
Method Article
יזם לכידת שלום-c: ברזולוציה גבוהה, הגנום כולו פרופיל של אינטראקציות יזם
* These authors contributed equally
In This Article
Summary
רכיבים רגולטוריות דנ א, כגון מוצרי טיפוח טבעיים, לשלוט ביטוי גנים פיזית שעת היעד ג'ין היזמים, לרוב באמצעות אינטראקציות כרומוזומלית ארוכי טווח הנמשכים מרחקים גדולים גנומית. יזם ללכוד Hi-C (PCHi-C) מזהה אינטראקציות משמעותיות בין היזמים ואזורים הדיסטלי, המאפשר את ההקצאה של פוטנציאל רגולטוריות רצפי הגנים היעד שלהם.
Abstract
הארגון תלת מימדי של הגנום הוא מקושר אל תפקידה. לדוגמה, רכיבים רגולטוריות כגון משפרי תעתיק לשלוט התבטאות הגנים היעד שלהם באמצעות מגע פיזי, לעיתים קרובות גישור מרחקים גנומית ניכרת (במקרים מסוימים מאות של kilobases), תוך עקיפת-עתיים גנים הסמוכה. הגנום האנושי בנמלים משפרי מיליון מוערך, הרוב המכריע של אשר יש נודע מטרות הגן. הקצאת אזורים רגולטוריות דיסטלי הגנים שלהם יעד חיוני ובכך להבין בקרת ביטוי גנים. פיתחנו יזם ללכוד Hi-C (PCHi-C) כדי לאפשר זיהוי הגנום כולו אזורים אינטראקציה-יזם הדיסטלי (חיישנים), עבור כל היזמים בניסוי יחיד. PCHi-c, ספריות ויטמינצ'יק מורכב במיוחד מועשר עבור יזם רצפים דרך הבחירה בפתרון היברידית עם אלפי פיתיונות biotinylated RNA משלים הקצוות של כל חלקי של הגבלת המכילים מקדם. המטרה היא ואז נפתחים יזם רצפים זוגם בתדירות גבוהה אינטראקציה כגון משפרי ורכיבים רגולטוריות פוטנציאליים אחרים. לאחר רצף לזווג-end תפוקה גבוהה, מבחן סטטיסטי מוחל על כל שבר ובין אם-לא יזם ההגבלה כדי לזהות חיישנים משמעותית ברמת פרגמנט ההגבלה. השתמשנו PCHi-C כדי ליצור אטלס של אינטראקציות יזם ארוכי טווח עשרות רבות של האדם, סוגי תאים העכבר. מפות interactome אלה יזם תרמו להבנה גדולה יותר של בקרת ביטוי גנים בתרבית של-ידי הקצאת בשם אזורים רגולטוריות הגנים שלהם היעד של חשיפת מועדף המרחבי יזם-יזם אינטראקציה עם רשתות. מידע זה כולל גם רלוונטיות גבוהה להבנת מחלות גנטיות אנושיות וזיהוי של גנים פוטנציאליים מחלה, על-ידי קישור ללא קידוד הקשורים למחלה רצף משתנים או בסמוך שליטה רצפי הגנים היעד שלהם.
Introduction
לצבירת ראיות מרמז כי הארגון תלת מימדי של הגנום ממלא תפקיד פונקציונלי חשוב במגוון תהליכים גרעיניים, כולל גנים הפעלה1,2,3, דיכוי4 ,5,6,7,8, רקומבינציה9,10, תיקון דנ א11,12,שכפול הדנ א13, ו הזדקנות ביולוגית הסלולר14. משפרי מרוחק נמצאים בסמיכות המרחבי היזמים שהם לווסת15,16,17, החיונית לבקרת ביטוי נאות גנים-עתיים. משפר מחיקות להראות כי משפרי דיסטלי הם חיוניים עבור היעד ג'ין שעתוק18,19,20,21,22, והכריח את ' כרומטין לולאה " מדגים כי קשירת מהונדסים בין שיפור של יזם היעד שלה במיקומה Hbb מספיקה לנהוג הפעלת גנים ברמת השעתוק23. עוד יותר, rearrangements הגנום המביאים גנים תחת השליטה של משפרי חוץ רחמי יכול לגרום הפעלת גנים לא הולם, מחלת24,25,26. יחד, דוגמאות אלה ממחישים כי יזם-enhancer אינטראקציות הם חיוניים עבור בקרת גנים, דורשים רגולציה הדוקה כדי להבטיח ביטוי גנים המתאימים. האדם ואת הגנום עכבר אחד מוערך למעגן משפרי מליון. עבור הרוב המכריע של אלה משפרי, היעד גנים אינם ידועים, ויש "חוקי המשחק" בין היזמים משפרי הם הבינו. הקצאת משפרי תעתיק את הגנים היעד שלהם ובכך נשאר אתגר גדול בפענוח בקרת ביטוי גנים יונקים.
ההבנה שלנו של אדריכלות הגנום תלת-ממדי יש כבר מהפכה על ידי המבוא של 3 ג27 (כרומוזום קונפורמציה לכידת) ו שלה משתנים28,29,30,31 . החזקים ביותר של טכניקות אלה, Hi-C (תפוקה גבוהה כרומוזום קונפורמציה לכידת) מיועד לזהות את ההרכב כולו של אינטראקציות כרומוזומלית בתוך אוכלוסיה התא. ספריות-C, בדרך כלל להפיק של מיליוני תאים, מורכבים מאוד במוצרים משוער 1011 מצדו עצמאית בין שברי ~ 4 kb ב הגנום האנושי32. כפי תוצאה, זיהוי לשחזור ואמין של אינטראקציות בין הגבלה בודדים שברים (כגון אלה המכילים של יזם או משפר) ויטמינצ'יק נתונים אינה אפשרית אלא אם ויטמינצ'יק ספריות ותוחלת רצף עמוק במיוחד, אשר אינה פתרון כלכלית עבור מעבדות הכנת ויטמינצ'יק ספריות באופן שגרתי. כדי לעקוף את זה חסרון, פיתחנו יזם ללכוד ויטמינצ'יק להעשיר במיוחד המכיל מקדם מכירות מוצרים מצדו מספריות ויטמינצ'יק. התמקדנו היזמים משתי סיבות. ראשית, יזם-שיפור הקשר הוכחו להיות קריטי עבור רמות ביטוי נאות גנים מחקרים רבים (ראו הפניות לעיל), שנית, כמו היזמים הם במידה רבה הקבועה בין סוגי תאים, באותה מערכת פיתיון לכידת יכול לשמש כדי לחקור המעגלים הרגולציה על פני מספר סוגי תאים והתנאים. הגישה שלנו מסתמך על הכלאה בבית-פתרון של ויטמינצ'יק ספריות עם עשרות אלפי 120mers biotinylated RNA משלים המכילים מקדם מוצרים מצדו של ויטמינצ'יק וללכוד עוקבות על מצופים streptavidin beads מגנטי. התוצאה PCHi-C ספריות עם הרבה מורכבות לעומת הספרייה ויטמינצ'יק המקורי, התמקדות רק הזיהוי של שברי כי מאתרים את היזמים בתדרים הגבוהים באופן משמעותי מופחתת.
השתמשנו PCHi-C מספר של האדם, סוגי תאים העכבר כדי לתרום הבנה טובה יותר של ג'ין בקרת ביטוי על ידי חשיפת יזם דיסטלי ארוכי טווח אזורים אינטראקציה עם פונקציית רגולציה בשם, כמו גם שאינן אקראיות אנשי קשר במרחב תלת-ממדי של הגרעין יזם-יזם. המחקרים מיפוי של מאות אלפי אנשי קשר יזם-משפר על פני רבים תא סוגי33,34,35,36,37,38, 39, זיהה ארגון הגנום המרחבי Polycomb דכאניים מורכבים בתיווך תאי גזע עובריים בעכבר7, הפגינו הטיפולי בקנה מידה גדול של יזם interactomes במהלך התמיינות37, 38 , 39, מקושר ללא קידוד הקשורים למחלה רצף גרסאות ל ג'ין היזמים35.
PCHi-C היא שיטה מתאים באופן אידיאלי כדי למפות את הגנום כולו אנסמבל של רצפי DNA אינטראקציה עם היזמים. גישות קשורים, כגון ללכוד ויטמינצ'יק האזורים גנומית רציפה (ראה דיון) הם השיטה של בחירה כדי להשיג פרופילים אינטראקציה ברזולוציה גבוהה עבור אזורים שנבחרו גנומית. PCHi-C ו- Hi-C ללכוד דומות מאוד ניסיוני נקודת מבט (ההבדל היחידי הוא הבחירה של מערכת לכידת), כך עצות והנחיות אנו מספקים ניתנים ליישום שתי הגישות. כאן, אנו מציגים תיאור מפורט של PCHi-סי אנחנו המתאר את הרציונל ואת העיצוב של ניסוי PCHi-C, מספקים פרוטוקול דור צעד אחר צעד של ספריית PCHi-C, להדגים כיצד ניתן לנטר את האיכות של ספריות PCHi-C-שלבים שונים בפרוטוקול להניב נתונים באיכות גבוהה.
Protocol
1. פורמלדהיד קיבוע
-
תא הכנה: להתחיל עם מינימום של 2 x 107 תאים לכל ניסוי.
- עבור תאים גדלו בתרבות, resuspend את התאים במדיום תרבות. עבור ex-vivo תאים, resuspend 1 x של Dulbecco ששינה נשר בינוני (DMEM), בתוספת 10% (vol/כרך) העובר סרום שור (FBS).
- עבור תאים חסיד, הסר בינוני תרבות ולהוסיף מל 30.625 בינוני טריים עם 10% (vol/כרך) FBS בטמפרטורת החדר (RT; 20 – 25 ° C).
- עבור תאים ההשעיה, לאסוף, צנטריפוגה תאים 400 x g ו- 20 ° C עבור מינימלית 3 הסרה תגובת שיקוע והשהה מחדש תא גלולה ב mL 30.625 בינוני עם 10% (vol/כרך) FBS-RT.
- בשביל לרקמות מוצקות, השתמש טריפסין (0.05% עד 2.5% הסופי ריכוז, בהתאם לסוג התא) או דאונס homogenizing כדי לקבל השעיה תא בודד. לאחר שלב נוסף זה, יטפל בתאים כמו תאים ההשעיה.
-
להוסיף 4.375 מ של 16% ללא מתנול paraformaldehyde (פתח המבחנה, רק לפני השימוש) ריכוז סופי של 2% (vol/כרך). תיקון עבור 10 דקות ב RT עם ערבוב עדין על כיסא נדנדה.
התראה: Paraformaldehyde הוא חומר כימי מסוכן. בצע את תקנות בטיחות וגהות המתאים. - להרוות את התגובה על-ידי הוספת 5 מ של 1 מ' טריות כקרח גליצין. לערבב במשך 5 דקות עם נדנדה עדין-RT ולאחר מכן דגירה על קרח למשך 15 דקות עם היפוך מפעם לפעם.
-
רוחצים את איסוף תאים קבוע.
- עבור תאים חסיד, להסיר את תגובת שיקוע, להוסיף 10 מ של קרח 1 x PBS pH 7.4 על הקיר צלחת ולהסיר אותו. להוסיף 1 מ"ל של קרח 1 x PBS pH 7.4, לאסוף תאים באמצעות תא במגרדת העברת לתוך צינור 50 מ. חזור על פעמיים כדי לאסוף תאים רבים ככל האפשר. להוסיף PBS קר כקרח לנפח סופי 50 מ.
- עבור השעיה תאים, התאים צנטריפוגה ב 760 x g ו- 4 מעלות צלזיוס למשך 5 דקות, להסיר את תגובת שיקוע, וכן מחדש להשעות תא גלולה ב 50 מ של PBS קר כקרח pH 7.4.
- צנטריפוגה תאים 400 x g ו- 4 מעלות צלזיוס למשך 10 דקות, הסר בזהירות את תגובת שיקוע. בגדר התא יכול להיות snap קפוא חנקן נוזלי, לאחר מכן מאוחסנים ב-80 מעלות צלזיוס למשך מספר חודשים.
2. פירוק התא
- מחדש להשעות צניפה תא ב 50 מ של מאגר טריות פירוק כקרח (10 מ מ טריס-HCl pH 8, 0.2% (vol/כרך) Igepal CA-630, 10 מ מ NaCl, ולא מעכב פרוטאז טבליה אחת קוקטייל) ומערבבים. דגירה על קרח למשך 30 דקות, מערבבים מדי פעם על-ידי היפוך. Centrifuge הגרעינים-760- g ו- 4 מעלות צלזיוס למשך 5 דקות ולהסיר תגובת שיקוע.
3. היןתרטר עיכול
- לשטוף את גרעין התא באמצעות מאגר הגבלת 1.25 x 2. מחדש להשעות תא גלולה ב 1 מ"ל מאגר הגבלת כקרח 1.25 x 2 והעברת לתוך צינור 1.5 מ. ספין הגרעינים-760-g ו- 4 מעלות צלזיוס למשך 5 דקות ולהסיר תגובת שיקוע.
- מחדש להשעות תא גלולה ב- 1790 µL מאגר הגבלת 1.25 x 2. להפוך 5 aliquots, שכל אחד מהם מכיל 5 – 10 מיליון תאים µL 358 של 1.25 x הגבלת מאגר 2.
- להוסיף 11 µL של 10% (wt/כרך) למען חברה דמוקרטית לכל aliquot ו- shake מהפכות 950 לכל מין (סל ד) למשך 30 דקות ב 37 מעלות צלזיוס ב thermomixer. אם מופיע תא גושים, מביצועם מאת pipetting, הימנעות בועות.
- להוסיף µL 75 של 10% X-100 טריטון (vol/כרך) לכל aliquot ו- shake-950 סל ד ו- 37 מעלות צלזיוס למשך 15 דקות thermomixer. אם מופיע תא גושים, מביצועם מאת pipetting, הימנעות בועות.
-
הוסף µL 12 של U/µL הין100 תרטר 100 (1,200 יחידות סה כ) לכל aliquot, דגירה ב 37 מעלות צלזיוס למשך הלילה (O/N) תוך טלטול-950 סל ד ב thermomixer.
- עבור הפקד עיכול, להעביר µL 25 מדגם (5 µL כל aliquot) צינור חדש לפני הוספת האנזים (פקד מעוכל) וחזור את ההליך לאחר הוספת האנזים (פקד מתעכל). דגירה שני צינורות באותו אופן כמו הספרייה ויטמינצ'יק.
- למחרת בבוקר, הוספת 5 µL של U/µL הין100 תרטר (500 יחידות סה כ) לפי aliquot, דגירה ב 37 מעלות צלזיוס במשך שעתיים תוך כדי טלטול-950 סל ד ב thermomixer.
-
בקרת מערכת העיכול: עבור הפקדים מעוכל, מעוכל (ראה 3.5.1), לבצע היפוך crosslink (שלב 6), פנול: כלורופורם חילוץ ו- DNA משקעים (שלב 7).
- עיצוב זוג תחל המתפרסים על אתר תרטר הין. באותו אזור, עיצוב עוד זוג תחל המתפרסים אל אתר תרטר הין. עיצוב תחל ה-PCR כמותי (Q-PCR) באמצעות Primer3 (http://bioinfo.ut.ee/primer3-0.4.0/) ואת הפרמטרים הבאים:
פריימר גודל: 20 אופטימלית (מינימלית: מקסימום 18,: 27); פריימר Tm: 60 אופטימלית (מינימלית: 57, מקסימום: 63); פריימר CG % תכנים: מינימלית: מקסימום 20: 80; אמפליקון גודל: RT-PCR ~ 100 bp (על-קונבנציונאלי PCR ~ 300 bp); Mispriming הספריה: האדם (primers אנושי) או מכרסם ופשוט (primers העכבר). - לבצע Q-PCR כדי לקבל 4 אומר Cts (סף מחזור): Ct [D; H], המתקבל המדגם מתעכל [ד] עם זוג תחל המתפרסים על אתר תרטר הין[H]; Ct [D;-], שהושג מדגם מתעכל [ד] עם זוג תחל המתפרסים אל אתר תרטר הין[-]; Ct [U; H], המתקבל המדגם מעוכל [U] עם זוג תחל המתפרסים על אתר תרטר הין; Ct [U;-], שהושג מדגם מעוכל [U] עם זוג תחל המתפרסים אל אתר תרטר הין[-]. לחשב את האחוזים של מערכת העיכול כמו: עיכול % = 100-100/2(Ct[D,H]-Ct[D,-]) - (Ct[U,H]-Ct[U,-]).
- עיצוב זוג תחל המתפרסים על אתר תרטר הין. באותו אזור, עיצוב עוד זוג תחל המתפרסים אל אתר תרטר הין. עיצוב תחל ה-PCR כמותי (Q-PCR) באמצעות Primer3 (http://bioinfo.ut.ee/primer3-0.4.0/) ואת הפרמטרים הבאים:
4. Biotinylation של הגבלת פרגמנט המסוכך
- להכין מיקס מאסטר biotinylation: 30.6 µL של 10 x הגבלת מאגר 2, µL 10.2 של H2O (ביולוגיה מולקולרית כיתה), 7.65 µL של 10 מ מ dCTP, µL 7.65 של 10 מ מ dGTP, µL 7.65 של 10 מ מ dTTP, µL 191.25 של 0.4 מ מ ביוטין-14-dATP ו- µL 51 של 5,000 U/mL DNA פולימראז אני (גדול פרגמנט Klenow).
- הוסף µL 60 של מיקס מאסטר biotinylation לכל aliquot, מיקס, דגירה ב 37 ° C עבור h 1 רועדת-700 סל ד (thermomixer) עבור 5 s, כל 30 s. לאחר 1 h, במקום aliquots על הקרח.
5. בגרעין מצדו
- להכין מיקס מאסטר מצדו: 510 µL של 10 x T4 DNA ליגאז מאגר, µL 51 של 10 מ"ג/מ"ל שור אלבומין (100 x BSA), µL 1754.4 של מים (ביולוגיה מולקולרית כיתה), µL 127.5 של 1 U/µL T4 DNA ליגאז (ראה טבלה של חומרים).
- להוסיף µL 479 של מיקס מאסטר מצדו לכל תערובת aliquot, דגירה ב 16 מעלות צלזיוס במשך 4 שעות רועדת-700 סל ד 5 s כל 2 דקות ב thermomixer.
- דגירה 30 דקות ב- RT.
6. Crosslink היפוך
- לשלב את כל aliquots ב שפופרת צנטרפוגה 50 מ ל (מתאים עבור צנטריפוגה במהירות גבוהה).
- להוסיף µL 62.5 של 10 מ"ג/מ"ל RNase A, מיקס, ולאחר תקופת דגירה של 30 דקות ב- 37 מעלות צלזיוס.
- להוסיף 300 µL של 10 מ"ג/מ"ל Proteinase K, מיקס, ולאחר תקופת דגירה של 30 דקות ב- 37 מעלות צלזיוס.
- דגירה התגובה O/N (או לפחות 4 שעות)-65 מעלות צלזיוס. למחרת בבוקר, להוסיף 300 µL של 10 מ"ג/מ"ל Proteinase K, מיקס, ולאחר תקופת דגירה של h 1-65 מעלות צלזיוס.
7. DNA טיהור
- הוסף µL 4337.5 של המאגר TLE (10 מ מ טריס-HCl pH 8.0; 0.1 מ מ EDTA pH 8.0), לערבב.
- להוסיף 1 נפח (10 מ"ל) פנול pH 8.0, מערבולת 10 s ו צנטריפוגה-RT ו- x 20,000 g עבור מינימלית 3 מ"ל להעביר 9 (מימית) השלב העליון צינור 50 מ.
התראה: פנול הוא חומר כימי מסוכן. בצע את תקנות בטיחות וגהות המתאים. - להוסיף 2 מ"ל TLE מאגר לשלב מימית הנותרים, מערבולת עבור 10 s ו צנטריפוגה-RT ו- x 20,000 g עבור מינימלית 3 להעביר 2.5 מ של שלב מימית לתוך הצינור החדש של צעד 7.2, שהופך את עוצמת הקול הסופי 11.5 מ ל. להתעלם שפופרת המכילה את השלב (אורגני) נמוך יותר.
- להוסיף נפח 1 (11.5 מ ל) של אלכוהול פנול: כלורופורם: isoamyl (25:24:1), מערבולת 10 s ו צנטריפוגה-RT ו- x 20,000 g עבור מינימלית 3 מ"ל 11 העברה (מימית) השלב העליון צינור 50 מ. חזור על שלב 7.3. אמצעי האחסון המדגם יהיה עכשיו 13.5 מ.
- להוסיף 1.35 מ של 3 מ' סודיום אצטט pH 5.2 ו 33.75 מ"ל אתנול 100% קר כקרח, מיקס, דגירה ב-80 מעלות צלזיוס במשך 45 דקות, או לחילופין בן לילה ב-20 ° C.
- צנטריפוגה-4 ° C ו- x 20,000 g 10 דקות להסיר את תגובת שיקוע, מחדש להשעות גלולה ב 1 מ"ל אתנול (vol/כרך) 70% טריות, להעביר צינור חדש.
- צנטריפוגה ב 4 ° C ו במהירות מלאה למשך 3 דקות ומפרידה benchtop ולאחר מכן להסיר את תגובת שיקוע.
- מחדש להשעות גלולה ב 1 מ"ל אתנול (vol/כרך) 70% קר קרח וחזור על שלב 7.7. יבש בגדר ב 37 מעלות צלזיוס למשך 10 דקות, להשעות מחדש ב µL 650 TLE המאגר. לקבוע את התשואה DNA על-ידי שימוש assay מבוסס על-ידי קרינה פלואורסצנטית לכמת כפול גדילי ה-DNA.
הערה: הפרוטוקול ניתן להשהות כאן על ידי הצמד הקפאה ואחסון המדגם ב-80 מעלות צלזיוס במשך מספר חודשים או ב-20 ° C לתקופה קצרה של זמן.
8. פקדים איכות
- הצג ספריית שלמות, מצדו על ידי אלקטרופורזה הדנ א. בניהול 200 ng הספרייה ב- 0.8% agarose/1 x TBE ג'ל. ה-DNA לפעול כלהקה מעל 10 kb.
- לאתר את סוג התא ידוע הקבועה קצר - ולא ארוך - range אינטראקציות באמצעות PCR קונבנציונלי. שימוש 100 ננוגרם של תבנית ה-DNA לפי תגובת ה-PCR. לעצב את תחל PCR מקרוב וגם כלפי באתרים מגבלת ההוראות לעיל (ראה 3.7.1). פריימר רצפים לבקרת איכות של העכבר וספריות ויטמינצ'יק האנושי מפורטים בטבלה 1.
-
מילוי ושליטה מצדו: לגזור את הלהקות ג'ל המכיל את amplicons של שליטה 8.2, ג'ל-תמצית ה-DNA ולהשתמש ה-DNA כתבנית עבור 4 תגובות PCR בודדים עם שילובים פריימר זהים.
- לטהר amplicons באמצעות ערכת טיהור PCR ולכמת את ריכוז הדנ א.
- להתכונן ארבע תגובות עיכול (היןתרטר [א], NheI [b], היןתרטר + NheI [c], אין אנזימים [d]) כל אמפליקון באמצעי אחסון הסופי של 15 µL: 500 ננוגרם של אמפליקון, µL 1.5 של 10 x הגבלת מאגר 2.1, µL 0.15 של 10 מ"ג/מ"ל שור אלבומין (100 x BSA) , ואת µL 0.1 (10 יחידות) של האנזים (היןתרטר [א], NheI [b], היןתרטר + מים [c] או NheI [ד]).
- לעיכול עבור h 1 ב 37 מעלות צלזיוס, ולאחר מכן להפעיל מערכת העיכול תגובות על 1.5% (wt/כרך) agarose/1 x TBE ג'ל.
9. DNA פיצול
- העברת µg 50.5 מדגם צינור חדש ולהוסיף TLE מאגר נפח סופי של 655 µL. פיצול הדגימה לתוך מבחנות sonication 5 (ראה טבלה של חומרים) על-ידי הוספת µL 130 של הספרייה (10 µg) כל מבחנה. להטיה לגודל של ~ 400 bp ב אולטרה-sonicator (ראה טבלה של חומרים) באמצעות הפרמטרים הבאים: פקטור חובה: 10%; שיא האירוע כוח (w): 140; מחזורים לכל פרץ: 200; זמן: 55 s.
- איסוף הדגימה sonicated בשפופרת mL 2 טריים.
10. בחירת גודל דו-צידית אלוהים אדירים-חרוז
- אלוהים אדירים מיקס (פאזה מוצקה הנייח הפיך) פתרון חרוז היטב על ידי היפוך, 1.85 מ של חרוז פתרון להעביר צינור ולהביא RT למשך 15 דקות.
- להוסיף 350 µL של מים (ביולוגיה מולקולרית כיתה) המדגם (נפח סופי 1 מ"ל).
- להוסיף 600 µL של אלוהים אדירים חרוז פתרון המדגם (נפח כולל מ ל 1.6; יחס של אלוהים אדירים פתרון ל- DNA: 0.6-1), תקופת דגירה של 5 דקות ב RT ו ספין מדגם צנטריפוגה benchtop עבור 2-3 s לאסוף דוגמה.
- פותחים את המכסה, למקם את הדגימה על הדוכן הפרדה מגנטית למשך 5 דקות, העברה ברורה supernatant לתוך צינור חדש ולבטל חרוזים.
- להתרכז חרוזים אלוהים אדירים עבור השלב השני של בחירת גודל: 930 להעביר µL של אלוהים אדירים חרוזים לתוך צינור חדש, במקום על הדוכן הפרדה מגנטית במשך 5 דקות, להשליך supernatant ברורה. מחדש להשעות את החרוזים ב µL 310 של אלוהים אדירים חרוז פתרון.
- להוסיף 300 µL של אלוהים אדירים מרוכז חרוזים (שלב 10.5) המדגם (נפח כולל 1.9 mL; יחס אלוהים אדירים פתרון ל- DNA הוא עכשיו 0.9 ל- 1), תקופת דגירה-RT של 5 דקות ו ספין לטעום ומפרידה benchtop במשך 2-3 ס פתוח בקפידה את המכסה , למקם את הצינורית על הדוכן הפרדה מגנטית למשך 5 דקות, ולמחוק את תגובת שיקוע.
- להוסיף 1 מ"ל אתנול 70% טריות (vol/כרך) הצינור מדגם על הדוכן הפרדה מגנטית, תקופת דגירה של s 30, ואת תגובת שיקוע להשליך. חזור על פעמיים.
- חרוזים יבש ב- 37 מעלות צלזיוס thermomixer (צינור המכסה פתוח) עבור לא יותר מ 5 דק להוסיף 300 µL מאגר TLE לדגימת, לערבב ולאחר תקופת דגירה של 10 דקות בטמפרטורת החדר.
- ספין מדגם צנטריפוגה benchtop עבור s 2-3, פותחים את המכסה, המקום הצינורית על הפרדה מגנטית עומד על 5 דק העברה לנקות את תגובת שיקוע לתוך צינור חדש ולמחוק את החרוזים.
11. ביוטין/Streptavidin נפתחים המוצרים מצדו
- להכין מאגרי: 1 x מאגר טרה-בתים (5 מ מ טריס-HCl pH 8.0 0.5 מ מ EDTA; 1 M NaCl; 0.05% Tween 20); 2 x NTB מאגר (10 מ מ טריס-HCl pH 8.0; 1 מ מ EDTA; 2 M NaCl); מאגר x NTB 1 (5 מ מ טריס-HCl pH 8.0; 0.5 מ מ EDTA; 1 M NaCl).
- להוסיף 200 µL של beads מגנטי מצמידים streptavidin, (ראה טבלה של חומרים) לתוך צינור חדש, למקם אותו על הדוכן הפרדה מגנטית עבור 1 דקות ולהסיר תגובת שיקוע.
-
רחץ חרוזים פעמיים עם µL 500 1 x טרה-בתים המאגר.
- עבור כל שלב שטיפת במהלך ביוטין נפתחים, סוף תיקון והסרה של ביוטין-קצוות הדנ א הלא-ובין אם לא, dATP עוקב, מתאם מצדו צעדים, מחדש להשעות את החרוזים למאגר המתאימים, סובב ב RT ו 15 סל"ד למשך 3 דקות, לסובב את הצינור ומפרידה benchtop עבור s 2-3, מקם את הצינורית על הדוכן הפרדה מגנטית למשך 3 דקות ולהסיר את תגובת שיקוע.
- להשעות מחדש חרוזים ב- 300 µL מאגר x NTB 2. לערבב חרוזים ודגימת (600 µL הנפח הכולל), דגירה-RT למשך 15 דקות על גלגל מסתובב במהירות של סל ד 3.
- להשיב חרוזים על הדוכן הפרדה מגנטית למשך 3 דקות ולהסיר את תגובת שיקוע ברורה. לשטוף חרוזים פעמיים ב- 500 µL 1 x NTB המאגר הראשון ולאחר מכן ב- µL 200 1 x מצדו המאגר. מחדש להשעות את החרוזים ב- 50 µL מאגר מצדו x 10.
12. סוף תיקון והסרה של ביוטין קצותיו DNA שאינם-ובין אם לא
- לשלב את הדגימה (50 µL סה כ) עם µL 50 של 2.5 מ מ dNTP מיקס (12.5 µL 10 מ מ של כל dNTP), 18.1 µL של 3,000 U/mL T4 DNA פולימראז, µL 18.1 של 10,000 U/mL T4 PNK, µL 3.7 של 5,000 U/mL DNA פולימראז אני גדול (Klenow) קטע , ואת µL 360.1 H2O.
- לערבב, דגירה ב- 20 ° C עבור h 1, רועד 5 s ב 700 סל ד כל 2 דקות ב thermomixer.
- להשיב חרוזים על הדוכן הפרדה מגנטית, להסיר את תגובת שיקוע ברורה, לשטוף חרוזים פעמיים ב µL 500 1 x מאגר טרה-בתים.
- לשטוף חרוזים ב µL 500 1 מאגר x NTB, ואחריו לרחוץ אחת 500 µL 1 x TLE.
- להשיב חרוזים על הדוכן הפרדה מגנטית, להסיר את תגובת שיקוע ברורה, וכן להשעות מחדש חרוזים ב 415 µL מאגר x TLE 1.
13. dATP עוקב
- לשלב לדוגמה (415 µL) עם 50 µL של 10 x הגבלת מאגר 2, 5 µL של 10 מ מ dATP, ו 30 µL של exo Klenow U/µL 5-מינוס....
- לערבב, דגירה ב 37 מעלות צלזיוס למשך 30 דקות, רועד 5 s ב 700 סל ד כל 2 דקות ב thermomixer.
- להשיב חרוזים על הדוכן הפרדה מגנטית, להסיר את תגובת שיקוע ברורה, לשטוף חרוזים פעמיים ב µL 500 1 x מאגר טרה-בתים.
- רחץ חרוזים ב 500 µL מאגר x NTB 1.
14. מתאם מצדו
- לשטוף חרוזים ב- µL 200 1 x מצדו תגובה מאגר (ראה טבלה של חומרים).
- להשעות מחדש חרוזים ב- µL 200 1 x מצדו תגובה המאגר. להוסיף 4 µL של ה-DNA ליגאז (ראה טבלה של חומרים) ו- µL 16 של 15 מיקרומטר מראש annealed מתאמי PE (anneal מראש על המתאמים PE על ידי ערבוב נפחים שווים של PE מתאם 1 ו- PE מתאם 2 (שניהם ב 30 מיקרומטר) ו המקננת לכמה דקות ב RT). דגירה-RT למשך 15 דקות.
- להשיב חרוזים על הדוכן הפרדה מגנטית, להסיר את תגובת שיקוע ברורה, לשטוף חרוזים פעמיים ב µL 500 1 x מאגר טרה-בתים.
- רחץ חרוזים ב 500 µL מאגר x NTB 1. אז, תשטוף חרוזי µL 100 1 x הגבלת מאגר 2, מחדש להשעות חרוזים ב- 50 µL 1 x הגבלת מאגר 2, ולהעביר לתוך צינור חדש.
15. היי-ספריית C הגברה
- להכין מיקס מאסטר PCR: µL 100 x Phusion 5 המאגר; 6 µL של מיקרומטר 25 PE PCR פריימר 1.0; 6 µL של מיקרומטר 25 PE PCR פריימר 2.0; µL 14 של dNTP מיקס (10 מ מ כל אחד); 6 µL של פולימראז Phusion; µL 318 של H2O.
- PCR מיקס מיקס מאסטר עם החרוזים (µL 500 סה כ), לחלק ב- 10 aliquots של 50 µL, ומגבירים מאת PCR תוך שימוש בתנאים הבאים:
בשנות ה-30 ב 98 ° C
7 מחזורים של: 10 s ב 98 ° C; 30 s ב- 65 מעלות צלזיוס; 30 s ב-72 מעלות
7 דקות ב-72 מעלות - לאסוף תגובות PCR לתוך צינור חדש, להשיב חרוזים על דוכן הפרדה מגנטית, ועל העברת supernatant (500 µL) לתוך צינור חדש.
-
לטהר את ה-DNA של הספרייה באמצעות חרוזים אלוהים אדירים.
- אלוהים אדירים מיקס חרוזים, להעביר µL 460 של חרוזי צינור חדש, להביא RT במשך 15 דקות להוסיף 450 µL של אלוהים אדירים חרוזים לתגובות PCR (נפח סופי 950 µL), תקופת דגירה של 5 דקות ב RT ו ספין מדגם צנטריפוגה benchtop עבור 2-3 s לאסוף דוגמה.
- פותחים את המכסה, למקם את הדגימה על הדוכן הפרדה מגנטית למשך 5 דקות, ולהסיר את תגובת שיקוע.
- שמירה על החרוזים על הדוכן הפרדה מגנטית, להוסיף 1 מ"ל של 70% אתנול (vol/כרך) מדגם צינור על פני שטח פנוי של חרוזים, להשאיר למשך 30 s, ואת תגובת שיקוע להשליך.
- חזור על שלב 15.4.3 עוד פעמיים.
- יבש חרוזים ב- 37 מעלות צלזיוס thermomixer (צינור המכסה פתוח) עבור לא יותר מ 5 דקות.
- להוסיף µL 51 TLE מאגר המדגם, מיקס, וכן תקופת דגירה של 10 דקות ב 37 מעלות צלזיוס, רועדת-950 סל ד ב thermomixer.
- ספין מדגם צנטריפוגה benchtop עבור s 2-3, פותחים את המכסה, המקום הצינורית על הפרדה מגנטית עומד על 5 דק העברה לנקות את תגובת שיקוע לתוך צינור חדש ולמחוק את החרוזים.
- לכמת את הריכוז של הספרייה ויטמינצ'יק. אחרי 7 סיבובים של ה-PCR-הגברה, אנו משיגים באופן שגרתי 500 – 1,500 ng של ויטמינצ'יק ספריה.
16. היברידית בפתרונות לכידת
הערה: חוסם ומאגר (SHS1-4) פתרונות להלן הם SureSelect קיט (ראה טבלה של חומרים).
- העברת 500 ng כדי µg 1 של ויטמינצ'יק ספריית לתוך צינור חדש ולא להתאדות דגימות ואקום רכז (ראה טבלה של חומרים; 45 ° C; לחץ ואקום: רמת 30.0, שיפוע 5) עד יבש.
- מחדש להשעות המתאיידים ויטמינצ'יק לספריה על-ידי הוספת 3.6 µL של H2O (ביולוגיה מולקולרית כיתה), 2.5 µL חוסם 1, µL 2.5 חוסם 2, ו- 0.6 µL חוסם מותאם אישית.
- העברת דגימה לבאר של רצועה חדשה של שפופרת PCR, לסגור עם רצועה קאפ PCR ומניחים על קרח. תוויות כמו "D" (עבור DNA-C).
- הכנת המאגר הכלאה: 12.5 µL של המאגר SHS1; µL 0.5 SHS2 המאגר; 5 µL SHS3 המאגר; µL 6.5 מאגר SHS4.
- דגירה-65 מעלות צלזיוס במשך 5 דקות thermomixer. להעביר לתוך טוב של רצועה חדשה של שפופרת PCR, לסגור עם רצועה קאפ PCR ולשמור על תווית RT. כמו "H" (עבור מאגר הכלאה).
- לבאר של רצועה חדשה של שפופרת PCR, לערבב µL 5 של 100 ng/µL biotinylated RNA רגשים (החנות ב- 80 ° C ו הפשרה על הקרח לפני השימוש); µL 0.5 מ B SRNase (מעכב RNase), 1.5 µL של H2O (ביולוגיה מולקולרית כיתה).
- סגור את רצועת צינור PCR עם רצועת קאפ PCR, המקום על קרח. תוויות כמו "R" (RNA).
- הגדרת ה-PCR מכונת באמצעות הפרמטרים הבאים:
5 דקות ב 95 מעלות צלזיוס; h 25-65 מעלות צלזיוס; המכסה מחוממת; 29 µL PCR התגובה נפח.
הערה: המשך מהר ככל האפשר במהלך כל ההליכים בעוד המכונה PCR פועל כדי למנוע אידוי הדגימה. - מניחים רצועת צינור "D" PCR המכונה PCR, סגור את המכסה המכונה PCR, ולהתחיל את תגובת ה-PCR. כאשר התוכנית PCR מגיעה 65 ° C, פותחים את המכסה המכונה PCR ולמקם רצועת צינור "H" PCR במכונת ה-PCR. סגור את מכסה המכונה PCR, תקופת דגירה של מינימום 3 פתח המכסה המכונה PCR, מקום ברכבת התחתית "R" PCR להתפשט על המכונה PCR, וסגור את המכונה PCR.
- לאחר 2 דקות, פתח שאת מכסה המכונה PCR ו- PCR כל צינור רצועות. להעביר µL 13 של טוב "H" לתוך טוב "R", ואז כל אמצעי האחסון של ד' טוב טוב "R". Pipet למעלה ולמטה 3 פעמים כדי לערבב את התגובה, לסגור את רצועת צינור PCR, הסר "H" ו- "D" PCR צינור רצועות וסגור מכסה המכונה PCR. דגירה התגובה ב 65 מעלות צלזיוס במשך 24 שעות ביממה.
17. בידוד של יזם רסיס המכיל מוצרים מצדו
הערה: השלבים הבאים מומלץ לסיים עם ערכת מתאם SureSelect וספריית (ראה טבלה של Materals).
- לחמם 1.5 מ של מאגר לשטוף 2 עבור דגימה ב 65 ° C מראש.
- להוסיף 60 µL של streptavidin מצמידים beads מגנטי (ראה טבלה של חומרים) לתוך צינור חדש, במקום על הדוכן הפרדה מגנטית עבור 1 דקות ולהסיר תגובת שיקוע.
- רחץ חרוזים שלוש פעמים עם µL 200 1 x מאגר מחייב.
הערה: עבור כל שלב שטיפת בתקופת הבידוד שלאחר לכידתו של מוצרים המכילים מקדם מצדו, מחדש להשעות חרוזים למאגר המתאימים, לסובב למשך 3 דקות ב RT, סל ד 15 על גלגל מסתובב, ברכות לסובב את הצינור ומפרידה benchtop עבור 2-3 s לאסוף לדוגמה, במקום הצינורית על הפרדה מגנטית לעמוד במשך 3 דקות, הסר תגובת שיקוע. - להשעות מחדש חרוזים ב- µL 200 1 x מאגר מחייב. פתח את המכונה PCR, רצועת צינור ה-PCR (בעוד עדיין פעולתה של תוכנית ה-PCR) ולהעביר את התגובה הכלאה לתוך הצינור עם החרוזים מגנטי. דגירה-RT במשך 30 דקות על גלגל מסתובב במהירות של סל ד 3.
- להשיב חרוזים על הדוכן הפרדה מגנטית ולהסיר את תגובת שיקוע ברורה. מחדש להשעות חרוזים לתוך µL 500 שטיפת מאגר 1, מיקס, ואת תקופת דגירה של 15 דקות ב- 20 ° C תוך טלטול-950 סל ד ב thermomixer.
- להשיב חרוזים על הדוכן הפרדה מגנטית למשך 3 דקות ולהסיר את תגובת שיקוע ברורה. מחדש להשעות חרוזים לתוך µL 500 שטיפת מאגר 2, לערבב, דגירה 10 דקות ב 65 ° C תוך טלטול-950 סל ד ב thermomixer. חזור על שלב 17.5 עוד פעמיים.
- להשיב חרוזים על הדוכן הפרדה מגנטית, להסיר את תגובת שיקוע ברור והשהה מחדש חרוזים ב- µL 200 1 x הגבלת מאגר 2. להשיב חרוזים על הדוכן הפרדה מגנטית, להסיר את תגובת שיקוע, מחדש להשעות חרוזים לתוך 30 µL של 1 x הגבלת מאגר 2.
18. PCHi-C Library הגברה
- להכין מיקס מאסטר PCR: 60 µL x PCR 5 המאגר (מאגר Phusion), µL 3.6 של 25 מיקרומטר PE PCR פריימר 1.0, 3.6 µL של 25 מיקרומטר PE PCR פריימר 2.0, µL 8.4 dNTP מיקס (10 מ מ כל אחד), µL 3.6 של פולימראז Phusion ו µL 190.8 של H2O.
- PCR מיקס מיקס מאסטר עם התהום (µL 300 סה כ), חרוזים ב 6 aliquots של 50 µL, ומגבירים PCR-שימוש בתנאים הבאים:
30 s ב 98 ° C
4 מחזורים של: 10 s ב 98 ° C, 30 s ב 65 ° C, 30 s ב-72 מעלות
7 דקות ב-72 מעלות - לאסוף את כל תגובות PCR צינור חדש, להשיב את החרוזים על המגנט, ולהעביר תגובת שיקוע (300 µL; מכיל ספריית PCHi-C) ל צינור חדש.
- לטהר הספרייה PCHi-C באמצעות חרוזים אלוהים אדירים, השלבים המתוארים לעיל תחת 15.4.
- לכמת את הריכוז של ספריית ה PCHi-C.
תוצאות
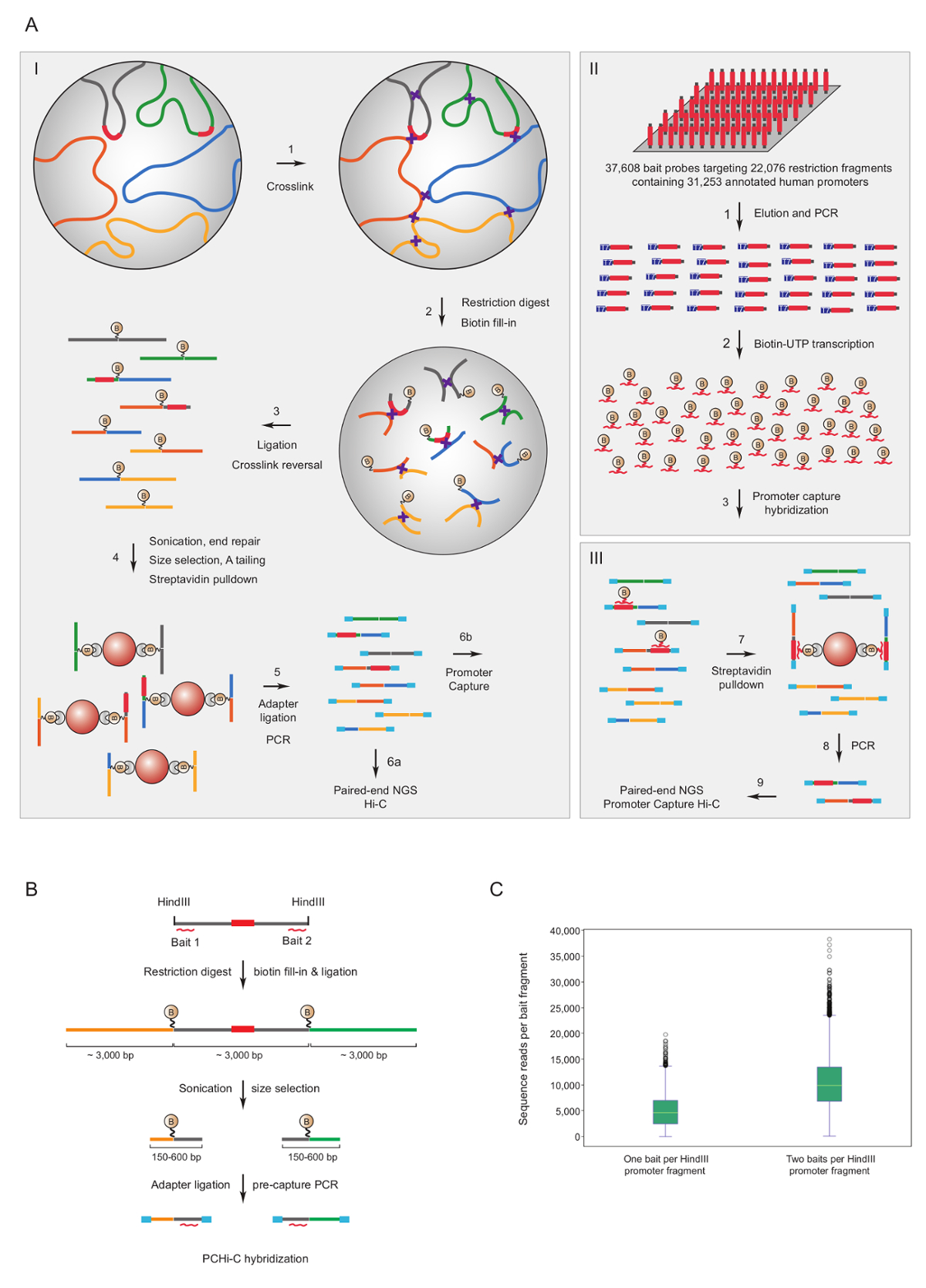
יזם ללכוד ויטמינצ'יק שימש כדי להעשיר את העכבר7,34,36,39 וספריות האנושי33,35,37,38 ויטמינצ'יק עבור מקדם אינטראקציות. פרוטוקול דומה (בשם HiCap) תואר על ידי קבוצה זנדברג40. איור 1A מראה סכימטי זרימת העבודה עבור יזם ללכוד היי-סי בפרוטוקול המתוארים כאן, ויטמינצ'יק ספריות נוצרות תוך שימוש בגרעין מצדו41, שתוצאתה מספר מוצרים מצדו כדין42מופחתת באופן משמעותי. עבור PCHi-C, העכבר מורכב או ספריות ויטמינצ'יק האנושית מוכתבת עליהם בבית-פתרון הכלאה, ללכוד באמצעות RNAs biotinylated 39,021 משלים 22,225 העכבר המכילים מקדם HindIII הגבלת קטעים, או 37,608 biotinylated RNAs מיקוד 22,076 אנושי המקדם המכיל HindIII הגבלת שברים, בהתאמה. ניתן לפלח המקדם המכיל קטעי הגבלת הקצוות או שניהם על ידי בודדים biotinylated RNAs (איור 1B). מצאנו כי לכידתו של שניהם מסתיים כיסוי משופרת של היזמים בודדים (איור 1C; קריאות רצף raw) כמעט כפולה, כצפוי. וכך, בכל פעם האפשר (קרי, באזורים שאינם חוזרות), אנו ממליצים להשתמש biotinylated RNAs משלים בשני הקצוות של קטע הגבלת ללכידה.
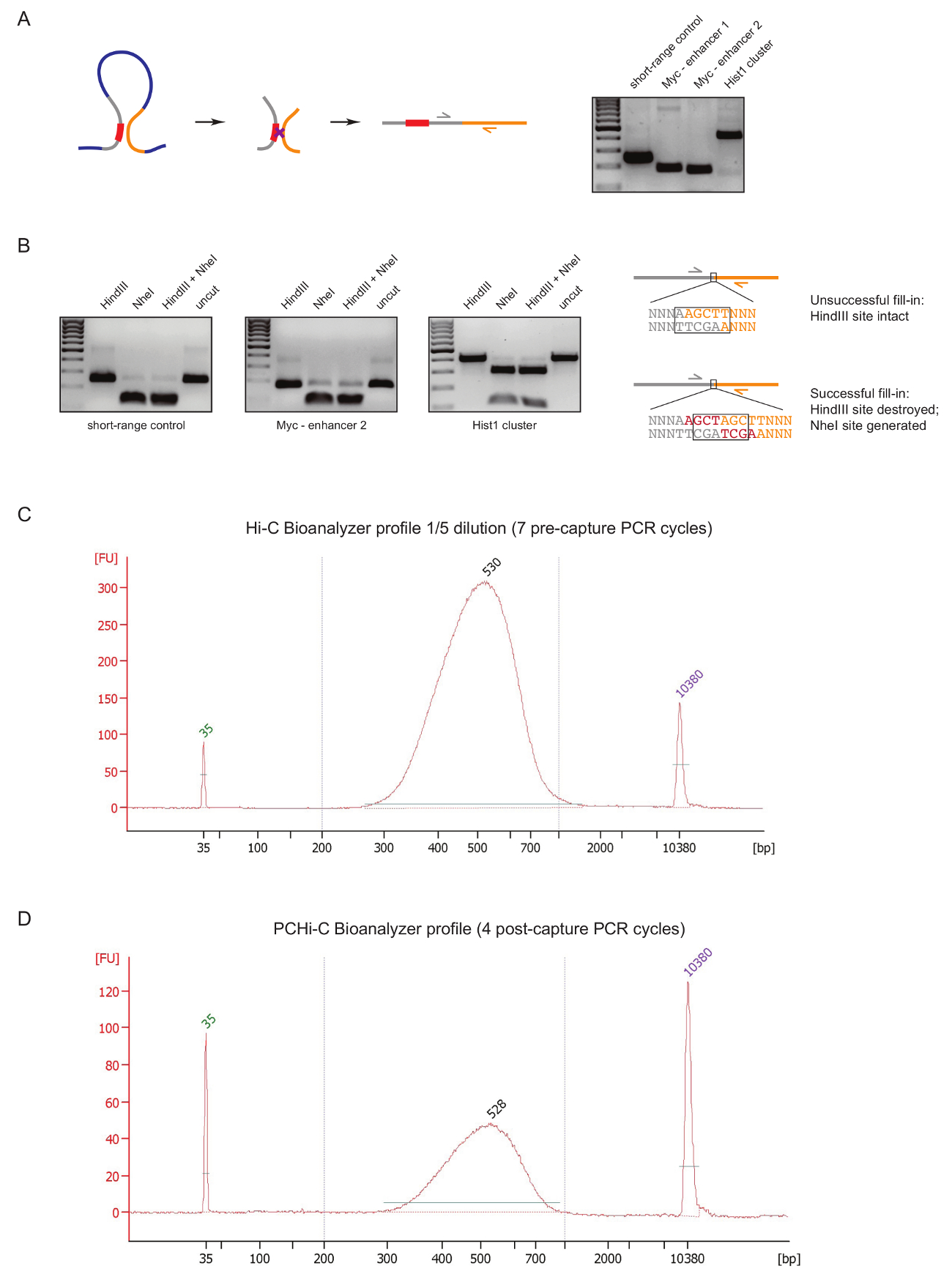
כדי להעריך איכות ספריית PCHi-C בשלב מוקדם במהלך הכנת ספריה, אנו מבצעים שני פקדים לאחר ה-DNA מצדו, טיהור, כפי שתואר לעיל31. הראשונה היא להשתמש זוגות פריימר ספציפיים כדי להגביר מצדו מוצרים כמו 3 ג27. אנו משתמשים פריימר זוגות (טבלה 1) כדי להגביר את סוג התא הקבועה מצדו ארוכי טווח מוצרים, כגון בין הגן Myc , משפרי הידוע שלה ממוקם כ 2 מגה-בתים משם (איור 2 א) או בין הגנים של (לוקוס Hist1 המופרדים 1.5 Mb), בין שני אזורים ממוקם בסמיכות ליניארי ('פקד לטווח קצר').
בקרת האיכות השנייה מתבצעת כדי לקבוע את היעילות של התאגדות ביוטין במהלך מילוי בתיווך Klenow של הגבלת המסוכך אתר עם ביוטין-dATP. מילוי Klenow מוצלחת ותוצאות מצדו בלנט-סיום עוקבות בהיעלמותו של אתר ההגבלה המקורי בין מולקולות הדנ א של מוצר מצדו, ובמקרה של HindIII להיווצרות של אתר הכרה NheI חדש (איור 2B ). היחס של HindIII למוצר מצדו מתעכל NheI הוא הבדיקה ישירה של ביוטין התאגדות יעילות. ספרייה ויטמינצ'יק באיכות ירודה יראה רמה גבוהה של עיכול HindIII, ואילו באיכות גבוהה ספריות יש כמעט מוחלטת NheI עיכול מצדו מוצרים (איור 2B).
לאחר ויטמינצ'יק ספריית הכנה (קרי, לאחר ביוטין-streptavidin למשוך למטה בגודל שנבחר ויטמינצ'יק מצדו, מתאם מצדו ומוצרי לכידת קדם PCR), התפלגות גודל של תקינות של הספרייה ויטמינצ'יק הוא מוערך על ידי Bioanalyzer (איור 2 C). באותו הפקד מתבצע בסוף PCHi-C ספריית הכנה (קרי, לאחר לכידתו הכלאה של פוסט-לכידת PCR ומוצרים מצדו המכילים מקדם). השוואה של הפרופילים Hi-C ו- Bioanalyzer PCHi-C עולה כי כצפוי, ויטמינצ'יק ספריות מרוכזים הרבה יותר מאשר ספריות PCHi-C המתאימה, אך התפלגות גודל של הספריות דומה מאוד, המציין הלכידה להתערב PCHi-C אינו מציג דעה קדומה גודל (איור 2C, יח).
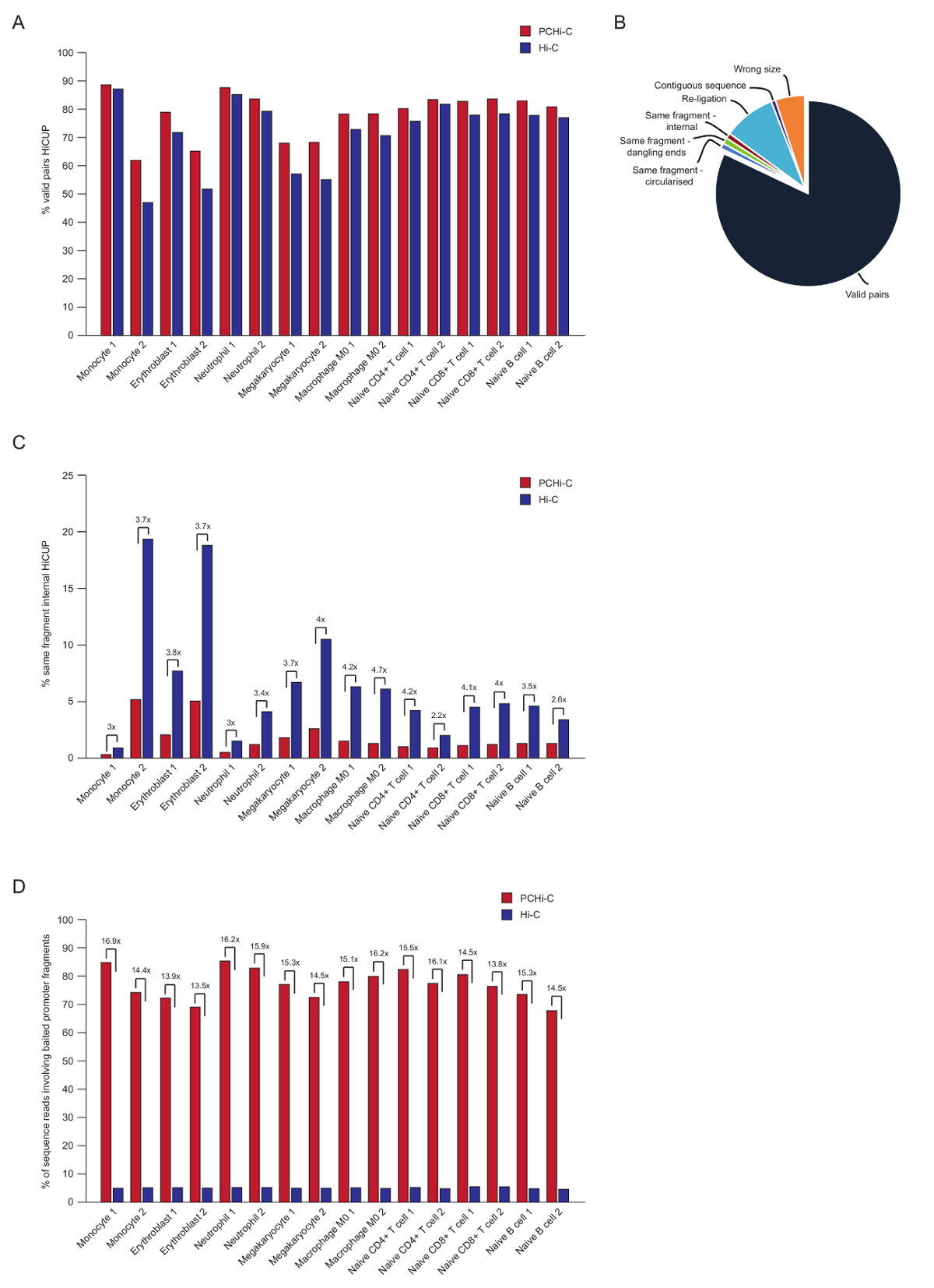
לאחר רצף לזווג-end, קריאות PCHi-C ממופים, איכות מבוקר, לסנן את צינור HiCUP43. באיכות גבוהה PCHi-C ספריות מכילים בין 70-90% 'זוגות חוקי"(קרי, סוף-לזווג רצף קורא בין שני קטעים ההגבלה זה לא שכנים על המפה גנומית ליניארית; איור 3 א, ב'). שימוש בגרעין מצדו פרוטוקול41,42, האחוז של טרנס לקרוא זוגות (קרי, סוף-לזווג רצף קורא בין שני קטעים הגבלת הנמצאות על כרומוזומים שונים) הם בדרך כלל נמוך, בין 5 ל- 25%, המשקף את קיומו של כרומוזום בשטחים, המציין ספריה טובה איכות. השוואה ישירה של אחוז 'זוגות חוקי' בין ספריות ויטמינצ'יק שלהם ספריות המתאימים PCHi-C35, מראה כי בכל המקרים האחוז של זוגות חוקי הוא גבוה בספריות PCHi-C (איור 3B). כל זה מלווה ירידה באחוז לא חוקית עבור 'אותו קטע פנימי' קריאות ב PCHi-C (איור 3C). זה צפוי, כמו השלב לכידת לא רק מעשיר למוצרים מצדו המכילים מקדם, אלא גם למטרות פרגמנט ההגבלה, בשל מיקומו של oligos ללכוד על הגבלות שברים (ראה איור 1B).
לאחר HiCUP סינון, נוכל לקבוע את היעילות הלכידה. PCHi-C ספריות מכילים שלושה סוגי הקריאות רצף חוקי לאחר סינון HiCUP:
1.) יזם: הגנום קורא (קרי, קריאות בין קטע יזם שנתפסו קטע ההגבלה HindIII הלא-יזם בכל מקום בגנום)
2.) יזם: יזם קורא (קריאות בין שני קטעים יזם שנתפסו)
גנום 3.): הגנום קורא (רקע ויטמינצ'יק מצדו המוצרים שבו אף אחד מבני הזוג המוצר מצדו ממפה אל מקדם שנתפסו). אלה יימחקו לפני ניתוחים במורד הזרם.
באיכות גבוהה PCHi-C ספריות יש לכידת יעילות (סכום של קטגוריות 1 ו- 2 לעיל) בין 65-90% (איור תלת-ממד). השוואה ישירה לספריות ויטמינצ'יק מראה כי PCHi-C תוצאות ~ 15-fold העשרה למוצרים המכילים מקדם מצדו (איור תלת-ממד), בחלק מהמקרים 17-fold. . זה קרוב למקסימום היפותטי (19.6-fold) העשרה עבור PCHi-C, אשר תלויה האחוז של השברים הגבלת הגנום מכוסה על ידי מערכת לכידת. העשרה גדולה יכולה להיות מושגת על ידי עיצוב מערכות הלכידה מיקוד פחות הגבלה שברי45,44,46.
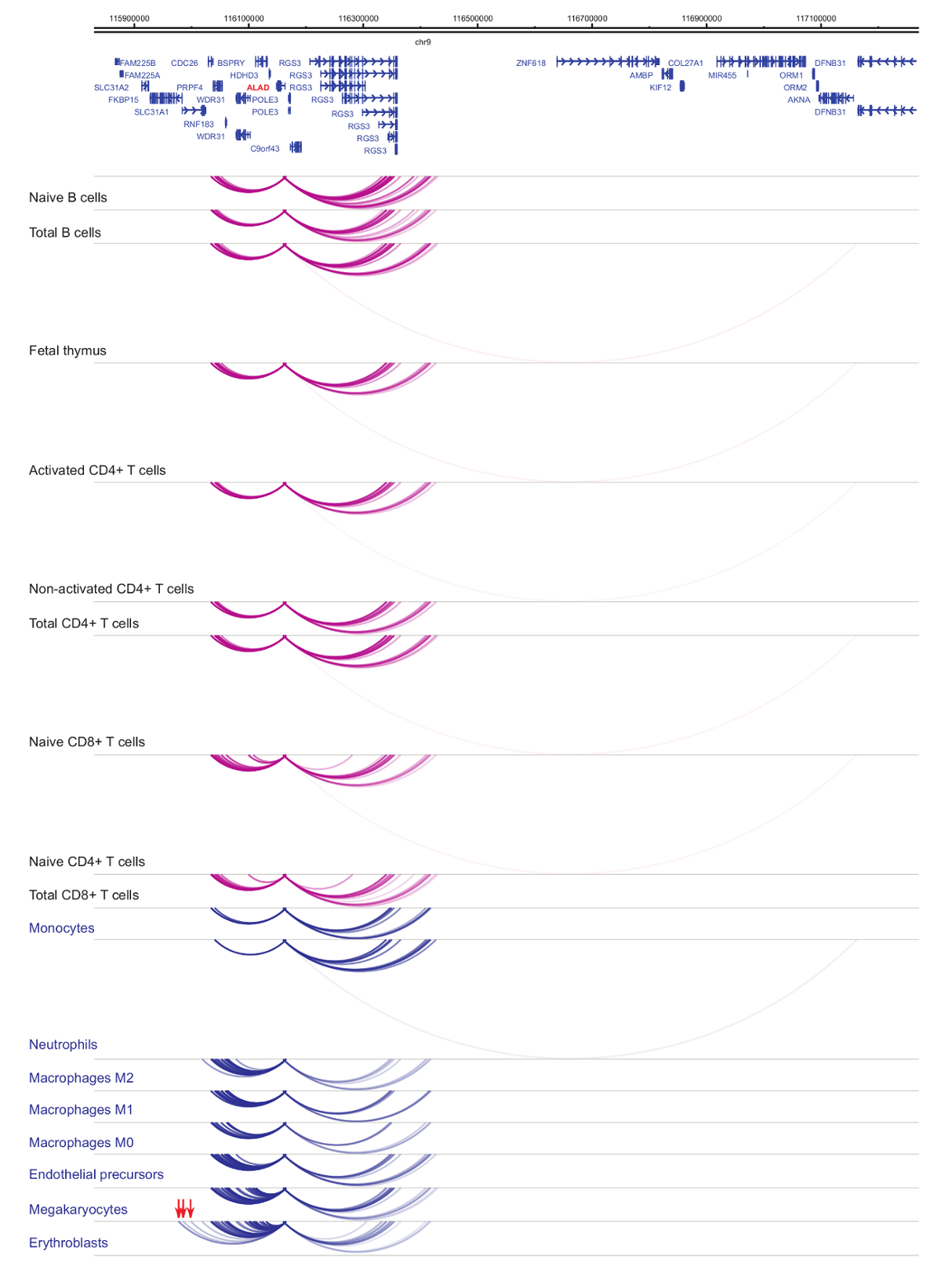
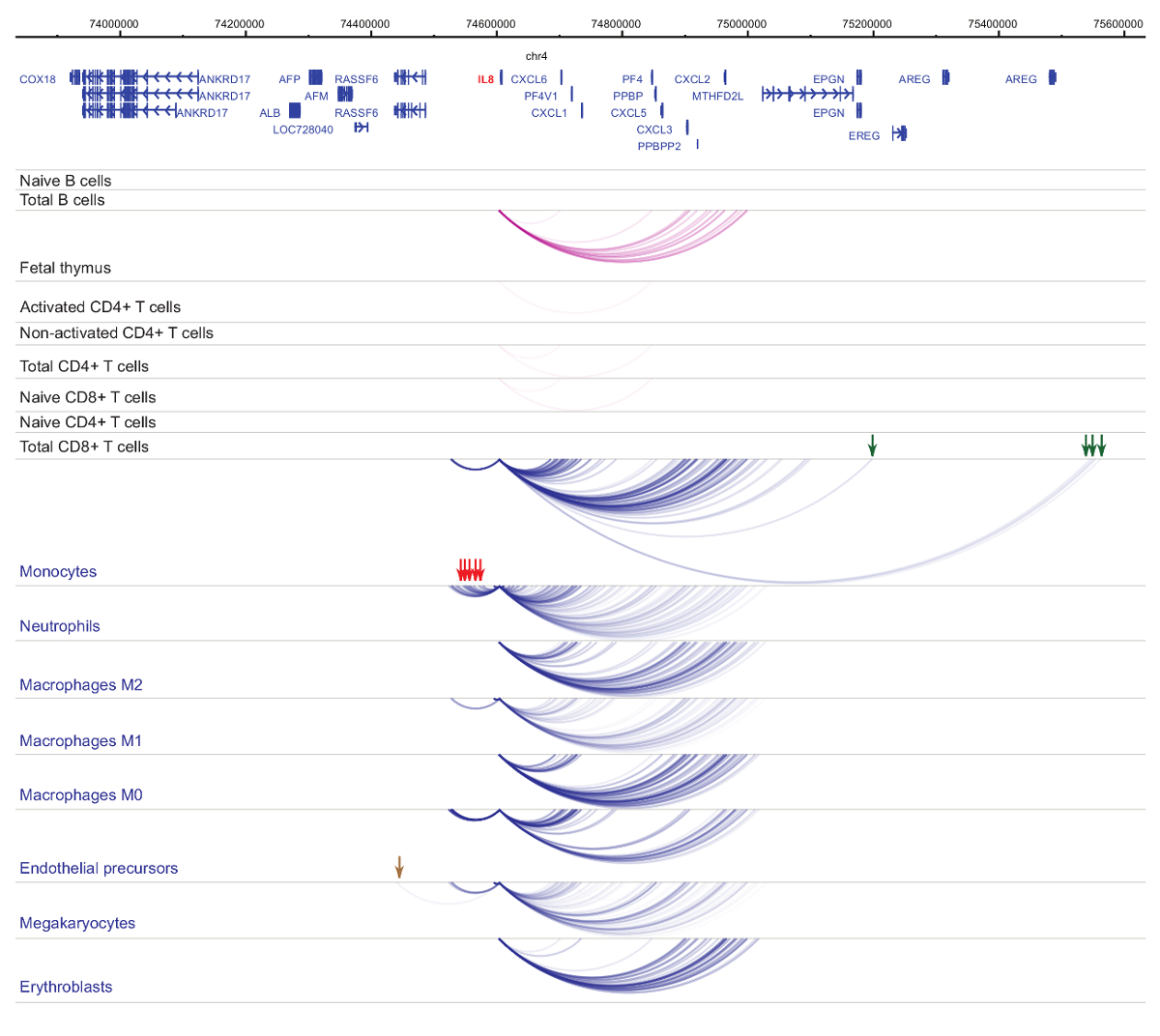
ניתוח של יזם interactomes מדגים תא סוג וספציפיות שושלת היוחסין33,34,35, בשינויים בולטת במהלך התמיינות37,38,39 . דמויות 4 ו- 5 להציג דוגמאות של שושלת היוחסין ירידה לפרטים ובידול dynamics היזמים ספציפיים. לדוגמה, ALAD מתבטאת צורונים כל התאים אבל הביטוי שלה הוא upregulated של erythroblasts47. יזם ALAD אנשי קשר מספר קטעים דיסטלי בכל תאי hematopoietic, עוסקת אינטראקציות נוספות בפרט erythroblasts (איור 4). IL-8 מראה אינטראקציות משמעותיות מבחינה סטטיסטית אין תאים B, מעט מאוד קשרי הגומלין של תאי T, אבל עשרות אינטראקציות בתאים של שושלת היוחסין מיאלואידית, כולל אינטראקציות ספציפיות סוג התא (monocytes, נויטרופילים, megakaryocytes איור 5). דוגמאות אלה מציגות כמה PCHi-C יכול לשמש כדי לפענח interactomes ספציפית סוג התא, לזהות אזורים יזם-אינטראקציה עם פוטנציאל רגולטוריות.

איור 1 : יזם ללכוד ויטמינצ'יק הרציונל וללכוד פיתיון עיצוב. (א) סכמטי של זרימת עבודה של PCHi-סי בגרעין מצדו ויטמינצ'יק41,42 (א) ואחריו בבית-פתרון הכלאה עם פיתיונות biotinylated RNA (II) מיקוד הרסיסים ההגבלה של כולנו בני אנוש (המתואר כאן) או עכבר ג'ין היזמים (III). (B) פיתיון עיצוב עבור PCHi-סי Biotinylated RNA לכידת פיתיונות (קווים מעוקלים אדום) מיועדים נגד הקצוות של קטעים הגבלת המכילים מקדם (אפור; הערה כי רצפי יזם את עצמם (אדום) הם רק מטרה פיתיונות ללכוד הרנ א אם הם ממוקמים-הגבלה המקטע מסתיים). המוצרים מצדו המורכב המכילים מקדם הגבלת קטעים (אפור) ו שברי הגבלת שמעצבת שלהם (צהוב וירוק) הם מבודדים באמצעות רצף-משלימים את החסר הכלאה בין פיתיון RNA DNA היעד, ובעקבות ביוטין-streptavidin הנפתח, כמוצג א (ג) השוואה של PCHi-C לכידת יעילות עבור יזם המכיל קטעים הגבלת מטרה אחת RNA פיתיון לכידת המכשיר לעומת שני RNA פיתיון לכידת רגשים (ראה מפרטים טכניים ב'). אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 2 : פקדים איכות רצף טרום PCHi-C. (א) שמאלה, סכמטי של סמיכות מרחבית בין יזם פיר, והתוצאה היא מוצר מצדו ויטמינצ'יק המורכב קטע הגבלת המכילים מקדם (אפור; יזם רצף באדום) ו קטע הגבלת פיר (צהוב). נכון, ה-DNA ג'ל אלקטרופורזה מציג דוגמאות של מוצרים מצדו ויטמינצ'יק מוגבר באמצעות פריימר ספציפי זוגות (כפי שהיא מתוארת מפרטים טכניים בצד השמאל). (B) שמאלה, דוגמאות מייצגות של ההגבלה HindIII, NheI ו- HindIII/NheI מעכל של ויטמינצ'יק מצדו מוצרים (PCR המוצרים המוצגים ב א). נכון, תיאור סכמטי של ה-DNA רצף לאחר ויטמינצ'יק מצדו בעקבות כושל (למעלה) או מילוי Klenow dNTP מוצלחת (למטה) של הגבלת צמתים וכן מצדו עוקבות. (ג) נציג ויטמינצ'יק ספריית bioanalyzer פרופיל (דילול 1/5). (ד) נציג PCHi-C ספריית bioanalyzer פרופיל (ללא דילול). אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 3 : PCHi-C שלאחר רצף פקדים איכות. (א) השוואה של אחוז רצף חוקי לקרוא זוגות לאחר HiCUP43 עיבוד ב- vs PCHi-C המתאים ויטמינצ'יק ספריות (נתונים מ. Javierre et al., 201635). (B) נציג HiCUP PCHi-C התוצאה מראה חוקי לקרוא זוגות וקטגוריות אחרות רצף זה יימחקו לפני ניתוחים במורד הזרם (נתונים מ. Javierre et al., 201635). (ג) השוואה של אחוז 'באותו מקטע פנימי' קורא לאחר HiCUP עיבוד ב- vs PCHi-C המתאים ויטמינצ'יק ספריות (נתונים מ. Javierre et al., 201635). (ד) השוואה של אחוז רצף הקראת קטעים יזם פתיונים מעורבים (לכידת יעילות) נגד PCHi-C המתאים ויטמינצ'יק ספריות (נתונים מ- Javierre. et al., 201635). אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 4: ALAD PCHi-C פרופיל בתאי אדם hematopoietic. מקדם אינטראקציות של סוגי התאים מיאלואידית מוצגים קשתות כחול, מקדם אינטראקציות של סוגי תאים הלימפה מוצגים קשתות סגול. אינטראקציות ספציפיות erythroblast מסומנים באמצעות חצים אדומים (נתונים מ. Javierre et al., 201635). אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 5 : IL8 PCHi-C פרופיל בתאי אדם hematopoietic. מקדם אינטראקציות של סוגי התאים מיאלואידית מוצגים קשתות כחול, מקדם אינטראקציות של סוגי תאים הלימפה מוצגים קשתות סגול. אינטראקציות ספציפיות מונוציט מסומנים באמצעות החצים הירוקים, אינטראקציות ספציפיות נויטרופילים מסומנים באמצעות חצים אדומים ו אינטראקציה מגקריוציט ספציפיים מסומן על ידי חץ חום (נתונים מ. Javierre et al., 201635). אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}
| אנושי | ||||||||
| שם | רצף | כרומוזום | סטרנד | תתחיל GRCh38/hg38 | סוף GRCh38/hg38 | פריימר שילובים כדי לבדוק אינטראקציות 3 ג של ביוטין התאגדות | ||
| hs AHF64 דקר | GCATGCATTAGCCTCTGCTGTTCTCTGAAATC | 11 | + | 116803960 | 116803991 | להשתמש בשילוב עם hs דקר AHF66 | ||
| hs AHF66 דקר | CTGTCCAAGTACATTCCTGTTCACAAACCC | 11 | + | 116810219 | 116810248 | להשתמש בשילוב עם hs דקר AHF64 | ||
| hs MYC לוקוס | GGAGAACCGGTAATGGCAAA | 8 | - | 127733814 | 127733833 | השתמש בשילוב עם hs MYC +1820 או hs MYC-538 | ||
| hs MYC +1820 | AAAATGCCCATTTCCTTCTCC | 8 | + | 129554527 | 129554547 | להשתמש בשילוב עם hs MYC לוקוס | ||
| hs MYC-538 | TGCCTGATGGATAGTGCTTTC | 8 | - | 127195696 | 127195716 | להשתמש בשילוב עם hs MYC לוקוס | ||
| hs HIST1 F | AAGCAGGAAAAGGCATAGCA | 6 | + | 26207174 | 26207193 | להשתמש בשילוב עם hs HIST1 R | ||
| hs HIST1 R | TCTTGGGTTGTGGGACTTTC | 6 | + | 27771575 | 27771594 | להשתמש בשילוב עם hs HIST1 F | ||
| העכבר | ||||||||
| רצף | כרומוזום | סטרנד | תתחיל GRCm38/mm10 | סוף GRCm38/mm10 | פריימר שילובים כדי לבדוק אינטראקציות 3 ג של ביוטין התאגדות | |||
| TCATGAGTTCCCCACATCTTTG | 8 | + | 84841090 | 84841111 | להשתמש בשילוב עם מ מ Calr2 | |||
| CTGTGGGCACCAGATGTGTAAAT | 8 | + | 84848519 | 84848541 | להשתמש בשילוב עם מ מ Calr1 | |||
| TATCAAGGGTGCCCGTCACCTTCAGC | 6 | + | 125163098 | 125163123 | להשתמש בשילוב עם Gapdh4 דקר | |||
| GGGCTTTTATAGCACGGTTATAAAGT | 6 | + | 125163774 | 125163799 | להשתמש בשילוב עם Gapdh3 דקר | |||
| GGAGGAGGGAAAAGGAGTGATT | 6 | + | 52212829 | 52212850 | להשתמש בשילוב עם מ מ Hoxa13 | |||
| CAGGCATTATTTGCTGAGAACG | 6 | - | 52253490 | 52253511 | להשתמש בשילוב עם מ מ Hoxa7 | |||
| GGGTAATGGTGTCACTAACTGG | 13 | + | 23571284 | 23571305 | להשתמש בשילוב עם מ מ Hist1h3e או מ מ Hist1h4i | |||
| GGGTTTGATGAGTTGGTGAAG | 13 | + | 23566541 | 23566561 | להשתמש בשילוב עם מ מ Hist1h2ae | |||
| TTGGGCCAAAGCCTATATGA | 13 | + | 22043085 | 22043104 | להשתמש בשילוב עם מ מ Hist1h2ae | |||
טבלה 1: פריימר רצפים לבקרת איכות של האדם וספריות העכבר ויטמינצ'יק.
Discussion
העיצוב המודולרי של יזם ללכוד Hi-C
יזם ללכוד ויטמינצ'יק מיועדת להעשיר באופן ספציפי ויטמינצ'יק ספריות עבור אינטראקציות מעורבים היזמים. אינטראקציות אלה מהווים רק ערכת משנה של מצדו שהמוצרים בספריה ויטמינצ'יק.
לכידת ויטמינצ'יק בקלות יכול להיות שונה כדי להעשיר את ויטמינצ'יק ספריות עבור כל אזור גנומית או אזורים של עניין על-ידי שינוי שיטת הלכידה. לכידת אזורים יכולים להיות מקטעי גנומית רציף44,45,46,48, משפרי כי כבר זיהו PCHi-C (' הפוך ללכוד Hi-C'35), או DNase אני רגיש אתרי49 . ניתן לכוונן את הגודל של מערכת לכידת בהתאם להיקף ניסיוני. לדוגמה, דריידן ואח. יעד 519 שברי פיתיון במדבריות שלושה גנים הקשורים עם סרטן השד44. מערכת לכידת מאת מרטין ואח. מטרות שני המקטעים גנומית רציפה ('לכידת אזור': 211 אזורים גנומית סה כ; 2,131 שברי ההגבלה) ונבחר היזמים (היזמים ג'ין 3,857)45.
SureSelect ספריות זמינים בטווחי גודל שונה: 1 kb עד 499 kb (5,190 – 4,806), 500 kb ל 2.9 Mb (5,190 – 4,816), ו- 3 Mb ל- 5.9 Mb (5,190 – 4,831). כמו כל לכידת הפרט ביוטין-RNA ארוכה 120 נוקלאוטידים, אלה ללכוד מערכות להכיל לכל היותר 4,158, 24,166 ו- 49,166 בודדים ללכוד הגששים, בהתאמה. זה תואם את 2,079, 12,083, 24,583 הגבלה יישוב שברים, בהתאמה (שימו לב כי המספרים עבור הגבלת קטעים ובכמה מבוסס על ההנחה כי ניתן לעצב שני רגשים לכידת נפרדים עבור כל הגבלה פרגמנט – במציאות עקב רצפים חוזרים, זה לא יהיה המקרה עבור כל הגבלה קטע (ראה גם איור 1B, C), וכתוצאה מכך מספר גבוה יותר של הגבלת targetable קטעים עבור מספר קבוע של הגששים לכידת זמין ).
הפרוטוקול המתואר כאן מבוססת על השימוש באנזים הגבלה עם אתר הכרה bp 6 לחשוף את האינטראקציות ארוכי טווח. שימוש באנזים הגבלה עם אתר הכרה bp 4 עבור רזולוציה גדולה יותר מאינטראקציות מקורב יותר הוא גם אפשרי40,49.
מגבלות של PCHi-C
מגבלה אחת הטבועה של כל כרומוזום קונפורמציה לכידת מבחני היא כי הרזולוציה שלהן נקבע על-ידי האנזים הגבלה בשימוש עבור הדור הספרייה. האינטראקציות המתרחשות בין רכיבי ה-DNA ממוקם על השבר הגבלה זהה הם בלתי נראה מבחני 'C-type'. יתר על כן, PCHi-c, במקרים מסוימים יותר מאשר אתר התחלת תיעתוק אחד יכול להימצא על השבר הגבלה זהה המכילים מקדם, חיישנים בחלק מהמקרים הנמל בשני סימני היסטון דכאניים האקטיביות, ולכן קשה לאתר שבו הרגולציה אלמנטים לתווך את האינטראקציות, וגם כדי לחזות את הפלט הרגולציה של מקדם אינטראקציות. באמצעות אנזימי הגבלה עם ארבעה אתרי זיהוי bp מפחית את הסיכון של בעיה זו אבל בא על חשבון ומעמד ויטמינצ'יק ספריית המורכבות (ויטמינצ'יק ספריות הנוצרת באמצעות אנזימי הגבלה אתר הכרה bp 4 לפחות פי 100 יותר מורכבות Hi-C ספריות שנוצר עם 6 bp זיהוי אתר אנזימי הגבלה), את עלויות עבור רצף הדור הבא.
מגבלה נוספת היא כי פרוטוקול PCHi-C הנוכחי דורש מיליוני תאים כמתחילה חומר, מסלק הניתוח של מקדם אינטראקציות סוגי תאים נדיר. גרסה מותאמת של PCHi-C כדי לאפשר חקירת אנשי קשר יזם אוכלוסיות תאים עם 10,000 עד 100,000 תאים (לדוגמה תאים במהלך התפתחות עוברית מוקדמת או תאי גזע hematopoietic) ולכן יהיה תוספת רבת ערך הלכידה שלום-C בארגז הכלים.
לבסוף, כמו כל שיטות המסתמכות על קיבוע פורמלדהיד, PCHi-C רק רשומות אינטראקציות 'קפואים' בנקודת זמן של קיבעון. לפיכך, כדי ללמוד את קינטיקה ואת הדינמיקה של מקדם אינטראקציות, שיטות כגון רזולוציה סופר תא חי מיקרוסקופ נדרשים לצד PCHi-סי
שיטות לנתח הארגון המרחבי כרומוזום ברזולוציה גבוהה
המורכבות העצום של ספריות אינטראקציה כרומוזומלית אוסר על זיהוי אמין של מוצרים אינטראקציה בין שני קטעים הגבלה ספציפית עם מובהקות סטטיסטית. כדי לעקוף בעיה זו, נעשה שימוש רצף לכידת להעשיר ויטמינצ'יק33,34,40,44 או 3 ג50,51 ספריות עבור אינטראקציות ספציפיות. היתרון העיקרי של שימוש בספריות ספריות מעל 3C ויטמינצ'יק לשלב העשרה הוא כי Hi-C, בניגוד 3C, כולל שלב העשרה למוצרים מקוריים מצדו. כתוצאה מכך, אחוז קריאות חוקיות בספריות PCHi-C הוא כ 10-fold גבוה יותר ב- C-לכידת ספריות50, אשר הכילה בסביבות 5%-8% חוקית קורא לאחר סינון HiCUP. . Sahlen et al. ישירות השוו לכידת-C כדי HiCap, אשר כמו PCHi-C משתמשת ויטמינצ'יק ספריות עבור לכידת העשרה, בניגוד לכידת-C העושה שימוש בספריות 3 ג. בקנה אחד עם הממצאים שלנו, הם מצאו כי לכידת-C ספריות מורכבים בעיקר קטעים משולחים שאינם מחוברים40. בנוסף, ספריות HiCap היה המורכבות גבוהה יותר מאשר ספריות לכידת-C40.
וריאציה של לכידת-C, שנקרא הדור הבא לכידת-C52 (NG לכידת-C) משתמשת oligo אחד בכל קצה קטע הגבלה, כמו שנוצרו קודם לכן ב- PCHi-C33,34, במקום חופפים הגששים בשימוש המקורי פרוטוקול לכידת-C50. פעולה זו מגדילה את אחוז חוקי קריאות בהשוואה לכידת-C בצניעות, אבל NG לכידת-C מעסיקה שני סיבובים רציפים של לכידת העשרה, מספר גבוה יחסית של ה-PCR מחזורי (20 עד 24 מחזורים בסך הכל, לעומת 11 מחזורים בדרך כלל עבור PCHi-C), אשר באופן בלתי נמנע תוצאות במספרים גבוהים יותר של רצף כפילויות והמורכבות ספריה נמוכה יותר. בניסויים הניסיון במהלך אופטימיזציה של PCHi-C, מצאנו כי אחוז ייחודי (קרי, אינם משוכפלים) לקרוא זוגות היה רק בסביבות 15% כאשר השתמשנו מחזורים PCR 19 (13 מחזורים מראש ללכוד + 6 מחזורים שלאחר ללכוד; נתוני לא מוצג), עם זאת אופטימיזציה למספר נמוך יותר של ה-PCR מחזורים, בדרך כלל מניבה 75-90% זוגות קריאה ייחודית. לפיכך, צמצום מספר מחזורים PCR באופן משמעותי מגדיל את כמות נתונים רצף אינפורמטיבי.
שיטה האחרונות משלב שבב עם ויטמינצ'יק להתמקד אינטראקציות כרומוזומלית מתווך על ידי חלבון ספציפי של עניין (HiChIP53). לעומת דשא54, אשר מבוסס על תירוץ דומה, HiChIP נתונים מכיל מספר גדול יותר של קריאות רצף מקיף, המאפשר אינטראקציה גבוהה יותר ביטחון עצמי מתקשר53. יהיה מאוד מעניין להשוות ישירות את HiChIP המתאים, ערכות נתונים ללכוד ויטמינצ'יק פעם הם הופכים לזמינים (לדוגמה HiChIP באמצעות נוגדן נגד Smc1a היחידה cohesin53 עם לכידת Hi-C עבור כל Smc1a מאוגדים הגבלה . שברי) בצד. הטבועה הבדל אחד בין שתי הגישות האלה היא ללכוד ויטמינצ'יק לא להסתמך על immunoprecipitation כרומטין, ולכן הוא מסוגל לחקור את האינטראקציות כרומוזומלית ללא התחשבות תפוסה של חלבון. פעולה זו מאפשרת השוואה של ארגון הגנום 3D ב נוכחות או היעדרות של גורם מסוים הכריכה כפי שימש לזיהוי PRC1 הרגולטור מפתח של העכבר ESC הגנום המרחבי אדריכלות7.
PCHi-C ו- GWAS
הגנום כולו האגודה מחקרים (GWAS) גילו כי גדול מ- 95% של מחלות הקשורות רצף גרסאות ממוקמים באזורים ללא קידוד של הגנום, לעיתים קרובות על מרחקים גדולים כדי חלבונים גנים55. GWAS משתנים לעיתים נמצא בסמיכות DNase אתרי רגיש, אשר מהווה סימן היכר של רצפי עם פוטנציאל לפעילות תקינה. PCHi-C ו- Hi-C ללכוד נעשה שימוש נרחב כדי לקשר היזמים GWAS הסיכון לוקוסים בהפרות סרטן השד44, סרטן המעי הגס48ומחלות אוטואימוניות-35,-45,-46. PCHi-C ללמוד על 17 תא שונים האנושי hematopoietic סוגי נמצאו SNPs הקשורים עם מחלות אוטואימוניות היו מועשר בחיישנים בתאים הלימפה, ואילו רצף משתנים הקשורים עם תכונות מיוחדות טסית דם, תא דם אדום נמצאו בעיקר מקרופאגים, erythroblasts, בהתאמה35,56. לפיכך, רקמות-סוג ספציפי יזם interactomes באמצעות PCHi-C עשוי לעזור להבין את התפקוד ללא קידוד הקשורים למחלה רצף משתנים ולזהות גנים המחלה פוטנציאליים חדשים התערבות טיפולית.
מאפייני האזורים אינטראקציה-יזם
מספר קווי ראיה לקשר מקדם interactomes בקרת ביטוי גנים. ראשית, מספר PCHi-C מחקרים הראו כי אזורים גנומית אינטראקציה עם היזמים של גנים (מאוד) ביטוי מועשרים בסימני המשויך משפר פעילות, כגון H3K27 acetylation p300 איגוד33,34 , 37. מצאנו מתאם חיובי בין רמת ביטוי הגן ואת המספר של משפרי אינטראקציה, רומז ההשפעות מוספים של משפרי תוצאה בביטוי הגנים מוגברת רמות34,35. שנית, שמן הביטוי אתרים (eQTLs) הם מועשרים בחיישנים המחוברים הגנים באותו ביטוי אשר מושפע eQTLs35. שלישית, על ידי שילוב טיול57 ונתונים PCHi-C, קיירנס. et al. מצא כי הטיול כתב גנים מיפוי חיישנים ב העכבר ESCs להראות כתב חזק יותר ביטוי גנים מאשר כתב גנים באתרים אינטגרציה באזורים הלא-יזם-אינטראקציה 58, המציין כי חיישנים בעלי פעילות רגולטורית תעתיק. יחד, ממצאים אלה מראים כי יזם interactomes שנחשפה על-ידי PCHi-C בהעכבר שונים, סוגי תאים אנושיים כולל מפתחות התקינה מודולים עבור בקרת ביטוי גנים.
. זה ראוי לציין כי מוצרי טיפוח טבעיים מייצגים רק חלק קטן (~ 20%) של חיישנים כל באמצעות PCHi-C33,34... . חיישנים אחרים יש תפקידים מבניים או טופולוגי ולא ישירה תעתיק פונקציות רגולטוריות. עם זאת, יש גם ראיות כי PCHi-C עלולה לחשוף את רכיבי ה-DNA עם פונקציית רגולציה לצבור קלאסית משפר סימני. בקו תאי הלימפה אדם, האמרגן BRD7 נמצאה אינטראקציה עם אזור ללא סימני שיפור שהוצג להחזיק משפר פעילות ב כתב ג'ין מבחני33. רכיבים תקינה עם מאפיינים דומים עשויים להיות שופע יותר מאשר כיום מוערך. לדוגמה, מסך מבוססי CRISPR לאלמנטים רגולטוריות DNA אלמנטים מזוהה לא מסומנים רגולטוריות (UREs) לשלוט ביטוי גנים, אבל הם חסרי משפר סימני59.
במקרים אחרים, חיישנים הוכחו הנמל כרומטין סימנים הקשורים עם דיכוי גנים ברמת השעתוק. חיישנים, שמעצבת היזמים מחויב על ידי PRC1 ב ESCs העכבר עסקו רשת מרחבית נרחבת של גנים מודחקים הנושאת שהדיכוי לסמן H3K27me37. בתאים lymphoblastoid האנושי, רכיב מרוחק אינטראקציה עם האמרגן BCL6 מודחקים transgene כתב ג'ין ביטוי33, רומז כי זה אולי מתפקד להדחיק שעתוק BCL6 בהקשרה מקורית.
. חיישנים מועשר עבור תפוסה של החלבון מבודד כרומטין CTCF של ESCs ו- NECs האנושי37 עשוי לייצג מחלקה נוספת של חיישנים באופן קולקטיבי, מראים תוצאות אלה חיישנים הנמל אוסף של פעילויות רגולטוריות ג'ין עדיין כדי להיות מאופיין באופן פונקציונלי.
Disclosures
לעורכים יש דבר לחשוף.
Acknowledgements
אנו מודים י ולריה Malysheva על קריאה ביקורתית של כתב היד, עזרה ממומחים עם איור 1. עבודה זו נתמכה על ידי המועצה למחקר רפואי, בריטניה (מר/L007150/1) ואת בריטניה ביוטכנולוגיה מחקר מדעי הביולוגיה המועצה, בריטניה (BB/J004480/1).
Materials
| Name | Company | Catalog Number | Comments |
| 16% (vol/vol) paraformaldehyde solution | Agar Scientific | R1026 | |
| Dulbecco's Modified Eagle Medium (DMEM) 1x | Life Technologies | 41965-039 | |
| Fetal bovine serum (FBS) sterile filtered | Sigma | F9665 | |
| Low-retention filter tips | Starlab | S1180-3810, S1180-1810, S1180-8810 and S1182-1830 | |
| 10x PBS pH 7.4 | Life Technologies | 70011-036 | |
| Molecular biology grade water | Sigma-Aldrich | W4502 | |
| 1 M Tris-HCl pH 8.0 | Life Technologies | 15568-025 | |
| IGEPAL CA-630 | Sigma-Aldrich | I8896 | |
| 5 M NaCl | Life Technologies | 24740-011 | |
| Protease inhibitor cocktail (EDTA-free) | Roche Diagnostics | 11873580001 | |
| Restriction buffer 2 (10x NEBuffer 2) | New England Biolabs | B7002 | |
| DNA LoBind tube, 1.5 mL | Eppendorf | 0030 108.051 | |
| DNA LoBind tube, 2 mL | Eppendorf | 30108078 | |
| 20% (wt/vol) SDS | Bio-Rad Laboratories | 161-0418 | |
| 20% (vol/vol) Triton X-100 | Sigma-Aldrich | T8787 | |
| HindIII, 100 U/uL | New England Biolabs | R0104 | |
| 10 mM dCTP | Life Technologies | 18253-013 | |
| 10 mM dGTP | Life Technologies | 18254-011 | |
| 10 mM dTTP | Life Technologies | 18255-018 | |
| 0.4 mM Biotin-14-dATP | Life Technologies | 19524-016 | |
| DNA polymerase I large (Klenow) fragment 5000 units/mL | New England Biolabs | M0210 | |
| 10x T4 DNA ligase reaction buffer | New England Biolabs | B0202 | |
| 100x 10mg/ml Bovine Serum Albumin | New England Biolabs | B9001 | |
| T4 DNA ligase, 1 U/μL | Invitrogen | 15224-025 | |
| RNase A | Roche | 10109142001 | |
| Proteinase K, recombinant, PCR grade | Roche | 3115836001 | |
| 20 000×g 50 ml centrifuge tube | VWR | 525-0156 | |
| 0.5 M EDTA pH 8.0 | Life Technologies | 15575-020 | |
| Phenol pH 8.0 | Sigma | P4557 | |
| Phenol: Chloroform: Isoamyl Alcohol 25:24:1 | Sigma | P3803 | |
| Sodium acetate pH 5.2 | Sigma | S7899 | |
| Quant-iT PicoGreen | Invitrogen | P7589 | |
| QIAquick Gel Extraction Kit | Qiagen | 28704 | |
| QIAquick PCR Purification Kit | Qiagen | 28104 | |
| Restriction buffer 2.1 (10x NEBuffer 2.1) | New England Biolabs | B7202 | |
| NheI, 100U/uL | New England Biolabs | R0131 | |
| Micro TUBE AFA Fiber Pre-slit snap cap 6x16mm vials | Covaris | 520045 | For sonication |
| SPRI beads (Agencourt AMPure XP) | Beckman Coulter | A63881 | |
| Dynabeads MyOne Streptavidin C1 beads | Invitrogen | 65001 | |
| Tween 20 | Sigma | P9416 | |
| 10 mM dATP | Life Technologies | 18252-015 | |
| T4 DNA polymerase 3000 units/mL | New England Biolabs | M0203 | |
| T4 PNK 10000 units/mL | New England Biolabs | M0201 | |
| Klenow exo minus 5000 units/mL | New England Biolabs | M0212 | |
| Quick ligation reaction buffer | New England Biolabs | B6058 | |
| NEB DNA Quick ligase | New England Biolabs | M2200 | |
| PE adapter 1.0 (5'-P-GATCGGAAGAGCGGTTCAGC AGGAATGCCGAG-3') | Illumina | ||
| PE adapter 2.0 (5'-ACACTCTTTCCCTACACGACGCT CTTCCGATCT-3') | Illumina | ||
| NEB Phusion PCR kit | New England Biolabs | M0530 | |
| PE PCR primer 1.0 (5'-AATGATACGGCGACCACCGA GATCTACACTCTTTCCCTAC ACGACGCTCTTCCGATCT-3') | Illumina | ||
| PE PCR primer 2.0 (5'-CAAGCAGAAGACGGCATACGA GATCGGTCTCGGCATTCCT GCTGAACCGCTCTTCCGATCT-3') | Illumina | ||
| PCR strips | Agilent Technologies | 410022 and 401425 | |
| SureSelect SSEL TE Reagent ILM PE full adaptor kit | Agilent Technologies | 931108 | |
| SureSelect custom 3-5.9 Mb library | Agilent Technologies | 5190-4831 | custom design mouse or human PCHi-C system |
| Dynabeads MyOne Streptavidin T1 beads | Invitrogen | 65601 | |
| E220 high-performance focused ultra-sonicator | Corvaris | E220 |
References
- Osborne, C. S., et al. Active genes dynamically colocalize to shared sites of ongoing transcription. Nature Genetics. 36, 1065-1071 (2004).
- Schoenfelder, S., et al. Preferential associations between co-regulated genes reveal a transcriptional interactome in erythroid cells. Nature Genetics. 42, 53-61 (2010).
- de Wit, E., et al. The pluripotent genome in three dimensions is shaped around pluripotency factors. Nature. 501, 227-231 (2013).
- Bantignies, F., et al. Polycomb-dependent regulatory contacts between distant Hox loci in Drosophila. Cell. 144, 214-226 (2011).
- Engreitz, J. M., et al. The Xist lncRNA exploits three-dimensional genome architecture to spread across the X chromosome. Science. 341, 1237973(2013).
- Denholtz, M., et al. Long-range chromatin contacts in embryonic stem cells reveal a role for pluripotency factors and polycomb proteins in genome organization. Cell Stem Cell. 13, 602-616 (2013).
- Schoenfelder, S., et al. Polycomb repressive complex PRC1 spatially constrains the mouse embryonic stem cell genome. Nature Genetics. 47, 1179-1186 (2015).
- Kundu, S., et al. Polycomb Repressive Complex 1 generates discrete compacted domains that change during differentiation. Molecular Cell. 65, 432-446 (2017).
- Skok, J. A., Gisler, R., Novatchkova, M., Farmer, D., de Laat, W., Busslinger, M. Reversible contraction by looping of the Tcra and Tcrb loci in rearranging thymocytes. Nature Immunology. 8, 378-387 (2007).
- Zhang, Y., et al. Spatial organization of the mouse genome and its role in recurrent chromosomal translocations. Cell. 148, 908-921 (2012).
- Aymard, F., et al. Genome-wide mapping of long-range contacts unveils clustering of DNA double-strand breaks at damaged active genes. Nature Structural & Molecular Biology. 24, 353-361 (2017).
- Ryba, T., et al. Evolutionarily conserved replication timing profiles predict long-range chromatin interactions and distinguish closely related cell types. Genome Research. 20, 761-770 (2010).
- Pope, B. D., et al. Topologically associating domains are stable units of replication-timing regulation. Nature. 515, 402-405 (2014).
- Chandra, T., et al. Global reorganization of the nuclear landscape in senescent cells. Cell Reports. 10, 471-483 (2015).
- Carter, D., Chakalova, L., Osborne, C. S., Dai, Y. F., Fraser, P. Long-range chromatin regulatory interactions in vivo. Nature Genetics. 32, 623-626 (2002).
- Tolhuis, B., Palstra, R. J., Splinter, E., Grosveld, F., de Laat, W. Looping and interaction between hypersensitive sites in the active beta-globin locus. Molecular Cell. 10, 1453-1465 (2002).
- Amano, T., Sagai, T., Tanabe, H., Mizushina, Y., Nakazawa, H., Shiroishi, T. Chromosomal dynamics at the Shh locus: limb bud-specific differential regulation of competence and active transcription. Developmental Cell. 16, 47-57 (2009).
- Zuniga, A., et al. Mouse limb deformity mutations disrupt a global control region within the large regulatory landscape required for Gremlin expression. Genes & Development. 18, 1553-1564 (2004).
- Sagai, T., Hosoya, M., Mizushina, Y., Tamura, M., Shiroishi, T. Elimination of a long-range cis-regulatory module causes complete loss of limb-specific Shh expression and truncation of the mouse limb. Development. 132, 797-803 (2005).
- D'Haene, B., et al. Disease-causing 7.4 kb cis-regulatory deletion disrupting conserved non-coding sequences and their interaction with the FOXL2 promotor: implications for mutation screening. PLoS Genet. 5, e1000522(2009).
- Sur, I. K., et al. Mice lacking a Myc enhancer that includes human SNP rs6983267 are resistant to intestinal tumors. Science. 338, 1360-1363 (2012).
- Herranz, D., et al. A NOTCH1-driven MYC enhancer promotes T cell development, transformation and acute lymphoblastic leukemia. Nature Medicine. 20, 1130-1137 (2014).
- Deng, W., et al. Controlling long-range genomic interactions at a native locus by targeted tethering of a looping factor. Cell. 149, 1233-1244 (2012).
- Groschel, S., et al. A single oncogenic enhancer rearrangement causes concomitant EVI1 and GATA2 deregulation in leukemia. Cell. 157, 369-381 (2014).
- Lupianez, D. G., et al. Disruptions of topological chromatin domains cause pathogenic rewiring of gene-enhancer interactions. Cell. 161, 1012-1025 (2015).
- Franke, M., et al. Formation of new chromatin domains determines pathogenicity of genomic duplications. Nature. 538, 265-269 (2016).
- Dekker, J., Rippe, K., Dekker, M., Kleckner, N. Capturing chromosome conformation. Science. 295, 1306-1311 (2002).
- Simonis, M., et al. Nuclear organization of active and inactive chromatin domains uncovered by chromosome conformation capture-on-chip (4C). Nature Genetics. 38, 1348-1354 (2006).
- Zhao, Z., et al. Circular chromosome conformation capture (4C) uncovers extensive networks of epigenetically regulated intra- and interchromosomal interactions. Nature Genetics. 38, 1341-1347 (2006).
- Dostie, J., et al. Chromosome Conformation Capture Carbon Copy (5C): A massively parallel solution for mapping interactions between genomic elements. Genome Research. 16, 1299-1309 (2006).
- Lieberman-Aiden, E., et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science. 326, 289-293 (2009).
- Belton, J. M., McCord, R. P., Gibcus, J. H., Naumova, N., Zhan, Y., Dekker, J. Hi-C: a comprehensive technique to capture the conformation of genomes. Methods. 58, 268-276 (2012).
- Mifsud, B., et al. Mapping long-range promoter contacts in human cells with high-resolution capture Hi-C. Nature Genetics. 47, 598-606 (2015).
- Schoenfelder, S., et al. The pluripotent regulatory circuitry connecting promoters to their long-range interacting elements. Genome Res. 25, 582-597 (2015).
- Javierre, B. M., et al. Lineage-specific genome architecture links enhancers and non-coding disease variants to target gene promoters. Cell. 167, 1369-1384 (2016).
- Wilson, N. K., et al. Integrated genome-scale analysis of the transcriptional regulatory landscape in a blood stem/progenitor cell model. Blood. 127, e12-e23 (2016).
- Freire-Pritchett, P., et al. Global reorganisation of cis-regulatory units upon lineage commitment of human embryonic stem cells. Elife. 6, (2017).
- Rubin, A. J., et al. Lineage-specific dynamic and pre-established enhancer-promoter contacts cooperate in terminal differentiation. Nature Genetics. 49, 1522-1528 (2017).
- Siersbaek, R., et al. Dynamic rewiring of promoter-anchored chromatin loops during adipocyte differentiation. Molecular Cell. 66, 420-435 (2017).
- Sahlen, P., et al. Genome-wide mapping of promoter-anchored interactions with close to single-enhancer resolution. Genome Biology. 16, 156(2015).
- Nagano, T., et al. Single-cell Hi-C reveals cell-to-cell variability in chromosome structure. Nature. 502, 59-64 (2013).
- Nagano, T., Varnai, C., Schoenfelder, S., Javierre, B. M., Wingett, S. W., Fraser, P. Comparison of Hi-C results using in-solution versus in-nucleus ligation. Genome Biology. 16, 175(2015).
- Wingett, S., et al. HiCUP: pipeline for mapping and processing Hi-C data. F1000 Res. 4, 1310(2015).
- Dryden, N. H., et al. Unbiased analysis of potential targets of breast cancer susceptibility loci by Capture Hi-C. Genome Research. 24, 1854-1868 (2014).
- Martin, P., et al. Capture Hi-C reveals novel candidate genes and complex long-range interactions with related autoimmune risk loci. Nature Communications. 6, 10069(2015).
- McGovern, A., et al. Capture Hi-C identifies a novel causal gene, IL20RA, in the pan-autoimmune genetic susceptibility region 6q23. Genome Biol.ogy. 17, 212(2016).
- Hodge, D., et al. A global role for EKLF in definitive and primitive erythropoiesis. Blood. 107, 3359-3370 (2006).
- Jager, R., et al. Capture Hi-C identifies the chromatin interactome of colorectal cancer risk loci. Nature Communications. 6, 6178(2015).
- Joshi, O., et al. Dynamic reorganization of extremely long-range promoter-promoter Interactions between two states of pluripotency. Cell Stem Cell. 17, 748-757 (2015).
- Hughes, J. R., et al. Analysis of hundreds of cis-regulatory landscapes at high resolution in a single, high-throughput experiment. Nature Genetics. 46, 205-212 (2014).
- Kolovos, P., et al. Targeted Chromatin Capture (T2C): A novel high-resolution high-throughput method to detect genomic interactions and regulatory elements. Epigenetics Chromatin. 7, 10(2014).
- Davies, J. O., et al. Multiplexed analysis of chromosome conformation at vastly improved sensitivity. Nature Methods. 13, 74-80 (2016).
- Mumbach, M. R., et al. HiChIP: efficient and sensitive analysis of protein-directed genome architecture. Nature Methods. 13, 919-922 (2016).
- Fullwood, M. J., et al. An oestrogen-receptor-alpha-bound human chromatin interactome. Nature. 462, 58-64 (2009).
- Maurano, M. T., et al. Systematic localization of common disease-associated variation in regulatory DNA. Science. 337, 1190-1195 (2012).
- Petersen, R., et al. Platelet function is modified by common sequence variation in megakaryocyte super enhancers. Nat. Commun. 8, 16058(2017).
- Akhtar, W., et al. Chromatin position effects assayed by thousands of reporters integrated in parallel. Cell. 154, 914-927 (2013).
- Cairns, J., et al. CHiCAGO: Robust detection of DNA looping interactions in Capture Hi-C data. Genome Biology. 17, 127(2016).
- Rajagopal, N., et al. High-throughput mapping of regulatory DNA. Nature Biotechnology. 34, 167-174 (2016).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved