
Method Article
启动子捕获高分辨率, 全基因组的启动子交互分析
* 这些作者具有相同的贡献
摘要
DNA 调控元素, 例如促进剂, 通过物理接触靶基因促进剂控制基因表达, 通常通过长距离染色体相互作用跨越大的基因组距离。启动子捕获高 c (PCHi) 确定促进者和远端区域之间的重要相互作用, 使潜在的调控序列分配给他们的目标基因。
摘要
基因组的三维组织与它的功能有关。例如, 转录增强剂等调控元素通过物理接触控制其目标基因的时空表达, 通常会在一定程度上弥合 (在某些情况下数以百计的 kilobases) 基因组距离和绕过附近的基因。人类基因组有大约100万促进剂, 其中绝大多数有未知的基因靶点。因此, 将远端调控区域分配给目标基因是理解基因表达控制的关键。我们开发了启动子捕获高 c (PCHi), 使全基因组的远端启动子相互作用区域 (PIRs) 的检测, 为所有发起人在一个单一的实验。在 PCHi 中, 高度复杂的高 C 库专为启动子序列, 通过在溶液混合选择与数以千计的生物素化 RNA 诱饵互补的所有启动子包含限制片段的两端特别丰富。其目的是, 然后拉下启动子序列和他们的频繁互动伙伴, 如增强剂和其他潜在的监管要素。在高通量配对结束排序后, 对每个启动子结扎的限制片段应用统计测试, 以确定限制片段级别上的重要 PIRs。我们使用 PCHi 生成了许多人和老鼠的细胞类型的远程启动子交互的图谱。这些启动子 interactome 映射通过将假定的调控区域分配给目标基因, 并揭示优先的空间启动子交互网络, 有助于更好地理解哺乳动物基因表达控制。这些信息也与了解人类遗传疾病和潜在疾病基因的识别有着很高的相关性, 通过将非编码疾病相关的序列变体或接近控制序列的基因链接到他们的目标。
引言
积累的证据表明, 基因组的三维组织在一系列核过程中扮演着重要的功能角色, 包括基因活化1、2、3、压制4 ,5,6,7,8, 重组9,10, dna 修复11, dna 复制12,13, 和细胞衰老14。遥远的促进剂被发现在接近的空间接近度对促进者他们调控15,16,17, 这是必要的适当的时空基因表达控制。增强剂的缺失表明, 远端促进因子是关键的目标基因转录18,19,20,21,22, 和 ' 强迫染色质循环 '表明, 在Hbb轨迹中的增强器与其目标启动子之间的设计, 足以驱动转录激活23。此外, 基因组重排在异位促进剂控制下的基因重组可能导致不适当的基因激活和疾病24,25,26。这些例子共同说明了促进剂的相互作用对基因控制至关重要, 需要严格的调控以确保适当的基因表达。人类和老鼠的基因组估计大约有100万促进剂。对于这些增强剂中的绝大多数, 目标基因是未知的, 并且促进者和促进剂之间的 "接触规则" 被理解得很差。因此, 将转录增强剂分配给其目标基因, 是破译哺乳动物基因表达控制的一个主要挑战。
我们对三维基因组体系结构的理解被引入 3C27 (染色体构象捕获) 和它的变种28,29,30,31的革命性改变。.最强大的这些技术, 高 C (高通量染色体构象捕获) 的设计, 以确定整个集合的染色体相互作用的细胞群内。高 C 库, 通常由数以百万计的细胞产生, 是高度复杂的, 大约 1011独立结扎产品之间的 4 kb 片段在人类基因组32。因此, 除非高 c 库接受超深测序, 否则, 从高 c 数据中对单个限制片段 (如包含启动器或增强器) 之间的相互作用进行可靠和可重现的识别是不可行的,这不是一个在经济上可行的解决方案, 为实验室编写的高 C 图书馆例行。为了规避这一缺陷, 我们开发了启动子捕获高 c, 专门从高 c 库中丰富的启动子包含结扎产品。我们专注于发起人有两个原因。首先, 促进剂的接触已被证明是关键的正确的基因表达水平在许多研究 (见上文参考), 第二, 由于推动者基本上不变的细胞类型, 同样的捕获诱饵系统可以用来审问跨多个单元类型和条件的调节电路。我们的方法依赖于高 c 库的解决方案杂交, 数以万计的生物素化 RNA 120mers 互补的促进剂包含的高 c 结扎产品和后续捕获链亲和素涂层磁性珠子。这将导致 PCHi 库的复杂性大大降低, 与原始的高 C 库相比, 它只集中于识别在显著高频率下被结扎到启动子的碎片。
我们已经使用 PCHi 在一些人和小鼠细胞类型, 以帮助更好地理解基因表达控制通过揭示远距离远端启动子互动区域的假定调节功能, 以及非随机启动子-启动子在原子核的三维空间中的接触。这些研究已经映射了成千上万的启动器-增强剂的联系横跨许多细胞类型33,34,35,36,37,38, 39, 确定了 Polycomb 压迫复合介导的小鼠胚胎干细胞的空间基因组组织7, 展示了大范围的启动子 interactomes 在细胞分化37,38,39, 并将非编码疾病相关序列变体与基因促进剂35联系起来。
PCHi 是一种理想的方法来映射基因组范围内的 DNA 序列与促进者互动的集合。相关的方法, 如捕获的连续基因组区域的高 C (参见讨论) 是选择的方法为选定的基因组区域的高度分辨率互作用概要。PCHi 和捕获的高 c 从实验的角度来看非常相似 (唯一的区别是捕获系统的选择), 因此我们提供的建议和指导方针都适用于这两种方法。在这里, 我们介绍了 PCHi 的详细描述。概述了 PCHi 实验的基本原理和设计, 提供了一步一步的 PCHi 库生成协议, 并说明了如何在协议的各个步骤中监视 PCHi 库的质量, 从而产生高质量的数据。
研究方案
1. 甲醛固定
-
细胞准备: 每项实验至少要从 2 x 107细胞开始。
- 对于在培养中生长的细胞, 并用重悬培养培养基中的细胞。对于前体细胞, 并用重悬在 1x Dulbecco 的改良鹰培养基 (DMEM), 补充 10% (卷/卷) 胎牛血清 (血清)。
- 对于黏附细胞, 去除培养基和添加30.625 毫升新鲜培养基与 10% (卷/卷) 的血清在室温下 (RT; 20–25°c)。
- 对于悬浮细胞, 收集和离心细胞在 400 x g 和20°c 3 分钟, 清除上清, 并重新悬浮细胞颗粒在30.625 毫升培养基与 10% (卷/卷) 血清在 RT。
- 对于固体组织, 使用胰蛋白酶 (0.05% 到2.5% 最终浓度, 取决于细胞类型) 或 dounce 均质, 以获得单个细胞悬浮。在这个额外的步骤后, 治疗细胞如悬浮细胞。
-
添加4.375 毫升16% 无甲醇多聚甲醛 (使用前打开安瓿) 到最后浓度 2% (卷/卷)。修正10分钟在 RT 与温和的混合在一个摇杆。
注意: 多聚甲醛是一种危险的化学物质。遵循适当的健康和安全规定。 - 通过加入5毫升新鲜制备的 1 M 冰冷甘氨酸, 淬火反应。混合5分钟, 在 RT 温和摇摆, 然后在冰上孵化15分钟, 偶尔反转。
-
清洗并收集固定的细胞。
- 对于黏附细胞, 去除上清, 添加10毫升冰冷 1x PBS pH 值7.4 在板墙上, 并删除它。添加1毫升冰冷 1x PBS pH 7.4, 收集细胞使用一个细胞刮刀和转移到50毫升管。重复两次以收集尽可能多的单元格。加入冰冷 PBS 多达50毫升最终体积。
- 对于悬浮细胞, 离心细胞在 760 x g 和4°c 为5分钟, 去除上清, 并在50毫升的冰冷 PBS pH 值7.4 中重新悬浮细胞颗粒。
- 离心细胞在 400 x g和4°c 为10分钟并且小心地去除上清。细胞颗粒可以在液氮中凝固, 随后储存在-80 摄氏度, 数月。
2. 细胞裂解
- 重新悬浮细胞颗粒在50毫升新鲜准备的冰冷裂解缓冲 (10 毫米三盐酸 pH 值 8, 0.2% (卷/卷) Igepal CA-630, 10 毫米氯化钠, 和一个片剂蛋白酶抑制剂鸡尾酒) 和混合。在冰上孵育30分钟, 偶尔通过反转混合。离心核在 760 x g和4°c 为5分钟和去除上清。
3.欣dIII 消化
- 用1.25x 限制缓冲器清洗细胞核2。在1毫升冰冷1.25x 限制缓冲器2中重新悬浮细胞颗粒, 并将其转移到1.5 毫升管中。旋转原子核在 760 x g 和4°c 为5分钟和去除上清。
- 在1790年µL 1.25x 限制缓冲器2中重新悬浮细胞颗粒。使5整除数, 每个包含5–10百万细胞在358µL 1.25x 限制缓冲2。
- 添加11µL 10% (重量/卷) SDS 每整除和动摇950转每分钟 (rpm) 30 分钟在37°c 在 thermomixer。如果细胞团簇出现, 由吹打离解, 避免气泡。
- 添加75µL 10% 海卫 X-100 (卷/卷) 每整除和动摇 950 rpm 和37°c 为15分钟在 thermomixer。如果细胞团簇出现, 由吹打离解, 避免气泡。
-
添加12µL 100 U/µL显dIII 100 (1200 单位总计) 每整除和孵化在37°c 隔夜 (O/N), 而颤抖在 950 rpm 在 thermomixer。
- 为消化控制, 转移25µL 样品 (5 µL 从每个整除) 在新的管子里加入酵素 (未消化的控制) 和重复同样做法在加入酵素以后 (消化的控制)。以与高 C 库相同的方式孵化两个管。
- 在第二天早晨, 增加5µL 100 U/µL欣dIII (总共500个单位) 每整除和孵化在37°c 为 2 h, 而颤抖在 950 rpm 在 thermomixer。
-
消化控制: 用于消化和未消化的控制 (见 3.5.1), 执行交联逆转 (步骤 6), 苯酚: 氯仿萃取, 和 DNA 沉淀 (步骤 7)。
- 设计一副引物横跨欣dIII 网站。在同一地区, 设计另一对引物, 不跨越一个欣dIII 网站。用 Primer3 (http://bioinfo.ut.ee/primer3-0.4.0/) 和以下参数进行定量 pcr (Q pcr) 的设计底漆:
底漆尺寸: 最佳 20 (最小:18, 最大值:27);底漆 Tm: 最佳 60 (最小:57, 最大值:63);底漆 CG% 含量: 最小:20, 最大值:80;扩增子尺寸: rt-pcr ~ 100 bp (常规 pcr ~ 300 bp);Mispriming 图书馆: 人类 (人类引物) 或啮齿动物和简单 (鼠标引物)。 - 执行 Q PCR 获得4平均 Cts (阈值周期): Ct [D;h], 从被消化的样品 [D] 得到与横跨欣dIII 站点的一对引物 [H];Ct [d;], 从被消化的样品 [d] 获得与不横跨欣dIII 站点的对引物 [-];Ct [U;H], 从未消化的样品 [U] 与横跨一个欣dIII 站点的一对引物获得;Ct [u;], 从未消化的样品 [u] 与不横跨显dIII 站点的一对引物 (-) 获得。计算消化率为:% 消化 = 100-100/2(ct [d, h]-ct [d,-])-(ct [u, h]-ct [u,-])。
- 设计一副引物横跨欣dIII 网站。在同一地区, 设计另一对引物, 不跨越一个欣dIII 网站。用 Primer3 (http://bioinfo.ut.ee/primer3-0.4.0/) 和以下参数进行定量 pcr (Q pcr) 的设计底漆:
4. 限制片段的 Biotinylation
- 准备 biotinylation 主组合: 30.6 µL 10x 限制缓冲器 2, 10.2 µL H2O (分子生物学级), 7.65 µL 10 毫米 dCTP, 7.65 毫米 dGTP, 10 µL 7.65 mm dTTP, 10 µL 191.25 毫米 biotin-14-dATP, 0.4 µL 51 U/毫升 DNA 聚合酶 I 大 (克莱诺) 片段。
- 每三十年代增加60µL 的 biotinylation 总混合, 每整除混合, 并孵化37摄氏度1小时, 在 700 rpm (thermomixer), 在 5 s, 每年的 h。1小时后, 将整除数放在冰上。
5. 髓内结扎术
- 准备结扎大师组合: 510 µL 10x T4 dna 连接酶缓冲器, 51 µL 10 毫克/毫升牛血清白蛋白 (100x BSA), 1754.4 µL 水 (分子生物学等级), 127.5 µL 1 U/µL T4 DNA 连接酶 (见材料表)。
- 添加479µL 的结扎主混合每整除混合和孵化在16°c 4 小时晃动 700 rpm 5 s 每2分钟在 thermomixer。
- 在 RT 孵化30分钟。
6. 交联反转
- 将所有整除数在50毫升离心管中组合 (适用于高速离心)。
- 添加62.5 µL 10 毫克/毫升 RNase A, 混合, 孵化30分钟在37摄氏度。
- 添加300µL 10 毫克/毫升蛋白酶 K, 混合, 孵化30分钟在37摄氏度。
- 孵育反应 O/N (或至少4小时) 在65摄氏度。第二天早上, 添加300µL 10 毫克/毫升蛋白酶 K, 混合, 并孵化1小时在65摄氏度。
7. DNA 纯化
- 添加4337.5 µL 缓冲 (10 毫米三盐酸 ph 8.0; 0.1 毫米 EDTA ph 值 8.0) 和混合。
- 添加1体积 (10 毫升) 苯酚 pH 8.0, 涡流为十年代, 和离心机在 RT 和 2万 x g为3分钟. 将上 (水) 相的9毫升转移到新的50毫升管。
注意: 苯酚是一种危险的化学物质。遵循适当的健康和安全规定。 - 将2毫升的缓冲液添加到剩余的水相中, 涡流为十年代, 在 RT 和 2万 x g的离心机上3分钟. 将水相的2.5 毫升从步骤7.2 转移到新管中, 使最终体积11.5 毫升。丢弃含有较低 (有机) 相的管。
- 增加1容量 (11.5 毫升) 的 phenol:chloroform:isoamyl 酒精 (25:24:1), 漩涡十年代, 和离心机在 RT 和 2万 x g为3分钟. 将上 (水) 相的11毫升转移到新的50毫升管。重复步骤7.3。总样本量现在是13.5 毫升。
- 加入1.35 毫升3米醋酸钠 pH 5.2 和33.75 毫升冰冷100% 乙醇, 混合, 并孵化在-80 °c 45 分钟, 或交替隔夜在-20 °c。
- 离心机在4°c 和 2万 x g为10分钟, 去除上清, 再悬浮颗粒在1毫升新准备的 70% (卷/卷) 乙醇, 并且转移到新的管子。
- 离心机在4°c 和全速为3分钟在台式离心机, 然后去除上清。
- 在1毫升冰冷 70% (卷/卷) 乙醇中重新悬浮颗粒, 重复步骤7.7。将颗粒干燥37°c 10 分钟, 并在650µL 的缓冲液中重新悬浮。用荧光法测定双链 dna 的 dna 产量。
注意: 该协议可以暂停冻结, 并存储样品在-80 °c 几个月或在-20 °c 较短的时间内。
8. 质量控制
- 监控图书馆的完整性, 并通过 DNA 电泳进行结扎。在 0.8% agarose/1x 的凝胶上运行200吴图书馆。DNA 应该以 10 kb 的波段运行。
- 通过常规 PCR 检测已知的细胞型不变短程和远距离相互作用。每个 PCR 反应使用 100 ng 模板 DNA。按照上面的说明, 设计 PCR 引物接近并朝向限制点 (见 3.7.1)。表 1列出了小鼠和人体高 C 库质量控制的底漆序列。
-
填充和结扎控制: 切出含有 amplicons 的凝胶带从控制 8.2, 凝胶提取 dna, 并使用 dna 作为模板, 4 个单独的 PCR 反应与相同的底漆组合。
- 用 PCR 纯化试剂盒净化 amplicons, 并量化 DNA 浓度。
- 准备四消化反应 (欣dIII [a], NheI [b],显dIII + NheI [c] 和没有酶 [d]) 为每个扩增子在最后的容量15µL: 500 ng 扩增子, 1.5 µL 10x 限制缓冲 2.1, 0.15 µL 10 毫克/毫升牛血清白蛋白 (100x BSA)和0.1 µL (10 单位) 的酶 (欣dIII [a], NheI [b],显dIII + NheI [c] 或水 [d])。
- 消化1小时在37°c, 然后跑消化反应在 1.5% (agarose/1x) 凝胶。
9. DNA 碎片
- 将50.5 µg 的样品转移到一个新的管子中, 并将其缓冲到最后的655µL. 将样本分成5个超声波瓶 (见材料表), 将130µL 的库 (10 µg) 添加到每个小瓶中。在超 sonicator (见材料表) 中, 用以下参数剪切至 400 bp 的尺寸: 工作因数: 10%;峰值入射功率 (w): 140;每爆裂周期: 200;时间: 五十五年代。
- 收集微气泡样品在一个新鲜的2毫升管。
10. 双面 SPRI-珠粒尺寸选择
- 混合 SPRI (固相可逆固定化) 珠解决方案井通过反转, 转移1.85 毫升的珠溶液到一个新的管, 并带来 RT 15 分钟。
- 添加350µL 的水 (分子生物学级) 的样品 (最终体积1毫升)。
- 在样品中加入600µL SPRI 珠溶液 (总容积1.6 毫升; SPRI 溶液与 DNA 的比值: 0.6 到 1), 在 RT 上孵化5分钟, 并在台式离心机中自旋取样以 2–3 s 收集样品。
- 打开盖子, 将样品放在磁分离架上5分钟, 将清清液转移到新管中, 丢弃珠子。
- 将 SPRI 珠浓缩为第二尺寸选择步骤: 将 SPRI 珠的930µL 转移到新管中, 在磁分离架上放置5分钟, 并丢弃清上清。在310µL SPRI 珠溶液中重新悬浮珠子。
- 添加300µL 的浓缩 SPRI 珠 (步骤 10.5) 到样品 (总容量1.9 毫升; SPRI 溶液的 DNA 现在是0.9 到 1), 在 RT 上孵化5分钟, 并在台式离心机中的旋转样本 2–3 s. 小心打开盖子, 将管放在磁分离架上5分钟, 并丢弃上清。
- 将1毫升新鲜制备的70% 乙醇 (卷/卷) 添加到磁分离架上的取样管上, 在三十年代孵化, 并丢弃上清。重复两次。
- 干燥珠在37°c 在一个 thermomixer (管盖子打开) 不超过5分钟, 增加300µL 的样本, 混合和孵化10分钟室温。
- 旋转样品在台式离心机为 2–3 s, 打开盖子并且放置管子在磁性分离立场为5分钟. 将清上清液转移到新管中并丢弃珠子。
11. 结扎产品的生物素/链亲和素拉下
- 准备缓冲器: 1x TB 缓冲器 (5mM 三盐酸 pH 值 8.0; 0.5mM EDTA; 1 米氯化钠; 0.05% 吐温 20);2x 非关税壁垒缓冲 (10 毫米三盐酸 pH 值 8.0; 1 毫米 EDTA; 2 米氯化钠);1x 非关税壁垒缓冲 (5 毫米三盐酸 pH 值 8.0; 0.5 毫米 EDTA; 1 米氯化钠)。
- 添加200µL 磁性链亲和素耦合珠 (见材料表) 进入一个新的管, 把它放在磁分离立场1分钟, 并删除上清。
-
用500µL 的 1x TB 缓冲器清洗珠子两次。
- 对于在生物素下拉下的每一个洗涤步骤, 在非结扎 DNA 末端、dATP 尾矿和适配器结扎步骤中, 停止对生物素的修复和去除, 在相应的缓冲器中重新悬浮珠子, 在 RT 旋转 15 rpm 3 分钟, 在台式离心机中旋转管对于 2–3 s, 将管放在磁分离架上3分钟, 取出上清。
- 在300µL 的2x 非关税壁垒缓冲中重新悬浮珠子。混合珠和样品 (600 µL 总体积) 和孵化在 RT 15 分钟的旋转轮在 3 rpm。
- 在磁分离架上回收珠子3分钟, 取出清清液。在500µL 的200µL 1x 结扎缓冲液中, 先洗两次, 再用1x 的无关税壁垒缓冲。在50µL 的10x 结扎缓冲器中重新悬浮珠子。
12. 在非结扎 DNA 末端修复和去除生物素
- 结合样品 (总共50µL) 与50µL 2.5 毫米 dNTP 混合 (12.5 µL 10 毫米每 dNTP), 18.1 µL 3000 u/毫升 T4 dna 聚合酶, 18.1 µL 1万 U/毫升 T4 PNK, 3.7 µL 5000 u/毫升 DNA 聚合酶 I 大 (克莱诺) 片段, 360.1 µL 的 H2O。
- 混合和孵化在20°c 1 小时, 摇晃 5 s 在 700 rpm 每2分钟在一个 thermomixer。
- 在磁分离架上回收珠子, 清除清上清, 并在500µL 1x TB 缓冲液中清洗两次珠子。
- 在500µL 的1x 的关税壁垒中洗涤珠子, 然后在500µL 的1x 里洗一遍。
- 在磁选架上回收珠子, 清除清上清液, 再将珠在415µL 的1x 缓冲器中重新悬浮。
13. dATP 尾矿
- 结合样品 (415 µL) 与50µL 10x 限制缓冲 2, 5 µL 10 毫米 dATP, 30 µL 5 U/µL 克莱诺外减。
- 混合和孵化在37°c 30 分钟, 摇晃 5 s 在 700 rpm 每2分钟在一个 thermomixer。
- 在磁分离架上回收珠子, 清除清上清, 并在500µL 1x TB 缓冲液中清洗两次珠子。
- 在500µL 1x 的关税壁垒中洗涤珠子。
14. 适配器结扎
- 在200µL 的1x 结扎反应缓冲器中洗涤珠子 (见材料表)。
- 重新悬浮珠在200µL 的1x 结扎反应缓冲。添加4µL 的 DNA 连接酶 (见材料表) 和16µL 15 µM 预退火 pe 适配器 (预退火 pe 适配器1和 pe 适配器 2 (在30µM) 和孵化几分钟在 RT)。在 RT 孵育15分钟。
- 在磁分离架上回收珠子, 清除清上清, 并在500µL 1x TB 缓冲液中清洗两次珠子。
- 在500µL 1x 的关税壁垒中洗涤珠子。然后, 在100µL 的1x 限制缓冲器2中清洗珠子, 在50µL 的1x 限制缓冲器2中重新悬浮珠子, 然后转入新管。
15. 高 C 库放大
- 准备 PCR 大师组合: 100 µL 5x Phusion 缓冲器;6µL 25 µM PE PCR 底漆 1.0;6µL 25 µM PE PCR 底漆 2.0;14µL dNTP 混合 (每) 10 毫米;6µL Phusion 聚合酶;318µL 的 H2O。
- 混合 pcr 大师组合与珠子 (总共500µL), 除以10整除数50µL, 并通过 PCR 放大使用以下条件:
30s 摄氏98摄氏度
7周期: 十年代在98°c;三十年代在65°c;三十年代在72°c
7分钟 at 72 °c - 将 PCR 反应收集到新管中, 在磁分离架上回收珠子, 将上清 (500 µL) 转化为新管。
-
用 SPRI 珠净化图书馆 DNA。
- 混合 SPRI 珠, 转移460µL 的珠子在一个新的管, 并带来 RT 15 分钟. 添加450µL SPRI 珠的 PCR 反应 (最终体积950µL), 孵化5分钟在 RT, 和旋转样品在台式离心机, 以收集样品。
- 打开盖子, 将样品放在磁分离架上5分钟, 取出上清。
- 将珠子保持在磁分离架上, 加入1毫升的70% 乙醇 (卷/卷), 在一个区域内的样品管上清除珠子, 离开三十年代, 并放弃上清。
- 重复步骤15.4.3 两次。
- 干珠在37°c 在一个 thermomixer (管盖子打开) 不超过5分钟。
- 增加51µL 的样本, 混合, 孵化10分钟37摄氏度, 颤抖在 950 rpm 在一个 thermomixer。
- 旋转样品在台式离心机为 2–3 s, 打开盖子并且放置管子在磁性分离立场为5分钟. 将清上清液转移到新管中并丢弃珠子。
- 量化高 C 库的集中度。经过7轮的 PCR 扩增, 我们经常获得500–1,500 的高 C 库。
16. 混合式解决方案捕获
注意: 下面的拦截器和缓冲 (SHS1-4) 解决方案来自 SureSelect 套件 (见材料表)。
- 将 500 ng 到1µg 的高 C 库转换成新管, 在真空集中器上蒸发样品 (见材料表; 45 °c; 真空压力: 30.0 级, 斜坡 5) 直到干燥。
- 通过添加3.6 µL2O (分子生物学级)、2.5 µL 阻滞剂1、2.5 µL 阻滞剂2和0.6 µL 自定义阻滞剂, 重新悬浮蒸发的高 C 库。
- 将样品转移到一个新的 pcr 管条井中, 与 pcr 盖条和冰上的地方紧密结合。标签为 "D" (为高 C 脱氧核糖核酸)。
- 制备杂交缓冲器: 12.5 µL SHS1 缓冲液;0.5 µL SHS2 缓冲器;5µL SHS3 缓冲器;6.5 µL SHS4 缓冲器。
- 在 thermomixer 中孵化65摄氏度, 5 分钟。转移到一个新的 pcr 管条井, 关闭与 pcr 帽条, 并保持在 RT. 标签为 "H" (用于杂交缓冲)。
- 入一个新的 PCR 管条的井, 混合5µL 100 ng/µL 生物素化 RNA 探针 (存放在-80 °c 和解冻在冰在使用之前);0.5 µL SRNase B (RNase 抑制剂) 和1.5 µL H2O (分子生物学级)。
- 用 pcr 盖条和冰上的地方关闭 pcr 管带。标签为 "R" (用于 RNA)。
- 使用以下参数设置 PCR 机:
5分钟在95°c;25 h 在65°c;盖加热;29µL PCR 反应量。
注意: 在所有过程中尽可能快地进行, 而 PCR 机运行, 以避免样品蒸发。 - 将 "D" pcr 管带放入 pcr 机中, 关闭 pcr 机盖, 开始 pcr 反应。pcr 程序达到65摄氏度时, 打开 pcr 机盖, 将 "H" pcr 管带放入 pcr 机中。关闭 pcr 机盖, 孵育3分钟. 打开 pcr 机盖, 将 "R" pcr 管带放在 pcr 机上, 并关闭 pcr 机。
- 2分钟后, 打开 pcr 机盖和所有 pcr 管条。转移13µL 井 "H" 入井 "r", 然后所有容量井 "D" 入井 "r"。吸管上下3次混合反应, 关闭 pcr 管条, 取出 "H" 和 "D" pcr 管条, 并关闭 pcr 机盖。孵育反应在65°c 为24小时。
17. 分离启动子片段含结扎产品
注意: 建议使用 SureSelect 适配器工具包和库执行以下步骤 (请参见原材料表)。
- 预热1.5 毫升的洗涤缓冲器2每样品在65°c 之前。
- 添加60µL 的链亲和素耦合磁珠 (见材料表) 进入一个新的管, 放置在磁分离立场1分钟, 并删除上清。
- 用200µL 1x 绑定缓冲器清洗珠子三次。
注: 对于每一个洗涤步骤在捕获后的启动器包含结扎产品, 重新悬浮珠在相应的缓冲, 旋转3分钟的 RT 和 15 rpm 的旋转轮, 软旋转管在一个台式离心机 2–3 s 收集样品, 将管放在磁分离架上3分钟, 取出上清。 - 在200µL 的1x 绑定缓冲区中重新挂起珠子。打开 pcr 机和 pcr 管条 (pcr 程序仍在运行), 用磁性珠子将杂交反应转移到管内。在 3 rpm 的旋转轮上孵育30分钟。
- 在磁选架上回收珠子, 清除清液。将珠子重新悬浮到500µL 的洗涤缓冲器 1, 混合, 并孵化15分钟20°c, 而颤抖在 950 rpm 在一个 thermomixer。
- 在磁分离架上回收珠子3分钟, 取出清清液。将珠子重新悬浮到500µL 的洗涤缓冲器 2, 混合和孵化10分钟65°c, 而颤抖在 950 rpm 在一个 thermomixer。重复步骤17.5 两次。
- 在磁选架上回收珠子, 清除清清液, 在200µL 1x 限制缓冲器2中重新悬浮珠。在磁选架上回收珠子, 去除上清, 再将珠子重新悬浮成30µL 1x 限制缓冲器2。
18. PCHi 图书馆扩增
- 准备 PCR 大师组合:60 µL 5x pcr 缓冲器 (Phusion 缓冲器), 3.6 µL 25 µM pe pcr 底漆 1.0, 3.6 µL 25 µM pe pcr 底漆 2.0, 8.4 µL 的 dNTP 混合 (10 毫米每), 3.6 µL 的 Phusion 聚合酶, 190.8 µL 的 H2O。
- 混合 pcr 大师组合与珠子 (总共300µL), 除以6整除数50µL 和 PCR 放大使用以下条件:
三十年代在98°c
4周期: 十年代在98°c, 三十年代在65°c, 三十年代在72°c
7分钟 at 72 °c - 收集所有 PCR 反应在一个新的管, 回收磁铁上的珠子, 转移上清 (300 µL; 包含 PCHi 库) 在一个新的管。
- 使用 SPRI 珠净化 PCHi 库, 按照上面描述的步骤15.4。
- 量化 PCHi 库的浓度。
结果
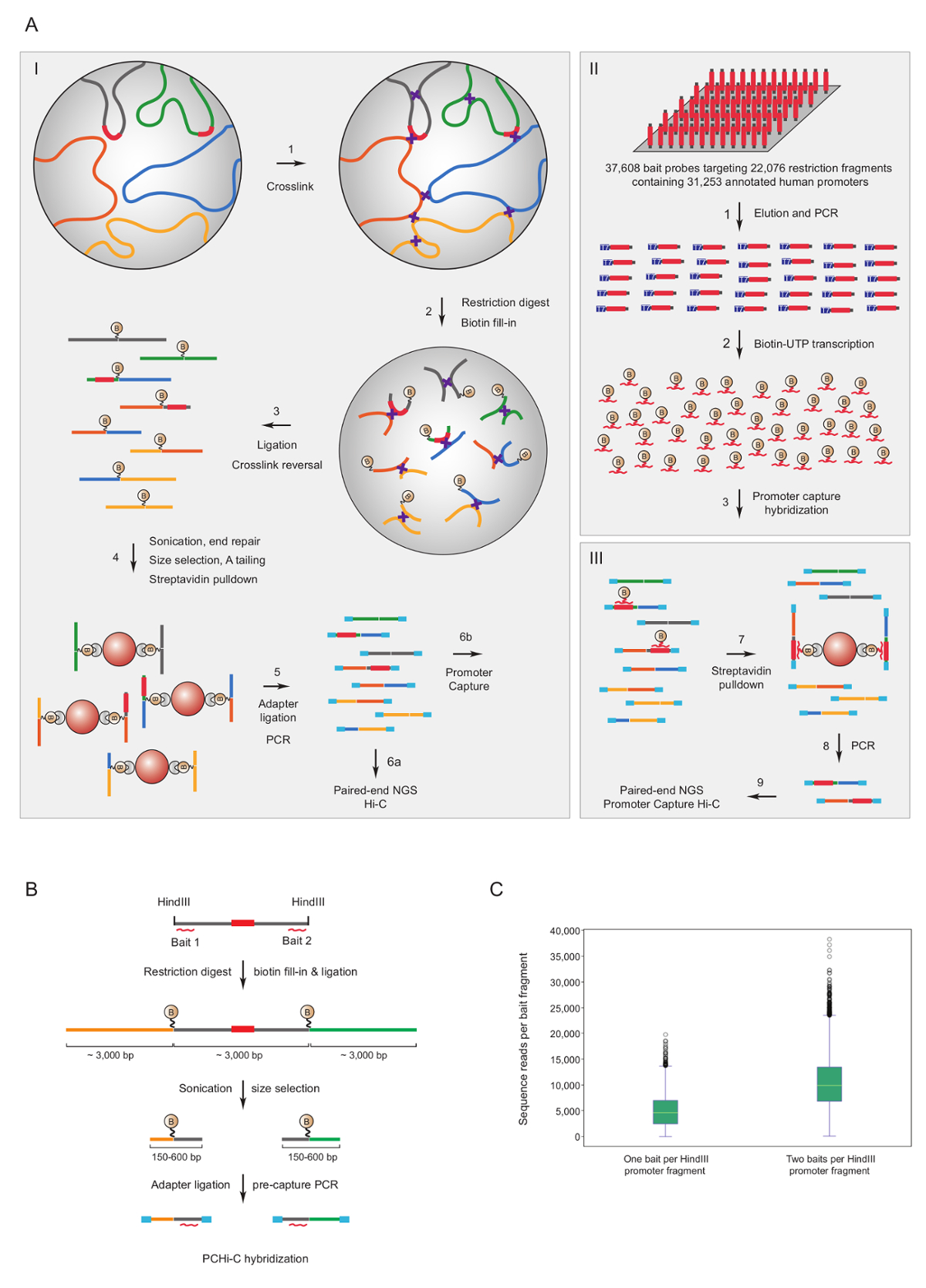
启动子捕获高 c 已被用来丰富的鼠标7,34,36,39和人类33, 35,37,38高 c 库启动子交互。一个类似的协议 (命名 HiCap) 已经描述了桑德伯格集团40。图 1A显示了启动子捕获高 C 的示意图工作流。在这里描述的协议中, 高 C 库是使用核结扎41产生的, 这导致了大量的假结扎产品的数量减少42。对于 PCHi, 高度复杂的小鼠或人的高 C 库在溶液中进行杂交和捕获, 使用39021生物素化 rna 补充22225个小鼠启动器-包含 HindIII 限制片段, 或37608生物素化 rna针对22076人启动子, 分别包含 HindIII 限制片段。包含限制片段的启动子可以以单个生物素化 rna (图 1B) 或两端为目标。我们发现两端的捕获改善了单个启动子的覆盖率 (图 1C; 原始序列读取) 几乎两倍, 如预期。因此, 只要有可能 (即在非重复区域), 我们建议使用生物素化 rna 互补的限制片段的两端被捕获。
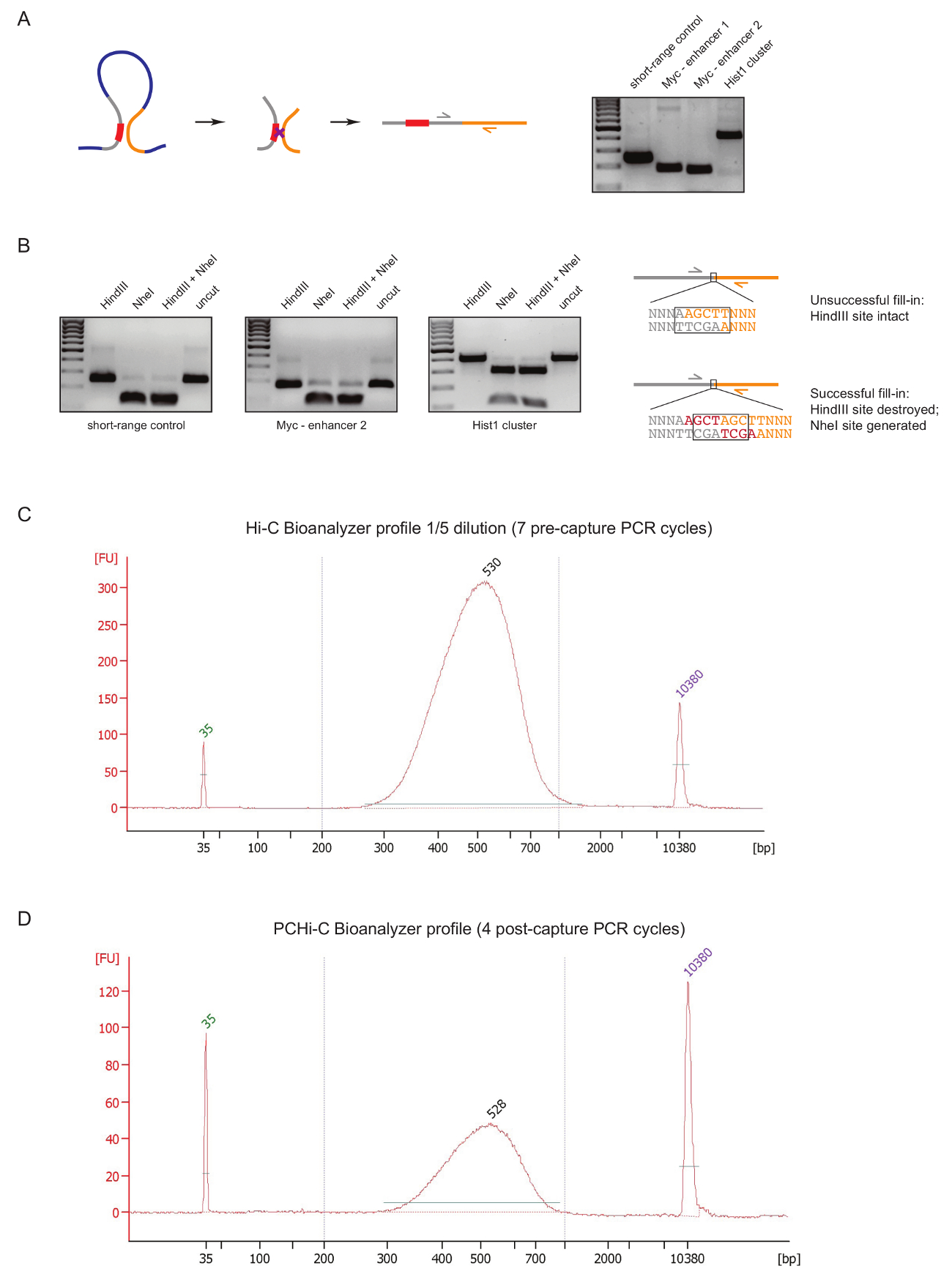
为评估图书馆准备工作初期 PCHi 图书馆的质量, 我们在 DNA 结扎和纯化后进行两种控制, 如前所述31。第一个是使用特定的底漆对放大结扎产品, 如在 3C27。我们使用底漆对 (表 1) 放大细胞类型不变的长距离结扎产品, 如c-myc基因和它的已知增强剂位于大约 2 Mb 以外 (图 2A) 或在Hist1轨迹的基因之间 (分离 1.5 Mb), 并在两个区域之间的接近线性接近 (' 短程控制 ')。
第二个质量控制是为了确定生物素在克莱诺介导的填充-dATP 的限制部位充填的效率。成功的克莱诺填充和随后的钝端结扎结果在原来的限制站点之间的结扎产品的 DNA 分子之间的消失, 并在 HindIII 的情况下形成一个新的 NheI 识别站点 (图 2B).HindIII 与 NheI 消化结扎产物的比值是生物素结合效率的直接读数。低质量的高 C 库将显示高水平的 HindIII 消化, 而高质量的图书馆有接近完全 NheI 消化的结扎产品 (图 2B)。
在高 c 库的准备 (即生物素-链亲和素拉下的大小选择的高 c 结扎产品, 适配器结扎和预捕获 PCR) 后, 高 c 库的完整性和大小分布由 Bioanalyzer 评估 (图2C). 同样的控制是在 PCHi 图书馆的准备结束时进行的 (即,在杂交捕获了启动子的结扎产品和后捕获 PCR 后)。比较了高 c 和 PCHi Bioanalyzer 的配置文件表明, 高 c 库比相应的 PCHi 库集中得多, 但库的大小分布非常相似, 表明捕获步骤在PCHi 不引入尺寸偏差 (图 2C, D)。
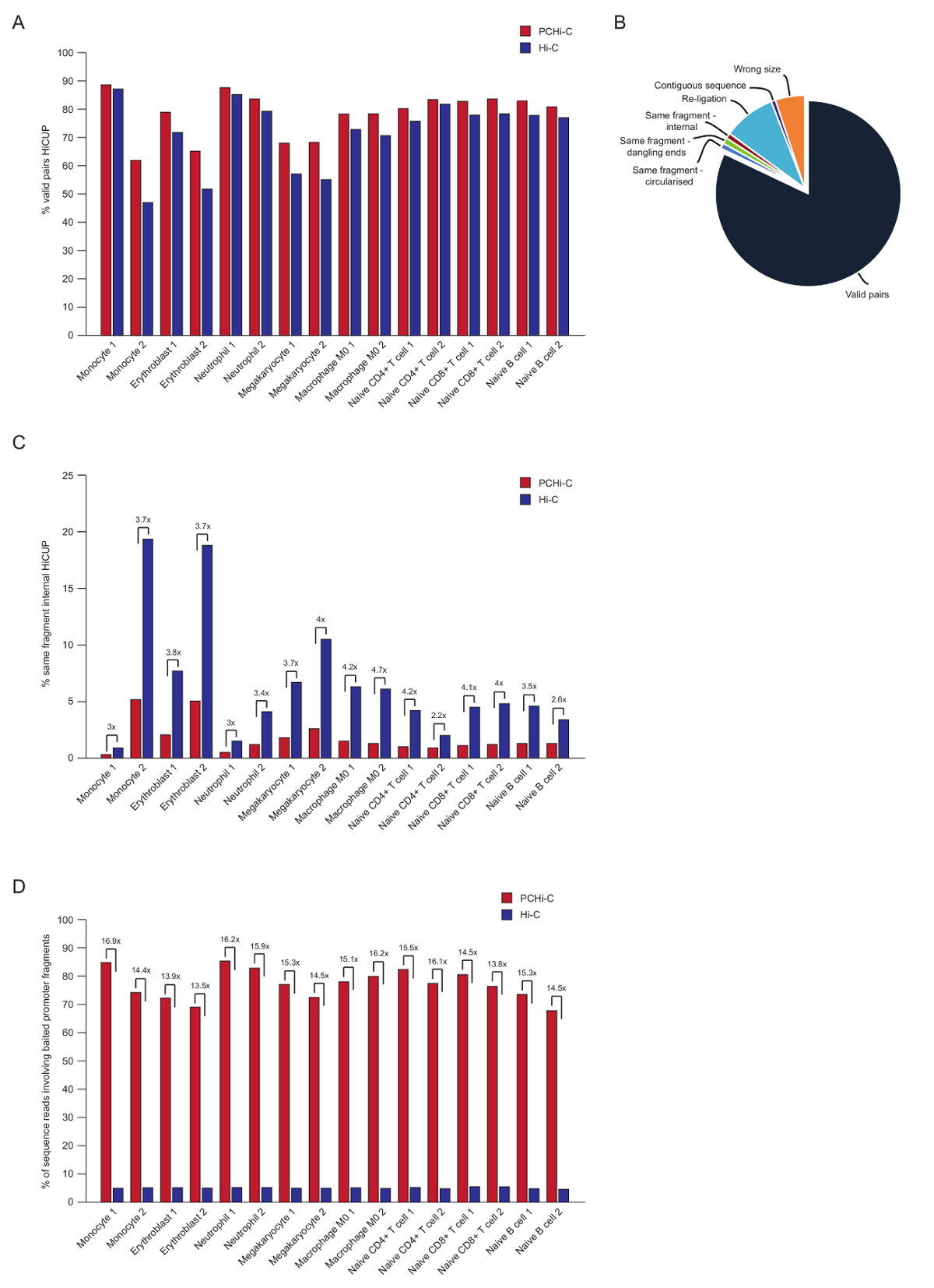
配对结束排序后, PCHi 读取被映射, 质量控制和过滤使用 HiCUP 管道43。高质量的 PCHi 库包含70-90% 个 "有效对" (即,在线性基因组映射上不相邻的两个限制片段之间的配对端序列读取;图 3A, B)。使用核内结扎协议41,42,反读对的百分比 (即,配对结束序列读取在两个限制片段之间位于不同的染色体) 通常是低的,5至 25%, 反映了染色体界的存在, 表明了良好的图书馆质量。对高 c 库与相应的 PCHi 库之间 "有效对" 百分比的直接比较35, 表明在所有情况下, PCHi 库中有效对的百分比都较高 (图 3B)。这伴随着 PCHi C 中非有效 "相同片段内部" 读取的百分比减少 (图 3C)。这是预期的, 因为捕获步骤不仅丰富了启动子包含结扎产品, 但也为限制片段结束, 由于捕获寡核苷酸在限制片段的位置 (见图 1B)。
HiCUP 滤波后, 确定捕获效率。在 HiCUP 筛选后, PCHi 库包含三种有效序列读取的类型:
1.) 启动子: 基因组读取 (即在基因组中任何地方的捕获启动子片段和非启动子 HindIII 限制片段之间读取)
2.) 促进者: 促进者读 (读在二被捉住的促进者片断之间)
3. 基因组: 基因组阅读 (背景的高 C 结扎产品, 没有结扎产品的合作伙伴映射到捕获的启动子)。这些都是在下游分析之前丢弃的。
高质量的 PCHi 库具有捕获效率 (上面的类别1和2的总和) 之间的 65–90% (图 3D)。与高 C 库的直接比较表明, PCHi-c 对含启动子的结扎产品有15倍的富集效果 (图 3D), 在某些情况下是17倍。这接近假设的最大 (19.6 倍) 富集为 PCHi, 这取决于基因组限制片段的百分比捕获系统。通过设计针对较少限制片段44、45、46的捕获系统, 可以实现更大的浓缩。
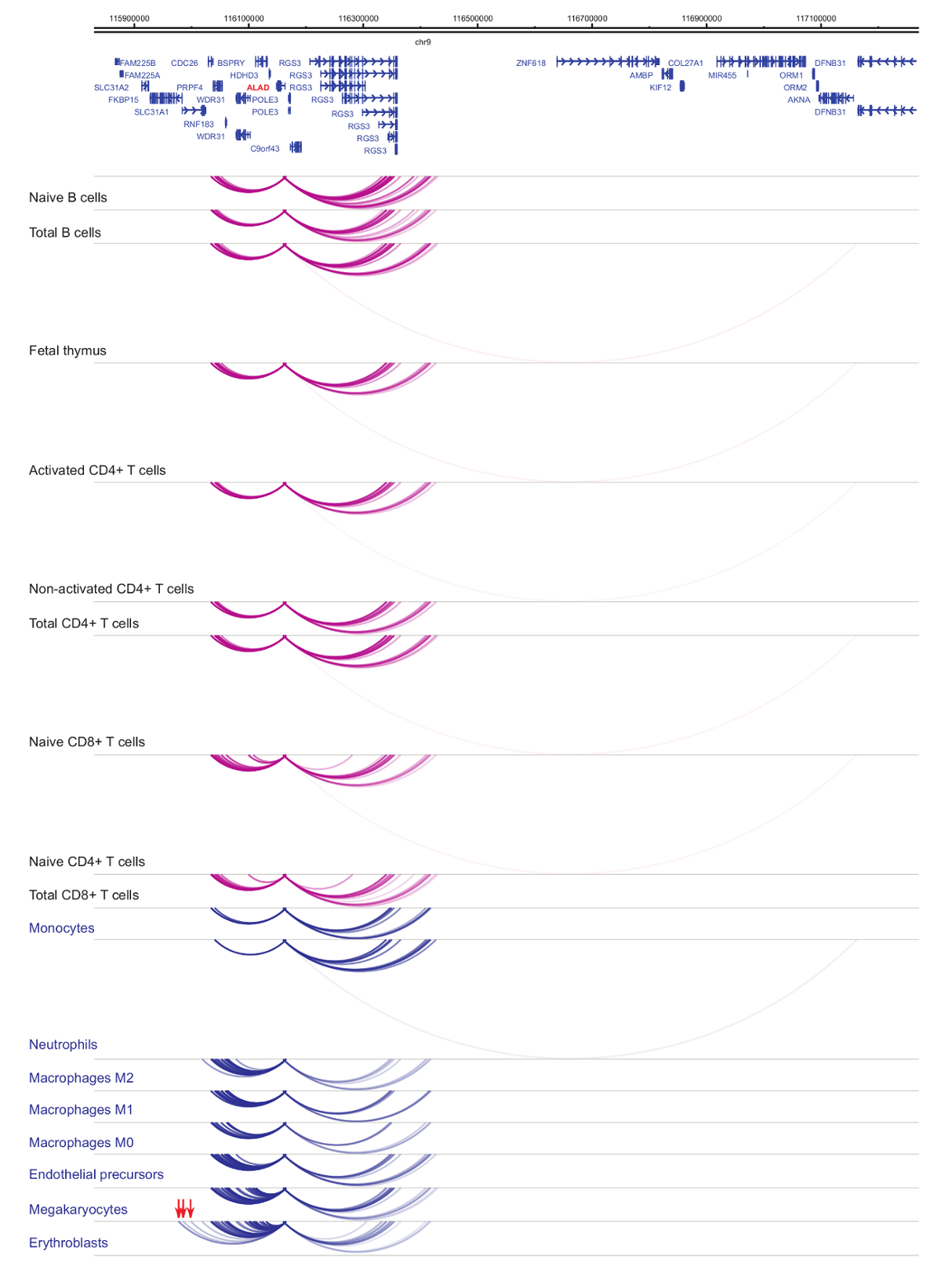
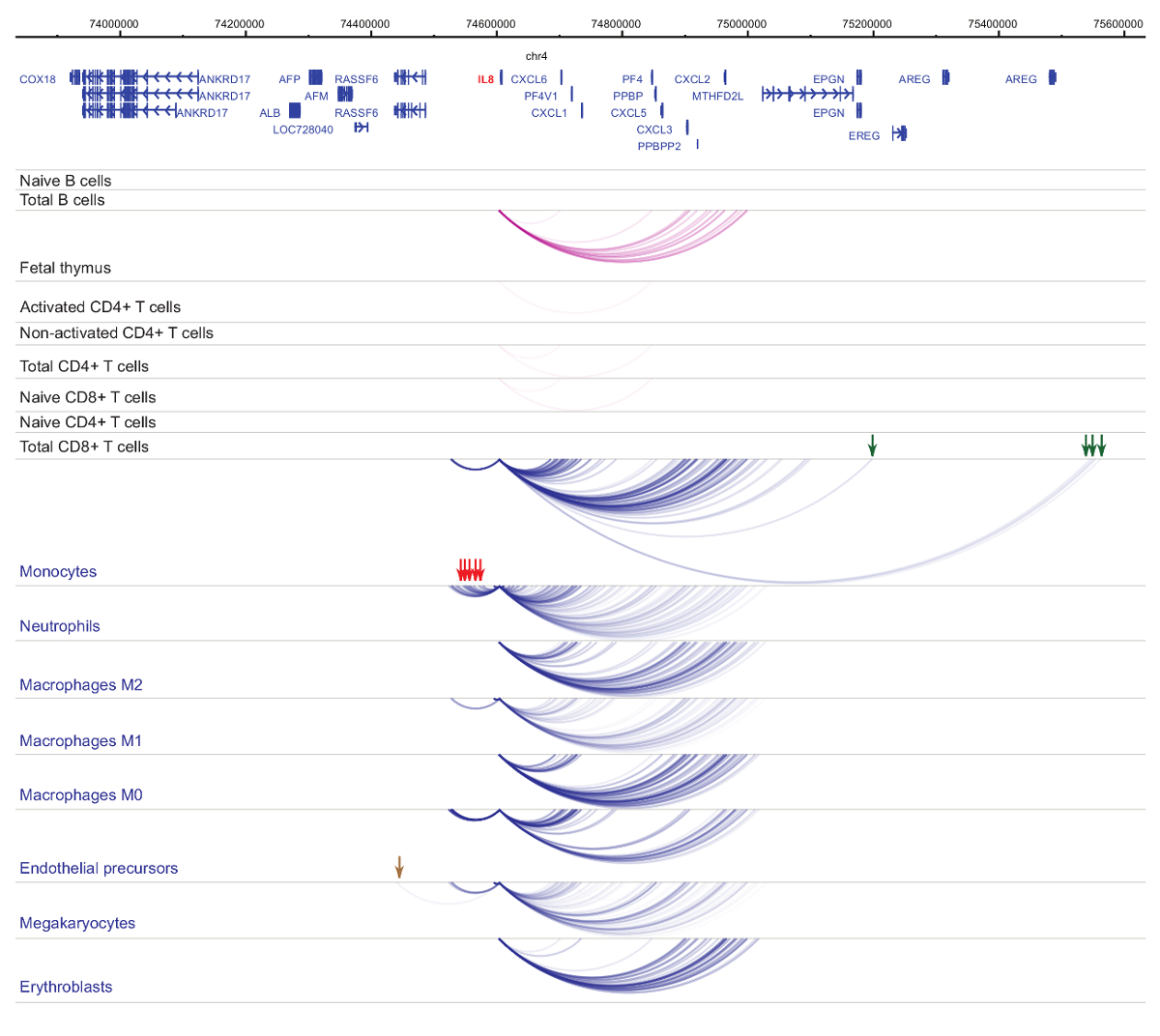
启动子 interactomes 的分析表明细胞类型和谱系特异性33,34,35, 在细胞分化期间有明显变化37,38,39.图 4和5显示了特定发起人的谱系特异性和分化动力学的例子。例如, ALAD组成性表达在所有单元格中, 但其表达式在孵育47中上调。ALAD启动子与所有造血细胞中的几个远端片段接触, 并在孵育中进行额外的相互作用 (图 4)。IL-8显示 B 细胞无统计学意义的相互作用, T 细胞的相互作用非常少, 但在髓系细胞中有许多相互作用, 包括细胞类型的特定相互作用在单核, 中性粒细胞和巨 (图 5)。这些例子说明了 PCHi 如何被用来解开细胞类型的特定 interactomes, 并识别具有监管潜力的启动子交互区域。

图 1: 启动子捕获高 C 理论和捕获诱饵设计.(A) PCHi 的示意图工作流程。在核结扎高 C41,42 (I) 后, 与生物素化 RNA 诱饵 (II) 的溶液杂交, 针对所有人类的限制片段 (这里描述) 或老鼠基因促进者 (III)。(B) PCHi 饵设计。生物素化 rna 捕获诱饵 (红色曲线) 是针对在启动子包含限制片段 (灰色; 注意, 启动子序列本身 (红色) 只针对 RNA 捕获诱饵, 如果他们位于限制片段结束)。通过 RNA 诱饵与 DNA 靶的序列互补杂交, 分离出含有启动子约束片段 (灰色) 及其相互作用限制片段 (黄色和绿色) 的结扎产品, 并随后生物素-链亲和素下拉, 如 A. (C)对一个 rna 诱饵捕获探针与两个 rna 诱饵捕获探针 (参见 B 中的示意图) 的启动子限制片段的 PCHi 捕获效率的比较。请单击此处查看此图的较大版本.
{kind=link}

图 2: PCHi 预排序质量控制.(A)左, 启动子与红外线之间的空间并列示意图, 导致一个高 C 结扎产品, 由一个包含启动子限制片段 (灰色; 启动程序序列红色) 和红外线限制片段 (黄色) 组成。右, DNA 凝胶电泳显示高 C 结扎产品的例子放大使用特定的底漆对 (如图示在左侧)。(B)左, 代表性的例子 HindIII, NheI 和 HindIII/NheI 限制文摘的高 C 结扎产品 (PCR 产品显示在 A)。右, 术后的 DNA 序列示意图失败后 (顶部) 或成功 (底部) dNTP 克莱诺填充的限制连接和随后结扎。(c)代表高 C 图书馆 bioanalyzer 简介 (1/5 稀释)。(D)代表 PCHi 图书馆 bioanalyzer 简介 (不稀释)。请单击此处查看此图的较大版本.
{kind=link}

图 3: PCHi 后测序质量控制。(A)在 PCHi HiCUP43处理后, 与相应的高 c 库 (来自 Javierre等的数据, 201635) 比较有效序列读对的百分比。(B)具有代表性的 HiCUP PCHi 结果显示有效的读对, 以及在下游分析之前丢弃的其他序列类别 (来自 Javierre等的数据, 201635)。(c) PCHi 与相应的 HiCUP 处理后的 "相同片段内部" 读取百分比的比较 (来自 Javierre等的数据, 201635)。(D) PCHi 与相应的高 c 库 (来自 Javierre等的数据, 201635) 的百分比序列读数的比较, 涉及诱饵启动子片段 (捕获效率)。请单击此处查看此图的较大版本.
{kind=link}

图 4: ALAD PCHi 在人类造血细胞中的分布.髓细胞类型的启动子相互作用表现为蓝色拱门, 淋巴细胞类型的启动子相互作用显示为紫色拱门。Erythroblast 特定的相互作用由红色箭头指示 (Javierre等的数据, 201635)。请单击此处查看此图的较大版本.
{kind=link}

图 5:IL8人类造血细胞中的 PCHi 剖面.髓细胞类型的启动子相互作用表现为蓝色拱门, 淋巴细胞类型的启动子相互作用显示为紫色拱门。单核细胞特定的相互作用由绿色箭头指示, 中性粒细胞特定的相互作用由红色箭头指示, 并且巨特定的相互作用由褐色箭头指示 (数据从 Javierre等, 201635)。请单击此处查看此图的较大版本.
{kind=link}
| 人类 | ||||||||
| 名字 | 序列 | 染色体 | 链 | 开始 GRCh38/hg38 | 结束 GRCh38/hg38 | 测试3C 相互作用和生物素结合的底漆组合物 | ||
| hs AHF64 | GCATGCATTAGCCTCTGCTGTTCTCTGAAATC | 11 | + | 116803960 | 116803991 | 与 hs AHF66 结合使用 | ||
| hs AHF66 | CTGTCCAAGTACATTCCTGTTCACAAACCC | 11 | + | 116810219 | 116810248 | 与 hs AHF64 结合使用 | ||
| c-myc 轨迹 | GGAGAACCGGTAATGGCAAA | 8 | - | 127733814 | 127733833 | 与 +1820 或-538 的 c-myc 结合使用 | ||
| c-myc +1820 | AAAATGCCCATTTCCTTCTCC | 8 | + | 129554527 | 129554547 | 与 c-myc 轨迹结合使用 | ||
| c-myc-538 | TGCCTGATGGATAGTGCTTTC | 8 | - | 127195696 | 127195716 | 与 c-myc 轨迹结合使用 | ||
| hs HIST1 F | AAGCAGGAAAAGGCATAGCA | 6 | + | 26207174 | 26207193 | 与 hs HIST1 R 结合使用 | ||
| hs HIST1 R | TCTTGGGTTGTGGGACTTTC | 6 | + | 27771575 | 27771594 | 与 hs HIST1 F 结合使用 | ||
| 鼠标 | ||||||||
| 序列 | 染色体 | 链 | 开始 GRCm38/mm10 | 结束 GRCm38/mm10 | 测试3C 相互作用和生物素结合的底漆组合物 | |||
| TCATGAGTTCCCCACATCTTTG | 8 | + | 84841090 | 84841111 | 与 mm Calr2 结合使用 | |||
| CTGTGGGCACCAGATGTGTAAAT | 8 | + | 84848519 | 84848541 | 与 mm Calr1 结合使用 | |||
| TATCAAGGGTGCCCGTCACCTTCAGC | 6 | + | 125163098 | 125163123 | 与 Gapdh4 结合使用 | |||
| GGGCTTTTATAGCACGGTTATAAAGT | 6 | + | 125163774 | 125163799 | 与 Gapdh3 结合使用 | |||
| GGAGGAGGGAAAAGGAGTGATT | 6 | + | 52212829 | 52212850 | 与 mm Hoxa13 结合使用 | |||
| CAGGCATTATTTGCTGAGAACG | 6 | - | 52253490 | 52253511 | 与 mm Hoxa7 结合使用 | |||
| GGGTAATGGTGTCACTAACTGG | 13 | + | 23571284 | 23571305 | 与 mm Hist1h3e 或 mm Hist1h4i 结合使用 | |||
| GGGTTTGATGAGTTGGTGAAG | 13 | + | 23566541 | 23566561 | 与 mm Hist1h2ae 结合使用 | |||
| TTGGGCCAAAGCCTATATGA | 13 | + | 22043085 | 22043104 | 与 mm Hist1h2ae 结合使用 | |||
表 1: 人类和小鼠高 C 库质量控制的引物序列。
讨论
启动子捕获的模块化设计
启动子捕获高 c 旨在专门丰富的高 c 库的互动涉及发起人。这些相互作用仅包括一个在高 C 库中存在的结扎产品的子集。
捕获高 c 可以很容易地被修改, 以丰富的高 c 库的任何基因组区域或感兴趣的地区通过改变捕获系统。捕获区域可以是连续的基因组片段44,45,46,48, 增强剂已确定在 PCHi (' 反向捕获高 c '35), 或 DNase I 敏感点49.捕获系统的大小可以根据实验范围进行调整。例如, 德莱顿等。目标519诱饵片段在三个基因沙漠与乳腺癌相关44。捕获系统由马丁等 al。目标两个连续基因组片段 (' 区域捕获 ': 211 基因组区域共计; 2131 限制片断) 和选择的促进者 (3857 基因促进者)45。
SureSelect 库可用于不同的大小范围: 1 kb 到 499 kb (5,190–4,806)、500 kb 到 2.9 mb (5,190–4,816) 和 3 mb 到 5.9 mb (5,190–4,831)。由于每个个体捕获生物素-RNA 是120核苷酸长, 这些捕获系统分别容纳 4158, 24166 和49166单独捕获探针。这对应于2079、12083和24583个目标限制片段, 分别 (注意, 限制片段的数字是下限的, 前提是两个单独的捕获探测器可以为每个限制设计片段-在现实中由于重复序列这将不是每个限制片段的情况 (请参见图 1B, C), 从而导致更多的多弹头限制片断为恒定的数量的可用的捕获探针).
此处描述的协议是基于使用限制酶与 6 bp 识别站点来发现远程交互。使用限制酶与 4 bp 识别站点更大的解决更近端的相互作用也是可能的40,49。
PCHi 的局限性
所有染色体构象捕获化验的一个固有的局限性是, 它们的分辨率是由用于库生成的限制酶决定的。在同一限制片段上的 DNA 元素之间发生的相互作用在 "C 型" 检测中是看不见的。此外, 在 PCHi, 在某些情况下, 一个以上的转录起始点可以位于相同的启动子包含限制片段, 和 PIRs 在某些情况下, 港口既活跃和压制组蛋白标记, 使难以确定哪些监管元素中介相互作用, 并预测启动子交互的调节输出。使用限制酶与 4 bp 识别站点缓解了这个问题, 但代价是大大提高了高 c 库的复杂性 (高 c 库生成的 4 bp 识别站点限制酶比高 c 更复杂至少100倍图书馆产生的 6 bp 识别站点限制酶), 以及相关成本的下一代测序。
另一个限制是, 当前的 PCHi 协议要求数以百万计的细胞作为起始材料, 从而排除了在稀有细胞类型中对启动子相互作用的分析。PCHi 的修改版本, 以便在1万到10万细胞 (例如早期胚胎发育或造血干细胞) 的细胞群中对启动子接触进行审问, 因此是捕获的重要补充。高 C 工具箱。
最后, 就像所有依赖于甲醛固定的方法一样, PCHi 只记录在固定时间点 "冷冻" 的相互作用。因此, 为了研究启动子相互作用的动力学和动力学, 需要在 PCHi 的同时, 采用超分辨率活细胞显微术。
高分辨率空间染色体组织的解剖方法
染色体相互作用库的巨大复杂性禁止了两个特定限制片段之间的相互作用产品的可靠识别, 具有统计学意义。为了规避这个问题, 序列捕获已经被用来丰富33,34,40,44或 3C50,51库的特定交互。在富集步骤中使用3C 库的高 c 库的主要优点是, 不同于3C 的高 c, 包括真正结扎产品的浓缩步骤。因此, PCHi 库中有效读取的百分比大约是10倍, 高于捕获 c 库50, 它包含在 HiCUP 筛选之后的5–8% 有效读取。Sahlen等直接将捕获 c 与 HiCap 进行了比较, 如 PCHi 使用高 c 库进行捕获浓缩, 与使用3C 库的捕获 c 形成对比。根据我们的发现, 他们发现捕获 C 库主要由未结扎的片段40组成。此外, HiCap 库的复杂性比捕获 C 库高40。
捕获 c (称为下一代捕获) 的变体-c52 (NG 捕获-c) 使用每个限制片段端的一个寡聚, 如以前在 PCHi33、34中建立的, 而不是原始的重叠探头。捕获-C 协议50。这增加了有效读数的百分比与捕获 c 比较谦虚, 但 NG 捕获-c 使用两个连续回合的捕获浓缩, 和一个相对较高数量的 PCR 周期 (20 到24周期的总和, 比较11周期通常为 PCHi), 这不可避免地会导致更多的序列重复和较低的库复杂性。在 PCHi 的优化过程中, 我们发现唯一的 (即不重复的) 读对的百分比只有大约 15%, 当我们使用 19 PCR 周期 (13 周期预捕获 + 6 周期后捕获; 数据未显示), 但优化到较低数量的 PCR 周期, 通常产生75–90% 唯一的读对。因此, 减少 PCR 周期的数量大大增加信息序列数据的数量。
最近的一种方法是将芯片与高 C 结合在一起, 以关注特定蛋白质介导的染色体相互作用 (HiChIP53)。与嘉-PET54, 这是基于类似的理由, HiChIP 数据包含更多的信息序列读取, 允许更高的信任交互调用53。这将是非常有趣的直接比较相应的 HiChIP 和捕获的高 C 数据集一旦可用 (例如 HiChIP 使用抗体对内聚力联合体单位 Smc1a53与捕获高 c 为所有 Smc1a 绑定限制片段) 并排。这两种方法的一个内在区别是, 捕获高 C 不依赖染色质免疫沉淀, 因此能够审问染色体的相互作用, 而不管蛋白质的占用情况如何。这使得3D 基因组组织在存在或没有特定的因子结合的情况下进行比较, 就像被用来识别 PRC1 作为鼠标 ESC 空间基因组结构的关键调节器7。
PCHi 和 GWAS
全基因组联合研究 (GWAS) 发现, 超过95% 的疾病相关序列变种位于基因组的非编码区域, 通常在很远的距离蛋白质编码基因55。GWAS 变异经常被发现在接近的接近度到 DNase I 敏感站点, 是序列的标志以潜在的调控活动。PCHi 和捕获的高 c 被广泛应用于连接促进剂到 GWAS 的风险位点在乳腺癌44, 大肠癌48, 和自身免疫性疾病35,45,46。对17种不同的人造血细胞类型的 PCHi 研究发现, 与自身免疫性疾病相关的 snp 在淋巴细胞中 PIRs 丰富, 而与血小板和红细胞特异性相关的序列变异主要见于巨噬细胞和孵育分别为35、56。因此, PCHi 发现的组织型特异启动子 interactomes 可以帮助了解非编码疾病相关序列变体的功能, 并确定新的潜在疾病基因进行治疗干预。
启动子相互作用区域的特征
几行证据链启动子 interactomes 基因表达控制。首先, 一些 PCHi 研究表明, 基因组区域与 (高度) 表达基因的促进者相互作用, 在与增强活性相关的标记中丰富, 如 H3K27 乙酰化和 p300 结合33,34,37. 我们发现基因表达水平与相互作用的促进剂数量呈正相关, 这表明促增素的添加剂效应会导致基因表达水平增加34,35。第二, 自然发生的表达数量性状基因位点 (eQTLs) 丰富的 PIRs, 是连接到相同的基因, 其表达受到影响的 eQTLs35。第三, 通过积分57和 PCHi 数据, 凯恩斯等发现, 旅行记者基因映射到 PIRs 在小鼠 ESCs 显示更强的报告基因表达比报告基因在整合站点的非促进者互动区域58、表明 PIRs 具有转录调控活性。这些发现表明, 在各种小鼠和人类细胞类型中 PCHi 发现的启动子 interactomes 包括基因表达控制的关键调控模块。
值得注意的是, 促促剂仅代表 PCHi-C33,34所揭示的所有 PIRs 中的一小部分 (~ 20%)。其他 PIRs 可以有结构或拓扑作用, 而不是直接转录调节功能。然而, 也有证据表明, PCHi 可能发现的 DNA 元素的监管功能, 不庇护经典的增强标记。在人类淋巴细胞系中, BRD7启动子被发现与一个没有增强标记的区域相互作用, 在报告基因检测33中显示有增强活性。具有类似特征的监管要素可能比目前所赞赏的更为丰富。例如, 一个基于 CRISPR 的管理 DNA 元素的屏幕识别了无标记的调控元素 (UREs), 控制基因表达, 但缺乏增强标记59。
在其他情况下, PIRs 已被证明与转录抑制相关的染色质标记。PIRs 和互动促进剂绑定的 PRC1 在小鼠 ESCs 参与了广泛的空间网络的压抑基因的压迫标记 H3K27me37。在人类永生细胞中, 与BCL6启动子相互作用的一个遥远元素抑制了转基因报告基因的表达33, 表明它可以在其原生语境中压制BCL6转录。
PIRs 丰富的染色质绝缘体蛋白 CTCF 在人类 ESCs 和 NECs37可能代表另一类 PIRs。总的来说, 这些结果表明, PIRs 港的基因调控活动的收集尚未功能特征。
披露声明
作者没有什么可透露的。
致谢
我们感谢 Valeriya Malysheva 的手稿和专家帮助的关键阅读图1。这项工作得到了英国医学研究委员会 (MR/L007150/1) 和英国生物技术和生物科学研究委员会 (BB/J004480/1) 的支持。
材料
| Name | Company | Catalog Number | Comments |
| 16% (vol/vol) paraformaldehyde solution | Agar Scientific | R1026 | |
| Dulbecco's Modified Eagle Medium (DMEM) 1x | Life Technologies | 41965-039 | |
| Fetal bovine serum (FBS) sterile filtered | Sigma | F9665 | |
| Low-retention filter tips | Starlab | S1180-3810, S1180-1810, S1180-8810 and S1182-1830 | |
| 10x PBS pH 7.4 | Life Technologies | 70011-036 | |
| Molecular biology grade water | Sigma-Aldrich | W4502 | |
| 1 M Tris-HCl pH 8.0 | Life Technologies | 15568-025 | |
| IGEPAL CA-630 | Sigma-Aldrich | I8896 | |
| 5 M NaCl | Life Technologies | 24740-011 | |
| Protease inhibitor cocktail (EDTA-free) | Roche Diagnostics | 11873580001 | |
| Restriction buffer 2 (10x NEBuffer 2) | New England Biolabs | B7002 | |
| DNA LoBind tube, 1.5 mL | Eppendorf | 0030 108.051 | |
| DNA LoBind tube, 2 mL | Eppendorf | 30108078 | |
| 20% (wt/vol) SDS | Bio-Rad Laboratories | 161-0418 | |
| 20% (vol/vol) Triton X-100 | Sigma-Aldrich | T8787 | |
| HindIII, 100 U/uL | New England Biolabs | R0104 | |
| 10 mM dCTP | Life Technologies | 18253-013 | |
| 10 mM dGTP | Life Technologies | 18254-011 | |
| 10 mM dTTP | Life Technologies | 18255-018 | |
| 0.4 mM Biotin-14-dATP | Life Technologies | 19524-016 | |
| DNA polymerase I large (Klenow) fragment 5000 units/mL | New England Biolabs | M0210 | |
| 10x T4 DNA ligase reaction buffer | New England Biolabs | B0202 | |
| 100x 10mg/ml Bovine Serum Albumin | New England Biolabs | B9001 | |
| T4 DNA ligase, 1 U/μL | Invitrogen | 15224-025 | |
| RNase A | Roche | 10109142001 | |
| Proteinase K, recombinant, PCR grade | Roche | 3115836001 | |
| 20 000×g 50 ml centrifuge tube | VWR | 525-0156 | |
| 0.5 M EDTA pH 8.0 | Life Technologies | 15575-020 | |
| Phenol pH 8.0 | Sigma | P4557 | |
| Phenol: Chloroform: Isoamyl Alcohol 25:24:1 | Sigma | P3803 | |
| Sodium acetate pH 5.2 | Sigma | S7899 | |
| Quant-iT PicoGreen | Invitrogen | P7589 | |
| QIAquick Gel Extraction Kit | Qiagen | 28704 | |
| QIAquick PCR Purification Kit | Qiagen | 28104 | |
| Restriction buffer 2.1 (10x NEBuffer 2.1) | New England Biolabs | B7202 | |
| NheI, 100U/uL | New England Biolabs | R0131 | |
| Micro TUBE AFA Fiber Pre-slit snap cap 6x16mm vials | Covaris | 520045 | For sonication |
| SPRI beads (Agencourt AMPure XP) | Beckman Coulter | A63881 | |
| Dynabeads MyOne Streptavidin C1 beads | Invitrogen | 65001 | |
| Tween 20 | Sigma | P9416 | |
| 10 mM dATP | Life Technologies | 18252-015 | |
| T4 DNA polymerase 3000 units/mL | New England Biolabs | M0203 | |
| T4 PNK 10000 units/mL | New England Biolabs | M0201 | |
| Klenow exo minus 5000 units/mL | New England Biolabs | M0212 | |
| Quick ligation reaction buffer | New England Biolabs | B6058 | |
| NEB DNA Quick ligase | New England Biolabs | M2200 | |
| PE adapter 1.0 (5'-P-GATCGGAAGAGCGGTTCAGC AGGAATGCCGAG-3') | Illumina | ||
| PE adapter 2.0 (5'-ACACTCTTTCCCTACACGACGCT CTTCCGATCT-3') | Illumina | ||
| NEB Phusion PCR kit | New England Biolabs | M0530 | |
| PE PCR primer 1.0 (5'-AATGATACGGCGACCACCGA GATCTACACTCTTTCCCTAC ACGACGCTCTTCCGATCT-3') | Illumina | ||
| PE PCR primer 2.0 (5'-CAAGCAGAAGACGGCATACGA GATCGGTCTCGGCATTCCT GCTGAACCGCTCTTCCGATCT-3') | Illumina | ||
| PCR strips | Agilent Technologies | 410022 and 401425 | |
| SureSelect SSEL TE Reagent ILM PE full adaptor kit | Agilent Technologies | 931108 | |
| SureSelect custom 3-5.9 Mb library | Agilent Technologies | 5190-4831 | custom design mouse or human PCHi-C system |
| Dynabeads MyOne Streptavidin T1 beads | Invitrogen | 65601 | |
| E220 high-performance focused ultra-sonicator | Corvaris | E220 |
参考文献
- Osborne, C. S., et al. Active genes dynamically colocalize to shared sites of ongoing transcription. Nature Genetics. 36, 1065-1071 (2004).
- Schoenfelder, S., et al. Preferential associations between co-regulated genes reveal a transcriptional interactome in erythroid cells. Nature Genetics. 42, 53-61 (2010).
- de Wit, E., et al. The pluripotent genome in three dimensions is shaped around pluripotency factors. Nature. 501, 227-231 (2013).
- Bantignies, F., et al. Polycomb-dependent regulatory contacts between distant Hox loci in Drosophila. Cell. 144, 214-226 (2011).
- Engreitz, J. M., et al. The Xist lncRNA exploits three-dimensional genome architecture to spread across the X chromosome. Science. 341, 1237973(2013).
- Denholtz, M., et al. Long-range chromatin contacts in embryonic stem cells reveal a role for pluripotency factors and polycomb proteins in genome organization. Cell Stem Cell. 13, 602-616 (2013).
- Schoenfelder, S., et al. Polycomb repressive complex PRC1 spatially constrains the mouse embryonic stem cell genome. Nature Genetics. 47, 1179-1186 (2015).
- Kundu, S., et al. Polycomb Repressive Complex 1 generates discrete compacted domains that change during differentiation. Molecular Cell. 65, 432-446 (2017).
- Skok, J. A., Gisler, R., Novatchkova, M., Farmer, D., de Laat, W., Busslinger, M. Reversible contraction by looping of the Tcra and Tcrb loci in rearranging thymocytes. Nature Immunology. 8, 378-387 (2007).
- Zhang, Y., et al. Spatial organization of the mouse genome and its role in recurrent chromosomal translocations. Cell. 148, 908-921 (2012).
- Aymard, F., et al. Genome-wide mapping of long-range contacts unveils clustering of DNA double-strand breaks at damaged active genes. Nature Structural & Molecular Biology. 24, 353-361 (2017).
- Ryba, T., et al. Evolutionarily conserved replication timing profiles predict long-range chromatin interactions and distinguish closely related cell types. Genome Research. 20, 761-770 (2010).
- Pope, B. D., et al. Topologically associating domains are stable units of replication-timing regulation. Nature. 515, 402-405 (2014).
- Chandra, T., et al. Global reorganization of the nuclear landscape in senescent cells. Cell Reports. 10, 471-483 (2015).
- Carter, D., Chakalova, L., Osborne, C. S., Dai, Y. F., Fraser, P. Long-range chromatin regulatory interactions in vivo. Nature Genetics. 32, 623-626 (2002).
- Tolhuis, B., Palstra, R. J., Splinter, E., Grosveld, F., de Laat, W. Looping and interaction between hypersensitive sites in the active beta-globin locus. Molecular Cell. 10, 1453-1465 (2002).
- Amano, T., Sagai, T., Tanabe, H., Mizushina, Y., Nakazawa, H., Shiroishi, T. Chromosomal dynamics at the Shh locus: limb bud-specific differential regulation of competence and active transcription. Developmental Cell. 16, 47-57 (2009).
- Zuniga, A., et al. Mouse limb deformity mutations disrupt a global control region within the large regulatory landscape required for Gremlin expression. Genes & Development. 18, 1553-1564 (2004).
- Sagai, T., Hosoya, M., Mizushina, Y., Tamura, M., Shiroishi, T. Elimination of a long-range cis-regulatory module causes complete loss of limb-specific Shh expression and truncation of the mouse limb. Development. 132, 797-803 (2005).
- D'Haene, B., et al. Disease-causing 7.4 kb cis-regulatory deletion disrupting conserved non-coding sequences and their interaction with the FOXL2 promotor: implications for mutation screening. PLoS Genet. 5, e1000522(2009).
- Sur, I. K., et al. Mice lacking a Myc enhancer that includes human SNP rs6983267 are resistant to intestinal tumors. Science. 338, 1360-1363 (2012).
- Herranz, D., et al. A NOTCH1-driven MYC enhancer promotes T cell development, transformation and acute lymphoblastic leukemia. Nature Medicine. 20, 1130-1137 (2014).
- Deng, W., et al. Controlling long-range genomic interactions at a native locus by targeted tethering of a looping factor. Cell. 149, 1233-1244 (2012).
- Groschel, S., et al. A single oncogenic enhancer rearrangement causes concomitant EVI1 and GATA2 deregulation in leukemia. Cell. 157, 369-381 (2014).
- Lupianez, D. G., et al. Disruptions of topological chromatin domains cause pathogenic rewiring of gene-enhancer interactions. Cell. 161, 1012-1025 (2015).
- Franke, M., et al. Formation of new chromatin domains determines pathogenicity of genomic duplications. Nature. 538, 265-269 (2016).
- Dekker, J., Rippe, K., Dekker, M., Kleckner, N. Capturing chromosome conformation. Science. 295, 1306-1311 (2002).
- Simonis, M., et al. Nuclear organization of active and inactive chromatin domains uncovered by chromosome conformation capture-on-chip (4C). Nature Genetics. 38, 1348-1354 (2006).
- Zhao, Z., et al. Circular chromosome conformation capture (4C) uncovers extensive networks of epigenetically regulated intra- and interchromosomal interactions. Nature Genetics. 38, 1341-1347 (2006).
- Dostie, J., et al. Chromosome Conformation Capture Carbon Copy (5C): A massively parallel solution for mapping interactions between genomic elements. Genome Research. 16, 1299-1309 (2006).
- Lieberman-Aiden, E., et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science. 326, 289-293 (2009).
- Belton, J. M., McCord, R. P., Gibcus, J. H., Naumova, N., Zhan, Y., Dekker, J. Hi-C: a comprehensive technique to capture the conformation of genomes. Methods. 58, 268-276 (2012).
- Mifsud, B., et al. Mapping long-range promoter contacts in human cells with high-resolution capture Hi-C. Nature Genetics. 47, 598-606 (2015).
- Schoenfelder, S., et al. The pluripotent regulatory circuitry connecting promoters to their long-range interacting elements. Genome Res. 25, 582-597 (2015).
- Javierre, B. M., et al. Lineage-specific genome architecture links enhancers and non-coding disease variants to target gene promoters. Cell. 167, 1369-1384 (2016).
- Wilson, N. K., et al. Integrated genome-scale analysis of the transcriptional regulatory landscape in a blood stem/progenitor cell model. Blood. 127, e12-e23 (2016).
- Freire-Pritchett, P., et al. Global reorganisation of cis-regulatory units upon lineage commitment of human embryonic stem cells. Elife. 6, (2017).
- Rubin, A. J., et al. Lineage-specific dynamic and pre-established enhancer-promoter contacts cooperate in terminal differentiation. Nature Genetics. 49, 1522-1528 (2017).
- Siersbaek, R., et al. Dynamic rewiring of promoter-anchored chromatin loops during adipocyte differentiation. Molecular Cell. 66, 420-435 (2017).
- Sahlen, P., et al. Genome-wide mapping of promoter-anchored interactions with close to single-enhancer resolution. Genome Biology. 16, 156(2015).
- Nagano, T., et al. Single-cell Hi-C reveals cell-to-cell variability in chromosome structure. Nature. 502, 59-64 (2013).
- Nagano, T., Varnai, C., Schoenfelder, S., Javierre, B. M., Wingett, S. W., Fraser, P. Comparison of Hi-C results using in-solution versus in-nucleus ligation. Genome Biology. 16, 175(2015).
- Wingett, S., et al. HiCUP: pipeline for mapping and processing Hi-C data. F1000 Res. 4, 1310(2015).
- Dryden, N. H., et al. Unbiased analysis of potential targets of breast cancer susceptibility loci by Capture Hi-C. Genome Research. 24, 1854-1868 (2014).
- Martin, P., et al. Capture Hi-C reveals novel candidate genes and complex long-range interactions with related autoimmune risk loci. Nature Communications. 6, 10069(2015).
- McGovern, A., et al. Capture Hi-C identifies a novel causal gene, IL20RA, in the pan-autoimmune genetic susceptibility region 6q23. Genome Biol.ogy. 17, 212(2016).
- Hodge, D., et al. A global role for EKLF in definitive and primitive erythropoiesis. Blood. 107, 3359-3370 (2006).
- Jager, R., et al. Capture Hi-C identifies the chromatin interactome of colorectal cancer risk loci. Nature Communications. 6, 6178(2015).
- Joshi, O., et al. Dynamic reorganization of extremely long-range promoter-promoter Interactions between two states of pluripotency. Cell Stem Cell. 17, 748-757 (2015).
- Hughes, J. R., et al. Analysis of hundreds of cis-regulatory landscapes at high resolution in a single, high-throughput experiment. Nature Genetics. 46, 205-212 (2014).
- Kolovos, P., et al. Targeted Chromatin Capture (T2C): A novel high-resolution high-throughput method to detect genomic interactions and regulatory elements. Epigenetics Chromatin. 7, 10(2014).
- Davies, J. O., et al. Multiplexed analysis of chromosome conformation at vastly improved sensitivity. Nature Methods. 13, 74-80 (2016).
- Mumbach, M. R., et al. HiChIP: efficient and sensitive analysis of protein-directed genome architecture. Nature Methods. 13, 919-922 (2016).
- Fullwood, M. J., et al. An oestrogen-receptor-alpha-bound human chromatin interactome. Nature. 462, 58-64 (2009).
- Maurano, M. T., et al. Systematic localization of common disease-associated variation in regulatory DNA. Science. 337, 1190-1195 (2012).
- Petersen, R., et al. Platelet function is modified by common sequence variation in megakaryocyte super enhancers. Nat. Commun. 8, 16058(2017).
- Akhtar, W., et al. Chromatin position effects assayed by thousands of reporters integrated in parallel. Cell. 154, 914-927 (2013).
- Cairns, J., et al. CHiCAGO: Robust detection of DNA looping interactions in Capture Hi-C data. Genome Biology. 17, 127(2016).
- Rajagopal, N., et al. High-throughput mapping of regulatory DNA. Nature Biotechnology. 34, 167-174 (2016).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。