלבון, תאים רקומביננטי המבטאים את החלבון היתוך נבצרו לאחר transfection, ומוכתם antibod תג FITC- מצומדות העכבר חד שבטיים אנטי-HA epitopey (איור 2). FITC הוא תרכובת אורגנית ניאון כי הוא קטן יותר באופן משמעותי מאשר כתבים האנזים ולכן לא צפויות כמו להפריע לתפקוד ביולוגי. mCherry- Ca V α2δ1-HA ביתר TSA-201cells, מייצר גידול של 3-יומן משמעותי את קרינת FITC ו mCherry הקרין על מגרשים דו ממדים 22. בהתחשב בכך epitope HA ממוקם בחלק התאי של החלבון, את עוצמת הקרינה עבור FITC שהושגה בנוכחות של תאים שלמים לשקף את המדד היחסי של הביטוי על פני קרום התא של חלבון HA-tagged. הנגישות של epitope HA של המבנים הוא תוקף באופן שיטתי על ידי מדידת האות FITC לאחר התא permeabilization. מדד זה משמש גם כדי לאשש את ביטוי חלבון הכולל המנורמל מאז עוצמות קרינה היחסיות עבור FITC מוערכות בתאי permeabilized הם איכותיים להשוות את ערכי הקרינה היחסיים FOr mCherry נמדד בתנאי permeabilized והלא-permeabilized 22,23. חשוב לציין כי ספקטרום קרינה הפנימי מוסט לכיוון ערכים גבוהים יותר לאחר permeabilization אבל זה הערך היחיד שדווח הוא השינוי בעוצמת קרינה לעומת המבנה השליט. שינויים יחסיים בעוצמת הקרינה עבור המבנים הבדיקה מחושבים באמצעות עוצמת הקרינה ΔMean (ΔMFI) ערכים עבור כל fluorophore (mCherry או FITC). ניסויים נועדו למדוד את עוצמת הקרינה של יחסי מבחן לבנות את עוצמת הקרינה של מבנה השליטה לידי ביטוי באותם תנאים להגביל וריאציות ניסיוני הקרינה הפנימית של הנוגדן מצומדות fluorophore. שני חלבונים בממברנה נחקרו בהצלחה באמצעות assay: למקטע יוצרים הנקבובי של תעלות סידן L- סוג המתח מגודרת Ca V 1.2 14,22 ו בסדרה שונה שלניסויים, למקטע תאיים עזר Ca V α2δ1 22,23. הפרוטוקול הבא שמש כדי לקבוע את הביטוי על פני קרום התא של למקטע Ca V α2δ1 של L-סוג הלב Ca V 1.2 ערוץ בתנאים מלאים לאחר מוטציות המשפיעות על שינוי posttranslational של ערוץ היון. תחת תנאי ניסוי סטנדרטיים, הקרינה פני התא של FITC מגבירה מעין-ליניארי עם הביטוי של cDNA מקודדי חלבוני mCherry-Ca V α2δ1-HA (האיור 5 בהפניה 22).

איור 2:. ייצוג סכמטי של תיוג סך קרום בזרימת cytometry פרוטוקול הניסוי התוכנית מתאר כמה מן השלבים העיקריים הדרושים כדי לכמת את הביטוי הכולל ותא השטח היחסי של תעלות יונים רקומביננטי ידי flow cytometry. תאים הם transfected עם בניית פעמים מתויגות mCherry-Ca V α2δ1-HA בתאי TSA-201 (1) ו מוכתמים לפני או אחרי permeabilization (2). נתוני פרמטרים מרובים נרכשים cytometer זרימה (3) עבור ניתוח רב משתנה (4). נא ללחוץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
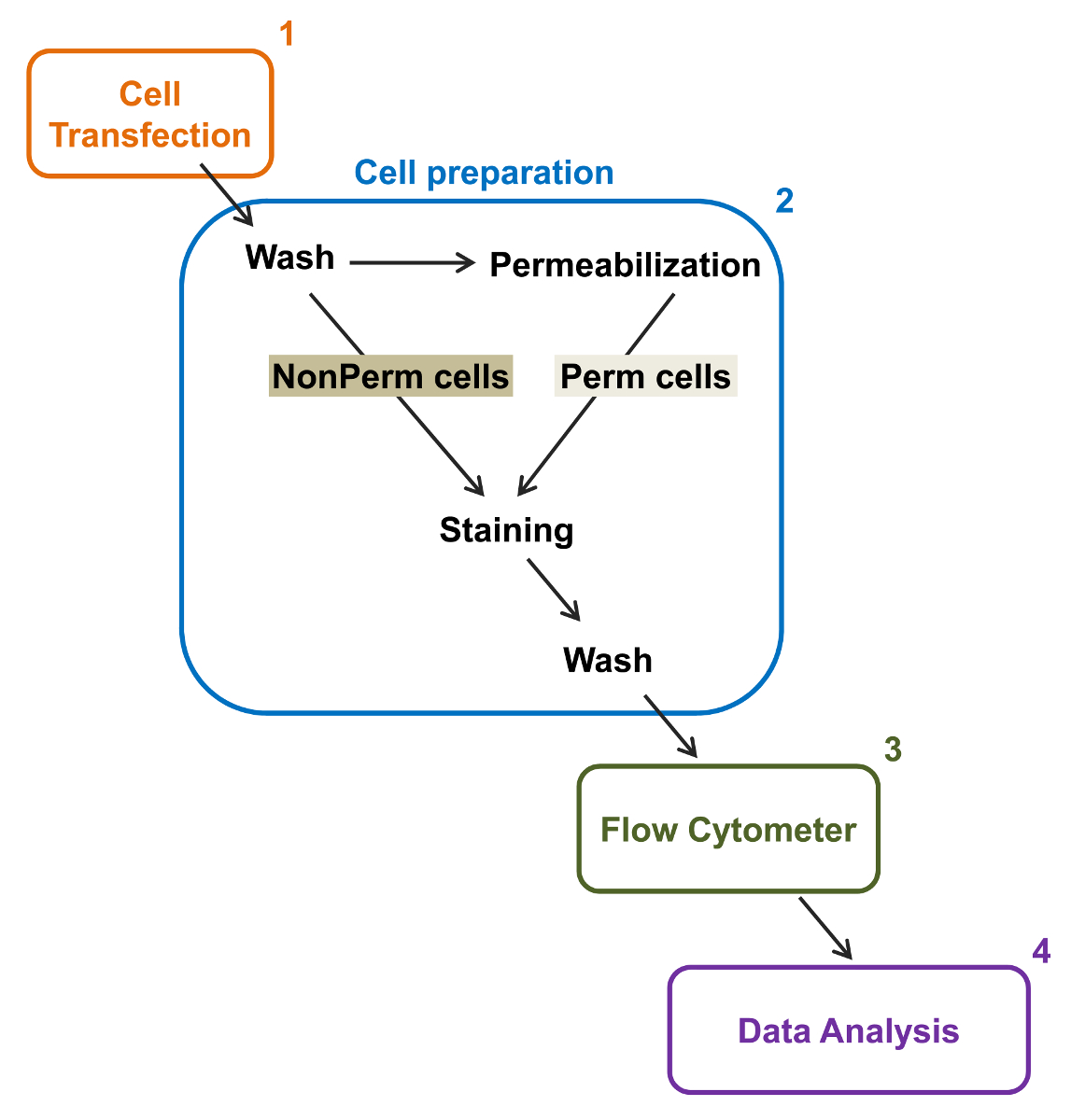{kind=link}
Protocol
1. בונה DNA Tagged כפלים
- הכנס את epitope HA (YPYDVPDYA) ב והמקשר התאי של α2δ1 Ca V בין D676 ו- R677 על ידי mutagenesis באתר ביים (איור 3 ב) 20. השתמש קדימה פריימר gccggattatgcgGGAAAACTCCAAACAACC הפוך acatcatacggataTCAATAAATTCATTGAAATTTAAAAGAAATTC פריימר.
- Subclone רצף cDNA של α2δ1 V HA Ca מתויג לתוך וקטור ביטוי pmCherry-N1 היונק נועד לבטא את החלבון התמזג N- הסופית של mCherry בין אתרי SACI ואת סאלי (איור 3B) 20.
הערה: תפקוד ערוץ מתאים צריכה להיבדק עם המבנה השליט באמצעות שיטות אלקטרו תקן 24.
2. Transfection חלוף בתיווך Liposome (30 דק ', כל הצעדים מבוצעים מתחת למכסת מנוע לזרום למינרית)
- יום 1: פלייט חצי מיליון תאים TSA-201 (או HEKT) ב35 מ"מ תרבות מנות עם 2 המדיום החיוני המינימום הגבוה גלוקוז של המ"ל של Dulbecco (DMEM-HG) בתוספת 10% עובריים שור סרום (FBS) ו -1% פניצילין, סטרפטומיצין (PS) בינוני תרבות. ספירת תאים באמצעות hemacytometer תקן. הערכת כדאיות התא מן שבריר של המדגם התא באמצעות Trypan כחול. פלייט תאים מספיק כדי להגיע למפגש 90% בזמן transfection.
- יום 2: שינוי התרבות בינוני עם 2 מ"ל של טרום התחמם טריים (37 ° C) בינוני תרבות ללא PS.
- עבור כל דגימה transfection, להכין שני צינורות 1.5 מ"ל. בשנת צינור 1, לדלל 4 מיקרוגרם של ה- DNA ב 250 μl של מדיום סרום מופחת. בשנת צינור 2, לערבב 10 μl של מגיב transfection בתיווך ליפוזום עם 250 בינוני תרבות μl-מופחת בסרום. מערבבים בעדינות מגיב transfection לפני השימוש.
- דגירה במשך 5 דקות בטמפרטורת החדר.
- מערבבים את תכולת שפופרת 1 צינור 2, מערבבים בעדינות דגירה לפחות 20 דקות בטמפרטורת החדר.
- מוסיפים את ליפוזומים / DNקומפלקסים אל התאים בתרבית בעדינות לטלטל את מנת התרבות לערבב.
- לדגור על 37 מעלות צלזיוס מתחת 5% CO 2 אווירה למשך 24 שעות.
3. צביעת תאים עבור cytometry זרימה (3 שעות)
- הכנת דגימות תאים
- יום 3: הסר בינוני מצלחת התרבות בזהירות לשטוף את התאים עם 400 μl של טרום התחמם (37 ° C) 0.05% טריפסין-1x EDTA (חומצה ethylenediaminetetraacetic).
- להוסיף 400 μl של טריפסין-EDTA דגירת הצלחת ב 37 מעלות צלזיוס מתחת 5% CO 2 אווירה במשך 5 דקות כדי לאפשר לתאים להתנתק המנה.
- עצור את העיכול אנזים על ידי הוספת 1 מ"ל של מדיום תרבות קר ללא PS ולשטוף את כל התאים מפני השטח על ידי pipetting בעדינות 4-5 פעמים. הימנע יתר העיכול יתר pipetting להפחית מוות של תאים.
- איסוף תאים ב 1.5 מ"ל צינורות ומניחים מיד על הקרח. השתמש בפתרונות קרים קרח ולשמור את התאים ב 4 ° C כדי למנוע ההפנמהשל פני השטח אנטיגנים. ירידת תאורה להגביל photobleaching של אות הניאון.
- צינורות צנטריפוגה ב 400 XG במשך 5 דקות ב 4 ° C. בזהירות לשאוב וזורקים supernatant.
- Re- להשעות גלולה ב 1 מ"ל של 1x שנאגרו מלוחים פוספט (PBS) כדי להכין ההשעיה תא בודד.
- בקצרה מערבולת הצינורות מאוד בעדינות וחזור על שלבים 3.1.5 ו 3.1.6 להסיר בינוני תרבות לחלוטין.
- Re- להשעות גלולה ב 600 μl של 1x PBS ולהתאים את ריכוז התאים עד למינימום של 3 x 10 6 תאים / מ"ל.
- מחלק את התאים בשני 1.5 מיליליטר צינורות חדשים עבור מכתים תאי ו תאי. כלול בקרות מתאימות להיפלות מכתים ספציפי מכתים הלא ספציפי.
הערה: נוגדן אלוטיפ עוזר בהערכת רמת מכתים רקע אידיאלי צריך להתאים לכל מין המארח של נוגדן ראשוני, אלוטיפ ו fluorophore. השתמש בפקד אלוטיפ נוגדנים מצומדות באותוריכוז החלבון.

טבלת 1: דגימות בקרת ניסוי-cytometry זרימה בלתי permeabilized ו permeabilized תאים כל ניסוי צריך לכלול את השולטת השלילית הבאה:. (1) תאי Nontransfected (ללא נוגדנים, עם אלוטיפ או עם נוגדן מצומדות). (2) תאי transfected עם החלבון של עניין subcloned פלסמיד ללא fluorochrome תאי פלורסנט מכונן (ע"א pCMV- α2δ1-HA) או עם הבנייה מתויגת כפליים (pmCherry-Ca V α2δ1 וטופח ללא נוגדנים, עם אלוטיפ או עם נוגדן מצומדות). צבע שולט יחיד משמש פיצוי של חפיפת פליטת fluorochrome. באותם אמצעי השליטה מנוהלת עבור הלא-permeabilized ו permeabilized תנאים בכל סדרה של ניסויים.
- מכתים פני התא של Intactבתאים חיים
- Aliquot 1 x 10 6 תאים / 100 μl ב 1.5 מ"ל צינורות.
- מוסיף את הנוגדן אנטי HA monoclonal FITC- מצומדות ב 5 מיקרוגרם / מיליליטר ו, מערבולת לפני דוגרי התאים על פלטפורמת נדנדה (200 סל"ד) בחושך ב 4 מעלות צלזיוס במשך 45 דקות.
הערה: הריכוז האופטימלי של נוגדנים נקבע בניסויי טיטרציה ראשוניים (איור 4). - הסרת תאים מן החשכה להוסיף 900 μl של / צינור 1x PBS. צנטריפוגה ב 400 XG במשך 5 דקות ב 4 ° C..
- לשאוב supernatant ו resuspend גלולה ב 1 מ"ל של PBS 1x, מערבולת ו צנטריפוגות ב 400 XG במשך 5 דקות ב 4 ° C..
- חזור על לשטוף (שלב 3.2.4) פעמים כדי להסיר כל נוגדנים מאוגדים. אם נוגדן ראשוני unconjugated משמש, דגירה עם נוגדנים משני המתאים.
- לאחר שטיפה של דבר, resuspend התאים 500 μl של PBS 1x ולהעביר את ההשעיה תא בודד ב 5 מ"ל cytometry זרימה צינורות. שמור התאים בחושך ב 4 ° C. עד פועל המדגם.
- הפעל את הדגימות על cytometer זרימה. לקבלת התוצאות הטובות ביותר, לנתח את התאים על זרימת cytometer בהקדם האפשרי ולא יאוחר מ -24 שעות לאחר.
- מכתים תאי: קיבוע, Permeabilization, מכתים
- Aliquot 1 x 10 6 תאים / 100 μl ב 1.5 מ"ל צינורות צנטריפוגות ב 400 XG במשך 5 דקות ב 4 ° C..
- מחק תאים supernatant ו resuspend ב 100 μl של פתרון קיבעון-permeabilization ישירות ממלאי.
- דגירה בחושך ב 4 מעלות צלזיוס במשך 20 דקות.
- הוספת 100 μl של חיץ permeabilization לשטיפת 1x מוכן טרי (לדלל 10x חיץ permeabilization לשטיפת ב O H 2 מזוקקים). תאים וורטקס ומשקע באמצעות צנטריפוגות שולחן ב 400 XG במשך 5 דקות ב 4 ° C.
- לשאוב וזורקים supernatant.
- חזור על שלבים 3.3.4 ו 3.3.5.
- להוסיף monoclonal FITC- מצומדות נוגדן אנטי HA ב 5מיקרוגרם / מ"ל ב 100 μl של חיץ permeabilization לשטיפת 1x ו, מערבולת לפני דוגרים תאים בחושך על 4 מעלות צלזיוס למשך 30 דקות.
הערה: המכתים התאי מתבצע בעקבות ההליך אותו לזו שבוצעה עבור שטח פני תא מכתים. סאפונין בתיווך התא permeabilization עם זאת, תהליך הפיך במהירות, ולכן חשוב להחליף PBS 1x עם חיץ 1x פרם / Wash לשמור על התאים בנוכחות מתמדת של saponin במהלך מכתים תאיים. - הסרת תאים מן החשכה להוסיף 100 μl חיץ לשטיפת permeabilization. צנטריפוגה ב 400 XG במשך 5 דקות ב 4 ° C..
- לשאוב בזהירות את supernatant ו resuspend גלולה ב 100 μl חיץ לשטיפת permeabilization, מערבולת ו צנטריפוגות ב 400 XG במשך 5 דקות ב 4 ° C..
- חזור על לשטוף (שלב 3.3.9) עוד פעם אחת כדי להסיר כל נוגדן מאוגד.
- לאחר שטיפה של דבר, resuspend התאים 500 μl של PBS 1x ולהעביר את החטאהשעית תא GLE עד 5 זרימת מיליליטר cytometry צינורות. שמור את התאים בחושך ב 4 מעלות צלזיוס עד הזרקת המדגם לתוך זרם cytometer.
- הפעל את הדגימות על cytometer זרימה. הפעל את הדגימות הקבועות על cytometer בהקדם האפשרי אך לא יאוחר מיום 1 שבוע לאחר צביעה. הפעל את התאים הלא-permeabilized ו permeabilized באותו יום.
4. Cytometry זרימה
- Cytometer זרימת הגדרת תא הסדרן יומי
- הפעל את תוכנת cytometry הזרימה. לפני להתנסות, לכייל ולהתקין את הזרימה cytometer סדרן תא כדי להבטיח ביצועים אופטימליים מכשיר (ליזר כלומר ואופטיקה מתפקדת למפרט, הליזר וזרימת תא מיושר כראוי) באמצעות חרוזי התקנה מכשירה.
- השתמש זרבובית 100 מיקרומטר עם לחץ נדן 20 psi.
הערה: הזרבובית אינה צריכה להיות שונה על ספסל לזרום cytometer. - הגדר את קצב הזרימה של cytometer פי manufacturמפרט אה. מאוד גבוהי זרימת שיעורים יקטנו רגישות זיהוי של וריאציות קרינה.
- בחר כחול (488 ננומטר לרגש והעמסת Isothiocayanate או FITC) ו-צהוב ירוק (561 ננומטר לרגש mCherry) לייזרים. אסוף רמות הקרינה FITC ו mCherry עם ננומטר 530/30 ועם מסנן bandpass 610/20 ננומטר בהתאמה.
- רוכש את הפיזור קדימה (FCS) לעומת פיזור בצד עלילת נקודה (SSC) עבור תאים בלא כתם באמצעות בקנה המידה ליניארי. התאם כל הגברה של גלאי לדמיין התאים ברבע השמאלי התחתון של העלילה נקודה.
- קריאת מדגם שלם ללא permeabilized תאים
- הגדר את P1gate עבור תאים שאינם permeabilized חיו על ידי התוויית צורה חופשית סביב התאים להיות מנותחים למעט פסולת תא אגרגטים תא, ובכך להגביל את אות הקרינה בתאים שלמים.
הערה: Live / צבעי הרחקה מתות ניתן להשתמש כדי להקל על מיקום שער על תאי חיים. גדר 10,000 אירועים להקליטב P1 שער עצירה. סמן את זה למספר גבוה יותר של אירועים אם יהיה בכך צורך. - רוכשת mCherry לעומת העלילה מתאר שתי פרמטר FITC לזהות autofluorescence הבסיס של תאים בלא כתם. השתמש בקנה מידה דו-לוגריתמי להראות ערכים שליליים ולשפר ברזולוציה בין אוכלוסיות 25. התאם כל של מתח גלאי להגדיר את התאים השליליים בלא הכתם בתוך החלק התחתון של עשר היחידות הראשונות של החלקות עוצמות קרינת יומן.
- לרכוש את כל דגימות השלמים שאינם permeabilized באמצעות הגדרות הוקמו בשנת 4.1.5 4.1.6 ולאסוף FSC, SSC ו אותות גלאי הקרינה.
- יצוא ולשמור * קבצי .fcs לשמש לניתוח באמצעות זרימת cytometry תוכנה לניתוח.
- הגדר את P1gate עבור תאים שאינם permeabilized חיו על ידי התוויית צורה חופשית סביב התאים להיות מנותחים למעט פסולת תא אגרגטים תא, ובכך להגביל את אות הקרינה בתאים שלמים.
- קריאת דגימת תאי permeabilized
- הזז את השער P1 כדי לבחור תאים חיים בדגימות permeabilized ולהתאים FSC ומתח SSC כמוצג 4.1.5 4.1.6.
- לרכוש את כל דגימות permeabilized ולאסוף FSC, SSCאותות nd ב גלאי הקרינה.
- יצוא ולשמור * קבצי .fcs לשמש לניתוח באמצעות זרימת cytometry תוכנה לניתוח.
- ניתוח נתונים
- הפעל את זרימת cytometry קבצים .fcs * ייבוא תוכנה וניתוח שנשמרו 4.2.4 ו 4.3.3.
- הקש על המדגם הראשון רשום בחלון סביבת העבודה. חלון חדש בשם אחרי מספר מזהה צינור נפתח אוטומטית. התחילו בתהליך gating בעלילה של SSC לעומת FSC. צייר שער (P1) באמצעות סמל האליפסה סביב תאי חיים ולחסל את כל פסולת, תאים מתים, או אגרגטים אשר יש פיזור קדימה שונה ולפזר לוואי מאשר תאי חיים
- כדי למשוך את העלילה מתאר שתי פרמטר של mCherry (ציר y) לעומת FITC (ציר x) עוצמת הקרינה של תאים חיים, ללחוץ תחילה על ציר ה- x ולבחור את FITC-A הערוץ ולאחר מכן לחץ על y ציר ולבחור את ערוץ PE-mCherry-A. לחץ על הסמל "Quad" כדי למקם את הסמן ברבע בקצה שלתאי utofluorescent בכל ערוץ קרינה.
הערה: השער להגדיר סביב התאים החיוביים FITC ו mCherry הוא שער P2. אוכלוסיית התאים השלילית הקרינה המכונית שער P3. ראה איור 5 עבור שיטת gating נציג שימוש במאמר זה. - P2 ובחר שערים P3 ולחץ על הסמל "הוספת סטטיסטיקות" בחלון סביבת העבודה המקורית. לחץ על "רוזן" (מספר תאים החיוביים) ולחץ על "Mean" (Mean קרינת עצמה של כל fluorochrome) או "חציון" (עוצמת קרינת החציון של כל fluorochrome) סטטיסטיקה בין רשימת האפשרויות. הקש על הסמל "הוסף הסטטיסטיקה" שוב. כל הערכים הללו מועברים אוטומטית בחלון סביבת העבודה המקורי.
הערה: "Mean" משמשת רק אם עוצמת הקרינה כדלקמן התפלגות נורמלית. בכל מקרה אחר, לחץ על הכרטיסייה "חציון". MFI ובכך יכול להתייחס Mean הקרינה Intensitעוצמת הקרינה y או חציון.
הערה: השלב הבא הוא ליישם את הפרמטרים של גייטס וסטטיסטיקה לכל הדגימות נחקרו על ידי cytometer. - בחלון סביבת העבודה, להשתמש בעכבר כדי לגרור ולשחרר את פרמטרי שערים וסטטיסטיקות על הקו מסומן כל דגימות.
- להפיק דוח אצווה של מגרשים מתאר דו מימדי (mCherry vs FITC) היסטוגרמות (תא לספור לעומת עוצמת הקרינה) עבור תאים שאינם permeabilized ו permeabilized (6A ומספרים - B).
- מהסטטיסטיקה שנוצרה בשלב 4.4.4, לחשב את עוצמת הקרינה הממוצעת (MFI) עבור כל fluorochrome עבור התאים המוכתמים. מתוך ערך זה, להפחית את ערך MFI המתקבל תאים בלא כתם לכמת את השטח וביטוי הכולל של החלבון של עניין.
- דווח על ערכי ΔMFI לכל fluorophore (mCherry ו FITC) (איור 6 ג - ד). לנרמל את ΔMFI נמדד על Ca V α2δ1 לבנות מוטציות לערך ΔMFI המתקבל עבור FITC ו mCherry עם מבנה WT.
הערה: הערך המוחלט של עוצמת קרינה יכולה להשתנות בחדות בהתאם תצווה של נוגדנים ואת היכולות הטכניות של כל עובד במעבדה, ומכאן הצורך לנרמל עוצמת קרינה של לבנות מוטנטים באמצעות מבנה WT.
תוצאות
מאמר זה מתאר פרוטוקול אמין לכמת פנים כולל ותא של תעלות יונים רקומביננטי לידי ביטוי TSA-201cells ידי זרימת שני צבעים cytometry assay. כדוגמה, פני התא ביחס וביטוי החלבון הכולל היה לכמת עבור α2δ1subunit ע"א. על מנת לבצע את זרימת שני צבעי cytometry assay, Ca V α2δ1 תוי...
Discussion
Assay cytometry מבוסס זרימה זו יושם בהצלחה על מדידת רמות פן כולל ותא היחסיים של fluorescently שכותרתו יוצרי הנקבוביות הקשורים יחידות המשנה של ערוצי סידן מגודר מתח 14,22,26. עדיף להשתמש כאשר חוקרים את ההשפעה של מוטציות גנטיות וכך מחייב כי עוצמת הקרינה הפנימית של המבנה fluorescently שכ?...
Disclosures
The authors declare that they have no competing financial interests.
Acknowledgements
We thank Mr. Serge Sénéchal and Dr. Jacques Thibodeau for sharing their expertise and granting us access to their flow cytometry and cell sorting platform. This work was completed with the operating grant 130256 from the Canadian Institutes of Health Research, a grant-in-aid from the Canadian Heart and Stroke Foundation, and support from the "Fondation de l'Institut de Cardiologie de Montréal" to L.P.
Materials
| Name | Company | Catalog Number | Comments |
| Q5 Site-Directed Mutagenesis Kit | New England Biolabs | E0554S | Can be substitute with QuickChange site-directed mutagenesis Kit (Agilent, #200523). |
| Tubes 1.5 ml | Sarstedt | 72-690-001 | |
| Tubes 15 ml | Sarstedt | 62-554-002 | |
| Disposable graduated Tranfer Pipets | VWR | 160001-192 | |
| 100 mm culture dish | Corning | 430167 | For standard culture of HEKT cells. |
| 35 mm culture dish | Falcon | 353001 | For standard culture of HEKT cells. |
| Serological pipette 1 ml | Sarstedt | 86.1251.001 | |
| Serological pipette 5 ml | Sarstedt | 86.1253.001 | |
| Serological pipette 10 ml | Sarstedt | 86.1254.001 | |
| Serological pipette 25 ml | Sarstedt | 86.1285.001 | |
| Dulbecco's high-glucose medium | Life Technologies | 12100-046 | Warm in 37 °C water bath before use. |
| Fetal Bovine Serum, qualified, heat inactivated, US origin | Life Technologies | 16140-071 | |
| Penicillin-Streptomycin (10,000 U/ml) | Life Technologies | 15140-122 | |
| Lipofectamine 2000 | Life Technologies | 11668-019 | For liposomal transfection. Can be substituted with calcium phosphate transfection. |
| Opti-MEM I Reduced Serum Medium | Life Technologies | 31985-070 | Warm in 37 °C water bath before use. |
| Trypsin-EDTA (1x) 0.05%, phenol red | Life Technologies | 25300-062 | |
| 1.5 ml microtubes | Sarstedt | 72.690.001 | |
| Phosphate Buffered Saline 1x | Fisher | BP661-10 | Can be "home-made". |
| Anti-HA FITC conjugated antibody | Sigma | H7411 | |
| IgG1−FITC Isotype Control antibody | Sigma | F6397 | |
| BD Cytofix/Cytoperm Fixation/Permeabilization Solution Kit | BD Biosciences | 554714 | Fixation/Permeabilization. Permeabilization/Wash solution, store at 4 °C. |
| Hemacytometer | Fisher | 49105161 | |
| Trypan Blue | Fisher | 15250061 | To access cell viability. |
| Refrigerated Microcentrifuge, 5430R | Eppendorf | A14H172200 | |
| Forma Steri-Cycle CO2 Incubator | Fisher | 370 | |
| Laboratory Platform Rocker | Fisher | 545034 | |
| Water Bath | VWR | 89032-216 | |
| BD FACSARIA III | BD Biosciences | 648282 | Flow cytometer. |
| FlowJo Software v10 | FlowJo | FlowJo v10 Dongle | For data analysis. |
References
- Delisle, B. P., Anson, B. D., Rajamani, S., January, C. T. Biology of Cardiac Arrhythmias: Ion Channel Protein Trafficking. Circ. Res. 94, 1418-1428 (2004).
- Birault, V., Solari, R., Hanrahan, J., Thomas, D. Y. Correctors of the basic trafficking defect of the mutant F508del-CFTR that causes cystic fibrosis. Curr Opin Chem Biol. 17, 353-360 (2013).
- Balijepalli, S. Y., Anderson, C. L., Lin, E. C., January, C. T. Rescue of Mutated Cardiac Ion Channels in Inherited Arrhythmia Syndromes. J. Cardiovas Pharm. 56, 113-122 (2010).
- Gargus, J. J. Unraveling Monogenic Channelopathies and Their Implications for Complex Polygenic Disease. Am. J. Hum. Genet. 72, 785-803 (2003).
- Abriel, H., Zaklyazminskaya, E. V. Cardiac channelopathies: Genetic and molecular mechanisms. Gene. 517, 1-11 (2013).
- Behr, E. R., et al. Sudden arrhythmic death syndrome: familial evaluation identifies inheritable heart disease in the majority of families. Eur Heart J. 29, 1670-1680 (2008).
- Catterall, W. A. Structure and regulation of voltage-gated Ca2+ channels. Annu. Rev. Cell Dev.Biol. 16, 521-555 (2000).
- Peterson, B. Z., DeMaria, C. D., Adelman, J. P., Yue, D. T. Calmodulin is the Ca2+ sensor for Ca2+ -dependent inactivation of L- type calcium channels. Neuron. 22, 549-558 (1999).
- Dolphin, A. C. Calcium channel diversity: multiple roles of calcium channel subunits. Curr.Opin.Neurobiol. 19, 237-244 (2009).
- Dai, S., Hall, D. D., Hell, J. W. Supramolecular assemblies and localized regulation of voltage-gated ion channels. Physiol Rev. 89, 411-452 (2009).
- Gao, T., et al. Identification and subcellular localization of the subunits of L-type calcium channels and adenylyl cyclase in cardiac myocytes. J. Biol. Chem. 272, 19401-19407 (1997).
- Carl, S. L., et al. Immunolocalization of sarcolemmal dihydropyridine receptor and sarcoplasmic reticular triadin and ryanodine receptor in rabbit ventricle and atrium. J. Cell Biol. 129, 673-682 (1995).
- Abriel, H., Rougier, J. S., Jalife, J. Ion Channel Macromolecular Complexes in Cardiomyocytes: Roles in Sudden Cardiac Death. Circ. Res. 116, 1971-1988 (2015).
- Bourdin, B., et al. Molecular Determinants of the Cavb-induced Plasma Membrane Targeting of the Cav1.2 Channel. J. Biol. Chem. 285, 22853-22863 (2010).
- Raybaud, A., et al. The Role of the GX9GX3G Motif in the Gating of High Voltage-activated Calcium Channels. J. Biol. Chem. 281, 39424-39436 (2006).
- Burashnikov, E., et al. Mutations in the cardiac L-type calcium channel associated with inherited J-wave syndromes and sudden cardiac death. Heart Rhythm. 7, 1872-1882 (2010).
- Hennessey, J. A., et al. A CACNA1C Variant Associated with Reduced Voltage-Dependent Inactivation, Increased Cav1.2 Channel Window Current, and Arrhythmogenesis. PLoS ONE. 9, e106982 (2014).
- Adan, A., Alizada, G., Kiraz, Y., Baran, Y., Nalbant, A. Flow cytometry: basic principles and applications. Crit Rev Biotechnol. , 1-14 (2016).
- Graham, M. D. The Coulter Principle: Foundation of an Industry. J. Lab. Autom. 8, 72-81 (2003).
- Baumgarth, N., Roederer, M. A practical approach to multicolor flow cytometry for immunophenotyping. J. Immunol. Methods. 243, 77-97 (2000).
- Rothe, G., Sack, U., Tarnok, A., Rothe, G. . Cellular Diagnostics. Basics, Methods and Clinical Applications of Flow Cytometry. , 53-88 (2009).
- Bourdin, B., et al. Functional Characterization of Cavalpha2delta Mutations Associated with Sudden Cardiac Death. J. Biol. Chem. 290, 2854-2869 (2015).
- Tetreault, M. P., et al. Identification of glycosylation sites essential for surface expression of the Cavalpha2delta1 subunit and modulation of the cardiac Cav1.2 channel activity. J. Biol. Chem. 291, 4826-4843 (2016).
- Senatore, A., Boone, A. N., Spafford, J. D. Optimized Transfection Strategy for Expression and Electrophysiological Recording of Recombinant Voltage-Gated Ion Channels in HEK-293T Cells. J Vis Exp. (47), (2011).
- Herzenberg, L. A., Tung, J., Moore, W. A., Herzenberg, L. A., Parks, D. R. Interpreting flow cytometry data: a guide for the perplexed. Nat.Immunol. 7, 681-685 (2006).
- Shakeri, B., Bourdin, B., Demers-Giroux, P. O., Sauve, R., Parent, L. A quartet of Leucine residues in the Guanylate Kinase domain of Cavbeta determines the plasma membrane density of the Cav2.3 channel. J Biol Chem. 287, 32835-32847 (2012).
- Morton, R. A., Baptista-Hon, D. T., Hales, T. G., Lovinger, D. M. Agonist- and antagonist-induced up-regulation of surface 5-HT3A receptors. Br. J. Pharmacol. 172, 4066-4077 (2015).
- Hoffmann, C., et al. Fluorescent labeling of tetracysteine-tagged proteins in intact cells. Nat. Protocols. 5, 1666-1677 (2010).
- Cockcroft, C. J., Gamper, N. . Ion Channels: Methods and Protocols. , 233-241 (2013).
- Gonzalez-Gutierrez, G., Miranda-Laferte, E., Neely, A., Hidalgo, P. The Src Homology 3 Domain of the beta-Subunit of Voltage-gated Calcium Channels Promotes Endocytosis via Dynamin Interaction. J. Biol.Chem. 282, 2156-2162 (2007).
- Galizzi, J. P., Borsotto, M., Barhanin, J., Fosset, M., Lazdunski, M. Characterization and photoaffinity labeling of receptor sites for the Calcium channel inhibitors d-cis-diltiazem, (+/-)-bepridil, desmethoxyverapamil, and (+)-PN 200-110 in skeletal muscle transverse tubule membranes. J. Biol.Chem. 261, 1393-1397 (1986).
- Bezanilla, F. The voltage sensor in voltage-dependent ion channels. Physiol.Rev. 80, 555-592 (2000).
- Sigworth, F. J. The variance of sodium current fluctuations at the node of Ranvier. J Physiol. 307, 97-129 (1980).
- Bailey, M. A., Grabe, M., Devor, D. C. Characterization of the PCMBS-dependent modification of KCa3.1 channel gating. J. Gen. Physiol. 136, 367-387 (2010).
- Fletcher, P. A., Scriven, D. R., Schulson, M. N., Moore, E. D. Multi-Image Colocalization and Its Statistical Significance. Biophys. J. 99, 1996-2005 (2010).
- Lizotte, E., Tremblay, A., Allen, B. G., Fiset, C. Isolation and characterization of subcellular protein fractions from mouse heart. Anal. Biochem. 345, 47-54 (2005).
- Mattheyses, A. L., Simon, S. M., Rappoport, J. Z. Imaging with total internal reflection fluorescence microscopy for the cell biologist. J. Cell Sci. 123, 3621-3628 (2010).
- Yamamura, H., Suzuki, Y., Imaizumi, Y. New light on ion channel imaging by total internal reflection fluorescence (TIRF) microscopy. J. Pharmacol. Sci. 128, 1-7 (2015).
- Wible, B. A., et al. HERG-Lite-R: A novel comprehensive high-throughput screen for drug-induced hERG risk. J. Pharmacol. Toxicol. Methods. 52, 136-145 (2005).
- Wilde, A. A. M., Brugada, R. Phenotypical Manifestations of Mutations in the Genes Encoding Subunits of the Cardiac Sodium Channel. Circ. Res. 108, 884-887 (2011).
- Milano, A., et al. Sudden Cardiac Arrest and Rare Genetic Variants in the Community. Circ Cardiovasc Genet. , (2016).
- Schnell, U., Dijk, F., Sjollema, K. A., Giepmans, B. N. G. Immunolabeling artifacts and the need for live-cell imaging. Nat. Meth. 9, 152-158 (2012).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved